
Submitted:
29 April 2023
Posted:
29 April 2023
You are already at the latest version
Abstract
A new spirurid nematode Protospirura canariensis n. sp., parasite of the black rat Rattus rattus (Rodentia: Muridae) in El Hierro Island (Canary Archipelago, Spain) is described by means of light (LM) and scanning electron microscopy (SEM). The most discriminant characteristics between the new species and the existing species in the genus Protospirura are: (a) the number of tooth-like prominences in the submedian and lateral lobes of pseudolabia, both in males and females (2 and 4, respectively), (b) the size of right and left spicules in males (643–715 µm and 309–412 µm, respectively), and (c) the number and arrangement of cloacal papillae in males. The new species has a total of 17 cloacal papillae (four large and pedunculated pairs of precloacal papillae, an unpaired precloacal papilla and four pairs of postcloacal papillae). The arrangement of postcloacal papillae are as follows: the first pair are large, pedunculated and placed near the posterior edge of cloaca; the three remaining postcloacal pairs are grouped and located near the posterior tip. In the latter group, papillae in the first pair are large and pedunculated. Parasitized host and geographical distribution are also useful criteria to distinguish P. canariensis n. sp. from the remaining species of the genus Protospirura. In addition, the cytochrome c oxidase subunit 1 (cox1) sequence of the new species is provided and compared with available data of related species.
Keywords:
Protospirura canariensis n. sp.
; Spiruridae
; Nematoda
; Rattus rattus
; Muridae
; Canary Islands
1. Introduction
The taxonomic status of the cosmopolitan genus Protospirura Seurat, 1914 (Spiruridae) has been confusing for a long time, particularly due to the morphological similitudes with the genus Mastophorus Diesing, 1853 (Spirocercidae). According to several authors [1,2], this confusion is due to the consideration of inappropriate characters. In this sense, Chitwood [1] established the differential characteristics between the genera Protospirura and Mastophorus: the number of teeth in the pseudolabia (two or four in Protospirura vs. three, five, seven or nine in Mastophorus), the morphology of the pharynx (laterally compressed in Protospirura vs. cylindrical in Mastophorus), the morphology of cloacal papillae in males (sessile in Protospirura vs. pedunculated in Mastophorus), the tail length in males (short in Protospirura vs. long in Mastophorus) and the position of the vulva in females (postequatorial in Protospirura vs. preequatorial in Mastophorus). However, some of these characteristics are not present in all the currently accepted species of the genus Protospirura. In fact, some Protospirura species have pedunculated cloacal papillae [3,4,6,10,16] or the vulva is located anteriorly to mid-body [5,11,16]. Also, the morphology of pseudolabia in the genera Protospirura and Mastophorus is quite different. In Protospirura the four submedian lobes are clearly less developed than the lateral lobes whereas in Mastophorus the oral opening is hexagonal and the submedian lobes of pseudolabia are well developed and quadrangular [17].
Currently, the genus Protospirura comprises 13 species parasites of mammals included in five orders and in 18 families: Artiodactyla (Bovidae), Carnivora (Canidae, Felidae and Viverridae), Eulipotyphla (Erinaceidae and Talpidae), Primates (Aotidae, Atelidae, Cebidae, Cercopythecidae, Hominidae and Lorisidae) and Rodentia (Bathyergidae, Cricetidae, Heteromyidae, Muridae, Nesomyidae and Sciuridae) [3,4,5,6,7,8,9,10,11,12,13,14,15,16,18,19]. Except for P. muricola, which has a worldwide distribution including Africa, the Caribbean region, Central and South America, Southeast Asia and Europe [14], the remaining species of this genus have a limited geographical distribution. With respect to the parasitized hosts, only seven species of Protospirura have been recorded in Muridae rodents. These are P. armeniana, P. chabaudi, P. kaindiensis, P. munimuniensis, P. muricola, P. okinavensis and P. siamensis [8,9,13,14,15,16,18,19]. Other species described as belonging to the genus Protospirura, namely P. ascaroidea, P. bestiarum, P. columbiana, P. glareoli, P. gracilis, P. labiodenta and P. marsupialis were considered synonyms of M. muris by Chitwood [1]. These synonymies were further confirmed by Wertheim [2]. Additionally, other species described posteriorly, namely P. chanchanensis, P. paucidentata and P. srivastavai should be included in the Spirocercidae because their pharynx is not laterally compressed [8]. Finally, P. bonnei was considered synonym of P. muricola and P. congolense was transferred to the genus Mastophorus by Quentin [20].
In the present study, we describe a new species, Protospirura canariensis n. sp., parasitizing the black rat (Rattus rattus) in El Hierro Island (Canary Archipelago, Spain). Additionally, the sequence of the mitochondrial cytochrome c oxidase subunit I gene (cox1) is provided and compared with data of related species.
2. Materials and Methods
2.1. Specimens
Specimens of Protospirura canariensis n. sp. were recovered from the stomach of several black rats Rattus rattus Linnaeus, 1758 (Rodentia, Muridae) captured in Lagartario-Frontera (27° 46' 29.9" N, 17° 59' 55.59" W), Camino-Frontera (27° 44' 38.07" N, 18° 2' 25.4" W) and Túnel-Valverde (27° 49' 12.41" N, 17° 57' 49.27" W) (El Hierro Island, Canary Archipelago, Spain) (Figure 1) during several trapping campaigns in 2008, 2009 and 2010. The studied rats were captured using Sherman traps (H.B. Sherman Traps, Inc., Tallahassee, FL, USA) and wire-mesh traps of Manufrance type (Saint-Étienne, France) or Firobind type (Besançon, France), sacrificed by cervical dislocation and then examined for gastrointestinal helminths under the stereomicroscope.
2.2. Light microscopy study
Specimens were mounted in Amann lactophenol on slides and then observed under the light microscope (LM) Leica DMLB (Leica Microsystems, Wetzlar, Germany). Drawings were made with the aid of a drawing tube and later modified using Adobe Illustrator software (Adobe, San José, CA, USA).
2.3. Scanning electron microscopy study
Some worms (three males and six females) were preserved for scanning electron microscopy (SEM) examination. Initially they were fixed in 70% ethanol in the field and later, in the laboratory, they were dehydrated in an ethanol series and critical point dried with carbon dioxide (Emitech K850X, Quorum Technologies Ltd., Laughton, East Sussex, UK). Finally, specimens were mounted on stubs with conductive adhesive tape and colloidal silver, coated with carbon in an Emitech K950X evaporator (Quorum Technologies Ltd.), and examined using a Field Emission Scanning Electron Microscope JSM-7001F (JEOL Ltd., Tokyo, Japan) at 10 kV in the "Centres Científics i Tecnològics" of the University of Barcelona (CCiTUB).
2.3. Molecular analyses and phylogenetic tree
Genomic DNA was extracted using fragments of Protospirura canariensis n. sp. specimens that had been reserved for this purpose. A total of 10 nematode fragments were extracted. The fragments were deposited in tubes containing a mixture of 250 µl of a lysis solution composed of 30 mM Tris-HCL pH 8.0, mM EDTA and 0.4% SDS. In addition, 3 µl of proteinase K (20 ml-1) (PanReac AppliChem ITW Reagents, Darmstadt, Germany) was added and then incubated at 56 °C overnight. The following day, 250 µl of NH4Ac 4 M was added, mixed thoroughly and subsequently incubated 30 min at room temperature (15 °C - 25 °C). The mix was spun for 10 min at 13,000 rpm, and the pellet was discarded. DNA was then precipitated from the supernatant with ethanol, and the pellet was resuspended in 200 µl of 1X TE (10 mM Tris-HCL pH 8.1 mM EDTA) [21]. The quantity and quality of genomic DNA was checked using DeNovix DS-11 + Spectrophotometer (DeNovix Inc., Wilmington, DE, USA).
PCR screening of DNA was based on cytochrome c oxidase subunit 1 (cox1) using the primers COIIntF and COIIntR described by Gaillard et al. [22]. The PCR amplification contained 1X Buffer (VWR), 0.2 mM of each dNTP (VWR), 1.5 mM MgCl2 (VWR), 20-40 ng of total genomic DNA in a total volume of 50 µl. Amplification was conducted with XP Cycler (Hangzhou Bioer Technology Co. Ltd., Hangzhou, China) using the following parameters: 94 °C for 3 min; 35 cycles at 94 °C for 30 s, 54 °C for 30 s, 68 °C for 45 s; and a final extension al 68 °C for 10 min [22]. The resulting amplifications were visualized on 1.5% agarose gel at 90 V for 1 hr.
The PCR products that presented the expected size (650-bp) were sequenced at Macrogen Spain Inc. (Madrid, Spain) with primers COIIntF/COIIntR [22]. The sequences obtained using the Sanger method were interpreted with the MEGA X software [23] using the multiple alignment program ClustalW included in MEGA X, and minor corrections were made by hand. Subsequently analyzed with the basic local alignment search tool (BLAST), and the identity confirmed by homology comparison.
Phylogenetic relationships based on the Maximum Likelihood method were carried out with the p-distance and Kimura 2-parameter model [24] and 1,000 bootstrap replications based on the cytochrome c oxidase subunit 1 (cox1) gene sequences exploring the relationships among species. The sequence Schistosoma mansoni NC002545 was used as the outgroup (Figure 2).
3. Results
3.1. Taxonomic summary
Family Spiruridae Oerley, 1885
Genus Protospirura Seurat, 1914
Protospirura canariensis n. sp. (Figure 3A–F, Figure 4A–D, Figure 5A–E, Figure 6A–C and Figure 7A–C)
Type host: Rattus rattus Linnaeus, 1758 (Rodentia: Muridae).
Type locality: Lagartario-Frontera (El Hierro Island, Canary Archipelago, Spain) (27° 46' 29.9" N, 17° 59' 55.59" W) (Figure 1).
Other localities: Camino-Frontera (El Hierro Island, Canary Archipelago, Spain) (27° 44' 38.07" N, 18° 2' 25.4" W), Túnel-Valverde (El Hierro Island, Canary Archipelago, Spain) (27° 49' 12.41" N, 17° 57' 49.27" W) (Figure 1).
Site of infection: stomach.
Type specimens: deposited in the "Muséum National d'Histoire Naturelle" (Paris, France) under the following accession numbers:
- holotype,  No. 1, MNHN HEL1905.
No. 1, MNHN HEL1905.
No. 1, MNHN HEL1905.- allotype,  No. 1, MNHN HEL1906.
No. 1, MNHN HEL1906.
No. 1, MNHN HEL1906.- 19 paratypes:
- 7 males: ♂︎No. 2, MNHN HEL1907; ♂︎No. 4, MNHN HEL1908; ♂︎No. 5, MNHN HEL1909; ♂︎No. 7, MNHN HEL1910; ♂︎No. 8, MNHN HEL1911; ♂︎No. 10, MNHN HEL1912 and ♂︎No. 11, MNHN HEL1913.
- 12 females: ♀︎No. 2, MNHN HEL1914; ♀︎No. 3, MNHN HEL1915; ♀︎No. 4, MNHN HEL1916; ♀︎No. 5, MNHN HEL1917; ♀︎No. 6, MNHN HEL1918; ♀︎No. 7, MNHN HEL1919; ♀︎No. 8, MNHN HEL1920; ♀︎No. 9, MNHN HEL1921; ♀︎No. 10, MNHN HEL1922; ♀︎No. 11, MNHN HEL1923; ♀︎No. 12, MNHN HEL1924 and ♀︎No. 13, MNHN HEL1925.
Mitochondrial cytochrome c oxidase subunit I gene (cox1) sequence: a fragment of 650-bp was obtained for the cox1. Four fragments of 409-bp were successfully sequenced and submitted to the GenBank database; accession numbers: OQ799521, OQ799522, OQ799523 and OQ799524.
Etymology: the specific name of this nematode refers to its geographical distribution.
3.2. Description
General: Large stout worms. Cuticle thick, with transverse striations. Anterior extremity with two highly developed pseudolabia raised above the mouth opening (Figure 3 3A,B, Figure 5A and Figure 6A–C). Each pseudolabium formed by a well-developed lateral lobe and two smaller submedian lobes (Figure 3B, Figure 5A and Figure 6A–C). Each lateral lobe has four tooth-like prominences in its internal side (Figure 3B, Figure 5A and Figure 7C). Each submedian lobe has two tooth-like prominences in its internal side (Figure 3B, Figure 5A and Figure 7C). Four pairs of submedian cephalic papillae at the base of the submedian lobes and two amphids in the lateral lobes (Figure 3B, Figure 5A and Figure 7A,B). Pharynx thick-walled, laterally compressed (Figure 5A and Figure 6B). Oesophagus divided into anterior muscular and posterior glandular portions (Figure 3A). Nerve ring surrounding oesophagus at the level of its muscular portion (Figure 3A). Excretory pore slightly posterior to nerve ring (Figure 3A and Figure 7A). Deirids placed between nerve ring and excretory pore (Figure 7A). Phasmids not seen.
Male (11 specimens measured, range, mean in parentheses, holotype measurements in brackets) (Figure 3C,E, Figure 4A–D and Figure 5A–E) (all measurements are given in micrometres, except where indicated): Oral opening surrounded by two tri-lobed pseudolabia (Figure 5A). Presence of four tooth-like prominences in the lateral lobes and two tooth-like prominences in the submedian lobes (Figure 5A). Body length 13.67–23.37 mm (18.43 mm) [15.02 mm]; width at the level of the oesophagus basis 330–526 (430) [382]. Pharynx length 39–69 (56) [>57]. Muscular oesophagus length 219–340 (280) [219]; width at base 75–103 (82) [75]. Glandular oesophagus length 2.95–4.67 mm (3.97 mm) [3.75 mm]; width at base 123–206 (167) [165]. Nerve ring located at 123–273 (217) [123] from the cephalic extremity. Deirids located at 213–308 (250) [231] from the cephalic extremity. Excretory pore located at 371–567 (474) [351] from the cephalic extremity. Posterior end of body strongly curved ventrally. The pericloacal surface is ornamented with cuticular markings (Figure 3C, Figure 4A and Figure 5B–E). Total of 17 caudal papillae; four pairs of large and pedunculated precloacal papillae (pairs 1 to 4) (Figure 3C, Figure 4A and Figure 5B), one large unpaired precloacal papilla (Figure 3C, Figure 4B and Figure 5B) and four pairs of postcloacal papillae (pairs 5 to 8) (Figure 3C, Figure 4C and Figure 5B,C). The postcloacal papillae in pair 5 are large, pedunculated and placed near the posterior edge of the cloaca (Figure 3C, Figure 4C and Figure 5B,C). The remaining three pairs of postcloacal papillae (pairs 6 to 8) are grouped and placed near the posterior tip (Figure 3C, Figure 4C and Figure 5B,C). The papillae of pair 6 are large and pedunculated (Figure 3C, Figure 4C and Figure 5B,C). The papillae of pairs 7 and 8 are smaller and apparently sessile (Figure 3C, Figure 4C and Figure 5B,C). Phasmids were not observed. Spicules unequal in size and shape; right spicule longer and slender 643–715 (675) [694] (Figure 3C and Figure 4D); left spicule shorter, stout and alate 309–412 (368) [360] (Figure 3C and Figure 4D). Gubernaculum, V-shaped, 116–159 (141) [123] (Figure 3D).
Female (13 specimens measured, range, mean in parentheses, allotype measurements in brackets) (Figure 3A,B,D,F and Figure 6A–C) (all measurements are given in micrometres, except where indicated): Oral opening surrounded by two tri-lobed pseudolabia (Figure 3A,B and Figure 6A–C). Presence of four tooth-like prominences in the lateral lobes and two tooth-like prominences in the submedian lobes (Figure 6C). Body length 25.52–41.99 mm (32.51 mm) [27.73 mm]; width at the level of the vulva 795–1455 (1137) [815]. Pharynx length 57–90 (74) [87]. Muscular oesophagus length 273–464 (346) [310]; width at base 108–175 (128) [113]. Glandular oesophagus length 3.49–5.79 mm (4.52 mm) [3.84 mm]; width at base 165–310 (230) [185]. Nerve ring located at 234–311 (278) [311] from the cephalic extremity (Figure 3A). Deirids located at 273–360 (317) [309] from the cephalic extremity. Excretory pore located at 464–660 (577) [309] from the cephalic extremity (Figure 3A). Vulva slightly preequatorial, located at a 11.92–19.43 mm (15.22 mm) [13.33 mm] distance from the cephalic extremity. Tail 258–423 (327) [289] (Figure 3D). Eggs, embryonated, 48.9–56.7 x 33.4–41.1 (51.8 x 38.2) [51.4 x 38.6] (Figure 3F).
L3 larva (1 specimen measured) (Figure 7A–C) (all measurements are given in micrometres, except where indicated): Body length 5.91 mm; width at the level of the oesophagus basis 162. Pharynx length 33. Muscular oesophagus length 159; width at base 39. Glandular oesophagus length 1.62 mm; width at base 67. Nerve ring located at 121 from the cephalic extremity. Deirids not observed. Excretory pore located at 270 from the cephalic extremity. Anus at 495 from the posterior extremity. Tail notched, with 10 pointed prominences arranged circularly in the same plan (Figure 7C).
3.3. Molecular analyses
All genomic DNA samples were amplified to the expected size (650-bp). Four of these amplicons were selected for sequencing as all the fragments had the same host species (R. rattus) and had the same origin (Frontera, El Hierro, Canary Islands, Spain), this was done in such a way for resource optimization. Four 409-bp fragments were successfully sequenced and sent to the GenBank database with accession numbers OQ799521, OQ799522, OQ799523 and OQ799524.
Phylogenetic analyses of the cox1 gene showed a tree composed of six monophyletic groups (Figure 2). We observe in the upper part a clade composed of the specimens of Protospirura canariensis n. sp. contributed by this study. Below is a clade made up of the species of Protospirura muricola with a robustness of 100%. Both clades are clearly separated from the rest of the included sequences.
4. Discussion
One of the most useful morphological characteristics in the recognition and differentiation of Protospirura species concerns the morphology of pseudolabia and their associated structures. The oral opening is surrounded by two pseudolabia, each with three lobes (a larger lateral lobe and two smaller submedian lobes). In the internal face of these lobes there are tooth-like prominences variable in number (usually two or four). Protospirura canariensis n. sp. has four tooth-like prominences in each lateral lobe and two tooth-like prominences in each submedian lobe. Only P. peromysci exhibit a similar arrangement of tooth-like prominences in pseudolabia lobes [3]. However, there are some discrepancies among earlier works that described several Protospirura species, including P. peromysci (see Table 1). Thus, Babero and Mathias [3] in the original description of the species mention the presence of four flat denticles in the lateral lobes and two triangular denticles in submedian lobes, while Smales [9] considers this species to have four teeth in both lateral and submedian lobes. In male specimens, P. canariensis n. sp. can be differentiated from P. peromysci in the size of the right spicule (respectively 643–715 µm vs. 820–1200 µm) and in the number of cloacal papillae (17 in P. canariensis n. sp. vs. 23 in P. peromysci) [3].
In male specimens, the size of spicules is another diagnostic character when the species of Protospirura are compared. Considering the size of both right and left spicules P. pseudomuris and P. siamensis present similar sized spicules: right spicule measures 643–715 µm in P. canariensis n. sp., 540–790 µm in P. pseudomuris and 469–701 µm in P. siamensis; left spicule measures 309–412 µm in P. canariensis n. sp., 270–380 µm in P. pseudomuris and 348–380 µm in P. siamensis [7,13]. Both species can be differentiated from the new species in the number of tooth-like prominences in lobes of pseudolabia (see Table 1). Additionally, the number of cloacal papillae also distinguish the new species from P. pseudomuris (17 vs. 21, respectively). In the remaining Protospirura species, spicule sizes are out of the range of those of P. canariensis n. sp. (see Table 1).
The number, morphology and disposition of cloacal papillae are other differential characters between P. canariensis n. sp. and the remaining species of the genus Protospirura. Thus, P. canariensis n. sp. has a total of 17 cloacal papillae, 4 pedunculated precloacal pairs, an unpaired precloacal papilla located in the front edge of cloaca and 4 postcloacal pairs (1 pedunculated pair near the cloaca and 1 pedunculated and 2 sessile pairs near the posterior tip). Only two species have a similar number of cloacal papillae. The first of them is P. mexicana, which has 17 cloacal papillae with a similar arrangement, although Falcón & Sanabria [4] described some variability in the arrangement, particularly for the postcloacal papillae. The second one is P. siamensis, with males that have 4 or 5 precloacal pairs of cloacal papillae and having a total of 17 or 19 cloacal papillae [13]. Both species clearly differ from P. canariensis n. sp. in the number of tooth-like prominences present in the submedian lobes of the pseudolabia (1 in P. mexicana, 4 in P. siamensis and 2 in P. canariensis n. sp.) and in the size of spicules, particularly the right spicule (340-465 µm in P. mexicana, 469-701µm in P. siamensis and 643-715 µm in P. canariensis n. sp.). The remaining species of Protospirura have a higher number of cloacal papillae, comprised between 18 and 25 papillae (see Table 1).
In females of P. canariensis n. sp., the vulva is located slightly before mid-body. Other Protospirura species, namely P. kaindiensis, P. okinavensis and P. pseudomuris also present a preequatorial placement of the vulva [7,8,9]. Females of the new species differ from those of these three species in the number of tooth-like prominences present in lateral and submedian lobes of pseudolabia (see Table 1).
Within the Protospirura genus, the life cycles of P. muricola and P. numidica criceticola have been elucidated [12,20,26]. Quentin et al. [12] described the life cycle of P. numidica criceticola with an L3 larva characterized by a notched posterior extremity having 12 to 15 pointed prominences. Posteriorly, Quentin [20] elucidated the life cycle of P. muricola and described a L3 larva presenting nine pointed prominences arranged in the same plan in its notched posterior extremity. In the present study, the single recovered L3 larva of P. canariensis n. sp. has 10 pointed prominences arranged circularly in the same plan. This morphological trait differs clearly from the two above-mentioned L3 larva of Protospirura [12,20] and is an additional feature differentiating these three species.
The parasitized host and the geographical distribution are additional criteria to distinguish P. canariensis n. sp. from the remaining species of Protospirura. There are seven Protospirura species parasites of Muridae rodents, namely P. armeniana, P. chabaudi, P. kaindiensis, P. munimuniensis, P. muricola, P. okinavensis and P. siamensis [6,8,9,13,14,15,16,18,19]. P. armeniana was found parasitizing the house mouse Mus musculus in Mongolia and Armenia [6,16,19]. Protospirura chabaudi was found parasitizing R. rattus in the Democratic Republic of Congo [18]. Both P. kaindiensis and P. munimuniensis were found in Papua New Guinea parasitizing Pseudohydromys murinus and Chiruromys lamia, respectively [9,15]. Protospirura okinavensis was reported in Japan parasitizing Mus caroli [8]. Protospirura muricola is the species with the widest geographical distribution parasitizing Muridae rodents, including numerous African and Asian countries and also with the largest host range, being recorded in diverse genera of murids such as Arvicanthis, Gerbilliscus, Hybomys, Lemniscomys, Malacomys, Mastomys, Mus, Praomys, Rattus and Uranomys [25,27,28,29,30,31]. Finally, P. siamensis was described in Thailand parasitizing several murids of genera Bandicota, Berylmys, Mus and Rattus [13], and more recently found in rats and mice from Pakistan [32]. However, these authors do not specify the species of murids parasitized by P. siamensis [32]. Thus, to our knowledge, apart from P. canariensis n. sp. presently recorded at the Canary Archipelago, only two species of Protospirura, namely P. chabaudi and P. muricola have been cited parasitizing the black rat R. rattus and reported in Democratic Republic of Congo, Nigeria, China and Taiwan [18,25,28,29].
The molecular results confirm the morphological observations showing that the specimens here described as a new species are clearly separated from the genus Mastophorus and cluster with P. muricola. The remaining species of the genus Protospirura should be reevaluated by integrating morphologic discriminant characters and molecular information.
In conclusion, the morphologic characteristics such as pseudolabia and the number of tooth-like prominences present in their lateral and submedian lobes, the pharynx, the particular characters of males (number and arrangement of cloacal papillae, and size of spicules) and females (position of the vulva), the molecular data, and the geographical distribution and parasitized host identify the discovered nematode as a new species of the genus Protospirura.
5. Conclusions
The present work contributes with the description of a new spirurid nematode of the genus Protospirura, P. canariensis n. sp. and provides further information on the taxonomic criteria useful to the characterization of species in this genus.
The most useful characteristics to differentiate P. canariensis n. sp. from the remaining species of the genus Protospirura are the number of tooth-like prominences present in lateral and submedian lobes of pseudolabia both in males and females, the size of spicules and the number and arrangement of cloacal papillae in male specimens, and the position of the vulva in female specimens. The host parasitized and the geographical distribution are additional and useful criteria. Thus, the current finding in El Hierro Island (Canary Archipelago, Spain) enlarges the geographical distribution of the genus Protospirura. After many years of trapping campaigns in all islands of the Canary Archipelago, P. canariensis n. sp. has been found only in El Hierro Island despite the existence of biotopes of similar biotic and abiotic characteristics in the other islands of the Archipelago. Unlike Tenerife and Gran Canaria islands, El Hierro is a small island without any commercial seaport, having only connections with the remaining islands through local planes and ferries. This may be the most feasible cause of the presence of the new species only in El Hierro. Finally, the sequence of the subunit I gene of the cytochrome c oxidase (cox1) also characterizes and differentiates P. canariensis n. sp. from P. muricola, the species with the widest geographical distribution and one of the two Protospirura species found parasitizing the black rat.
Author Contributions
Conceptualization, J.M., C.F. and P.F.; Methodology, J.M., N.M.-C., A.R. and P.F.; Investigation, J.M., N.M.-C., A.R., S.S.-V., C.F. and P.F.; Resources, J.M., N.M.-C. and P.F.; Writing—Original Draft Preparation, J.M., N.M.-C. and P.F.; Writing—Review & Editing, J.M., C.F. and P.F.; Supervision, J.M., C.F. and P.F.; Project Administration, C.F. and P.F.; Funding Acquisition, P.F. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by ProID2021010013 from "Consejería de Economía, Industria, Comercio y Conocimiento, Gobierno de Canarias", "Fondos FEDER-FSE Canarias 2014-2020", "Programa de Apoyo a la Investigación María del Carmen Betencourt y Molina" and "Gobierno de Canarias-ULL" agreement ("Estudio de patógenos en aves migratorias y en especies exóticas en un escenario de cambio climático").
Institutional Review Board Statement
Animal work was approved in accordance with the Spanish Government Laws 42/2007 and RD 630/2013, the Canary Government law 151/2001 (references FYF141/10, FYF205/09, EEI-007/2019, ADL/mjb, MRR/rsh, A/EST-030/2016, AFF115/16 and EEI-007/2019) and the Ethic Committees of Research and Animal Wellness of "Universidad de La Laguna" (CEIBA2018-0330).
Data Availability Statement
The type specimens are available upon request from MNHN Paris. Additional specimens are available upon request from the corresponding author.
Acknowledgments
The authors wish to thank the staff of the "Centres Científics i Tecnològics de la Universitat de Barcelona (CCiTUB)" for their assistance in the preparation of SEM samples. A.R. and J.M. are members of the 2021-SGR-00359 research group.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Chitwood, B.G. The status of Protospirura vs. Mastophorus with a consideration of the species of these genera. In Livro Jubilar do Professor Lauro Travassos; Silva, B., de Almeida, B.J., Ferreira, N., Gonçalves, A., Strommer, S., Gonçalves, O., de Almeida, C.A., Cantizano dos Santos, A., Eds.; Typographia do Instituto Oswaldo Cruz: Rio de Janeiro, Brazil, 1938; pp. 115–118. [Google Scholar]
- Wertheim, G. A study of Mastophorus muris (Gmelin, 1790) (Nematoda: Spirurida). Trans. Am. Microsc. Soc. 1962, 81, 274–279. [Google Scholar] [CrossRef]
- Babero, B.B.; Matthias, D. Protospirura peromysci n. sp. (Nematoda: Spiruridea) and other helminths from Peromyscus spp. in Nevada. Proc. Helminthol Soc. Wash. 1967, 34, 255–261. [Google Scholar]
- Falcón Ordaz, J. , Sanabria Espinoza, M.A. Especie nueva del género Protospirura (Nemata: Spiruridae) de Peromyscus difficilis (Rodentia: Cricetidae) de Hidalgo, México. An. Inst. Biol., UNAM, Ser. Zool. 1995, 66, 17–26. [Google Scholar]
- Seurat, L.-G. Sur un nouveau spiroptère du Chat ganté. C. R. Soc. Biol., Paris 1914, 77, 344–347. [Google Scholar]
- Skryabin, K.I.; Sobolev, A.A. Spirurata of Animals and Man and the Diseases Caused by Them. Spirurata, Part 5, Spiruroidea. In Osnovy Nematologii; Skryabin, K.I., Ed.; Nauka: Moscow, Russia, 1963; Vol. 11. (In Russian) [Google Scholar]
- Yokohata, Y.; Abe, H. Two new Spirurid Nematodes in Japanese Moles, Mogera spp. Jpn. J. Parasitol. 1989, 38, 92–99. [Google Scholar]
- Hasegawa, H. Protospirum okinavensis sp. n. (Nematoda: Spiruridae) from Mus caroli on Okinawa Island, Japan. J. Helminthol. Soc. Wash. 1990, 57, 153–156. [Google Scholar]
- Smales, L.R. Protospirura kaindiensis n. sp. (Spirura: Spiruridae) and other helminths from Pseudohydromys (Muridae: Hydromyinae) from Papua New Guinea. J. Parasitol. 2001, 87, 169–172. [Google Scholar] [CrossRef]
- Kreis, H.A. Beitrage zur Kenntnis parasitischer nematoden. VII. Parasitische Nematoden der schweizerischen wissenschaftlichen Expedition nach Angola (Afrika) in Jahre 1932. Ctbl. Bakt. I. Orig. 1938, 142, 90–105. [Google Scholar]
- Pence, D.B.; Windberg, L.A. Population dynamics across selected habitat variables of the helminth community in coyotes, Canis latrans, from South Texas. J. Parasitol. 1984, 70, 735–746. [Google Scholar] [CrossRef]
- Quentin, J.-C.; Karimi, Y.; Rodríguez de Almeida, C. Protospirura numidica criceticola, n. subsp. parasite de Rongeurs Cricetidae du Brésil. Cycle évolutif. Ann. Parasitol. (Paris) 1968, 43, 583–596. [Google Scholar] [CrossRef]
- Ribas, A.; Veciana, M.; Chaisiri, K.; Morand, S. Protospirura siamensis n. sp. (Nematoda: Spiruridae) from rodents in Thailand. Syst. Parasitol. 2012, 82, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Smales, L.R.; Harris, P.D.; Behnke, J.M. A redescription of Protospirura muricola Gedoelst, 1916 (Nematoda: Spiruridae), a parasite of murid rodents. Syst. Parasitol. 2009, 72, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Smales, L.R. The gastrointestinal nematodes of Chiruromys forbsei Thomas and C. lamia (Thomas) (Rodentia: Muridae) with the description of a new species of Helgenema (Heligmonellidae) and a new species of Protospirura (Spiruridae) from Papua New Guinea. Trans. R. Soc. S. Aust. 2021, 145, 60–76. [Google Scholar] [CrossRef]
- Tinnin, D.S.; Ganzorig, S.; Gardner, S.L. Helminths of small mammals (Erinaceomorpha, Soricomorpha, Chiroptera, Rodentia, and Lagomorpha) of Mongolia. Fac. Publ. Harold W. Manter Lab. Parasitol., Special Publ., Mus. Texas Tech. Univ. 2011, 59, 1–50. [Google Scholar]
- Chabaud, A.G. No. 3 Keys to genera of the order Spirurida. Part 2. Spiruroidea, Habronematoidea and Acuarioidea. In CIH Keys to the Nematode Parasites of Vertebrates; Anderson, R.C., Chabaud, A.G., Willmott, S., Eds.; CAB: Farnham Royal, Bucks, UK, 1975; pp. 29–58. [Google Scholar]
- Vuylsteke, C. Mission de Zoologie médicale au Maniema (Congo, Léopoldville) (P.L.G. Bennoit, 1959). 3. Vermes–Nematoda. Ann. Mus. R. Afr. Centr., Ser. 8 Zool. 1964, 132, 41–66. [Google Scholar]
- Movsesyan, S.O.; Nikoghosian, M.A.; Petrosian, R.A.; Vlasov, E.A.; Kuznetsov, D.N. Nematodes of rodents of Armenia. Ann. Parasitol. 2018, 64, 173–180. [Google Scholar] [PubMed]
- Quentin, J.-C. Cycle biologique de Protospirura muricola Gedoelst, 1916 Nematoda Spiruridae. Ann. Parasitol. (Paris) 1969, 44, 485–504. [Google Scholar] [CrossRef]
- López, C.; Clemente, S.; Almeida, C.; Brito, A.; Hernández, M. A genetic approach to the origin of Millepora sp. in the eastern Atlantic. Coral Reefs 2015, 34, 631–638. [Google Scholar] [CrossRef]
- Gaillard, C.M.; Pion, S.D.; Hamou, H.; Sirima, C.; Bizet, C.; Lemarcis, T.; Locatelli, S. Detection of DNA of filariae closely related to Mansonella perstans in faecal samples from wild non-human primates from Cameroon and Gabon. Parasites Vectors 2020, 13, 1–13. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Baylis, H.A. On a collection of nematodes from Nigerian mammals (chiefly rodents). Parasitology 1928, 20, 280–304. [Google Scholar] [CrossRef]
- Anderson, R.C. Nematode Parasites of Vertebrates. Their Development and Transmission, 2nd ed.; CABI Publishing: Wallingford, Oxon, UK, 2000; pp. 413–427. [Google Scholar]
- Brouat, C.; Kane, M.; Diouf, M.; Bâ, K.; Sall-Dramé, R.; Duplantier, J.M. Host ecology and variation in helminth community structure in Mastomys rodents from Senegal. Parasitology 2007, 134, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.T. A preliminary report on a survey of animal parasites of Canton, China, rats. Lignan Sci. J. (Canton) 1933, 12, 65–74. [Google Scholar]
- Hasegawa, H.; Kobayashi, J.; Otsuru, M. Helminth parasites collected from Rattus rattus on Lanyu, Taiwan. J. Helminthol. Soc. Wash. 1994, 61, 95–102. [Google Scholar]
- Smales, L.R. The gastrointestinal helminths of Rattus niobe (Rodentia: Muridae) with descriptions of two new genera and three new species (Nematoda) from Papua New Guinea and Papua Indonesia. Zootaxa 2016, 4117, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Temme, M. Stomach nematodes of the Polynesian rat Rattus exulans in the Northern Marshall Islands, Pacific Ocean. Z. Angew. Zool. 1983, 70, 463–472. [Google Scholar]
- Rajper, M.; Birmani, N.A. New record of Protospirura siamensis Ribas, 2012 (Nematoda: Spiruridae) recovered from Rat and Mice of district Hyderabad, Sindh, Pakistan. In Proceedings of the 40th Pakistan Congress of Zoology, virtual, 14-19 December 2021; p. 74. [Google Scholar]
Figure 1.
Localities where Rattus rattus parasitized by Protospirura canariensis n. sp. were captured on El Hierro Island (Canary Archipelago, Spain). (1) Camino-Frontera (27° 44' 38.07" N, 18° 2' 25.4" W); (2) Lagartario-Frontera (type locality) (27° 46' 29.9" N, 17° 59' 55.59" W); (3) Túnel-Valverde (27° 49' 12.41" N, 17° 57' 49.27" W).
Figure 1.
Localities where Rattus rattus parasitized by Protospirura canariensis n. sp. were captured on El Hierro Island (Canary Archipelago, Spain). (1) Camino-Frontera (27° 44' 38.07" N, 18° 2' 25.4" W); (2) Lagartario-Frontera (type locality) (27° 46' 29.9" N, 17° 59' 55.59" W); (3) Túnel-Valverde (27° 49' 12.41" N, 17° 57' 49.27" W).
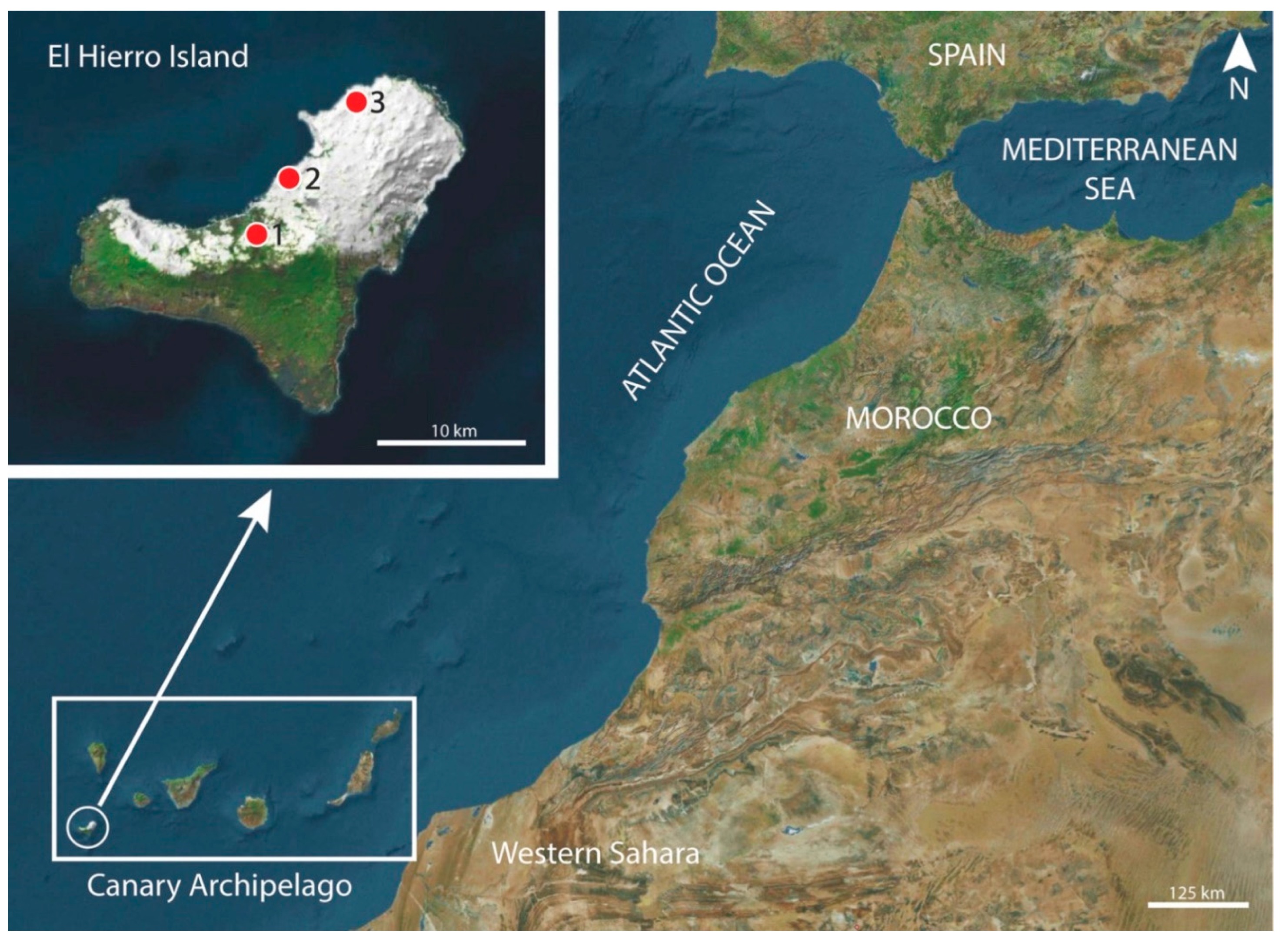
Figure 2.
Phylogenetic analysis using the Maximum Likelihood method with p-distance and 1,000 bootstrap replications based on the mitochondrial cytochrome c oxidase subunit I gene (cox1) sequence. Sequences obtained in the present study are shown in bold. Schistosoma mansoni was used as the outgroup.
Figure 2.
Phylogenetic analysis using the Maximum Likelihood method with p-distance and 1,000 bootstrap replications based on the mitochondrial cytochrome c oxidase subunit I gene (cox1) sequence. Sequences obtained in the present study are shown in bold. Schistosoma mansoni was used as the outgroup.
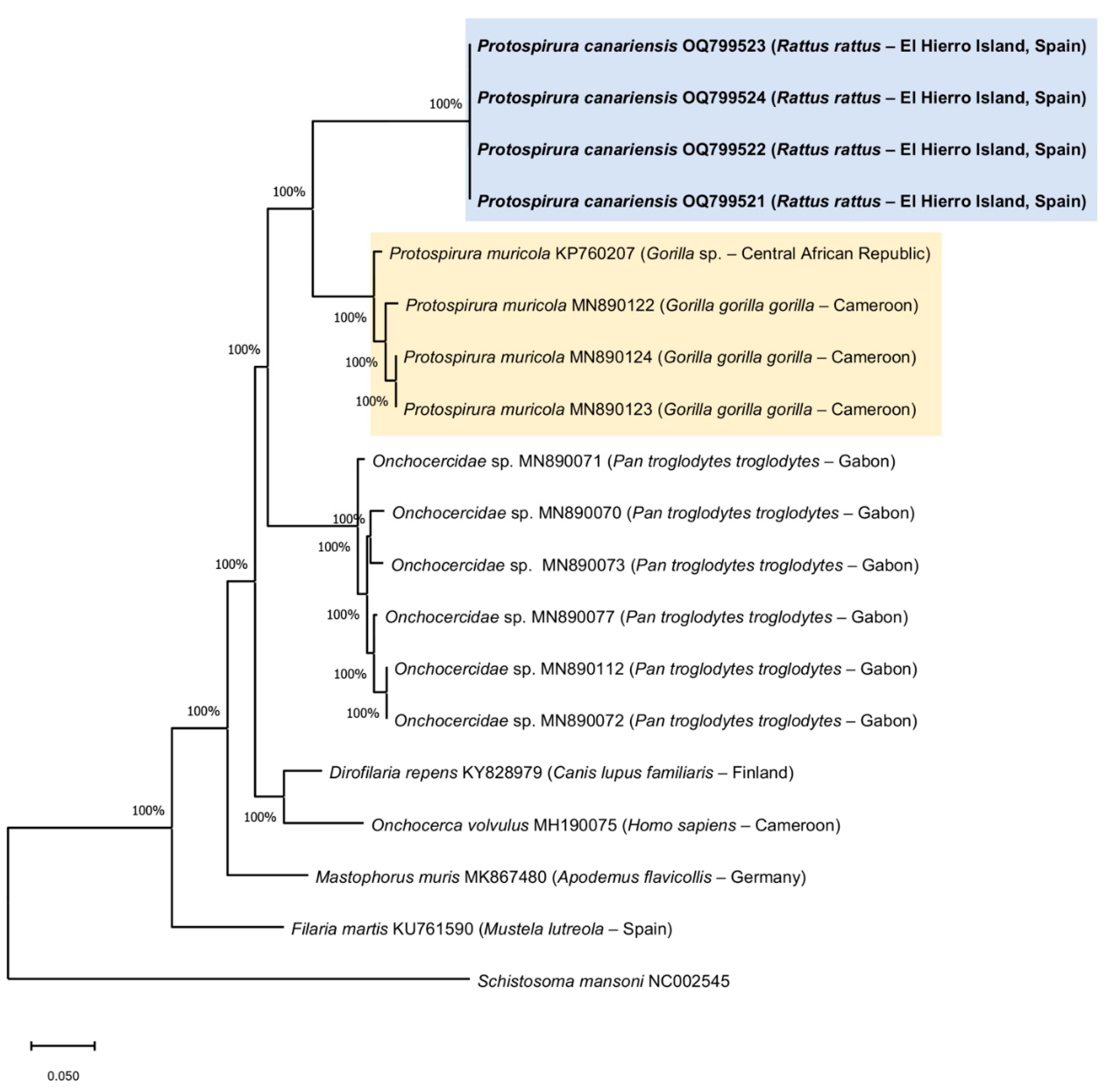
Figure 3.
Protospirura canariensis n. sp., male and female. A Cephalic extremity of female, lateral view. B Detail of submedian and lateral lobes of one of the pseudolabia of female, lateral view. C Caudal extremity of male showing the two spicules and the cloacal papillae, lateral view. D Tail of female, lateral view. E V-shaped gubernaculum. F Embryonated egg.
Figure 3.
Protospirura canariensis n. sp., male and female. A Cephalic extremity of female, lateral view. B Detail of submedian and lateral lobes of one of the pseudolabia of female, lateral view. C Caudal extremity of male showing the two spicules and the cloacal papillae, lateral view. D Tail of female, lateral view. E V-shaped gubernaculum. F Embryonated egg.
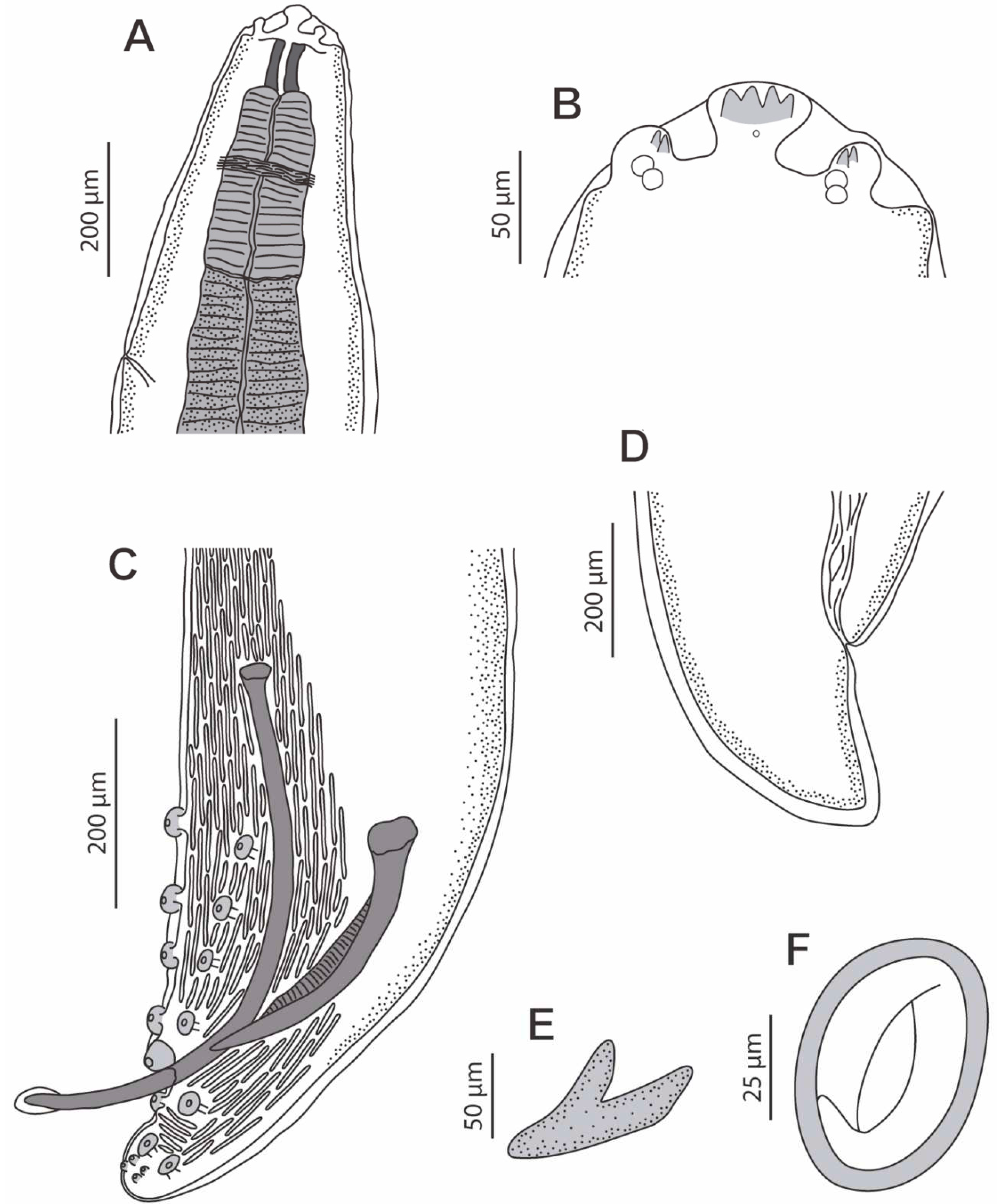
Figure 4.
Protospirura canariensis n. sp., male, LM. A Caudal extremity showing the four pairs of precloacal papillae (1 to 4). B Caudal extremity showing the unpaired precloacal papilla (up). C Caudal extremity showing the four pairs of postcloacal papillae (5 to 8). D Detail of right (rSp) and left spicule (lSp).
Figure 4.
Protospirura canariensis n. sp., male, LM. A Caudal extremity showing the four pairs of precloacal papillae (1 to 4). B Caudal extremity showing the unpaired precloacal papilla (up). C Caudal extremity showing the four pairs of postcloacal papillae (5 to 8). D Detail of right (rSp) and left spicule (lSp).
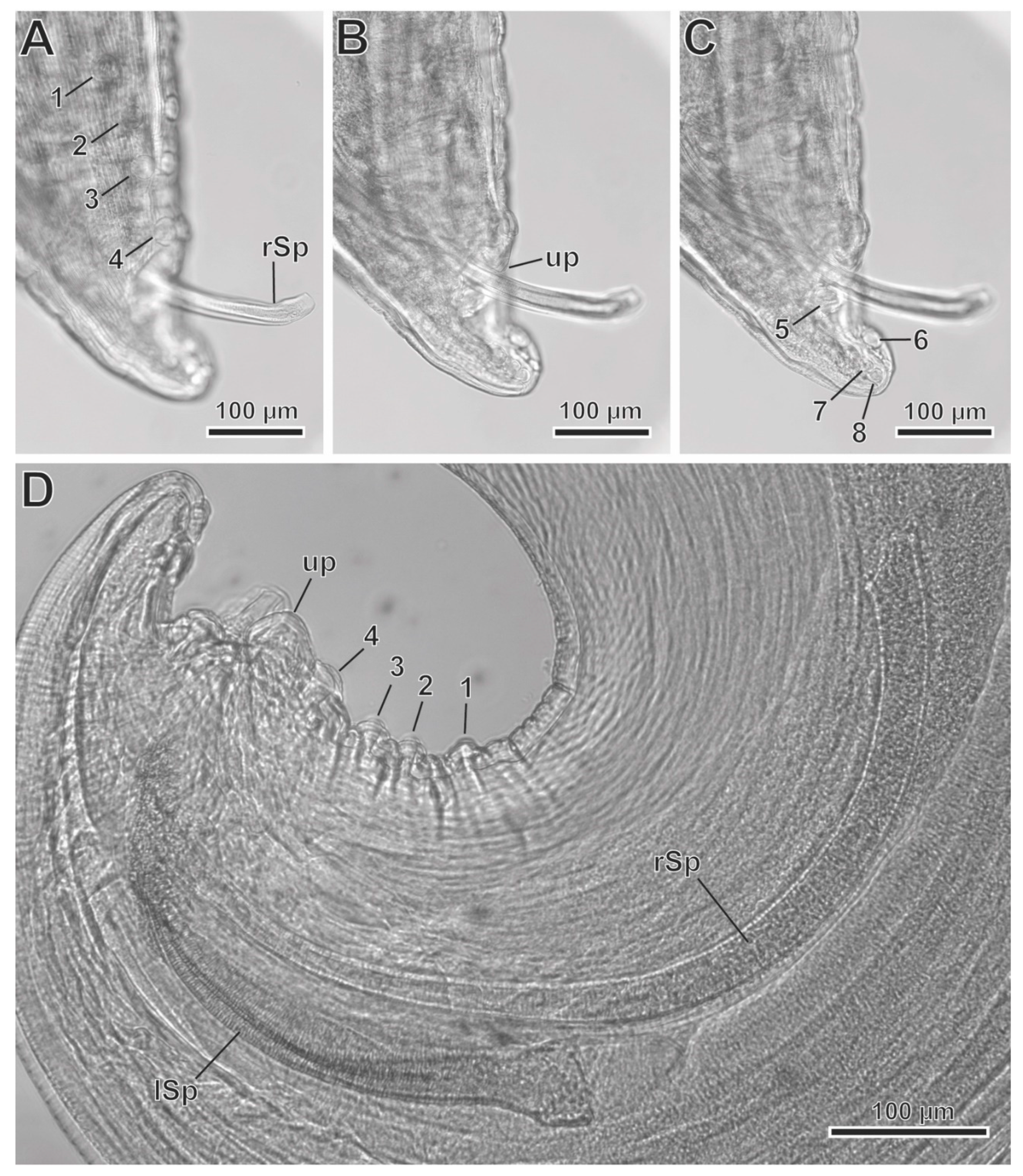
Figure 5.
Protospirura canariensis n. sp., male, SEM. A Apical view of the cephalic extremity showing the two tri-lobed pseudolabia. B Caudal extremity illustrating the distribution of cloacal papillae. C Postcloacal pairs of papillae. D Detail of right spicule (rSp). E Detail of left spicule (lSp). (arrows) tooth-like prominences; (*) submedian lobes; (**) lateral lobes; (1–4) pairs of precloacal papillae; (5–8) pairs of postcloacal papillae; (am) amphid; (cp) cephalic papillae; (ph) pharynx; (up) unpaired precloacal papilla.
Figure 5.
Protospirura canariensis n. sp., male, SEM. A Apical view of the cephalic extremity showing the two tri-lobed pseudolabia. B Caudal extremity illustrating the distribution of cloacal papillae. C Postcloacal pairs of papillae. D Detail of right spicule (rSp). E Detail of left spicule (lSp). (arrows) tooth-like prominences; (*) submedian lobes; (**) lateral lobes; (1–4) pairs of precloacal papillae; (5–8) pairs of postcloacal papillae; (am) amphid; (cp) cephalic papillae; (ph) pharynx; (up) unpaired precloacal papilla.
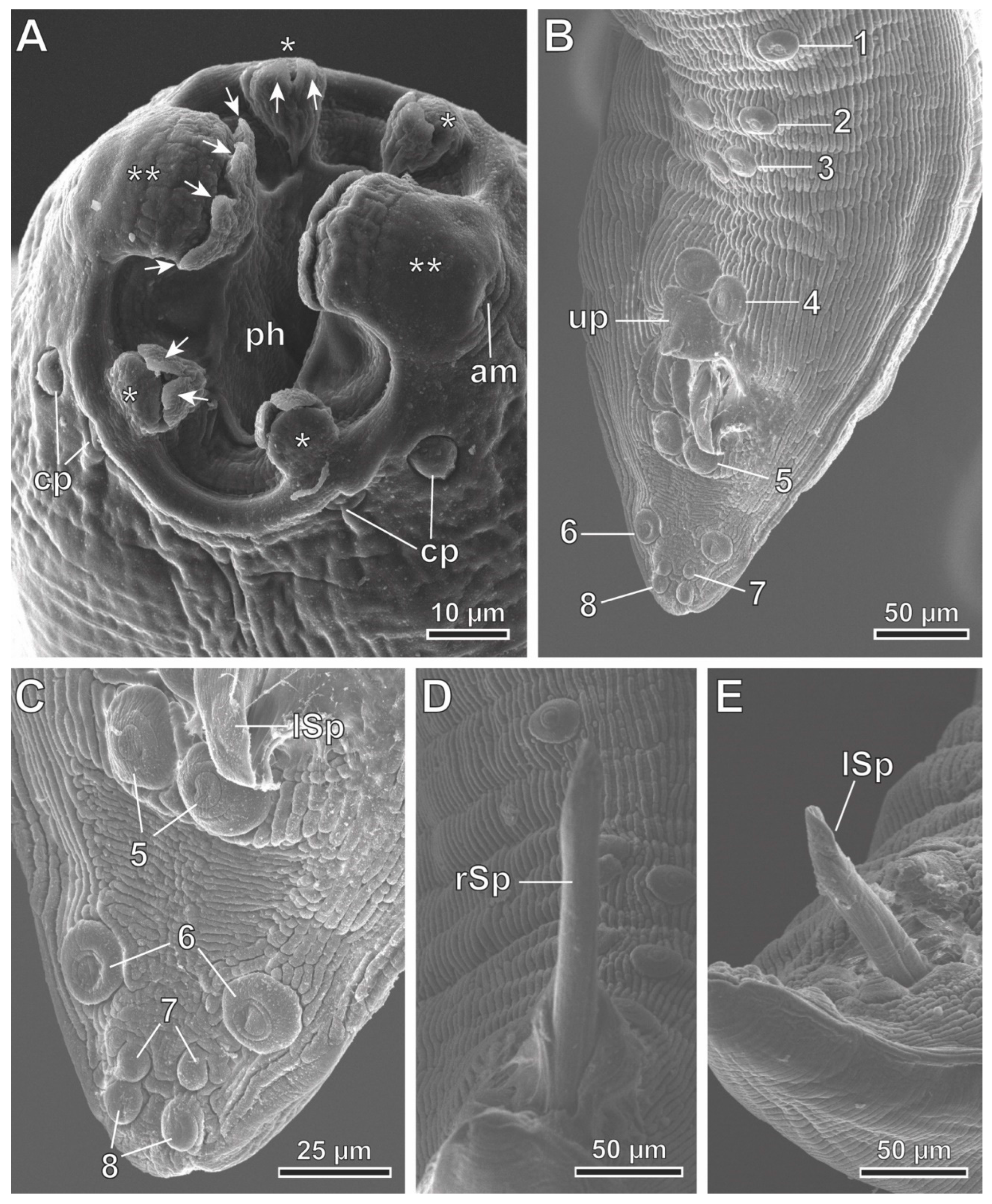
Figure 6.
Protospirura canariensis n. sp., female, SEM. A Lateroventral view of the cephalic extremity showing the right deirid (d) and the excretory pore (ep). B Apical view of the cephalic extremity showing the two tri-lobed pseudolabia. C Detail of one of the pseudolabia showing tooth-like prominences (arrows). (*) submedian lobes; (**) lateral lobes; (am) amphid; (cp) cephalic papillae; (ph) pharynx.
Figure 6.
Protospirura canariensis n. sp., female, SEM. A Lateroventral view of the cephalic extremity showing the right deirid (d) and the excretory pore (ep). B Apical view of the cephalic extremity showing the two tri-lobed pseudolabia. C Detail of one of the pseudolabia showing tooth-like prominences (arrows). (*) submedian lobes; (**) lateral lobes; (am) amphid; (cp) cephalic papillae; (ph) pharynx.
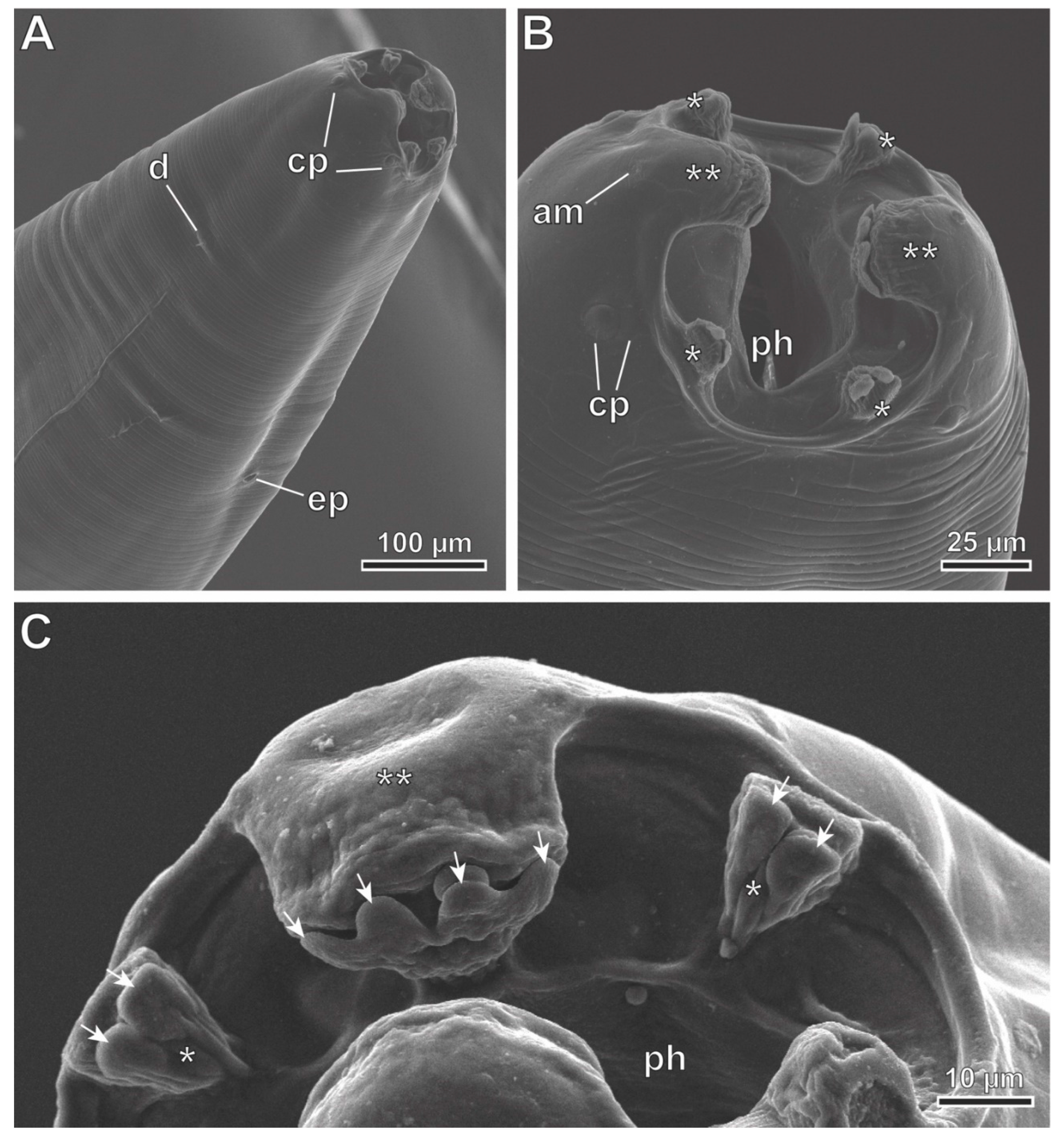
Figure 7.
Protospirura canariensis n. sp., L3 larva, LM. A Cephalic extremity showing the entire oesophagus (o). B Detail of cephalic extremity showing the pharynx (ph). C Detail of the notched posterior extremity showing caudal pointed prominences (arrowheads). (arrow) posterior extremity of the oesophagus; (a) anus.
Figure 7.
Protospirura canariensis n. sp., L3 larva, LM. A Cephalic extremity showing the entire oesophagus (o). B Detail of cephalic extremity showing the pharynx (ph). C Detail of the notched posterior extremity showing caudal pointed prominences (arrowheads). (arrow) posterior extremity of the oesophagus; (a) anus.
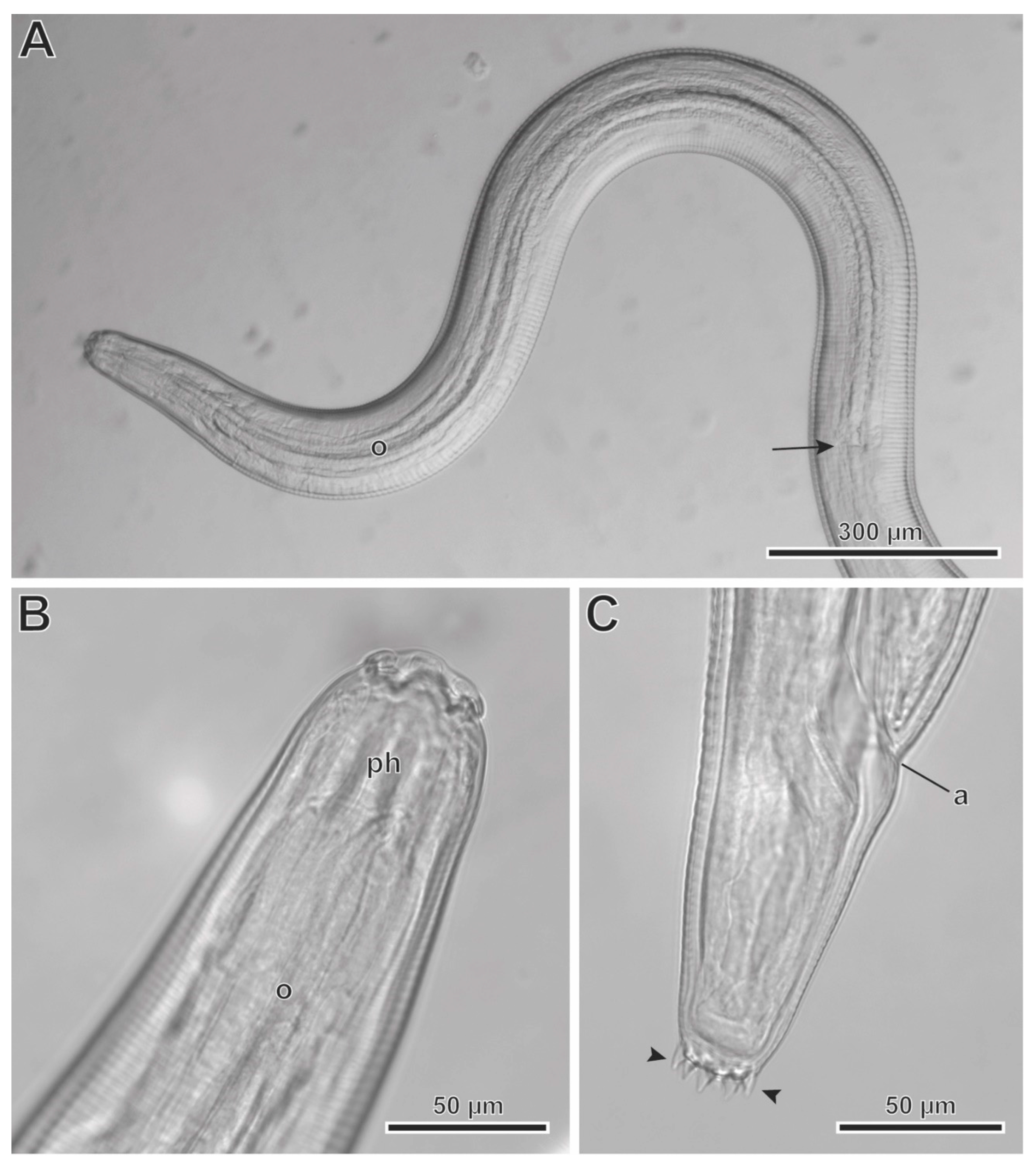

Table 1.
Some differential characteristics of Protospirura species.
| Species | Pseudolabia | Spicules | Cloacal papillae | Vulva | References | ||
|---|---|---|---|---|---|---|---|
| subm | lat | right | left | ||||
| Protospirura anopla | 01 42 |
01 42 |
287 | 632 | 4 pre, 5 post (18) |
posteq | [6,9] |
| Protospirura armeniana | 21 42 |
21 42 |
620–639 | 370–411 | 4 pre, upre, 7 post, upost (24) |
posteq | [6,9] |
| Protospirura canariensis n. sp. | 2 | 4 | 643–715 (675) |
309–412 (368) |
4 pre, upre, 4 post (17) |
preeq | Present study |
| Protospirura chabaudi | 23 42 |
03 42 |
980 | 420 | 4 pre, 5 post (18) |
posteq | [9,18] |
| Protospirura kaindiensis | 2 | 2 | 450–480 | 310–330 | 4 pre, upre, 5 post (19) |
preeq | [9] |
| Protospirura mexicana | 1 | 0 | 340–465 (404) |
420–527 (481) |
4 pre, upre, 4 post (17) |
posteq | [4] |
| Protospirura munimuniensis | 2 | 2 | 602–603 | 430–455 | 4 pre, upre, 6 post (21) |
? | [15] |
| Protospirura muricola | 2 | 2 | 268–430 (352) |
290–501 (411) |
4 pre, upre, 6 post (21) |
posteq | [14,25] |
| Protospirura numidica | 34 42 |
34 45 42 |
830 | 420 | 4 pre, upre, 5 post (19) |
posteq | [5,9,12] |
| Protospirura numidica criceticola | ? | 4 | 1250 | 470 | 4 pre, upre, 7 post (23) |
posteq | [12] |
| Protospirura okinavensis | 4 | 4 | 600–650 (620) |
320–350 (320) |
5–6 pre, upre, 4 post (19 or 21) |
preeq | [8] |
| Protospirura peromysci | 26 42 |
46 42 |
820–1200 | 330–380 | 4 pre, upre, 7 post (23) |
posteq | [3,9] |
| Protospirura pseudomuris | 17 22 |
17 22 |
540–790 (680) |
270–380 (330) |
4 pre, upre, 6 post (21) |
preeq | [7,8,9] |
| Protospirura siamensis | 4 | 4 | 469–701 (572.6) |
348–380 (368.7) |
4–5 pre, upre, 4 post (17 or 19) |
posteq | [13] |
| Protospirura suslica | 21 42 |
21 42 |
315 | 873 | 5 pre, upre, 6 post (23) |
posteq | [6,9] |
Mean size of spicules in parentheses. Total number of cloacal papillae in parentheses. 1-7Characters as described by several authors: 1Skryabin and Sobolev [6], 2Smales [9], 3Vuylsteke [18], 4Seurat [5], 5Quentin et al. [12], 6Babero and Matthias [3], 7Yokohata and Abe [7]. (lat) tooth-like prominences in lateral lobes; (post) postcloacal pairs of papillae; (posteq) postequatorial; (pre) precloacal pairs of papillae; (preeq) preequatorial; (subm) tooth-like prominences in submedian lobes; (upost) unpaired postcloacal papilla; (upre) unpaired precloacal papilla; (?) doubtful or unknown data.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



