Submitted:
19 April 2023
Posted:
20 April 2023
You are already at the latest version
Abstract
Cytokines and chemokines (chemotactic cytokines) are soluble extracellular proteins that bind to specific receptors and play an integral role in the cell-to-cell signaling network, in addition can promote the homing of cancer cells into different organs. We investigated the potential relationship between human hepatic sinusoidal endothelial cells (HHSECs) and several melanoma cell lines for the expression of chemokine and cytokine ligand and receptor expression during invasion of melanoma cells. In order to define the invasion related gene expression differences, we selected invasive and non-invasive subpopulations of cells after co-culturing with HHSECs. In addition, we determined protein expression of the endothelial cells before and after co-culturing with melanoma cell lines originated from primary tumors. Cell lines with increased invasive capacity after culturing with conditioned medium showed a set of receptor genes (CXCR1, IL1RL1, IL1RN, IL3RA, IL8RA, IL11RA, IL15RA, IL17RC, and IL17RD) with significantly different expression. It is very important note, that the IL11RA gene expression level was also significantly increased in primary melanoma tissues with liver metastasis as well. Proteome arrays revealed 15 differentially expressed proteins (including CD31, VCAM-1, ANGPT2, CXCL8, and CCL20) in the hepatic endothelial cells after co-cultured with melanoma cells. Our findings clearly indicate the interaction between liver endothelial- and melanoma cells. Based on our data, we assume that overexpression of the IL11RA gene might has key role in the formation of organ specific metastasis to the liver by primary melanoma cells.
Keywords:
malignant melanoma
; liver metastasis
; cytokines
; IL11RA
; hepatic endothelial cells
1. Introduction
Efficient communication between cells is one of the most important aspects of cell condition in multicellular organisms [1]. Metastasis formation of primary tumors is also a balance of the host (‘soil’) and the tumor (‘seed’) cellular interactions, an altered cross-talk between the cells, which mechanism is determined by both the intrinsic properties of the tumor cells and the host response [2,3,4]. According to the ‘seed and soil hypothesis’, the primary tumor contributes to the preparation of secondary sites for tumor cell invasion and development, known as the pre-metastatic niche (PMN) [5,6]. The distribution of the targeted organ is especially variable depending on the cancer type, however, melanoma has the unique property of having a fully comprehensive metastatic potential, hence any organ or tissue can host melanoma metastasis [7]. Metastasis is the main factor limiting survival in most patients with cancer. Therefore, understanding the molecular mechanisms that control the metastatic behavior of tumor cells is an important key to the successful treatment of cancer.
Cytokines and chemokines (chemotactic cytokines) are soluble extracellular proteins that are an integral part of the cell-to-cell signaling network and can bind to specific receptors to control cellular growth, development, hematopoiesis, lymphocyte recruitment, inflammation and immune regulation [8]. There are numerous observations that endothelial cells coating the blood vessels of various organs express a number of different cytokines, chemokines and adhesion molecules that bind to specific receptors on the cell surface to promote the homing of cancer cells [9,10,11]. In this way, the site of secondary tumor formation depends on the chemokine receptors expressed on the tumor cell surface and the chemokine expression specific to each tissue. The use of cytokines in the clinic as monotherapy due to their need for high and frequent dosing is often resulted in toxicity [12]. Combinations of cytokines with various checkpoint inhibitors may provide promising aspects for tumor treatment including melanoma [13].
The most common sites of distant metastases in melanoma patients include skin, liver, lung, and brain; however, the initial sites of melanoma cell spread are typically lymph nodes [7,14]. Liver metastases are observed in 10-20% of patients with cutaneous melanoma, which occur relatively late during disease progression with an average 2-4 months survival [14,15,16]. The liver has a rich blood supply and therefore provides fertile "soil" for metastatic spread [17]. Liver sinusoidal endothelial cells (LSECs) constitute approximately 50% of the non-parenchymal cells of the liver and form the fenestrated wall of the hepatic sinusoids with minimal basement membrane [18,19]. LSECs are the first cells to contact the blood flow in hepatic sinusoids and they have several pro-metastatic properties [20]. LSECs express numerous cellular adhesion molecules including ICAM-1, VCAM-1, endothelial (E)-selectin, and CD31, which can facilitate cancer cell migration, and promote the tumor cells adherence to the LSECs, extravasation and metastasis formation [21]. Mendt et al. have described the possible role of the CXCL12/CXCR4 axis in liver metastasis of melanoma, however, little is known about the specific factors that regulate the growth of liver metastases [22]. Importantly, different cytokines and chemokines are also expressed by the LSECs, which play an important role in recruiting distinct leukocytes to contribute to the immune microenvironment within the liver [20,23].
Different chemokine and cytokine secretion by endothelial cells may have different effects on the invasion properties of melanoma cells expressing specific receptors. Therefore, we aimed to investigate the potential relation between human hepatic sinusoidal endothelial cells (HHSECs) and several different melanoma cell lines in terms of chemokine and cytokine ligand and receptor expression during in vitro invasion of melanoma cells. We hypothesize that the study of the ligand/receptor axis expressed during melanoma cell invasion targeting hepatic endothelial cells may provide new insights into the understanding of the mechanism of liver-specific metastasis of melanoma.
2. Results
2.1. Effect of HHSEC on melanoma cell invasion
To investigate the impact of HHSEC cells on melanoma cell invasion, invasion assay was implemented using human hepatic sinus endothelial cells conditioned media (HHSEC-CM) as chemoattractant or unconditioned media containing 10% FBS as control (Supplementary Table S1). We observed that all cell lines showed invasive property, however, significantly increased cell invasion was detected in three cases (WM983A, WM3248, and WM1360) after co-culturing melanoma cells with HHSEC-CM compared to culturing in unconditioned medium (Figure 1). Cell lines (WM1366, WM278 and WM793B) that did not change their invasive properties regardless of the medium (conditioned or not) were referred as stable invasive.
2.2. Chemokine- and cytokine receptor expression of melanoma cells
In order to define the gene expression differences related to melanoma cell invasion, we selected the invasive and the non-invasive cells after co-culturing with HHSEC in case of every cell lines. In this way, we were able to establish selected invasive cell populations (WM1366-HINV, WM278-HINV, WM793B-HINV, WM983A-HINV, WM1361-HINV, and WM3248-HINV; H = hepatic, INV = invasive), and selected non-invasive cell populations (WM1366-HNON-INV, WM278-HNON-INV, WM793B-HNON-INV, WM983A-HNON-INV, WM1361-HNON-INV, and WM3248-HNON-INV; H = hepatic, NON-INV = non-invasive). After the selection, we determined the gene expression patterns of six invasive and six non-invasive cell populations (Supplementary Table S2).
To investigate the gene-expression differences between the invasive and non-invasive cells triggered by the hepatic endothelial cells, we analyzed the expression changes in the two main groups generated according to the response to the HHSEC conditioned medium. Melanoma cell lines with stable invasive properties exhibited a small cohort of genes (XCR1, IL1RN, IL11RA, TNFRSF4, TNFRSF12A, and TNFRSF13C) that were significantly altered in the separated invasive cells compared to the non-invasive ones (Figure 2Aa). The same comparison was made for cell lines that responded to the conditioned medium with increased invasion, and a different group of altered genes (CXCR1, IL1RL1, IL1RN, IL3RA, IL8RA, IL11RA, IL15RA, IL17RC, and IL17RD) were found in the separated invasive cells (Figure 2Ab and 2C). In the union of the two groups there was a single gene: IL11RA, which expression was significantly higher in invasive cells than in the non-invasive cells in both comparisons; however, the difference was relatively low in cell lines with stable invasion, and much more remarkable in cell lines that showed increased invasiveness after culturing with HHSEC conditioned medium (Figure 2B).
2.3. Correlation between gene expression and invasiveness
We analyzed the correlation between invasive potential and altered expression of the receptor genes resulting from co-cultivation with HHSEC-CM. We correlated the invasion changes with the ratio of the gene expression data between invasive and non-invasive populations after culturing the cells with HHSEC-CM. Significantly correlated genes included: four chemokine receptor-, two tumor necrosis factor receptor- and seven interleukin receptor genes (Supplementary Table S2). Interestingly, we found significant positive correlation between the expression of IL15RA, IL17RC, and IL17RD genes and the invasiveness of cell lines (R = 0.933, p = 0.007; R = 0.872, p = 0.024; R = 0.938, p = 0.006, respectively, Figure 3), those three receptor genes displayed significantly increased expression in the selected invasive melanoma cells after culturing with HHSEC-CM as well.
2.4. Proteome profile of HHSECs
Proteome arrays (Proteome Profiler Human Chemokine Array Kit and Proteome Profiler Human XL Cytokine Array Kit) were used to detect protein expression differences between HHSEC before (HHSEC-Control) and after co-culturing with different melanoma derived primary cell lines (HHSEC+WM1366, HHSEC+WM278, HHSEC+WM793B, HHSEC+WM983A, HHSEC+WM1361, and HHSEC+WM3248). Proteins with detectable differences (>10%) in at least one cell line revealed 15 differentially expressed proteins (including CD31, VCAM-1, ANGPT2, CXCL8, and CCL20) in the co-cultured HHSECs compared to the control (Figure 4). Furthermore, correlation analysis revealed that CCL20 protein expression was significantly correlated with the invasive capacity of melanoma cells after the co-culturing (R = 0.918, p = 0.01).
2.5. Gene expression of chemokine- and cytokine receptors in melanoma tissue samples
We were able to analyze the gene expression differences of chemokine- and cytokine receptor genes for a small cohort of primary melanoma tissue samples (non-metastatic, N=4; primary melanoma with liver metastasis, N=5). We examined the expression of genes (CXCR1, IL1RL1, IL1RN, IL3RA, IL8RA, IL11RA, IL15RA, IL17RC, and IL17RD) that showed significant differences between invasive and non-invasive cells in the cell lines with increased invasiveness after culturing with HHSEC-CM. According to our qRT-PCR data, the relative expression level of the IL11RA gene was significantly higher in primary melanoma tissues with liver metastasis compared to primary melanoma without metastasis (Figure 5A). On the hand, we analyzed the association of the gene expression of these receptor genes with the Breslow thickness (mm) of primary melanoma tissues. This analysis, revealed a significant positive correlation for IL17RD gene expression (R = 0.777, p = 0.014; Figure 5B).
3. Discussion
The mechanisms of melanoma metastasis have been the subject of extensive studies for decades, as melanoma has one of the highest metastatic potential among human cancers with diverse secondary tumor sites including liver [14,25,26]. However, most of the recent studies are focusing on the most common primary cancers that metastasizes to the liver, including colorectal- and pancreatic cancers, as well as uveal melanoma [15,27,28,29]. The organotropism of the liver is influenced by different factors such as blood flow pattern, tumor stage and histological subtype of the tumor [17]. On the other hand, it is well established that targeted organs of the future metastasis are not passively participate in this process, but modified by the primary tumor via intercellular communication [5,30]. Several studies have shown that a wide range of ligands and receptors including integrins, selectins, adherins, and immunoglobulin superfamily receptors contribute to heterotypic adhesion processes between tumor cells and endothelial cells [31,32,33]. In a model of liver metastasis it was shown that CXCR4 chemokine receptor on tumor cells and its ligand CXCL12 expressed by endothelial cells significantly promote tumor cell transendothelial migration [34]. Mendt et el. also highlighted the role of CXCR4/CXCL12 in vitro migration, invasion, proliferation and adhesion of melanoma cells during liver metastasis [22].
We aimed to investigate the possible interaction and communication between human hepatic sinusoidal endothelial cells (HHSECs) and melanoma cells, and analyze the effect of HHSEC on the invasion of the different primary tumor originated melanoma cell lines. As it was expected, not all of the cell lines had the same response during co-culturing with HHSEC, three cell lines showed significantly increased invasive potential compared to the invasiveness before the co-culture. After co-culturing, we investigated the gene expression differences between the invasive and non-invasive melanoma cells, and examined the protein expression changes of the HHSEC cells before and after the co-culturing as well.
In good concordance with previous findings, we observed expression changes of several cellular adhesion molecules, cytokines and chemokines, including VCAM-1, VEGF, CD105 or VCAM-1 in the HHSEC cells after interacting with melanoma cells [21]. CD105 (Endoglin), which is predominantly expressed by endothelial cells and is involved in tumor angiogenesis, showed higher protein expression in HHSEC cells in four cell lines after co-culture [35]. Li et al. have interpreted that CD105 promotes the invasion and metastases of liver cancer cells by increasing VEGF expression [36]. Vascular cell adhesion molecule-1 (VCAM-1) was also overexpressed in HHSEC as a result of co-culturing in 3 melanoma cell lines. Previously it was described that IL-18 is able to increase the expression of VCAM-1 in the hepatic sinusoidal epithelium promoting the adherence of melanoma cells [37]. Blocking IL-18 with a soluble factor can decrease the adhesion of melanoma cells by inhibiting this mechanism [38]. On the other hand, increased protein expression of Angiopoietin-2 (ANGPT2) was also observed in HHSEC after co-culturing in all of the cell lines. ANGPT2 is a nearly endothelial cell-specific cytokine that promotes vascular remodeling in an autocrine pathway, and its circulating level has been reported to be in association with the progression of metastasis in melanoma patients, particularly in stage III/IV melanoma patients [39,40,41]. Furthermore, Urosevic et al. suggested that ANGPT2 is important for metastatic outgrowth in the liver in colorectal cancer by binding to the TIE2 receptor [42].
Interleukins, such as IL-2, IL-10, IL-12, IL15, IL-21, and their receptors have been shown be efficient mediators of anti-tumor immunity in preclinical cancer models [43,44,45,46]. However, our aim was to find specific receptors which has a possible role in invasion toward hepatic endothelial cells, therefore possibly promote the liver metastasis of melanoma cells, which mechanism is not well-studied yet. According to our results, the conditioned medium produced by hepatic endothelial cells had different effect on the invasion of melanoma cell. Half of the cell lines (WM983A, WM1361, and WM3248) showed increased invasion property after co-culturing with HHSEC-CM compared cells grown in unconditioned medium as control, while three cell lines (WM1366, WM278, and WM793B) had stable invasiveness compared to the controls. In all cell lines, we selected the invasive melanoma cells from non-invasive cells, and compared the chemokine and cytokine receptor gene expression of the two separated subpopulations based on their response to HHSEC conditioned media. Cell lines with increased invasive capacity revealed a set of receptor genes (CXCR1, IL1RL1, IL1RN, IL3RA, IL8RA, IL11RA, IL15RA, IL17RC, and IL17RD) with significantly different expression pattern.
On the other hand, we examined the cytokine and chemokine expression of the hepatic endothelial cells, and found 14 cytokines (CD105, CD147, CXCL8, EGF, CXCL16, Midkine, CD26, CXCL1, CD31, VEGF, CCL20, ANGPT2, VCAM-1, and TSP-1) that showed at least 10% difference after the co-culture with melanoma cells compared to the original protein profile of the hepatic endothelial cells.
CXCR1 (C-X-C chemokine receptor 1) interacting with CXCL8 (Interleukin-8, IL-8) ligand has a well-known role in the initiation and development of various cancers including melanoma [47,48]. Several studies have indicated that the upregulated CXCR1 is associated with enhanced proliferation and invasiveness in melanoma cells, moreover, it is suggested that IL-8 and its receptor promotes liver metastasis in colorectal cancer [49,50,51]. In agreement with these studies, we found that the relative mRNA level of the CXCR1 gene was significantly higher in the selected invasive cells compared to the non-invasive ones, especially in melanoma cells with increased invasiveness after cultured with conditioned medium from HHSEC. On the other hand, CXCL8 showed variable expression pattern with no strong association with melanoma cell invasiveness, however, the highest expression was found in HHSEC cells co-cultured with the WM3248 cell line, which had the highest increased invasiveness of all cell lines.
In case of the IL1RL1 gene, which is coding a specific receptor (ST2) for IL-33 interleukin, it was reported that IL-33/ST2 axis increases the migration and invasion of melanoma cells through ERK1/2 signaling [52]. Interestingly, Luo et al. have been found that the IL-33/ST2 inhibits colon cancer growth and metastasis to lung and liver; on the other hand, Zhang et al. described that increased expression of IL-33 in colorectal cancer cells enhance tumor growth and liver metastasis [53,54]. In our experiments, we found higher expression level of the IL1RL1 gene in melanoma cell lines with increased invasion after HHSEC-CM treatment, but we have not observed any IL-33 expression in the hepatic endothelial cells. The antitumor function of IL-33 is controversial, since Gao et al. suggested that IL-33 accelerate tumor growth in the absence of lymphocytes, and inhibit tumor growth by IL-33-driven immune responses [55].
The role of IL-11 expression was first studied in primary breast cancer which developed metastasis into the bone [56]. IL-11 and its receptor, interleukin-11 receptor α (IL-11Rα) activates STAT3 signaling, which correlates with poor patient prognosis in most human cancers [57,58,59]. In addition, Yamazumi et al. described that the expression of IL11Rα significantly correlates colorectal carcinoma invasion [60]. The overexpression of IL11RA gene in association with melanoma liver metastasis has not been described yet, but important to note that we found significantly higher mRNA level in the selected invasive melanoma cells after HHSEC-CM treatment. We had the opportunity to compare the IL11RA expression level of primary melanoma tissues with liver metastasis and without metastasis, similarly to our model experiments, we found significant increase of the IL11RA gene expression in primary melanoma tissues with liver metastasis. Unfortunately, the access to primary melanoma samples is highly limited, as indicated by the small sample size of our analysis. According to our results, we assume that IL11RA has a possible role in the liver metastasis of primary melanoma cell which requires further investigation.
4. Materials and Methods
4.1. Cell lines and culturing
Primary tumor-derived melanoma cell lines were obtained from the Coriell Institute for Medical Research (Camden, New Jersey, USA). The characteristics and the origins of the cell lines are summarized in Table 1. The cells were cultured in RPMI 1640 medium (Lonza Group Ltd.) supplemented with 10% fetal bovine serum (Gibco, Carlsbad, CA, USA) at 37 °C in 5% CO2.
Human hepatic sinusoidal endothelial cells (HHSEC) were obtained from ScienCell Research Laboratories, Inc. (Carlsbad, CA, USA) and cultured in a 37oC, 5% CO2 incubator according to the recommended protocol. HHSEC cell were maintained using Endothelial Cell Medium in a 2 μg/cm2 fibronectin-coated culture vessel (ScienCell Research Laboratories, Inc., Carlsbad, CA, USA). Conditioned media were collected as described before [22]. Briefly, after the endothelial cells were grown to 90% confluence, culture medium was replaced with fresh medium and incubated for 48 hours. The conditioned medium was centrifuged at 2000g for 15 min, and passed through 0.2 mm filter.
4.2. Melanoma tissue samples used for qRT-PCR
Fresh/frozen melanoma tissues were obtained from the Department of Dermatology at the University of Debrecen (Debrecen, Hungary) from patients who did not undergo therapy before surgical removal of their primary lesions. This study was approved by the Regional and Institutional Ethics Committee of the University of Debrecen [document no.: 25364-1/2012/ EKU (449/PI/12)] and was carried out according to all relevant regulations. Lesions were diagnosed on the basis of formalin-fixed paraffin-embedded tissue sections stained with hematoxylin–eosin. A total of 9 primary melanoma samples were used for the qRT-PCR. Clinical–pathological parameters of the tumors are summarized in Table 2.
4.3. In vitro invasion assay
The invasive potential of melanoma cells was observed using BD Biocoat Matrigel invasion chambers (pore size: 8 μm, 24 wells; BD Biosciences, Bedford, Massachusetts, USA). In the control experiments, the upper chamber was filled with 500 μl of the cell suspension in serum-free media (5 × 104 cells/well) and 750 μl medium containing 10% FBS was applied to the lower chamber as a chemoattractant. To examine the effect of HHSEC on the invasion of melanoma cells, the lower chamber was filled with 750 μl HHSEC conditioned medium (HHSEC-CM) as it was described before [22]. After the cells were incubated for 24 h at 37 °C, the cells in the lower layer were fixed and stained. The invaded cells were counted using a light microscope in seven different visual fields at 200X magnification; the data are presented as the means ± SD of three independent experiments.
4.4. Co-culturing of melanoma cell lines and endothelial cells
For co-culturing, BD Biocoat Matrigel invasion chambers (pore size: 8 μm, 6 wells; BD Biosciences, Bedford, Massachusetts, USA) were used. Melanoma cells (2 × 105 cells/well) were placed to the upper chamber and carefully inserted into 6 well plate containing HHSEC monolayer. Cells were incubated together for 24 hours, then the insert containing melanoma cells were moved to new plate. The invading melanoma cells and the non-invasive cells were treated separately with a 0.5% trypsin/0.2% EDTA solution (Sigma-Aldrich Inc., St. Louis, MO, USA) for recovery from the membrane and cultured using standard protocols.
4.5. Real-time quantitative PCR analysis
RNA isolation was performed using an RNeasy Plus Mini Kit (Qiagen GmbH, Hilden, Germany) according to the manufacturer’s protocol. RNA concentration and quality were assessed using a NanoDrop (Agilent Technologies, Palo Alto, CA, USA). RNA samples with a 260/280 ratio ≥ 1.8 were included in further analyses. Reverse transcription of total RNA (1000 ng) was performed using a High Capacity cDNA Archive Kit (Applied Biosystems, Carlsbad, California, USA) according to the manufacturer’s protocol. Relative expression level of 96 genes (20 chemokine receptor genes, 43 interleukin receptor genes, 25 tumor necrosis factor receptor genes, and eight housekeeping genes) were determined using a LightCycler® 480 Real-Time PCR System (Roche Diagnostics, GmbH, Mannheim, Germany). Primer sets (Human Cytokine and Chemokine Receptor Primer Library) were provided by RealTime Primers (RealTimePrimers.com, Elkins Park, PA) and performed as described before [61]. The data are presented as the mean of 2–ΔCt values from three independent experiments.
4.6. Protein Expression Analysis
Protein expression analyses of HHSEC cells were performed before and after co-culturing with different melanoma cell lines (WM983A, WM793B, WM1366, WM1361, WM278) as described in detail before [62]. RIPA Lysis and Extraction Buffer (Thermo Fisher Scientific Inc., Waltham, MA, USA) containing 20 µL protease and phosphatase inhibitor cocktail (Thermo Fisher Scientific Inc., Waltham, MA, USA) was used for extraction, and the protein concentration was determined using the Pierce™ Coomassie (Bradford) Protein Assay Kit (Thermo Fisher Scientific Inc., Waltham, MA, USA). Proteome Profiler Human Chemokine Array Kit was used to determine the expression of 31 different chemokines, and Proteome Profiler Human XL Cytokine Array Kit was used to analyze 105 cytokines simultaneously (R&D Systems, Inc., Minneapolis, MN, USA). All the necessary reagents and the array procedure was performed according to the manufacturer’s detailed protocol. The labelled proteins were detected and visualized by the Azure c300 Chemiluminescent Imaging System (Dublin, CA, USA) using Chemi Reagent Mix (R&D Systems Inc., Minneapolis, MN, USA). Data were analyzed with AzureSpot (Version: 2.2.167) software. The intensity of the positive control (reference spot) was considered 100%.
4.7. Statistical analysis
IBM SPSS Statistics 26 (IBM Corp., Armonk, NY, USA) software was used for the statistical analyses. The Shapiro–Wilk test was used to evaluate the normality of the data. The Spearman’s and Pearson’s correlation coefficient was calculated to correlate the qPCR data with the invasive capacity of the melanoma cells and with Breslow thickness of primary melanoma samples. The Mann–Whitney–Wilcoxon test and Wilcoxon Signed Ranks Test were used to compare the qPCR data. p < 0.05 was considered to be statistically significant.
5. Conclusions
We found several potential genes and proteins with altered expression in melanoma cells that were co-cultured with human hepatic sinusoidal endothelial cells. Our findings clearly indicate the interaction between liver endothelial- and melanoma cells. Based on our data, overexpression of the IL11RA gene might has key role in the formation of organ specific metastasis to the liver induced by primary melanoma cells.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table S1, Supplementary Table S2. Supplementary Table S3.
Author Contributions
Conceptualization, V.K.. and M.B.; methodology, V.K., and I.Sz.; validation, V.K..; formal analysis, V.K..; investigation, V.K.; resources, M.B. and V.K..; data curation, V.K.; writing—original draft preparation, V.K..; writing—review and editing, V.K. and M.B.; visualization, V.K..; supervision, M.B..; project administration, V.K. and M.B.; funding acquisition, V.K. and M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by y the National Research Development and Innovation Fund (grant number K-135752), the European Regional Development Fund (GINOP-2.3.2-15-2016-00005), the Hungarian Academy of Sciences (TK2016-78), and the Eötvös Loránd Research Network (TKCS-2021/32). The work was also supported by the ÚNKP-22-4 New National Excellence Program of the Ministry for Culture and Innovation from the Source of National research, Development and Innovation Fund (ÚNKP-22-4-II-DE-86).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, X.; Lupardus, P.; Laporte, S.L.; Garcia, K.C. Structural biology of shared cytokine receptors. Annu Rev Immunol 2009, 27, 29–60. [Google Scholar] [CrossRef]
- Talmadge, J.E.; Fidler, I.J. AACR centennial series: the biology of cancer metastasis: historical perspective. Cancer Res 2010, 70, 5649–5669. [Google Scholar] [CrossRef]
- Cucanic, O.; Farnsworth, R.H.; Stacker, S.A. The cellular and molecular mediators of metastasis to the lung. Growth Factors 2022, 40, 119–152. [Google Scholar] [CrossRef]
- Paget, S. The distribution of secondary growths in cancer of the breast. 1889. Cancer Metastasis Rev 1989, 8, 98–101. [Google Scholar]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: organ-specific homes for metastases. Nat Rev Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Gerashchenko, T.S.; Schegoleva, A.A.; Khozyainova, A.A.; Choinzonov, E.L.; Denisov, E.V. Metastasis prevention: How to catch metastatic seeds. Biochim Biophys Acta Rev Cancer 2023, 1878, 188867. [Google Scholar] [CrossRef] [PubMed]
- Obenauf, A.C.; Massague, J. Surviving at a distance: organ specific metastasis. Trends Cancer 2015, 1, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Signore, A.; Chianelli, M.; Bei, R.; Oyen, W.; Modesti, A. Targeting cytokine/chemokine receptors: a challenge for molecular nuclear medicine. Eur J Nucl Med Mol Imaging 2003, 30, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.; Homey, B.; Soto, H.; Ge, N.; Catron, D.; Buchanan, M.E.; McClanahan, T.; Murphy, E.; Yuan, W.; Wagner, S.N.; et al. Involvement of chemokine receptors in breast cancer metastasis. Nature 2001, 410, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.X.; Bos, P.D.; Massague, J. Metastasis: from dissemination to organ-specific colonization. Nat Rev Cancer 2009, 9, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Amos, S.E.; Choi, Y.S. The Cancer Microenvironment: Mechanical Challenges of the Metastatic Cascade. Front Bioeng Biotechnol 2021, 9, 625859. [Google Scholar] [CrossRef] [PubMed]
- Floros, T.; Tarhini, A.A. Anticancer Cytokines: Biology and Clinical Effects of Interferon-alpha2, Interleukin (IL)-2, IL-15, IL-21, and IL-12. Semin Oncol 2015, 42, 539–548. [Google Scholar] [CrossRef]
- Ugurel, S.; Rohmel, J.; Ascierto, P.A.; Flaherty, K.T.; Grob, J.J.; Hauschild, A.; Larkin, J.; Long, G.V.; Lorigan, P.; McArthur, G.A.; et al. Survival of patients with advanced metastatic melanoma: the impact of novel therapies-update 2017. Eur J Cancer 2017, 83, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Damsky, W.E.; Rosenbaum, L.E.; Bosenberg, M. Decoding melanoma metastasis. Cancers (Basel) 2010, 3, 126–163. [Google Scholar] [CrossRef]
- Kastelan, S.; Mrazovac Zimak, D.; Ivankovic, M.; Markovic, I.; Gverovic Antunica, A. Liver metastasis in uveal melanoma - treatment options and clinical outcome. Front Biosci (Landmark Ed) 2022, 27, 72. [Google Scholar] [CrossRef]
- Burns, D.; George, J.; Aucoin, D.; Bower, J.; Burrell, S.; Gilbert, R.; Bower, N. The Pathogenesis and Clinical Management of Cutaneous Melanoma: An Evidence-Based Review. J Med Imaging Radiat Sci 2019, 50, 460–469. [Google Scholar] [CrossRef]
- Tsilimigras, D.I.; Brodt, P.; Clavien, P.A.; Muschel, R.J.; D'Angelica, M.I.; Endo, I.; Parks, R.W.; Doyle, M.; de Santibanes, E.; Pawlik, T.M. Liver metastases. Nat Rev Dis Primers 2021, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- DeLeve, L.D.; Maretti-Mira, A.C. Liver Sinusoidal Endothelial Cell: An Update. Semin Liver Dis 2017, 37, 377–387. [Google Scholar] [CrossRef]
- Poisson, J.; Lemoinne, S.; Boulanger, C.; Durand, F.; Moreau, R.; Valla, D.; Rautou, P.E. Liver sinusoidal endothelial cells: Physiology and role in liver diseases. J Hepatol 2017, 66, 212–227. [Google Scholar] [CrossRef]
- Wilkinson, A.L.; Qurashi, M.; Shetty, S. The Role of Sinusoidal Endothelial Cells in the Axis of Inflammation and Cancer Within the Liver. Front Physiol 2020, 11, 990. [Google Scholar] [CrossRef]
- Clark, A.M.; Ma, B.; Taylor, D.L.; Griffith, L.; Wells, A. Liver metastases: Microenvironments and ex-vivo models. Exp Biol Med (Maywood) 2016, 241, 1639–1652. [Google Scholar] [CrossRef] [PubMed]
- Mendt, M.; Cardier, J.E. Activation of the CXCR4 chemokine receptor enhances biological functions associated with B16 melanoma liver metastasis. Melanoma Res 2017, 27, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Neumann, K.; Erben, U.; Kruse, N.; Wechsung, K.; Schumann, M.; Klugewitz, K.; Scheffold, A.; Kuhl, A.A. Chemokine Transfer by Liver Sinusoidal Endothelial Cells Contributes to the Recruitment of CD4+ T Cells into the Murine Liver. PLoS One 2015, 10, e0123867. [Google Scholar] [CrossRef] [PubMed]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: web-enabled heat mapping for all. Nucleic Acids Res 2016, 44, W147–153. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, D.C.; Pavan, W.J.; Bastian, B.C. The melanomas: a synthesis of epidemiological, clinical, histopathological, genetic, and biological aspects, supporting distinct subtypes, causal pathways, and cells of origin. Pigment Cell Melanoma Res 2011, 24, 879–897. [Google Scholar] [CrossRef]
- Damsky, W.E.; Theodosakis, N.; Bosenberg, M. Melanoma metastasis: new concepts and evolving paradigms. Oncogene 2014, 33, 2413–2422. [Google Scholar] [CrossRef]
- Kim, D.; Gupta, B.; Wong, G.Y.M. Prognostic circulating proteomic biomarkers in colorectal liver metastases. Comput Struct Biotechnol J 2023, 21, 2129–2136. [Google Scholar] [CrossRef]
- Kitsel, Y.; Cooke, T.; Sotirchos, V.; Sofocleous, C.T. Colorectal Cancer Liver Metastases: Genomics and Biomarkers with Focus on Local Therapies. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Rossi, E.; Croce, M.; Reggiani, F.; Schinzari, G.; Ambrosio, M.; Gangemi, R.; Tortora, G.; Pfeffer, U.; Amaro, A. Uveal Melanoma Metastasis. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Karami Fath, M.; Azargoonjahromi, A.; Jafari, N.; Mehdi, M.; Alavi, F.; Daraei, M.; Mohammadkhani, N.; Mueller, A.L.; Brockmueller, A.; Shakibaei, M.; et al. Exosome application in tumorigenesis: diagnosis and treatment of melanoma. Med Oncol 2022, 39, 19. [Google Scholar] [CrossRef]
- Bendas, G.; Borsig, L. Cancer cell adhesion and metastasis: selectins, integrins, and the inhibitory potential of heparins. Int J Cell Biol 2012, 2012, 676731. [Google Scholar] [CrossRef] [PubMed]
- Reymond, N.; d'Agua, B.B.; Ridley, A.J. Crossing the endothelial barrier during metastasis. Nat Rev Cancer 2013, 13, 858–870. [Google Scholar] [CrossRef]
- Strilic, B.; Offermanns, S. Intravascular Survival and Extravasation of Tumor Cells. Cancer Cell 2017, 32, 282–293. [Google Scholar] [CrossRef]
- Gassmann, P.; Haier, J.; Schluter, K.; Domikowsky, B.; Wendel, C.; Wiesner, U.; Kubitza, R.; Engers, R.; Schneider, S.W.; Homey, B.; et al. CXCR4 regulates the early extravasation of metastatic tumor cells in vivo. Neoplasia 2009, 11, 651–661. [Google Scholar] [CrossRef]
- Ceausu, R.A.; Cimpean, A.M.; Gaje, P.; Gurzu, S.; Jung, I.; Raica, M. CD105/Ki67 double immunostaining expression in liver metastasis from colon carcinoma. Rom J Morphol Embryol 2011, 52, 613–616. [Google Scholar] [PubMed]
- Li, Y.; Zhai, Z.; Liu, D.; Zhong, X.; Meng, X.; Yang, Q.; Liu, J.; Li, H. CD105 promotes hepatocarcinoma cell invasion and metastasis through VEGF. Tumour Biol 2015, 36, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Vanaclocha, F.; Fantuzzi, G.; Mendoza, L.; Fuentes, A.M.; Anasagasti, M.J.; Martin, J.; Carrascal, T.; Walsh, P.; Reznikov, L.L.; Kim, S.H.; et al. IL-18 regulates IL-1beta-dependent hepatic melanoma metastasis via vascular cell adhesion molecule-1. Proc Natl Acad Sci U S A 2000, 97, 734–739. [Google Scholar] [CrossRef]
- Carrascal, M.T.; Mendoza, L.; Valcarcel, M.; Salado, C.; Egilegor, E.; Telleria, N.; Vidal-Vanaclocha, F.; Dinarello, C.A. Interleukin-18 binding protein reduces b16 melanoma hepatic metastasis by neutralizing adhesiveness and growth factors of sinusoidal endothelium. Cancer Res 2003, 63, 491–497. [Google Scholar]
- Helfrich, I.; Edler, L.; Sucker, A.; Thomas, M.; Christian, S.; Schadendorf, D.; Augustin, H.G. Angiopoietin-2 levels are associated with disease progression in metastatic malignant melanoma. Clin Cancer Res 2009, 15, 1384–1392. [Google Scholar] [CrossRef]
- Augustin, H.G.; Koh, G.Y.; Thurston, G.; Alitalo, K. Control of vascular morphogenesis and homeostasis through the angiopoietin-Tie system. Nat Rev Mol Cell Biol 2009, 10, 165–177. [Google Scholar] [CrossRef]
- Abdul Pari, A.A.; Singhal, M.; Hubers, C.; Mogler, C.; Schieb, B.; Gampp, A.; Gengenbacher, N.; Reynolds, L.E.; Terhardt, D.; Geraud, C.; et al. Tumor Cell-Derived Angiopoietin-2 Promotes Metastasis in Melanoma. Cancer Res 2020, 80, 2586–2598. [Google Scholar] [CrossRef]
- Urosevic, J.; Blasco, M.T.; Llorente, A.; Bellmunt, A.; Berenguer-Llergo, A.; Guiu, M.; Canellas, A.; Fernandez, E.; Burkov, I.; Clapes, M.; et al. ERK1/2 Signaling Induces Upregulation of ANGPT2 and CXCR4 to Mediate Liver Metastasis in Colon Cancer. Cancer Res 2020, 80, 4668–4680. [Google Scholar] [CrossRef]
- Bentebibel, S.E.; Diab, A. Cytokines in the Treatment of Melanoma. Curr Oncol Rep 2021, 23, 83. [Google Scholar] [CrossRef] [PubMed]
- Dranoff, G. Cytokines in cancer pathogenesis and cancer therapy. Nat Rev Cancer 2004, 4, 11–22. [Google Scholar] [CrossRef]
- Lee, S.; Margolin, K. Cytokines in cancer immunotherapy. Cancers (Basel) 2011, 3, 3856–3893. [Google Scholar] [CrossRef] [PubMed]
- Ardolino, M.; Hsu, J.; Raulet, D.H. Cytokine treatment in cancer immunotherapy. Oncotarget 2015, 6, 19346–19347. [Google Scholar] [CrossRef]
- Liu, Q.; Li, A.; Tian, Y.; Wu, J.D.; Liu, Y.; Li, T.; Chen, Y.; Han, X.; Wu, K. The CXCL8-CXCR1/2 pathways in cancer. Cytokine Growth Factor Rev 2016, 31, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Raimondo, M.; Woodward, T.A.; Wallace, M.B.; Gill, K.R.; Tong, Z.; Burdick, M.D.; Yang, Z.; Strieter, R.M.; Hoffman, R.M.; et al. CXC-chemokine/CXCR2 biological axis promotes angiogenesis in vitro and in vivo in pancreatic cancer. Int J Cancer 2009, 125, 1027–1037. [Google Scholar] [CrossRef]
- Singh, S.; Nannuru, K.C.; Sadanandam, A.; Varney, M.L.; Singh, R.K. CXCR1 and CXCR2 enhances human melanoma tumourigenesis, growth and invasion. Br J Cancer 2009, 100, 1638–1646. [Google Scholar] [CrossRef]
- Shang, F.M.; Li, J. A small-molecule antagonist of CXCR1 and CXCR2 inhibits cell proliferation, migration and invasion in melanoma via PI3K/AKT pathway. Med Clin (Barc) 2019, 152, 425–430. [Google Scholar] [CrossRef]
- Bie, Y.; Ge, W.; Yang, Z.; Cheng, X.; Zhao, Z.; Li, S.; Wang, W.; Wang, Y.; Zhao, X.; Yin, Z.; et al. The Crucial Role of CXCL8 and Its Receptors in Colorectal Liver Metastasis. Dis Markers 2019, 2019, 8023460. [Google Scholar] [CrossRef]
- Yang, F.; Wen, M.; Pan, D.; Lin, X.; Mo, J.; Dong, X.; Liao, S.; Ma, Y. IL-33/ST2 Axis Regulates Vasculogenic Mimicry via ERK1/2-MMP-2/9 Pathway in Melanoma. Dermatology 2019, 235, 225–233. [Google Scholar] [CrossRef]
- Luo, P.; Deng, S.; Ye, H.; Yu, X.; Deng, Q.; Zhang, Y.; Jiang, L.; Li, J.; Yu, Y.; Han, W. The IL-33/ST2 pathway suppresses murine colon cancer growth and metastasis by upregulating CD40 L signaling. Biomed Pharmacother 2020, 127, 110232. [Google Scholar] [CrossRef]
- Zhang, Y.; Davis, C.; Shah, S.; Hughes, D.; Ryan, J.C.; Altomare, D.; Pena, M.M. IL-33 promotes growth and liver metastasis of colorectal cancer in mice by remodeling the tumor microenvironment and inducing angiogenesis. Mol Carcinog 2017, 56, 272–287. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, X.; Yang, Q.; Zhao, X.; Wen, W.; Li, G.; Lu, J.; Qin, W.; Qi, Y.; Xie, F.; et al. Tumoral expression of IL-33 inhibits tumor growth and modifies the tumor microenvironment through CD8+ T and NK cells. J Immunol 2015, 194, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Sotiriou, C.; Lacroix, M.; Lespagnard, L.; Larsimont, D.; Paesmans, M.; Body, J.J. Interleukins-6 and -11 expression in primary breast cancer and subsequent development of bone metastases. Cancer Lett 2001, 169, 87–95. [Google Scholar] [CrossRef]
- Putoczki, T.L.; Thiem, S.; Loving, A.; Busuttil, R.A.; Wilson, N.J.; Ziegler, P.K.; Nguyen, P.M.; Preaudet, A.; Farid, R.; Edwards, K.M.; et al. Interleukin-11 is the dominant IL-6 family cytokine during gastrointestinal tumorigenesis and can be targeted therapeutically. Cancer Cell 2013, 24, 257–271. [Google Scholar] [CrossRef]
- To, S.Q.; Dmello, R.S.; Richards, A.K.; Ernst, M.; Chand, A.L. STAT3 Signaling in Breast Cancer: Multicellular Actions and Therapeutic Potential. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Xu, L.; Liu, H.; Zhang, W.; Liu, W.; Liu, Y.; Fu, Q.; Xu, J. High expression of interleukin-11 is an independent indicator of poor prognosis in clear-cell renal cell carcinoma. Cancer Sci 2015, 106, 592–597. [Google Scholar] [CrossRef]
- Yamazumi, K.; Nakayama, T.; Kusaba, T.; Wen, C.Y.; Yoshizaki, A.; Yakata, Y.; Nagayasu, T.; Sekine, I. Expression of interleukin-11 and interleukin-11 receptor alpha in human colorectal adenocarcinoma; immunohistochemical analyses and correlation with clinicopathological factors. World J Gastroenterol 2006, 12, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Koroknai, V.; Szasz, I.; Jambor, K.; Balazs, M. Cytokine and Chemokine Receptor Patterns of Human Malignant Melanoma Cell Lines. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Szasz, I.; Koroknai, V.; Kiss, T.; Vizkeleti, L.; Adany, R.; Balazs, M. Molecular alterations associated with acquired resistance to BRAFV600E targeted therapy in melanoma cells. Melanoma Res 2019, 29, 390–400. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The effect of human hepatic sinusoidal endothelial cells conditioned media (HHSEC-CM) on the invasion of melanoma cell lines (WM1366, WM278, WM793B, WM983A, WM1361, and WM3248) compared to the effect of unconditioned medium as control. Results are presented as the ratio (mean ± SE, n=3) of HHSEC-CM induced and control invaded cells. The asterix indicates statistically significant difference between HHSEC-CM induced data and control data (Mann-Whitney test: p < 0.05). The results of the invasion assay are shown as the mean of three independent experiments.
Figure 1.
The effect of human hepatic sinusoidal endothelial cells conditioned media (HHSEC-CM) on the invasion of melanoma cell lines (WM1366, WM278, WM793B, WM983A, WM1361, and WM3248) compared to the effect of unconditioned medium as control. Results are presented as the ratio (mean ± SE, n=3) of HHSEC-CM induced and control invaded cells. The asterix indicates statistically significant difference between HHSEC-CM induced data and control data (Mann-Whitney test: p < 0.05). The results of the invasion assay are shown as the mean of three independent experiments.
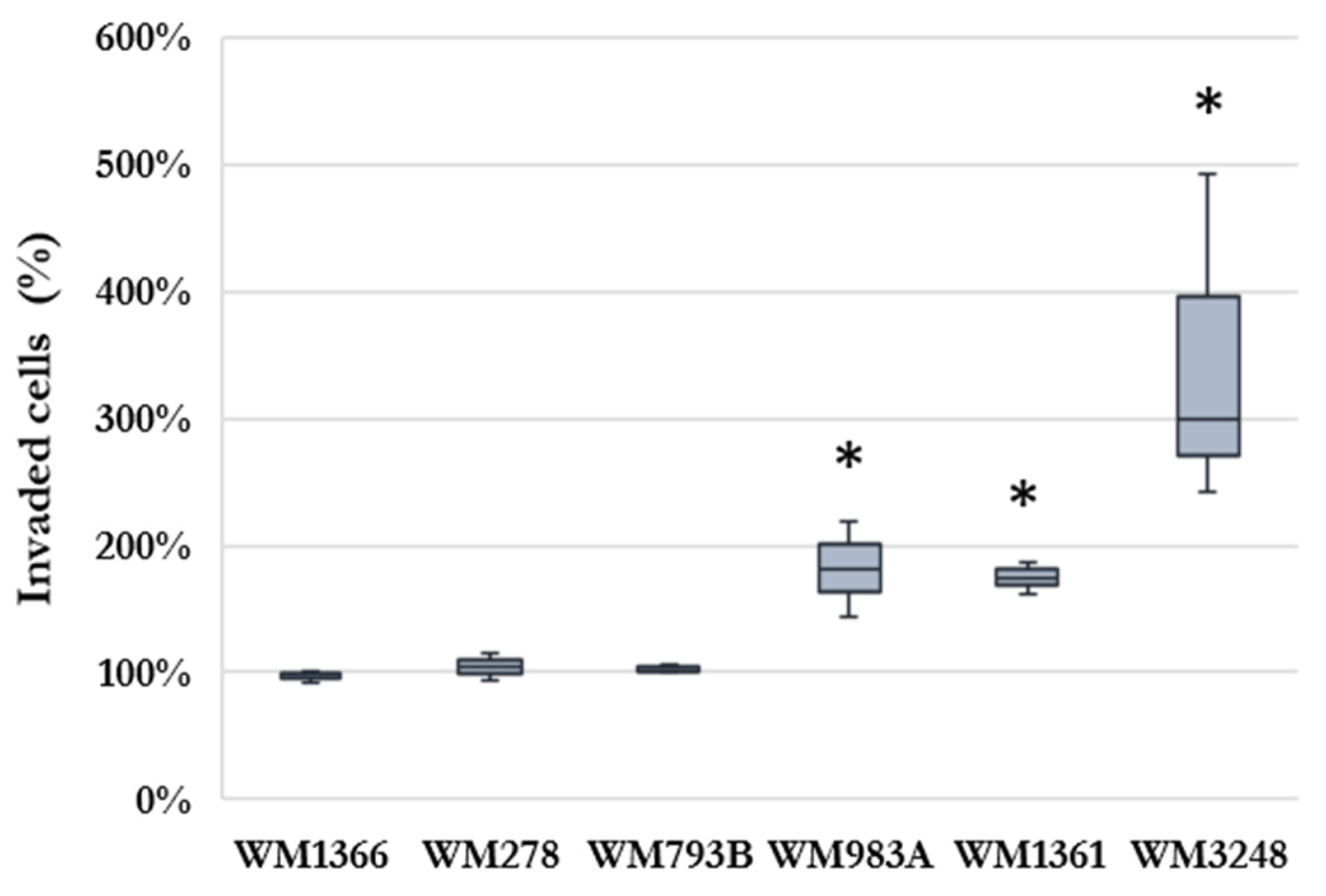
Figure 2.
Association between chemokine- and cytokine receptor expression of melanoma cells and the invasive capacity responding to human hepatic sinusoidal endothelial cells conditioned media (HHSEC-CM). (A) Venn diagram showing the gene expression alterations in cell lines with stable invasiveness (a), and in cell lines with increased invasiveness (b). Arrows indicate over-expression (↑) and down-regulation (↓) in the selected invasive cells compared to the non-invasive populations (B) Comparison of the relative mRNA expressions of IL11RA gene in selected non-invasive and invasive cells according to the invasive capacity of the cell lines. Asterix indicates significant differences. (C) Unsupervised hierarchical clustering of the significantly differently expressed genes in the melanoma cell lines with increased invasiveness. The separated non-invasive (NON-INV) and invasive (INV) cells are displayed vertically, and the genes are displayed horizontally. The heat map was generated from the seven significantly expressed genes using www.heatmapper.ca [24].
Figure 2.
Association between chemokine- and cytokine receptor expression of melanoma cells and the invasive capacity responding to human hepatic sinusoidal endothelial cells conditioned media (HHSEC-CM). (A) Venn diagram showing the gene expression alterations in cell lines with stable invasiveness (a), and in cell lines with increased invasiveness (b). Arrows indicate over-expression (↑) and down-regulation (↓) in the selected invasive cells compared to the non-invasive populations (B) Comparison of the relative mRNA expressions of IL11RA gene in selected non-invasive and invasive cells according to the invasive capacity of the cell lines. Asterix indicates significant differences. (C) Unsupervised hierarchical clustering of the significantly differently expressed genes in the melanoma cell lines with increased invasiveness. The separated non-invasive (NON-INV) and invasive (INV) cells are displayed vertically, and the genes are displayed horizontally. The heat map was generated from the seven significantly expressed genes using www.heatmapper.ca [24].
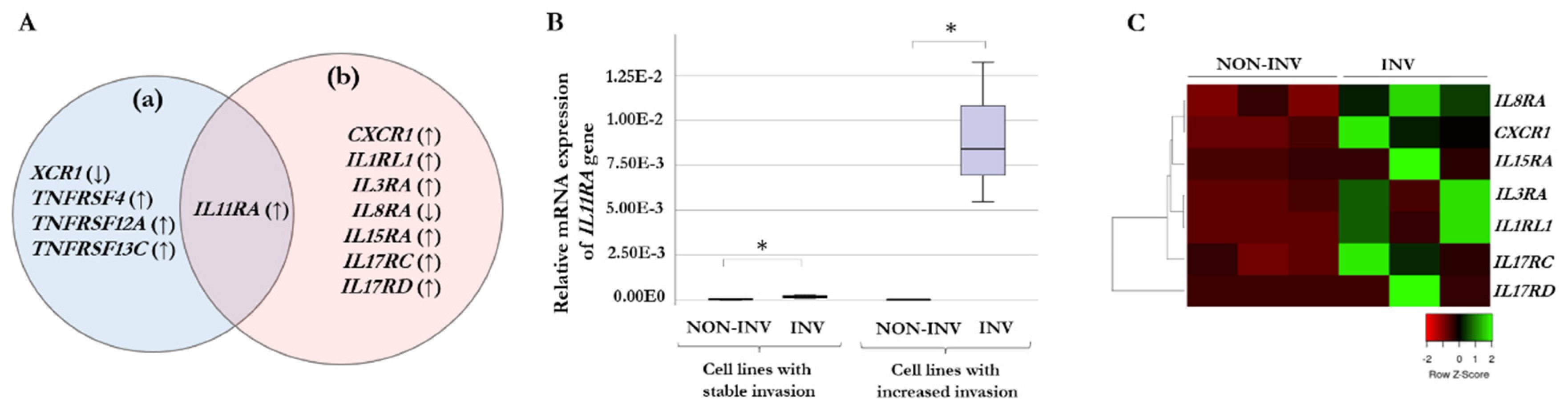
Figure 3.
Correlation between gene expression and invasiveness in melanoma cell lines. Significant positive correlations (p < 0.05) between the relative expression of the IL15RA, IL17RC, IL17RD genes and the HHSEC-CM induced invasion changes (%). The qRT-PCR results are presented as the mean ratio of 2−ΔCt values between invasive and non-invasive populations (log2-transformed data) of three independent experiments.
Figure 3.
Correlation between gene expression and invasiveness in melanoma cell lines. Significant positive correlations (p < 0.05) between the relative expression of the IL15RA, IL17RC, IL17RD genes and the HHSEC-CM induced invasion changes (%). The qRT-PCR results are presented as the mean ratio of 2−ΔCt values between invasive and non-invasive populations (log2-transformed data) of three independent experiments.
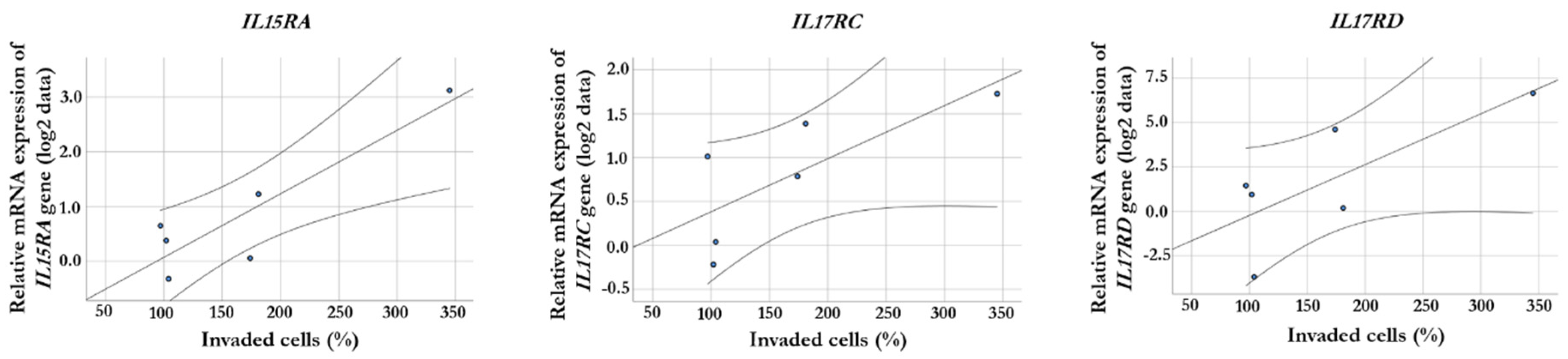
Figure 4.
Relative protein expression profiles of human hepatic sinusoidal endothelial cells (HHSECs) co-cultured with different melanoma cell lines. The intensity of the reference is displayed as 100%. Numbers beside the columns indicate the protein expression as a percentage of the intensity of the reference spots on the array. Protein expression in control HHSECs is indicated in green color, protein expression in HHSECs co-cultured with melanoma cell lines with stable invasiveness is indicated in blue color, and protein expression in HHSECs co-cultured with cell lines with increased invasiveness is indicated in red color.
Figure 4.
Relative protein expression profiles of human hepatic sinusoidal endothelial cells (HHSECs) co-cultured with different melanoma cell lines. The intensity of the reference is displayed as 100%. Numbers beside the columns indicate the protein expression as a percentage of the intensity of the reference spots on the array. Protein expression in control HHSECs is indicated in green color, protein expression in HHSECs co-cultured with melanoma cell lines with stable invasiveness is indicated in blue color, and protein expression in HHSECs co-cultured with cell lines with increased invasiveness is indicated in red color.
Figure 5.
Association between cytokine receptor gene expression of primary melanoma tumor samples and the clinical pathological characteristics. (A) Significantly increased relative expression level of the IL11RA gene in primary melanoma samples with liver metastasis compared to non-metastatic primary melanoma samples. Asterix indicates significant difference (p = 0.049). (B) Significantly positive correlation between IL17RD gene expression and Breslow thickness (mm) of the primary tumors (p = 0.014). The qRT-PCR results are presented as 2−ΔCt values.
Figure 5.
Association between cytokine receptor gene expression of primary melanoma tumor samples and the clinical pathological characteristics. (A) Significantly increased relative expression level of the IL11RA gene in primary melanoma samples with liver metastasis compared to non-metastatic primary melanoma samples. Asterix indicates significant difference (p = 0.049). (B) Significantly positive correlation between IL17RD gene expression and Breslow thickness (mm) of the primary tumors (p = 0.014). The qRT-PCR results are presented as 2−ΔCt values.
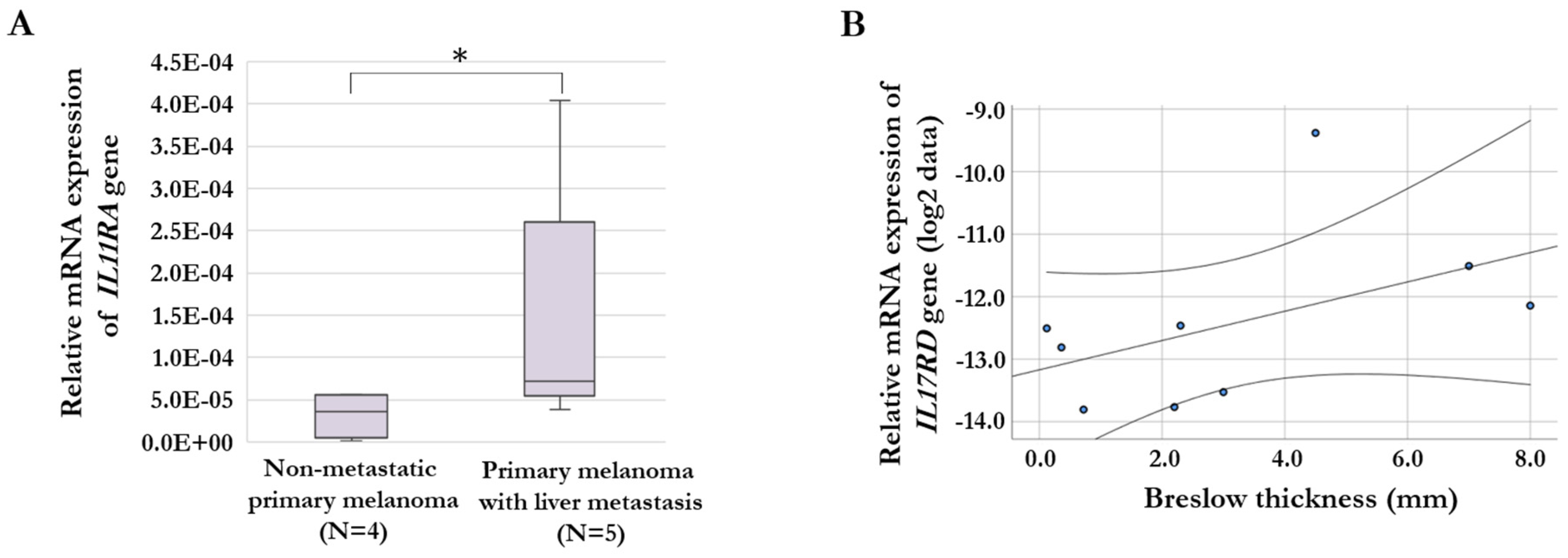

Table 1.
Characteristics of primary tumor originated melanoma cell lines.
| Cell line | Growth phase1 | Histologic subtype2 | BRAF mutation status3 | NRAS mutation status4 |
| WM793B | RGP/VGP | SSM | V600E | wt |
| WM1361 | VGP | SSM | wt | Q61L |
| WM278 | VGP | NM | V600E | wt |
| WM983A | VGP | n.a. | V600E | wt |
| WM1366 | VGP | n.a. | wt | Q61L |
| WM3248 | VGP | n.a. | V600E | wt |
| 1RGP: radial growth phase, VGP: vertical growth phase; 2SSM: superficial spreading melanoma, NM: nodular melanoma, n.a.: data not available; 3V: valine, E: glutamic acid, wt: wild-type; 4Q: glutamine, L: leucine | ||||
Table 2.
Clinical-pathological data of melanoma tissue samples used in qRT-PCR analysis.
| Sample number | Gender1 | Age at initial diagnosis (years) | Location | Histological subtype2 | Breslow thickness (mm) | Ulceration |
| Primary melanoma with no metastasis3 | ||||||
| 1 | F | 64 | Extremities | SSM | 0.4 | No |
| 2 | M | 67 | Head | NM | 0.1 | No |
| 3 | M | 72 | Trunk | NM | 4.5 | No |
| 4 | M | 59 | Trunk | SSM | 0.7 | No |
| Primary melanoma with liver metastasis3 | ||||||
| 5 | M | 71 | Trunk | SSM | 2.3 | No |
| 6 | M | 40 | Extremities | NM | 3.0 | No |
| 7 | M | 69 | Trunk | SSM/NM | 8.0 | Yes |
| 8 | M | 63 | Trunk | SSM | 2.2 | No |
| 9 | F | 71 | Trunk | NM | 7.0 | Yes |
| 1F: female, M: male; 2SSM: superficial spreading melanoma, NM: nodular melanoma; 3Patients with at least 5-years follow-up period | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



