Submitted:
03 July 2026
Posted:
06 July 2026
You are already at the latest version
Abstract
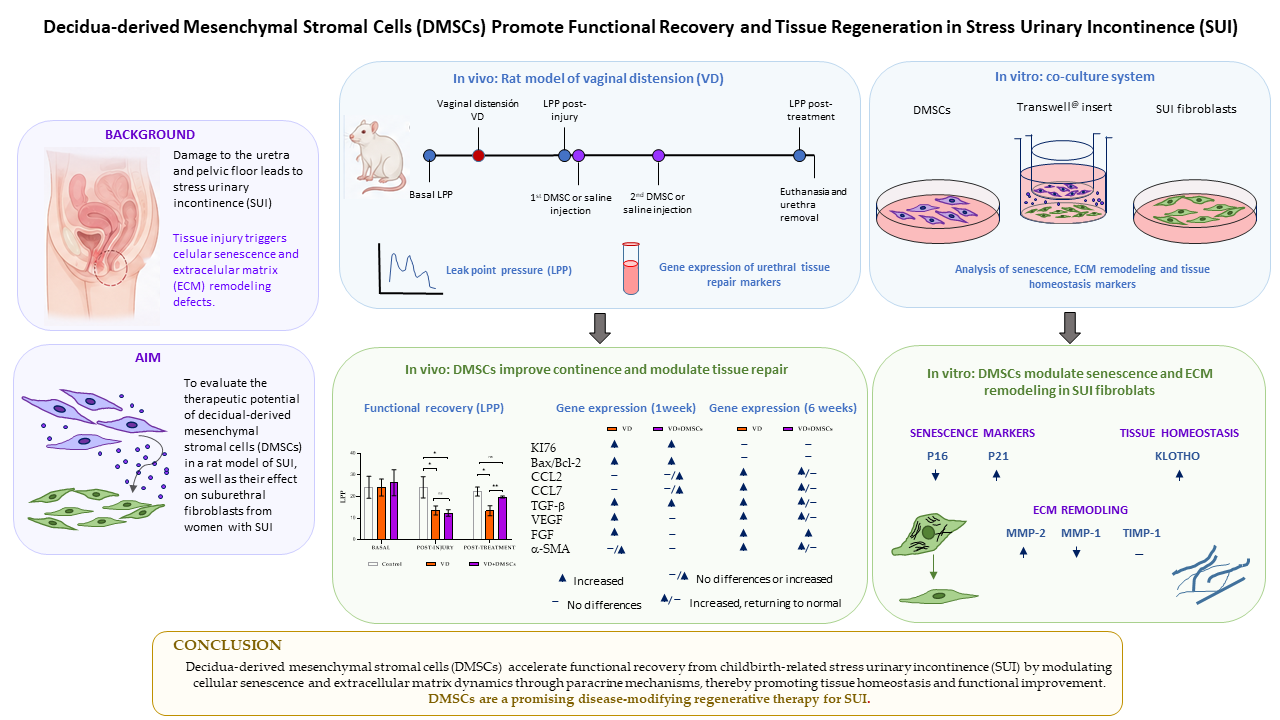
Stress urinary incontinence (SUI) is a highly prevalent condition associated with pelvic floor damage, fibroblast dysfunction, and impaired extracellular matrix (ECM) remodeling. This study aimed to investigate the regenerative potential and underlying molecular mechanisms of decidua-derived mesenchymal stromal cells (DMSCs) in a rat model of vaginal distension (VD) and in human suburethral fibroblasts from SUI patients. Adult female rats were subjected to VD and treated with periurethral DMSC injections, followed by functional and transcriptomic analyses, one and six weeks after VD. In parallel, an in vitro co-culture system was used to evaluate the paracrine effects of DMSCs on SUI fibroblasts. DMSC treatment demonstrated a substantial improvement in continence in rats, as measured by leak point pressure (LPP) and accelerated recovery, while modulating key pathways associated with tissue repair. Gene expression analyses revealed transient increases in proliferative and inflammatory markers, followed by earlier normalization compared to untreated animals. In vitro, DMSCs reduced p16 expression, increased p21 and Klotho levels, and rebalanced eECM remodeling by decreasing MMP-1 and increasing MMP-2. These findings indicate that DMSCs promote a regenerative microenvironment by modulating senescence and ECM dynamics. Overall, DMSCs may represent a promising disease-modifying strategy for SUI by enhancing tissue repair and functional recovery.
Keywords:
perinatal mesenchymal stromal cells
; decidua-derived mesenchymal stromal cells (DMSCs)
; stress urinary incontinence (SUI)
; vaginal distension
; urethral function
; endopelvic fascia
; tissue regeneration
; regenerative medicine
; senescence
; extracellular matrix
1. Introduction
Stress urinary incontinence (SUI) is defined as the unintentional loss of urine following an effort due to a weakened pelvic floor or urethral sphincter muscles. SUI is a prevalent condition among women, affecting up to one in three of them, particularly following childbirth or as part of the aging process [1]. SUI has been demonstrated to result in a multitude of psychosocial consequences, including social isolation, diminished self-esteem, sexual dysfunction, and augmented financial strain. The implementation of efficacious management regimens frequently requires a multifaceted approach, encompassing behavioral strategies in conjunction with, on occasion, surgical interventions. These measures are undertaken with the objective of restoring functionality and enhancing quality of life [2]. SUI is the most common type of female incontinence, with a prevalence that varies from the 23%in Spain to over 40% in France (44%), Germany (41%), and the UK (42%) [3].
The endopelvic fascia is a complex, three-dimensional network of connective tissue that lines the pelvic walls and floor. This fascia plays a critical role in maintaining pelvic organ support, including the vagina, uterus, and bladder. It also serves as a conduit for nerves and blood vessels [4]. Fibroblasts are the predominant cellular element of this tissue. They are responsible for producing and remodeling of the extracellular matrix (ECM), a complex network of proteins that provides structural support [5]. The ECM is primarily composed of collagen, which contributes to tensile strength, and elastin, which facilitates elasticity. Weakness in the endopelvic fascia, resulting from abnormal fibroblast activity, leads to compromised support of the pelvic organs [6].
Vaginal distension (VD) models are primarily used in preclinical animal studies to simulate pelvic floor trauma related to childbirth and SUI. The VD model has been reported to cause external urethral sphincter dysfunction and various physiological changes, including genitourinary organ hypoxia [7], loss of external urethral sphincter function [8], and reduced leakage pressure [9]. In response to VD, affected tissues activate cellular and molecular mechanisms to restore homeostasis. Animals subjected to a VD procedure typically show partial recovery of urethral function over time [10]. The time required for spontaneous recovery in the VD model has been reported to be inversely proportional to the duration of distension. While functional improvement may occur, some structural effects of VD can persist for up to six weeks after a four-hour procedure in rats [11], and up to 12 weeks in ovariectomized rats [12]. The most significant biochemical changes associated with tissue repair following VD have been observed in the urethra compared to neighboring organs such as the bladder, vagina, and rectum, at least immediately after VD. This is attributed to the urethra’s lack of exposure to tensile forces and its lower vascularization. Consequently, the urethra appears to be more susceptible to damage caused by stretch overload and acute hypoxia [13].
Regenerative medicine, defined as the use of stem cells to repair or revitalize damaged tissues, has emerged as a promising therapeutic approach for stress urinary incontinence (SUI). Among the various cell types that have been studied, mesenchymal stem cells (MSCs)—a population of adult stem cells—represent one of the most promising and extensively investigated sources for stem cell–based research and therapy [14,15]. MSCs can be obtained from several tissues, including bone marrow, adipose tissue, dental pulp, and various perinatal tissues, such as the placenta, Wharton’s jelly, the amniotic membrane, the decidua, and others. Perinatal tissues have emerged as particularly promising sources, as they are typically discarded after childbirth, making them readily available, easy to obtain, and free from ethical concerns. Among these, cells derived from the maternal portion of the placenta (the decidua) have been identified as mesenchymal in nature and are known as decidua-derived stromal mesenchymal cells (DMSCs) [16]. DMSCs are a relatively homogeneous population of mesenchymal cells that have demonstrated therapeutic potential in various disease models, both in vivo and in vitro. This potential is largely attributable to their migratory behavior, antitumor effects, low immunogenicity, and immunomodulatory properties.
In a previous study conducted by our laboratory, we observed that six weeks after six hours of vaginal distension, functional recovery was significantly greater in animals treated with two injections of DMSCs than in untreated animals [9]. Histological analysis of the urethras revealed that DMSC treatment resulted in a significant increase in the number and integrity of muscle fibers in the external urethral sphincter. We also observed reduced extracellular matrix infiltration and larger elastic fibers in the urethral tissue of the DMSC-treated animals. In the present study, we conducted a transcriptomic analysis of these urethras to identify the molecular mechanisms underlying the superior recovery observed in the treated animals. Rather than emphasizing structural outcomes, this approach sought to identify the dynamic molecular mechanisms that are activated during tissue repair, as well as the potential contribution of DMSCs in modulating this process. The study examined apoptotic balance (Bax/Bcl-2), cell proliferation (Ki67), angiogenic signaling (VEGF and FGF), tissue remodeling (TGF-β and α-SMA), and chemokine-mediated recruitment (CCL2 and CCL7) of inflammatory and immune cells.
As most changes caused by vaginal distension become evident shortly after injury, we included another group of animals that we studied one-week post-injury. We believe this will allow us to evaluate the early effect of DMSCs on the urethra, both at the functional and molecular levels. Furthermore, the study will allow us to identify molecular pathways involved in urethral repair following VD and compare molecular changes in the short (one week) and long (six weeks) term. This could enhance our understanding of the temporal progression of the urethral repair process and help determine whether therapeutic intervention with DMSCs influences this remodeling and recovery process.
2. Results and Discussion
2.1. The Effect of DMSCs on Urodynamic Parameters and on a Stress Urinary Incontinence Model in Rats Induced by Vaginal Distension
We developed two procedures to study the therapeutic use of DMSCs in adult, virgin Sprague-Dawley rats that were subjected to vaginal distension. The first procedure, which has been reported previously, evaluated the functional and histological recovery of the urethra six weeks after injury [9]. Here, we present the results obtained using the second procedure, where the recovery was evaluated one week after injury (Figure 1A). Baseline LPP values were determined two or three days before vaginal distension (VD). Damage was assessed by measuring leak point pressure (LPP) after VD and one-week post-procedure (see Figure 1A). The animals were divided into three groups prior to the experimental intervention: a control group (CONTROL) with no damage or treatment, an untreated group with vaginal distension but without cell treatment (VD), and a group with vaginal distension and cell treatment (VD+DMSCs). The flowchart of the design for LPP measurements, VD, and DMSC treatment at six weeks can be viewed in our previous publication [9]. LPP measurements were performed in a blinded manner, without knowledge of the group assignment of each animal, in order to minimize bias. The impact of VD on LPP was evident, with a significant decrease observed in both groups of animals compared to the control group (Figure 1B; post-injury). However, the results of the one-week measurement indicated that the administration of two doses of DMSC following injury led to a significant improvement in LPP in the treated group compared to the untreated group (Figure 1B; post-treatment).
2.2. Expression of Genes Related to the Urethral Repair Process
To characterize the molecular response to vaginal distension (VD) and DMSC treatment, we analyzed the expression of genes involved in tissue injury and repair in urethral samples obtained from both experimental procedures (one-week and six-week procedures).
2.2.1. Expression of Genes Related to Apoptosis and Proliferation
First, we have evaluated apoptosis and cell proliferation, as these processes will allow us to understand the dynamics of urethral tissue turnover. Apoptosis was evaluated using the Bax/Bcl-2 expression ratio, while Ki67 expression was evaluated as a marker of cell proliferation. One week after vaginal distension (VD), a significant increase in Ki67 expression was observed in both experimental groups compared to the control group. An increase in the Bax/Bcl-2 ratio was also observed, though it was only significant in the urethras of the treated animals (Figure 2). The increase in these parameters one week after VD is consistent with the ongoing phase of tissue remodeling following injury. This phase is characterized by the controlled death of damaged cells (apoptosis) and an increased rate of cell proliferation to replace lost cells [17]. The changes observed in the animals treated with DMSC (VD+DMSCs) were significant compared to those in the untreated animals. This suggests that the treated animals had a more potent regenerative response at this time.
To analyze the repair process over time, Bax/Bcl-2 expression ratio and Ki67 expression were examined in the sixth-week group. However, no significant differences were observed between the treated (VD+DMSCs) and untreated groups (VD) at this stage. The Bax/Bcl-2 ratio approached baseline levels, indicating that the damage-induced apoptotic response had been resolved (Figure 2). Furthermore, Ki67 expression decreased in the experimental groups by the sixth week. Evaluating these parameters alone at six weeks does not allow us to determine the extent to which tissue repair has been completed. However, the behavior of these parameters by the sixth week suggests that urethral tissue in both groups may have reached an advanced stage of repair and resolution. In conclusion, these findings suggest that DMSC treatment does not necessarily alter the final outcome. Rather, it enhances the magnitude of the initial repair response, consequently accelerating the overall repair process as seen at the functional level.
2.2.2. Chemokine Gene Expression
Chemokines play a pivotal role in tissue repair [18]. These proteins regulate homeostasis and are produced by various cell types in response to tissue damage [19]. While their presence was initially associated with the onset of the inflammatory response, evidence suggests that they also play a significant role in the resolution phase of tissue damage [20]. By recruiting and guiding immune and progenitor cells to the site of injury, they coordinate the initial inflammatory phase and facilitate the transition to the remodeling stages [17,21].
We focused on the chemokines CCL2 (MCP-1) and CCL7 (MCP-3) because they are both strongly upregulated in urethral and bladder tissues after ischemic, mechanical, or childbirth-related injuries [22]. In week 1, there were no significant differences observed between the two treatment groups (VD and VD+DMSCs) and the control group for either chemokine (Figure 3A,B). Though, an increase in the levels of both CCL2 and CCL7 was observed in some animals in the DMSC treated group (VD+DMSCs), but not in the untreated group (VD). By week 6, the expression of CCL2 and CCL7 was above control levels in most animals, both treated (VD+DMSCs) and untreated (VD, Figure 3A,B). The increase was significant for CCL7 (**p < 0.01) and nearly significant for CCL2 (p = 0,0508) in the untreated (VD) group. In the treated group (VD+DMSCs), levels of both chemokines were higher compared to the control group, although the difference was not statistically significant. It is noteworthy that these levels were lower compared to the untreated group (VD), suggesting that the repair process was more advanced in the animals treated with DMSCs.
Studies by other authors have shown that CCL7 is rapidly upregulated in the urethral tissue of rats immediately after vaginal distension, and returns to baseline within 24 hours [23]. In light of this, our data on CCL2 and CCL7 expression may suggest that the acute phase of inflammation has likely subsided in most animals after the one-week experimental procedure. The elevation observed at week six is more consistent with a secondary phase of chemokine activity associated with tissue remodeling. During this phase, chemokines contribute to the mobilization of progenitor cells, the deposition of the extracellular matrix, and angiogenesis, in addition to immune cell recruitment [20,24].
Both CCL2 and CCL7 are involved in the recruitment of mesenchymal stromal cells (MSCs) through the CCL2/CCR2 and CCL7/CCR1 signaling pathways [25,26,27,28,29,30]. Experimental models have shown that activating these pathways improves functional recovery and enhances stem cell homing after urogenital injury [27]. In this context, the increased expression of CCL2 and CCL7 by week 6 may indicate an active reparative microenvironment that promotes progenitor cell recruitment and tissue remodeling, rather than ongoing acute inflammation.
Importantly, chemokine signaling must be tightly regulated during the repair process. CCL2 and CCL7 are essential for effective regeneration; however, their sustained overexpression can promote chronic inflammation, fibrosis, and pathological remodeling. Insufficient expression, on the other hand, impairs macrophage recruitment and delays healing [18,20,31]. In our study, chemokine levels in several DMSC-treated animals approached control levels by week 6, whereas untreated animals maintained higher expression levels. These results imply that DMSC therapy could speed up the resolution of the inflammatory-reparative response and encourage a more efficient transition to tissue homeostasis [32].
2.2.3. Growth Factors Associated with Regeneration
Transforming growth factor beta (TGF-β) plays a key role in maintaining homeostasis and promoting tissue renewal under physiological conditions [33]. In pathological conditions, TGF-β signaling plays a key role in regulating inflammation and wound healing [34]. TGF-β has been shown to stimulate collagen formation and extracellular matrix (ECM) synthesis while decreasing its degradation [35]. Dysregulation during the tissue repair process can lead to fibrosis or poor wound healing. In the present study, TGF-β expression increased in both VD groups (VD and VD+DMSCs) at the week 1 procedure, reaching significantly higher levels than in the control group. By week 6, TGF-β expression remained elevated in both groups compared to the control group. However, in the treated group (VD+DMSCs), TFG-β levels remained at levels similar to those observed in week 1, while in the untreated animals (VD), they were higher (Figure 4).
The TGF-β induction observed in week 1 in both groups (VD and VD+DMSCs) after injury is consistent with its role in activating the repair response, which requires the synthesis of extracellular matrix components. Subsequent attenuation of TGF-β is essential for maintaining collagen balance during injury recovery and preventing the excessive and disorganized accumulation of ECM, which could lead to fibrosis [36]. In fact, TGF-β levels six weeks after VD in half of the DMSC-treated animals (VD+DMSCs) approached the levels observed in the control animals. Other authors have linked fibromuscular damage in the urethra with activation of the TGF-β/Smad pathway four weeks after postpartum vaginal dilation and bilateral ovariectomy [37]. At the six-week procedure, it was observed that the increase in TGF-β had already ceased in the DMSC-treated group and was no longer rising, as the relative expression levels were closer to the control levels (Figure 4). It is evident that, in some of the animals within this group (VD+DMSCs), TGF-β expression levels have converged towards those observed in the control group. This finding could suggest that some animals in the DMSC-treated group may be entering a subsequent remodeling phase. However, this remodeling phase has not yet occurred in the untreated group (VD), which at six weeks has higher TGF-β levels than in the week one (Figure 4). This suggests that DMSC treatment (VD+DMSC) may restore the balance between ECM degradation and synthesis more rapidly, leading to enhanced remodeling compared to untreated animals (VD).
Vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF) are key proteins involved in tissue repair. Research indicates that they act synergistically [38]. VEGF is responsible for promoting the formation of new blood vessels that will perfuse the injured area, while FGF promotes cell proliferation, tissue regeneration, and modulates the inflammatory process [38,39]. In our study, an increase in both factors was observed in urethral tissue in response to VD. This early response was attenuated by DMSC treatment. At week 1, the mean expression level of these factors was approximately equivalent in DMSC-treated animals (VD+DMSCs) and controls, while untreated animals (VD) exhibited increased VEGF and FGF expression (Figure 5). By week six, expression levels were higher than in the control group in both VD groups, though the increase was statistically significant only in the untreated animals (Figure 5).
The increased expression of VEGF and FGF after vaginal distension is consistent with the activation of endogenous repair mechanisms, as previously described in these experimental models of vaginal distension [10]. Persistent VEGF expression during the later stages of healing may be a reflection of ongoing tissue remodeling [40]. Because VD causes hypoxic injury, the timing of VEGF expression is particularly important. VEGF plays a dual role in tissue repair. In the acute phase, VEGF is upregulated, increasing vascular permeability and contributing to edema and inflammation. During the reparative phase, VEGF promotes angiogenesis and tissue regeneration [41]. Our findings suggest that DMSC treatment may modulate the temporal response of VEGF. The lower VEGF expression observed in DMSC-treated animals after one week may have limited the early increase in vascular permeability and tissue edema. This, in turn, may have contributed to the improved urethral function observed at this stage. Conversely, the increase in VEGF expression at six weeks likely reflects the activation of angiogenic pathways involved in tissue remodeling and regeneration, as previously described in hypoxic injury models [42]. However, VEGF activation should be transient. Persistent elevation may indicate incomplete healing. As demonstrated in animal models, the pharmacological inhibition of angiogenesis has been shown to improve healing outcomes [43]. At week 6, the expression levels of the cytokines VEGF and FGF remain above the control levels (Figure 5). However, in some animals treated with DMSCs (VD+DMSCs), these values are approaching that of the control group. This result has not yet been observed in the untreated (VD) group, suggesting that the animals treated with DMSCs appear to have an accelerated repair process [44].
Successful tissue repair depends on the activation of multiple growth factors and cytokines, as well as their coordinated temporal expression. Previous studies have shown that wound healing improves when immunomodulatory signals, such as TGF-β, predominate in the early phase. This is followed by the sequential activation of pro-angiogenic and regenerative factors, including VEGF and FGF [45]. In untreated VD animals, TGF-β, VEGF, and FGF were induced early after injury. In contrast, DMSC-treated animals exhibited delayed increases in VEGF and FGF at week 6, which coincided with declining TGF-β expression in some animals. This temporal shift suggests that DMSC treatment may promote more coordinated progression through the phases of tissue repair.
Smooth muscle alpha-actin (α-SMA) plays a critical role in the field of molecular biology. In addition to its function as a cytoskeletal protein associated with contractility, it is also a molecular indicator of tissue repair and remodeling. The expression of α-SMA can be altered by mechanical stress and hypoxia, as occurs after VD [46]. A slight early increase in α-SMA expression was observed in several of the untreated (VD) animals, which persisted and was significant at six weeks (Figure 6). In contrast, DMSC-treated (VD+DMSCs) animals exhibited a delayed induction of α-SMA, with lower expression levels even at 6 weeks compared to untreated animals (VD) (Figure 6).
In the urethra, α-SMA expression can originate from both resident smooth muscle cells and activated myofibroblasts involved in tissue remodeling after injury [47]. An increase in α-SMA expression could indicate improved smooth muscle restoration, but it could also indicate profibrotic remodeling, highlighting the dual role of α-SMA during urethral repair. According to the literature on fibrotic disease models in the urinary and genital tracts, tissue injury promotes a reparative response, in which TGF-β stimulates the activation and differentiation of resident fibroblasts into α-SMA-expressing myofibroblasts. This results in increased ECM deposition and tissue contraction [48]. While this response may initially contribute to structural repair, persistent activation of the TGF-β/myofibroblast axis and sustained α-SMA expression can drive pathological fibrosis [49]. As with the other processes studied in this work, ensuring adequate resolution and self-limitation of the TGF-β /α-SMA response is essential to prevent the loss of normal tissue. In this context, DMSCs may contribute to fine-tuning this response, promoting timely attenuation of profibrotic signaling and favoring proper tissue remodeling.
In conclusion, our findings in the animal model of incontinence mediated by VD demonstrate the efficacy of DMSCs in promoting functional recovery. As outlined in our previous publication, DMSC administration enhances the structure of urethral tissue [9]. The present study suggests that DMSC treatment may promote faster urethral repair after VD. In the long-term study (six weeks), the untreated group (VD) demonstrated significant and sustained dysregulation of most markers associated with injury and remodeling. Increased levels of α-SMA and TGF-β could also contribute to abnormal ECM remodeling six weeks after VD induction in this group. However, in the group treated with DMSCs (VD+DMSCs), a tendency to return to normal levels was observed in some animals. These findings suggest that DMSCs may promote proper and controlled remodeling of ECM components, reducing the risk of developing fibrosis.
2.3. The Paracrine Effects of DMSCs on Suburethral Fibroblasts Isolated from Incontinent Women
In our previous work, we thoroughly characterized suburethral fibroblasts isolated from patients with stress urinary incontinence (SUI) [9]. SUI fibroblasts were found to have reduced proliferative capacity and increased senescence markers when compared to fibroblasts from women without incontinence [9]. Cellular senescence is a pro-inflammatory condition closely associated with defective tissue remodeling [50]. A notable increase in cellular senescence markers has been observed in SUI tissues [51]. Based on these observations, we hypothesized that DMSC injection could promote a rejuvenating or senescence-modulating effect on damaged urethral tissue in animals following VD [9]. Such an effect could contribute to the improved functional recovery previously observed after DMSC treatment [9], and may also explain the trend toward normalization of several biochemical markers reported in the present study. To test this hypothesis, we established an in vitro co-culture model using fibroblasts from women with SUI and DMSCs. We then evaluated the expression of senescence-associated markers in fibroblasts cultured in the presence or absence of DMSCs, focusing on pathways involved in cell cycle arrest, extracellular matrix remodeling, and tissue homeostasis.
The expression levels of the genes encoding p16 (CDKN2A) and p21 (CDKN1A) two key regulators of cell cycle, were evaluated. These genes encode two proteins that are activated in response to DNA damage, oxidative stress, and other cellular stressors. In the pathophysiology of pelvic floor disorders, particularly pelvic organ prolapse, and stress urinary incontinence (SUI), hypoxia [52], and mechanical overload [53] are two convergent stimuli capable of inducing cellular senescence by activating the p16 and p21 pathways [51]. As illustrated in Figure 7, the co-culture of SUI fibroblasts with DMSCs led to a significant reduction in p16 expression, while p21 expression levels exhibited an increase in most co-culture samples. These findings suggest that DMSCs may be exerting only a partial modulatory effect on SUI cell senescence and/or that the effects of DMSCs on senescence are independent of p21. Although p16 and p21 are both well-established markers of senescence, they are not functionally equivalent. As such, they may indicate different biological conditions [54]. Published reports generally associate p16-related senescence with a persistent, stable, and difficult-to-reverse arrest of cellular proliferative capacity. In contrast, signaling via p21 has been described as a more dynamic and potentially reversible process [55,56]. In addition to its well-established role in cell cycle arrest, p21 has been shown to contribute to skin wound healing due to its ability to regulate cell migration, differentiation, and ECM remodeling [57]. Furthermore, some of the beneficial effects of senescence on tissue formation are mediated by p21, which modulates ECM-associated genes and facilitates the synthesis of a functional ECM [58]. Therefore, DMSCs would modulate SUI cell senescence in co-culture by attenuating the more persistent senescent phenotype and promoting a transient stress response program potentially linked to tissue remodeling and repair.
Tissue homeostasis is crucially influenced by Klotho, a longevity-associated protein involved in regulating cellular aging and tissue repair processes [59]. Our research indicates that Klotho expression is elevated in SUI fibroblasts when they are co-cultured with DMSCs compared to non-co-cultured SUI fibroblasts (Figure 8). The cytoprotective effects of Klotho are linked to the modulation of oxidative stress, inflammation, and cell cycle regulatory pathways, including the p53/p21 and Wnt/β-catenin signaling pathways [59]. Klotho deficiency in fibroblasts has been linked to cellular senescence and dysregulation of extracellular matrix remodeling, as evidenced by studies on pelvic floor disorders [60,61]. Recent clinical evidence indicates a correlation between the severity of SUI and serum Klotho levels [62]. According to data from the National Health and Nutrition Examination Survey (NHANES), middle-aged and older women with severe urinary incontinence had significantly lower serum levels of α-Klotho compared to continent controls [62]. The data on reduced Klotho expression in pelvic floor disorders indicate that it contributes to impaired tissue repair and dysregulation of the extracellular matrix. The increased Klotho expression following co-culture of SUI fibroblasts with DMSCs may reflect an effect of DMSCs contributing to improved regulatory mechanisms of the ECM and tissue homeostasis. A recent study in this line indicates that FGF21-mediated activation of cardiac FGFR1-β-Klotho induces cardiomyocyte secretion of CCL2/MCP-1, thereby mitigating inflammation in the diabetic heart [63]. Additionally, increased Klotho levels suppress the proliferation of interstitial fibroblasts and attenuate kidney fibrosis. Furthermore, Klotho has protective effects against inflammation and reduces fibrosis due to its ability to block TGF-β [64]. Therefore, Klotho acts as a systemic TGF-β inhibitor by targeting the TGF-β/Smad signaling pathways, influencing fibrosis and inflammation [64]. In our study, the Klotho elevations were associated with the protective effects through ECM remodeling and, in part, through reduced TGF-β levels in the SUI-DMSC co-culture.
The matrix metalloproteases (MMPs) play a fundamental role in the physiological processes of tissue remodeling. However, their deregulated expression during senescence can contribute to the structural and functional deterioration of tissues. Matrix metalloproteinase-1 (MMP-1) and matrix metalloproteinase-2 (MMP-2) orchestrate extracellular matrix remodeling through collagen degradation, whereas tissue inhibitor of metalloproteinases-1 (TIMP-1) modulates proteolytic activity to maintain structural and cellular homeostasis. It is widely accepted that increased collagen breakdown is a pathological cause of SUI [65]. The results obtained here demonstrate that co-culture with DMSCs significantly increases MMP-2 expression in SUI fibroblasts compared to cells without co-culture (Figure 9). In the case of MMP-1, DMSCs have been observed to decrease it in SUI cells, while TIMP-1 remains unchanged or shows a slight decrease. These results indicate that DMSCs may potentially restore certain molecular pathways associated with reduced cellular senescence.
Subsequent to tissue injury, ECM remodeling is a dynamic process that relies on a precise balance between matrix synthesis and proteolytic degradation, in which matrix metalloproteinases play a central role [66]. In this context, MMP-2 functions as a key gelatinase by degrading basement membrane and extracellular matrix components, thereby facilitating matrix turnover required for tissue repair [67]. In wound healing, MMP-2 demonstrates a delayed and localized expression pattern, primarily in fibroblasts, with peak levels occurring 8–12 weeks after injury. This finding indicates a predominant role during the remodeling phase rather than the early inflammatory response [68]. The activity of MMP-2 is subject to stringent regulation by tissue inhibitors of metalloproteinases (TIMPs). Disruption of this regulatory balance can result in disorganized or chronic remodeling [67,69,70]. The collective evidence indicates that MMP-2 functions not only as a marker of tissue damage but also as an active and essential regulator of post-injury ECM reorganization [67,70].
The decrease in MMP-1 gene expression observed in cells isolated from patients with SUI after co-culture with perinatal DMSCs can be interpreted as a potentially beneficial effect in the context of tissue regeneration. Although MMP-1 is necessary in the early stages of repair to facilitate controlled collagen degradation and cell migration [71,72,73], its elevated or sustained activity has been associated with excessive extracellular matrix degradation, loss of structural support, and poor healing in tissues subjected to chronic damage [74,75]. In this context, the reduction of MMP-1 induced by perinatal DMSCs could contribute to preserving collagen integrity and promoting more stable and functional tissue remodeling, consistent with a more balanced regenerative environment [76]. These findings indicate that DMSCs have a regulatory effect on matrix remodeling, adjusting MMP-1 expression to levels more suitable for regeneration. The reduction of MMP-1 could contribute to preserving the mechanical integrity of the tissue, while the increase in MMP-2 would allow the matrix plasticity necessary for functional repair. The observation that TIMP-1 does not increase, but rather exhibits a slight decrease, suggests that DMSCs do not induce a global inhibition of proteolytic activity. Instead, they appear to promote a dynamic balance between matrix degradation and stabilization.
These results may suggest that DMSCs prevent excessive degradation and promote the remodeling necessary for regeneration [76,77]. This coordinated pattern of MMPs and TIMPs is consistent with a regulatory paracrine effect of DMSCs on the matrix microenvironment, particularly relevant in a chronically altered tissue such as the pelvic floor.
3. Materials and Methods
3.1. Isolation and Culture of Mesenchymal Stromal Cells from the Decidua
Human placentas were collected from healthy mothers in the Department of Obstetrics and Gynecology after childbirth. The Ethics Committee of Hospital Universitario 12 de Octubre approved the collection, which was performed with written informed consent. The placental membranes were dissected and the cells obtained according to the well-established protocol [16]. In summary, the tissue was digested twice with 0.05% trypsin-EDTA (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) for 30 minutes each time. The isolated cells were cultivated in Dulbecco’s Modified Eagle Medium (DMEM) containing 2 mM GlutaMAX and 0.1 mM sodium pyruvate (Gibco, Thermo Fisher Scientific, Waltham, MA), further enriched with 1% nonessential amino acids (NEAA) (STEMCELL Technologies, Vancouver, BC, Canada), 1% penicillin/streptomycin (Gibco, Thermo Fisher Scientific, Waltham, MA), 10% fetal bovine serum (FBS) (Biowest, Nuaillé, France), and 10 ng/mL epidermal growth factor (EGF) (Sigma-Aldrich, St. Louis, MO). The cells were maintained at 37 °C in a controlled environment with 5% CO2 and 95% humidity. The cells used in this study were characterized by flow cytometry and were found to be positive for the markers DC90, CD44, CD105, and CD73. They were negative for the hematopoietic markers CD34 and CD45, as previously demonstrated [9,16,78,79,80]. These cells were termed decidual-derived mesenchymal stromal cells (DMSCs).
3.2. Animal Model of Stress Urinary Incontinence Due to Childbirth
Vaginal distension was induced in twelve 17-week-old virgin female Sprague-Dawley rats (300-350 g) to mimic maternal injuries during childbirth, according to our previously published method (3). Briefly, isoflurane and ketamine-xylazine were used as anesthetics. After accommodating the vagina with lubricated bougie dilators, a modified 10-Fr Foley catheter was inserted and the balloon was inflated with 2 mL of PBS over half an hour, later increasing to 3 mL. The rats were positioned in the anti-Trendelenburg position, and the balloon was maintained for six hours. Post-procedure pain was managed with subcutaneous administration of buprenorphine. A Foley catheter was inserted into the vagina of the sham animals for six hours, though the balloon was not inflated.
3.3. DMSC Treatment
After VD, the animals were randomly assigned to two groups of six: a control group and a DMSC-treated group. The DMSC-treated group received two doses of DMSCs resuspended in Hank’s Balanced Salt Solution (HBSS), at a concentration of 2x106 DMSCs per dose, administered two days apart. The doses were administered one day after VD and two days apart. The control group received injections of the same volume of HBSS without cells. The DMSC or HBSS injections were administered into the periurethral region via bilateral injections at the 3 and 9 o’clock positions.
3.4. Leak Point Pressure (LPP) Measurement
The Leak Point Pressure (LPP) measurement was performed while the animals were under anesthesia with inhaled isoflurane (5% for induction and 2% for maintenance), as previously described [9]. A PE-50 catheter was transurethrally inserted into the bladder. After emptying the bladder, its capacity was determined by filling it with PBS and methylene blue until leakage occurred. Three filling cycles were completed to calculate the average capacity. To measure the minimum pressure causing leakage (LPP), the bladder was filled to 50% capacity. A data acquisition device was connected to the catheter, and the intravesical pressure was recorded using AcqKnowledge software (Biopac). The LPP was determined by manually applying abdominal pressure [81]. Three measurements were carried out for each animal, and the mean pressure was calculated from these measurements.
3.5. Isolation and Culture of Fibroblasts from Suburethral Tissue of Patients with Stress Urinary Incontinence (SUI)
This study examined eight suburethral tissue samples obtained from patients with SUI. Demographic and clinical characteristics of the patients are included in Table S1. The samples were obtained during a surgical procedure to insert a suburethral mesh sling in women with SUI at the Pelvic Floor Unit of the Department of Obstetrics and Gynecology at 12 de Octubre University Hospital. The Ethics Committee of the 12 de Octubre University Hospital approved the protocol, and informed consent was obtained from the patients. The cells were isolated by digesting the tissue enzymatically with 100 units/μL of collagenase type II (GIBCO) in HBSS for 16 hours. The digestion product was then centrifuged at 250 g for 10 minutes. The resulting pellet was then resuspended in complete DMEM medium for cell seeding. The cells were maintained at 37 °C with 5% CO2 throughout the expansion process.
3.6. Indirect DMSC-SUI Fibroblast Co-Culture System
To evaluate the effects of DMSCs on SUI fibroblasts in vitro, an indirect co-culture system was set up using Transwell inserts (Costar) with 0.4-μm-pore-size polycarbonate membranes. On the same day, fibroblasts were seeded in 6-well plates at a density of 3 × 105 cells/well and DMSCs were seeded in the inserts at a density of 1.5 × 105 cells/insert. After 24 hours, the inserts were transferred to the fibroblast-containing wells to initiate co-culture. The cells were then maintained under co-culture conditions for an additional 24 hours prior to analysis.
3.7. Gene Expression Analysis
3.7.1. RNA Isolation
Total RNA was extracted from the samples using the NZY Total RNA Isolation Kit (NZYTech, Ltd., Lisbon, Portugal) according to the manufacturer’s instructions. The concentration and purity of the RNA were assessed spectrophotometrically using a Nanodrop (Thermo Scientific), and the extracted RNA was stored at −80 °C until further use.
3.7.2. Retrotranscription
Complementary DNA (cDNA) was synthesized from 1 µg of total RNA using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) according to the manufacturer’s protocol. The resulting cDNA was stored at −20 °C and used as a template for subsequent gene expression analyses.
3.7.3. Quantitative Polymerase Chain Reaction (qPCR)
Gene expression analysis in urethral tissues was performed using quantitative PCR (qPCR) with TaqMan probes (Table S2), Fast TaqMan Master Mix (Applied Biosystems, Fisher Scientific, Spain), and a 7500 Fast Real-Time PCR System (Applied Biosystems, CA, USA). PCR was performed under the following cycling conditions: an initial denaturation step at 95 °C for 2 min, followed by 40 cycles of denaturation at 95 °C for 3 s and annealing/extension at 60 °C for 30 s. TaqMan probes used in the study are included in Table S2. Target gene expression was normalized to TATA-box binding protein (TBP) and Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as reference genes by calculating the geometric mean of their expression levels. Relative gene expression levels were calculated using the 2^−ΔΔCt method, with the mean expression level of the control animals used as the calibrator.
For the study of SUI fibroblasts, quantitative PCR was performed using Fast SYBR Green Master Mix (Applied Biosystems, Fisher Scientific, Spain) and the 7500 Fast Real-Time PCR System (Applied Biosystems, CA, USA). PCR was performed under the following cycling conditions: an initial denaturation step at 95 °C for 20 s, followed by 40 cycles of 95 °C for 3 s and annealing/extension at 60 °C for 30 s. Target gene expression was normalized to TBP as the reference gene. Relative gene expression levels were calculated using the 2^−ΔΔCt method. Gene expression in cocultured cells was compared with that in the corresponding non-cocultured cells. Primer sequences are listed in Table S3.
Klotho analysis in SUI fibroblasts was performed using Taqman technology with Fast TaqMan Master Mix (Applied Biosystems, Fisher Scientific, Spain) and a 7500 Fast Real-Time PCR System (Applied Biosystems, CA, USA). The conditions of PCR were the same as those described for urethral gene expression. Target gene expression was normalized to TBP as the reference gene. Relative gene expression levels were calculated using the 2^−ΔΔCt method. Gene expression in cocultured cells was compared with that in the corresponding non-cocultured cells.
3.8. Statistical Analysis
Data were analyzed using GraphPad Prism software (version 8.0; GraphPad Software, San Diego, CA, USA). Data are presented as mean ± standard deviation (SD). Normality was assessed prior to statistical testing. Differences between groups were analyzed using an unpaired Student’s t-test for normally distributed data or the Mann–Whitney U test for non-normally distributed data, as appropriate. A p value < 0.05 was considered statistically significant.
4. Conclusions
Treatment with DMSCs has been shown to promote functional recovery in animals subjected to vaginal distension (VD), a model of childbirth-related damage. Functional improvements were evident at one and six-weeks post-treatment. Transcriptomic analysis indicated that all animals after VD exhibited damage-repair responses, though only some recovered continence. The majority of displayed persistent changes in gene expression were linked to tissue remodeling, suggesting ongoing repair even six weeks after injury. While significant functional recovery differences were noted between treated and untreated groups, molecular differences were less pronounced, revealing individual variability reflective of women’s varying susceptibility to postpartum urinary incontinence and recovery processes. Gene analysis indicated that DMSCs influence repair dynamics, promoting accelerated regeneration without altering the repair process itself. At six weeks, untreated animals demonstrated sustained elevated growth factor levels, while treated animals exhibited a trend towards normalization, suggesting accelerated regeneration. However, the VD model has limitations due to its simplified representation of human childbirth, resulting in a different neuromuscular injury and quicker recovery, which may underestimate human chronic urinary incontinence. Future studies should combine this model with pudendal nerve injury to create a more accurate simulation of postpartum incontinence.
The findings suggest that DMSCs have therapeutic potential in stress urinary incontinence (SUI) through paracrine effects that improve the regenerative environment by reducing senescence in suburethral fibroblasts and enhancing tissue dynamics, as demonstrated by changes in molecular markers like p16, p21, and Klotho. In addition, DMSCs have been shown to restore the balance of the extracellular matrix. This, in turn, promotes structural integrity while enabling controlled turnover. In summary, DMSCs are a promising disease-modifying therapy that enhances tissue repair in SUI, supporting improved clinical outcomes as a regenerative intervention.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1. Demographic and clinical characteristics of the patients with stress urinary incontinence (SUI) included in the study; Table S2. Primer sequences for human gene expression analysis; Table S3. TaqMan probes used in the study.
Author Contributions
Conceptualization, A.I.F and P.d.l.T.; methodology, P.d.l.T. and J.C.; P.d.l.T., J.C., and A.I.F; resources, L.F., A.R.M-C, A.G., J.M.P., E.M.; writing—original draft preparation, P.d.l.T., M.J.M.-J., J.J.M., and A.I.F; funding acquisition, A.I.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research has also been funded by INSTITUTO DE SALUD CARLOS III (ISCIII), through the projects PI15/01803, PI18/01278, and DTS22-00166, and co-funded by the European Union. This research was also funded by Grant PID2022-140785OB-I00 funded by MCIN/AEI/10.13039/501100011033/ FEDER, UE. Moreover, the project has received two grants for the hiring of Laboratory Technicians from the Comunidad de Madrid in 2016 and 2020 (Youth Employment Operational Program (YEI): Contracts for Research assistants and laboratory technicians of the Community of Madrid and the European Social Fund, PEJ16/MED/TL-1388 and PEJ-2020-TL/BMD-19230), and a grant for the contract of a research assistant (PEJ-2024-AI/SAL-GL-33566).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Ethics Committee of HOSPITAL UNIVERSITARIO 12 DE OCTUBRE (Nº CEI: 16/275, date of approval 27 September 2016; Nº CEIm: 18/525, date of approval 15 January 2019; Nº CEIm: 22/568, date of approval 17 January 2023).
The: animal study protocol was approved by the Bioethics Committee of the Autonomous University of Madrid and the Directorate General of Agriculture and Environment, Community of Madrid (protocol code PROEX 109/19 and date of approval 2019/05/16) and (protocol code PROEX 008.3/24 and date of approval 2024/02/09), according to the Guide for the Care and Use of Laboratory Animals and Ethical Care and Welfare Guidelines (2013/175) of the European Union (2010/63/EU).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
During the preparation of this manuscript, the author(s) utilized ChatGPT, DeepL, and M365 Copilot to refine the text’s language and assist in developing the concept for the graphical abstract. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| SUI | Stress urinary incontinence |
| VD | Vaginal distension |
| DMSCs | Decidua-derived mesenchymal stromal cells |
| LPP | Leak point pressure |
| ECM | Extracellular matrix |
| CCL2 | Monocyte Chemoattractant Protein-1 (MCP-1) |
| CCL7 | Monocyte Chemoattractant Protein-3 (MCP-3) |
| TGF-β | Transforming growth factor beta |
| VEGF | Vascular endothelial growth factor |
| FGF | Fibroblast growth factor |
| α-SMA | Smooth muscle alpha-actin |
| MMPs | Matrix metalloproteases |
| TIMP-1 | Tissue inhibitor of metalloproteinases 1 |
References
- Luber, K.M. The definition, prevalence, and risk factors for stress urinary incontinence. Rev. Urol. 2004, 6 Suppl 3, S3–9. [Google Scholar]
- Dutta, I.; Pathak, B. Quality of Life of Patients With Stress Urinary Incontinence Before and After Transobturator Tape Surgery: An Observational Study. Cureus 2024, 16, e76092. [Google Scholar] [CrossRef]
- Hampel, C.; Artibani, W.; Espuna Pons, M.; Haab, F.; Jackson, S.; Romero, J.; Gavart, S.; Papanicolaou, S. Understanding the burden of stress urinary incontinence in Europe: a qualitative review of the literature. Eur. Urol. 2004, 46, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Herschorn, S. Female pelvic floor anatomy: the pelvic floor, supporting structures, and pelvic organs. Rev. Urol. 2004, 6 Suppl 5, S2–S10. [Google Scholar] [PubMed]
- Guler, Z.; Roovers, J.P. Role of Fibroblasts and Myofibroblasts on the Pathogenesis and Treatment of Pelvic Organ Prolapse. Biomolecules 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Vetuschi, A.; D’Alfonso, A.; Sferra, R.; Zanelli, D.; Pompili, S.; Patacchiola, F.; Gaudio, E.; Carta, G. Changes in muscularis propria of anterior vaginal wall in women with pelvic organ prolapse. Eur. J. Histochem 2016, 60, 2604. [Google Scholar] [CrossRef] [PubMed]
- Damaser, M.S.; Whitbeck, C.; Chichester, P.; Levin, R.M. Effect of vaginal distension on blood flow and hypoxia of urogenital organs of the female rat. J. Appl. Physiol. (1985) 2005, 98, 1884–1890. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.H.; Gustilo-Ashby, A.M.; Salcedo, L.B.; Pan, H.Q.; Sypert, D.F.; Butler, R.S.; Damaser, M.S. Electrophysiological function during voiding after simulated childbirth injuries. Exp. Neurol. 2009, 215, 342–348. [Google Scholar] [CrossRef] [PubMed]
- De La Torre, P.; Perez-Lorenzo, M.J.; Alcazar-Garrido, A.; Collado, J.; Martinez-Lopez, M.; Forcen, L.; Masero-Casasola, A.R.; Garcia, A.; Gutierrez-Velez, M.C.; Medina-Polo, J.; et al. Perinatal mesenchymal stromal cells of the human decidua restore continence in rats with stress urinary incontinence induced by simulated birth trauma and regulate senescence of fibroblasts from women with stress urinary incontinence. Front Cell Dev. Biol. 2022, 10, 1033080. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Liu, G.; Li, M.; Xiao, N.; Daneshgari, F. Recovery of continence function following simulated birth trauma involves repair of muscle and nerves in the urethra in the female mouse. Eur. Urol. 2010, 57, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.Q.; Kerns, J.M.; Lin, D.L.; Liu, S.; Esparza, N.; Damaser, M.S. Increased duration of simulated childbirth injuries results in increased time to recovery. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1738–1744. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Shindel, A.W.; Banie, L.; Deng, D.; Wang, G.; Hayashi, N.; Lin, C.S.; Lue, T.F. Molecular mechanisms related to parturition-induced stress urinary incontinence. Eur. Urol. 2009, 55, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Wood, H.M.; Kuang, M.; Woo, L.; Hijaz, A.; Butler, R.S.; Penn, M.; Rackley, R.; Damaser, M.S. Cytokine expression after vaginal distention of different durations in virgin Sprague-Dawley rats. J. Urol. 2008, 180, 753–759. [Google Scholar] [CrossRef] [PubMed]
- De La Torre, P.; Pérez-Lorenzo, M.J.; Flores, A.I. Human Placenta-Derived Mesenchymal Stromal Cells: A Review from Basic Research to Clinical Applications. In Stromal Cells-Structure, Function, and Therapeutic Implications; IntechOpen, 2018. [Google Scholar]
- Pinzariu, A.C.; Moscalu, R.; Soroceanu, R.P.; Maranduca, M.A.; Drochioi, I.C.; Vlasceanu, V.I.; Timofeiov, S.; Timofte, D.V.; Huzum, B.; Moscalu, M.; et al. The Therapeutic Use and Potential of MSCs: Advances in Regenerative Medicine. Int. J. Mol. Sci. 2025, 26. [Google Scholar] [CrossRef] [PubMed]
- Macias, M.I.; Grande, J.; Moreno, A.; Dominguez, I.; Bornstein, R.; Flores, A.I. Isolation and characterization of true mesenchymal stem cells derived from human term decidua capable of multilineage differentiation into all 3 embryonic layers. Am. J. Obstet. Gynecol. 2010, 203, 495 e499–495 e423. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Adit, G.N.; Srikanth, K.; Harithpriya, K.; Ganesan, K.; Ramkumar, K.M. Modulation of Chemokine Activity for Enhanced Angiogenesis and Tissue Regeneration in Chronic Wounds. Int. J. Mol. Sci. 2026, 27. [Google Scholar] [CrossRef] [PubMed]
- Lenis, A.T.; Kuang, M.; Woo, L.L.; Hijaz, A.; Penn, M.S.; Butler, R.S.; Rackley, R.; Damaser, M.S.; Wood, H.M. Impact of parturition on chemokine homing factor expression in the vaginal distention model of stress urinary incontinence. J. Urol. 2013, 189, 1588–1594. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, V.L.S.; Proost, P.; Struyf, S. Chemokines in the resolution of inflammation: key players and targets for therapeutic modulation. Front Immunol. 2025, 16, 1717666. [Google Scholar] [CrossRef] [PubMed]
- Andreas, K.; Sittinger, M.; Ringe, J. Toward in situ tissue engineering: chemokine-guided stem cell recruitment. Trends Biotechnol. 2014, 32, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, E.J.; Arms, L.; Vizzard, M.A. The role(s) of cytokines/chemokines in urinary bladder inflammation and dysfunction. BioMed Res. Int. 2014, 2014, 120525. [Google Scholar] [CrossRef] [PubMed]
- Woo, L.L.; Hijaz, A.; Kuang, M.; Penn, M.S.; Damaser, M.S.; Rackley, R.R. Over expression of stem cell homing cytokines in urogenital organs following vaginal distention. J. Urol. 2007, 177, 1568–1572. [Google Scholar] [CrossRef] [PubMed]
- Ridiandries, A.; Tan, J.T.M.; Bursill, C.A. The Role of Chemokines in Wound Healing. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Hijaz, A.K.; Grimberg, K.O.; Tao, M.; Schmotzer, B.; Sadeghi, Z.; Lin, Y.H.; Kavran, M.; Ozer, A.; Xiao, N.; Daneshgari, F. Stem cell homing factor, CCL7, expression in mouse models of stress urinary incontinence. Female Pelvic Med. Reconstr. Surg. 2013, 19, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Ito, H.; Kitaori, T.; Murata, K.; Shibuya, H.; Furu, M.; Yoshitomi, H.; Fujii, T.; Yamamoto, K.; Matsuda, S. MCP/CCR2 signaling is essential for recruitment of mesenchymal progenitor cells during the early phase of fracture healing. PLoS ONE 2014, 9, e104954. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.H.; Ji, L.X.; Li, H.Y.; Song, Q.X.; Bano, Y.; Chen, L.; Liu, G.; Wang, M. Combined Treatment With CCR1-Overexpressing Mesenchymal Stem Cells and CCL7 Enhances Engraftment and Promotes the Recovery of Simulated Birth Injury-Induced Stress Urinary Incontinence in Rats. Front Surg. 2020, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.S.; Lue, T.F. Stem cell therapy for stress urinary incontinence: a critical review. Stem Cells Dev. 2012, 21, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, L.; Sopko, N.; Jiang, H.H.; Damaser, M.; Penn, M.; Zutshi, M. Chemokine upregulation in response to anal sphincter and pudendal nerve injury: potential signals for stem cell homing. Int. J. Colorectal Dis. 2011, 26, 1577–1581. [Google Scholar] [CrossRef] [PubMed]
- Schenk, S.; Mal, N.; Finan, A.; Zhang, M.; Kiedrowski, M.; Popovic, Z.; McCarthy, P.M.; Penn, M.S. Monocyte chemotactic protein-3 is a myocardial mesenchymal stem cell homing factor. Stem Cells 2007, 25, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Liu, Y.; Peng, X.; Li, Z.; Lei, M. Chronic Wound Healing: Research Advances from Pathological Mechanisms to Natural Herbal Active Ingredients and Material Delivery Systems. Molecules 2026, 31. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Guo, H.; Dong, X.; Wang, Z.; Yang, Z.; Shang, Q.; Wang, Q. Regulation of inflammation during wound healing: the function of mesenchymal stem cells and strategies for therapeutic enhancement. Front Pharmacol. 2024, 15, 1345779. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zheng, L.; Yuan, Q.; Zhen, G.; Crane, J.L.; Zhou, X.; Cao, X. Transforming growth factor-beta in stem cells and tissue homeostasis. Bone Res. 2018, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Tie, Y.; Tang, F.; Peng, D.; Zhang, Y.; Shi, H. TGF-beta signal transduction: biology, function and therapy for diseases. Mol. BioMed 2022, 3, 45. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.B.; Sporn, M.B.; Assoian, R.K.; Smith, J.M.; Roche, N.S.; Wakefield, L.M.; Heine, U.I.; Liotta, L.A.; Falanga, V.; Kehrl, J.H.; et al. Transforming growth factor type beta: rapid induction of fibrosis and angiogenesis in vivo and stimulation of collagen formation in vitro. Proc. Natl. Acad. Sci. U S A 1986, 83, 4167–4171. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Sheppard, D.; Chapman, H.A. TGF-beta1 Signaling and Tissue Fibrosis. Cold Spring Harb. Perspect. Biol. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Y.; Cui, W.S.; Zhou, F.; Gao, Z.Z.; Xin, H.; Liu, T.; Li, W.R.; Gong, Y.Q.; Bai, G.Y.; Guo, Y.L.; et al. Pathology of urethral fibromuscular system related to parturition-induced stress urinary incontinence and TGF-beta1/Smad pathway. Mol. Cell Biochem 2012, 364, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, M.J.; Kumar, A.; Lee, H.W.; Yang, Y.; Kim, Y. Vascular endothelial growth factor signaling in health and disease: from molecular mechanisms to therapeutic perspectives. Signal Transduct. Target Ther. 2025, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Khan, A.W.; Kim, M.S.; Choi, S. The Role of Fibroblast Growth Factor (FGF) Signaling in Tissue Repair and Regeneration. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Nissen, N.N.; Polverini, P.J.; Koch, A.E.; Volin, M.V.; Gamelli, R.L.; DiPietro, L.A. Vascular endothelial growth factor mediates angiogenic activity during the proliferative phase of wound healing. Am. J. Pathol. 1998, 152, 1445–1452. [Google Scholar] [PubMed]
- Crafts, T.D.; Jensen, A.R.; Blocher-Smith, E.C.; Markel, T.A. Vascular endothelial growth factor: therapeutic possibilities and challenges for the treatment of ischemia. Cytokine 2015, 71, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Phull, H.S.; Pan, H.Q.; Butler, R.S.; Hansel, D.E.; Damaser, M.S. Vulnerability of continence structures to injury by simulated childbirth. Am. J. Physiol. Ren. Physiol. 2011, 301, F641–649. [Google Scholar] [CrossRef] [PubMed]
- DiPietro, L.A. Angiogenesis and wound repair: when enough is enough. J. Leukoc. Biol. 2016, 100, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Galiano, R.D.; Tepper, O.M.; Pelo, C.R.; Bhatt, K.A.; Callaghan, M.; Bastidas, N.; Bunting, S.; Steinmetz, H.G.; Gurtner, G.C. Topical vascular endothelial growth factor accelerates diabetic wound healing through increased angiogenesis and by mobilizing and recruiting bone marrow-derived cells. Am. J. Pathol. 2004, 164, 1935–1947. [Google Scholar] [CrossRef] [PubMed]
- Jimi, S.; Jaguparov, A.; Nurkesh, A.; Sultankulov, B.; Saparov, A. Sequential Delivery of Cryogel Released Growth Factors and Cytokines Accelerates Wound Healing and Improves Tissue Regeneration. Front Bioeng. Biotechnol. 2020, 8, 345. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Guan, H.; Liu, J.Q.; Zheng, Z.; Zhou, Q.; Zhang, J.; Su, L.L.; Hu, D.H. Hypoxia drives the transition of human dermal fibroblasts to a myofibroblast-like phenotype via the TGF-beta1/Smad3 pathway. Int. J. Mol. Med. 2017, 39, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.; Woods, E.L.; Dally, J.; Kong, D.; Steadman, R.; Moseley, R.; Midgley, A.C. Myofibroblasts: Function, Formation, and Scope of Molecular Therapies for Skin Fibrosis. Biomolecules 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Rahardjo, H.E.; Marker, V.; Tsikas, D.; Kuczyk, M.A.; Uckert, S.; Bannowsky, A. Fibrotic Diseases of the Human Urinary and Genital Tract: Current Understanding and Potential Strategies for Treatment. J. Clin. Med. 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Lyu, L.; Xing, C.; Chen, Y.; Hu, S.; Wang, M.; Ai, Z. The pivotal role of TGF-beta/Smad pathway in fibrosis pathogenesis and treatment. Front Oncol. 2025, 15, 1649179. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Yamamoto, S.; Chikenji, T.S. Role of cellular senescence in inflammation and regeneration. Inflamm. Regen. 2024, 44, 28. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Chen, M.; Li, L.; Chen, L.; Tian, X.; Huang, X.; Zhu, F.; Li, B.; Hong, L. PIM1 induces hypoxia-related fibroblast senescence in a mouse model of stress urinary incontinence. PLoS ONE 2025, 20, e0335501. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Nepovimova, E.; Heger, Z.; Valko, M.; Wu, Q.; Kuca, K.; Adam, V. Role of hypoxia in cellular senescence. Pharmacol. Res. 2023, 194, 106841. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Z.; Li, T.; Xu, H.; Zhang, H. Senescence in osteoarthritis: from mechanism to potential treatment. Arthritis Res. Ther. 2022, 24, 174. [Google Scholar] [CrossRef] [PubMed]
- Moiseeva, V.; Cisneros, A.; Cobos, A.C.; Tarrega, A.B.; Onate, C.S.; Perdiguero, E.; Serrano, A.L.; Munoz-Canoves, P. Context-dependent roles of cellular senescence in normal, aged, and disease states. FEBS J. 2023, 290, 1161–1185. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Chen, S.; Yi, Z.; Zhao, R.; Zhu, J.; Ding, S.; Wu, J. The role of p21 in cellular senescence and aging-related diseases. Mol. Cells 2024, 47, 100113. [Google Scholar] [CrossRef] [PubMed]
- Roger, L.; Tomas, F.; Gire, V. Mechanisms and Regulation of Cellular Senescence. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Kozyrska, K.; Pilia, G.; Vishwakarma, M.; Wagstaff, L.; Goschorska, M.; Cirillo, S.; Mohamad, S.; Gallacher, K.; Carazo Salas, R.E.; Piddini, E. p53 directs leader cell behavior, migration, and clearance during epithelial repair. Science 2022, 375, eabl8876. [Google Scholar] [CrossRef] [PubMed]
- Brauer, E.; Lange, T.; Keller, D.; Gorlitz, S.; Cho, S.; Keye, J.; Gossen, M.; Petersen, A.; Kornak, U. Dissecting the influence of cellular senescence on cell mechanics and extracellular matrix formation in vitro. Aging Cell 2023, 22, e13744. [Google Scholar] [CrossRef] [PubMed]
- Hajare, A.D.; Dagar, N.; Gaikwad, A.B. Klotho antiaging protein: molecular mechanisms and therapeutic potential in diseases. Mol. BioMed 2025, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Fang, M.; Luo, J.; Fan, X.; Zhao, Y. Klotho Regulates the Extracellular Matrix Metabolism via TGF-beta1/Wnt Signaling Pathway of Fibroblasts in Pelvic Organ Prolapse. J. Biochem Mol. Toxicol. 2025, 39, e70358. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Qin, M.; Fan, B.; Chen, X. Klotho Protein Reduced the Expression of Matrix Metalloproteinase-1 (MMP-1) and Matrix Metalloproteinase-3 (MMP-3) in Fibroblasts from Patients with Pelvic Organ Prolapse (POP) by Down-Regulating the Phosphorylation of ERK1/2. Med. Sci. Monit. 2019, 25, 3815–3824. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Song, Y.; Liang, H.; He, W.; Chen, Z.; Yang, L.; Yang, C.; Du, R. The relationship between serum alpha-klotho levels and urinary incontinence in middle-aged and older females: insights from NHANES. Sci. Rep. 2024, 14, 28667. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Gare, S.R.; Ruiz-Velasco, A.; Miller, J.M.; Abouleisa, R.R.E.; Ou, Q.; Shen, J.; Soran, H.; Mohamed, T.M.A.; Liu, W. FGF21/FGFR1-beta-KL cascade in cardiomyocytes modulates angiogenesis and inflammation under metabolic stress. Heliyon 2023, 9, e14952. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Han, D.; Zhao, C.; Yang, F.; Wang, Z.; Gao, Y.; Jin, M.; Tao, R. New insights into the role of Klotho in inflammation and fibrosis: molecular and cellular mechanisms. Front Immunol. 2024, 15, 1454142. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.H.; Wen, Y.; Li, H.; Polan, M.L. Collagen metabolism and turnover in women with stress urinary incontinence and pelvic prolapse. Int. Urogynecol J. Pelvic Floor Dysfunct. 2002, 13, 80–87; discussion 87. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, I. Extracellular matrix remodelling: the role of matrix metalloproteinases. J. Pathol. 2003, 200, 448–464. [Google Scholar] [CrossRef] [PubMed]
- Wolosowicz, M.; Prokopiuk, S.; Kaminski, T.W. The Complex Role of Matrix Metalloproteinase-2 (MMP-2) in Health and Disease. Int. J. Mol. Sci. 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Gillard, J.A.; Reed, M.W.; Buttle, D.; Cross, S.S.; Brown, N.J. Matrix metalloproteinase activity and immunohistochemical profile of matrix metalloproteinase-2 and -9 and tissue inhibitor of metalloproteinase-1 during human dermal wound healing. Wound Repair Regen. 2004, 12, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, B.; Hakkinen, L.; Larjava, H. Proteolytic events of wound-healing--coordinated interactions among matrix metalloproteinases (MMPs), integrins, and extracellular matrix molecules. Crit. Rev. Oral Biol. Med. 2001, 12, 373–398. [Google Scholar] [CrossRef] [PubMed]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Rohani, M.G.; Parks, W.C. Matrix remodeling by MMPs during wound repair. Matrix Biol. 2015, 44-46, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Caley, M.P.; Martins, V.L.; O’Toole, E.A. Metalloproteinases and Wound Healing. Adv. Wound Care (New Rochelle) 2015, 4, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-F.; Esam, G.; Lin, Y.-C.; Chung, R.-J. From destruction to protection: rethinking MMP-1 in skin aging and anti-aging strategies. Arch. Dermatol. Res. 2026, 318, 56. [Google Scholar] [CrossRef]
- Bellayr, I.H.; Mu, X.; Li, Y. Biochemical insights into the role of matrix metalloproteinases in regeneration: challenges and recent developments. Future Med. Chem. 2009, 1, 1095–1111. [Google Scholar] [CrossRef] [PubMed]
- Brew, K.; Nagase, H. The tissue inhibitors of metalloproteinases (TIMPs): an ancient family with structural and functional diversity. Biochim Biophys. Acta 2010, 1803, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Coto-Vilcapoma, A.; Sanchez-Carretero, L.; Arenas-Gonzalez, D.; Molina, J.A.; Moran-Jimenez, M.J.; Merino, J.J.; Torre, P.; Flores, A.I. Protective Effect of Placental Mesenchymal Stromal Cells in an In Vitro Model of Parkinson’s Disease Using Differentiated Neuroblastoma Cells. Int. J. Mol. Sci. 2026, 27. [Google Scholar] [CrossRef] [PubMed]
- de la Torre, P.; Fernandez-de la Torre, M.; Flores, A.I. Premature senescence of placental decidua cells as a possible cause of miscarriage produced by mycophenolic acid. J. BioMed Sci. 2021, 28, 3. [Google Scholar] [CrossRef] [PubMed]
- Serrano, L.J.; de la Torre, P.; Liras, A.; Flores, A.I. Cell therapy for factor V deficiency: An approach based on human decidua mesenchymal stem cells. BioMed Pharmacother. 2021, 142, 112059. [Google Scholar] [CrossRef] [PubMed]
- Damaser, M.S.; Kim, F.J.; Minetti, G.M. Methods of testing urethral resistance in the female rat. Adv. Exp. Med. Biol. 2003, 539, 831–839. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Periurethral injection of DMSCs significantly improved continence in rats one week after vaginal distension (VD). (A) Flowchart of the design for leakage point pressure (LPP) measurements, vaginal distension, and DMSC treatment. (B) Leak point pressure (LPP) was measured in animals before and after vaginal distension (VD) and treatment with DMSCs (VD+DMSCs) or a saline solution (VD). “Basal” refers to the LPP measurements taken during the two days prior to VD. “Post-injury” refers to the LPP measurement taken one day after VD and before the injection of DMSCs. “Post-treatment” refers to the LPP measurements taken one week after injury, and following two injections of DMSCs or saline. A significant decrease in LPP was observed in the saline group (VD) and in the DMSC treated group (VD+DMSCs) one day after VD (post-injury). DMSC injection restores normal LPP values in comparison with rats that have been injected with saline (post-treatment). One week after the procedure, the incidence of incontinence was comparable between the DMSC group and the control group. Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01; ns, not significant.
Figure 1.
Periurethral injection of DMSCs significantly improved continence in rats one week after vaginal distension (VD). (A) Flowchart of the design for leakage point pressure (LPP) measurements, vaginal distension, and DMSC treatment. (B) Leak point pressure (LPP) was measured in animals before and after vaginal distension (VD) and treatment with DMSCs (VD+DMSCs) or a saline solution (VD). “Basal” refers to the LPP measurements taken during the two days prior to VD. “Post-injury” refers to the LPP measurement taken one day after VD and before the injection of DMSCs. “Post-treatment” refers to the LPP measurements taken one week after injury, and following two injections of DMSCs or saline. A significant decrease in LPP was observed in the saline group (VD) and in the DMSC treated group (VD+DMSCs) one day after VD (post-injury). DMSC injection restores normal LPP values in comparison with rats that have been injected with saline (post-treatment). One week after the procedure, the incidence of incontinence was comparable between the DMSC group and the control group. Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01; ns, not significant.

Figure 2.
Expression of genes related to proliferation (Ki67) and apoptosis (Bax and Bcl-2) in rats one and six weeks after vaginal distension (VD). (A) Ki67 gene expression, as determined by real-time qPCR. (B) Gene expression of Bax and Bcl-2, as determined by qPCR and expressed as the Bax/Bcl-2 ratio. The genes were analyzed in animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01; ***p < 0.001.
Figure 2.
Expression of genes related to proliferation (Ki67) and apoptosis (Bax and Bcl-2) in rats one and six weeks after vaginal distension (VD). (A) Ki67 gene expression, as determined by real-time qPCR. (B) Gene expression of Bax and Bcl-2, as determined by qPCR and expressed as the Bax/Bcl-2 ratio. The genes were analyzed in animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01; ***p < 0.001.

Figure 3.
Expression of CCL2 and CCL7 genes in rats one or six weeks after vaginal distension (VD). (A) CCL2 gene expression, as determined by real-time qPCR. (B) Gene expression of CCL7, as determined by qPCR. The genes were analyzed in animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01; ***p < 0.001.
Figure 3.
Expression of CCL2 and CCL7 genes in rats one or six weeks after vaginal distension (VD). (A) CCL2 gene expression, as determined by real-time qPCR. (B) Gene expression of CCL7, as determined by qPCR. The genes were analyzed in animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01; ***p < 0.001.

Figure 4.
Expression of TGF-β gene in rats, one and six weeks after vaginal distension (VD). The TGF-β gene was analyzed by qPCR in the urethral tissue of animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01; ***p < 0.001.
Figure 4.
Expression of TGF-β gene in rats, one and six weeks after vaginal distension (VD). The TGF-β gene was analyzed by qPCR in the urethral tissue of animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01; ***p < 0.001.

Figure 5.
Expression of VEGF and FGF genes in rats one or six weeks after vaginal distension (VD). (A) VEGF gene expression, as determined by real-time qPCR. (B) Gene expression of FGF, as determined by qPCR. The genes were analyzed in animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01.
Figure 5.
Expression of VEGF and FGF genes in rats one or six weeks after vaginal distension (VD). (A) VEGF gene expression, as determined by real-time qPCR. (B) Gene expression of FGF, as determined by qPCR. The genes were analyzed in animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05; **p < 0.01.

Figure 6.
Expression of smooth muscle alpha-actin (α-SMA) gene in rats one or six weeks after vaginal distension (VD). α-SMA gene expression was determined by real-time qPCR. The gene was analyzed in animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05.
Figure 6.
Expression of smooth muscle alpha-actin (α-SMA) gene in rats one or six weeks after vaginal distension (VD). α-SMA gene expression was determined by real-time qPCR. The gene was analyzed in animals that were either control (CONTROL), untreated (VD), or DMSC-treated (VD+DMSC). Data are presented as the mean ± standard deviation (SD). *p < 0.05.

Figure 7.
Analysis of senescence markers in SUI fibroblasts after co-culture with DMSCs. Paracrine effect of DMSCs on SUI fibroblast co-cultured with on Transwell® culture inserts. Fold changes in CDKN1A (p16) and CDKN2A (p21) in SUI fibroblasts co-cultured with DMSC (CC) in relation to non-co-cultured SUI fibroblasts (SUI) (n=8). Data are presented as the mean ± standard deviation (SD). *p < 0.05.
Figure 7.
Analysis of senescence markers in SUI fibroblasts after co-culture with DMSCs. Paracrine effect of DMSCs on SUI fibroblast co-cultured with on Transwell® culture inserts. Fold changes in CDKN1A (p16) and CDKN2A (p21) in SUI fibroblasts co-cultured with DMSC (CC) in relation to non-co-cultured SUI fibroblasts (SUI) (n=8). Data are presented as the mean ± standard deviation (SD). *p < 0.05.

Figure 8.
Analysis of Klotho in SUI fibroblasts after co-culture with DMSCs. Paracrine effect of DMSCs on SUI fibroblast co-cultured with on Transwell® culture inserts. Fold changes in Klotho gene expression in SUI fibroblasts co-cultured with DMSCs (CC) relative to the corresponding SUI fibroblasts (SUI) (n=7). Data are presented as the mean ± standard deviation (SD).
Figure 8.
Analysis of Klotho in SUI fibroblasts after co-culture with DMSCs. Paracrine effect of DMSCs on SUI fibroblast co-cultured with on Transwell® culture inserts. Fold changes in Klotho gene expression in SUI fibroblasts co-cultured with DMSCs (CC) relative to the corresponding SUI fibroblasts (SUI) (n=7). Data are presented as the mean ± standard deviation (SD).

Figure 9.
Analysis of matrix metalloproteases (MMPs) in SUI fibroblasts after co-culture with DMSCs. Paracrine effect of DMSCs on SUI fibroblast co-cultured with on Transwell® culture inserts. Fold changes in the expression of matrix metalloprotease-2 (MMP-2) and -1 (MMP-1) genes, as well as tissue metalloprotease inhibitor-1 (TIMP-1) gene, in SUI fibroblasts co-cultured with DMSCs (CC) relative to the corresponding SUI fibroblast (SUI) (n=7). Data are presented as the mean ± standard deviation (SD).
Figure 9.
Analysis of matrix metalloproteases (MMPs) in SUI fibroblasts after co-culture with DMSCs. Paracrine effect of DMSCs on SUI fibroblast co-cultured with on Transwell® culture inserts. Fold changes in the expression of matrix metalloprotease-2 (MMP-2) and -1 (MMP-1) genes, as well as tissue metalloprotease inhibitor-1 (TIMP-1) gene, in SUI fibroblasts co-cultured with DMSCs (CC) relative to the corresponding SUI fibroblast (SUI) (n=7). Data are presented as the mean ± standard deviation (SD).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



