Submitted:
09 June 2026
Posted:
10 June 2026
You are already at the latest version
Abstract
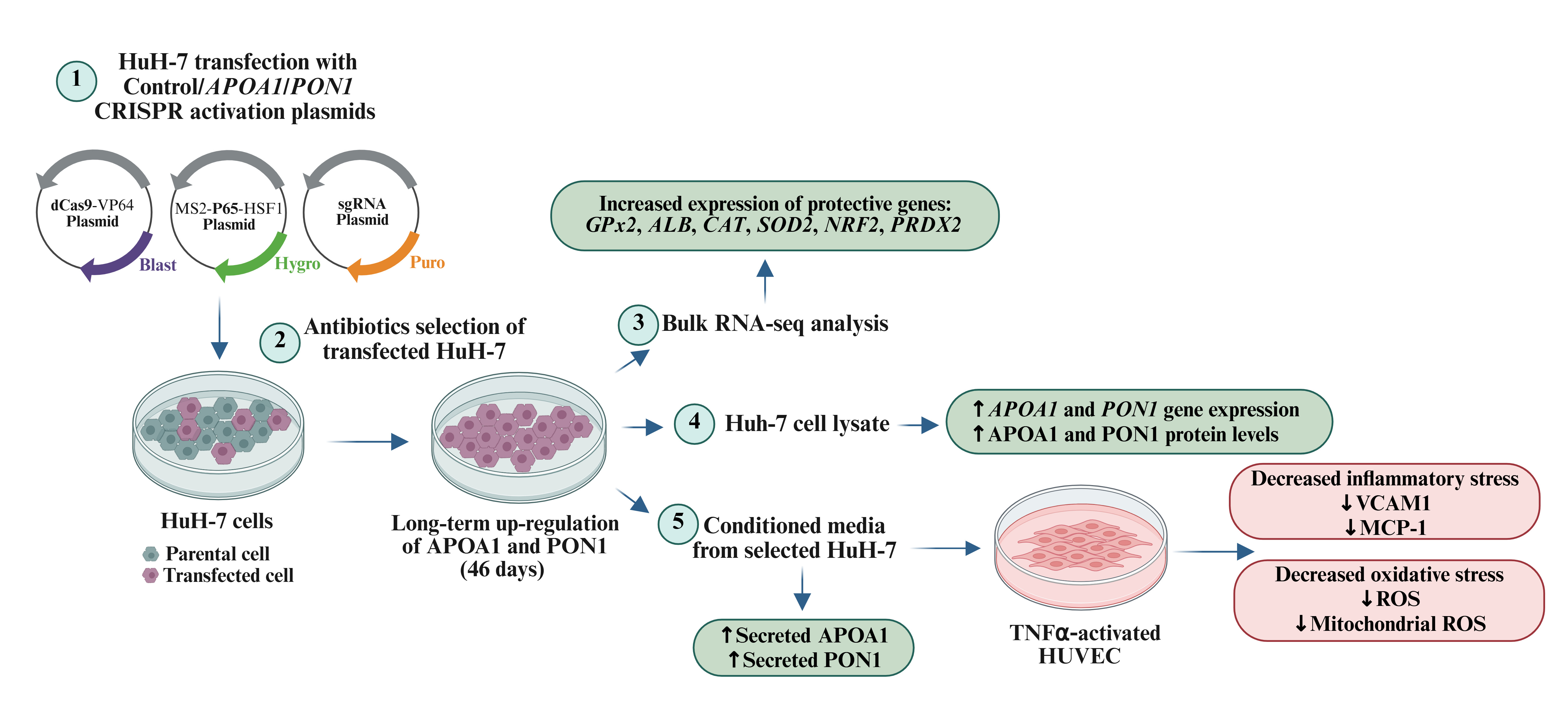
Apolipoprotein A1 (APOA1) and paraoxonase 1 (PON1) are key proteins of high-density lipoprotein (HDL). The aim of the present study was to obtain and characterize an in vitro model for endogenous APOA1 and PON1 long-time up-regulation in hepatocytes that can be further used to decipher the mechanism of their protective action. Cultured human hepatocytes (HuH-7 cell line) were transfected with CRISPR/dCas9 activation plasmids targeting APOA1/PON1 genes. Following selection with specific antibiotics, RNA sequencing was used for the transcriptomic characterization of the transfected hepatocytes. The functionality of the secreted APOAI/PON1 was evaluated as the capacity of the conditioned medium (CM) from transfected HuH-7 to modulate the oxidative and inflammatory stress in TNFα-activated primary human umbilical endothelial cells (HUVEC). The results showed that: (1) a robust, long-time up-regulation (46 days) of endogenous APOA1/PON1 was obtained after CRISPR/dCas9 transfection and antibiotics selection; (2) APOA1/PON1 up-regulation led to a modified transcriptomic profile and increased the expression of several antioxidant genes in transfected hepatocytes as demonstrated by RNAseq analysis; (3) secreted APOA1/PON1 were functional as demonstrated by the CM ability to reduce the levels of reactive oxygen species and inflammatory markers (VCAM-1, MCP-1) in TNF-α-activated HUVEC. In conclusion, we achieved an experimental model of successful long-term up-regulation of endogenous APOA1 and PON1 in human hepatocytes. The targeted proteins are secreted in a functional form and can be used for deciphering their complex mechanism of protective action in various pathological conditions.
Keywords:
1. Introduction
2. Results
2.1. Time-Course of APOA1 and PON1 Expression in Selected and Non-Selected Transfected HuH-7 Cells
2.2. Transcriptome Profiling of HuH-7 Cells with Long-Term Upregulated APOA1 or PON1
2.3. Enrichment Analysis of Differentially Expressed Genes Associated with Oxidative Signaling Pathways
2.4. APOA1 and PON1 Enriched Conditioned Media from Transfected HuH-7 Cells Alleviate TNFα-Induced Oxidative and Inflammatory Stress in Human Primary Endothelial Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cells and Culture Conditions
4.3. Transfection of HuH-7 Cells to Activate the Transcription of APOA1 and/or PON1 Genes Using CRISPR/dCas9 System
4.4. Selection of Transfected HuH-7 to Obtain Long-Term Upregulation of APOA1 and/or PON1
4.5. Bulk RNA-Seq Analysis of APOA1 and PON1 Transfected Hepatocytes
4.6. Preparation of Conditioned Media from Selected HuH-7 with Transcriptional Activated APOA1 and PON1
4.7. TNFα Activation of HUVECs and Incubation with CM from HuH-7 Cells
4.8. Determination of Total Intracellular ROS
4.9. Measurement of Mitochondrial ROS Levels
4.10. Quantitative Real-Time PCR Analysis of Gene Expression
4.11. Quantitative Western Blot Analysis of Protein Expression
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APOA1 | Apolipoprotein A1 |
| PON1 | Paraoxonase 1 |
| CP | Control plasmid |
| CM | Conditioned media |
| TNFα | Tumor necrosis factor α |
| HUVEC | Human umbilical vascular endothelial cells |
| VCAM1 | Vascular cell adhesion molecule 1 |
| MCP1 | Monocyte chemoattractant protein 1 |
| CVD | Cardiovascular disease |
| HDL | High density lipoproteins |
| dCas9 | Dead Cas9 |
| sgRNA | Single guide RNA |
| ROS | Reactive oxygen species |
References
- Capone, F.; Vacca, A.; Bidault, G.; Sarver, D.; Kaminska, D.; Strocchi, S.; Vidal-Puig, A.; Greco, C.M.; Lusis, A.J.; Schiattarella, G.G. Decoding the Liver-Heart Axis in Cardiometabolic Diseases. Circ. Res. 2025, 136, 1335–1362. [Google Scholar] [CrossRef] [PubMed]
- Stadler, J.T.; Marsche, G. Dietary Strategies to Improve Cardiovascular Health: Focus on Increasing High-Density Lipoprotein Functionality. Front Nutr. 2021, 8, 761170. [Google Scholar] [CrossRef] [PubMed]
- Lewoń-Mrozek, D.; Kurzynoga, J.; Jędrzejewski, P.; Kędzierska, K.; Partyka, A.; Kuriata-Kordek, M.; Ściskalska, M. Molecular Structure of Paraoxonase-1 and Its Modifications in Relation to Enzyme Activity and Biological Functions-A Comprehensive Review. Int. J. Mol. Sci. 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Diaz, L.; Bielczyk-Maczynska, E. High-density lipoprotein cholesterol: how studying the 'good cholesterol' could improve cardiovascular health. Open Biol. 2025, 15, 240372. [Google Scholar] [CrossRef]
- Kanafi, M.M.; Tavallaei, M. Overview of advances in CRISPR/deadCas9 technology and its applications in human diseases. Gene 2022, 830, 146518. [Google Scholar] [CrossRef]
- Cai, R.; Lv, R.; Shi, X.; Yang, G.; Jin, J. CRISPR/dCas9 Tools: Epigenetic Mechanism and Application in Gene Transcriptional Regulation. Int. J. Mol. Sci. 2023, 24. [Google Scholar] [CrossRef]
- Siew, W.S.; Tang, Y.Q.; Kong, C.K.; Goh, B.H.; Zacchigna, S.; Dua, K.; Chellappan, D.K.; Duangjai, A.; Saokaew, S.; Phisalprapa, P.; et al. Harnessing the Potential of CRISPR/Cas in Atherosclerosis: Disease Modeling and Therapeutic Applications. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Luo, N.; Zhong, W.; Li, J.; Lu, J.; Dong, R. CRISPR/dCas9 for hepatic fibrosis therapy: implications and challenges. Mol. Biol. Rep. 2022, 49, 11403–11408. [Google Scholar] [CrossRef]
- Rubin, E.M.; Ishida, B.Y.; Clift, S.M.; Krauss, R.M. Expression of human apolipoprotein A-I in transgenic mice results in reduced plasma levels of murine apolipoprotein A-I and the appearance of two new high density lipoprotein size subclasses. Proc. Natl. Acad. Sci. U S A 1991, 88, 434–438. [Google Scholar] [CrossRef]
- Lebherz, C.; Sanmiguel, J.; Wilson, J.M.; Rader, D.J. Gene transfer of wild-type apoA-I and apoA-I Milano reduce atherosclerosis to a similar extent. Cardiovasc Diabetol. 2007, 6, 15. [Google Scholar] [CrossRef]
- Shakweer, W.M.E.; Krivoruchko, A.Y.; Dessouki, S.M.; Khattab, A.A. A review of transgenic animal techniques and their applications. J. Genet Eng. Biotechnol. 2023, 21, 55. [Google Scholar] [CrossRef]
- Karlson, C.K.S.; Mohd-Noor, S.N.; Nolte, N.; Tan, B.C. CRISPR/dCas9-Based Systems: Mechanisms and Applications in Plant Sciences. Plants 2021, 10. [Google Scholar] [CrossRef]
- Dorr, C.R.; Remmel, R.P.; Muthusamy, A.; Fisher, J.; Moriarity, B.S.; Yasuda, K.; Wu, B.; Guan, W.; Schuetz, E.G.; Oetting, W.S.; et al. CRISPR/Cas9 Genetic Modification of CYP3A5 *3 in HuH-7 Human Hepatocyte Cell Line Leads to Cell Lines with Increased Midazolam and Tacrolimus Metabolism. Drug Metab. Dispos. 2017, 45, 957–965. [Google Scholar] [CrossRef]
- Li, H.; Song, C.; Li, Y.; Zhang, T.; Yang, X.; Wang, H. Genome-wide CRISPR screen reveals host factors for gama- and delta-coronavirus infection in Huh7 cells. Int. J. Biol. Macromol. 2025, 304, 140728. [Google Scholar] [CrossRef]
- Lee, H.K.; Lim, H.M.; Park, S.H.; Nam, M.J. Knockout of Hepatocyte Growth Factor by CRISPR/Cas9 System Induces Apoptosis in Hepatocellular Carcinoma Cells. J. Pers. Med. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Soubeyrand, S.; Nikpay, M.; Turner, A.; Dang, A.T.; Herfkens, M.; Lau, P.; McPherson, R. Regulation of MFGE8 by the intergenic coronary artery disease locus on 15q26.1. Atherosclerosis 2019, 284, 11–17. [Google Scholar] [CrossRef]
- Toma, L.; Barbălată, T.; Sanda, G.M.; Niculescu, L.S.; Sima, A.V.; Stancu, C.S. CRISPR/dCas9 Transcriptional Activation of Endogenous Apolipoprotein AI and Paraoxonase 1 in Enterocytes Alleviates Endothelial Cell Dysfunction. Biomolecules 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Bai, X. Apolipoprotein A-I mimetic peptides (ApoAI MP) improve oxidative stress and inflammatory responses in Parkinson's disease mice. Front Pharmacol. 2022, 13, 966232. [Google Scholar] [CrossRef]
- Jangale, N.M.; Devarshi, P.P.; Dubal, A.A.; Ghule, A.E.; Koppikar, S.J.; Bodhankar, S.L.; Chougale, A.D.; Kulkarni, M.J.; Harsulkar, A.M. Dietary flaxseed oil and fish oil modulates expression of antioxidant and inflammatory genes with alleviation of protein glycation status and inflammation in liver of streptozotocin-nicotinamide induced diabetic rats. Food Chem. 2013, 141, 187–195. [Google Scholar] [CrossRef]
- Moustafa, E.M.; Thabet, N.M. Beta-sitosterol upregulated paraoxonase-1 via peroxisome proliferator-activated receptor-γ in irradiated rats. Can. J. Physiol. Pharmacol. 2017, 95, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, M.; Hoekstra, M.; Hildebrand, R.B.; Yaong, Y.; Stengel, D.; Kruijt, J.K.; Sattler, W.; Tietge, U.J.; Ninio, E.; Van Berkel, T.J.; et al. Increased oxidative stress in scavenger receptor BI knockout mice with dysfunctional HDL. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Zhao, M.; Wu, C.; Zhang, W.; Niu, C.; Yu, B.; Jin, J.; Ji, L.; Willard, B.; Mathew, A.V.; et al. Apolipoprotein A-1 mimetic peptide 4F promotes endothelial repairing and compromises reendothelialization impaired by oxidized HDL through SR-B1. Redox Biol. 2018, 15, 228–242. [Google Scholar] [CrossRef]
- Rosenbaum, M.A.; Chaudhuri, P.; Abelson, B.; Cross, B.N.; Graham, L.M. Apolipoprotein A-I mimetic peptide reverses impaired arterial healing after injury by reducing oxidative stress. Atherosclerosis 2015, 241, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Mackness, B.; Hine, D.; Liu, Y.; Mastorikou, M.; Mackness, M. Paraoxonase-1 inhibits oxidised LDL-induced MCP-1 production by endothelial cells. Biochem Biophys. Res. Commun. 2004, 318, 680–683. [Google Scholar] [CrossRef]
- Yuhanna, I.S.; Zhu, Y.; Cox, B.E.; Hahner, L.D.; Osborne-Lawrence, S.; Lu, P.; Marcel, Y.L.; Anderson, R.G.; Mendelsohn, M.E.; Hobbs, H.H.; et al. High-density lipoprotein binding to scavenger receptor-BI activates endothelial nitric oxide synthase. Nat. Med. 2001, 7, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Mineo, C.; Shaul, P.W. HDL stimulation of endothelial nitric oxide synthase: a novel mechanism of HDL action. Trends Cardiovasc Med. 2003, 13, 226–231. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: a web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–w221. [Google Scholar] [CrossRef]
- Thomas, P.D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L.P.; Mi, H. PANTHER: Making genome-scale phylogenetics accessible to all. Protein Sci. 2022, 31, 8–22. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–d646. [Google Scholar] [CrossRef]
- Toma, L.; Sanda, G.M.; Deleanu, M.; Stancu, C.S.; Sima, A.V. Glycated LDL increase VCAM-1 expression and secretion in endothelial cells and promote monocyte adhesion through mechanisms involving endoplasmic reticulum stress. Mol. Cell Biochem 2016, 417, 169–179. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.



