Submitted:
08 June 2026
Posted:
10 June 2026
You are already at the latest version
Abstract
Natural products have long contributed to oncology by providing structurally diverse metabolites and pharmacologically relevant scaffolds. Despite advances in precision oncology, immunotherapy, and targeted therapies, cancer remains a leading cause of morbidity and mortality worldwide. Plant-derived metabolites continue to attract attention due to their ability to modulate key processes such as apoptosis, metabolic adaptation, epigenetic regulation, oxidative stress, inflammation, and tumor microenvironment signaling. However, current evidence is heterogeneous and often based on simplified in vitro models or insufficiently characterized extracts. This review critically distinguishes pharmacologically validated findings from preliminary claims with limited translational relevance. A narrative synthesis of global advances was conducted, focusing on mechanistic pathways, major phytochemical classes, and emerging technologies shaping compound discovery and preclinical validation. The potential contribution of Chilean biodiversity was also examined. Although several Chilean plant species demonstrate cytotoxic, anti-inflammatory, and antioxidant activities relevant to oncology, most findings remain preclinical, limited by inadequate taxonomic identification, lack of phytochemical standardization, scarce in vivo validation, and insufficient pharmacological characterization. The future impact of natural products in oncology will depend on stricter standards for chemical characterization, biological validation, and translational relevance. Chilean biodiversity represents a promising yet largely prospective source for cancer-related drug discovery.
Keywords:
natural products
; phytochemicals
; cancer therapy
; Chilean endemic plants
; immunomodulation
1. Introduction
Cancer remains a major global health challenge, with nearly 20 million new cases diagnosed each year and a persistently high mortality rate despite significant progress in prevention, diagnosis, and treatment [1]. Over the past decade, precision oncology, molecularly targeted therapies, and immunotherapeutic strategies have transformed the management of several cancers. However, tumor heterogeneity, adaptive resistance, immune escape, and treatment-related toxicity continue to limit durable benefits in many settings [2].
Within this context, natural products remain highly relevant to oncology. Historically, they have yielded some of the most influential anticancer agents in clinical use, including vincristine, vinblastine, paclitaxel, and camptothecin-derived drugs [3]. Their ongoing importance is based on structural diversity and the ability to interact with multiple biologic targets. Plant-derived metabolites are still studied as modulators of apoptosis, cell-cycle regulation, oxidative stress, inflammatory signaling, angiogenesis, epigenetic remodeling, metabolic adaptation, and tumor-microenvironment interactions [4,5].
Nevertheless, growing enthusiasm for natural products in cancer research has not always been matched by equivalent pharmacological rigor. Much of the current literature still relies on early-stage in vitro observations involving poorly characterized extracts, supraphysiological concentrations, limited dose-response analysis, or mechanistic claims based on a narrow panel of molecular markers [6,7]. A more critical assessment requires distinguishing between clinically validated compounds, robust preclinical leads, and preliminary bioactivity reports with uncertain pharmacology.
At the same time, natural-product research is undergoing significant technological change. High-resolution metabolomics, molecular networking, multi-omics integration, and computational target prediction are improving the identification and characterization of bioactive metabolites [8]. Advanced experimental platforms such as three-dimensional spheroids, organoids, co-culture systems, and organ-on-a-chip models address longstanding limitations of traditional two-dimensional cell culture [9,10]. This evolving landscape is especially important for countries with unique biological resources such as Chile, whose geographic isolation, diverse climate, and high plant endemism offer a distinctive source of botanical diversity with potential biomedical relevance [11].
The purpose of this review is not just to list reported bioactivities but to critically evaluate the current state of natural product-based cancer research both globally and locally. We discuss the main mechanisms through which plant-derived metabolites may influence cancer biology, followed by key chemical classes shaping the field, new discovery technologies, opportunities and challenges related to Chilean biodiversity, and the main conceptual, methodological, and translational gaps that slow progress in this area.
2. Global Advances in Natural Product-Based Cancer Therapy
Natural products continue to hold a unique position in cancer research because of their structural diversity, evolutionary refinement, and broad biological relevance. Their historical contribution to oncology is well established: agents such as paclitaxel, vincristine, and camptothecin-derived drugs illustrate how naturally occurring metabolites can yield clinically effective therapies [3,5]. However, the modern field extends far beyond these landmark examples; plant-derived metabolites are now studied as modulators of proteostatic stress, senescence, metabolism, chromatin state, inflammatory signaling, and immune function within the tumor microenvironment [4,5].
As an overview, plant-derived natural compounds exert anticancer effects through multiple and interconnected biological mechanisms. These compounds influence key cellular processes involved in tumor initiation, progression, and response to therapy, including the regulation of cell survival, stress responses, metabolism, and immune function.
2.1. Mechanistic Pathways of Natural Compounds in Cancer Biology
A main reason for the ongoing interest in plant-derived metabolites is their ability to influence multiple biological processes simultaneously. Unlike highly selective synthetic inhibitors, many phytochemicals act on interconnected signaling networks, affecting cell survival, inflammation, metabolic adjustment, and stress-response pathways at once. In diseases characterized by redundancy and adaptive plasticity, this multimodal behavior can offer a significant biological advantage. Figure 1 illustrates the key mechanistic pathways by which plant-derived natural compounds modulate cancer biology, including their effects on cell proliferation, apoptosis, inflammation, angiogenesis, metastasis, and immune regulation. However, it also raises the risk of overinterpretation, especially when mechanistic claims are based on a limited set of molecular or phenotype data.
2.1.1. ER Stress and Apoptosis
One of the most common mechanistic themes in natural-product oncology is the induction of endoplasmic reticulum (ER) stress and its progression toward apoptosis. This is biologically plausible because many malignant cells operate near the limits of proteostatic tolerance due to rapid proliferation, increased protein synthesis, hypoxia, and metabolic stress. Under these conditions, disruption of protein folding or ER homeostasis can shift the unfolded protein response (UPR) from an adaptive process to a terminal signaling pathway. The three main UPR branches—PERK-eIF2alpha-CHOP, IRE1-XBP1, and ATF6—are key to this transition and have thus become important mechanistic targets in natural-product research [12,13].
For example, triptolide has been reported to cause sustained ER stress in pancreatic cancer models, with activation of PERK-eIF2alpha and IRE1-XBP1 signaling leading to apoptosis rather than recovery [14]. Flavonoids, terpenoids, and alkaloids have all been linked to ER stress-related apoptosis in cancer cells, although the extent of mechanistic support varies considerably among different compounds and models [12]. However, ER stress in cancer should not be viewed as a universally favorable therapeutic mechanism; its importance depends heavily on the model context, the duration and severity of stress, and whether the observed response truly indicates a targetable vulnerability rather than nonspecific injury [15].
2.1.2. Senescence Induction
Cellular senescence has become a key mechanistic theme because it offers a potential alternative to direct cytotoxicity. Instead of rapidly killing tumor cells, some metabolites may induce a stable state of growth arrest, along with characteristic changes such as altered morphology, senescence-associated beta-galactosidase activity, DNA damage response signaling, and modified telomerase regulation. Recent research has consistently linked several natural metabolites, including baicalein, berberine, and pterostilbene, to this phenotype [16].
In colorectal cancer cells, baicalin was reported to induce a pro-senescent phenotype characterized by reduced hTERT expression, diminished telomerase activity, and features consistent with durable growth arrest, with ERK and p38MAPK signaling involved [17]. In non-small cell lung cancer, pterostilbene was shown to promote senescence through p53-dependent reduction of hTERT and persistent DNA damage response signaling [18]. Berberine has also been associated with senescence-like arrest in glioblastoma models by suppressing EGFR and attenuating the EGFR-MEK-ERK pathway [19]. However, senescence is not inherently beneficial in all circumstances; its significance depends heavily on tumor type, stromal context, immune surveillance, and the composition of the senescence-associated secretory phenotype (SASP) [20].
2.1.3. Metabolic Reprogramming
Metabolic reprogramming has become a key focus of natural product research, especially regarding glycolytic dependence. Polyphenolic metabolites such as resveratrol, quercetin, and epigallocatechin gallate (EGCG) have been repeatedly linked to glucose uptake, lactate production, glycolytic enzymes, and signaling pathways involved in metabolic adaptation, including AMPK, Akt/mTOR, and HIF-1alpha-related pathways [4,21].
In ovarian cancer cells, resveratrol inhibited glycolysis via AMPK/mTOR-related regulation of metabolic flux while also reducing proliferation [22]. In hepatocellular carcinoma models, quercetin decreased hexokinase 2-dependent glycolysis and dampened Akt-mTOR signaling [23]. In preclinical breast cancer models, EGCG reduced tumor growth in association with decreases in glucose uptake and lactate levels [24]. However, this area is often overstated in literature; reduced glucose uptake or changes in a single enzyme do not alone prove comprehensive metabolic rewiring. Many studies still lack metabolic flux analyses or broader metabolomic datasets to establish whether tumor cells have genuinely shifted their metabolic state.
2.1.4. Epigenetic Regulation
Epigenetic regulation has become an increasingly important theme because it connects diet-derived and botanical metabolites to reversible mechanisms that influence tumor plasticity and treatment response. The compounds most consistently discussed include curcumin, sulforaphane (SFN), and genistein, all of which have been linked to modulation of DNA methylation, histone deacetylation activity, and non-coding RNA programs [21].
Curcumin has repeatedly been associated with suppression of HDAC activity and increased histone acetylation in cancer cells [25,26]. SFN provides a particularly clear example of promoter-level epigenetic regulation; in human colon cancer cells, it inhibited DNMT1 expression and facilitated demethylation of the Nrf2 (NFE2L2) promoter, enabling the restoration of antioxidant transcriptional programs relevant for chemoprevention [27]. Genistein induced demethylation at CpG sites near the miR-200c/miR-141 loci in prostate cancer cells, supporting the reactivation of EMT-restraining programs linked to epithelial identity [28]. Overall, epigenetic effects induced by natural metabolites should be viewed as biologically informative and potentially relevant for prevention or combination strategies, but not yet as sufficient evidence for broad epigenetic normalization in cancer.
2.1.5. Immune Modulation
Immune modulation has become a key theme in natural-product oncology, expanding the discussion beyond direct tumor killing to include the tumor immune microenvironment (TIME). Natural products are increasingly studied as immunomodulatory adjuvants capable of influencing antigen presentation, effector lymphocyte activity, myeloid-cell programming, checkpoint pathways, and immunogenic cell death [29]. However, these effects depend heavily on dose, timing, tumor type, and the immune environment under investigation.
Curcumin has been reported to shift Foxp3+ regulatory T cells toward a Th1-like phenotype in lung cancer patients, along with increased IFN-gamma [30]. Resveratrol suppressed tumor growth in a murine renal tumor model in a CD8+ T-cell-dependent manner [31]. Curcumin also inhibited the buildup of myeloid-derived suppressor cells (MDSCs) and promoted their differentiation [32]. Withaferin A has been reported to induce immunogenic cell death (ICD) in non-small cell lung cancer cells while increasing PD-L1 expression, further supporting the concept of combining natural-product regimens with immune checkpoint inhibitors [33]. Overall, the current evidence supports certain natural metabolites as promising leads for combination immunotherapy research, but their clinical relevance still requires stronger validation in immunocompetent tumor models.
2.2. Key Phytochemical Classes: Structural and Functional Perspectives
Beyond a mechanism-based organization, it is also useful to consider natural products according to their principal phytochemical classes, since structural families often display recurrent pharmacological tendencies as well as characteristic translation limitations. These classes are not homogeneous in their clinical relevance; some include metabolite scaffolds that have already shaped oncology practice, whereas others are composed mainly of compounds that remain mechanistically informative but largely preclinical.
2.2.1. Polyphenols and Flavonoids
Polyphenols and flavonoids are among the most thoroughly researched plant metabolites in cancer studies. Resveratrol, quercetin, curcumin, piceatannol, EGCG, and genistein have all been linked to the regulation of inflammatory signaling, apoptosis, epithelial-mesenchymal transition, oxidative stress, and transcriptional programs associated with proliferation and metastasis [34,35,36].
Their appeal partly stems from their pleiotropy; resveratrol has been linked to metabolic regulation, inflammatory signaling, and checkpoint-related effects, while quercetin is more specifically associated with glycolytic inhibition and redox modulation [4,23]. Curcumin is discussed in relation to inflammatory, epigenetic, and immune pathways, while EGCG has been connected to both angiogenic and metabolic signaling [4,37]. However, many polyphenols exhibit low oral bioavailability, extensive metabolism, and in vitro activity at concentrations that may not be achievable in the body. Their redox behavior and wide-ranging protein interactions raise concerns about specificity and assay interference. Polyphenols should therefore be viewed as mechanistically rich metabolites whose most practical value may lie in chemopreventive applications, rational combinations, or lead optimization.
2.2.2. Terpenoids
Terpenoids hold a significant role in oncology, especially because this group includes one of the clearest examples of successful natural-product application. Paclitaxel (Taxol), derived from Taxus brevifolia, remains a key chemotherapeutic agent due to its ability to stabilize microtubules, interfere with dynamic remodeling, and trigger mitotic arrest followed by tumor cell death [3,38,39]. This example highlights the clinical importance of terpenoid scaffolds.
Beyond taxanes, research on terpenoids is still largely exploratory. Triterpenoids such as ursolic acid, betulinic acid, and oleanolic acid have been linked to mitochondrial depolarization, cytochrome c release, regulation of Bcl-2 family proteins, inhibition of NF-kB and STAT3, and growth suppression in various tumor models [40,41,42,43]. While these compounds demonstrate meaningful biological activity, their development remains limited by poor solubility, unfavorable pharmacokin[40–43etics, and the ongoing gap between mechanistic plausibility and consistent therapeutic effectiveness. Terpenoids are therefore best discussed in two categories: taxane-based agents as validated anticancer drugs, and non-taxane terpenoids as promising but predominantly preclinical candidates.
2.2.3. Alkaloids and Lactones
Alkaloids and sesquiterpene lactones represent another example of uneven development within a structural class. Camptothecin and its clinically used derivatives remain among the strongest examples of successful natural-product translation, functioning through topoisomerase I (TOP1) poisoning and stabilization of TOP1-DNA cleavage complexes, leading to replication-associated DNA damage and cell-cycle arrest [44,45]. Their clinical relevance is well established and should be clearly distinguished from the more exploratory compounds often grouped within the same category.
Berberine illustrates the more experimental side of alkaloid research. It has been linked to AMPK activation, suppression of metastatic behavior, interference with epithelial-mesenchymal transition, and inhibition of proliferative and migratory programs in gastrointestinal and other cancer models [46,47,48]. Sesquiterpene lactones such as parthenolide, artemisinin-related metabolites, and leptocarpin have also garnered attention because of their electrophilic nature and their ability to influence redox-sensitive and inflammatory signaling pathways, especially NF-kB [49,50,51]. While this electrophilicity may be mechanistically beneficial, it also raises concerns abo[46–48ut promiscuity and non-specific reactivity.
2.3. Technological Advances in Natural-Product Discovery
The current progress in natural-product oncology is closely linked to a new generation of technologies that have transformed the discovery, characterization, and translational development of bioactive metabolites. High-resolution metabolomics, molecular networking, and integrated bioinformatics platforms have significantly improved metabolite annotation, dereplication, and the identification of biologically relevant compounds within highly complex botanical extracts [8,52]. Simultaneously, artificial intelligence (AI) and machine learning approaches are expanding opportunities for target prediction, drug–target interaction modeling, structure–activity relationship analysis, and candidate prioritization, accelerating the transition from natural-product screening to drug development [53]. In parallel, advances in experimental cancer models—including three-dimensional spheroids, patient-derived organoids, co-culture systems, and organ-on-a-chip technologies—provide a more physiologically relevant context for evaluating the anticancer effects of natural compounds. These platforms more accurately reproduce tumor architecture, cellular heterogeneity, microenvironmental gradients, immune interactions, and drug penetration dynamics than conventional two-dimensional cultures, thereby improving the predictive value of preclinical studies and reducing the translational gap between laboratory findings and clinical outcomes [9,10].
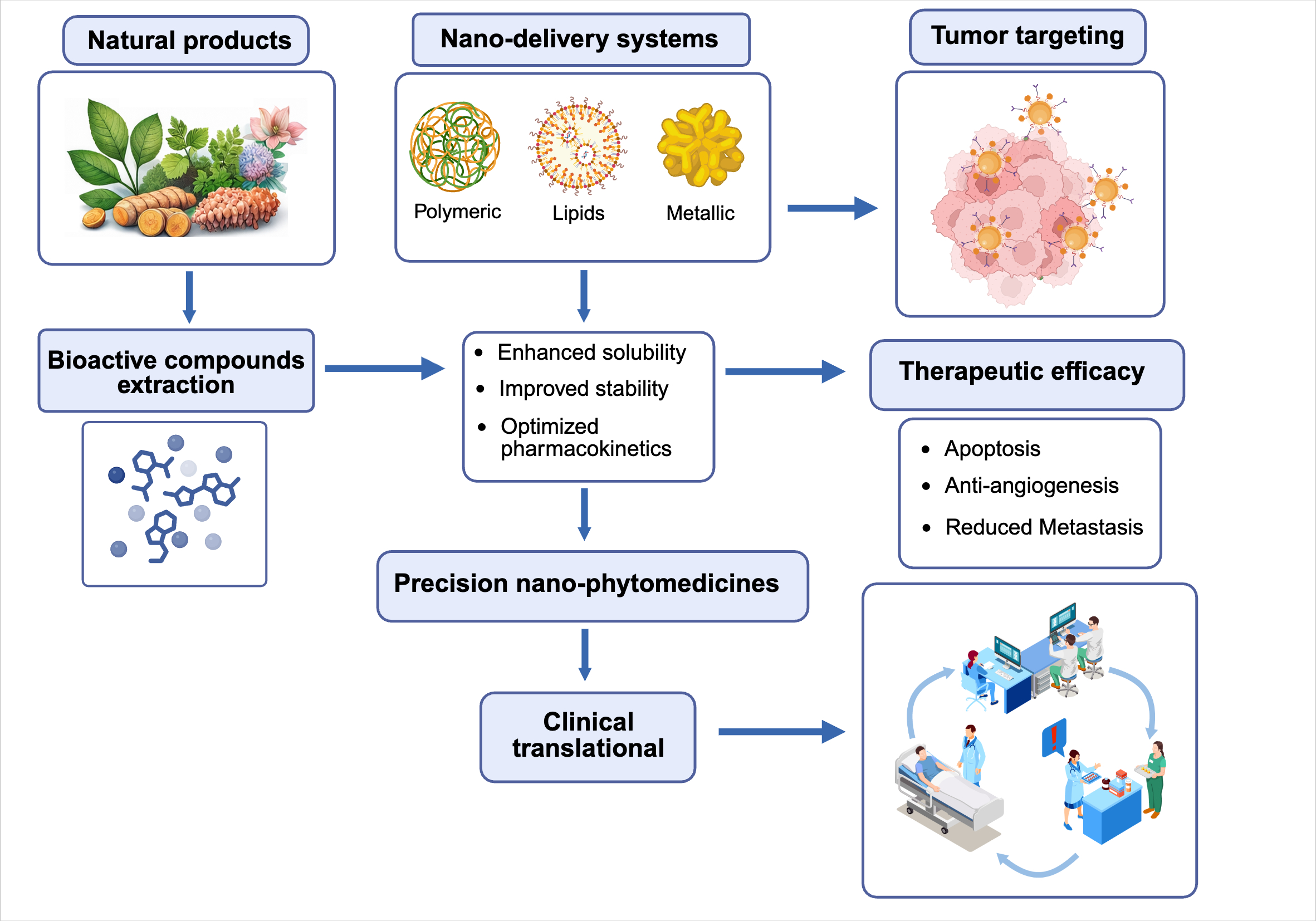
Despite these advances, one of the major barriers limiting the clinical application of natural-product-derived therapeutics remains their unfavorable pharmacological properties. Many promising metabolites exhibit poor aqueous solubility, limited chemical stability, rapid systemic clearance, extensive metabolism, and low bioavailability, resulting in insufficient concentrations at tumor sites. Nanotechnology-based delivery systems have emerged as a powerful strategy to overcome these limitations by encapsulating natural compounds within polymeric nanoparticles, lipid-based carriers, liposomes, micelles, solid lipid nanoparticles, nanostructured lipid carriers, or metallic nanomaterials. These platforms not only improve solubility and stability but also prolong circulation time, protect bioactive molecules from degradation, facilitate controlled drug release, and enhance preferential accumulation within tumors through passive and active targeting mechanisms.
As summarized in Table 1 and Figure 2, an increasing number of nanoformulated natural metabolites have demonstrated superior therapeutic performance compared with their free counterparts across multiple cancer types. Curcumin-loaded polymeric micelles have shown enhanced cellular uptake, increased induction of apoptosis, and greater suppression of colorectal tumor growth in vivo [54]. TPGS-resveratrol solid lipid nanoparticles have exhibited improved intracellular delivery, stronger pro-apoptotic effects, and reduced migratory capacity in paclitaxel-resistant breast cancer cells [55]. Similarly, quercetin-loaded solid lipid nanoparticles have enhanced antiproliferative activity and apoptosis induction in triple-negative breast cancer models [56]. Beyond improving drug delivery, nanoformulations can modulate the tumor microenvironment, inhibit angiogenesis, reduce metastatic dissemination, and increase sensitivity to chemotherapy and immunotherapy. Importantly, the clinical success of albumin-bound paclitaxel (nab-paclitaxel) demonstrates that nanotechnology can facilitate the translation of bioactive compounds into clinically meaningful therapies, leading to improved pharmacokinetics, treatment response, and progression-free survival in patients with metastatic breast cancer [57].
Collectively, these technological and pharmaceutical advances are reshaping the landscape of natural-product-based oncology. The integration of metabolomics, AI-driven discovery platforms, advanced tumor models, and nano-delivery technologies is creating a more robust translational pipeline capable of transforming promising natural metabolites into precision therapeutics. While rigorous biological validation, pharmacological characterization, and well-designed clinical studies remain essential, nano-phytomedicine is increasingly recognized as a critical enabling strategy for unlocking the full therapeutic potential of natural products in cancer prevention and treatment.
3. Chilean Biodiversity as an Emerging Resource for Anticancer Compounds
Chile provides a biologically unique environment for natural product research because of its clear latitudinal gradient, strong biogeographic isolation, and high level of plant endemism [11,58]. These ecological factors have promoted the diversification of stress-adaptive compounds—phenolics, terpenoids, alkaloids, and sesquiterpene lactones—many of which are now being studied for their antioxidant, anti-inflammatory, cytotoxic, or chemopreventive effects [11,59]. Ethnopharmacological traditions from the Mapuche, Aymara, and Rapa Nui communities further highlight the importance of this biodiversity [60,61,62].
3.1. Species with Documented Anticancer Potential
Among the Chilean taxa currently discussed in oncology-oriented literature, Leptocarpha rivularis DC. (Asteraceae) is one of the most promising candidates because its evidence goes beyond general antioxidant activity and includes direct observations in tumor models. Its major sesquiterpene lactone, leptocarpine (LTC), has been reported to exert antiproliferative and pro-apoptotic effects in gastric cancer cell lines [51]. Flower extracts obtained with dichloromethane (DCM), ethyl acetate (EtOAc), hexane (Hex), and ethanol (EtOH) were tested in AGS and MKN-45 gastric cancer cells; under the most active conditions, DCM-, EtOAc-, and Hex-derived extracts reduced proliferation, altered cell-cycle distribution, promoted mitochondrial membrane depolarization, increased DEVDase activity, impaired clonogenic capacity, induced senescence, and decreased migration and invasion [51].
Although the available evidence remains predominantly preclinical, several native and endemic plant species have demonstrated promising anticancer activity in cellular and experimental tumor models. Table 2 summarizes the current evidence regarding Chilean plant species investigated for cancer-related applications, including the tested biological material, tumor models evaluated, observed biological effects, and the principal limitations that must be addressed to advance their translational potential.
In micropropagated material, leptocarpine and clonal plant extracts were tested on HeLa cervical cancer cells; after 48 hours, leptocarpine at 1-1.2 ppm more effectively reduced metabolic activity in HeLa cells compared to non-tumor CoN cells, and LTC significantly lowered IL-6 and MMP-2 expression [63]. More recently, ovatifolin isolated from the aerial parts of L. rivularis was tested on A-2058 and A-375 melanoma cells, revealing live-cell IC50 values of 27.6 and 18.4 microg/mL, respectively [64]. These findings position L. rivularis among the most promising Chilean sources of cancer-related sesquiterpenoids, while also highlighting that current evidence remains mainly preclinical and model-dependent.
Ugni molinae Turcz. (Myrtaceae) has been discussed in cancer-related contexts, although current evidence is more convincing for its antioxidant and anti-inflammatory activities than for direct anticancer effects. Extracts prepared after various drying processes were tested at 0.25 mg/mL for 48 hours in NCI-H1975 non-small cell lung cancer cells, with the fresh extract decreasing viability by approximately 13% [65]. Aqueous leaf extracts also reduced viability in AGS gastric adenocarcinoma cells at concentrations from 62.5 microg/mL [66]. The available evidence more consistently supports U. molinae as a source of antioxidant and anti-inflammatory metabolites with possible chemopreventive relevance [67,68].
Berberis microphylla G.Forst. (Berberidaceae) can be more directly linked to cancer-related evidence. Crude and anthocyanin-rich calafate fruit extracts were tested on AGS and G415 gallbladder carcinoma cells; both extracts reduced cell viability and migration, with the anthocyanin-rich extract showing up to 70% inhibition mainly associated with delphinidin derivatives [69]. Azorella compacta Phil. (Apiaceae) demonstrated cancer-related activity at both the extract and metabolite levels; a methanolic extract inhibited growth of HL60, HepG2, SNU-1, MCF-7, HT1080, and A549 cell lines, while isolated diterpenoids reduced viability in MCF-7 cells, with azorellane-type diterpenoids reported as more active than mulinanes [70,71].
Lithraea caustica (Molina) Hook. & Arn. (Anacardiaceae) demonstrates a different type of cancer-related evidence. A standardized litre extract (LexT) was tested in a murine B16 melanoma model; both topical treatment and intratumoral injection slowed tumor growth, and topical treatment caused tumor regression in 15% of treated animals, supporting an antitumor immune response rather than a traditional direct cytotoxic mechanism [72]. This highlights that cancer-related activity in Chilean species may also occur through immune-mediated pathways
3.2. Biotechnology, Standardization, and Sustainability as Enabling Conditions
Biotechnology, chemical standardization, and sustainability should be viewed as essential enabling conditions that influence whether cancer-relevant metabolites can be studied in a reproducible and meaningful translational way. Plant tissue culture and related biotechnological methods are increasingly seen as practical strategies for large-scale production, conservation, and standardization of plant secondary metabolites [73,74].
Among the Chilean examples discussed, L. rivularis offers one of the clearest cases of how biotechnology can enhance a pharmacological research process. A micropropagation system allowed for optimizing introduction, propagation, and rooting stages, with regenerated plants achieving an 83% acclimatization rate under greenhouse conditions [63]. This propagation system was combined with phytochemical and biological analysis, demonstrating that leptocarpine remained the main metabolite and that extracts from clonal plants retained cancer-relevant activity in vitro [63].
In U. molinae, different drying methods modified phenolic content along with antioxidant, anti-inflammatory, and tumor-cell viability effects, indicating that pre-analytical processing can influence the apparent pharmacological behavior of an extract before any mechanistic interpretation is made [65]. Sustainable access enhances reproducibility and minimizes ecological impact, but it does not address deeper issues such as dose feasibility, tumor selectivity, pharmacokinetics, or model relevance. These strategies only provide true value when combined with more robust experimental design and more informative biological systems.
Governance and access-and-benefit-sharing also represent important dimensions. In Latin America, access-and-benefit-sharing challenges under the Convention on Biological Diversity and the Nagoya framework have been identified as major barriers to collaborative ethnopharmacological research [75]. Future Chilean natural-product oncology should integrate traceability, community recognition, and fair benefit-sharing into the research process itself, rather than addressing them only as regulatory requirements.
4. Gaps, Limitations, and Future Research Priorities
The renewed interest in natural products in oncology reflects a broader shift from reductionist, single-target drug discovery toward a more integrated view of pharmacology. Advances in metabolomics and molecular networking have greatly enhanced dereplication, metabolite annotation, and the understanding of complex botanical mixtures [8,52]. However, these advancements have not addressed a key issue: the evidence supporting many claims of anticancer relevance remains highly inconsistent, often based on simplified in vitro systems or mechanistic interpretations that extend beyond the strength of the available data [6,7].
4.1. Preclinical Validation: Methodological Gaps
One of the most ongoing challenges in natural-product pharmacology is the lack of a clear definition of the material being studied. Extract-based studies are still often published without enough information about botanical identity, plant part, extraction solvent, batch consistency, or analytical composition. Current guidance stresses the need for thorough botanical verification, transparent documentation, fit-for-purpose analytical characterization, and batch-to-batch consistency control [6]. Orthogonal analytical methods such as LC-MS/MS and NMR are increasingly seen as critical tools for metabolite fingerprinting and reliable chemical profiling [6,76].
A large part of the literature still depends on traditional two-dimensional monolayer systems exposed to high and sometimes poorly contextualized concentrations. These assays remain helpful for preliminary screening, but they do not mimic tumor architecture, extracellular matrix constraints, microenvironmental gradients, or adaptive metabolic states that are highly relevant to treatment response [9]. More advanced systems, including organ-on-a-chip platforms, provide a better approximation of tissue complexity and drug-response dynamics [10].
Selectivity and exposure feasibility are additional weaknesses. Many studies evaluate tumor cell lines without including non-tumor comparators or clarifying whether the reported active concentrations are pharmacologically realistic. A reduction in tumor cell viability is not inherently meaningful if it occurs at similarly toxic concentrations in non-malignant cells, or at exposure levels unlikely to be achieved in vivo. These gaps are worsened by the fact that non-clinical evaluation is not always aligned with internationally recognized safety-testing frameworks such as OECD Good Laboratory Practice [77].
Many compounds frequently discussed in phytochemical oncology fall into categories associated with pan-assay interference or promiscuous behavior—especially redox-active polyphenols and electrophilic sesquiterpene lactones—whose apparent activity may partly stem from assay interference or non-specific reactivity [78]. Strong in vitro activity should not be equated with translational relevance. The field must shift from solely reporting bioactivity to properly evaluating evidence through a clearer hierarchy supported by appropriate positive and negative controls and non-tumor system comparators.
4.2. Challenges and Opportunities in Chilean Biodiversity Research
The case of Chilean biodiversity demonstrates both the potential and the current immaturity of natural-product oncology. Chile has one of the most unique floras in the region, shaped by distinct ecological gradients, geographic isolation, and high endemism [11]. However, most evidence supporting Chilean native or endemic species is still preliminary and scattered, often limited by non-standardized extraction methods, incomplete chemical annotation, weak reproducibility, and limited validation in physiologically relevant tumor models [8,9,11].
The top priority is ensuring taxonomic and chemical accuracy. Full taxonomic validation—including accepted species names, authorities, and families—is crucial for reproducibility, comparability, and regulatory trustworthiness. Current guidelines highlight that botanical authentication and clearly defined analytical preparations are necessary to support meaningful biological interpretation [6]. Without this foundation, cross-laboratory comparisons remain weak and translational extrapolation becomes fragile.
A second priority is advancing the experimental process through a more systematic workflow: starting with taxonomic and phytochemical analysis, then conducting concentration-aware screening in relevant tumor and non-tumor systems, followed by mechanistic studies, validation in three-dimensional or co-culture platforms, and only then moving into justified in vivo models. A third priority focuses on infrastructure and reproducibility through integration of metabolomics-guided annotation, phytochemical traceability, and biologically informative testing [8]. The most valuable investment is probably interdisciplinary: platforms that connect taxonomy, phytochemistry, analytical chemistry, advanced biological models, and early pharmacology.
Ultimately, the future of Chilean biodiversity research depends on clearer prioritization. Not every native or endemic species should be viewed as a potential anticancer agent. A more practical approach would be to differentiate between taxa with specific metabolite-level potential for lead discovery, those with stronger evidence for prevention or anti-inflammatory applications, and taxa whose current value mainly lies in exploratory phytochemistry. The true potential of Chilean biodiversity is not in the immediate development of new anticancer agents, but in the opportunity to establish a more rigorous, taxonomically precise, chemically defined, and translationally cohesive research framework.
5. Conclusions
Natural products continue to play a vital role in oncology, not only because they have historically led to some of the most influential anticancer agents in clinical practice, but also because they remain a rich source of chemically diverse metabolites capable of uncovering biologically relevant vulnerabilities in tumor cells and the tumor microenvironment [3,5]. Simultaneously, the current landscape is characterized by a persistent tension between biological potential and scientific rigor. While some compounds and scaffold families have clear pharmacological and translational value, a significant portion of the current literature still relies on preliminary findings based on simplified in vitro systems, incompletely characterized extracts, or mechanistic interpretations that are broader than what the available data can convincingly support.
A key conclusion of this review is that natural products should not be considered a single, uniform therapeutic category. Instead, they should be viewed as a diverse research area that includes clinically validated agents, biologically meaningful lead compounds, prevention-focused metabolites, and many exploratory compounds whose importance remains tentative. Improved standards for taxonomic validation, phytochemical analysis, dose-aware pharmacology, mechanistic understanding, and model selection will be necessary to distinguish truly actionable candidates from metabolites that are simply biologically intriguing [6,78].
In the Chilean context, the true value of biodiversity-based research will only become apparent when biological potential is linked to chemical understanding, sustainability, and institutional governance. Chilean biodiversity should be regarded not as an immediately actionable source of anticancer agents, but as a strategically valuable research platform whose future impact on oncology depends on integrating rigorous pharmacology with ecological and institutional stewardship.
Author Contributions
Conceptualization, R.G.-S. and E.S.-M.; methodology, R.G.-S.; writing—original draft preparation, R.G.-S.; writing—review and editing, J.R.-A. and E.S.-M.; supervision, E.S.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors thank the Fundación Arturo López Pérez (FALP) for institutional support.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AMPK | AMP-activated protein kinase |
| DNMT | DNA methyltransferase |
| EGCG | Epigallocatechin gallate |
| EMT | Epithelial-mesenchymal transition |
| ER | Endoplasmic reticulum |
| HDAC | Histone deacetylase |
| ICD | Immunogenic cell death |
| LTC | Leptocarpine |
| MDSC | Myeloid-derived suppressor cell |
| SASP | Senescence-associated secretory phenotype |
| SFN | Sulforaphane |
| TAM | Tumor-associated macrophage |
| TIME | Tumor immune microenvironment |
| TOP1 | Topoisomerase I |
| UPR | Unfolded protein response |
References
- World Health Organization. Global Cancer Statistics 2024; WHO: Geneva, 2024. [Google Scholar]
- Izzo, A.A.; et al. Limitations of natural product research. Pharmacol. Res. 2025, 201, 107080. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over nearly four decades. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Talib, W.H.; et al. Plant-derived anticancer agents. Molecules 2024, 29, 1234. [Google Scholar]
- Atanasov, A.G.; et al. Natural products in drug discovery. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.; et al. Quality standards in ethnopharmacology. J. Ethnopharmacol. 2022, 285, 114815. [Google Scholar]
- Izzo, A.A.; et al. Critical view on natural product bioactivity claims. Eur. J. Pharmacol. 2025, 960, 176080. [Google Scholar]
- Wolfender, J.L.; et al. Metabolomics in natural product research. Nat. Prod. Rep. 2019, 36, 855–868. [Google Scholar] [CrossRef]
- Langhans, S.A. Three-dimensional in vitro cell culture models. Front. Pharmacol. 2018, 9, 6. [Google Scholar] [CrossRef]
- Bhatia, S.N.; Ingber, D.E. Microfluidic organs-on-chips. Nat. Biotechnol. 2014, 32, 760–772. [Google Scholar] [CrossRef]
- Otero, G.; et al. Chilean medicinal plants: a bibliometric review. J. Ethnopharmacol. 2023, 300, 115731. [Google Scholar]
- Kim, C.; Kim, B. Anti-cancer natural products and their bioactive compounds. Evid. Based Complement. Altern. Med. 2018, 2018, 8654397. [Google Scholar]
- Hertz, N.T.; et al. ER stress and UPR in cancer. Cell 2018, 175, 1403–1416. [Google Scholar]
- Mujumdar, N.; et al. Triptolide induces ER stress in pancreatic cancer. Mol. Oncol. 2014, 8, 1208–1219. [Google Scholar]
- Limonta, P.; et al. ER stress in cancer: context-dependent relevance. Cancers 2019, 11, 1563. [Google Scholar]
- Liu, J.; et al. Senescence and natural metabolites. Cancer Res. 2020, 80, 3155–3168. [Google Scholar]
- Dou, Q.P.; et al. Baicalin induces senescence in colorectal cancer. Aging Cell 2018, 17, e12754. [Google Scholar]
- Chen, P.N.; et al. Pterostilbene induces senescence in lung cancer. Food Chem. Toxicol. 2017, 103, 112–121. [Google Scholar]
- Liu, B.; et al. Berberine induces senescence in glioblastoma. J. Neurooncol. 2015, 124, 25–34. [Google Scholar]
- Lee, B.Y.; et al. Senescence-associated secretory phenotype. Nat. Cell Biol. 2019, 21, 394–405. [Google Scholar]
- Khan, H.; et al. Phytochemicals and cancer metabolic reprogramming. Biomed. Pharmacother. 2024, 170, 116021. [Google Scholar]
- Liu, M.; et al. Resveratrol and glycolysis in ovarian cancer. Molecules 2018, 23, 3011. [Google Scholar]
- Wu, Q.J.; et al. Quercetin and HK2 in hepatocellular carcinoma. Cancers 2019, 11, 810. [Google Scholar]
- Wei, R.; et al. EGCG inhibits glycolytic metabolism in breast cancer. J. Agric. Food Chem. 2018, 66, 3091–3099. [Google Scholar]
- Hassan, F.U.; et al. Curcumin and HDAC in cancer. Curr. Drug Targets 2019, 20, 1249–1264. [Google Scholar]
- Ming, H.; et al. Curcumin modulates histone acetylation. Front. Pharmacol. 2022, 13, 853908. [Google Scholar]
- Zhou, C.; et al. Sulforaphane demethylates Nrf2 in colon cancer. Carcinogenesis 2019, 40, 977–985. [Google Scholar]
- Lynch, S.M.; et al. Genistein and miR-200c demethylation in prostate cancer. Oncotarget 2016, 7, 62553–62566. [Google Scholar]
- Alqathama, A.; et al. Immunomodulatory natural products. Phytomedicine 2024, 123, 155181. [Google Scholar]
- Zou, J.; et al. Curcumin and Treg cells in lung cancer. J. Clin. Invest. 2018, 128, 1516. [Google Scholar]
- Chen, X.; et al. Resveratrol and CD8+ T cells in renal tumor model. Cancer Immunol. Immunother. 2015, 64, 1333. [Google Scholar]
- Tu, S.P.; et al. Curcumin inhibits MDSCs and tumor growth. Cancer Prev. Res. 2012, 5, 205–215. [Google Scholar] [CrossRef]
- Khalil, H.; et al. Withaferin A induces ICD and PD-L1 in NSCLC. Front. Immunol. 2023, 14, 1198661. [Google Scholar]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef]
- Reuter, S.; et al. Oxidative stress, inflammation, and cancer. Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Sethi, G. Bioactive natural compounds in cancer prevention. Semin. Cancer Biol. 2016, 40, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; et al. EGCG inhibits angiogenesis. Oncol. Rep. 2014, 31, 1657–1664. [Google Scholar]
- Yang, C.H.; Horwitz, S.B. Taxol: the first microtubule stabilizing agent. Int. J. Mol. Sci. 2017, 18, 1733. [Google Scholar] [CrossRef]
- Kingston, D.G. Structural and synthetic studies on taxol. J. Org. Chem. 2007, 72, 3249–3259. [Google Scholar]
- Shishodia, S.; et al. Ursolic acid inhibits NF-kB activation. J. Biol. Chem. 2003, 278, 52148–52163. [Google Scholar]
- Fulda, S. Betulinic acid for cancer therapy. Curr. Drug Targets 2009, 10, 1545–1548. [Google Scholar]
- Mullauer, F.B.; et al. Oleanolic acid induces apoptosis in cancer cells. Apoptosis 2009, 14, 191–202. [Google Scholar] [CrossRef]
- Yin, H.; et al. Triterpenoids in cancer. Front. Pharmacol. 2018, 9, 1375. [Google Scholar]
- Pommier, Y. Topoisomerase I inhibitors: camptothecins and beyond. Nat. Rev. Cancer 2006, 6, 789–802. [Google Scholar] [CrossRef]
- Talukdar, R.; et al. Camptothecin and derivatives in cancer. Bioorg. Med. Chem. 2022, 65, 116789. [Google Scholar]
- Kim, S.; et al. Berberine inhibits cancer cell migration. Phytomedicine 2012, 19, 1166–1173. [Google Scholar]
- Hu, M.; et al. Berberine suppresses EMT in gastrointestinal cancer. Oncotarget 2018, 9, 22490–22498. [Google Scholar]
- Chu, S.C.; et al. Berberine inhibits proliferative programs. Am. J. Chin. Med. 2014, 42, 275–292. [Google Scholar]
- Babaei, G.; et al. Parthenolide and NF-kB signaling in cancer. Biomed. Pharmacother. 2018, 98, 826–833. [Google Scholar]
- Bosio, P.; et al. Sesquiterpene lactones in cancer biology. Nat. Prod. Rep. 2015, 32, 1112–1129. [Google Scholar]
- Carrasco, F.; et al. Leptocarpine exerts antiproliferative effects in gastric cancer. Phytomedicine 2023, 112, 154697. [Google Scholar]
- Gaudencio, S.P.; et al. Natural product discovery from the ocean. Nat. Rev. Chem. 2023, 7, 295–313. [Google Scholar]
- Mullowney, M.W.; et al. AI and machine learning in natural product discovery. Nat. Rev. Drug Discov. 2023, 22, 895–916. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; et al. Curcumin polymeric micelles in colorectal cancer. Int. J. Nanomed. 2015, 10, 2387–2398. [Google Scholar]
- Wang, T.; et al. Resveratrol solid lipid nanoparticles in resistant breast cancer. J. Control. Release 2021, 331, 326–340. [Google Scholar]
- Hatami, E.; et al. Quercetin nanoparticles in TNBC. J. Drug Deliv. Sci. Technol. 2023, 80, 104101. [Google Scholar]
- Xiong, Y.; et al. Nab-paclitaxel in metastatic breast cancer. Breast Cancer Res. Treat. 2024, 205, 345–355. [Google Scholar]
- Rodriguez, R.; et al. Plantas endemicas de Chile. Gayana Bot. 2018, 75, 1–131. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef]
- Holdsworth, D.K. Traditional medicinal plants of Rapa Nui. Int. J. Pharmacog. 1992, 30, 129–136. [Google Scholar]
- Estomba, D.; et al. Medicinal plants used by Mapuche communities. J. Ethnopharmacol. 2006, 107, 87–94. [Google Scholar]
- Rubio-Astudillo, J.; et al. Micropropagation of Leptocarpha rivularis and leptocarpine production. Ind. Crops Prod. 2022, 177, 114475. [Google Scholar]
- Burgos, V.; et al. Ovatifolin from Leptocarpha rivularis in melanoma. Phytochemistry 2025, 220, 114008. [Google Scholar]
- Lopez, V.; et al. Drying method effects on Ugni molinae extracts. Food Chem. 2023, 401, 134169. [Google Scholar]
- Avello, M.; et al. Ugni molinae aqueous extract in AGS cells. J. Ethnopharmacol. 2020, 248, 112316. [Google Scholar]
- Rubilar, M.; et al. Polyphenol content of Ugni molinae. J. Agric. Food Chem. 2006, 54, 7688–7693. [Google Scholar]
- Aguirre, M.C.; et al. Anti-inflammatory triterpenes from Ugni molinae. J. Ethnopharmacol. 2006, 108, 199–209. [Google Scholar]
- Calderon-Reyes, P.; et al. Calafate extracts in gastric and gallbladder cancer cells. Food Chem. Toxicol. 2020, 146, 111768. [Google Scholar]
- Sung, B.; et al. Azorella compacta extract induces apoptosis in HL60. Phytomedicine 2015, 22, 1062–1071. [Google Scholar]
- Burgos, V.; et al. Diterpenoids from Azorella compacta in MCF-7 cells. Phytochemistry 2016, 121, 45–52. [Google Scholar]
- Robles-Planells, C.; et al. Lithraea caustica extract in murine melanoma model. Front. Pharmacol. 2019, 10, 888. [Google Scholar]
- Danova, K.; et al. Biotechnology for sustainable production of plant metabolites. Biotechnol. Adv. 2022, 60, 108004. [Google Scholar]
- Ozyigit, I.I.; et al. Plant tissue culture for secondary metabolite production. Planta 2023, 258, 117. [Google Scholar]
- Heinrich, M.; et al. Access and benefit-sharing in ethnopharmacology. J. Ethnopharmacol. 2020, 246, 112221. [Google Scholar]
- Vinayagam, R.; et al. NMR metabolomics in phytochemical research. Phytochem. Rev. 2025, 24, 1–19. [Google Scholar]
- OECD. Principles of Good Laboratory Practice; OECD: Paris, 1998. [Google Scholar]
- Bolz, S.; et al. Pan-assay interference compounds in phytochemistry. J. Med. Chem. 2021, 64, 7346–7358. [Google Scholar]
Figure 1.
Mechanistic pathways of plant-derived natural compounds in cancer biology. Key mechanisms include ER stress and apoptosis (via PERK/IRE1/CHOP), senescence (p53-p21 pathway leading to cell-cycle arrest), and metabolic and immune reprogramming (involving dendritic cells, CD8+ T cells, and M1 macrophages), all contributing to disrupted cancer cell hallmarks and relative preservation of normal cell homeostasis.
Figure 1.
Mechanistic pathways of plant-derived natural compounds in cancer biology. Key mechanisms include ER stress and apoptosis (via PERK/IRE1/CHOP), senescence (p53-p21 pathway leading to cell-cycle arrest), and metabolic and immune reprogramming (involving dendritic cells, CD8+ T cells, and M1 macrophages), all contributing to disrupted cancer cell hallmarks and relative preservation of normal cell homeostasis.

Figure 2.
Nano-delivery systems for natural product-based cancer therapy. Bioactive compounds extracted from natural products are incorporated into polymeric, lipid-based, or metallic nano-delivery systems, enhancing solubility, stability, and pharmacokinetics. These precision nano-phytomedicines improve tumor targeting and therapeutic efficacy (apoptosis, anti-angiogenesis, reduced metastasis), supporting clinical translational applications.
Figure 2.
Nano-delivery systems for natural product-based cancer therapy. Bioactive compounds extracted from natural products are incorporated into polymeric, lipid-based, or metallic nano-delivery systems, enhancing solubility, stability, and pharmacokinetics. These precision nano-phytomedicines improve tumor targeting and therapeutic efficacy (apoptosis, anti-angiogenesis, reduced metastasis), supporting clinical translational applications.


Table 1.
Selected nanoformulation natural metabolites in cancer research and translation.
| Natural metabolite | Nano-system | Evidence type | Cancer model | Main outcome | Reference |
|---|---|---|---|---|---|
| Curcumin | Polymeric micelles | in vitro / in vivo | CT26; subcutaneous CT26 in BALB/c mice | Increased uptake and apoptosis; improved in vivo antitumor efficacy vs. free curcumin | Yang et al. (2015) |
| Resveratrol | TPGS solid lipid nanoparticles | in vitro / in vivo | SKBR3/PR; xenograft in BALB/c nude mice | Increased uptake, apoptosis; reduced migration/invasion; improved antitumor efficacy | Wang et al. (2021) |
| Quercetin | Solid lipid nanoparticles | in vitro | MDA-MB-231; MCF-7 | Increased antiproliferative activity and apoptosis; reduced colony formation and angiogenesis | Hatami et al. (2023) |
| EGCG | Encapsulated nanoparticles | in vitro | HCT-116, HT-29, HCT-15 | Increased uptake and anticancer activity; enhanced ROS-associated damage, DNA fragmentation, and apoptosis | Das et al. (2024) |
| Genistein | Chitosan nanoparticles | in vitro | HeLa | Increased cellular uptake and anticancer activity | Cai et al. (2017) |
| Paclitaxel | Albumin-bound nanoparticles (nab-paclitaxel) | Clinical study | Taxane-pretreated metastatic breast cancer | Improved clinical activity, PFS, ORR, and disease control | Xiong et al. (2024) |
Abbreviations: CT26, colon tumor 26 murine colon carcinoma cells; TPGS, D-alpha-tocopheryl polyethylene glycol 1000 succinate; SKBR3/PR, paclitaxel-resistant SKBR3 breast cancer cells; BALB/c, Bagg and Albino laboratory-bred mouse strain; MDA-MB-231, triple-negative human breast cancer cells; MCF-7, human breast adenocarcinoma cells; EGCG, (-)-epigallocatechin-3-gallate; ROS, reactive oxygen species; HeLa, human cervical cancer cells; PFS, progression-free survival; ORR, objective response rate.
Table 2.
Chilean plant species with documented cancer-related activity: summary of tested material, tumor models, biological outcomes, and main evidence limitations.
Table 2.
Chilean plant species with documented cancer-related activity: summary of tested material, tumor models, biological outcomes, and main evidence limitations.
| Species (authority, family), common name | Tested material / major bioactive constituents | Tumor cell model(s) | Biological activities / mechanisms | References |
|---|---|---|---|---|
| Leptocarpha rivularis DC. (Asteraceae), palo negro | Flower extracts (DCM, EtOAc, Hex, EtOH); leptocarpine (LTC); ovatifolin | AGS, MKN-45, HeLa, A2058, A375 | Antiproliferative and pro-apoptotic effects; mitochondrial depolarization; increased DEVDase activity; senescence; reduced migration/invasion; decreased IL-6 and MMP-2 | Carrasco et al. (2023); Rubio et al. (2022); Burgos et al. (2025) |
| Ugni molinae Turcz. (Myrtaceae), murta | Berry and aqueous leaf extracts; gastrointestinal digestion products; catechin, pyrogallol, alphitolic, corosolic, and asiatic acids | AGS, Caco-2 | Modest tumor-cell viability reduction; antioxidant and anti-inflammatory-associated bioactivity | Lopez et al. (2023); Avello et al. (2020); Avello et al. (2021); Aguirre et al. (2006); Rubilar et al. (2006) |
| Berberis microphylla G.Forst. (Berberidaceae), calafate | Crude and anthocyanin-rich fruit extracts; delphinidin derivatives | AGS, G415 | Reduced viability and migration; antiproliferative activity associated with anthocyanin-rich extract | Calderon-Reyes et al. (2020); Speisky et al. (2012); Lamy et al. (2006) |
| Drimys winteri J.R.Forst. & G.Forst. (Winteraceae), canelo | Ethyl acetate bark extract; drimane sesquiterpenes (drimenol, nordrimenone, isonordrimenone, polygodial); leaf essential oil | A375, MCF-7, 786-O, ACHN | Antigrowth and apoptosis-related effects; ROS-associated response; reduced Hsp70 expression; antiproliferative activity in melanoma, breast, and renal tumor cells | Russo et al. (2019); Bruna et al. (2022) |
| Peumus boldus Molina (Monimiaceae), boldo | Purified boldine | MDA-MB-231, MDA-MB-468 | Apoptosis; mitochondrial dysfunction; cytochrome c release; G2/M arrest | Paydar et al. (2014) |
| Escallonia spp. (Escalloniaceae) | Active stem extracts; major compounds not fully resolved | MCF-7, HT-29, PC-3 | Selective cytotoxicity; ROS generation; redox imbalance; mitochondrial dysfunction | Jara-Gutierrez et al. (2024) |
| Azorella compacta Phil. (Apiaceae), llareta | Methanolic extract; mulinane and azorellane diterpenoids | HL60, HepG2, SNU-1, MCF-7, HT1080, A549 | Growth inhibition; apoptosis-related signaling; active diterpenoids in breast cancer cells | Sung et al. (2015); Borquez et al. (2016) |
| Lithraea caustica (Molina) Hook. & Arn. (Anacardiaceae), litre | Standardized Litre extract (LexT) | Murine B16 melanoma model | Delayed tumor growth; topical treatment induced regression in 15% of animals; evidence points to immune-mediated antitumor response | Robles-Planells et al. (2019) |
Abbreviations: DCM, dichloromethane; EtOAc, ethyl acetate; Hex, hexane; EtOH, ethanol; ROS, reactive oxygen species; AGS, human gastric adenocarcinoma cells; MKN-45, human gastric carcinoma cells; HeLa, human cervical adenocarcinoma cells; A2058/A375, human melanoma cells; Caco-2, human colorectal adenocarcinoma cells; G415, human gallbladder carcinoma cells; MCF-7, human breast adenocarcinoma cells; 786-O, human renal cell carcinoma cells; ACHN, human renal adenocarcinoma cells; MDA-MB-231/MDA-MB-468, triple-negative human breast cancer cells; PC-3, human prostate cancer cells; HT-29, human colorectal adenocarcinoma cells; HL60, human promyelocytic leukemia cells; HepG2, human hepatocellular carcinoma cells; SNU-1, human gastric carcinoma cells; HT1080, human fibrosarcoma cells; A549, human lung carcinoma cells; B16, murine melanoma cells; LexT, standardized Lithraea caustica extract.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



