Submitted:
04 June 2026
Posted:
09 June 2026
You are already at the latest version
Abstract
Abstract: Turtles are facing worldwide declines for many reasons including emergent diseases, including shell disease. A lab population of Painted Turtles experienced an outbreak of shell disease, which created carapacial lesions, and I used the opportunity to test the resource trade-off hypothesis, which posits that carotenoids are used in colorful animals as either an integumentary colorant or for boosting health, but not both, before I treated the affected turtles for the infection and returned them back to their original habitats. The results of the experiment showed that turtles that were supplemented with dietary xanthophylls did not differ in forelimb stripe color regardless of whether they had shell disease or not, whereas turtles that lacked dietary xanthophylls with shell disease lost yellow chroma in the orange-red forelimb stripe, compared to xanthophyll supplemented turtles that did not have shell disease. Painted turtles that are prized as pets for their striking colors lose richness of stripe and spot colors when they lack access to carotenoids and experience shell disease. Turtle conservation should include turtle health and disease treatment.
Keywords:
carotenoids
; integumentary stripe color
; shell Disease
; Resource trade-off hypothesis
1. Introduction
Emergent diseases are exacerbating challenges among wild turtle populations (Frederickson et al. 2024, Fey et al., 2015; Adamovicz et al., 2018; Zhang et al., 2019; Waltzek et al., 2022). Turtles already face declines in wild population numbers because of Illegal pet trade, habitat loss & destruction, harvesting as a human food source (Nardoni and Mancianti, 2023). Turtles have been on earth for hundreds of millions of years and are among the oldest extant reptile (Nardoni and Mancianti, 2023). Despite this, many species are currently faced with extinction (King et. al, 2026; Nardoni and Mancianti, 2023).
Painted Turtles (Chrysemys picta picta) are model organisms for the study of physiology, behavior, and evolution (Valenzuela, 2009). Painted Turtles are common North American turtles whose skin possess a variety of integumentary stripes and spots that are red, yellow and orange. Turtles are prized by humans world-wide because of these colorful stripes and spots.
Painted Turtle stripe and spot colors are generated by pterins and carotenoids bound in dermal xanthophores of the integument (Bagnara, 1983; Bagnara and Matsumoto, 2006), and the colors are dependent on access to carotenoids. Painted turtle forelimb stripes are orange-red in color to the unaided human eye, chin stripes are orange-red and post-orbital spots are yellow to the human eye. These colors are in part due to access to xanthophyll carotenoids (Steffen et al., 2015, 2019).
Carotenoids are lipid-soluble photosynthetic pigments in plants that animals must acquire from the diet (Goodwin, 1984). In contrast, pterins are nitrogen-rich, ultraviolet- (UV) fluorescent compounds that animals synthesize from basic purine (e.g., guanine) precursors (McGraw, 2006). Carotenoids play diverse and important roles in photoprotection, free-radical scavenging, immunomodulation, visual tuning, and animal coloration (Vershinin, 1999), and are purported to function as honesty-reinforcing mechanisms that underlie costly yet beneficial colorful ornamental traits (Lozano, 1994; Hill, 2002; Ibanez et al., 2014).
The Hamilton-Zuk hypothesis posits that animals use secondary sex characters, like spots and stripes, because they are costly to the individual who possesses them and reflect the possessor’s health state or condition (1982). This cost may be related to maintaining the immune function (Wedekind, 1992; Wedekind and Folstad, 1994). Carotenoids are used as an integumentary colorant but because they are also known to be important to modulating the immune system (reviewed in Svensson and Wong, 2011), they mediate a trade-off in colorful animals that consume them, so that animals either use specific carotenoid molecules as an integumentary colorant or a health enhancement, but not both (Koch and Hill, 2018; Hill, 2019; Hill et al., 2023). This is known as the Resource trade-off hypothesis.
Shell Disease is a general term for lesions that occur on turtle carapaces and skin, especially in captivity (Hatt, 2010; Zwart et al., 1985). Here I present experimental research that opportunistically provides support for the Resource trade-off hypothesis of carotenoids. I was performing a behavioral lab experiment in the lab when an accidental outbreak of shell disease (actual biological cause was unknown) in a lab population, spread to several (but not all) individuals in the lab. I decided to opportunistically test the hypothesis that if shell lesions grow on the carapace of captive turtles, then half of the turtles in the lab which have been supplemented with xanthophylls, the other half which have not, then turtles will use xanthophylls for either an integumentary colorant or as an immunological agent (but not both), and I predicted that there will be an interaction between turtles who lack dietary access to carotenoids and the carotenoids use as an integumentary colorant.
I show that turtles who have been supplemented xanthophylls have lower forelimb stripe brightness than turtles who have not been xanthophyll supplemented, but that there is an interaction: In turtles with xanthophyll supplementation but who are free of shell disease forelimb (whose stripes are orange-red) yellow chroma is similar to yellow chroma in turtles who have been supplemented xanthophylls but experience shell disease. But turtles who experience shell disease but lack xanthophyll supplementation have significantly lower forelimb stripe yellow chroma compared to those that have been supplemented xanthophylls. This indicates that carotenoids are used by Painted Turtles as either an integumentary colorant or for some unidentified immunological function.
2. Methods and Materials.
Turtle collection and care. Turtles were captured with 4-foot diameter hoop nets (baited with sardines & dry pellet dog chow) in a variety of ponds throughout Jefferson County, West Virginia. Turtles were then transported back to a lab in Shepherd University upon capture, and cared for according to Shepherd University IACUC protocol # 2018-001. Turtles were assigned unique identities by carving notches along the marginal scutes with a bastard file. Turtles were then transferred into 36.8 cm×22.2cm×24.8cm (20.379L3) plastic Critter Keepers™ tanks (LLL Reptile Large Exo Terra Faunarium Critter Keeper). The Critter Keeper tanks were filled with tap water and placed on shelving units with full spectrum fluorescent tubes (Vitalite T8, 32 watt) suspended 30 cm above each tank top. The room was maintained on a14:10 hour light: dark photoperiod, at an air temperature of 28.0°C, with relative humidity ranging between 40 and 60%. The Critter Keeper water was changed every other day and scrubbed with an abrasive brush.
Initial experimental protocol: Maintenance & husbandry: Upon initial entry into the lab, turtles were fed Tetra ReptoMin™ floating food sticks ad libitum, 3-4 times per week and housed for variable lengths of time before the experiment (range May 5-June 9th, 2019). On June 10, 2019 a sufficient number of turtles were in the lab to start the experiment (N = 46), and turtles were randomly assigned to one of two diet groups: 1) fed nutritional gel with no xanthophyll supplementation (i.e., the negative control level of the independent variable, or IV hereafter) or 2) fed a nutritional gel enriched with xanthophyll supplements (i.e., the experimental level of the IV).
Once the experiment started (i.e., when a sufficient number of turtles were placed in the lab, N = 46), turtles were maintained by feeding individuals prepared and pre-cut squares of Mazuri™ Aquatic Turtle Gel (formula code 5M7N, Purina Mills, LLC, St. Louis, Missouri) ranging in mass between 3.8 and 6.8 grams. The base Mazuri Turtle Gel 5M7N (i.e., with no xanthophyll supplementation) contained 38 ppm beta-carotene and consisted of 55% protein, 15% fat, and 5% fiber. The Xanthophyll supplementation level of the experimental level of the IV consisted of the base Mazuri Turtle Gel 5M7N enriched with 40 ppm lutein in addition to the beta-carotene already present in the base diet. The experimental feeding regiment was performed for 9 weeks and 2 days (or 65 calendar days, June 10th – August 13th, 2019).
Spectrometry.—I measured spectral variation (300-700 nm) of the chin, post-orbital spot, and right forelimb stripes of Painted Turtles using an Ocean Optics (UV-Vis) S2000 spectrometer (Dunedin, Florida) coupled to a tungsten–deuterium light source. I used a bifurcated fiber-optic cable (400 µm inner diameter) mounted in a black plastic sleeve that was fastened to the probe tip and that was placed at an angle of 90° to, and 2 mm away from, the measured skin surface. I dried the turtles’ skin surfaces before the reflectance measurement, and I took three unique, adjacent, and non-overlapping measurements in the colorful stripe or spot of each body region. Color data were gathered as percent reflectance at 1 nm wavelength increments from 300 to 700 nm (integration time = 2 ms, scans to average = 22) and this output was smoothed and processed using CLR software v 1.0 (Montgomerie, 2006). From the smoothed spectral reflectance data, I constructed spectral reflectance curves for chin, forelimb and post-orbital color patches. The following wavelength ranges delimit hue for three wavebands: 300–399 nm = UV, 550–599 nm = yellow, and 650–700 nm = red. UV chroma (i.e., UVC) describes spectral purity in the UV waveband (300–399 nm), yellow chroma (i.e., YC, provided as S1Y in the Montgomerie CLR output) describes spectral purity in the yellow waveband and red chroma (RC, provided as S1R in the Montgomerie CLR output) describes spectral purity in the red waveband. Total reflectance from 300-700 nm describes overall brightness, B1 (Montgomerie, 2006). Initial spectrometric measurements were made on dates ranging from June 10 to 13, and we made a final series of spectrometric measurements on August 13th as a post-experimental measure of stripe and spot color.
Shell lesions were discovered on captive turtles on July 11, 2019. I present results of forelimb stripes only, to simplify presentation of the results, and because they were the only stripe or spot region that showed a significant effect.
Treatment of Shell Disease Before Turtles were Released Back to the Wild
Once shell disease was detected on the carapace of a turtle, the turtle was handled by me or my lab students with latex gloves. The lesion was washed with mild liquid soap with a toothbrush. The toothbrush was rinsed to ensure that the lesion causing agent was not spread to another turtle in the future. Then the lesion was dried off with a paper towel, and covered with Betadine iodine. The turtle was placed in a dry terrarium for 15 minutes. Then the turtle was placed back into its Critter Keeper tank that was scrubbed and re-filled with fresh tap water. The turtle was subjected to this treatment every day until the lesion disappeared.
Statistical analysis.—I studied the effects of shell disease lesions (IV1) on post-experimental skin stripe and spot color (the dependent variables, or DVs) when xanthophyll supplementation is a second indpendent variable, (or IV2) and pre-experimental skin stripe and spot color was treated as a covariate. By using this covariate approach, I could determine if the IV had significant effects on the DV after the covariate influence was removed.
Before this can be done, however, data need to meet all assumptions of ANOVA and ANCOVA, multivariate significance needs to be investigated, and covariates need to be assessed for reliability. If covariates are reliable, then one usually wants to determine the ability of specific covariates to adjust specific DV’s. Only after these steps have been performed can the researcher reliably evaluate the dependent variables, and infer statistical significance of the effects of factors on DV’s (Tabachnik and Fidell, 2001).
I used SPSS v. 20 to evaluate normality of distributions of spectral variation. Several variables did not meet assumptions of normality; when yellow, red or UV chroma or overall brightness was non-normally distributed, the values were Box-Cox- transformed. The transformations were retained and used for the analyses only if they improved the normality of the variable (Shapiro-Wilks W test, available in SPSS). Otherwise, the original, untransformed variables were used. After appropriate transformations were retained or rejected, the data met all assumptions of homogeneity of variance-covariance matrices.
The following analyses were all performed in SPSS in syntax mode (with syntax provided by Tabachnik and Fidell, 2001). SPSS’s MANOVA command was used to perform MANOVA and determine multivariate significance. MANOVA was also used to assess the reliability of covariates. In this assessment, Wilks’ lambda was used to test for significant relationships between the linear combinations of DV’s (representing post-experimental skin stripe and spot color) and the covariates (i.e., pre-experimental skin stripe and spot color).
I also used SPSS MANOVA to investigate the ability of covariates to adjust specific wavebands of post-experimental spectral variation (i.e., the DV’s) by performing multiple regressions for each DV in turn, with the covariates acting as IV’s in the model. Regression coefficients (beta coefficients) and statistical tests of covariate significance were provided as output in SPSS (Table 1). All pre-experimental wavebands were able to adjust post-experimental wavebands (i.e., all post-experimental spectral variation were influenced by pre-experimental spectral variation).
I used MANCOVA to analyze the effects of carotenoid supplementation on post treatment skin color (post-yellow chroma, post-red chroma, post-UV chroma, post-carotenoid chroma, or post-brightness) in Painted Turtles. Pre-experimental measures of color (pre-carotenoid chroma, pre-yellow chroma, pre-UV chroma or pre-brightness) were used as covariates. Data were Box-Cox transformed when their distributions did not meet assumptions of normality or homogeneity of variance. I measured the colorful stripes along the chin (yellow to the human eye) and fore-leg (orange-red to the human eye) and post-orbital eyespots (yellow to the human eye).
Once the contributions of covariates to adjust the DV were evaluated, I wanted to see if the DV’s (post-experimental stripe and spot color) were affected by the IV’s (shell disease lesions and xanthophyll supplementation) once the effect of covariates (pre-experimental color) were removed. Because many of the DV’s were significantly correlated to each other (i.e., they showed a correlation coefficient > 0.30, see Tabachnick and Fidell, 2001), we used SPSS MANOVA to perform Roy-Bargmann step-down analysis. In Roy-Bargmann step-down analysis, DV’s are assigned priority based on theoretical or practical considerations, and the highest priority DV is analyzed as a univariate ANOVA where the covariates are entered as IV’s. The second priority DV is then entered into ANCOVA as a DV, while the highest priority DV is entered as a covariate (along with covariates from previous ANOVA). Following a first ANCOVA, a second ANCOVA is performed, with the higher priority DV from the previous ANCOVA being entered as a covariate along with previous covariates. This succession of ANCOVA’s is performed until the lowest priority DV is entered as a DV, and all previous DV’s are entered into ANCOVA as covariates (along with initial covariates, so that adjustment is made for the previous DV’s as well as the pre-experimental spectral variation (i.e the covariates). I assigned yellow chroma as the highest priority in red-striped patches, and red chroma the second highest priority (because xanthophylls likely influence portions of the red spectral curve, i.e., yellow and red make orange). UV chroma was assigned third highest priority, H4c (carotenoid chroma, as designated by Montgomerie, 2006) and overall brightness was assigned the fourth and lowest priority.
One assumption of MANOVA is that the regression between different covariates and each DV have similar slopes, and use of the average regression to investigate the effect of the IV on the relationship between covariates and the DV’s are therefore justified (Tabachnik & Fidell, 2001). If there is heterogeneity of regression for any step (i.e., P > 0.01) within the Stepdown analysis , then one cannot reliably interpret any of the steps following the heterogeneous step. I tested the Homogeneity of regression assumption of MANOVA according to Tabachnik & Fidell (2001)) and described in detail in Steffen et al. (2019). The assumption of homogeneity of regression was satisfactory for all steps and combinations of covariates and DV’s, and the DV’s were judged to be sufficiently reliable to act as covariates Table 2.
After performing Roy-Bargmann stepdown analysis and testing for homogeneity of regression I used SPSS MANOVA to perform univariate ANCOVA on each DV, with xanthophyll supplementation as IV, and pre-experimental measures of yellow, red, and UV chroma as well as overall brightness as covariates. To test for significant effects of xanthophyll supplementation on the linear combination of DV’s that represent post-experimental skin stripe and spot spectral variation, one needs to compare significant differences in spectral variation between univariate ANCOVA and stepdown F tests. According to Tabachnik and Fiddell (2001), DV’s that show both significant univariate and stepdown F statistics are statistically pure tests of significance because they demonstrate that there is unique variability shared by the treatments after adjustments for differences created by the covariates. I combined F statistics, and effect and error degrees of freedom from the univariate and stepdown analyses and present them side by side in Table 3 to facilitate comparison and evaluate the significance of each analysis type. When results show significant univariate but not significant stepdown F statistics, there is not unique treatment variability after covariate adjustment. Thus, significance is not reliable. If there is a significant stepdown F but not a significant univariate F, significant DV’s take on importance in the presence of higher order DV’s, and the results may be related to the order in which DV’s were entered into the stepdown analysis. Thus, significance is not reliable in this case, either. I controlled for increased likelihood of Type I inflation error due to multiple comparisons by dividing a typical P value indicating an experiment-wise error rate of 0.05 by 5 (i.e., P < 0.01) to accommodate the increased probability of making type I errors among 5 separate tests (i.e., one separate test for each DV).
3. Results
Roy-Bargmann Step down Analysis showed the following results. Pre-experimental yellow chroma (preS1y) was a significant covariate of post-experimental yellow chroma in painted turtle forelimb stripes (Table 3).
There was a marginal main effect of xanthophyll supplementation on yellow chroma (Table 3, Figure 1). Xanthophyll supplementation created a marginal decrease in yellow chroma (S1y) of forelimb stripes. Forelimb stripes of xanthophyll deprived turtles (C-) had an average yellow chroma of 0.199 (S.E. = 0.004) while forelimb stripes of xanthophyll supplemented turtles (C+) had an average yellow chroma of 0.188 (S.E.=0.005). Univariate F = 2.077, Stepdown F = 2.077, P = 0.073.
There was also a significant effect of shell disease state on forelimb stripe brightness (Table 3, Figure 2). Forelimb stripes of disease-free turtles showed lower average overall brightness of 46.71 (S.E. = 7.02), and forelimb stripes of turtles with shell disease showed an overall brightness of 65.67 (S.E.= 6.9). Univariate F = 3.145, Stepdown F = 5.024, P = 0.031.
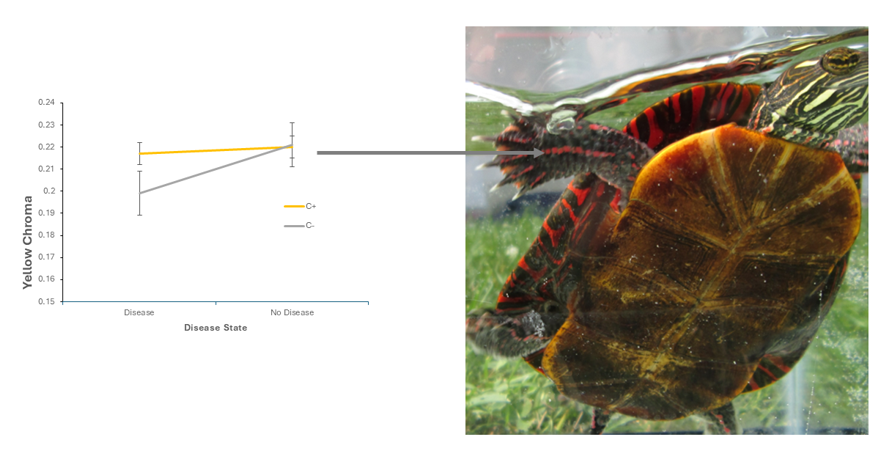
There was a marginally significant interaction between xanthophyll supplementation and shell disease on forelimb stripe yellow chroma. In turtles free of shell disease, forelimb yellow chroma is similar between turtles who are supplemented with xanthophylls and in turtles who are supplemented with xanthophylls, but experienced shell disease. But turtles who experience shell disease but lack xanthophyll supplementation have significantly lower forelimb stripe yellow chroma compared to those that were supplemented with xanthophylls (Figure 3).
Turtles that were disease free, but who were xanthophyll supplemented (i.e. C+) had an average yellow chroma of 0.217 (S.E. = 0.01), and disease free turtles who lacked xanthophyll supplementation (i.e. C-) had an average yellow chroma of 0.220 (S.E.=0.009). Among turtles with shell disease, those who were xanthophyll supplemented (i.e., C+) had an average yellow chroma of 0.199 (S.E.= 0.01) and turtles who lacked xanthophyll supplementation (i.e., C-) had an average yellow chroma of 0.221 (S.E. = 0.009). Univariate F = 3.516, Step-down F = 3.886, P = 0.056
4. Discussion
The research presented here adds to published literature demonstrating that turtles, archosaur reptiles that have been on the earth for 200 million years (Ernst and Lovich, 2009), must ingest carotenoids to maintain their colorful spots & stripes (Steffen et.al 2019). Other reptiles that have been shown to maintain color with carotenoids, which also respond to some aspect of homeostatic physiology include Frillneck lizards (Chlamydosaurus kingii, McLean et al., 2019), Common lizards (Lacerta vivipara, Fitze et al., 2009; Cote et al., 2010), Iberian Green lizards (Lacerta schreiberi, Kopena et al., 2014) and even a 50% probability of tissue-based expression in Archelosauria, Archosauria, and Aves (i.e., a clade joining turtles and archosaurs—birds and crocodilians—and their closest relatives, Davis and Clarke, 2022).
In Painted Turtles, carotenoids appear to be used for maintaining an (undetermined) aspect of health in turtles that have shell lesions, because their orange-red forelimb stripes lose chroma from the yellow waveband (i.e., spectral purity in 550-559 nm, Steffen et. al., 2019). This finding supports the resource trade-off hypothesis (Koch and Hill, 2018; Koch et al., 2019) of carotenoids as colorant of stripes and spots in turtles. The disease that provides this support is a common illness experienced by turtles in captivity (Rothstein and Steffen, 2020). Shell disease increases the overall brightness (or overall reflectance) of Painted Turtle forelimb stripes.
Painted Turtle color generating mechanisms have not been comprehensively studied, but Red-eared sliders (Trachemys scripta) and River Cooters (Pseudemmys concinna) have been studied with respect to cellular (i.e., pigmentary), ultra- and nano-structure color generating elements (Brejcha et al., 2019). How pigments (e.g. xanthophyll) interact with other mechanisms can have important outcomes for reflectance patterns of spots or stripes in turtles. The yellow forelimb stripes of both P. concinna and T. scripta contain xanthophores and iridophores, but the post-orbital regions of the two species differ in cell-type distribution. Turtles also employ novel color-generating mechanisms (e.g., nano-organization of dermal collagen).
When carotenoids are diverted from the dermal chromatophore unit to some other location, and for some other purpose (e.g. health enhancement) overall brightness increases because carotenoid saturation decreases (Shawkey et al., 2005, 2017). Finally, this research showed a marginal interaction between shell disease state, xanthophyll supplementation and yellow chroma of the orange-red forelimb stripes. Turtles who were supplemented with xanthophylls had similar forelimb stripe yellow chroma regardless of whether the turtles had shell disease or not. Presumably xanthophyll supplementation provided a sufficient amount of xanthophylls to allow turtles to use carotenoids for integumentary color maintenance and immunological purposes. But among turtles who lacked xanthophyll supplementation, forelimb stripes from turtles who had shell disease had lower yellow chroma than turtles who were free of shell disease. When carotenoids are diverted from the dermal chromatophore unit to some other location, and for some other purpose (e.g. health enhancement) overall brightness increases because carotenoid saturation decreases (Shawkey et al., 2005, 2017).
Several pieces of evidence indicate that these disease-based changes in color are perceptible to other Painted Turtles (Steffen et. al., 2019). Red-eared slider turtles (and all Emydid turtles) have a well-developed tetra-chromatic visual system (Loew and Govardovskii, 2001). Emydid turtle retina are capable of complex visual processing (Loew and Govardovskii, 2001; Zana et al., 2001). Turtles possess UVS, SWS, MWS and LWS retinal photoreceptor cones (Loew and Govardovskii, 2001). These retinal cones contain keto-carotenoid pigmented oil droplets, which aid in wavelength discrimination by reducing scattered light and improving contrast, as well as signal-to-noise ratio (Loew and Govardovskii, 2001; Neumeyer and Jäger, 1985, 2001; Zana et al., 2001; Lopes et al., 2016; Twyman et al., 2016).
Fresh water lakes and ponds that Painted Turtles inhabit transmit light maximally at longer wavelengths (Lythgoe, 1984). When viewed against low-reflecting brown colors of the carapace or skin, the increases in yellow and red spots and stripes are likely to be highly visible to conspecific individuals. Finally, Painted Turtles are a quasi-social species of reptile, and they show these stripes and spots in close contact social encounters such as basking or in water during courtship (Bury, 1979; Lovich, 1988).These results provide support for the Resource Trade-off hypothesis of carotenoids. Turtles suffering shell disease presumably lost yellow chroma in their forelimb stripes because the xanthophyll supplementation was diverted to an undetermined health use instead of the integument as a colorant (Koch and Hill, 2018).
I did not identify the cause of the shell lesions that appeared in the lab and I noted shell disease by observing lesions on the keratin-covered scutes of the carapace only. Lesions could have been caused by fungi, bacteria, and/or ecto parasites. In previous studies, wild-caught River Cooters (Pseudemmys concinna) and Yellow-Bellied Turtles (Trachemys scripta) lesions correlated with bacterial and trematode infections (Lovich et.al 1996; Garner et al., 1997). In map turtles (Graptemys species) maintained under natural conditions but brought to University of Georgia’s Veterinary Teaching Hospital, six biopsies of lesions revealed yielded aerobic isolates (Chryseobacterium indologenes, Aeromonas hydrophila, Ralstonia pickettii, and Morganella morganii), whereas eleven shell samples grew various clostridial anerobes (Hernandez-Divers et al., 2009). Emydomyces testavorans is not likely to be the causative agent of the lesions because it was found at low prevalence when investigated as causes to lesions in aquatic Painted Turtles (Frederickson et al., 2024, White et. al., 2025).
5. Conclusions
The cause of shell lesions in the present study is presumably due to poor turtle husbandry and omission of aquatic animal water conditioner in the lab (Hatt, 2010, Rothstein and Steffen, 2020, Nardoni and Manchianti, 2023). No hygienic practice was used in the present study that might harm turtles (e.g., turtle housing was not washed with bleach to sterilize tanls), and turtles were treated with standard veterinary care procedures to alleviate disease and suffering. Nevertheless, Painted Turtle color, one of the most aesthetically pleasing aspects of Painted Turtle appearance which leads to Painted Turtles to be sold through the pet trade and renders Painted Turtles as common pets, is affected by shell disease that is common to anthropogenic landscapes (i.e., captivity). In other words, shell disease, a common disease of human captivity reduces the vibrancy of Painted Turtle stripe and spot color.
Ulcerative shell disease has been found on Geomydid fossil turtles from the early Eocene in southwestern Wyoming (Zonneveld and Bartels, 2023). Thus, turtles have been co-evolving with shell disease for millions of years. Nonetheless, turtles are facing extinction at a very high rate from so many causes, that shell disease due to human captivity or other indirect anthropogenic causes needs to be better understood and treated. Chelonian conservation efforts should continue to include turtle health and disease treatment.
References
- Adamovicz, L.; Allender, M. C.; Archer, G.; Rzadkowska, M.; Boers, K.; Phillips, C.; Chu, C. Investigation of multiple mortality events in eastern box turtles (Terrapene carolina carolina). PLoS ONE 2018, 13(4), e019617. [Google Scholar] [CrossRef]
- Bagnara, J.T. Developmental aspects of vertebrate chromatophores. Amer. Zool. 1983, 23, 465–478. [Google Scholar] [CrossRef]
- Bagnara, J.T.; Matsumoto, J. Comparative anatomy and physiology of pigment cells in non-mammalian tissues. In The Pigmentary System; Nordlund, J.J., Boissy, R.E., Hearing, V.J., King, R., Oetting, W., Ortonne, P.J., Eds.; Blackwell Publishing: Malden, MA, 2006; pp. 11–59. [Google Scholar]
- Cote, J.; Meylan, S.; Clobert, J.; Voituron, Y. Carotenoid-based coloration, oxidative stress and corticosterone in common lizards. J. Exp. Biol. 2010. 213(12), 2116–2124. [CrossRef]
- Davis, S. N.; Clarke, J. A. Estimating the distribution of carotenoid coloration in skin and integumentary structures of birds and extinct dinosaurs. Evolution 2022, 76(1), 42–57. [Google Scholar] [CrossRef] [PubMed]
- Ernst, C.H.; Lovich, J.E. Turtles of the United States and Canada, 2nd ed.
- Johns Hopkins University Press; 2009; p. 7.
- Fitze, P. S.; Cote, J.; San-Jose, L. M.; Meylan, S.; Isaksson, C.; Andersson, S.; Clobert, J. Carotenoid-based colours reflect the stress response in the common lizard. PLoS ONE 2009, 4(4), e5111. [Google Scholar] [CrossRef] [PubMed]
- Fredrickson, K.; Adamovicz, L.; Terio, K.; Davidson, A.; Ryan, M.; Waligora, M.; Allender, M. C. Emydomyces testavorans surveillance in multiple free-ranging terrestrial and aquatic chelonian species in Illinois, USA. J. Wild. Dis. 2024, 60(4), 850–859. [Google Scholar] [CrossRef]
- Fey, S.B.; Siepielski, A.M.; Nusslé, S.; Cervantes-Yoshida, K.; Hwan, J.L.; Huber, E.R.; Fey, M.J.; Catenazzi, A.; Carlson, S.M. Recent shifts in the occurrence, cause, and magnitude of animal mass mortality events. Proc. Nat. Acad. Sci. 2015, 112(4), 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, T. W. Carotenoid—protein complexes. In The Biochemistry of the Carotenoids: Volume II Animals.; Springer Netherlands: Dordrecht, 1984; pp. 1–21. [Google Scholar]
- Hamilton, W. D.; Zuk, M. Heritable true fitness and bright birds: a role for parasites? Science 1982, 218(4570), 384–387. [Google Scholar] [CrossRef]
- Hatt, J. M. Dermatological diseases in reptiles. Schweiz. Arch. Fur Tierheilkd. 2010, 152(3), 123–130. [Google Scholar] [CrossRef]
- Hill, G. E.; Inouye, C. Y.; Montgomerie, R. Dietary carotenoids predict plumage coloration in wild house finches. Proc.Roy. Soc. London. Ser. B Biol. Sci. 2002. 269(1496), 1119–1124. [CrossRef]
- Hill, G. E.; Weaver, R. J.; Powers, M. J. Carotenoid ornaments and the spandrels of physiology: a critique of theory to explain condition dependency. Biol. Rev. 2023, 98(6), 2320–2332. [Google Scholar] [CrossRef]
- Ibáñez, A.; Polo-Cavia, N.; López, P.; Martín, J. Honest sexual signaling in turtles: experimental evidence of a trade-off between immune response and coloration in red-eared sliders Trachemys scripta elegans. Naturwissenschaften 2014, 101(10), 803–811. [Google Scholar] [CrossRef]
- King, R. B.; Edmonds, D. A.; Adamovicz, L.; Ballard, S.; Blankenship, R.; Blogg, R.; Towey, J. B. Quantification of human-caused mortality in an imperiled turtle. Wild.Biol. 2026, e01686. [Google Scholar] [CrossRef]
- Koch, R. E.; Hill, G. E. Do carotenoid-based ornaments entail resource trade-offs? An evaluation of theory and data. Funct. Ecol. 2018, 32(8), 1908–1920. [Google Scholar] [CrossRef]
- Kopena, R.; López, P.; Martín, J. What are carotenoids signaling? Immunostimulatory effects of dietary vitamin E, but not of carotenoids, in Iberian green lizards. Naturwissenschaften 2014, 101(12), 1107–1114. [Google Scholar] [CrossRef]
- Lozano, G. A. Carotenoids, parasites, and sexual selection; Oikos, 1994; pp. 309–311. [Google Scholar]
- McLean, C. A.; Lutz, A.; Rankin, K. J.; Elliott, A.; Moussalli, A.; Stuart-Fox, D. Red carotenoids and associated gene expression explain colour variation in frillneck lizards. Proc. Roy. Soc. Ser. B Biol. Sci. 2019, 286, 1907. [Google Scholar] [CrossRef] [PubMed]
- McGraw, K.J. The mechanics of uncommon bird colors: pterins, porphyrins, and psittacofulvins. In Bird Coloration. I. Mechanisms and Measurements.; Hill, G.E., McGraw, K.J., Eds.; Harvard University Press: Cambridge, MA, 2006; pp. 354–398. [Google Scholar]
- Montgomerie, R. Analyzing colors. In Bird Coloration. I. Mechanisms, Function and Evolution.; Hill, G.E., McGraw, K.J., Eds.; Harvard University Press: Cambridge, MA, 2006; pp. 90–147. [Google Scholar]
- Nardoni, S.; Mancianti, F. Mycotic diseases in chelonians. J. Fungi 2023, 9(5), 518. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, L.; Steffen, J. Influences of husbandry on coloration of turtles. Radiata 2020, 29(1), 10–21. [Google Scholar]
- Shawkey, M. D.; Hill, G. E. Carotenoids need structural colours to shine. Biol. Lett. 2005, 1(2), 121–124. [Google Scholar] [CrossRef]
- Shawkey, M. D.; D'Alba, L. Interactions between colour-producing mechanisms and their effects on the integumentary colour palette. Phil. Trans. Roy. Soc. B Biol. Sci. 2017, 372, 1724. [Google Scholar]
- Steffen, J. E.; Hultberg, J.; Drozda, S. The effect of dietary carotenoid increase on painted turtle spot and stripe color. Comp. Biochem. Physiol. B Biochem. Molec. Biol. 2015, 229, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Steffen, J. E.; Quigley, R.; Whibley, I.; McGraw, K. J. Carotenoid deprivation and beta-carotene's effects on male and female turtle color. In Comp. Biochem. Physiol. B: Biochem. and Molec. Biol. 2019. 253; Tabachnik and Fidell, Ed.; 2001; p. 110546. [Google Scholar]
- Svensson, Wong. Carotenoid-based signals in behavioural ecology: a review. Behaviour 2011, 148(2), 131–189. [Google Scholar] [CrossRef]
- Valenzuela, N. The painted turtle, Chrysemys picta: A model system for vertebrate evolution, ecology, and human health. Cold Spring Harb. Protoc. 2009, 7, pdb–emo124. [Google Scholar] [CrossRef]
- Vershinin, A. Biological functions of carotenoids-diversity and evolution. Biofactors 1999, 10(2-3), 99–104. [Google Scholar] [CrossRef]
- Waltzek, T. B.; Stacy, B. A.; Ossiboff, R. J.; Stacy, N. I.; Fraser, W. A.; Yan, A.; Shender, L. A. A novel group of negative-sense RNA viruses associated with epizootics in managed and free-ranging freshwater turtles in Florida, USA. PLoS Path 2022, 18(3), e1010258. [Google Scholar] [CrossRef] [PubMed]
- Wedekind, C. Detailed information about parasites revealed by sexual ornamentation. Proc. Roy. Soc. London. Ser. B Biol. Sci. 1992, 247(1320), 169–174. [Google Scholar]
- Wedekind, C.; Folstad, I. Adaptive or nonadaptive immunosuppression by sex hormones? Am. Nat. 1994, 143(5), 936–938. [Google Scholar] [CrossRef]
- Z Zhang, W.; Chen, B.; Niu, C.; Yuan, L.; Jia, H.; Storey, K. B. Response of the Chinese soft-shelled turtle to acute heat stress: insights from the systematic antioxidant defense. Front. Physiol. 2019, 10, 710. [Google Scholar] [CrossRef]
- Zonneveld, J. P.; Bartels, W. S. The occurrence and significance of ulcerative Shell disease (USD) on geoemydid turtles, middle Eocene, southwestern Wyoming, United States. Palaios 2023, 38(11), 453–473. [Google Scholar] [CrossRef]
- Zwart, P.; Schroder, H.D. Reptilien. In Handbuch der Zootierkrankheiten; Ippen, R., Schroder, H.D., Elze, K., Eds.; Akademia Verlag: Berlin, Germany, 1985; p. 349. [Google Scholar]
Figure 1.
Xanthophyll supplementation had a main effect on forelimb stripe yellow chroma. Xanthophyll supplementation decreased yellow chroma of forelimb stripes. Univariate F = 2.077, Stepdown F = 2.077, P = 0.073.
Figure 1.
Xanthophyll supplementation had a main effect on forelimb stripe yellow chroma. Xanthophyll supplementation decreased yellow chroma of forelimb stripes. Univariate F = 2.077, Stepdown F = 2.077, P = 0.073.

Figure 2.
The effect of shell disease on forelimb overall brightness, B1. Shell disease increases forelimb stripe brightness. Shell disease increased overall brightness of forelimb stripes. Univariate F = 3.145, Stepdown F = 5.024, P = 0.031.
Figure 2.
The effect of shell disease on forelimb overall brightness, B1. Shell disease increases forelimb stripe brightness. Shell disease increased overall brightness of forelimb stripes. Univariate F = 3.145, Stepdown F = 5.024, P = 0.031.

Figure 3.
The effect of shell disease on Painted Turtle forelimb yellow chroma when turtles are not supplemented with xanthophylls (i.e., C-). C+ shows forelimb yellow chroma when turtles are supplemented with xanthophylls. Univariate F = 3.516, Step-down F = 3.886, P = 0.056.
Figure 3.
The effect of shell disease on Painted Turtle forelimb yellow chroma when turtles are not supplemented with xanthophylls (i.e., C-). C+ shows forelimb yellow chroma when turtles are supplemented with xanthophylls. Univariate F = 3.516, Step-down F = 3.886, P = 0.056.


Table 1.
Multiple regression Beta coefficients to assess ability of specific covariates to adjust DV's in painted turtle color stripes and patches.
Table 1.
Multiple regression Beta coefficients to assess ability of specific covariates to adjust DV's in painted turtle color stripes and patches.
| Region | Covariate | Post YC | Post RC | Post UVC | Hue (H4b) | Post B1 |
|---|---|---|---|---|---|---|
| Fore | YC | 1.081** | 0.418 | -0.75 | 0.38 | 0.381 |
| RC | 0.212 | 0.06 | -0.167 | 0.139 | -0.208 | |
| UVC | 0.924* | 0.461 | -0.816 | 0.392 | 0.207 | |
| Hue (h4b) | 0.064 | 0.165 | -0.07 | -0.219 | -0.061 | |
| B1 | -0.371 | -0.063 | 0.197 | 0.139 | 0.016 |
Pre=pre-experimental spectral measurements, post=post experimental; spectral measurements. YC=yellow chroma, RC=red chroma, and UVC=ultra-violet chroma. B1=overall brightness. *represents beta coefficient values where P<0.05 and specific covariates significantly adjust the DV.
Table 2.
Results of SPSS test of Homogeneity of regression. The assumption of homogeneity of regression was satisfactory for all steps and combinations of covariates and DV’s, and the DV’s were judged to be sufficiently reliable to act as covariates.
Table 2.
Results of SPSS test of Homogeneity of regression. The assumption of homogeneity of regression was satisfactory for all steps and combinations of covariates and DV’s, and the DV’s were judged to be sufficiently reliable to act as covariates.
| Region | Source (Interaction between IV & Covariates | Step | DV | F | P |
|---|---|---|---|---|---|
| Forelimb Stripe | PreS1y*PreS1R*PreS1UV*PreH4C*PreB1 BY Lesions | 1 | Post S1y | 0.63 | 0.431 |
| PostS1y*PreS1y*PreS1R*PreS1UV*PreH4C*PreB1 BY Lesions | 2 | Post S1 R | 0.42 | 0.522 | |
| PostS1R*PostS1y*PreS1y*PreS1R*PreS1UV*PreH4C*PreB1 BY Lesions | 3 | Post S1UV | 4.37 | 0.044 | |
| PostS1UV*PostS1R*PostS1y*PreS1y*PreS1R*PreS1UV*PreH4C*PreB1 BY Lesions | 4 | Post H4c | 0.14 | 0.709 | |
| PostH4c*PostS1UV*PostS1R*PostS1y*PreS1y*PreS1R*PreS1UV*PreH4C*PreB1 BY Lesions | 5 | Post B1 | 1.83 | 0.185 |
Table 3.
Univariate and Stepdown F tests for assessment of statistically significant differences in post-experimental painted turtle stripe and spot color (DV) due to manipulation of the IV (C+, xanthophyll supplementation). Covariates (Cov) are the pre-experimental stripe and spot spectral variables (and are the same spectral variables as described for the DV below).
Table 3.
Univariate and Stepdown F tests for assessment of statistically significant differences in post-experimental painted turtle stripe and spot color (DV) due to manipulation of the IV (C+, xanthophyll supplementation). Covariates (Cov) are the pre-experimental stripe and spot spectral variables (and are the same spectral variables as described for the DV below).
| Color Region | Source | DV | Univariate F | df | Stepdown F | df | POBS | PPRED |
|---|---|---|---|---|---|---|---|---|
| Fore | Covar | YC | 0.58 | 5,40 | 0.58 | 5,40 | 0.002 | 0.01 |
| RC | 4.967 | 5,40 | 4.702 | 5,39 | 0.715 | 0.01 | ||
| UVC | 1.44 | 5,40 | 0.761 | 5,38 | 0.584 | 0.01 | ||
| Hue (h4b) | 0.483 | 5,40 | 1.3382 | 5,37 | 0.254 | 0.01 | ||
| B1 | 0.216 | 5,40 | 0.31 | 5,36 | 0.904 | 0.01 | ||
| C+ | YC | 2.077 | 1,40 | 2.077 | 1,40 | 0.073 | 0.01 | |
| RC | 5.542 | 1,40 | 3.391 | 1,39 | 0.157 | 0.01 | ||
| UVC | 2.048 | 1,40 | 0.905 | 1,38 | 0.348 | 0.01 | ||
| Hue (h4b) | 0.128 | 1,40 | 0.24 | 1,37 | 0.627 | 0.01 | ||
| B1 | 0.59 | 1,40 | 0.947 | 1,36 | 0.337 | 0.01 | ||
| Shell Disease | YC | 0.307 | 1,40 | 0.307 | 1,40 | 0.294 | 0.01 | |
| RC | 1.475 | 1,40 | 1.13 | 1,39 | 0.583 | 0.01 | ||
| UVC | 1.102 | 1,40 | 0.401 | 1,38 | 0.53 | 0.01 | ||
| Hue (h4b) | 0.249 | 1,40 | 0.029 | 1,37 | 0.866 | 0.01 | ||
| B1 | 3.145 | 1,40 | 5.024 | 1,36 | 0.031 | 0.01 | ||
| C+ x Shell Disease | YC | 0.063 | 1,40 | 0.063 | 1,40 | 0.059 | 0.01 | |
| RC | 3.516 | 1,40 | 3.775 | 1,39 | 0.803 | 0.01 | ||
| UVC | 0.921 | 1,40 | 0.355 | 1,38 | 0.555 | 0.01 | ||
| Hue (h4b) | 0.401 | 1,40 | 0.142 | 1,37 | 0.709 | 0.01 | ||
| B1 | 0.193 | 1,40 | 0.975 | 1,36 | 0.33 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



