
Submitted:
02 January 2023
Posted:
06 January 2023
You are already at the latest version
Abstract
The genus Musa spp. contains commercially important fleshy fruit producing plants including plantains and bananas with a strong potential of providing food security and source of revenue to farmers. Occasionally, plantlets are often not available to fulfill the supply demand of farmers particularly in Caribbean region. Concerns with the quality of vegetative tissues along with the possibility of the transmission of phytopathogens makes availability of plantlets limited to farm-ers. Micropropagation of plantains offers an alternative to producing large number of in-vitro plantlets. However, conventional methods of micropropagation techniques require high produc-tion costs and are labor-intensive. Recently, Temporary Immersion Bioreactor (TIB) has emerged as an alternative to conventional micropropagation methods. Our work utilized SEM (Scanning Electron Microscope), molecular and biochemical tools (qRT-PCR and ICP-OES) to characterize and compare the morphological, elemental composition, and photosynthetic gene expression of plantains cultured on TIB. Additionally, morphological features of growth and propagation rates were analyzed for comparing outputs obtained from temporary immersion bioreactor with conventional micropropagation (CM) techniques. Results showed higher growth and multiplication rates for plantlets cultivated in TIB. Gene expression analysis of selected photosynthetic genes demonstrated high transcript abundance of phosphoenol pyruvate carboxylase (PEPC) in plantain tissues obtained by TIB. Elemental composition analysis showed higher content of iron in plantains grown in TIB suggesting a potential correlation with PEPC expression. These results demonstrate the potential of TIB to be an efficient method to produce healthy in-vitro plantains.
Keywords:
Temporary Immersion Bioreactor
; Micropropagation
; Plantain
; plant tissue culture
1. Introduction
Musa spp. is a plant genus of very valuable horticultural crops because of the rich nutritional components these possess and includes bananas and plantains [1]. Musa spp. is native to Asian regions, and nowadays these can also be found throughout the subtropical and other tropical regions [2]. The genus Musa is one of the major food crops produced all over the world including ~130 countries [3]. Though in Puerto Rico plantain crops lead the category of agricultural production with 55.2%, these crops were significative impacted by hurricanes Maria in 2017 and Fiona (2022) [4]. These environmental events led to shortage of plantains in Puerto Rico which affected the availability of fleshy fruit in the region. Identifying food insecurity factors and associated risks can help to improve programs and enhance techniques to fight food insecurity both at local and global levels [5]. Since plantain have a very high demand in the Caribbean region, high dependence on imported fleshy fruits have been affecting the local farmers [6]. The production and the availability of propagating new plantlets are limited by the lack of starting plant material [7]. Another concern for farmers is the transmission of lethal pests including, viruses and fungi (for example, Black Sigatoka disease) using the conventional field–grown shoots in their farms. The limited availability of starting plant material and challenges associated with pest prevalence has increased the interest in plant micropropagation [7]. Plant tissue culture provides a solution for mass-propagation of disease-free plantlets in a short period under laboratory conditions [3]. Conventional micropropagation of Musa spp. demands high production costs that eventually may limit its commercial use [7]. The significance and prominence of the plant's biotechnological advances rely on the search for novel methods to improve crop production in plants [8]. For example, replacing solid plant tissue culture media with liquid plant tissue culture media has provided a solution for automation and decreased the production budget in commercial plant tissue culture facilities [7,8]. Temporary Immersion Bioreactor (TIB) allows temporary immersion of the explants in the liquid media [9]. Also, the positive and consistent effects of TIB on in-vitro shoot regeneration have been proved for a variety of plants, indicating TIB as a promising new technique for in-vitro plantain propagation [7]. TIB system promotes certain advantages which allows the normal development of the plantlets [10]. Also, TIB promotes optimal conditions for nutrient acquisition and humidity with minimum liquid contact [11]. Additionally, enhanced oxygen transport helps to develop a better gas exchange by reducing oxygen limitation. The controlled environmental conditions provided by TIB system protect the tissues which preserves the tissue integrity and improves the morphology and physiology of the organs [12]. This method of automated propagation has provided a possible solution for reducing conventional micropropagation costs [13]. Additionally, TIB has been functional and successful in several valuable crops such as banana [14], pineapple [10], and plantain [7]. Further, TIB cultivated plants have demonstrated increased plant growth with significant increase in the yield [15]. Studying the developmental stages of plantain in a temporary immersion bioreactor environment is essential for evaluating plant physiology at a molecular and biochemical level. Analyzing photosynthetic gene expression may help us understand how TIB environment can alter plants response during growth.
Rubisco is a multifunctional gene that catalyzes both reactions: carboxylation and oxygenation, where carboxylation is the first stage in which CO2 fixation occurs [16,17], and is a rate-limiting factor of photosynthesis [18]. Previous reports have suggested that the availability of the small subunit Rubisco enzyme upregulates the transcripts levels of the larger subunits of Rubisco enzyme [19]. These small subunits of Rubisco enzymes promote conformational changes in the other subunits resulting in the improvement of catalytic rate [19]. Additionally, on C4 plant species, the phosphoenol-pyruvate carboxylase catalyzes the first carboxylation reaction of photosynthesis in the mesophyll cells [20]. Also, phosphoenolpyruvate carboxylase (PEPC) plays a key role in C4 photosynthesis since it is involved in a variety of mechanisms such as anaplerotic metabolism, stomatal opening, and pH regulation [21]. However, during micropropagation, PEPC is responsible for the mobilization of sugars through an anaplerotic route to guarantee the supply of carbon skeletons for amino acid synthesis [22]. Besides, it has been documented [23] that in temporary immersion bioreactors systems PEPC activity is higher and it is suggested to be because the TIB system mimics outdoors conditions leading to high activity of PEPC enzyme and photosynthetic rate [23]. Additionally, examining the elemental composition could reveal metal ions that potentially act as cofactors during the expression of the interesting genes. Subsequently, in plant biology, metals possess certain functions that help molecular and biosynthesis pathways act as cofactors. For example, metals such as iron, zinc, manganese, and copper are very important and vital for organisms in this case plants genus; this is because of its numerous biological actions that are essential for the host to grow [24]. Past research has revealed that iron and copper are found in high quantities in the chloroplast of plants [24]. This high content of transition metals on the plant chloroplast is because of the metalloproteins that strictly require the interaction of these metals in the photosynthetic electron transport chain [25,26,27]. To the best of our knowledge, there is no knowledge on Rubisco small units and PEPC expression on in-vitro plantains in general as well as no biochemical characterization that can help to elucidate how metal ions can potentially interact with photosynthetic genes expression in tissues originating in TIB.
We characterized the growth of plantain in TIB at a molecular, biochemical, and morphological level to consider as an alternative and cost-effective method of producing plantlets under laboratory conditions. This will provide a viable model for the mass production of plantain for farmers and reduce the challenges of food availability in future. The implementation of micropropagation of plants using bioreactors could help to establish an alternate protocol allowing plantain to be cultured at a mass scale.
2. Materials and Methods
2.1. Plant Material
Explant selection was based on traits with commercial value, then explants were disinfected and cultured in a controlled environment. Selected plant materials were propagated using the method as described in Aragón et al. [22] with some minor modifications. Conventional seeds (also known as corm) of Musa maricongo were selected and bought from a plantain farmer in Barranquitas, Puerto Rico. The collected corm was washed with water and disinfected with a solution containing Sodium hypochlorite. Corms were clipped for obtaining the meristematic tissue and rinsed, before putting in a 250ml beaker with disinfecting solution (3% Sodium hypochlorite and 1 drop of tween 20) and were left overnight. Then, tissues were processed in the cleanroom inside a biological cabinet. After the first disinfection, solution was discarded from the beaker and the second disinfection was made using a 10% Sodium hypochlorite solution for 30 minutes. Next, 3 washes of approximately 2-3 minutes with distilled water were carried out. The tissue was placed in an initiation media in a dark environment for 15 days. Subsequently, after the 15 days the tissues were subdivided into two fragments and transplanted into propagation media for further development. Finally, the tissues were continually transplanted, after 3 subcultures of ~22 days the plantlets were ready for the first experimental trials on the bioreactors and test tubes for comparison. Then, plantain shoots were micro propagated in a gelled medium (GM) for creating stock for experimental plantlets.
2.2. Temporary Immersions Bioreactor (TIB) and Micropropagation Assays
The temporary immersion protocol used was adapted from Aragon et al. [22], with certain modifications which included using 5 biological replicates (n=15) of different sizes and stages of development to study the progress of the tissue under bioreactors systems to see at what stage of growth the tissue can be integrated into the TIB system. Additionally, 6-Benzylaminopurine (BAP) was added to both culture media.
2.3. Growth Comparison in TIB vs Conventional Micropropagation
The growth comparison was carried out according to protocols established in Aragon et al. [23] with some modifications. In brief, plantlets were harvested at the end of the second cycle (1 cycle = 22-25 days) for morphological and physiological parameter determinations. At each time point, the following growth parameters were evaluated in plantlets: shoot length, leaves per shoot, number of roots per shoot, multiplication rate, and photosynthetic activity. The harvests time of the tissue varied from weekly to every 22-25 days to characterize development of plantains in 1 complete cycle on TIB systems.
2.4. RNA Isolation and cDNA Preparation:
The protocol for RNA extraction from plantain was adapted from Valderrama -Cháirezet al., [28] previously modified by Rodríguez-García et al.[29]. Before converting the isolated RNA into cDNA, purification of RNA was performed using Invitrogen DNA-free kit following manufacturer procedure as suggested for DNase treatment. cDNA was synthesized using Invitrogen SuperScript VILO master mix in which 4µl of this master mix were added to 16µl of each sample as per manufacturer’s instructions. Then the samples were put in the thermal cycler with the default settings for qRT-PCR.
2.5. Oligonucleotide Design
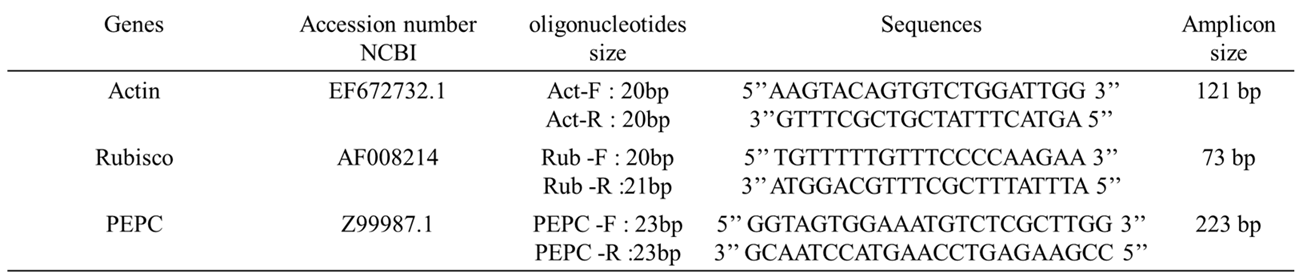
Oligonucleotides for analyzing gene expressions were synthesized according to bioinformatic analysis using the NCBI primers blast. Oligonucleotides pairs were designed for Rubisco (AF008214) and Actin (EF672732.1) using NCBI nucleotide sequences from Musa spp. The oligonucleotides pairs synthesized for phosphoenol pyruvate carboxylase (PEPC) gene were obtained from Aragón, et al., [23]. All oligos were synthesized by Sigma, Inc. (Table 1).

Table 1.
Primers sequences for qRT-PCR.
2.6. Quantitative Real Time- Polymerase Chain Reaction (qRT-PCR)
Reactions were carried out on Studio 12k Flex (Applied Biosystems). Actin was used for normalization. The qRT-PCR reaction mixture consisted of cDNA, 0.1 µM gene-specific primers, and a master mix containing SBYR green. The PCR parameters were: 95 ºC for 3 min, 40 cycles at 95 ºC for 15 s, 59 ºC for 30 s, and 72 ºC for 20 s followed by melt curve analysis. PCR products were resolved on 2 % (m/v) agarose gels run at 72 V in a TBE 1x buffer together with a low molecular DNA-standard ladder to confirm the amplicon size (Supplementary Figure 1). A comparison of relative gene expression was performed using delta-delta Ct method.
2.7. Photosynthetic Activity Measurement
After in-vitro tissue culture experiments, plantains developed on test tubes and bioreactors were successfully acclimatized under controlled conditions inside the greenhouse. After several weeks of plant acclimatization, photosynthetic activity was measured on plantains developed in test tubes and a bioreactor using a JUNIOR-PAM instrument. A light curve analysis was made following the manufactures instruction to obtain the Relative electron transport rate. Finally, crude data of Electron Transport chain Rate (ETR) and Photosynthetically Active Radiation (PAR) were plotted in excel (ETR on Y-axis and PAR on X-axis) to construct a graph to visualize the data obtained from the analysis, also a statistical test was made to verify its significance.
2.8. ICP-OES Qualitative Analysis
A semi-quantitative /qualitative metal analysis was applied using ICP-OES (Shimadzu ICPE-9820). In brief, fresh plantain tissue was taken from the experimental plants cultured on test tubes (n=3) and bioreactors (n=3) for elemental composition analysis. Approximately 0.5grams of tissue were weighed and digested in 10 ml of 2% nitric acid solution. Samples were then placed in the microwave digestion and extraction system (CEM Mars 6). Digested samples were passed to ICP-OES and parameters were adjusted for the injections of the samples for measuring the elemental distribution and composition.
2.9. Scanning Electron Microscopy
Fresh leaves and roots tissues of plantains cultured on TIB, and CM were washed with distilled water and attached to microscopic metal stubs. Samples were dried for 30 minutes, and gold coated with 10nm thickness on Luxur Gold coater. Samples were analyzed on Scanning Electron Microscope (Thermo Scientific Phenom Pro X) with adjusted parameters.
3. Results
3.1. Comparison of Growth Parameters in TIB vs Conventional Micropropagation
The regenerants were evaluated in-vitro using two different methodologies: Temporary immersion bioreactor (TIB) and conventional micropropagation (CM). The morphological characteristics and data obtained exhibited differences between the tissue growth in TIB as compared to CM. Morphological characteristics demonstrated a more developed plantlet using the bioreactors as a culture system. Furthermore, the growth rate (Figure 2. A), multiplication rate (Figure 2. B,) and total leaves per plantlet (Figure 2C) of plantains showed a higher development (Figure 3. A-B) and propagation (Figure 4A) on bioreactors plants. SEM images show differences only in the subterranean system specifically roots (Figure 3. C-D). In contrast, the aerial part did not show any morphological differences (Figure 3. E-F). Subsequently, a t-Student test was applied to verify the statistical significance of the morphological characteristic of plantain growth in both systems. Statistical results suggest that there’s no significant difference (P= 7.9309) in the growth rate but there is a significant difference in the multiplication rate (P= 0.01676) and the total leaves per plantlets (P= 0.0017) of plantains cultured in temporary immersion bioreactors.
To the best of our knowledge, we report here the first plantain tissues developed through a temporary immersion bioreactor system in Puerto Rico (Figure 3.D). Also, the efficient acclimatization of plantains cultivated on bioreactors showed that approximately 75% of plantains from bioreactors fully acclimatized to external environment. The acclimatization rate indicates that using TIB systems increases the viability to develop and acclimatize plantains faster than the conventional method.
3.3. Gene Expression
qRT-PCR data obtained showed a high transcript abundance of both PEPC and Rubisco genes on plantains cultivated on bioreactors, compared to the ones grown in test tubes (Figure 5). Statistical analysis using one-way ANOVA suggested a significant difference in PEPC transcript abundances between TIB and CM culitvated tissues (P= 0.0310).
3.4. Photosynthetic Activity
After ~3 weeks of the plants being fully acclimatized under controlled conditions in the greenhouse, plantains culitvated on TIB showed a higher relative electron transport chain rate on photosynthetic active pathways compared to the ones cultivated in conventional micropropagation (Figure 6). However, this analyzed data using the T-student test demonstrated that there are no significant differences (P= 0.5476) in the photosynthetic activity between the plant’s growth in variable growth settings.
3.5. Digestion and ICP-OES Qualitative Analysis
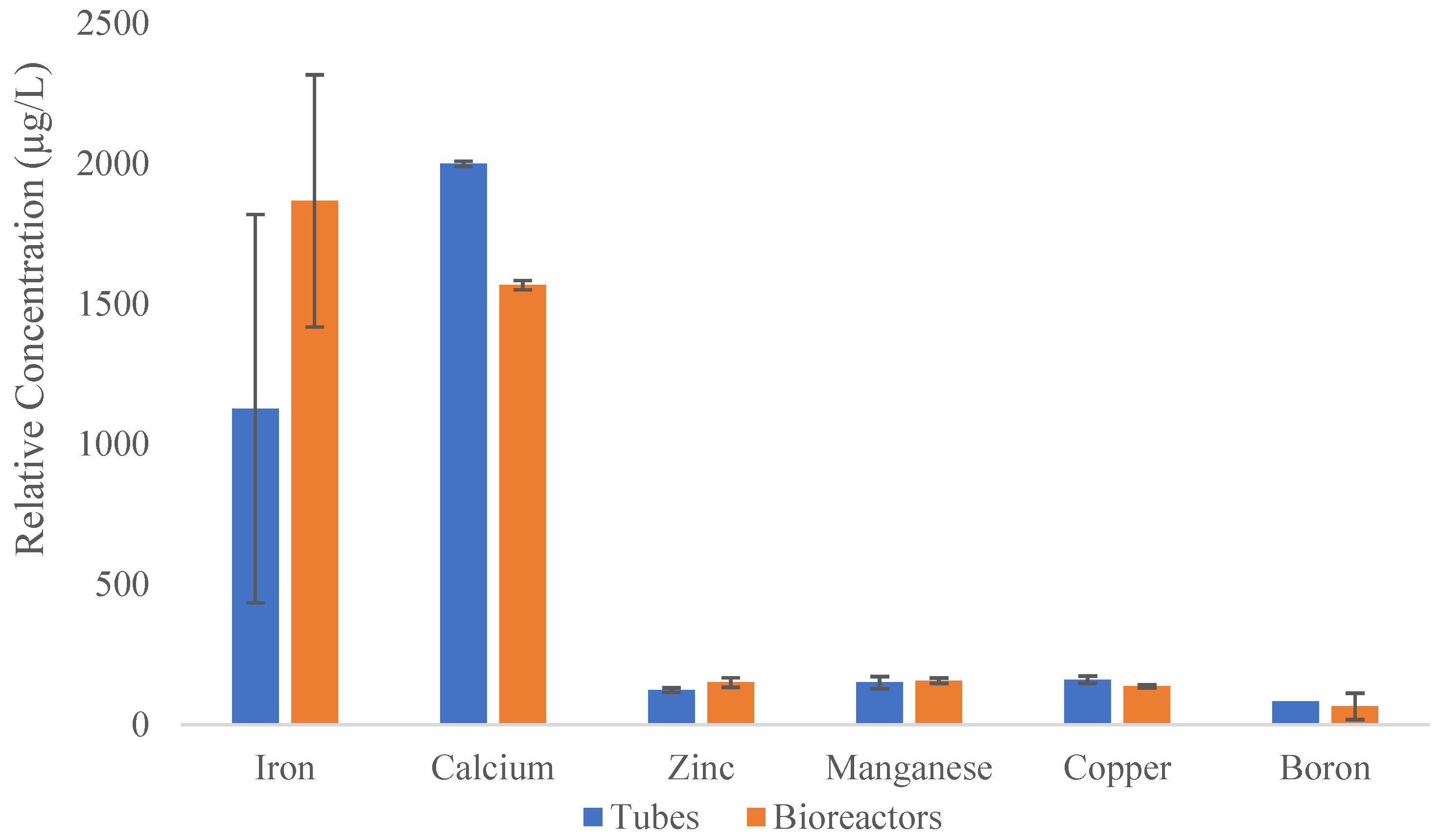
A comparative semi-quantitative assay detects the presence of all metals in plantains tissues (Supplementary Figure 2; Table 1). However, the relative concentration of the detected transition metals shows a higher presence of Iron (Fe) in plants growing on bioreactors than the ones grown in the tubes. However, a statistical test was applied to verify the significant difference of Fe in both methods. Statistical analysis, t-student test, determined that there are no significant differences (P= 0.2736) between the Fe content in plantains grown on TIB and CM. Besides, the presence of Fe was in higher quantity in comparison with the other elements of interest studied (Figure 7). However, t-student test shows a significant difference in calcium content (P= 0.0226).
4. Discussion
Plantain growth comparison using CM and TIB demonstrated better morphological characteristics on TIB. Shoot elongation, length of the roots, multiplication rate, and growth rate were similar to previously documented reports [23,24,25,26,27,28,29,30]. We tested the inoculation of different stages of plantain tissue to test the development efficiency on temporary immersion bioreactor, showing the proliferation and differentiation of all the tissues (clusters of shoots, small, rounded shape tissues, small shoots, small and big plantlet). All these tissues were able to proliferate/multiply. Results can potentially explain TIB efficacy, TIB provides numerous attributes for optimum development of plant tissue, such as oxygen supply/transport, space, nutrient acquisition, and immersion ranges [11,12]. We saw during the experimental procedure that the headspace, air supply, and temporary immersion of liquid media with a photoperiod of 16/8h improved plants health and development [22], thus showing the development of a complex root at day 5 on the TIB system. Also, SEM images highlighted morphological differences in the root structures between plantains growth on TIB and CM. Additionally, stomatal structures showed similarity in during tissue growth on both methods suggesting limited effect on stomatal morphology. Scanning stomata at various developmental stages could provide an insight into the effect of growth conditions on morphology of stomata. Additionally, air supply increased the interchanges of gases in the headspace of the bioreactors helping to simulate a natural environment, thus helping the plant to pass minimal stress during in-vitro and ex-vitro acclimatization [23,24,25,26,27,28,29,30,31,32]. Also, the interchange of gases is linked to an enzyme that is involved in carbon assimilation during photosynthetic pathways [21]. Uma et al., [30] demonstrated a 2.7-fold increase in tissue multiplication on TIB in comparison with CM. These features and our obtained data suggested an improvement in plantain multiplication rate on TIB. In brief, we provide data that support the enhancement and upscaling of plantain propagation using TIB over CM.
In addition, molecular analysis was made to study how representative photosynthetic genes express during in-vitro development of plantains under two different methodologies. Results show no significant difference in Rubisco transcripts abundances. The region of Rubisco studied for the first time on TIB plants, was the small subunit that is related to producing conformational changes in the other subunits promoting the catalytic rate [19]. However, there was significant up-regulation of PEPC gene on plantains cultivated on TIB. The conditions inside bioreactor influence the expression of phosphoenolpyruvate carboxylase (PEPC) [22]. PEPC gene shows a significantly high transcript abundance on plantains cultivated in TIB than CM, demonstrating the efficacy of carbon assimilation on plants inside TIB [23]. The carbon concentration produced by the PEPC pathway controls the process of photorespiration thus promoting the efficiency of photosynthesis [33] under TIB conditions. Additionally, as we mention before PEPC enzyme is related to the interchange of gases on plant tissue specifically on leaves, and when this enzyme is up- regulated, the rates of stomata opening increase significantly, suggesting that interchange of gases between the plant and TIB is effective and promote plant organ differentiation/multiplication. High transcripts abundances of this gene on plantains produced in TIB can explain the healthy and vigorous growth of these plantlets [23]. This suggests that plantains produced on TIB perhaps are more photosynthetically active than the ones produced on CM. Additionally, relative electron transport rate measurement on ~3 weeks of fully acclimatized plantains obtained from TIB and CM suggests that TIB plantains continue to possess high photosynthetic activity weeks after acclimatization. Though there’s no statistical difference in photosynthetic rate in plantains growth on TIB and CM during ex-vitro conditions, further analysis during in-vitro conditions needs to be applied. However, previous data shows a relation between light intensity and gases such as CO2 with photosynthetic rate on in-vitro plant development and PEPC activation [21]. Proving earlier documented data which explain the presence [34,35] and essential [36] role of gas interchanges on TIB headspace for improved growth development.
Elemental composition of plantains on TIB was carried out to study presence of certain metals of interest that potentially interact with photosynthesis pathway. All metals of the periodic table were tested for plantains growth on CM and TIB. Substantial differences in some elements of interest including Boron, Calcium, and transition metals, Iron (Fe), Manganese (Mg), Zinc (Zn), and Copper (Cu) were obtained and can provide an explanation for the difference in plant development on CM and TIB. Iron was present in more quantity on TIB. This was expected since previous data [24] suggest that copper, zinc, manganese, and specifically iron act as a cofactor on some metabolic pathways in plants. These features can positively suggest that Fe can be potentially linked to the high transcript abundance of PEPC on TIB plantains. Also, is possible that Fe can be involved in chloroplast where carbon metabolism occurs. Most of the pathways that occur in this organelle require the presence of metals as cofactors [24]. The higher content of Fe found in plantains developed on TIB suggested a highly potential correlation with PEPC proteins. It is known that Fe acts as a cofactor of some photosynthetic pathways [24] and a correlation between iron and PEPC may occur during the activation/signaling of transcription factors related to these two photosynthetic genes. Previous research [25] revealed that iron and copper are present in high quantities in chloroplast, and the high content of these metals is linked to metalloproteins that strictly require these cofactors for photosynthetic electron transport chain. We tested the relative electron transport rate on acclimatized plantains produced on test tubes and bioreactors and the results confirm previous data which explained the crucial and important roles that metals play as cofactors and regulators on photosynthetic pathways [25,26,27,28,29,30,31,32,33,34,35,36,37]. Transition metals are essential for various metabolic paths that are crucial for healthy plant development [20]. These features potentially explain why plantains produced on bioreactors possess a successful acclimatization rate because presence of these elements can also be involved in developing certain resistances to biotic and abiotic factors [37,38]. Additionally, content of boron on TIB was lower than CM, this has a reason to be and is that excess or deficiency of boron can be toxic for plants producing interruption and damage to metabolic pathways [39]. It can be suggested that concentration of boron on plantains cultivated on TIB was optimum in comparison with CM. Boron is linked to play a role in the cell wall composition and synthesis as well as cell division [39] demonstrating and explaining why plantains produced in TIB growth vigorous as well the possess high multiplication rate. Manganese content on TIB plantains shows a higher relative content in comparison with the plantain produced CM. This might provide a probable correlation with photosynthetic gene expression as this metal acts on the oxygen evolving complex and, in the photosystem II [40].
In addition, other important elements such as copper and calcium show a higher relative concentration in leaf tissues of plantains grown on CM in comparison to the TIB. Differences in plantain pseudostem development between TIB and CM can be an effect of calcium concentrations because of its essential role in cell wall synthesis and structure [41]. We suggest that CM plantains are compressed in a small environment and transport elements such as Ca develop a rigid cell wall. Subsequently, the lower relative concentration of Ca on TIB plantains suggested an explanation why TIB plantains show a dynamic/flexible pseudostem development. However, quantitative elemental analysis could reveal more specific quantification than the applied semi-quantitative/qualitative analysis. In brief, our data suggest, that morphological, biochemical, and molecular analysis can help to understand how plants develop in-vitro and how the method of cultures can be improved for producing high-quality plants.
5. Conclusions
Evaluation of both in-vitro methods demonstrates an improvement in plant development, growth and multiplication rate using TIB system over CM. Morphological and imaging aspects need validation through molecular techniques for further validating the functions. Differences between data obtained from the TIB and CM suggest an enhancement and optimum development of plantain using TIB system. Photosynthetic gene expression analysis shows high transcripts abundance of phosphoenolpyruvate carboxylase (PEPC) in plants cultured on TIB. Also, to the best of our knowledge Rubisco small subunits and PEPC expression were tested for the first time in TIB. The photosynthetic activity measurement using the Junior PAM system helps to validate the gene expression analysis giving an overview of this activity after plant acclimatization. These result suggest that those experimental genes can still be up-regulated during the acclimatization of plants obtained from the TIB. Finally, we conclude that TIB is a viable tool for increasing plantain propagation, health, and strength faster than CM. Also, using TIB show a potential tool for food security and a more cost-effective and less time-consuming.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1. title: Melting Curves and Electrophoresis results; Figure S2. title: ICP-OES element composition; Table S1: title: ICP-OES raw data.
Author Contributions
Conceptualization: A.A; methodology: C.S, S.M.M, D.A.S &A.R.C; formal analysis, A.A.; investigation: C.S.; data curation, A.A; writing—original draft preparation: C.S; writing—review and editing, A.A & C.S; visualization, C.S; supervision J.A.N.B, A.N. &A.A; project administration, J.N. All authors have read and agreed to the published version of the manuscript.
Funding
Agro-Evolution project funded by Banco Popular Puerto Rico and Hispanic Federation of Puerto Rico thought the award of Big Ideas Challenges.
Institutional Review Board Statement
Not applicable
Data Availability Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest.
References
- Huang, H.; Jing, G.; Wang, H.; Duan, X.; Qu, H.; Jiang, Y. The combined effects of Phenylurea and gibberellins on quality maintenance and shelf life extension of banana fruit during storage. Science Horticulture 2014, 167, 36–42. [Google Scholar] [CrossRef]
- Nelson, S., Ploetz, R.C, & Kepler, A.K. (2006). Musa Species (bananas and plantains) Species Profiles for Pacific Island Agroforestry. Permanent Agriculture Resource. https://agroforestry.org/images/pdfs/Musa-banana-plantain.pdf.
- Kumar, N.; Reddy, M. In vitro Plant Propagation: A Review. Journal of Forest Science 2011, 27, 61–72. [Google Scholar] [CrossRef]
- Rodríguez Cruz, L., & Niles, M. (2018). Hurricane Maria’s Impacts on Puerto Rican Farmers: Experience, Challenges, and Perceptions. University of Vermont: Burlington.
- Smith, M., & Meade, B. (2019, June 3). Global Food Security.USDA. https://www.ers.usda.gov/amber-waves/2019/june/who-are-the-world-s-food-insecure-identifying-the-risk-factors-of-food-insecurity-around-the-world/.
- Bellido, M. S. (September 10, 2014). Food Security in Puerto Rico: Vulnerable Food Supply. American Society for Nutrition. https://nutrition.org/food-security-puerto-rico-vulnerable-food-supply/.
- Roels, S.; Escalona, M.; Cejas, I.; Noceda, C.; Rodriguez, R.; Canal, M.; Debergh, P. Optimization of plantain (Musa AAB) micropropagation by temporary immersion system. Plant Cell, Tissue and Organ Culture 2005, 82, 57–66. [Google Scholar] [CrossRef]
- Nhut, D. T., Trinh Don, N., Hong Vu, N., Quoc Thien, N., Thi Thu Thuy, D., Duy, N., & A. Teixeira da Silva, J. (2006). Advanced Technology in Micropropagation of Some Important Plants. Global Science Book.
- Escalona, M.; Lorenzo, J.C.; Gonza´ lez, B.; Daquinta, M.; Gonza´ lez, J.; Desjardins, Y.; Borroto, C.G. Pineapple (Ananas comosus L. Merr) micropropagation in temporary immersion systems. Plant Cell Reports 1999, 18, 743–748. [Google Scholar] [CrossRef]
- Florez, S.; Curtis, M.S.; Shaw, S.E.; Hamaker, N.K.; Larsen, J.S.; Curtis, W.R. A Temporary Immersion Plant Propagation Bioreactor with Decoupled Gas and Liquid Flows for Enhanced Control of Gas Phase. American Institute of Chemical Engineers 2016, 32, 337–345. [Google Scholar] [CrossRef]
- Albarrán, J.; Bertrand, B.; Lartaud, M.; Etienne, H. Cycle characteristics in a temporary immersion bioreactor affect regeneration, morphology, water, and mineral status of coffee (Coffea arabica) somatic embryos. Plant Cell, Tissue and Organ Culture 2005, 81, 27–36. [Google Scholar] [CrossRef]
- Georgiev, V.; Schumann, A.; Pavlov, A.; Bley, T. Temporary immersion systems in plant biotechnology. Engineering in Life Sciences 2014, 14, 607–621. [Google Scholar] [CrossRef]
- Paek, K.; Chakrabarty, D.; Hahn, E. Application of bioreactor systems for large-scale production of horticultural and medicinal plants. Plant Cell, Tissue and Organ Culture 2005, 81, 287–300. [Google Scholar] [CrossRef]
- Alvard, D.; Cote, F.; Teisson, C. Comparison of methods of liquid medium culture for banana micropropagation. Plant Cell, Tissue and Organ Culture 1993, 32, 55–60. [Google Scholar] [CrossRef]
- Etienne, H.; Berthouly, M. Temporary immersion in plant micropropagation. Plant Cell Tissue and Organ Culture 2002, 69, 215–231. [Google Scholar] [CrossRef]
- Tumer, N.; Clark, W.; Tabor, G.; Hironaka, C.; Fraley, R.; Shah, D. The genes encoding the small subunit of ribulose-1,5-bisphosphate carboxylase are expressed differentially in petunia leaves. Nucleic Acids Research 1986, 14, 3325–3342. [Google Scholar] [CrossRef]
- Scafaro, A.P.; Bautsoens, N.; Boer, B.; Rie, J.; Gallé, A. A Conserved Sequence from Heat-Adapted Species Improves Rubisco Activase Thermostability in Wheat1. Plant Physiology 2019, 181, 43–54. [Google Scholar] [CrossRef]
- Marcus, Y., Altman-Gueta, H., Snir, A., Wolff, Y., & Gurevitz, M. Does Rubisco Limit the Rate of Photosynthesis? Photosynthesis. Energy from the Sun: 14th International Congress on Photosynthesis 2008, 863-866. Springer. [CrossRef]
- Suzuki, Y.; Makino, A. Availability of Rubisco small subunit up-regulates the transcript levels of large subunit for stoichiometric assembly of its holoenzyme in rice. Plant Physiology 2012, 160, 533–540. [Google Scholar] [CrossRef]
- Kanai, R., & Edward, G. (1999). The biochemistry of C4 photosynthesis. In R Monson, ed, C4 Plant Biology. Academic Press.
- Cousins, A.; Baroli, I.; Badger, M.; Ivakov, A.; Lea, P.; Leegood, R.; Caemmerer, S. The Role of Phosphoenolpyruvate Carboxylase during C4 Photosynthetic Isotope Exchange and Stomatal Conductance. Plant Physiology 2007, 145, 1006–1017. [Google Scholar] [CrossRef]
- Aragón, C.; Escalona, M.; Capote, I.; Pina, D.; Cejas, I.; Rodriguez, R.; Canal, M.; Sandoval, J.; Roels, S.; Debergh, P.; Gonzalez - Olmedo, J. Photosynthesis and carbon metabolism in plantain (Musa AAB) growing in temporary immersion bioreactor (TIB) and ex vitro acclimatization. In Vitro Cellular & Developmental Biology - Plant 2005, 41, 550–554. [Google Scholar] [CrossRef]
- Aragón, C.; Sanchez, C.; Gonzalez - Olmedo, J.; Escalona, M.; Carvalho, L.; Amancio, S. Comparison of plantain plantlets propagated in temporary immersion bioreactors and gelled medium during in vitro growth and acclimatization. Biologia Plantarum 2014, 1, 29–38. [Google Scholar] [CrossRef]
- Ravet, K.; Pilon, M. Copper and Iron Homeostasis in Plants: The Challenges of Oxidative Stress. Antioxidants & Redox Signaling 2013, 19. [Google Scholar] [CrossRef]
- Yruela, I. Transition metals in plant photosynthesis. Metallomics. 5 2013, 1090–1109. [Google Scholar] [CrossRef]
- Merchant, S., Wise, R., & Hoober, J. Trace metal utilization in chloroplasts. The Structure and Function of Plastids 2006, 199– 218. Springer. [CrossRef]
- Raven, J.; Evans, M.; Korb, R. The role of trace metals in photosynthetic electron transport in O2-evolving organisms. Photosynthesis Research 1999, 60, 111–149. [Google Scholar] [CrossRef]
- Valderrama-Cháirez, M.L.; Cruz-Hernández, A.; Paredes-López, O. Isolation of functional RNA from cactus fruit. Plant Mol. Biol. Rep. 2002, 20, 279–286. [Google Scholar] [CrossRef]
- Rodríguez-García, C.; Peraza-Echeverría, L.; Islas-Flores, I.; Canto-Canché, B.; Grijalva, R. Isolation of retro-transcribed RNA from in vitro Mycosphaerella fijiensis-infected banana leaves. Genetics and molecular research: GMR. 2010, 9, 1460–1468. [Google Scholar] [CrossRef]
- Uma, S.; Karthic, R.; Kalpana, S.; Backiyarani, S.; Saraswathi, M.S. A novel temporary immersion bioreactor system for large-scale multiplication of banana (Rasthali AAB-Silk). Scientific Reports 2021, 11, 20371. [Google Scholar] [CrossRef]
- Yang, L.; Zambrano, Y.; Hu, C.-J.; Carmona, E. Sugarcane metabolites produced in CO2-rich temporary immersion bioreactors (TIBs) induce tomato (Solanum lycopersicum)resistance against bacterial wilt (Ralstonia solanacearum) In Vitro Cellular and Developmental Biology. Plant 2010, 46, 558–568. [Google Scholar] [CrossRef]
- Aragón, C.; Escalona, M.; Rodriguez, R.; Cañal, M. Effect of sucrose, light, and carbon dioxide on plantain micropropagation in temporary immersion bioreactors. In Vitro Cellular & Developmental Biology - Plant 2009, 46, 89–94. [Google Scholar] [CrossRef]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Pérez-Pérez, M.; Francia, F.; Lemaire, S. Redox regulation of the Calvin–Benson cycle: something old, something new. Frontiers in Plant Science 2013, 4, 1–21. [Google Scholar] [CrossRef]
- Matthijs, D., Gielis, J., & Debergh, P. (1995). Automation and Environmental Control in Plant Tissue Culture. Kluwer Academic Publishers. [CrossRef]
- Maes, K.; Debergh, P. Volatiles emitted from in vitro grown tomato shoots during abiotic and biotic stress. Plant Cell, Tissue and Organ Culture. 2003, 75, 73–78. [Google Scholar] [CrossRef]
- Buddendorf-Joosten, J.; Woltering, E. Components of the gaseous environment and their effect on plant growth and development in-vitro. Plant Growth Regulation. 1994, 15, 1–16. [Google Scholar] [CrossRef]
- González-Guerrero, M.; Escudero, V.; Saéz, Á.; Tejada-Jiménez, M. Transition Metal Transport in Plants and Associated Endosymbionts: Arbuscular Mycorrhizal Fungi and Rhizobia. Frontiers in Plants Science 2016, 7, 1088. [Google Scholar] [CrossRef]
- Gonzalez, E. (2005). Mass propagation of tropical crops in temporary immersion systems. Springer. [CrossRef]
- Seyed Ali, G., & Hassan, E. (2020). The Importance of Boron in Plant Nutrition. In Metalloids in Plants. John Wiley & Sons, Inc. [CrossRef]
- Santiago, A.; Höller, S.; Meier, B.; Peite, E. Manganese in Plants: From Acquisition to Subcellular Allocation. Frontiers in Plant Science 2020, 11, 1–23. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: a central regulator of plant growth and development. The Plant cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef]
Figure 1.
Musa sp. cultured on conventional micropropagation (A), and temporary immersion bioreactors (A). [n=15].
Figure 1.
Musa sp. cultured on conventional micropropagation (A), and temporary immersion bioreactors (A). [n=15].

Figure 2.
In-vitro plantains development comparison using conventional micropropagation and temporary immersion bioreactors. [n=15]. (A.) Growth rate, (B). Multiplication rate, (C). Number of leaves per plantlet.
Figure 2.
In-vitro plantains development comparison using conventional micropropagation and temporary immersion bioreactors. [n=15]. (A.) Growth rate, (B). Multiplication rate, (C). Number of leaves per plantlet.
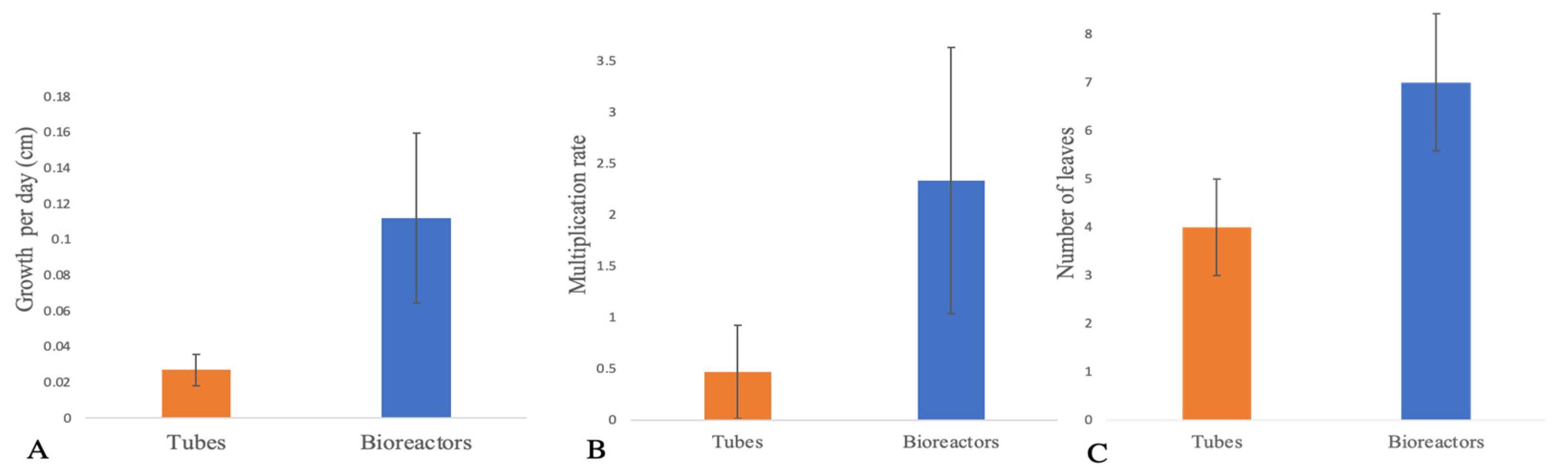
Figure 3.
Morphological comparison of plantains growth on TIB (A) and CM (B). SEM images of root apex of tissues grown in TIB (C) and CM (D) and stomata in TIB (E) and CM (F).
Figure 3.
Morphological comparison of plantains growth on TIB (A) and CM (B). SEM images of root apex of tissues grown in TIB (C) and CM (D) and stomata in TIB (E) and CM (F).

Figure 4.
Shoots cluster of plantains multiplication in TIB (A). First plantain obtained through TIB demonstrating successful acclimatization (B).
Figure 4.
Shoots cluster of plantains multiplication in TIB (A). First plantain obtained through TIB demonstrating successful acclimatization (B).

Figure 5.
Relative gene expression of photosynthetic genes, Rubisco (A); PEPC (B), comparison between Musa sp. growth on conventional micropropagation and bioreactors [n=9].
Figure 5.
Relative gene expression of photosynthetic genes, Rubisco (A); PEPC (B), comparison between Musa sp. growth on conventional micropropagation and bioreactors [n=9].
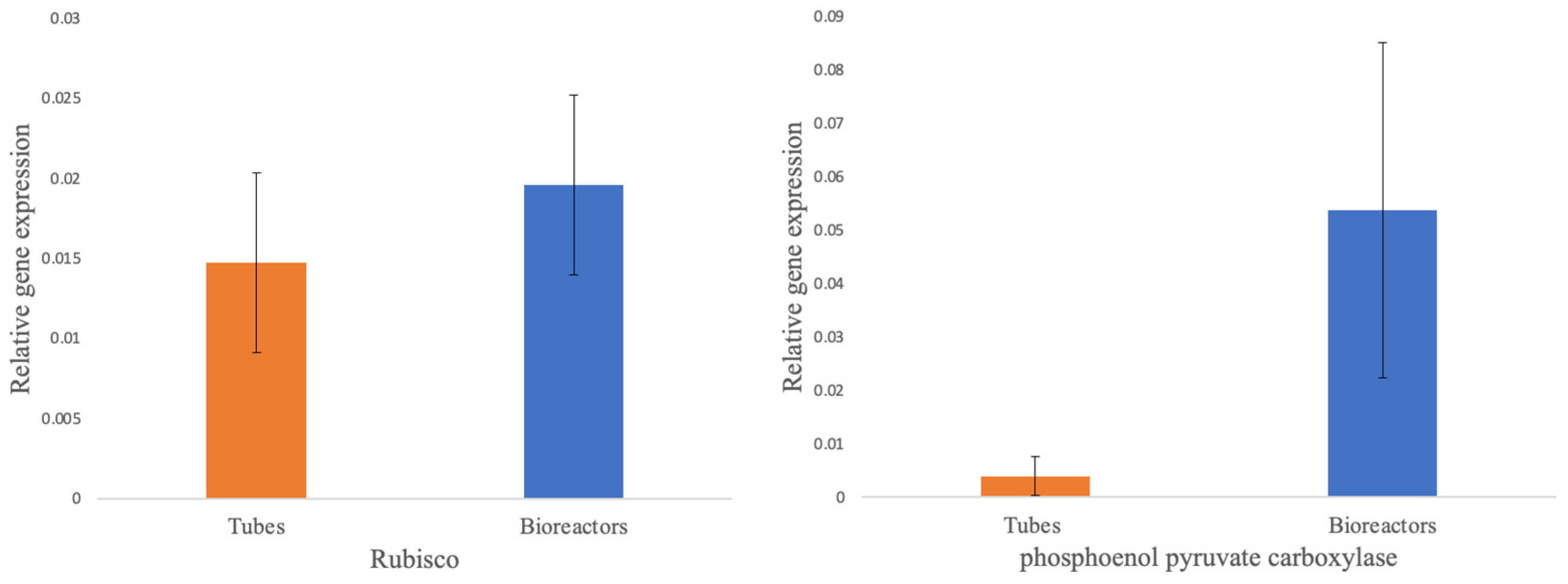
Figure 6.
Photosynthetic activity comparison between acclimatized Musa sp. growth on bioreactors and conventional micropropagation.
Figure 6.
Photosynthetic activity comparison between acclimatized Musa sp. growth on bioreactors and conventional micropropagation.
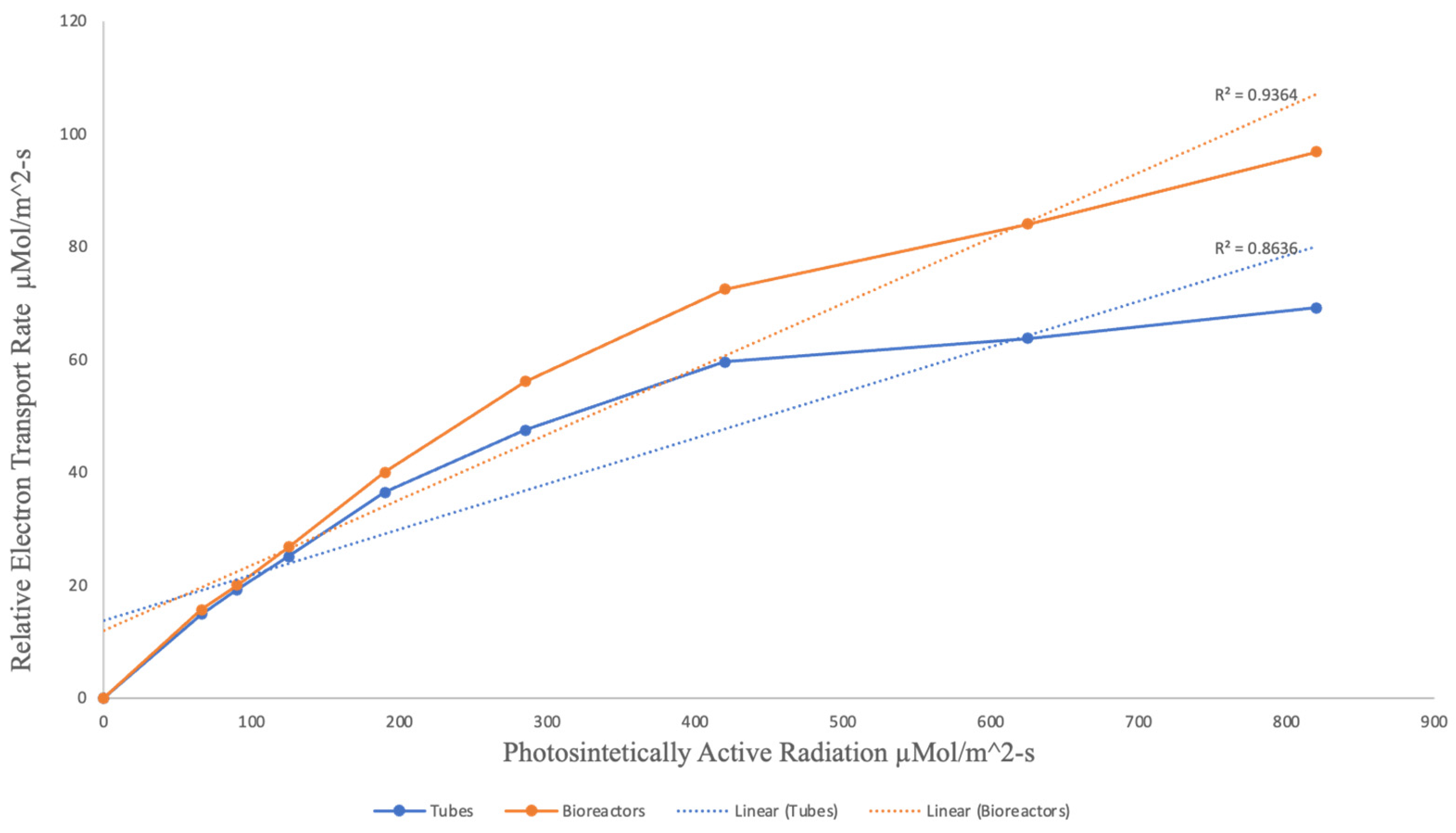
Figure 7.
Comparison of the transition metals present on Musa sp. growth in-vitro [n=3].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



