
Submitted:
19 April 2023
Posted:
20 April 2023
You are already at the latest version
Abstract
The physiological mechanisms that underlie the generation and maintenance of pain are diverse. There are afferent fibers that receive sensory information and transfer the information to the central nervous system. However, when trying to classify and determine their molecular characteristics, it becomes a difficult task due to the variety of markers that they share. With current technological advances, it is possible to specifically modulate the fibers that process pain signaling through optogenetics and chemogenetics. Nonetheless, to accomplish this aim it is necessary a specific identification of each neuron subtypes involved. This review addresses the molecular markers present in the nociceptive fibers responsible for sensory signals and the use of these molecular markers as optogenetics targets to modulate nociceptive behavior.
Keywords:
Pain
; Vglut2
; optomodulation
; chemogenetic
1. Introduction
Pain is a complex experience that involves not only the physical sensation of tissue damage or injury, but also emotional and cognitive components. It is important to distinguish between pain and nociceptive behavior, which refers to the body's automatic and reflexive response to tissue damage or injury. While nociceptive behavior helps protect the body from further injury and promotes healing, pain is a subjective experience that is influenced by a variety of factors, including emotions, past experiences, and cognitive processes. Understanding the differences between pain and nociceptive behavior is crucial to understanding the pathophysiological processes underlying the pathophysiology of painful conditions and developing effective therapeutic strategies.
The physiological process for pain perception is generated through the transmission of information from the site of damage to the central nervous system (CNS). The neurons that capture signals are essential elements in the perception of pain. These terminal endings confront a pseudo unipolar neuron that is in the dorsal root ganglion. The axon of this neuron is divided into two branches, one projecting to the periphery and the other to the CNS [1,2]. All sensory information is captured by the primary afferent fibers of various types, that carry the signal to the dorsal horn of the spinal cord. The sensory primary afferents are categorized into three main groups, according to their diameter, degree of myelination and driving speed. Neurons whose cell body is large in diameter are A-β neurons, they have myelinated axons which gives them the characteristic of fast conduction. The medium-sized neurons are called A-δ and have lightly myelinated axons and moderate conduction velocities, while neurons with small diameter are C fibers, whose axon is unmyelinated and their conduction velocity is the slowest [3].
With recent techniques as optogenetics or chemogenetics is possible to activate or inactivate neuronal circuits in a specific way; thus, is essential to identify molecular targets from each fiber, such as channels, receptors and transcription factors that allow a selective transfection and expression of opsins to modulate their activity, like the vesicular transporters that recently had been described. Herein we review the molecular targets and their colocalization with the vesicular transporter of glutamate. Also, we will review how optical control of specific sensory fibers could drive the nociceptive process.
2. Neural Types in the Dorsal Ganglia Root and Their Colocalization with the Vesicular Transporter of Glutamate
The A-β fibers have an axon diameter greater than the others, are highly myelinated and therefore their velocity of conduction is between 30-80 m/s [4], and respond to innocuous mechanical stimulation. The projection of these neurons makes synaptic contacts in the III-V layers with second order neurons in the dorsal horn of the spinal cord. The molecular profile (Table 1, section 1) include the following protein markers that are characteristic of large sized neurons in dorsal root ganglion (DRG) such as NF200 (neurofilament 200), NEFH (neurofilament heavy), Thy-1 (Thymocyte differentiation antigen 1), TrKC (Tropomyosin receptor kinase C), Cdh12 (Cadherin 12), parvalbumin [5-8], biomarkers that are correlated with the survival and stability of nerve terminals related to dynamic touch, gentle skin stroke, kinesthetic sense of position, skin indentation, poke, pressure on guard hairs, or texture perception by fingertips [9].
The A-β fibers neurons also express specific channels as Nav1.1 and 1.6 (voltage-gated sodium channel), ASIC1 (Acid Sensing Ion Channel Subunit 1), HCN1 (Hyperpolarization Activated Cyclic Nucleotide Gated Potassium Channel 1) and HCN2 (Hyperpolarization-Activated Cyclic Nucleotide Gated Potassium Channel 2) [6,10-12]. Although these HCN channels are expressed in a variety of sensory neuron types, their expression in A-β neurons is related to their conduction velocities and frequencies due to their current properties [13].
Transcription factors are another set of important markers that act as regulators of lineage-specific gene expression. Among A-β fibers, RunX3 (runt-related transcription factor 3), ETV1 (ETS Variant Transcription Factor 1), ETV4 (ETS Variant Transcription Factor 4) [6,14-16], c-Ret and MafA/c-Maf. At the same time three subpopulations can be differentiated, those that express Ret+ MafA+, Ret+ MAfA+ TrkC+, and MAFA+ TrkB+ Shox2+. The presence of these markers conditions the expression of TrkC, and the differentiation to A-β fibers [17,18].
In addition to the fibers that receive harmless sensory information, there are neuronal pathways that detect painful information such as A-δ and C-fibers. The A-δ fibers are lightly myelinated and have a relatively fast conduction velocity of 5-25 m/s. The diameter of Aδ-fibers is about 2–5 µm, and are responsive of short lasting and pricking nociceptive behavior, transmitting the immediate response to pain stimuli, which generates a withdrawal reaction from the source of stimulation. The markers are difficult to distinguish in A-δ neurons, since they express overlapped markers with other subtypes of proteins, channels, and metabotropic receptors. The A-δ and A-β fibers share markers, such as NF200 and NEFH, but also have proteins on their surface such as substance P and CGRP that are also shared in C fibers [5]. One of the markers by which they can be differentiated is the presence of the TrkB receptor and the voltage-dependent calcium channel 3.2 (Cav 3.2) [19]. Other notable differences are the presence of transcription factors such as Contactin Associated Protein 2 (CNTNAP2) and Family with Sequence Similarity 19 Member A1 (FAM19A1) [12].
The C-fibers have an important role in the pathophysiology of pain transmit, [20]. C- fibers are unmyelinated with less than 2 µm in diameter and have a relatively slow conduction velocity of approximately 0.5-2 m/s. C-fibers terminate in laminae I and II in the dorsal root ganglia, and correspond to polymodal nociceptors, which are activated by thermal, mechanical and chemical stimuli. They have diverse markers that allow the identification of different neuronal subpopulations. The peptidergic neurons express substance P (SP) and the calcitonin-gene related protein (CGRP) while neurons that bind to isolectin B4 (IB4) are considered IB4+ neurons or non-peptidergic [12,21-23].
There are protein markers or transcription factors that allow the differentiation of C fibers. One important protein family is the transient receptor potential (TRP) channel, a cationic channel that senses the injurious information. There are 6 families of TRP, the canonical (TRPC), vanilloid (TRPV), ankyrin (TRPA), melastatin (TRPM), polycystin (TRPP), and mucolipin (TRPML).
C-peptidergic neurons are distinguished by the expression of TRPV1, a channel that participates in high threshold thermal sensitivity and harmful chemical capsaicin [22,39]. These fibers also express the TRPA1 and TRPM8 receptors, which function as channels that can be activated in the presence of irritating agents, inflammatory mediators, and cold temperatures. Nonpeptidergic IB4+ type-C fibers also have TRPA1; however, these fibers are differentiated by the expression of TRPM3, which is characterized by its activation in the cold thermal stimuli (-40 °C) and TRPC3, which plays an important role in sensitization to noxious stimuli [46].
The sodium channels play an important role in the generation of excitability for the transmission of pain towards the dorsal horn of the spinal cord and are distributed within the C fibers. There are nine isoforms of this protein (Nav1.1 to Nav1.9), of which Nav 1.8 is found in peptidergic and nonpeptidergic fibers, while the Nav 1.7 channel is expressed in peptidergic C fibers, and Nav 1.9 in IB4+ neurons [28].
The P2X family are ligand-gated ion channels mediating the actions of extracellular ATP and other nucleotides released after tissue damage. The activation of these channels contributes to the generation and maintenance of pain, due to a distribution in primary afferent neurons. A total of seven P2X receptors have been described so far (P2X1-P2X7), with C-nonpeptidergic neurons expressing the purinergic receptor P2X3 [47,48].
Neurons
In addition, there are metabotropic protein receptors in C-fibers as opioid, dopamine (DA), the Mas related G protein coupled receptor (Mrg), or Tropomyosin receptor kinase. Opioid receptors play an important role in modulating the nociceptive signal and are distributed in the nerve circuits of pain. MOR receptor (µ opioid receptor) colocalizes with substance P and CGRP in peptidergic fibers, while the DOR receptor (δ opioid receptor) is located in neurons that do not express neuropeptides or TRPV1 [30,49-51] (Figure 1). Similarly, there is also a clear difference in expression in the subpopulation of C fibers with the dopamine receptor. It has been suggested that presynaptic and postsynaptic effects are responsible for the antinociceptive effect of DA and are mediated mainly by D2-like receptors [52-54]. A D2-like receptor agonist, produces antinociception to mechanonociceptive stimuli but not to thermonociceptive stimuli [55,56], denoting that D2 receptor expressed mainly in IB4+ small DRG neurons. Lkewise, Y1R is found in peptidergic neurons. IB4+ fibers express both the Y1R and Y2R receptors.
Pituitary adenylate cyclase-activating polypeptide (PACAP) belongs to the VIP/secretin/glucagon superfamily. Three distinct G-protein-coupled receptors mediate the actions of PACAP and VIP, present in the spinal dorsal horn and dorsal root ganglia, suggesting an important role of PACAP signaling systems in the modulation of spinal nociceptive transmission [57]. Its effects are mainly produced through the cAMP-protein kinase A (PKA) pathway by PACAP type I (PAC1) receptor [41], inducing the cAMP responsive element binding protein (CREB) activation. In C-fibers, PACAP and their agonists induce phosphorylation of both extracellular signal regulated kinase (ERK) via PKA in the dorsal horn neurons. This phenomenon is associated with spontaneous nociceptive behavior. The Jun N-Terminal Kinase (JNK) is activated subsequently in astrocytes directly by PAC1 receptor signaling, or via ERK signaling pathway in the dorsal horn of the spinal cord, which are involved in the long-term maintenance of pain [58].
C-fibers also differ in their central terminals in the spinal cord. The peptidergic subset terminates more superficially, in lamina I and the outer part of lamina II, while the IB4+-nociceptors project mainly to inner lamina II of the dorsal horn of the spinal cord [9]. A subset of C-fiber has been identified, whose characteristic is to be a low-threshold tactile mechanoreceptor (C-LTMR). These afferents are characterized by the expression of thyroxine hydroxylase (TH) and TrkB (a receptor for both brain- derived neurotrophic factor (BDNF), neurotrophin-4 (NT4), Ret and vGlut3 [59-61].
Different combinations of molecular markers are used to separate the variety of neuronal subtypes that process various forms of nociceptive information. Understanding the etiology of pain requires research into how these molecular markers alter along each of these routes or subcircuits. Although there is more than one marker for each type of fiber, the subpopulation is defined by a specific combination of markers.
In addition to these neurochemical markers and transcription factors, other characteristics have been found in these fibers, such as the vesicular transporter. Glutamate is considered the main excitatory amino acid within the nervous system and has been identified as an important factor in the generation of central pain sensitization [31,62]. To maintain glutamate homeostasis there are two main transporters, the excitatory amino acid transporters subtype 2 (EAAT2) and vesicular glutamate transporter (VGLUT). Therefore, it is important to identify the neurons that release glutamate, especially those related to the transmission of pain. There are three VGLUT subtypes that have been identified so far. The differences in vesicle subtypes have been associated with the colocalization of the primary afferent fibers. One example is the VGLUT1 that is present in most of the large-sized neurons of the DRG; these neurons originate from the A-β fibers [63,64]. Moreover, it has been demonstrated that VGLUT2 colocalizes with CGRP and IB4+ neurons, suggesting that C-peptidergic and C-non-peptidergic package glutamate by this subtype of vesiculator [21,63]; and C-LTMR are Vglut-3 [65].
3. Pain Pathway Alteration
One of the main signaling pathways involved in the inhibitory process of neuronal excitability is the GABAergic system. The GABA receptor is known as the main regulator of the painful signal, promoting hyperpolarization of neurons in the spinal cord [66]. The injury to the nerve promotes anatomical changes that cause a redutction in the number of inhibitory synapses in the dorsal spinal cord. In addition to this, molecular changes are generated that modify the inhibition process through the GABA receptor [67-69]. The excitability of CNS is regulated by intracellular Cl-, mainly by changes in its intracellular concentrations. These changes are sensed by the protein kinase WNK1 which will activate the SPAK kinase and subsequently control the permeability of the KCC2 and NKCC1 transporters. When these transporters are phosphorylated, the internal concentration of Cl- is higher during chronic pain. This change in internal Cl- concentration impairs the hyperpolarizing function of GABA, so that in the chronic stage of neuropathic pain GABA now becomes depolarizing (Figure 2). These changes in the GABAergic function modify the intrinsic excitability of projection neurons in the spinal cord [69-70]. This notion raises the question whether this change in Cl- concentrations is occurring in all or only some neurons, and is a subject of debate that should be addressed in future studies.
4. Optomodulation of Peripheral Nerve Activity
Among the problems to understand neuronal circuits was the control of a group of neurons in a specific way without affecting neighboring cells and understanding cell physiology. This was possible by the application of optogenetics control on physiological processes. Optogenetics is a technique that involves the use of light to control neuronal activity through ion channels [72,73]. This novel method allows the activation or inactivation of excitable cells through the expression of types of optical activated channels called opsins, of which the main are channelrhodopsin (ChR2), halorhodopsin (NpHR) and archeorodopsin (ArchT). The study and understanding of nerve circuits that are related to the perception and transmission of pain toward the spinal cord have been favored by the selective modulation of neuronal subsets by optogenetics.
ChR2 is a protein that is activated by a luminous beam with a wavelength of 450-480 nm (blue light) and its stimulation allows the passage of cations such as Na+, K+ and Ca++ into the cell, which in the case of a neuronal cell, it generates a depolarization that creates an action potential (i.e., activation) [74]. The other protein, NpHR, is activated with a luminous beam with a wavelength of 570-600 nm. Its activation generates the entry of Cl- to the neuron, causing a negative membrane potential that results in a phenomenon of hyperpolarization, having the inactivation of neuron as final effect [75]. Similar effect is observed with the expression of ArchT channel, however this hyperpolarization is given by the H+ output. In the case of this protein, the activation is also given by the exposure to yellow light with a wavelength of 520-560 nm [76]. The expression of these proteins can be achieved by viral transfection or breeding strategies through molecular markers, which will allow the control of the neuronal activity in a specific way.
One of the first methods used is through the non transgenic (i.e., wild type) animal and the use of viral vectors not CRE-dependent. This is achieved by endowing the virus with the opsin gene together with a non-specific promoter, such as the human synapsin 1 gene (hsyn), when the aim is to perform a generalized (i.e., non-specific) neuronal transfection. Other widely used non-specific promoters are CaMKII, that directs transfection mainly towards glutamatergic neurons, or VGAT, for GABAergic neurons [77]. The use of specific promoters provide specificity to the cell and as a consequence, to the model.
The other method for transfecting viral vectors is by using transgenic animals, such as Cre, Dre or FLP recombinase systems. These recombinases are enzymes that catalyze recombination between two relatively close sites and the genetic material between them. When a viral vector containing the opsin gene between the recognition sites is introduced in animals whose cells have the recombinase driven by a specific promoter, they will express the opsin in specific neurons of interest [78]. Nevertheless, in order to be specific for the neuronal subtype, it is necessary to use more than one promoter with at least two recombinases, this is the so-called intersection technique (i.e. Vglut2 Cre intersection with TH Flp). However, to date this intersection technique has not been used in nociceptive pathways and the published research until now still uses only one promoter.
Optomodulation of Primary Afferent Fibers with Viral Transfections and Constitutive Expression
There are studies that have used these strategies to demonstrate the modulation of sensory pathways [19,23,50,79], which are summarized in Table 2. One of the first transfection attempts was using the Nav1.8 promoter, which is present in all sensory pathways. Therefore this strategy was not specific of a fiber subtype, using a constitutive strategy composed by a homozygous Nav1.8 –Cre mice crossed with a heterozygous Ai32 mice, which carry the ChR2(H134R)–EYFP gene in their Gt(ROSA)26Sor locus getting Nav.18-ChR2. The selective stimulation with blue light in freely moving mice, exhibit nocifensive behaviors, paw withdrawal, paw licking, jumping, and audible vocalization, this wide variety of behaviors are explained by the global activation of nociceptive fibers, which present the expression of the Nav1.8 channel in both peptidergic and non-peptidergic neurons [79]. This work showed that the sensitization is evocated in similar way to the painful stimuli when ChR2 activate the Nav1.8+ neurons, which is present in a generalized way in afferent sensory fibers.
On the other hand, in Nav1.8-Arch mice, obtained with heterozygous Nav1.8-Cre mice crossed with homozygous Ai35 mice carrying the floxed stop-Arch-EGFP gene in the ROSA26, the mechanical and thermal sensitization was completely prevented in the stimulation of yellow light. Additionally, optical stimulation reduced capsaicin- and zymosan-induced mechanical allodynia [39]. These results together suggest that afferent fibers with Nav1.8+ play an important role in the onset and maintenance of pain. These results are expected; however, it is difficult to attribute the effects to a neuronal subtype involved in pain transmission, since Nav 1.8 has a widespread distribution in primary afferent fibers.
To transfect peptidergic fibers, the Cre-dependent strategy has been used under TRPV1 promoter control. Beaudry [23] transfected a neuronal subgroup using homozygous TRPV1-Cre mice injected intrathecally with an adeno-associated virus (AAV) 2/8 virus (AAV2/8-CAG-floxed stop-ChR2[H134R]-tdTomato-WPRE) to generate TRPV1-ChR2 mice. The behavior was reported as paw withdrawal, paw lifting, and paw licking [23]. In the same group in neurons but in constitutive strategy cross-breeding heterozygous rosa-CAG-LSL-hChR2(H134R)-tdTomato-WPRE (Ai27D) mice with heterozygous mice with Cre recombinase inserted downstream to the TRPV1 specific expression of ChR2 in the TRPV1-Cre gene showed no place preference in the absence of photostimulation, whereas its activation with light blue triggered marked aversion of the stimulation-paired area, with a corresponding increase in time spent in the non-stimulation-paired area [80]. In opposite way, the inhibitions of TRPV1 by ArchT with an injection in the DRG of adeno-associated virus (AAV5-TRPV1-ArchT-eGFP) increase mechanical paw withdrawal threshold and the latency to thermal stimulation [19]. Thus, the modulation of c-peptidergic fibers (TRPV1+) produces burning pain or itch that generates aversion.
To activate non-peptidergic fibers, Beudry, et al., [23] used the Mas-related G protein–coupled receptor subtype D (MrgD) with homozygous MrgD::CreERT2 mice crossed with homozygous Ai32 carry in the ROSA26 locus the floxed stop-ChR2(H134R)-EYFP construct mice to generate the MrgD-ChR2 heterozygous transgenic mouse. To induce opsin expression in MrgD-ChR2 mice with Cre-inducible, tamoxifen was injected intraperitoneally. The optical stimulation with blue light evoked paw withdrawal and paw lifting mainly and lesser paw licking [23]. Overall, TRPV1 and MRGD fiber activation showed similar conducts. However, in quantitative analysis, TRPV1-ChR2 mice spent more time licking whilst MRGD-ChR2 mice spent more time lifting, reflecting different somatosensory perception differences that agree with the evidence of the participation of peptidergic fibers in thermoception and burning sensations and non-peptidergic in mechanical nociception [86].
A second clearly defined methodology to generate the expression of opsins that can be controlled by light, in non transgenic mice with no CRE dependent virus is the intraneural viral injection (adeno-associated virus serotype-6) express the step-function inhibitory ChR2 used by Iyer, et al. [83], a variant of the opsins that allows neurons to be activated with less frequent light pulses, thereby reducing the exposure traditionally required. Using SwiChR2, a variant that is permeable to Cl- or the chloride-conducting inhibitory channelrhodopsin (iC1C2) by AAV6-hSyn-biC1C2-TS-eYFP intraneural injection to generate hyperpolarization, the blue light stimulation produced large increases in mechanical withdrawal thresholds and thermal latency measures, because all fibers were transfected but mainly unmyelinated despite using a non-specific promoter such as hsyn-I. In the evaluation of the long-term inhibition, the sensitivity of mechanical withdrawal threshold and thermal latency decreased, and mice exhibited ‘post-light’ inhibition property (inhibition after light stimulus has disappeared). Furthermore, when the expression of SwiChR was evaluated in sensitized mice through the injection of formalin and subsequently stimulated with blue light on the plantar paw, a reduction in behavior was observed in phase 1, but not in phase II in formalin model [83].
To transfect halorhodopsin in unmyelinated cells of the primary afferents, an injection of AAV6-hSyn-eNpHR3.0-eYFP into the sciatic nerve was used to have halorhodopsin in unmyelinated fibers. When mechanical sensitivity was evaluated in mice, stimulation with yellow light generated an inhibition of nociceptors observed through an increase in the response threshold with stimulation with the Von Frey filament and an increase in latency to thermal stimulation. If the effect is analyzed in mice with neuropathic pain symptoms generated by the CCI in mice expressing halorhodopsin model, the optogenetic inhibition in these same fibers evoke a reversal of mechanical allodynia and an increase in thermal threshold [82]. With these results it is demonstrated that light delivered transdermally raises the hyperpolarization of unmyelinated fibers and decreases the activity of nociception.
AHTMR-neurons, known as high threshold mechanoreceptors nociceptors, are responsible for transmitting the initial signals to the spinal cord for pain perception. When optogenetically manipulated through the injection of (AAV8)/CAG-ArchT-GFP in spinal nerves L4-L5 it generates an increase in mechanical response thresholds in Sprague-Dawley rats with neuropathic pain through the SNL model, suggesting that this neuronal subpopulation contributes to the transmission of pain after damage to the nervous system [85]. Although the promoter and AAV8 are thought to be nonselective, the expression of ArchT was preferentially in fast-conducting high-threshold mechanoreceptors (AHTMR), mainly in neurons NF200+. Possibly this restriction of expression is due to the presence of specific glycoproteins on myelinated sensory neurons that allow AAV8 greater binding and possibly improved access to the cell.
5. Long-Term Chemogenic Modulation of Pain
Another strategy to modulate neural activity is through designer receptors exclusively activated by designer drugs (i.e., DREADDs), which are modified G coupled-protein receptors typically introduced into cells using viral vectors. One of the applications is the selective modulation using specific ligands (Salvinorin B or clozapine-N-Oxide) [87]. There are DREADDs that can modulate the neural activity through G couple-proteins, either to activate by using Gq/Gs, or to inactivate by using Gi proteins.
Iyer et al., [83] carried out a chemogenetic modulation on unmyelinated peripheral fibers with injection of AAV6-hSyn-HA-hM4D(Gi)-IRES-mCitrine and subsequent intraperitoneal administration of clozapine-N-oxide increasing mechanical withdrawal thresholds and thermal sensation in mice expressing the hM4D receptor. The authors reported the expression of mCitrine in small diameter neurons despite using the non-specific hsyn promoter.
As stated previously, the neurons involved in the transmission of pain from the periphery to the spinal cord are VGluT2. The approaches to modulate the activity of this neuronal subtype have been carried out by the generation of a transgenic model by cross breeding VlguT2-Cre mice with tdTomatoFlox / flox mice (Vglut2::Cre) and for chemogenetic modulation, with the injection of the viral construct AAV8- hSyn-DIO-hM3Dq-mCherry or AAV8-hSyn-DIO-HA-KORD-IRES-mCitrine in spinal dorsal horn neurons [81]. The activation of the Gq-coupled human M3 muscarinic receptor DREADD (hM3D) with intraperitoneally administration of clozapine-N-Oxide activates VGluT2-expressing glutamatergic dorsal horn neurons reduce the mechanical and thermal withdrawal threshold eliciting spontaneous flinching and licking. Therefore, there is an increase in nociceptive activity. Moreover, a wide decrease in the mechanical response threshold was observed, but not in thermal stimulation in Vglut2-Cre mice with spared nerve injury and KORD-mediated inhibition induced by Salbinorin B [81]. This indicates that VGluT2-expressing dorsal horn neurons are critically involved in maintaining pain hypersensitivity.
6. Conclusions
In chronic neuropathic pain there are situations where pharmacological treatments are not completely effective, so the search for promising strategies is an impending need. With current technological advances, it is possible to modulate the activity of pain-processing neurons at an experimental level. This modulation could be fast by using photosensitive ionotropic proteins with optogenetics, or slow, with metabotropic receptors with DREADDs. Optogenetics is a promising tool, however, the activation/inactivation of neuronal subgroups that participate in pain transmission can still be made more specific by using more than one promoter. It is well-described that there are specialized mechanisms that trigger the appearance of painful behaviors, such as allodynia or hyperalgesia. An important question yet to be answered is what neuronal circuit participates in either allodynia or hyperalgesia. To date, there are no studies using specific promoters directed to a particular neuronal population that could be important to delineate the pathophysiological processes mediating chronic neuropathic pain.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication Funding. JJ-AB, JV-EJ wrote the manuscript draft, and JJ-AB, JV-EJ, E-C, and B-E reviewed the manuscript, participated in editing, and provided suggestions for revision. Additionally, JJ-AB, JV-EJ, E-C, and B-E made revisions for the final version of the manuscript.
Funding
Please add: “This research received no external funding”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This is a short text to acknowledge the contributions of specific colleagues, institutions, or agencies that aided the efforts of the authors. Fundación Miguel Aleman.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Campbell, J. N., and Meyer, R. A. (2006). Review Mechanisms of Neuropathic Pain. Neuron 52, 77–92. [CrossRef]
- Treede, R.D. , Jensen, T.S., Campbell, J.N., Cruccu, G., Dostrovsky, J.O., Griffin, J.W., Hansson, P., Hughes, R., Nurmikko, T., Serra, J. (2008). Neuropathic pain: redefinition and a grading system for clinical and research purposes. Neurology. 70, 1630-1635. [CrossRef]
- Basbaum, A. I., Bautista, D. M., Scherrer, G., and Julius, D. (2009). Cellular and Molecular Mechanisms of Pain. Cell. 139, 267-284. [CrossRef]
- Zhang, J. M. , Song, X. J., and LaMotte, R. H. (1997). An in vitro study of ectopic discharge generation and adrenergic sensitivity in the intact, nerve-injured rat dorsal root ganglion. Pain. 72 51-57. [CrossRef]
- Ruscheweyh, R. , Forsthuber, L., Schoffnegger, D., and Sandkühler, J. (2007). Modification of classical neurochemical markers in identified primary afferent neurons with Aβ-, Aδ-, and C-fibers after chronic constriction injury in mice. J. Comp. Neurol. [CrossRef]
- Chiu, I.M., Barrett, L.B., Williams, E.K., Strochlic, D.E., Lee, S., Weyer, A.D., Lou, S., Bryman, G.S., Roberson, D.P., Ghasemlou, N., Piccoli, C., Ahat, E., Wang, V., Cobos, E.J., Stucky, C.L., Ma. Q., Liberles, S.D., Woolf, C.J. (2014). Transcriptional profiling at whole population and single cell levels reveals somatosensory neuron molecular diversity. Elife. 3, e04660. [CrossRef]
- Medici, T., and Shortland, P. J. (2015). Effects of peripheral nerve injury on parvalbumin expression in adult rat dorsal root ganglion neurons. BMC Neurosci. 16, 1–10. [CrossRef]
- Taylor-Clark, T. E. , Wu, K. Y., Thompson, J. A., Yang, K., Bahia, P. K., and Ajmo, J. M. (2015). Thy1.2 YFP-16 transgenic mouse labels a subset of large-diameter sensory neurons that lack TRPV1 expression. PLoS One. 10, 1–16. [CrossRef]
- Crawford, L. T. K. , and Caterina, M. J. (2020). Functional Anatomy of the Sensory Nervous System: Updates From the Neuroscience Bench. Toxicol. Pathol. 48, 174–189. [CrossRef]
- Amaya, F. , Decosterd, I., Samad, T. A., Plumpton, C., Tate, S., Mannion, R. J., et al. (2000). Diversity of expression of the sensory neuron-specific TTX-resistant voltage-gated sodium ion channels SNS and SNS2. Mol. Cell. Neurosci. 15, 331–342. [CrossRef]
- Smith, T. , Al Otaibi, M., Sathish, J., and Djouhri, L. (2015). Increased expression of HCN2 channel protein in L4 dorsal root ganglion neurons following axotomy of L5- and inflammation of L4-spinal nerves in rats. Neuroscience 295, 90–102. [CrossRef]
- Usoskin, D. , Furlan, A., Islam, S., Abdo, H., Lönnerberg, P., Lou, D., Hjerling-Leffler, J., Haeggström, J., Kharchenko, O., Kharchenko, P.V., Linnarsson, S., Ernfors, P. (2015). Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 18, 145–153. [CrossRef]
- Acosta, C. , McMullan, S., Djouhri, L., Gao, L., Watkins, R., Berry, C., Dempsey, K., Lawson, S.N. (2012). HCN1 and HCN2 in Rat DRG neurons: levels in nociceptors and non-nociceptors, NT3-dependence and influence of CFA-induced skin inflammation on HCN2 and NT3 expression. PLoS One. 7, e50442. [CrossRef]
- Levanon, D. , Bettoun, D., Harris-Cerruti, C., Woolf, E., Negreanu, V., Eilam, R., Bernstein, Y., Goldenberg, D., Xiao, C., Fliegauf, M., Kremer, E., Otto, F., Brenner, O., Lev-Tov, A., Groner, Y. (2002). The Runx3 transcription factor regulates development and survival of TrkC dorsal root ganglia neurons. EMBO J. 21, 3454-3463. [CrossRef]
- Nakamura, S. , Senzaki, K., Yoshikawa, M., Nishimura, M., Inoue, K. I., Ito, Y., Ozaki, S., Shiga, T. (2008). Dynamic regulation of the expression of neurotrophin receptors by Runx3. 135, 1703-1711. Development. [CrossRef]
- de Nooij, J. C., Doobar, S., and Jessell, T. M. (2013). Etv1 Inactivation Reveals Proprioceptor Subclasses that Reflect the Level of NT3 Expression in Muscle Targets. Neuron 77, 1055–1068. [CrossRef]
- Yoshikawa, M., Senzaki, K., Yokomizo, T., Takahashi, S., Ozaki, S., and Shiga, T. (2007). Runx1 selectively regulates cell fate specification and axonal projections of dorsal root ganglion neurons. Dev. Biol. 303, 663–674. [CrossRef]
- Hu, J. , Huang, T., Li, T., Guo, Z., and Cheng, L. (2012). C-Maf is required for the development of dorsal horn laminae III/IV neurons and mechanoreceptive DRG axon projections. J. Neurosci. 32, 5362–5373. [CrossRef]
- Li, B., Yang, X., Qian, F., Tang, M., and Ma, C. (2015). A novel analgesic approach to optogenetically and speci fi cally inhibit pain transmission using TRPV1 promoter. Brain Res. 1609, 12–20. [CrossRef]
- Costigan, M. , and Woolf, C. J. (2000). Pain: molecular mechanisms. J Pain 1, 35–44. [CrossRef]
- Brumovsky, P. , Watanabe, M., and Hökfelt, T. (2007). Expression of the vesicular glutamate transporters-1 and -2 in adult mouse dorsal root ganglia and spinal cord and their regulation by nerve injury. Neuroscience 147, 469–490. [CrossRef]
- Lagerström, M. C. , Rogoz, K., Abrahamsen, B., Persson, E., Reinius, B., Nordenankar, K., Olund, C., Smith, C., Mendez, J.A., Chen, Z.F., Wood, J.N., Wallén-Mackenzie, A., Kullander, K. (2010). VGLUT2-Dependent Sensory Neurons in the TRPV1 Population Regulate Pain and Itch. Neuron. [CrossRef]
- Beaudry, H. , Daou, I., Ase, A. R., Ribeiro-Da-Silva, A., and Séguela, P. (2017). Distinct behavioral responses evoked by selective optogenetic stimulation of the major TRPV1+ and MrgD+ subsets of C-fibers. Pain 158, 2329–2339. [CrossRef]
- Duan, B. , Cheng, L., Bourane, S., Britz, O., Padilla, C., Garcia-Campmany, L., Krashes, M., Knowlton, W., Velasquez, T., Ren, X., Ross, S., Lowell, B.B., Wang, Y., Goulding, M., Ma. Q. (2014). Identification of spinal circuits transmitting and gating mechanical pain. Cell. 59(6):1417-1432. [CrossRef]
- Häring, M. , Zeisel, A., Hochgerner, H., Rinwa, P., Jakobsson, J.E.T., Lönnerberg, P., La Manno, G., Sharma, N., Borgius, L., Kiehn, O., Lagerström, M.C., Linnarsson, S., Ernfors, P. (2018). Neuronal atlas of the dorsal horn defines its architecture and links sensory input to transcriptional cell types. Nat Neuroscience, 21(6):869-880. [CrossRef]
- Jiang, Y.Q., Sun, Q., Tu, H.Y., Wan, Y. (2008) Characteristics of HCN channels and their participation in neuropathic pain. Neurochem Research. 33, 1979-1989. [CrossRef]
- Kobayashi, K. , Fukuoka, T., Obata, K., Yamanaka, H., Dai, Y., Tokunaga, A., Noguchi, K. (2005) Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptors. J Comp Neurol. 26;493(4):596-606. [CrossRef]
- Patil, M.J. , Hovhannisyan, A.H., Akopian, A.N. (2018) Characteristics of sensory neuronal groups in CGRP-cre-ER reporter mice: Comparison to Nav1.8-cre, TRPV1-cre and TRPV1-GFP mouse lines. PLoS One. 13(6):e0198601. [CrossRef]
- Peirs, C. , Williams, S.P., Zhao, X., Walsh, C.E., Gedeon, J.Y., Cagle, N.E., Goldring, A.C., Hioki, H., Liu, Z., Marell, P.S., Seal, R.P. (2015). Dorsal Horn Circuits for Persistent Mechanical Pain. Neuron. 87(4):797-812. [CrossRef]
- Scherrer, G., Imamachi, N., Cao, Y. Q., Contet, C., Mennicken, F., O’Donnell, D., Kieffer, B.L., Basbaum, A.I. (2009). Dissociation of the opioid receptor mechanisms that control mechanical and heat pain. Cell 137, 1148–1159. [CrossRef]
- Scherrer, G. , Low, S.A., Wang, X., Zhang, J., Yamanaka, H., Urban, R., Solorzano, C., Harper, B., Hnasko, T.S., Edwards, R.H., Basbaum. A.I. (2010). VGLUT2 expression in primary afferent neurons is essential for normal acute pain and injury-induced heat hypersensitivity. Proc Natl Acad Sci U S A. 107, 22296-22301. [CrossRef]
- Wright, D.E. , Snider, W.D. (1995). Neurotrophin receptor mRNA expression defines distinct populations of neurons in rat dorsal root ganglia. J Comp Neurol. 351(3):329-38. [CrossRef]
- Bardoni, R., Tawfik, V.L., Wang, D., François, A., Solorzano, C., Shuster, S.A., Choudhury, P., Betelli, C., Cassidy, C., Smith, K., de Nooij, J.C., Mennicken, F., O'Donnell, D., Kieffer, B.L., Woodbury, C.J., Basbaum, A.I., MacDermott, A.B., Scherrer, G. (1014). Delta opioid receptors presynaptically regulate cutaneous mechanosensory neuron input to the spinal cord dorsal horn. Neuron. 81, 1312-1327. [CrossRef]
- Heinke, B. , Gingl, E., Sandkühler, J. (2011). Multiple targets of μ-opioid receptor-mediated presynaptic inhibition at primary afferent Aδ- and C-fibers. J Neurosci. 31, 1313-1322. [CrossRef]
- Li, L. , Rutlin, M., Abraira, V. E., Cassidy, C., Kus, L., Gong, S., Jankowski, M.P., Luo, W., Heintz, N., Koerber, H.R., Woodbury, C.J., Ginty, D.D. (2011). The functional organization of cutaneous low-threshold mechanosensory neurons. Cell. 1615. [Google Scholar] [CrossRef]
- Abrahamsen, B. , Zhao, J., Asante, C.O., Cendan, C.M., Marsh, S., Martinez-Barbera, J.P., Nassar, M.A., Dickenson, A.H., Wood, J.N. (2008). The cell and molecular basis of mechanical, cold, and inflammatory pain. Science. 321, 702-705. [CrossRef]
- Acton, D. , Ren, X., Di, Costanzo, S., Dalet, A., Bourane, S., Bertocchi, I., Eva, C., Goulding, M. (2019). Spinal neuropeptide Y1 receptor-expressing neurons form an essential excitatory pathway for mechanical itch. Cell Rep. 28, 625–639. [CrossRef]
- Brumovsky, P., Stanic, D., Shuster, S., Herzog, H., Villar, M., Hökfelt, T. (2005). Neuropeptide Y2 receptor protein is present in peptidergic and nonpeptidergic primary sensory neurons of the mouse. J Comp Neurol. 489, 328-48.
- Daou, I. , Beaudry, H., Ase, A. R., Wieskopf, J. S., Ribeiro-da-Silva, A., Mogil, J. S., Séguela, P. (2016). Optogenetic silencing of Nav1.8-positive afferents alleviates inflammatory and neuropathic pain. eNeuro 3, 702–705. [CrossRef]
- Szabo NE, da Silva RV, Sotocinal SG, Zeilhofer HU, Mogil JS, Kania A. Hoxb8 intersection defines a role for Lmx1b in excitatory dorsal horn neuron development, spinofugal connectivity, and nociception. J Neurosci. 2015 Apr 1;35(13):5233-46.
- Takasaki, I. , Watanabe, A., Yokai, M., Watanabe, Y., Hayakawa, D., Nagashima, R., Fukuchi, M., Okada, T., Toyooka, N., Miyata, A., Gouda, H., Kurihara, T. (2018). In silico screening identified novel small-molecule antagonists of PAC1 receptors. J. Pharmacol. Exp. Ther. 365, 1–8. [CrossRef]
- Wende, H. , Lechner, S.G., Cheret, C., Bourane, S., Kolanczyk, M.E., Pattyn, A., Reuter, K., Munier, F.L., Carroll, P., Lewin, G.R., Birchmeier, C. (2012). The transcription factor c-Maf controls touch receptor development and function. Science. 335, 1373-1376. [CrossRef]
- Xiang, C.X., Zhang, K.H., Johnson, R.L., Jacquin, M.F., Chen, Z.F. (2012). The transcription factor, Lmx1b, promotes a neuronal glutamate phenotype and suppresses a GABA one in the embryonic trigeminal brainstem complex. Somatosens Mot Res. 29, 1-12. [CrossRef]
- Xu, Y., Lopes, C., Wende, H., Guo, Z., Cheng, L., Birchmeier, C., Ma, Q. (2013). Ontogeny of excitatory spinal neurons processing distinct somatic sensory modalities. J Neurosci. 33, 14738-14748. [CrossRef]
- Draxler, P. , Honsek, S.D., Forsthuber, L., Hadschieff, V., Sandkühler, J. (2014). VGluT3⁺ primary afferents play distinct roles in mechanical and cold hypersensitivity depending on pain etiology. J Neurosci. 34, 12015-12028. [CrossRef]
- Duitama, M. , Vargas-López, V., Casas, Z., Albarracin, S.L., Sutachan, J.J., Torres, Y.P. TRP Channels Role in Pain Associated With Neurodegenerative Diseases. Frontiers in Neuroscience. 2020 Aug 4;14:782. [CrossRef]
- Collo, G. , North, R.A., Kawashima, E., Merlo-Pich, E., Neidhart, S., Surprenant, A., Buell, G. (1996). Cloning of P2X5 and P2X6 receptors and the distribution and properties of an extended family of ATP-gated ion channels. J Neurosci. 16, 2495–2507. [CrossRef]
- Teixeira, J.M. , Bobinski, F., Parada, C.A., Sluka, K.A., Tambeli, C,H. (2017) P2X3 and P2X2/3 receptors play a crucial role in articular hyperalgesia development through infammatory mechanisms in the knee joint experimental synovitis. Mol Neurobiol. 54, 6174-6186. [CrossRef]
- Rau, K. K., Caudle, R. M., Cooper, B. Y., and Johnson, R. D. (2005). Diverse immunocytochemical expression of opioid receptors in electrophysiologically defined cells of rat dorsal root ganglia. J. Chem. Neuroanat. 29, 255–264. [CrossRef]
- Wang, D. , Tawfik, V. L., Corder, G., Low, S. A., François, A., Basbaum, A. I., Scherrer, G. (2018). Functional Divergence of Delta and Mu Opioid Receptor Organization in CNS Pain Circuits. Neuron 98, 90-108.e5. [CrossRef]
- Zhang, X.Y. , Dou, Y.N., Yuan, L., Li, Q., Zhu, Y.J., Wang, M., Sun, Y.G. (2020). Different neuronal populations mediate inflammatory pain analgesia by exogenous and endogenous opioids. Elife. 9:e55289. [CrossRef]
- Tamae, A. , Nakatsuka, T., Koga, K., Kato, G., Furue, H., Katafuchi, T., Yoshimura, M. (2005). Direct inhibition of substantia gelatinosa neurones in the rat spinal cord by activation of dopamine D2-like receptors. J. Physiol. 568, 243–253. [CrossRef]
- Taniguchi, W. , Nakatsuka, T., Miyazaki, N., Yamada, H., Takeda, D., Fujita, T., Kumamoto, E., Yoshida, M. (2011). In vivo patch-clamp analysis of dopaminergic antinociceptive actions on substantia gelatinosa neurons in the spinal cord. Pain 152, 95–105. [CrossRef]
- Lu, Y., Doroshenko, M., Lauzadis, J., Kanjiya, M. P., Rebecchi, M. J., Kaczocha, M., Puopolo, M. (2018). Presynaptic inhibition of primary nociceptive signals to dorsal horn lamina I neurons by dopamine. J. Neurosci. 38, 8809–8821. [CrossRef]
- Almanza, A. , Simón-Arceo, K., Coffeen, U., Fuentes-García, R., Contreras, B., Pellicer, F., et al. (2015). A D2-like receptor family agonist produces analgesia in mechanonociception but not in thermonociception at the spinal cord level in rats. Pharmacol. Biochem. Behav. 137, 119–125. [CrossRef]
- Mercado-Reyes, J., Almanza, A., Segura-Chama, P., Pellicer, F., and Mercado, F. (2019). D2-like receptor agonist synergizes the μ-opioid agonist spinal antinociception in nociceptive, inflammatory and neuropathic models of pain ni ht e art. Eur. .J Pharmacol. 853, 56–64. [CrossRef]
- Ohnou T, Yokai M, Kurihara T, Hasegawa-Moriyama M, Shimizu T, Inoue K, Kambe Y, Kanmura Y, Miyata A (2016). Pituitary adenylate cyclase-activating polypeptide type 1 receptor signaling evokes long-lasting nociceptive behaviors through the activation of spinal astrocytes in mice. J Pharmacol Sci. 130,194-203. [CrossRef]
- Yokai, M., Kurihara, T., and Miyata, A. (2016). Spinal astrocytic activation contributes to both induction and maintenance of pituitary adenylate cyclase-activating polypeptide type 1 receptor-induced longlasting mechanical allodynia in mice. Mol. Pain 12:1744806916646383. [CrossRef]
- Brumovsky, P. , Hygge-Blakeman, K., Villar, M. J., Watanabe, M., Wiesenfeld-Hallin, Z., and Hökfelt, T. (2006). Phenotyping of sensory and sympathetic ganglion neurons of a galanin-overexpressing mouse-Possible implications for pain processing. J. Chem. Neuroanat. 31, 243–262. [CrossRef]
- François, A. , Schüetter, N., Laffray, S., Sanguesa, J., Pizzoccaro, A., Dubel, S., Mantilleri, A., Nargeot, J., Noël, J., Wood, J.N., Moqrich, A., Pongs, O., Bourinet, E. (2015). The Low-Threshold Calcium Channel Cav3.2 Determines Low-Threshold Mechanoreceptor Function. Cell Rep. 10 370-382. [CrossRef]
- Seal, R. P. (2016). Do the distinct synaptic properties of VGLUTs shape pain?. Neurochem. Int. 98, 82–88. [CrossRef]
- Liu, X.J. , Salter, M.W. (2010) Glutamate receptor phosphorylation and trafficking in pain plasticity in spinal cord dorsal horn. Eur J Neurosci. 2010 Jul;32(2):278-89. [CrossRef]
- Todd, A.J., Hughes, D.I., Polgár, E., Nagy, G.G., Mackie, M., Ottersen, O.P., Maxwell, D.J (2003). The expression of vesicular glutamate transporters VGLUT1 and VGLUT2 in neurochemically defined axonal populations in the rat spinal cord with emphasis on the dorsal horn. The European Journal of Neuroscies. 17, 13-27. [CrossRef]
- Alvarez, F.J. , Villalba, R.M., Zerda, R., Schneider, S.P. (2004). Vesicular glutamate transporters in the spinal cord, with special reference to sensory primary afferent synapses. J Comp Neurol. 472(3):257-80. [CrossRef] [PubMed]
- Larsson, M. , and Broman, J. (2019). Synaptic Organization of VGLUT3 Expressing Low-Threshold Mechanosensitive C Fiber Terminals in the Rodent Spinal Cord. eNeuro 6. [CrossRef]
- Deuchars SA, Milligan CJ, Stornetta RL, Deuchars J. (2005). GABAergic neurons in the central region of the spinal cord: a novel substrate for sympathetic inhibition. J Neurosci. 25:1063-70. [CrossRef]
- Coull, J. A. , Boudreau, D., Bachand, K., Prescott, S. A., Nault, F., Sik, A., De Koninck, P., & De Koninck, Y. (2003). Trans-synaptic shift in anion gradient in spinal lamina I neurons as a mechanism of neurophatic pain Nature, 424, 938–942.
- Paul, J., Zeilhofer, H. U., & Fritschy, J. M. (2012). Selective distribution of GABAA receptor subtypes in mouse spinal dorsal horn neurons and primary afferents. Journal of Comparative Neurology, 520, 3895–3911. [CrossRef]
- Kahle, K. T. , Schmouth, J. F., Lavastre, V., Latremoliere, A., Zhang, J., Andrews, N., Omura, T., Laganière, J., Rochefort, D., Hince, P., Castonguay, G., Gaudet, R., Mapplebeck, J.C., Sotocinal, S.G., Duan, J., Ward, C., Khanna, A.R., Mogil, J.S., Dion, P.A., Woolf, C.J., Inquimbert, P., Rouleau, G.A. (2016). Inhibition of the kinase WNK1/HSN2 ameliorates neuropathic pain by restoring GABA inhibition. Sci. Signal. 9, ra32. [CrossRef]
- Lee, H. K., Ahmed, M. M., King, K. C., Miranpuri, G. S., Kahle, K. T., Resnick, D. K., Sun, D. (2014). Persistent phosphorylation of NKCC1 and WNK1 in the epicenter of the spinal cord following contusion injury. Spine J. 14, 777–781. [CrossRef]
- Tillman, L. , Zhang, J. Crossing the Chloride Channel: The Current and Potential Therapeutic Value of the Neuronal K+-Cl- Cotransporter KCC2. Biomed Res Int. 2019 ;2019:8941046. 21 May. [CrossRef]
- Deisseroth, K. , Feng, G., Majewska, A. K., Miesenböck, G., Ting, A., and Schnitzer, M. J. (2006). Next-Generation Optical Technologies for Illuminating Genetically Targeted Brain Circuits. J Neurosci. 26, 10380-10386. [CrossRef]
- Deisseroth, K. , and Hegemann, P. (2017). The form and function of channelrhodopsin. Science 15,357:6356. [CrossRef]
- Boyden, E. S. , Zhang, F., Bamberg, E., Nagel, G., and Deisseroth, K. (2005). Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 8, 1263–1268. [CrossRef]
- Chow, B. Y., Han, X., Dobry, A. S., Qian, X., Chuong, A. S., Li, M., Henninger, M.A., Belfort, G.M., Lin, Y., Monahan, P.E., Boyden, E.S. (2010). High-performance genetically targetable optical neural silencing by light-driven proton pumps. Nat. 2010 4637277 463, 98–102. [CrossRef]
- Han, X. , Chow, B. Y., Zhou, H., Klapoetke, N. C., Chuong, A., Rajimehr, R., Yang, A,. Baratta, M.V., Winkle, J., Desimone, R., Boyden, E.S. (2011). A high-light sensitivity optical neural silencer: Development and application to optogenetic control of non-human primate cortex. Front. Syst. Neurosci. 5, 18. [CrossRef]
- Watakabe, A., Sadakane, O., Hata, K., Ohtsuka, M., Takaji, M., Yamamori, T. (2017). Application of viral vectors to the study of neural connectivities and neural circuits in the marmoset brain. Dev Neurobiol. 77, 354-372. (. [CrossRef]
- Anastassiadis, K. , Fu, J., Patsch, C., Hu, S., Weidlich, S., Duerschke, K., Buchholz, F., Edenhofer, F., Stewart, A.F. (2009). Dre recombinase, like Cre, is a highly efficient site-specific recombinase in E. coli, mammalian cells and mice. Disease models & mechanisms. 9,508-515. [CrossRef]
- Daou, I., Tuttle, A. H., Longo, G., Wieskopf, J. S., Bonin, R. P., Ase, A. R., et al. (2013). Remote Optogenetic Activation and Sensitization of Pain Pathways in Freely Moving Mice. 33, 18631–18640. [CrossRef]
- Michoud, F., Seehus, C., Schönle, P., Brun, N., Taub, D., Zhang, Z., Jain, A., Furfaro, I., Akouissi, O., Moon, R., Meier, P., Galan, K., Doyle, B., Tetreault, M., Talbot, S., Browne, L.E., Huang, Q., Woolf, C.J., Lacour, S.P. (2021). Epineural optogenetic activation of nociceptors initiates and amplifies inflammation. Nat. Biotechnol. 39, 179–185. [CrossRef]
- Wang, L. , Chen, S. R., Ma, H., Chen, H., Hittelman, W. N., and Pan, H. L. (2018). Regulating nociceptive transmission by VGluT2-expressing spinal dorsal horn neurons. J. Neurochem. 147, 526–540. [CrossRef]
- Iyer, S. M. , Montgomery, K. L., Towne, C., Lee, S. Y., Ramakrishnan, C., Deisseroth, K., Delp, S.L. (2014). Virally mediated optogenetic excitation and inhibition of pain in freely moving nontransgenic mice. Nat. Biotechnol. 32, 274–278. [CrossRef]
- Iyer, S. M. , Vesuna, S., Ramakrishnan, C., Huynh, K., Young, S., Berndt, A., Lee, S.Y., Gorini, C.J., Deisseroth, K., Delp, S.L. (2016). Optogenetic and chemogenetic strategies for sustained inhibition of pain. Sci. Rep. 6, 1–10. [CrossRef]
- Tashima, R. , Koga, K., Sekine, M., Kanehisa, K., Kohro, Y., Tominaga, K., Matsushita, K., Tozaki-Saitoh, H., Fukazawa, Y., Inoue, K., Yawo, H., Furue, H., Tsuda, M. (2018). Optogenetic Activation of Non-Nociceptive Aβ Fibers Induces Neuropathic Pain-Like Sensory and Emotional Behaviors after Nerve Injury in Rats. eNeuro. 5, ENEURO.0450-17.2018.
- Boada, M. D. , Martin, T. J., Peters, C. M., Hayashida, K., Harris, M. H., Houle, T. T., et al. (2014). Fast-conducting mechanoreceptors contribute to withdrawal behavior in normal and nerve injured rats. Pain 155, 2646–2655. [CrossRef]
- Cavanaugh, D. J., Lee, H., Lo, L., Shields, S. D., Zylka, M. J., Basbaum, A. I., Anderson, D.J. (2009). Distinct subsets of unmyelinated primary sensory fibers mediate behavioral responses to noxious thermal and mechanical stimuli. Proc. Natl. Acad. Sci. U. S. A. 106, 9075–9080. [CrossRef]
- Armbruster, B. N. , Li, X., Pausch, M. H., Herlitze, S., and Roth, B. L. (2007). Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. 104, 5163–5168. [CrossRef]
Figure 1.
Main proteins expressed in peptidergic (left side) and non-peptidergic (right side) VgluT2 neurons, there are molecular markers for their differentiation as CGRP, SP and TRPV1 in peptidergic neurons and in the other hand MrgprD and IB4 in non peptidergic. (Image developed from table -1, C-fiber section).
Figure 1.
Main proteins expressed in peptidergic (left side) and non-peptidergic (right side) VgluT2 neurons, there are molecular markers for their differentiation as CGRP, SP and TRPV1 in peptidergic neurons and in the other hand MrgprD and IB4 in non peptidergic. (Image developed from table -1, C-fiber section).
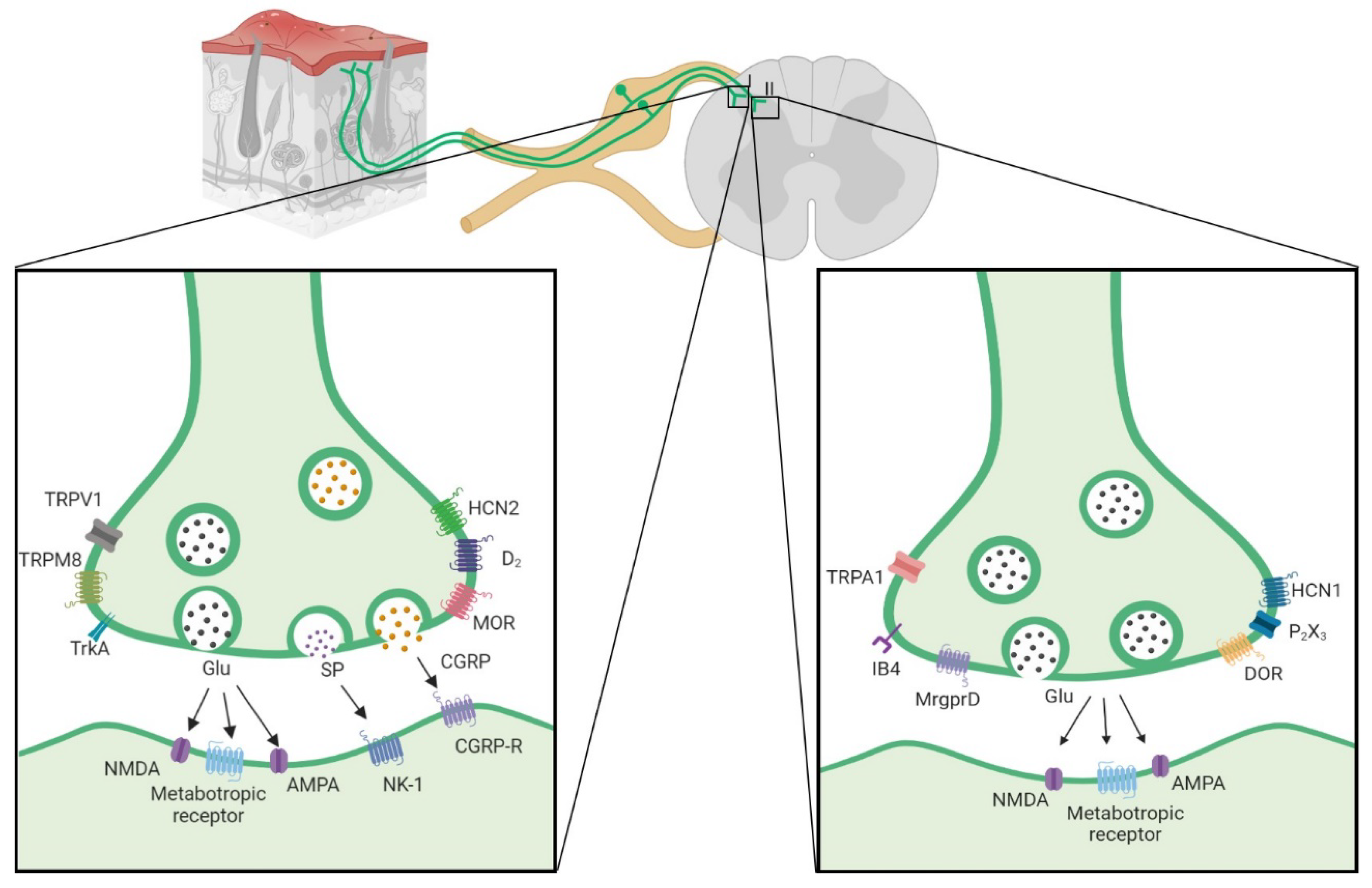
Figure 2.
GABA impairment in neuropathic pain is caused by shift in chloride concentration. (a) Normal NKCC1 and KCC2 activity maintains a lower concentration inside, allowing chloride to enter the cell and hyperpolarizing when GABA is activated; (b) in neuropathic pain, phosphorylation of NKCC1 and KCC2 inverts the concentration of chloride, and when GABA is activated, there is an efflux of chloride depolarizing the neuron. (Image developed from [67-71]).
Figure 2.
GABA impairment in neuropathic pain is caused by shift in chloride concentration. (a) Normal NKCC1 and KCC2 activity maintains a lower concentration inside, allowing chloride to enter the cell and hyperpolarizing when GABA is activated; (b) in neuropathic pain, phosphorylation of NKCC1 and KCC2 inverts the concentration of chloride, and when GABA is activated, there is an efflux of chloride depolarizing the neuron. (Image developed from [67-71]).
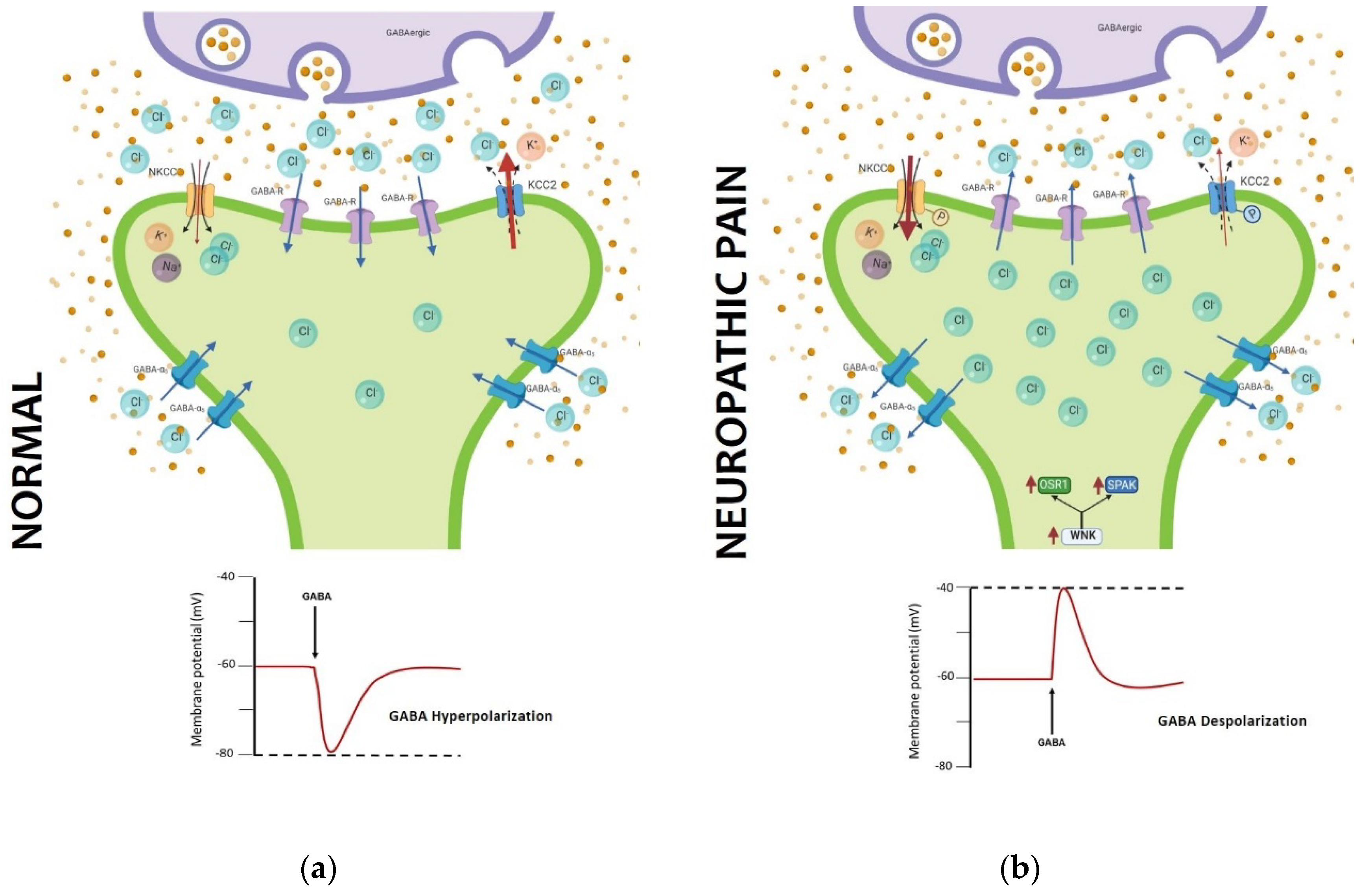

Table 1.
Main markers of primary afferent fibers.
| Fiber Type | Conduction velocity | Vesiculator type | Protein Markers | Channels | Membrane receptors | Dorsal Laminar distribution | Transcription factor | References | |
|---|---|---|---|---|---|---|---|---|---|
| Aβ | 30-80 m/s | Vglut 1 | NF200 Thy-1 parvalbumin Caderina SPP1 NEFH |
Nav1.1 Nav1.6 HCN3 ASIC1 Nav1.8 |
DOR TrkC |
III, IV, V | RunX3 ETV1 Etv4 Cdh12 |
[4-8,11,12,24-32] | |
| Aδ | 5-25 m/s | Vglut 2 | NF200 NEFH CGRP Substance P |
TRPM8 Nav 1.8 HCN1 HCN3 Cav 3.2 |
TrkA TRKB MOR DOR |
I, II | CNTNAP2 FAM19A RET |
[5,11,12,24- 29,31-35] |
|
| C | Peptidergic | 0.5-2 m/s | Vglut 2 | Substance P CGRP Cdk 5mTOR |
TRPV1 TRPA1 TRPM8 Nav1.7 Nav1.8 HCN2 HCN3 |
D2 MOR MrgprA3 mGlu2 Ntrk1 PAC1 TrkA Y1R Y2R |
I | cMAF GDNF HOB8 LMBX1 RET TAC1 |
[5,6,21-26,29,30,36-44] |
| Non-peptidergiC | 0.5-2 m/s | Vglut 2 | IB4 Cdk 5mTOR |
TRPA1 TRPM3 TRPC3 Nav1.7 Nav1.8 Nav1.9 HCN1 HCN3 P2X3 |
DOR MrgprA3 MrgprB4 MrgprD Y1R Y2R |
II | GfrA1 LMBX1 NGF PLXNC1 RET RunX |
[5,6,11,21,23,24,25, 28- 30,35,36-38,43-45] |
|
| LTMR | <0.5 m/s | Vglut 3 | TH | TRPM8 Nav 1.8 Nav 1.9 |
TrkB | I, II, III | GfrA2 PIEZO2 RET |
[12,25,28,29,35,45] | |
Table 2.
Modulation of sensory pathways with optogenetic and chemogenetic.
| Specificity strategy | Construct | Fiber and Cell type | Injection site | Place of stimulation | Behavioral phenotype | Painful condition | Reference | |
| Cre dependent |
Nav1.8 ::Cre / Ai32 ( carry the ChR2(H134R)–EYFP in Gt(ROSA)26Sor locus ) Product: Nav1.8–ChR2+ |
Aβ Fiber Aδ Fiber C- fibers: -Peptidergic and -nonpeptidergic |
NA | The plantar surface of the hindpaw blue light (473 nm) |
paw withdrawal and paw licking | Not sensitized |
[79] | |
|
TRPV1::Cre / AAV5-TRPV1-ArchT-eGFP Product: TRPV1-Arch+ |
C-fiber Peptidergic |
DRG injection | The plantar surface of the hindpaw Green light (532 nm) |
Increases in mechanical withdrawal threshold and thermal latency | [19] | |||
|
TRPV1::Cre / AAV2/8-CAG-floxed stop-ChR2[H134R]-tdTomato-WPRE Product: TRPV1-ChR2 |
C-fiber Peptidergic |
Intrathecal | The plantar surface of the hindpaw Blue light (473 nm) |
Increases in paw withdrawal, paw lifting and paw licking | [23] | |||
|
TRPV1::Cre/Ai27D (carry Rosa-CAG-LSL-hChR2(H134R)-tdTomato-WPRE) Product: TRPV1-ChR2 |
C-fiber Peptidergic |
NA | Epineural in sciatic nerve Blue light (470 nm) |
Thermal and mechanical sensitivity | [80] | |||
|
MrgD::CreERT2 / Ai32 (carry in the ROSA26 locus the floxed stop-ChR2(H134R)-EYFP) Product: MrgD-ChR2 (Opsin induced by tamoxiofen) |
C-fiber Nonpeptidergic |
NA | The plantar surface of the hindpaw Blue light (473 nm) |
Paw withdrawal and lifting. | [23] | |||
|
VGluT2::Cre / AAV8-hSyn-DIO-hM3Dq-mCherry Product: VGluT2- hM3D-Gq |
C-fiber: -Peptidergic and –nonpeptidergic (Vglut2-dorsal horn) |
Dorsal horn of the spinal cord | clozapine N-oxide intraperitoneally | Increases in mechanical and thermal sensitivities | [81] | |||
| No Cre dependent | AAV6-hSyn-ChR2-eYFP Product: Chr2-eYFP |
C- fiber: -Unmyelinated primary afferent |
intrasciatic | The plantar Surface Blue light (473 nm) |
Decreases in paw withdrawal Place aversion |
[82] |
||
| AAV6-hSyn-SwiChR-eYFP Product: SwiChR-eYFP |
C- fiber: -Unmyelinated primary afferent |
intraneural | The plantar surface of the hindpaw Blue light (473 nm) |
Increases in mechanical withdrawal threshold and thermal withdrawal latency | [83] | |||
| AAV6-hSyn-biC1C2-TS-eYFP Product: iC1C2-eYFP |
C- fiber: -Unmyelinated primary afferent |
intraneural | The plantar surface of the hindpaw Blue light (473 nm) |
Increases in mechanical withdrawal threshold and thermal withdrawal latency | [83] |
|||
| AAV6-hSyn-eNpHR3.0-eYFP |
C- fiber: -Unmyelinated primary afferent |
Intrasciatic | The plantar Surface Yellow light (590 nm) |
Increases in mechanical withdrawal threshold and thermal withdrawal latency | [82] |
|||
| AAV6-hSyn-HA-HM4D(Gi)-IRES-mCitrine Product:hM4D-Gi |
C- fiber: -Unmyelinated primary afferent |
Intraneural | Clozapine-N-oxide intraperitoneally |
Increases in mechanical withdrawal threshold and thermal withdrawal latency | [83] |
|||
| Cre-dependetn |
VGluT3::Cre/Ai32 carry ChR2 Product: VGluT3-ChR2 |
LTMR Fiber: VGluT3+ |
NA | The plantar surface of the hindpaw Blue light (470 nm) |
Elicit nociceptive behaviour |
sensitized |
Oxaliplatin-induced neuropathy | [45] |
|
Nav1.8::Cre / Ai35 (carry the floxed stop-Arch-EGFP gene in the ROSA26 locus) Product: Nav1.8-Arch+ |
C-fiber Peptidergic |
NA | The plantar surface of the hindpaw yellow light |
Decrease in mechanical allodynia |
Spared nerve injury | [39] | ||
|
VGluT2::Cre / AAV8-hSyn-DIO-HA-KORD-IRES-mCitrine Product: VGluT2-KORD-Gi |
Peptidergic and nonpeptidergic (Vglut2-dorsal horn) |
Dorsal horn of the spinal cord | Salvinorin B intraperitoneal | Decrease in tactile allodynia / Decrease in pain hipersensibility |
SNI / CFA | [81] | ||
| Thy1-COP4/YFP / W-TChR2V4 | Aβ fibers | NA | The plantar surface of the hindpaw Yellow light |
Lifting and flinching behaviors. | PNI | [84] | ||
| No Cre-dependetn |
AAV6-hSyn-SwiChR-eYFP Product: SwiChR-eYFP |
C-fiber: Unmyelinated primary afferent |
intrasciatic | The plantar surface of the hindpaw Blue light (473 nm) |
Reduced pain behavior in phase I | Formalin test | [83] | |
| AAV6-hSyn-eNpHR3.0-eYFP Product: eNpHR-eYFP |
C-fiber unmyelinated primary afferent |
intraneural | The plantar surface of the hindpaw Yellow light (490 nm) |
Reduction of mechanical allodynia and thermal hyperalgesia | CCI | [82] | ||
| AAV8/CAG-ArchT-GFP In myelinated neurons. Product: ArchT-GFP |
AHTMR-fiber myelinated primary afferent |
intrathecal | The plantar surface of the hindpaw Green light |
Increase in paw withdrawal threshold | SNL | [85] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



