Submitted:
04 June 2026
Posted:
08 June 2026
You are already at the latest version
Abstract
Background/Objectives: Parvovirus infection cause sever diseases in both feline and canine species, mostly affects adult cats and dogs, but cause higher risk in the kittens and puppies. This virus is known to be contagious; the simple way of spreading occurs through food and shelter as well as hands and clothing of people. The recovered species may continue to shed parvovirus in their feces for an extended period, leading to severe environmental contamination. There is no universal vaccine available that protect dogs and cats against parvovirus infections. The main goal of this study is to design a pan parvovirus multiepitope DNA based vaccine that could be administered to dog and cats. Methods: We utilized AI-machine learning incorporated server tools such as IEDB and NetMHCpan to predict B-cell and T-cell epitopes. VaxiJen and ToxinPred were used to analyze immune characteristic features and docking with feline alleles using HADdock server. Following, the immune response and stability of vaccine construct was confirmed with disulfide engineering, Normal mode analysis and molecular docking, and dynamic simulations were done with Toll like receptors of both feline and canine (TLR4 and TLR5) for 50ns. The triggered immune response was determined with immuno-simulation (ImmSim) and their activity in biological environment was reinforced with in silico cloning. Results: The B cell epitopes (NS1 - 9, NS2 - 4, VP1- 12 and VP2- 9) predicted with IEDB database were subjected to antigenicity prediction. MHC class I and IFN prediction and MHC class II and IL-4 prediction were done with IEDB and NetMHCpan. The T cell epitopes showed high binding affinities with the feline alleles. The final vaccine was designed by combining the top-ranked B-cell epitopes T- cell epitopes, filtered from high antigenic, non-allergic, non-toxic and good solubility, and with the better binding affinity score of the structural and non- structural proteins (NS1, NS2, VP1 and VP2) of feline and canine parvoviruses through linkers and adjuvants. The disulfide bond prediction and Normal mode analysis showed our vaccine construct are stable and flexible. The molecular docking analysis was performed between the designed vaccine epitopes and the TLRs (TLR4 – feline and TLR5 – canine) with Biovia Discovery Studio using Zdocker, it showed the better binding interaction with value of 22.26 (Zdock score), -47.409 (Zrank score) for feline and 16.54 (Zdock score), -134.295 (Zrank score) for canine. Conclusions: The pan multi-epitope DNA based vaccine combining the four major structural proteins (NS1, NS2, VP1 and VP2) possess dual purpose to protect both feline and canine species against the parvovirus were designed and constructed. The molecular docking and dynamic simulation analysis showed higher binding affinities and stable conformation with canine (TLR5) and feline (TLR4) toll like receptors. Though computational analysis will support us to predict the more precise top-ranked epitopes and their immuno-antigenic properties, further experimental validation will be required to be used against those viruses.
Keywords:
feline
; canine
; parvovirus
; multi-epitope DNA vaccine
; B-cell and T-cell epitope predictions
; disulfide bond and normal mode analysis
; molecular docking and dynamic simulation
; Immune simulation
; In silico cloning
1. Introduction
The parvovirus causes acute infections that are highly contagious and potentially life-threatening diseases affecting both feline and canine species in adult animals especially in kittens and puppies. It is primarily characterized by non-suppurative myocarditis and acute hemorrhagic enteritis in both the species, producing high contagiousness and lethality [1]. Both the feline and canine behave quite similarly, researchers found that in early 1970s, the parvovirus originally evolved from feline and begin infecting the canine species with slight modifications [2,3]. The main disadvantage is that the virus becomes infectious during incubation and before clinical signs become evident, results in finding it difficult to differentiate between the healthy and infectious ones. This is the reason that animals can enter or leave the shelter being healthy, even though they are affected. This concept is very important when it comes to implementing outbreak control measures.
The existing way of measures is through parvo diagnosis with IDEXX brand SNAP test, though positive results should be considered even the animals are vaccinated. The strains of parvovirus aren’t harder to diagnose or prevent, which could be detected with in-house fecal ELISA tests. Feline parvovirus (FPV) and canine parvovirus (CPV) don’t mutate rapidly the way influenza viruses and other RNA viruses does, thus the available vaccine in the market produced by major manufacturers, provide excellent immunity to all variants [4]. The adult animals could contract kittens and puppies are most at risk of this serious disease [5]. The protection is not increased by multiple vaccination, as the small species will be getting only few times, which makes them harder to fight against those viruses [6].
The conventional vaccines such as live attenuated and inactivated vaccines have been extensively used to protect animals and birds for many viral diseases. Despite the benefits and the efficacy of some of these vaccines in the protection of animals and birds against these viral diseases, they have some concerns particularly reversion to virulence in case of the live attenuated vaccines in some cases and the pos vaccination reactions in case of the live attenuated vaccines. Some of these vaccines are outdated and prepared long time ago thus the used viral strains in these vaccines do not match the currently circulating filed strains which hamper the success of these vaccines. The ideal vaccine should offer a good humeral and cell mediated immunity against he currently circulating strains of the viral pathogen of interest. The emergence of the new artificial intelligence tools particularly the machine learning, the molecular docking and simulation enhance the vaccine design and development and shorten the required time for vaccine validation pipelines for the human and veterinary uses. The next generation sequencing technology offers a strong support to the screening of some antiviral therapy and vaccine design and development through decoding the genetic materials of the currently circulating strains of the viral pathogens of interest [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22]. Integration of the NGS and AI-driven tools will enhance the vaccine design and development and increase the efficacy of the designed vaccines and minimize the number of animals used for the vaccine pipelines [23,24,25,26,27,28].
The current vaccines such as inactivated vaccines, live-attenuated, nucleic acid and subunit vaccines provide vast measures with few limitations. Those vaccines don’t have intact pathogens, eliminating the risk of viral shedding and avoiding allergic reactions caused by irrelevant antigenic components. Erin et al. developed modified live FPV (MLV) vaccines and their results showed induced immunity effect in kittens compared to inactivated vaccines [29]. Though there are more effective vaccines, animals still lose their lives to these diseases due to misconceptions among pet owners and even some veterinary professionals. Overvaccination provides concern among pet owners and veterinary professionals, choose to vaccinate less frequently or not vaccinating at all. Indeed, the American Veterinary Medical Association’s handout (2010) boldly announced that in the past, these viruses was a leading cause of death in cats and dogs, but today it is an uncommon disease, due in large part to the availability and use of very effective vaccines. Wang et al., developed a vaccine with FPV virus forming them like particle using recombinant baculovirus system, produces strong immunogenicity and protection against FPV in cats [30].
The other form of vaccine is epitope vaccines, a type of subunit vaccine, are composed of multiple neutralizing antigenic epitopes, results with the production of higher level of antibodies. Unlike traditional vaccines, these vaccines not only induce immune responses in hosts, but it also excludes the pathogen itself, which prevents viral transmission or mutations, results in enhanced safeness [31]. The sizes of CPV and FPV are approximately 25nm and its genome has two open reading frames (ORFs). The first ORFs have two non-structural proteins (NS1 and NS2), while the second ORFs have two structural proteins (VP1 and VP2). In this, VP2 is the most critical, that induces the antibody production, making it better candidate for vaccine design [6]. Many researchers have used this structural protein for developing genetically engineered vaccines and have been experimentally validated or predicted as potential antigenic sites. The mapping of these antigenic epitopes is very important and current methods for evaluating these epitopes is done through chemical cleavage or enzymatic digestion, X-ray crystallography and nuclear magnetic resonance (NMR) analysis and Computational approaches [32,33].
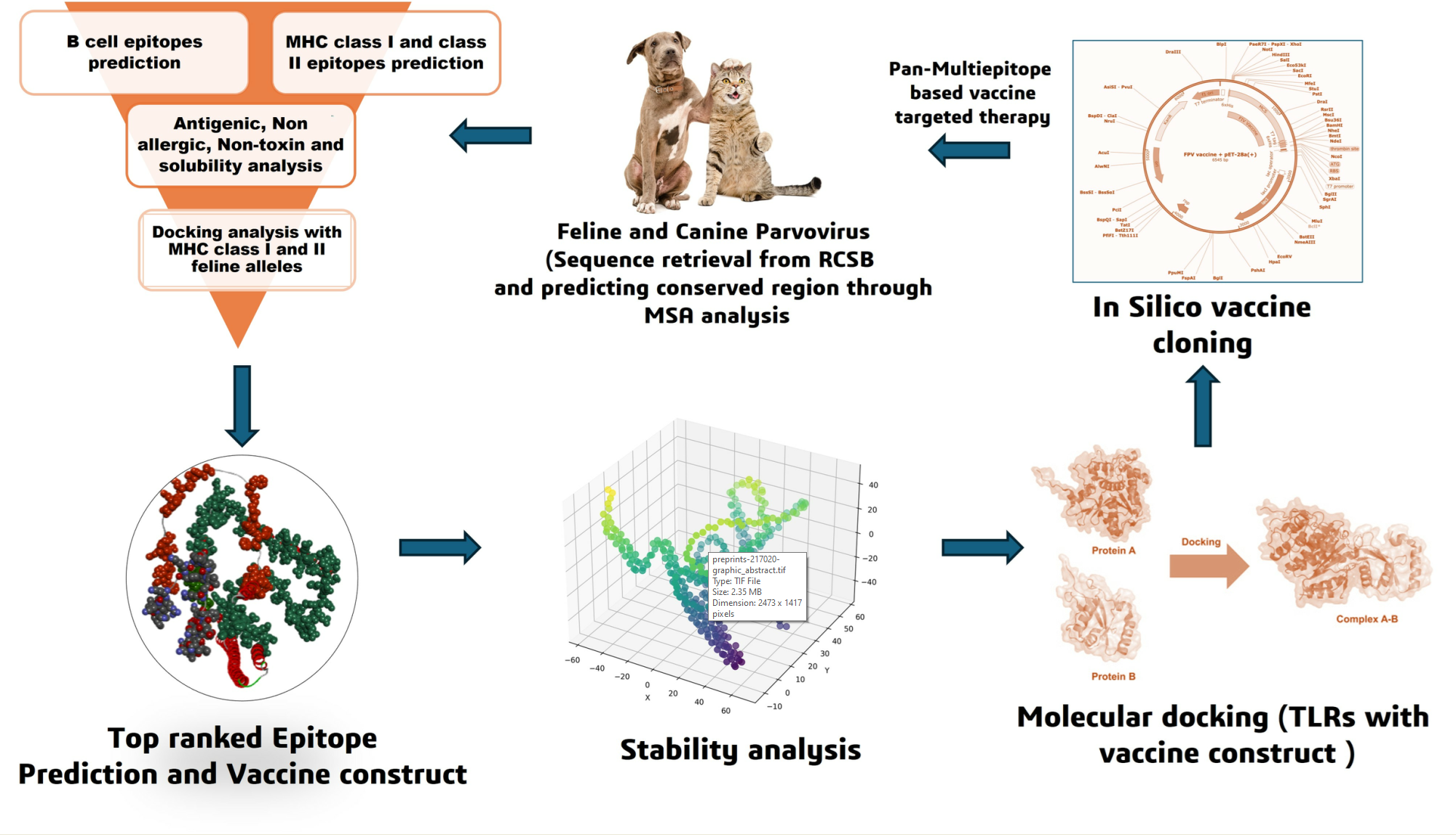
In the current study, we intend to develop a pan multiepitope based vaccine to possess dual purpose for protecting both canine and feline species from dreadful parvovirus infections in dogs and cats. We incorporated different computational AI tools to predict the top ranked epitopes and validated them through analyzing immunogenic characteristic features of predicted epitopes and binding affinity scoring from docking studies with feline alleles. The immune receptor such as TLRs were docked with vaccine construct and its stability were evaluated with Molecular dynamic simulations. The vaccine construct was cloned through in silico cloning to understand its behavior in the biological living environment. This achievement has laid a solid foundation for the development of highly effective and safe epitope vaccines, and meanwhile, it has provided an important basis for the prevention and control of viral infectious diseases.
2. Materials and Methods
2.1. Retrieval of the Canine and Feline Parvoviruses Protein Sequences fron the GenBank
A total of 143 isolate sequences belong to the two structural and two non-structural proteins of canine and feline species from different regions such as china, USA, Brazil, South Korea, Uk and Australia were retrieved from the National Center for Biotechnology (NCBI) database (https://www.ncbi.nlm.nih.gov/protein). The Supplementary Table 1 presents information about these sequences.
2.2. The Multiple Sequence Alignment (MSA) Analysis
The Geneious software (https://www.geneious.com/) and the clustal Omega server tool (https://www.ebi.ac.uk/jdispatcher/msa/clustalo) were used to analysis MSA for all the structural proteins (NS1, NS2, VP1 and VP2) of feline and canine species [34]. The most conserved region between the structural proteins of both the species were further considered for the predicting B cell and T cell (MHC class I and class II of molecules) class of epitopes.
2.3. Representing B Cell Epitopes Within the Conserved Region of Structural Proteins of (NS1, NS2, VP1 and VP2)
2.3.1. Mapping of the Linear B-Cell Epitopes Across Parvoviruses Protein Sequences
The linear B-cell epitopes were identified from the sequences of non-structural and structural proteins (NS1, NS2, VP1 and VP2) of both the feline and canine species. We used BCPREDS (BepiPred 2.0) (http://services.healthtech.dtu.dk/services/BepiPred-2.0/) [35] and the IEDB analysis resource server (http://tools.iedb.org/bcell/) for the prediction and kept the length of targeted epitopes to 20mers. The predicted epitopes were subjected to filtration based on their antigenicity, allergenicity, toxicity, and solubility as previously explained.
2.3.2. Mapping the Discontinuous/ Conformational B-Cell Epitopes Across Parvoviruses Protein Sequences
The identification of discontinuous/conformational B cell epitopes was performed with CBTOPE (https://webs.iiitd.edu.in/raghava/cbtope/) web server [36]. The threshold value was set to default, and the study was conducted using the specific amino acid composition as an input feature for support vector machine (SVM) with prediction accuracy of more than 85% and area under curve (AUC) 0.9. The Biovia discovery studio was used to represent the position of epitopes on 3D structures of all the structural proteins.
2.4. Mapping the T-Cell Epitopes Within the Non-Structural and Structural Proteins of Canine and Feline Species (NS1, NS2, VP1 and VP2)
2.4.1. Identification of the MHC Class-I Epitopes (Cytotoxic T-Lymphocyte)
The Cytotoxic T-lymphocyte epitopes were identified using the IEDB server (http://tools.iedb.org/main/tcell). The epitope binding predictor NetMHCpan 4.1 BA (version 2023.09) was used to sort the peptides by the percentile rank score and IC50 value and they were subjected to fundamental characteristic analysis to be selected as best ranking epitopes [37]. The source species were entered as canine with the peptide length of ~9 to 10mers and the associate canine alleles (DLA -8803401, DLA -8850101, and DLA -8850801). We used canine alleles to predict feline species as their sequences were conserved and we confirmed with MSA.
2.4.2. Identification of MHC Class-II Epitopes (helper T-Lymphocyte)
The helper T-lymphocyte epitopes were identified using NetMHCIIpan-4.3 (https://services.healthtech.dtu.dk/services/NetMHCIIpan-4.3/), pan-specific binding of peptides to MHC class II molecules of know sequence using the experimentally predicted feline alleles (DRB1_0103, DRB1_0102 and DRB1_0107) in comparison with canine and human alleles [38,39]. The characteristic analysis was used as previously described and the filtrated epitopes used for further analysis.
2.5. Evaluation of the Cytokines Producton Potenial (IFNs and IL-4)
The immune response of HTL epitopes have been evaluated through their cytokines inducing potential (IFNs) utilizing the IFN prediction software (https://webs.iiitd.edu.in/raghava/ifnepitope) [40]. This prediction helps us to understand the intensive upregulating effect on exhibiting antigen present on both MHC class I and class II class of molecules. Its primarily increases the expression of MHC class I molecules compared to MHC class II molecules. Followed by, analysis of Interleukin -4(IL-4) (https://webs.iiitd.edu.in/raghava/il4pred) [41] were employed to predict the capacity of filtered epitopes to induce the IL-4, which are essential for regulating immune homeostasis. Along with this, the Th2 associated cytokine, increases the expression of MHC class II molecules on antigen presenting cells such as B cell and macrophages, promoting the antigen presentation causing minimum impact on MHC class I.
2.6. Analysis of the Interaction Between the T-Cell Epitopes (MHC Class I and MHC Class II Molecules ) with Feline Alleles (MHC I and MHC II Class) Through the Molecular Docking
The PEP-FOLD3 (De novo peptide structure prediction) server (https://mobyle.rpbs.univ-paris-diderot.fr/cgi-bin/portal.py#forms::PEP-FOLD3), using sOPEP energy function were used to cluster peptide conformation (3D structures). The sequence of feline alleles (MHC class I – Q95481, MHC class II – Q19430) were reterived from Uniprot and were generated using Biovia discovery studio. The molecular docking analysis were performed between the selected epitopes and feline alleles (MHC class I and II epitopes) [42]. The PDBsum server were used to visualize the peptide – ligand interaction and key residues contact. The epitopes with best binding score were processesd to design the vaccine construct.
2.7. Assembly of the Multi-Epitope DNA Panparvoviruses Vaccine Using the Top-Ranked Epitopes
The construction of final vaccine was done by linking the B-cell and T-cell predicted epitopes that were filtered out based on the predicted (antigenic, non-allergic, non-toxic, and good solubility) of the selected epitopes. The N-terminal ends of the vaccine construct were linked with Salmonella enterica flagellin FliC adjuvants (GGS), along with the PADRE sequence, by EAAAK. The top ranked B and T cell epitopes were linked using KK, AAY, GPGPG as linkers. And the sequence was provided with 6×His-tag (H) and stop codon (TAA) attached to the C-terminus for purification and identification of the vaccine upon expression.
2.8. Disulfide Bond Engineering of the Multi-Epitope DNA Panparvoviruses Vaccine
The importance of creating the disulfide bond between the cysteine residues would be essential to keep the protein more stable. After refinement, the vaccine protein was further submitted to the Disulfide by Design v2.12 web server (http://cptweb.cpt.wayne.edu/DbD2/) for disulfide bridging [43]. The χ3 value and the Cα-Cβ-Sγ angle were kept as default parameters. As previous studies suggested, the χ3 angle must range between −87 and +97°, and the energy score must not exceed 2.2 kcal/mol for disulfide bridging. In the end, to allow bridging between possible residue pairs, they had undergone mutation to cysteine residues applying the server’s “Create/View Mutant” option.
2.9. Evaluation of the Stability of the Designed Vaccine Using the Normal-Mode Analysis (NMA)
The stability and physical mobility of the atoms from the constructed vaccine molecules were analyzed using normal mode analysis (NMA). It was conducted in iMODS server tools (https://imods.iqf.csic.es/) where multiple descriptors such as eigenvalues, covariance, B-factors, and deformability and intrinsic motions of a multiplex of proteins were analyzed [44]. The assessment of motif stiffness was done using eigenvalues, whereas the deformity of the main chain was predicted from the biological targets’ efficacy measurement.
2.10. Assessment of the Physiochemical Properties of the Designed Pan Parvovirus Multi-Epitope DNA Based Vaccine Construct
We utilized the same methodology described in our previous work, the physicochemical properties of the designed protein were assessed using the Protparam server (https://web.expasy.org/protparam/) . The potent antigenicity of selected proteins was predicted by using VaxiJen v2.0 server (http://www.ddg-pharmfac.net/vaxijen/VaxiJen/VaxiJen.html) with a default threshold of 0.4. The allergenicity and toxicity of proteins were assessed by using AllerTOP v.2.1 server (http://ddg- pharmfac.net/AllerTOP/), and ToxinPred2 server (webs.iiitd.edu.in/raghava/toxinpred2/index.html), respectively. The same server was later used to assess the physiochemical, antigenicity, allergenicity, and toxicity properties of the selection of epitopes as well as for the designed vaccine construct. The solubility nature of the proteins, epitopes, and final vaccine construct was analyzed using the Innovagen solubility check server (https:// innovagen.com/proteomics-tools).
2.11. Prediction of the Secondary and Tertiary Structures of the Designed Pan Parvovirus DNA Multi-Epitopes Based Vaccine Construct
We utilized PDBsum server tool (https://www.ebi.ac.uk/thornton-srv/databases/pdbsum/) to predict and visualize the secondary structure, topology and folds of the construct. And we used Biovia discovery studio to predict and model the tertiary structure. The stability was analyzed and compared through the Ramachandran plot from both Biovia Discovery Studio as well as the PDBSum server tool.
2.12. Molecular Docking and Molecular Dynamic Simulation Anlaysis of the Designed Multi-Epitope Panparvovirus DNA Based Vaccine Construct with the Feline and Canine Toll-Like Receptors (TLRs)
The toll like receptors such as TLR4 (feline, Uniprot ID: ) and TLR5 (canine, Uniprot ID:) were selected to analysis the interaction between the final pan feline-canine vaccine construct and immune receptors. The protein structures were retrieved from Uniprot database, and the structure was visualized through Biovia discovery studio (v22.1.021297). Upon protein preparation, the active binding sites were projected before performing the docking studies, which is crucial for greater binding affinity. We utilized Zdocker docking tool and the detailed analysis of the binding interfaces were characterized by PDBSum server tool. Among the top ten poses, we selected the best pose with higher binding energy to perform Molecular dynamic simulation studies for 50 ns [45].
2.13. Codon Optimization and In-Silico Cloning of the Pan Parvovirus Multi-Epitope DNA Based Vaccine Construct
As previously described, the multiepitope vaccine was optimized and cloned into the expression vector to ensure the potential effective cloning. Hence, the reverse translation of the vaccine protein sequences into a respective DNA sequence was performed using the Vector builder software (https://en.vectorbuilder.com/tool/codon-optimization.html). The codon adaptive index (CAI) value and the GC content of the multi-epitope construct were also calculated as described previously. The restriction enzyme sequences BamHI and EcoRI were added at the DNA’s 3’ and 5’ ends, respectively. Along with this, the Kozak sequence was added to ensure efficient translational initiation in eukaryotic expression systems, which surround the starting codon. The restriction cloning module from Snapgene V.6.0.2 software was used to incorporate the multi-epitope construct into the pET28a(+) plasmids using the indicated restriction enzyme sites.
2.14. In-Silico Immune Simulation of the Designed Pan Parvovirus Multi-Epitope DNA Based Vaccine Construct
The immune response activation was predicted to the designed multiepitope vaccine through in-silico immune simulation, C-ImmSim server (https://150.146.2.1/C-IMMSIM/index.php). The C-ImmSim server simulates the influence of vaccine construct on B and T lymphocytes by modeling the immune system of three major immune organs (bone marrow, thymus, and spleen). The parameters were set as a default with 50 and 1000 simulation steps [46]. We are proposing the administration of the designed multiepitope vaccine construct three times using 4-week intervals. During simulation, each step indicates eight hours of real-time with periods of 1, 84, and 168 hours. Subsequently, this server also predicts the host cellular immune response and cytokine expression levels induced by the multi-epitope vaccine candidates in silico as previously described.
3. Results
3.1. Results of the Prediction of the B Cell Epitopes (Linear and Discontinuous) Within the Major Proteins of Feline and Canine Parvoviruses (NS1, NS2, VP1 and VP2)
The IEDB and BCpred server tool were utilized as same as that of our previously published and compared the results of those two servers keeping the default threshold value of 0.75. The length of the peptides was kept nearly 20mers and the analyzed epitopes (NS1 - 9, NS2 - 4, VP1- 12 and VP2- 9) were subjected to antigenicity prediction. Among those peptides, the top ranked epitopes with high antigenic score were taken into consideration for final vaccine construct (Table 1).

Table 1.
Predicted antigenic linear B cell epitopes across the conserved region of major proteins of feline and canine parvoviruses (NS1, NS2, VP1 and VP2).
Table 1.
Predicted antigenic linear B cell epitopes across the conserved region of major proteins of feline and canine parvoviruses (NS1, NS2, VP1 and VP2).
| No | Start | End | Peptide (IEDB) | Length | Antigen score |
| NS1 | |||||
| 1 | 349 | 361 | ADNTKLTNFDLAN | 13 | 1.3722 |
| 2 | 491 | 506 | TIVRIGCEERPEHTQP | 16 | 0.8492 |
| 3 | 519 | 531 | KLPGDFGLVDKEE | 13 | 1.3432 |
| NS2 | |||||
| 11 | 67 | 80 | MDQTEEEEMDWESE | 14 | 1.1828 |
| VP1 | |||||
| 12 | 5 | 13 | AKRARRGLV | 9 | 1.0456 |
| 13 | 65 | 73 | QRFIDQTKD | 9 | 1.0456 |
| 15 | 228 | 241 | NNMDKTAVNGNMAL | 14 | 0.6846 |
| 16 | 298 | 309 | ESATQPPTKVYN | 12 | 0.4705 |
| 18 | 602 | 606 | NVPPV | 5 | 0.5012 |
| 19 | 617 | 624 | FDTDLKPR | 8 | 1.5012 |
| 20 | 650 | 662 | TNEYDPDASANMS | 13 | 0.7027 |
| 21 | 683 | 689 | RASHTWN | 7 | 0.6420 |
| 22 | 696 | 703 | INVDNQFN | 8 | 1.0518 |
| 23 | 710 | 724 | GGMKIVYEKSQLAPR | 15 | 0.7348 |
| VP2 | |||||
| 24 | 5 | 13 | AKRARRGLV | 9 | 1.0456 |
| 26 | 188 | 199 | FNNQTEFKFLEN | 12 | 0.6846 |
| 30 | 644 | 656 | TNEYDPDASANMS | 13 | 0.7027 |
| 31 | 677 | 698 | RASHTWNPIQQMSINVDNQFNY | 22 | 0.6793 |
| 32 | 707 | 715 | VYEKSQLAP | 9 | 1.1655 |
*Probable antigen Results from the cbtope server to predict the discontinuous epitopes from the fasta sequence of proteins with minimum score of -0.3. The list of the predicted discontinuous B cell epitopes was recognized at different exposed surface areas shown in (Table 2). The position of each predicted epitope on the surface of 3D structure of all the considered proteins could be visualized using Biovia discovery studio visualizer.
Table 2.
Predicted antigenic discontinuous/conformational B cell epitopes across the conserved region of major proteins of feline and canine parvoviruses (NS1, NS2, VP1 and VP2).
Table 2.
Predicted antigenic discontinuous/conformational B cell epitopes across the conserved region of major proteins of feline and canine parvoviruses (NS1, NS2, VP1 and VP2).
| S.No |
Gene Type |
Discontinuous/Conformational B cell epitopes |
| 1 | NS1 | MSGNQYTEEV MEGVNWLKKH AENEAFSFVF KCDNVQLNGK DVRWNNYTKP IQNEELTSLI RGAQTAMDQT EEEEMDWESE VDSLAKKQVQ TFDALIKKCL FEVFVSKNIE PNECVWFIQH EWGKDQGWHC HVLLHSKNLQ QATGKWLRRQ MNMYWSRWLV TLCSVNLTPT EKIKLREIAE DSEWVTILTY RHKQTKKDYV KMVHFGNMIA YYFLTKKKIV HMTKESGYFL STDSGWKFNF MKYQDRQIVS TLYTEQMKPE TVETTVTTAQ ETKRGRIQTK KEVSIKCTLR DLVSKRVTSP EDWMMLQPDS YIEMMAQPGG ENLLKNTLEI CTLTLARTKT AFELILEKAD NTKLTNFDLA NSRTCQIFRM HGWNWIKVCH AIACVLNRQG GKRNTVLFHG PASTGKSIIA QAIAQAVGNV GCYNAANVNF PFNDCTNKNL IWIEEAGNFG QQVNQFKAIC SGQTIRIDQK GKGSKQIEPT PVIMTTNENI TIVRIGCEER PEHTQPIRDR MLNIKLVCKL PGDFGLVDKE EWPLICAWLV KHGYESTMAN YTHHWGKVPE WDEWAEPKI QEGINSPGCK DLETQAASNP QSQDQVLTPL TPDVVDLALE PWSTPDTPIA ETANQQSNQL GVTHKDVQAS PTWSEIEADL RAIFTSEQLE EDFRDDLD |
| 2 | NS2 | MSGNQYTEEV MEGVNWLKKH AENEAFSFVF KCDNVQLNGK DVRWNNYTKP IQNEELTSLI RGAQTAMDQT EEEEMDWESE VDSLAKKLQR LRDTSGKQSS ESRPSSNSSD SGRSGPCTGT VEYSRYAYCR NCKSTIKPTW RYSQRRASES DMVRNRGRPE SHLYF |
| 3 | VP1 | MAPPAKRARR GLVPPGYKYL GPGNSLDQGE PTNPSDAAAK EHDEAYAAYL RSGKNPYLYF SPADQRFIDQ TKDAKDWGGK IGHYFFRAKK AIAPVLTDTP DHPSTSRPTK PTKRSKPPPH IFINLAKKKK AGAGQVKRDN LAPMSDGAVQ PDGGQPAVRN ERATGSGNGS GGGGGGGSGG VGISTGTFNN QTEFKFLENG WVEITANSSR LVHLNMPESE NYRVVVNNMD KTVNGNMALD DIHQIVTPWS LVDANAWGVW FNPGDWQLIV NTMSELHLVS FEQEIFNVVL KTVSESATQP PTKVYNNDLT ASLMVALDSN NTMPFTPAAM RSETLGFYPW KPTIPTPWRY YFQWDRTLIP SHTGTSGTPT NYHGTDPDDV QFYTIENSVP VHLLRTGDEF ATGTFFFDCK PCRLTHTWQT NRALGLPPFL NSLPQSEGAT NFGDIGVQQD KRRGVTQMGN TYITEATIMR PAEVGYSAPY YSFEASTQGP FKTPIAAGRG GAQTDENQAA DGPRYAFGRQ HGQKTTTTGE TPERFTYIAH QDTGRYPEGD WIQNINFNLP VTNDNVLLPT DPIGGKTGIN YTNIFNTYGP LTALNNVPPV YPNGQIWDKE FDTDLKPRLH VNAPFVCQNN CPGQLFVKVA PNLTNEYDPD ASANMSRIVT YSDFWWKGKL VFKAKLRASH TWNPIQQMSI NVDNQFNYVP NIGMKIVYEK SQLAPRKLY |
| 4 | VP2 | MSDGAVQPDG GQPAVRNERA TGSGNGSGGG GGGGSGGVGI STGTFNNQTE FKFLENGWVE ITANSSRLVH LNMPESENYR RVVVNNLDKT AVKGNMALDD IHAQIVTPWS LVDANAWGVW FNPGDWQLIV NTMSELHLVS FEQEIFNVVL KTVSESATQP PTKVYNNDLT ASLMVALDSN NTMPFTPAAM RSETLGFYPW KPTIPTPWRY YFQWDRTLIP SHTGTSGTPT NIYHGTDPDD VQFYTIENSV PVHLLRTGDE FATGTFFFDC KPCRLTHTWQ TNRALGLPPF LNSLPQAEGG TNFGYIGVQQ DKRRGVTQMG NTNYITEATI MRPAEVGYSA PYYSFEASTQ GPFKTPIAAG RGGAQTDENQ AADGDPRYAF GRQHGQKTTT TGETPERFTY IAHQDTGRYP EGDWIQNINF NLPVTNDNVL LPTDPIGGKA GINYTNIFNT YGPLTALNNV PPVYPNGQIW DKEFDTDLKP RLHVNAPFVC QNNCPGQLFV KVAPNLTNEY DPDASANMSR IVTYSDFWWK GKLVFKAKLR ASHTWNPIQQ MSINVDNQFN YLPNNIGAMK IVYEKSQLAP RKLY |
3.2. Results of the Prediction of the Cytotoxic T Lymphocyte Epitopes (MHC Class-I molecules) Within the Major Proteins of the Feline and Canine Parvoviruses (NS1, NS2, VP1 and VP2)
As previously stated in materials and methods, MHC class I molecules and its associate cytokine induction (IFN) were analyzed using IEDB and IFN server tool. Table 3 shows the predicted MHC class I epitopes with the binding affinity (IC50; IC50 < 50 nM) and percentile rank. Table 3 also shows the parameters of the top-ranked epitopes, taking into consideration the allergenicity, antigenicity, non-toxic, and solubility per each listed epitope. Table 4 shows the predicted IFN epitopes using SVM method, the epitopes which shows the positive value with their desired score were considered for final vaccine construct.
3.4. Results of the Prediction of the Helper T Lymphocyte Epitopes Prediction Within the Major Proteins of the Feline and Canine Parvovirus (NS1, NS2, VP1 and VP2)
The prediction of MHC class II molecules and the cytokine inducing epitopes (IL-4) were analyzed and the values are tabulated. And the epitopes were filtered based on their physicochemical properties (Table 5) and subjected to cytokines induction based on SVM method (Table 6). The values which showed the positive were a final candidate to be used in vaccine construct.
3.5. Results of the Molecular Docking of the Selected MHC Classes (I and II) Epitopes with the Within Major Proteins of the Feline and Canine Parvovirus (NS1, NS2, VP1 and VP2)
The interaction analysis between the Feline MHC I and MHC II antigen (DRB- (Uniprot ID : Q95481 – MHC class I and Uniport ID: Q19430 – MHC class II)) and the selected epitopes were studied using HADdock server tool using peptide-binding groove affinity (Figure 1 and Figure 2). Initially, we utilized PEPFOLD3 server to design the structure of epitopes/peptides and import pdb files into HADdock server. The feline DRB files were used as receptors and the MHC class I and MHC class II peptides listed in Table 3 were considered as ligands. Results show the binding affinity and confidence score as listed in Table 7 and the top-ranked epitopes showing the highest binding affinity score were chosen for the design of the final vaccine construct.
3.6. Structure and Design of the Pan-Multiepitope Vaccine Against the Major Proteins of the Feline and Canine Parvovirus (NS1, NS2, VP1 and VP2)
The final vaccine were designed by combining the top-ranked B-cell epitopes T- cell epitopes of both MHC I and MHC II class of molecules, filtered from high antigenic, non-allergic, non-toxic and good solubility, and with the better binding affinity score of the structural proteins (NS1, NS2, VP1 and VP2) of feline and canine parvovirus as listed in the Table 5. The N-terminus were linked with adjuvants, PADRE using GGS and EAAAK, where C-terminus were with 6X H-tag (HHHHHH) and stop codon using EAAAK. Linkers such as KK, AAY, GPGPG were used to connect top ranked B cell and T cell epitopes (Figure 3).
3.7. Results of the Physiochemical Properties of the Designed Multiepitope Vaccine Against the Feline and Canine Parvoviruses (NS1, NS2, VP1 and VP2)
The physiochemical properties such as molecular weight, Theoretical pI, number of amino acids, formation of Cys residues, instability index and nature of solubility of constructed vaccine were analyzed using Protparam server tool. The results showed that vaccine weighed 43165.02 Dalton and theoretical isoelectric point of 9.71, indicating the alkaline nature. The number of positively charged residues (Arg + Lys) were 62 and the number of negatively charged residues (Arg + Glu) were 41 and the extinction coefficient measured in water at 280 nm was shown to be 0.706. The instability index (II) was about 10.30, showing the structure of the vaccine protein was stable. The aliphatic index was about 69.19, indicating the hydrophilic nature with a value of -0.736.
3.8. Results of the Secondary and Tertiary Structures of the Designed Multiepitope Pan Parvovirus DNA Vaccine Construct
The secondary and tertiary structure of the multiepitope-based vaccine construct were analyzed and modeled through the Biovia discovery studio and Psipred server tool (Figure 4a and Figure 4b). We utilized Alpha fold prediction model to predict the structural features with 3D XYZ coordinates plot (Figure 4c). The coordinate plot shows a well-organized and compact spatial distribution of C-alpha atoms, including the epitopes, linkers and adjuvant elements fold into a stable conformation. The stability of the constructed vaccine was analyzed by Ramachandran plot as previously described (Figure 4d).
3.9. Results of the Disulphide Bond Engineering
The analysis from Disulfide by Designv2.12 webserver showed that the six residue pairs such as ASN-SER, LEU-ALA, PRO-GLY, PRO-TYR, GLU-SER and PRO-ASP were observed based on chi3 and B-factor energy parameters.
In each scenario, the formation of disulfide bonds was initiated through the introduction of cysteine residues, and the evaluation was done within the range of −87 to +97 chi3 values and ensuring an energy value of < 2.2. These steps were employed to identify the suitable residues for the formation of disulfide bond, enhancing the stability of the vaccine construct (Figure 5).
3.10. Results of the Assessment of the Stability of the Designed Vaccine Construct Using the Normal Mode Analysis and Prediction
The stability and physical movement of the vaccine complex were analyzed with Normal mode analysis using iMODS server tool. Figure showed the B factor (Figure 6a) represents the atomic fluctuations in normal mode (NMA) comparing them with experimental mode of vibration (PDB), the values less than 50 shows stable proteins where more than that show flexible proteins. The variance plot showed the restricted motion through
Purple and green color reflect the cumulative variance (Figure 6b). The eigen value of the complex showed the values of about 1.904111×10−7 as shown in the (Figure 6c) and the deformability plot (Figure 6d) showed the capability of bending during the motion. Figure 6e shows the covariance map showing the motion correlations between residues, where red indicates the correlated movements, blue indicates the anti-correlation and white shows no movements between them. Lastly, the elastic network showed the stable region with dense spring and flexible region with sparse ones (Figure 6f).
3.11. Results of the Molecular Docking of the Designed Vaccine Construct with the Feline and Canine Toll-Like Receptors (TLR4 and TLR5)
The exploration of the interaction between vaccine construct and toll like immunoreceptor of both feline and canine, to be as the better pan vaccine candidate for preventing the parvovirus among the species is much important. Hence, the molecular docking analysis were performed the vaccine construct and Toll-like receptors (TLR4 – feline and TLR5 – canine). Initially, the sequence of TLR4 (Uniprot ID:AF-P58727) and TLR5 (Uniprot ID: A0A1P8NLQ2) were retrieved from the database and modeled through Biovia discovery studio, followed by preparing the protein for the docking study by removing water molecules, adding hydrogen and performing energy minimization. Docking analysis was performed using the Zdocker, and the results obtained indicate the strong binding affinity between the vaccine construct and Toll-like receptors (TLR4 and TLR5). The best-ranked complexes with their respective ZDock scores provide us with confirmation of the strong and stable interaction between them, Figure 7. The interaction residues, multiple hydrogen bonds, and hydrophobic bonds were analyzed through the PDBsum server tool. Figure 7(a-b) shows the topology visualization of TLR4 and TLR5, shows the docking interaction analysis between the vaccine construct and Toll-like receptors, and finally, Supp-Figure 1(c-d) illustrates the interaction amino acid residues and formation of multiple hydrogen bonds, hydrophobic bonds, etc., through PDBsum server tool.
3.12. In Silico Cloning of the Pan Based Multi Epitope Vaccine Spanning Key Epitopes Within the Major Proteins (NS1, NS2, VP1 and VP2) Proteins
The vaccine construct was cloned using the Vector builder from the decoded amino acid sequence of each epitope’s respective DNA sequences to mimic the vaccine’s expression in the E.coli K12 expression system. The GC content and codon adaptation index values generated by the vector builder server represent the level of expression in the E.coli system. Finally, Snap gene software was used to clone the constructed vaccines in the pET-28a (+) expression vector between the restriction enzyme cutting locations of BamHI and EcoRI, and the results obtained are shown in Supp.Figure 2(a-b).
3.13. In Silico Immune Simulation of the Designed Pan Based Multi Epitope Vaccine Spanning Key Epitopes Within the Major Proteins (NS1, NS2, VP1 and VP2) Proteins
The predicted immune response of the constructed vaccine was analyzed through the interaction between the antigens and B cells, T cells, and cytokines (Supp.Figure 3 a-s) utilizing C-ImmSim immune simulation methods. The primary and secondary humoral response were shown by significant increase in IgM and IgG. Similar from our IFN-γ and IL-4 database prediction, the marked expansion and robust cellular immune response were evaluated with this immune simulation as well.
4. Discussion
The canine and feline parvovirus has become one of the most dreaded viral infections caused in dogs and cats, though it is uncommon now (2026) due to larger part of vaccine availability, it’s been a dangerous infectious disease in early 2010’s. This infection is characterized by marked leukopenia, high fever and hemorrhagic diarrhea often resulting in severe dehydration, secondary infections and mortality rates exceeding 80% in susceptible animals [1]. The dogs or cats under two years of age with acute bloody diarrhea should be considered at high risk for parvoviruses, particularly if the vaccine history is incomplete. Since both feline panleukopenia virus (FPV) and its close corollary canine parvovirus (CPV) are antigenically stable i.e., they won’t change rapidly as other virus does, the existing vaccines remain very effective. But the animals in shelters across the country are still losing their lives daily to these diseases [47,48]. Greater than 90% of dogs with CPV-2 enteritis will survive if administered supportive care shortly after clinical signs develop. Feline panleukopenia often has a higher fatality rate. Hence, there is an urgent need to develop some effective vaccines that could protect both the species against those dreadful viruses. As described in our previously published articles, the incorporation of AI into the drug development process has vastly improved. The multi-epitope based vaccine provides not only the long-term protection through trained immune system but also safer and cost effective and their limits is due to its extensive validation requirements [49,50].
The project mainly focuses on utilizing AI integrated tools to develop a pan multiepitope DNA-based vaccine against both feline and canine parvovirus. Initially, it begins with (i) retrieving the desired sequence of feline and canine sub structural proteins (NS1, NS2, VP1 and VP2) from RCSB database and performed MSA analysis for the conserved region. (ii) The B-cell and MHC class I epitopes were predicted from IEDB database and MHC class II epitopes were predicted with NetMHCIIpan-4.3, using the experimentally predicted feline alleles (DRB1_0103, DRB1_0102 and DRB1_0107) in comparison with canine and human alleles (percentile score (<4) for MHC class I molecules and (<10) for MHC class II molecules with the IC50 value of (<50nM)). Followed by the (iii) analysis of immunogenetic characteristic features such as antigenic, non-allergic, non-toxic and better solubility was performed (VaxiJen 2.0 and AllerTop). (iv) The top ranking epitopes were docked with feline MHC class I and II alleles to analysis their binding interaction between them (HADdock docking server). (v) The final vaccine was constructed with top ranked epitopes through linkers and adjuvants, and the physicochemical properties of the designed vaccine were studied. (vi) The molecular docking was performed between vaccine construct and TLRs (TLR4 and TLR5) of feline and canine toll like receptors to study the binding interaction between them, followed by molecular dynamic simulation was done for 100 ns to check their long-term astability (Biovia discovery studio). (vii) immunosimulation of the final vaccine construct to assess its potential potency in the activation of the humoral and cell-mediated immunity of both feline and canine species (Immsim).
The major challenges in this study were during the prediction of MHC class II epitopes; there is the lack of data about the feline alleles in the IEDB database server. This has been rectified through alternative strategies by using the experimentally predicted alleles align with human and canine allele in identifying the epitopes with NetpanMHCII class server tool [49,50,51]. The parameters were default with the polymerase length of 12mer. This approach successfully provided the best score data and matched the feline alleles with the corresponding epitopes.
The interferons (IFNs) and Interleukins (IL-4) have the most important part in triggering the immune response in both MHC class I and MHC class II molecules of T-cell expression. The filtered epitopes of MHC class I molecules were subjected to interferon SVM method of prediction, and the epitopes such as FVFKCDNVQL (0.45722211), KVAPNLTNEY (0.43379691), RGLVPPGYKY (0.45733135) and KRSKPPPHIF (0.44701719) showed the positive values. Like, the filtered epitopes of MHC class II molecules were subjected to interleukin SVM method of prediction, and the epitopes such as FMKYQDRQI (1.24), IACVLNRQG (0.29), WVTILTYRH (0.28), YSQRRASES (0.30), WGGKIGHYF (0.28) and LDDIHAQIV (0.24) resulted in positive values. Those epitopes were utilized in final vaccine construct and confirmed their binding interaction with Feline MHC class I and MHC class II molecules through molecular docking studies using HADdock server.
The top-ranked peptide was selected based on their binding score and high antigenic score for all structural genomes we considered for the study, for MHC class I of molecules – FVFKCDNVQL (0.7526 and -193.10 docking score – NS1), KVAPNLTNEY (0.8912 and -157.61 docking score – NS2), RGLVPPGYKY (0.5816 and -214.32 docking score – VP1), KRSKPPPHIF (1.1015 and -201.88 docking score – VP2) and for MHC class II of molecules – FMKYQDRQI (0.8792 and docking score of -190.38- NS1), YSQRRASES - (0.5799 and docking score of -227.71 – NS2), WGGKIGHYF -(1.0920 and docking score of -222.16 – VP1) and LDDIHAQIV – (0.9059 and docking score of -168.88 – VP2). Finally, these epitopes were used in the vaccine construct and were designed using linkers and adjuvants. The interaction residues between them were identified through Tamarind bio (PDBsum) and were displayed in the figure, which results in multiple hydrogen bonds and hydrophobic bonds, especially to capture their better binding interactions.
The disulfide bond analysis was done to predict the presence of cysteine residues, which contains a reactive thiol (-SH) group in its side chain, and once incorporated into a protein, this thiol would participate in important biochemical functions. Normally, the disulfide bonds form due to the oxidation reaction happens between two cysteine residues, which makes the vaccine structure to be more stabilized. The conformational classification χ₃ dihedral angle (C–S–S–C), describes the rotation around S-S bond and determines the spatial orientations of the two cysteines linked by it [52]. When the angle is approximately ±90°, the bonds show the right- or left-handed spiral conformations, and when it is close to 180°, they correspond to trans conformation, shows the extended and linear arrangement of cysteines. And finally, the angle close to 0°, it shows the cis conformation, due to steric clashes and unfavorable geometry that make it energetically unstable. Since all the resultant values were nearly 102° to 113°, the protein structures making the right- or left-hand conformations with their corresponding energy values.
Next, further validation on the stability and flexibility of the vaccine construct were done with Normal mode analysis (NMA). The mobility plot provides the residues with higher peaks (0.5 – 2 arbitrary units) shows the flexible loop or linker region, while the lower values (0-0.3) give the stable structural region. The regions which bend during the motion were shown in deformability plot, falls within the range of 0.05 – 0.15. Next, the atomic fluctuations were seen with B-factor plot, where the stable proteins occur in the range below 50 and above it shows the flexible proteins. Normally, the eigen values falling within 1×10⁻⁶ to 1×10⁻⁴, represents the stiffness of the protein, since the obtained values were higher than the standard range, means the vaccine construct has greater structural rigidity and must need higher energy needed for deformation. The individual (purple) and cumulative (green) variances were observed through variance plot , where lower individual variance values (<0.05) represent restricted motion of each mode and the accumulation of modes were represented with rise in cumulative curve. Finally, the elastic network model gives the stable region with dense springs and flexible areas with sparse ones.
The molecular docking analysis was performed with Biovia Discovery Studio using Zdocker, it showed the better binding interaction between the designed vaccine epitopes and the TLRs (TLR4 – feline and TLR5 – canine), facilitating effective immune recognition and the initiation of the robust immune response. The binding affinity was calculated based on ZDock score and ZRank score for both feline and canine and they are 22.26 (Zdock score), -47.409 (Zrank score) for feline and 16.54 (Zdock score), -134.295 (Zrank score) for canine. The PDBsum results were analyzed and the interaction between the residues possess hydrogen bonds (Arg 578-His 384), (Gly 617 – Lys 383), (Asp 596 – Ser 120), (Ser 589 – Ser 124), (Lys 632 – Glu 131) for feline TLR4 and (Cys 24 – Arg 53) for canine TLR5 with the vaccine construct.
Next, the vaccine construct was cloned using the optimized codons with vector builders to express in the E.coli K12 system by converting the amino acid sequence into a DNA sequence. Through Snap gene, the insertion of vaccine was done into the pET-28a(+) vector between BamHI and EcoRI sites confirmed proper insertion and orientation of the gene, results the vaccine construct can be efficiently expressed in E.coli for production of recombinant protein [53,54]. Finally, in silico immune simulations using C-ImmSim provided critical insights about the potential immune responses elicited by the designed four structural proteins (NS1, NS2, VP1 and VP2) of the pan multi-epitope based (canine and feline) vaccine constructs [55,56,57]. The simulation results revealed robust activation of T-cell populations, including cytotoxic T cells and helper T cells, crucial for cellular and humoral immunity. As described in our previously published article, this analysis would tell us the vaccine construct in this study would induce strong humoral and cell-mediated immunity [58].
From these results, we would like to convey that our constructed pan parvovirus multi-epitope-based vaccine combining the four major structural proteins (NS1, NS2, VP1 and VP2) will be effective in the protection of both canine and feline species against the parvovirus. Though computational analysis will support us to predict the more precise top-ranking epitopes and their immuno-antigenic properties, further experimental validation is required to be used against those viruses.
5. Conclusions
We successfully designed a pan-multi-epitope-based vaccine compiling the top ranked immunogenic and antigenic epitopes. The interferon and interleukin will enhance/trigger immuno-response of those MHC class I and MHC class II of molecules. The T cell epitopes showed high binding affinities with the feline alleles. The disulfide bond prediction and Normal mode analysis showed our vaccine construct are stable and flexible. The molecular docking and dynamic simulation analysis showed higher binding affinities and stable conformation with canine (TLR5) and feline (TLR4) toll like receptors. The designed vaccine construct showed high immunogenic potential in terms of the production of humoral and cell-mediated immunity in feline and canine using an immune simulation approach. We believe the designed vaccine in the current study will protect both the feline and canine species against this dreadful parvovirus.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
ND: Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Writing – original draft, Writing – review & editing. MK: Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Writing – original draft, Writing – review & editing. AS: Data curation, Investigation, Methodology, Software, Validation, Writing – original draft, Writing – review & editing, Resources. MC: Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing, Conceptualization, Funding acquisition. MH: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
This study was funded by a seed grant (PI: MGH) from Long Island University (Grant no: 40262) and funds from the USDA-NIFA Animal Health and Disease Research grant (NI24AHDRXXXXG066).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors upon request.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The author(s) declared that they were editorial board members of Scientific report, at the time of submission. This had no impact on the peer review process and the final decision.
References
- Truyen, U.; Parrish, C.R. Canine and feline host ranges of canine parvovirus and feline panleukopenia virus: distinct host cell tropisms of each virus in vitro and in vivo. J Virol 1992, 66, 5399-5408. [CrossRef]
- Parrish, C.R.; O’Connell, P.H.; Evermann, J.F.; Carmichael, L.E. Natural variation of canine parvovirus. Science 1985, 230, 1046-1048. [CrossRef]
- Parrish, C.R.; Holmes, E.C.; Morens, D.M.; Park, E.C.; Burke, D.S.; Calisher, C.H.; Laughlin, C.A.; Saif, L.J.; Daszak, P. Cross-species virus transmission and the emergence of new epidemic diseases. Microbiol Mol Biol Rev 2008, 72, 457-470. [CrossRef]
- Parrish, C.R.; Kawaoka, Y. The origins of new pandemic viruses: the acquisition of new host ranges by canine parvovirus and influenza A viruses. Annu Rev Microbiol 2005, 59, 553-586. [CrossRef]
- Sun, Y.; Zhang, C.; Liu, J.; Guo, D.; Li, S.; Xu, L.; Hu, B.; Zeng, H.; Liu, Y.; Fu, J.; et al. Development of a potent protective felinized chimeric antibody C8A8 targeting feline and canine parvovirus. Vet Microbiol 2026, 312, 110817. [CrossRef]
- Tang, A.; Li, B.; Zhu, M.; Zhu, S.; Zhang, D.; Li, N.; Zhang, M.; Zhu, Y.; Li, C.; Meng, C.; et al. A novel feline herpesvirus vector subunit FCV VP1 and FPV VP2 vaccine protects cats against FHV-1 and FPV challenge and induces serum neutralizing antibody responses against FCV. Front Immunol 2025, 16, 1636514. [CrossRef]
- Al-Mubarak, A.I.A.; Al-Kubati, A.A.G.; Sheikh, A.; Abdelaziz, A.M.; Hussen, J.; Kandeel, M.; Falemban, B.; Hemida, M.G. Detection of Avian Orthoavulavirus-1 genotypes VI.2.1 and VII.1.1 with neuro-viscerotropic tropism in some backyard pigeons (Columbidae) in Eastern Saudi Arabia. Front Vet Sci 2024, 11, 1352636. [CrossRef]
- Al-Mubarak, A.I.A.; Al-Kubati, A.A.G.; Skeikh, A.; Hussen, J.; Kandeel, M.; Flemban, B.; Hemida, M.G. A longitudinal study of bovine viral diarrhea virus in a semi-closed management dairy cattle herd, 2020-2022. Front Vet Sci 2023, 10, 1221883. [CrossRef]
- Al-Mubarak, A.I.A.; Hussen, J.; Kandeel, M.; Al-Kubati, A.A.G.; Falemban, B.; Hemida, M.G. Avian encephalomyelitis virus in backyard chickens. Vet World 2023, 16, 1866-1870. [CrossRef]
- Alsultan, M.A.; Alhammadi, M.A.; Hemida, M.G. Infectious bronchitis virus from chickens in Al-Hasa, Saudi Arabia 2015-2016. Vet World 2019, 12, 424-433. [CrossRef]
- Ba Abduallah, M.M.; Hemida, M.G. Comparative analysis of the genome structure and organization of the Middle East respiratory syndrome coronavirus (MERS-CoV) 2012 to 2019 revealing evidence for virus strain barcoding, zoonotic transmission, and selection pressure. Rev Med Virol 2021, 31, 1-12. [CrossRef]
- Chu, D.K.W.; Hui, K.P.Y.; Perera, R.; Miguel, E.; Niemeyer, D.; Zhao, J.; Channappanavar, R.; Dudas, G.; Oladipo, J.O.; Traore, A.; et al. MERS coronaviruses from camels in Africa exhibit region-dependent genetic diversity. Proc Natl Acad Sci U S A 2018, 115, 3144-3149. [CrossRef]
- Gomaa, M.H.; Barta, J.R.; Ojkic, D.; Yoo, D. Complete genomic sequence of turkey coronavirus. Virus Res 2008, 135, 237-246. [CrossRef]
- Hemida, M.G.; Al-Hammadi, M.A.; Daleb, A.H.S.; Gonsalves, C.R. Molecular characterization and phylogenetic analyses of virulent infectious bronchitis viruses isolated from chickens in Eastern Saudi Arabia. Virusdisease 2017, 28, 189-199. [CrossRef]
- Hemida, M.G.; Ali, M.; Alhammadi, M.; Alnaeem, A. The Middle East respiratory syndrome coronavirus in the breath of some infected dromedary camels (Camelus dromedarius). Epidemiol Infect 2020, 148, e247. [CrossRef]
- Hemida, M.G.; Alnaeem, A.A. Betaretrovirus infections in dromedary camels in Saudi Arabia. Vet Med Sci 2022, 8, 1330-1336. [CrossRef]
- Hemida, M.G.; Chu, D.K.; Poon, L.L.; Perera, R.A.; Alhammadi, M.A.; Ng, H.Y.; Siu, L.Y.; Guan, Y.; Alnaeem, A.; Peiris, M. MERS coronavirus in dromedary camel herd, Saudi Arabia. Emerg Infect Dis 2014, 20, 1231-1234. [CrossRef]
- Hemida, M.G.; Chu, D.K.W.; Chor, Y.Y.; Cheng, S.M.S.; Poon, L.L.M.; Alnaeem, A.; Peiris, M. Phylogenetic Analysis of MERS-CoV in a Camel Abattoir, Saudi Arabia, 2016-2018. Emerg Infect Dis 2020, 26, 3089-3091. [CrossRef]
- Hemida, M.G.; Perera, R.; Chu, D.K.W.; Alnaeem, A.A.; Peiris, M. Evidence of equine influenza A (H3N8) activity in horses from Eastern and Central Saudi Arabia: 2013-2015. Equine Vet J 2019, 51, 218-221. [CrossRef]
- Malik, Y.S.; Kumar, P.; Ansari, M.I.; Hemida, M.G.; El Zowalaty, M.E.; Abdel-Moneim, A.S.; Ganesh, B.; Salajegheh, S.; Natesan, S.; Sircar, S.; et al. SARS-CoV-2 Spike Protein Extrapolation for COVID Diagnosis and Vaccine Development. Front Mol Biosci 2021, 8, 607886. [CrossRef]
- Shah, A.U.; Esparza, B.; Illanes, O.; Hemida, M.G. Comparative Genome Sequencing Analysis of Some Novel Feline Infectious Peritonitis Viruses Isolated from Some Feral Cats in Long Island. Viruses 2025, 17. [CrossRef]
- Shah, A.U.; Gauger, P.; Hemida, M.G. Isolation and molecular characterization of an enteric isolate of the genotype-Ia bovine coronavirus with notable mutations in the receptor binding domain of the spike glycoprotein. Virology 2025, 603, 110313. [CrossRef]
- Duraisamy, N.; Khan, M.Y.; Shah, A.U.; Elalaoui, R.N.; Cherkaoui, M.; Hemida, M.G. Machine learning tools used for mapping some immunogenic epitopes within the major structural proteins of the bovine coronavirus (BCoV) and for the in silico design of the multiepitope-based vaccines. Front Vet Sci 2024, 11, 1468890. [CrossRef]
- Goud, M.D.; Ramos, E.; Shah, A.U.; Hemida, M.G. Artificial Intelligence Driven Framework for the Design and Development of Next-Generation Avian Viral Vaccines. Microorganisms 2025, 13. [CrossRef]
- Khan, M.Y.; Shah, A.U.; Duraisamy, N.; Cherkaoui, M.; Hemida, M.G. Repurposing of Some Nucleoside Analogs Targeting Some Key Proteins of the Avian H5N1 Clade 2.3.4.4b to Combat the Circulating HPAI in Birds: An In Silico Approach. Viruses 2025, 17. [CrossRef]
- Khan, M.Y.; Shah, A.U.; Duraisamy, N.; ElAlaoui, R.N.; Cherkaoui, M.; Hemida, M.G. Leveraging Artificial Intelligence and Gene Expression Analysis to Identify Some Potential Bovine Coronavirus (BCoV) Receptors and Host Cell Enzymes Potentilly Involved in the Viral Replication and Tissue Tropism. Int J Mol Sci 2025, 26. [CrossRef]
- Khan, M.Y.; Shah, A.U.; Duraisamy, N.; Moawad, N.; ElAlaoui, R.N.; Cherkaoui, M.; Hemida, M.G. Identification of potential inhibitors of the main protease from feline infectious peritonitis virus using molecular docking and dynamic simulation approaches. PeerJ 2025, 13, e19744. [CrossRef]
- Michalka, T.; Shah, A.U.; Liang, T.; Hemida, M.G. Multi-Epitope DNA-Based Feline Immunodeficiency Virus Vaccine Construct Designed by Immunoinformatic and Machine Learning Tools as a Surrogate Model for HIV Vaccine Development. Pathogens 2026, 15. [CrossRef]
- Patterson, E.V.; Reese, M.J.; Tucker, S.J.; Dubovi, E.J.; Crawford, P.C.; Levy, J.K. Effect of vaccination on parvovirus antigen testing in kittens. J Am Vet Med Assoc 2007, 230, 359-363. [CrossRef]
- Wang, T.; Wu, H.; Wang, Y.; Guan, Y.; Cao, Y.; Wang, L.; Wang, M.; Tan, F.; Pang, W.; Tian, K. Virus-like Particle Vaccine for Feline Panleukopenia: Immunogenicity and Protective Efficacy in Cats. Vaccines (Basel) 2025, 13. [CrossRef]
- Mortazavi, B.; Molaei, A.; Fard, N.A. Multi-epitopevaccines, from design to expression; an in silico approach. Hum Immunol 2024, 85, 110804. [CrossRef]
- Shawan, M.; Sharma, A.R.; Halder, S.K.; Arian, T.A.; Shuvo, M.N.; Sarker, S.R.; Hasan, M.A. Advances in Computational and Bioinformatics Tools and Databases for Designing and Developing a Multi-Epitope-Based Peptide Vaccine. Int J Pept Res Ther 2023, 29, 60. [CrossRef]
- Basmenj, E.R.; Pajhouh, S.R.; Ebrahimi Fallah, A.; Naijian, R.; Rahimi, E.; Atighy, H.; Ghiabi, S.; Ghiabi, S. Computational epitope-based vaccine design with bioinformatics approach; a review. Heliyon 2025, 11, e41714. [CrossRef]
- Duraisamy, N.; Shah, A.U.; Khan, M.Y.; Cherkaoui, M.; Hemida, M.G. A Pan-H5N1 Multiepitope DNA Vaccine Construct Targeting Some Key Proteins of the Clade 2.3.4.4b Using AI-Assisted Epitope Mapping and Molecular Docking. Viruses 2025, 17. [CrossRef]
- Chauhan, V.; Singh, M.P. Immuno-informatics approach to design a multi-epitope vaccine to combat cytomegalovirus infection. Eur J Pharm Sci 2020, 147, 105279. [CrossRef]
- Ansari, H.R.; Raghava, G.P. Identification of conformational B-cell Epitopes in an antigen from its primary sequence. Immunome Res 2010, 6, 6. [CrossRef]
- Karosiene, E.; Lundegaard, C.; Lund, O.; Nielsen, M. NetMHCcons: a consensus method for the major histocompatibility complex class I predictions. Immunogenetics 2012, 64, 177-186. [CrossRef]
- Jain, P.; Joshi, A.; Akhtar, N.; Krishnan, S.; Kaushik, V. An immunoinformatics study: designing multivalent T-cell epitope vaccine against canine circovirus. J Genet Eng Biotechnol 2021, 19, 121. [CrossRef]
- Yuhki, N.; Beck, T.; Stephens, R.M.; Nishigaki, Y.; Newmann, K.; O’Brien, S.J. Comparative genome organization of human, murine, and feline MHC class II region. Genome Res 2003, 13, 1169-1179. [CrossRef]
- Dhanda, S.K.; Vir, P.; Raghava, G.P. Designing of interferon-gamma inducing MHC class-II binders. Biol Direct 2013, 8, 30. [CrossRef]
- Dhanda, S.K.; Gupta, S.; Vir, P.; Raghava, G.P. Prediction of IL4 inducing peptides. Clin Dev Immunol 2013, 2013, 263952. [CrossRef]
- Yan, Y.; Tao, H.; He, J.; Huang, S.Y. The HDOCK server for integrated protein-protein docking. Nat Protoc 2020, 15, 1829-1852. [CrossRef]
- Soltan, M.A.; Elbassiouny, N.; Gamal, H.; Elkaeed, E.B.; Eid, R.A.; Eldeen, M.A.; Al-Karmalawy, A.A. In Silico Prediction of a Multitope Vaccine against Moraxella catarrhalis: Reverse Vaccinology and Immunoinformatics. Vaccines (Basel) 2021, 9. [CrossRef]
- Yousaf, M.; Ismail, S.; Ullah, A.; Bibi, S. Immuno-informatics profiling of monkeypox virus cell surface binding protein for designing a next generation multi-valent peptide-based vaccine. Front Immunol 2022, 13, 1035924. [CrossRef]
- Lopes, T.S.; Gheno, B.P.; Miranda, L.D.S.; Detofano, J.; Khan, M.A.A.; Streck, A.F. In silico designing of multi-epitope vaccine against canine parvovirus using reverse vaccinology. Braz J Microbiol 2024, 55, 2953-2968. [CrossRef]
- Pang, M.; Tu, T.; Wang, Y.; Zhang, P.; Ren, M.; Yao, X.; Luo, Y.; Yang, Z. Design of a multi-epitope vaccine against Haemophilus parasuis based on pan-genome and immunoinformatics approaches. Front Vet Sci 2022, 9, 1053198. [CrossRef]
- Haseeb, A.; Yousaf, W.; Cao, Z.; Fan, K.; Sun, N.; Sun, P.; Sun, Y.; Yang, H.; Yin, W.; Zhang, H.; et al. Parvoviruses NS1 oncolytic attributes: mechanistic insights and synergistic anti-tumor therapeutic strategies. Front Microbiol 2025, 16, 1631433. [CrossRef]
- Wen, Y.; Tang, Z.; Wang, K.; Geng, Z.; Yang, S.; Guo, J.; Chen, Y.; Wang, J.; Fan, Z.; Chen, P.; et al. Epidemiological and Molecular Investigation of Feline Panleukopenia Virus Infection in China. Viruses 2024, 16. [CrossRef]
- Negahdaripour, M.; Nezafat, N.; Eslami, M.; Ghoshoon, M.B.; Shoolian, E.; Najafipour, S.; Morowvat, M.H.; Dehshahri, A.; Erfani, N.; Ghasemi, Y. Structural vaccinology considerations for in silico designing of a multi-epitope vaccine. Infect Genet Evol 2018, 58, 96-109. [CrossRef]
- Rahman, S.; Huang, D.-W.; Shah, M.; Almutairi, M.M.; Liaqat, I.; Tanaka, T.; Chen, C.-C.; Bahadar, S.; Ali, A. Vaccinomics-based identification of immunodominant epitopes in the tick-borne encephalitis virus polyprotein for multi-epitope vaccine development. Computational and Structural Biotechnology Reports 2025, 2, 100047. [CrossRef]
- Kuwahara, Y.; Kitoh, K.; Kobayashi, R.; Iwata, J.; Ohne, R.; Hosokawa-Kanai, T.; Matsumoto, Y.; Kitagawa, H.; Sasaki, Y. Genotyping of feline MHC (FLA) class II DRB by PCR-RFLP method using group-specific primers. J Vet Med Sci 2000, 62, 1283-1289. [CrossRef]
- Paul, B.; Alam, J.; Hossain, M.M.K.; Hoque, S.F.; Bappy, M.N.I.; Akter, H.; Ahmed, N.; Akter, M.; Ali Zinnah, M.; Das, S.; et al. Immunoinformatics for Novel Multi-Epitope Vaccine Development in Canine Parvovirus Infections. Biomedicines 2023, 11. [CrossRef]
- Deepthi, V.; Sasikumar, A.; Mohanakumar, K.P.; Rajamma, U. Computationally designed multi-epitope vaccine construct targeting the SARS-CoV-2 spike protein elicits robust immune responses in silico. Sci Rep 2025, 15, 9562. [CrossRef]
- Naveed, M.; Husnain, M.; Aziz, T.; Qadir, P.; Asim, M.; Majeed, M.N.; Aloufi, A.S.; Shami, A.; Alwethynani, M.S.; Tombozara, N. Immunoinformatics-based design and evaluation of a multi-epitope vaccine against Vibrio fluvialis. Sci Rep 2026, 16, 4100. [CrossRef]
- Ishwar, D.; Padavu, S.; Kumar, M.; Gollapalli, P.; Ballamoole, K.K.; Kumar, A.; Rai, P. In silico design of a multi-epitope vaccine targeting DENV-1 and DENV-3. Sci Rep 2026, 16, 5308. [CrossRef]
- Naveed, M.; Asim, M.; Aziz, T.; Athar, A.; Majeed, M.N.; Tombozara, N.; Al-Zaban, M.I.; Shami, A.; Alwethaynani, M.S.; Al-Joufi, F.A. In silico design and immunoinformatics assessment of a multiepitope vaccine targeting borealpox virus. Sci Rep 2026, 16, 3885. [CrossRef]
- Wu, H.; Zhao, C.; Cheng, Z.; Huang, W.; Yu, Y. In Silico Epitope-Based Peptide Vaccine Design Against Influenza B Virus: An Immunoinformatics Approach. Processes 2025, 13, 681. [CrossRef]
- Lu, Q.; Wu, H.; Meng, J.; Wang, J.; Wu, J.; Liu, S.; Tong, J.; Nie, J.; Huang, W. Multi-epitope vaccine design for hepatitis E virus based on protein ORF2 and ORF3. Front Microbiol 2024, 15, 1372069. [CrossRef]
Figure 1.
Docking analysis between MHC class I feline allele and selected epitopes (a) WVTILTYRH NS1 (b) IACVLNRQG NS1 (c) YSQRRASES NS2 (d) FRAKKAIAP VP1 (e) WGGKIGHYF VP1 (f) WIQNINFNL VP1 (g) FVKVAPNLT VP2 (h) LTALNNVPP VP2 (i) WIQNINFNL VP2.
Figure 1.
Docking analysis between MHC class I feline allele and selected epitopes (a) WVTILTYRH NS1 (b) IACVLNRQG NS1 (c) YSQRRASES NS2 (d) FRAKKAIAP VP1 (e) WGGKIGHYF VP1 (f) WIQNINFNL VP1 (g) FVKVAPNLT VP2 (h) LTALNNVPP VP2 (i) WIQNINFNL VP2.

Figure 2.
Docking analysis between MHC class II feline allele and selected epitopes (a) FMKYQDRQI NS1 (b) YSQRRASES NS2 (c) WGGKIGHYF VP1 (d) LDDIHAQIV VP2.
Figure 2.
Docking analysis between MHC class II feline allele and selected epitopes (a) FMKYQDRQI NS1 (b) YSQRRASES NS2 (c) WGGKIGHYF VP1 (d) LDDIHAQIV VP2.

Figure 3.
Schematic representation of the multiepitope vaccine construct targeting top-ranked epitopes from the major structural proteins (NS1, NS2, VP1 and VP2) of feline and canine parvovirus, (Top) Mapping of predicted B cell, MHC class I, and MHC class II epitopes across viral proteins. (Below-left side) Design of the final multiepitope vaccine construct comprising selected epitopes genetically linked to the adjuvants to enhance immunogenicity, (Below – right side) the red color represents the B cell epitopes, grey color represents the MHC class I epitopes and green color represents the MHC class II epitopes highlighted in vaccine construct.
Figure 3.
Schematic representation of the multiepitope vaccine construct targeting top-ranked epitopes from the major structural proteins (NS1, NS2, VP1 and VP2) of feline and canine parvovirus, (Top) Mapping of predicted B cell, MHC class I, and MHC class II epitopes across viral proteins. (Below-left side) Design of the final multiepitope vaccine construct comprising selected epitopes genetically linked to the adjuvants to enhance immunogenicity, (Below – right side) the red color represents the B cell epitopes, grey color represents the MHC class I epitopes and green color represents the MHC class II epitopes highlighted in vaccine construct.

Figure 4.
Structural analysis of the final multi-epitope vaccine construct: (A) shows the 3D structure of the vaccine construct modeled through Biovia discovery studio using template sequence alignment method, (B) shows the multi-epitope vaccine construct’s topology diagram to visualize the secondary structure elements arrangements, (C) 3D representation of spatial arrangement of C-alpha atoms predicted using Alpha fold prediction model and (D) Ramachandran plot confirms stability, a greater number of green dots on the respective region, confirms the quality of protein conformations, and ensures its accuracy.
Figure 4.
Structural analysis of the final multi-epitope vaccine construct: (A) shows the 3D structure of the vaccine construct modeled through Biovia discovery studio using template sequence alignment method, (B) shows the multi-epitope vaccine construct’s topology diagram to visualize the secondary structure elements arrangements, (C) 3D representation of spatial arrangement of C-alpha atoms predicted using Alpha fold prediction model and (D) Ramachandran plot confirms stability, a greater number of green dots on the respective region, confirms the quality of protein conformations, and ensures its accuracy.

Figure 5.
Stability of the vaccine construct by disulfide bond engineering in (a) original form and (b) mutant form. Six pairs of amino acids highlighted with the red stick shows the S-S bond and its respective X3 and energy value (left side).
Figure 5.
Stability of the vaccine construct by disulfide bond engineering in (a) original form and (b) mutant form. Six pairs of amino acids highlighted with the red stick shows the S-S bond and its respective X3 and energy value (left side).

Figure 6.
Normal-mode analysis of the vaccine protein, (a) B-factor - NMA mobility, (b) variance (purple color indicates individual variances, and green color indicates cumulative variances), (c) eigenvalues, (d) deformability (e) covariance map (correlated (red), uncorrelated (white) or anti-correlated (blue) motions), and (f) elastic network.
Figure 6.
Normal-mode analysis of the vaccine protein, (a) B-factor - NMA mobility, (b) variance (purple color indicates individual variances, and green color indicates cumulative variances), (c) eigenvalues, (d) deformability (e) covariance map (correlated (red), uncorrelated (white) or anti-correlated (blue) motions), and (f) elastic network.

Figure 7.
Molecular docking analysis of the multiepitope vaccine construct targeting final multi-epitope vaccine construct with the feline and canine Toll-like receptor (TLR4 and TLR5) using BIOVIA Discovery Studio. (a) Docked complex showing the binding interaction between the vaccine construct and feline TLR4, (b) Docked complex showing the binding interaction between the vaccine construct and canine TLR5.
Figure 7.
Molecular docking analysis of the multiepitope vaccine construct targeting final multi-epitope vaccine construct with the feline and canine Toll-like receptor (TLR4 and TLR5) using BIOVIA Discovery Studio. (a) Docked complex showing the binding interaction between the vaccine construct and feline TLR4, (b) Docked complex showing the binding interaction between the vaccine construct and canine TLR5.

Table 3.
Predicted MHC class-I epitopes across the conserved region of major proteins of feline and canine parvoviruses (NS1, NS2, VP1 and VP2) and their relevant information (percentile ranks and allele specification).
Table 3.
Predicted MHC class-I epitopes across the conserved region of major proteins of feline and canine parvoviruses (NS1, NS2, VP1 and VP2) and their relevant information (percentile ranks and allele specification).
| allele | Start | end | Peptide | Antigenic Score | Solubility |
| NS1 | |||||
| DLA-8850101 | 28 | 37 | FVFKCDNVQL | 0.7526 | 0.4731 |
| NS2 | |||||
| DLA-8850101 | 28 | 37 | FVFKCDNVQL | 0.7526 | 0.4731 |
| VP1 | |||||
| DLA-8850801 | 638 | 647 | KVAPNLTNEY | 0.8912 | 0.5638 |
| DLA-8803401 | 10 | 19 | RGLVPPGYKY | 0.5816 | 0.5708 |
| DLA-8803401 | 113 | 122 | KRSKPPPHIF | 1.1015 | 0.5288 |
| VP2 | |||||
| DLA-8850801 | 638 | 647 | KVAPNLTNEY | 0.8912 | 0.5638 |
| DLA-8803401 | 10 | 19 | RGLVPPGYKY | 0.5816 | 0.5708 |
| DLA-8803401 | 113 | 122 | KRSKPPPHIF | 1.1015 | 0.5288 |
*Non antigen, non-allergen, non-toxin.
Table 4.
Prediction of the ability of the designed multiepitope DNA pan parvovirus vaccine to trigger the IFN – interferon gamma production.
Table 4.
Prediction of the ability of the designed multiepitope DNA pan parvovirus vaccine to trigger the IFN – interferon gamma production.
| Type | Allele | Method of prediction | Inducer | Score |
| NS1/NS2 | FVFKCDNVQL | SVM based | POSITIVE | 0.45722211 |
| VP1/VP2 | KVAPNLTNEY | SVM based | POSITIVE | 0.43379691 |
| RGLVPPGYKY | SVM based | POSITIVE | 0.45733135 | |
| KRSKPPPHIF | SVM based | POSITIVE | 0.44701719 |
Table 5.
Predicted MHC class-II epitopes across the conserved region of major proteins of feline and canine parvoviruseses (NS1, NS2, VP1 and VP2) and their relevant information (percentile ranks and allele specification).
Table 5.
Predicted MHC class-II epitopes across the conserved region of major proteins of feline and canine parvoviruseses (NS1, NS2, VP1 and VP2) and their relevant information (percentile ranks and allele specification).
| Pos | MHC class II | Core | %Rank EL | Antigenic score | Solubility |
| NS1 | |||||
| 237 | DRB1_0103 | FMKYQDRQI | 6.79 | 0.8792 | 0.524 |
| 380 | DRB1_0104 | IACVLNRQG | 9.66 | 1.0045 | 0.4634 |
| 181 | DRB1_0103 | WVTILTYRH | 9.36 | 0.8681 | 0.4719 |
| 308 | DRB1_0102 | YIEMMAQPG | 0.31 | 0.6113 | 0.5619 |
| NS2 | |||||
| 139 | DRB1_0103 | YSQRRASES | 3.14 | 0.5799 | 0.528 |
| VP1 | |||||
| 488 | DRB1_0103 | FKTPIAAGR | 9.35 | 0.4738 | 0.6051 |
| 83 | DRB1_0103 | FRAKKAIAP | 1.85 | 1.0426 | 0.7233 |
| 80 | DRB1_0103 | HYFFRAKKA | 2.91 | 0.5722 | 0.5058 |
| 703 | DRB1_0102 | IVYEKSQLA | 0 | 0.7683 | 0.4781 |
| 588 | DRB1_0102 | LTALNNVPP | 6.16 | 0.9624 | 0.5226 |
| 155 | DRB1_0102 | VRNERATGS | 1.88 | 0.7287 | 0.5693 |
| 74 | DRB1_0103 | WGGKIGHYF | 4.25 | 1.0920 | 0.4548 |
| 548 | DRB1_0103 | WIQNINFNL | 4.54 | 1.1052 | 0.4649 |
| 643 | DRB1_0107 | YDPDASANM | 8.66 | 0.5605 | 0.6031 |
| 633 | DRB1_0102 | FVKVAPNLT | 2.19 | 0.9309 | 0.5289 |
| VP2 | |||||
| 350 | DRB1_0103 | FKTPIAAGR | 9.35 | 0.4738 | 0.6051 |
| 556 | DRB1_0102 | FNYLPNNIG | 0.63 | 0.9149 | 0.4604 |
| 496 | DRB1_0102 | FVKVAPNLT | 2.19 | 0.9309 | 0.5289 |
| 568 | DRB1_0102 | IVYEKSQLA | 0 | 0.7683 | 0.4781 |
| 94 | DRB1_0102 | LDDIHAQIV | 3.4 | 0.9059 | 0.4939 |
| 451 | DRB1_0102 | LTALNNVPP | 6.16 | 0.9624 | 0.5226 |
| 411 | DRB1_0103 | WIQNINFNL | 4.54 | 1.1052 | 0.4649 |
| 276 | DRB1_0102 | WQTNRALGL | 7.5 | 0.6719 | 0.5207 |
| 506 | DRB1_0107 | YDPDASANM | 8.66 | 0.5605 | 0.6031 |
| 448 | DRB1_0102 | YGPLTALNN | 7.91 | 0.4988 | 0.4919 |
*Non antigen, non-allergen, non-toxin.
Table 6.
Prediction of IL4 – interleukin4 inducing epitopes.
| Type | Allele | Method of prediction | Inducer | Score |
| NS1 | FMKYQDRQI | SVM based | POSITIVE | 1.24 |
| IACVLNRQG | SVM based | POSITIVE | 0.29 | |
| WVTILTYRH | SVM based | POSITIVE | 0.28 | |
| NS2 | YSQRRASES | SVM based | POSITIVE | 0.30 |
| VP1 | WGGKIGHYF | SVM based | POSITIVE | 0.28 |
| VP2 | LDDIHAQIV | SVM based | POSITIVE | 0.24 |
Table 7.
List of top ranked selected epitopes used for the construction of the multi-epitope vaccine against major proteins of the feline and canine parvovirus (NS1, NS2, VP1 and VP2).
Table 7.
List of top ranked selected epitopes used for the construction of the multi-epitope vaccine against major proteins of the feline and canine parvovirus (NS1, NS2, VP1 and VP2).
| Types | Epitopes | Antigenic Score | Docking Score |
Confidence Score |
| B cell | ADNTKLTNFDLAN | 1.3722 | ||
| TIVRIGCEERPEHTQP | 0.8492 | |||
| KLPGDFGLVDKEE | 1.3432 | |||
| MDQTEEEEMDWESE | 1.1828 | |||
| AKRARRGLV | 1.0456 | |||
| QRFIDQTKD | 1.0456 | |||
| FDTDLKPR | 1.5012 | |||
| AKRARRGLV | 1.0456 | |||
| TNEYDPDASANMS | 0.7027 | |||
| VYEKSQLAP | 1.1655 | |||
| MHC class I | FVFKCDNVQL | 0.7526 | -193.10 | 50.95 |
| KVAPNLTNEY | 0.8912 | -157.61 | 46.53 | |
| RGLVPPGYKY | 0.5816 | -214.32 | 50.43 | |
| KRSKPPPHIF | 1.1015 | -201.88 | 54.58 | |
| MHC class II | FMKYQDRQI | 0.8792 | -190.38 | 46.98 |
| YSQRRASES | 0.5799 | -227.71 | 51.50 | |
| WGGKIGHYF | 1.0920 | -222.16 | 45.80 | |
| LDDIHAQIV | 0.9059 | -168.88 | 51.17 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



