Submitted:
03 June 2026
Posted:
04 June 2026
You are already at the latest version
Abstract
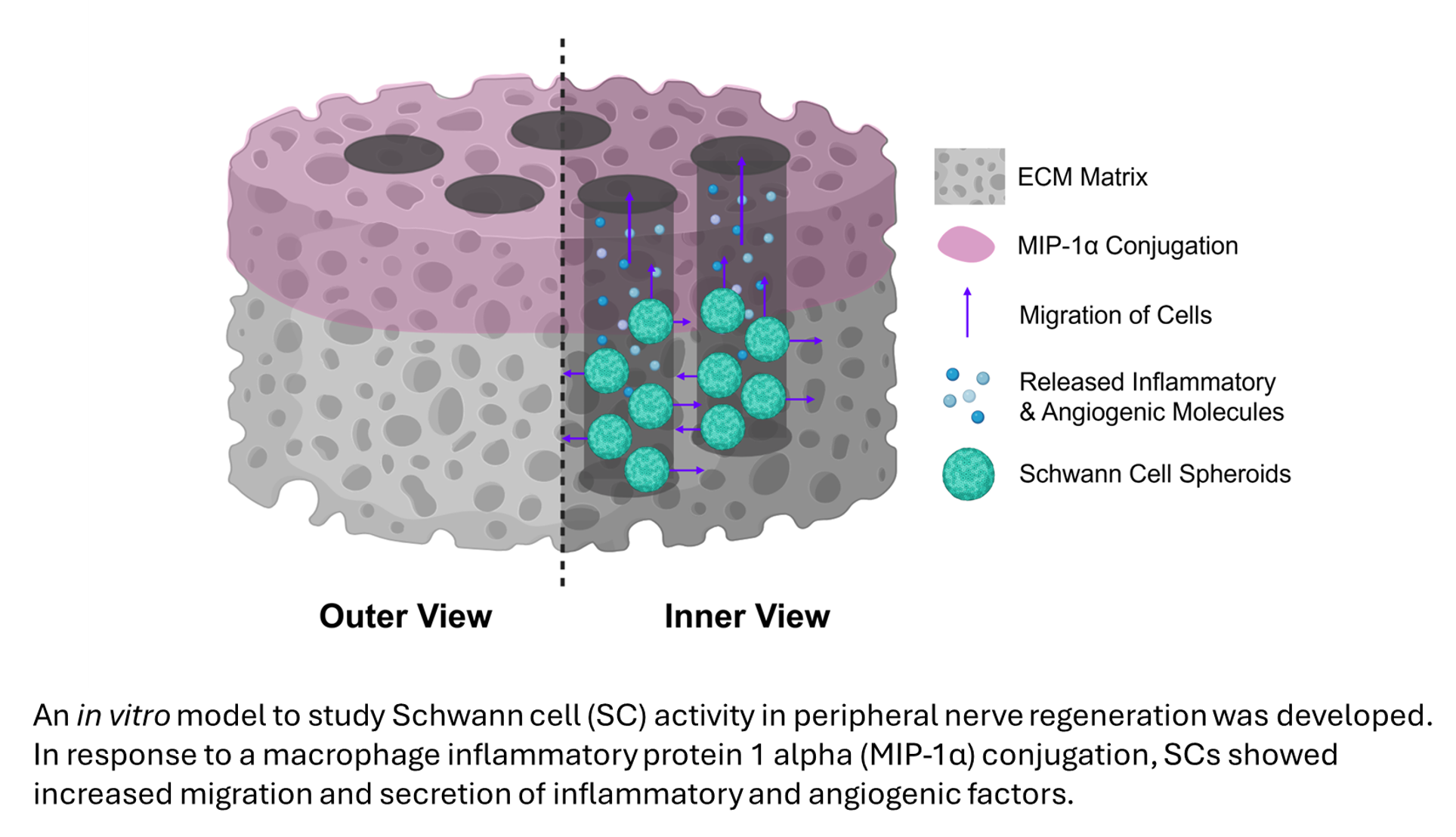
Peripheral nerve injuries represent significant clinical challenges, often resulting in lifelong motor function loss and disability. Evidence suggests regenerating axons cannot cross nerve gaps without Schwann Cell (SC) assistance. However, FDA-approved bio-material conduits for peripheral nerve repair lack bioactivity and structural complexity needed to facilitate SC migration. To address this, we developed a three-dimensional biomimetic in vitro model to simulate complex cellular interactions within the nerve bridge. The model features lyophilized hydrogel bioscaffolds with longitudinal channels to recapitulate the nerve microenvironment. To encourage directional SC migration, the top ~30% of the lyophilized hydrogel was conjugated with macrophage inflammatory protein-1α (MIP-1α). Cell line-derived SCs were seeded within no-MIP-1α- and MIP-1α-conjugated bioscaffold channels as spheroids and cultured for nine days. Histology demonstrated that MIP-1α Conjugation retained more SC spheroids during culture with greater cellular distribution. Cytokine profiling revealed MIP-1α conjugation activated SCs into a pro-repair phenotype characterized by increases in VEGF, ICAM-1, IL-6, and CINC-1 production, alongside downregulation of IL-1β, IL-10, and IL-13. The SC-derived pro-inflammatory and pro-angiogenic mediators did not inhibit NSC-34 motor neurite extension compared to controls. This study establishes an in vitro model that serves as both a screening platform and mechanistic tool, advancing our understanding of peripheral nerve repair.
Keywords:
Schwann cells
; spheroids
; neurogenesis
; bioscaffold
; in vitro modeling
; hydrogel
; lyophilization
1. Introduction
The peripheral nervous system (PNS) is comprised of a mixture of motor and sensory nerves which contribute to motor function and movement output [1]. Peripheral nerve injury (PNI) is a common occurrence following accidents in civil and military populations, which affects 2-5% of trauma patients worldwide [2,3,4,5,6]. After PNI, the regenerative window remains open only for a limited time. If robust regeneration is not achieved within this critical period, prolonged denervation can lead to demyelination of the nerves with detrimental and often permanent motor deficits [7,8,9,10]. This is especially crucial in complete transection injuries, known as laceration PNI or LPNI, as these injuries are clinically significant, result in extremely challenging recovery, and can lead to diminished quality of life [2,5].
Schwann cells (SCs) are responsible for myelinating large-diameter axons, are vital for saltatory conduction, and are the main mediators of peripheral nerve regeneration [4,7,11,12]. Directly following LPNI, damaged axons in the nerve bridge begin to breakdown in a process known as Wallerian degeneration, in which SCs respond to the degenerating axons signals and become reprogrammed into a progenitor-like phenotype that has multiple roles throughout nerve regeneration [8,13,14,15]. Reprogrammed SCs first assist ganglia macrophages in the controlled breakdown of myelin and axonal debris; then SCs dedifferentiate a second time where they migrate as cellular cords along regenerating nerve vasculature and support axon growth across the nerve bridge [13,15]. SC cord migration is guided by directional chemoattractant signaling from the chemokine CCL3, also known as macrophage inflammatory protein-1α (MIP-1α), which is secreted by hypoxic macrophages in the nerve bridge following laceration nerve injury and forms a chemotactic gradient [15]. Previous studies have shown that sufficient expression of the MIP-1α gradient following nerve injury is required for functional nerve regeneration as the loss of MIP-1α causes SCs to become misdirected, leading to less efficient migration and guidance of regenerating axons [15].
It has been demonstrated that migrating cords of SCs cannot navigate through the nerve bridge matrix if nerve vasculature has been compromised, indicating how SC mediated regeneration may not be feasible without surgical intervention in cases of LPNI [15,16]. Traditional nerve grafting surgical treatments, such as autographs, face many limitations in feasibility and efficacy, often leading to substantial donor-site deficits and complications [16]. To combat these limitations, biomaterial nerve conduits (BNCs) are a promising approach as they alleviate donor-site morbidity concerns and are widely available [5,10,16,17,18]. Current BNCs are made from a variety of materials, ranging from synthetic polymers to decellularized natural proteins, and often contain guiding pores, microchannels, and specific patterning to further encourage axon regeneration by providing physical support [7,9,10,16,17,18,19]. These scaffolds are primarily first- and second-generation biomaterials, as they provide physical support for the regenerating nerve and may be resorbable [16]. BNCs have shown great success in clinical applications, as evidence of their FDA approval, and have demonstrated improvements for nerve grafting surgery, such as improving surgical time and reducing difficulty [16]. However, BNCs have primarily enhanced functional efficacy in small nerve defects, with further research needed to progress treatments in more complex, larger nerve deficits [16].
Despite the success of current BNCs for nerve regeneration, FDA-approved nerve conduits lack directional biosignaling cues secreted by cells during natural SC-mediated regeneration, as demonstrated by MIP-1α [13,16]. To more closely mimic native nerve tissue, development of models that incorporate extracellular matrix proteins (ECM), factors, and cellular components can improve treatment efficacy for small and large nerve deficits alike, leading to development of enhanced third-generation BNCs [4,16,20]. Our lab has previously established a biomimetic, lyophilized hydrogel scaffold comprised of type 1 collagen, laminin-111, and type A gelatin crosslinked with 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC) and N-hydroxysuccinimide (NHS), which has demonstrated regenerative capabilities in skeletal muscle injuries [21]. Modifying the established biomimetic scaffold with guidance microchannels will mimic native nerve fascicle structure and provide physical support for cells within the matrix while satisfying biocompatibility considerations. Furthermore, the addition of MIP-1α onto the bioscaffold will provide the required chemical signaling needed to direct and initiate SC migration, which we aim to replicate. Implementing cellular components into this bioscaffold, such as culturing SCs, will further enhance regenerative capabilities and support biosignaling pathways [16]. Seeding the SCs as cellular spheroids will more closely mimic tissue density and microenvironment, as well as upregulate regenerative effects of SCs [8]. This is demonstrated by a previous study that investigated the injection of SCs in spheroidal geometry in a murine sciatic nerve injury model, which found enhanced pro-repair SC phenotype acquisition and regenerative potential [8]. Utilizing a comprehensive model of peripheral nerve regeneration will validate how the individual mechanisms that drive nerve repair work together to provide functional outcomes and will improve surgical solutions for LPNI.
To improve upon current treatment options, the development of a three-dimensional in vitro model of SC-mediated peripheral nerve regeneration is necessary to understand how to facilitate SC reprogramming. Using this in vitro system to model the nerve bridge during regeneration, we can identify the required chemical and physical guidance cues that current BNCs and traditional nerve grafts may be lacking, leading to less efficient patient outcomes. The main goal of this study is to create a three-dimensional biomimetic in vitro model of peripheral nerve regeneration utilizing a lyophilized hydrogel conduit comprised of ECM proteins with the addition of longitudinal channels for physical guidance, a chemotactic MIP-1α gradient for chemical guidance, and seeded SC spheroids for cellular investigation. We hypothesize that the creation of longitudinal channels through the bioscaffold, coupled with a MIP-1α chemotactic gradient, will provide both physical and chemical guidance cues to support the directional growth and migration of SCs through the bioscaffold’s matrix to closely replicate native nerve repair.
2. Results and Discussion
To assess the feasibility of the experimental model and examine the SC pro-repair phenotype acquisition, a variety of in vitro analyses were conducted. Formation of the BioGroove scaffold channels was verified using scanning electron microscopy (SEM) imaging. Model feasibility and optimization was conducted using SC spheroids with a density of 600 cells/spheroid. Spheroids were assessed for viability after four days of culture using a Live/Dead assay. Quantification of DAPI⁺ Area Fraction (%) and spheroid presence was determined via histological analysis. Distribution of seeded channels was examined using stitching of fluorescent histological images and Fast Fourier Transform analysis. Characterization of SC secretome in response to MIP-1α conjugation was determined using cytokine arrays and appropriate enzyme-linked immunosorbent assays (ELISAs) of collected culture supernatants. Analysis of neurite differentiation and extension in response to collected secretome to determine neuroregenerative potential was also performed.
2.1. Results
2.1.1. BioGroove Scaffold Characterization
SEM visualization of BioGroove scaffolds revealed that channels remain throughout the scaffold sterilization and lyophilization process, verifying successful GelGrooving (Figure 1). Channels were visually consistent in size and shape across the surface of each bioscaffold that was observed, demonstrating sufficient repetitions of GelGrooving had been achieved using the device (Figure 1A). Quantification of BioGroove channels determined an approximate diameter of 500 µm, which corresponds to the schematic design of the GelGroover device and confirmed that channels did not collapse throughout the drying process (Figure 1B). Transverse cross-sections of BioGroove scaffolds revealed that channels had obtained a depth of 3.5-4.5 mm (~70% of 5-7 mm bioscaffold height) (Figure 1C). The transverse view also demonstrated that the GelGroover device did not remove ECM material during channel formation and instead pushed the material aside.
2.1.2. Spheroid Viability
Spheroid viability at four days of culture was assessed utilizing a Live/Dead assay (Figure 2A). The percentage of living cells was significantly (p < 0.0001) greater than the percentage of dead cells, demonstrating viability throughout spheroid formation (Figure 2B). In addition, the average viability and average percentage of dead cells were calculated from the per field of view (PFV) percentages and determined to be approximately 98.75% and 1.25%, respectively. The diameter of four-day cultured spheroids was also determined, and the average spheroid diameter was determined to be 159.35 µm (Figure 2C).
2.1.3. Cellular Quantification and Analysis
Following nine days of culture, preserved spheroid seeded bioscaffolds were imaged at various magnifications for quantification (Figure 3). Representative histological images depict unorganized cellular content in the non-MIP-1α conjugated bioscaffolds and well-retained spheroids in the MIP-1α conjugated bioscaffolds following day 9 of culture (Figure 3A). The enhanced retention of spheroids in the MIP-1α conjugated bioscaffolds is indicative of reparative phenotype expression in the SCs, as the non-conjugated bioscaffolds contain a greater presence of single-cell suspension. MIP-1α conjugated bioscaffolds contained significantly (p = 0.0075) greater DAPI⁺ nuclei in the BioGroove channels compared to the non-conjugated controls (Figure 3B). Similar to DAPI⁺ area fraction, the number of spheroids was significantly (p = 0.0035) greater in response to the MIP-1α conjugation than in the non-conjugated controls (Figure 3C). These results verify increased spheroid retention in the presence of MIP-1α and further indicate enhanced reparative capabilities in the SC population due to the chemotactic conjugation.
2.1.4. Cellular Distribution Analysis
To visualize cellular distribution throughout the BioGroove channels, a series of 5X magnification DAPI images were taken of the seeded spheroids on day 9 (Figure 4). Stitched channel images revealed individual cells obtained vertical and lateral migration from the spheroids in both experimental groups (Figure 4A). Additionally, Fast Fourier Transform (FFT) analysis revealed a single discrete peak in the no-MIP-1α group, indicating that the cells remained sequestered within the primary longitudinal channel (Figure 4B). However, in the MIP-1α group, three distinct FFT peaks suggest that the cells showed significant lateral migration into the surrounding Biosponge matrix (Figure 4C). The multi-peak profile indicates a divergence from the initial seeding axis, reflecting a highly motile cellular phenotype.
2.1.5. SC Spheroid Secretome Characterization
Secretion of vascular endothelial growth factor (VEGF) and interleukin-1β (IL-1β) was determined on days 3, 5, 7 and 9 to further characterize cellular response in the BioGroove scaffolds (Figure 5). The release of IL-1β from SCs in cell culture supernatants was quantified (interaction p = 0.0245). IL-1β production was significantly increased on day 7 (post-hoc, p = 0.006) and day 9 (post-hoc, p = 0.0257) in the non-MIP-1α conjugated bioscaffolds, indicating a pro-inflammatory response (Figure 5A). Furthermore, ELISA quantification of VEGF showed a significant increase between treatment groups (column factor, p = 0.0125) but no significant interaction (p = 0.5415), which is further suggestive of an inflammatory response in the SCs due to MIP-1α exposure (Figure 5B). The significance of IL-1β and VEGF quantified results strongly indicate MIP-1α conjugation enhanced reparative secretions in the seeded SCs spheroids, peaking around day 7 of culture.
To further assess trophic factor secretion in response to MIP-1α conjugation, proteome profiling of day 1 (Figure 6A) and day 7 (Figure 6B) supernatants was performed. The release of MIP-1α from the conjugated bioscaffolds showed a roughly 4 log2 fold increase over the unconjugated ones on both days 1 and 7 indicating successful conjugation of the cytokine with sustained release. Quantification also revealed that MIP-1α conjugated bioscaffolds activated the SCs, leading to an increase (> 0.5 fold change) in Thymus Chemokine, RANTES (Regulated on Activation Normal T-Cell Expressed and Secreted), IL-6, and VEGF, alongside a downregulation (< -0.5 fold change) of GM-CSF (Granulocyte-Macrophage Colony-Stimulating Factor), IL-1ra (Interleukin-1 Receptor Antagonist), IL-2 at day 1 (Figure 6C). Quantification also revealed that MIP-1α conjugated bioscaffolds continued to activate the SCs, leading to an increase (> 0.5 log2 fold change) in ICAM-1 (Intercellular Adhesion Molecule-1), CINC-1 (Cytokine-induced Neutrophil Chemoattract 1), IL-6, and VEGF, alongside a downregulation (< -0.5 log2 fold change) of IL-13, Fractalkine, IL-17 at day 7 (Figure 6C). These secreted factors are indicative of an early, reparative response from the SCs in the presence of MIP-1α.
2.1.6. Motor Neuron Extension
Neuroregenerative potential in response to SC secretome was examined using a Mouse Motor Neuron-Like Hybrid Cell Line (NSC-34; RRID:CVCL_D356; CLU140, Cedarlane Cellutions Biosystems Inc.) extension assay (Figure 7). NSC-34 cells were differentiated in three different groups of media: positive control with regular differentiation media, a 50/50 mix of non-MIP-1α conjugated media with regular differentiation media, and a 50/50 mix of MIP-1α conjugated media with regular differentiation media (Figure 7A). While the mean neurite extension increased significantly over time (p < 0.0001), no significant differences were observed between treatment groups (interaction p = 0.9351) suggesting that the treatment had no effect on the rate of outgrowth (Figure 7B). Treatment did not alter neurite size distributions on days 1 and 3 but did on day 2 (interaction p = 0.0398) in the smallest size bin (Figure 7C-E). At this time point, MIP-1α conjugated treatment significantly increased the frequency of neurites in the smallest size bin (0–50 µm) relative to controls. No significant differences between the differentiation groups suggest that MIP-1α exposure does not hinder neurite growth despite its inflammatory nature.
2.2. Discussion
To address the spatial and biosignaling requirements of peripheral nerve regeneration, this study developed a 3D in vitro platform. A primary challenge in neuroengineering is replicating the anisotropic architecture of native nerve fascicles [22,23,24]. Without directional cues, regenerating axons disperse, leading to poor functional recovery and neuroma formation. By employing a straightforward technique, we successfully created longitudinal microchannels within a 3D lyophilized ECM hydrogel. This simple approach achieves biomimicry without complex equipment or scalability limitations often associated with microfluidic systems or high-resolution 3D printing. In characterizing the lyophilized hydrogel, SEM imaging demonstrated that the GelGroover device successfully created longitudinal channels for cell seeding with sufficient diameter and depth for spheroids.
Another key feature of the in vitro platform was the consideration of cellular density and microenvironmental signaling. Traditional models utilizing 2D monolayers or cell seeded 3D hydrogels fail to capture the cell-to-cell signaling at the injury site. To overcome this limitation, we introduced SCs in the form of high-density spheroids. This approach effectively mimics the dense packing of cells observed within the Bands of Büngner during Wallerian degeneration [25]. By grouping the cells into spheroids, we expected to positively modulate cell survival, proliferation, and the collective migratory behaviors essential for bridging large nerve gaps. SCs were aggregated into spheroids (600 cells/spheroid), seeded into the BioGroove scaffold channels, and cultured for nine days in vitro. SC spheroids have been shown to increase ECM protein deposition and cell-ECM interactions when compared to single-cell suspension, which enhances their viability when cultured in ECM-based hydrogel scaffolds [8]. A previous study determined SC spheroid viability within the range of 120 µm, but the spheroids created for this model were larger in diameter and cultured for longer durations, so a Live/Dead assay was necessary [8]. Proof of viability is crucial as large (> 200 µm) 3D spheroidal geometries can often lead to restricted diffusion of nutrients, oxygen, and growth factors into the spheroid core during long term culture, which may induce cell death within the inner most regions known as necrotic core formation [26].
To provide localized biochemical signaling, we conjugated MIP-1α directly to the lyophilized hydrogel. Following peripheral nerve injury, the sequential recruitment and activation of cells are primarily driven by chemokine gradients. Immobilizing MIP-1α through chemical crosslinking likely prevented the rapid, burst-release diffusion typical of soluble factors, thereby maintaining a sustained availability of MIP-1α within the bioscaffold. Histological analysis of seeded SC spheroids demonstrated significantly more identified nuclei in MIP-1α conjugated channels as well as higher retention of spheroid morphology. Furthermore, MIP-1α conjugated bioscaffolds also showed higher lateral cellular migration compared to the unconjugated bioscaffolds. These results suggest that the MIP-1α immobilization provides stabilizing cues that reinforce cell-matrix and cell-cell adhesion and results in a more conducive environment for SC growth.
Beyond mimicking physical architecture and supporting cellular proliferation and migration, this engineered microenvironment also modulated the secretory profile of the SCs. Peripheral nerve repair requires a coordinated cascade of biochemical signals to orchestrate cellular recruitment, growth, differentiation, and maturation [27]. In our model, cells cultured within the MIP-1α conjugated bioscaffolds released significantly higher levels of trophic factors specifically associated with inflammation and angiogenesis. This enhanced secretome is highly relevant to the in vivo injury response as localized inflammatory signals are required to promote macrophage infiltration, while angiogenic factors stimulate the neovascularization necessary to support axonal migration and cellular survival [28].
SC culture supernatants collected and pooled on day 1 revealed the highest expression of Thymus Chemokine, RANTES, IL-6, and VEGF, which are indicative of early inflammation due to MIP-1α conjugation. Thymus chemokines are heavily involved in neural-immune crosstalk mechanisms, including the modulation of T-cell recruitment and thymocyte migration to nerve injury sites [29]. RANTES, or CCL5, is secreted by SCs following nerve transection injuries and contributes to various nerve repair processes, including macrophage recruitment, immune cell recruitment, Wallerian degeneration, SC regulation, and promotion of nerve growth [30]. IL-6 is secreted by activated SCs and plays a key role in the regulation of immune responses, acute-phase responses, and inflammatory processes following nerve injury, further demonstrating that SCs are activated by this system [31,32]. Additionally, there was upregulation of VEGF on day 1, indicating a pro-reparative, angiogenic response throughout the entirety of culture aligning with the ELISA findings. The cytokine array also confirmed successful MIP-1α conjugation, as that was the factor with the greatest increase in fold change on day 1 which persisted through day 7. However, we acknowledge the possibility that exogenous immobilization of MIP-1α may have increased endogenous MIP-1α/CCL3 secretion through a potential positive autocrine or paracrine feedback loop mediated by the engineered matrix. Therefore, the elevated levels of MIP-1α may be attributed to release from both bioscaffolds and SCs.
Quantification of pooled day 7 SC supernatant revealed the greatest upregulations of ICAM-1, CINC-1, IL-6, and VEGF, which are indicative of early inflammation due to MIP-1α conjugation. ICAM-1 is a cell-surface adhesion receptor known for driving inflammation in epithelial injury-resolution responses [33,34]. In PNIs, ICAM-1 plays a role in recruiting endothelial cells to the nerve gap following injury. CINC-1 is typically upregulated following ischemia and helps to mediate neutrophil infiltration following nerve injury, also contributing to inflammation [35]. Similarly to the day 1 results, an increase in IL-6 production was determined and is likely due to the activation of SCs in response to MIP-1α exposure which works to modulate immune responses.
To further corroborate these findings, we determine the release of IL-1β and VEGF at various time-points of culture. For instance, IL-1β plays a key role in the differentiation mechanisms of SCs and is involved in the recruitment of immune cells, such as neutrophils and proinflammatory M1 macrophages [36,37]. We observed significantly elevated levels of IL-1β in the unconjugated bioscaffolds on days 7 and 9. These results suggest a reparative phenotype acquisition at an acute inflammatory timepoint as high levels of IL-1β can inhibit regeneration and cause systemic inflammation [31,38]. Previous studies have established that MIP-1α can impede the release of IL-1β during highly inflammatory reactions, particularly in immune cells like macrophages, as secretion of this interleukin is highly dependent on MIP-1α mechanisms [38,39]. Since SCs are differentiating into a macrophage-like phenotype when activated by this model, we theorize that IL-1β mechanisms may similarly be “turned off” by the cells within this modeling system due to high levels of MIP-1α exposure and acquired immune cell-like behavior. Furthermore, there has been discourse about the existence of an IL-1β feedback loop following high inflammation, suggesting that SCs are not adopting a hyper inflammatory or cytotoxic phenotype in response to MIP-1α conjugated bioscaffolds [39]. The reduction in IL-1β levels paired with an increase in VEGF highlights the selective amplification of pro-angiogenic and reparative signaling necessary for tissue bridge formation.
Taken together, these results indicate that the MIP-1α conjugated bioscaffolds drive an acute inflammatory SC response geared toward functional repair. Characterization of the day 1 and day 7 secretome revealed that MIP-1α conjugated bioscaffolds drive an acute inflammatory SC response geared toward functional repair. Because this study concluded at day 9, these data capture a critical early window in SC biology characterized by an acute phase response. Throughout this culture period, SCs persistently signal to recruit neutrophils, macrophages, and T-cells for myelin debris clearance, while concurrently signaling endothelial cells to initiate neurovascular remodeling. Crucially, the cells promote this microenvironment by suppressing anti-inflammatory signals (e.g., IL-13 and IL-10) to allow for thorough debris removal and also chronic inflammatory markers like IL-1β to prevent cytotoxicity.
Our results also show that SC conditioned media from the conjugated bioscaffolds did not inhibit or impair neurite outgrowth compared to the unconjugated control. Studies have shown that high concentrations of inflammatory cytokines can often exert neurotoxic or inhibitory effects on axonal growth. The observation that neurite extension remained unaffected indicates that the secretory profile induced by MIP-1α conjugated sbiocaffold does not compromise neuronal activity. This establishes the platform as a stable, biocompatible microenvironment capable of supporting the multi-lineage demands of the regenerative niche.
Future studies will investigate longer time points to assess long-term bioscaffold degradation and its impact on SC activity. By integrating endothelial cells into the culture system alongside SCs, this platform can help unravel the complex cellular and molecular crosstalk that drives peripheral nerve repair.
3. Conclusions
In summary, we have developed a biomimetic 3D platform that successfully integrates native ECM composition, directional topographies, and localized chemokine signaling. Because native ECM molecules like laminin and collagen are preserved within our lyophilized hydrogel, the conjugated MIP-1α works synergistically with these native cell-adhesive proteins to stabilize dense spheroidal cellular structures and enhance their proliferative and migratory capacity. It was also shown that the acute inflammatory environment induced by MIP-1α conjugation did not hinder neurite extension, further establishing the reparative nature of the SC phenotype acquisition. Ultimately, this integrated system serves as a valuable in vitro tool for studying SC mediated peripheral nerve repair in response to various neurotrophic factors and cytokines.
4. Materials and Methods
4.1. Bioscaffold Synthesis
Autoclaved deionized (DI) water was heated in a water bath to 75 °C to make a 3 wt% porcine skin gelatin (Sigma Aldrich, St. Louis, MO) solution. After the solution was completely dissolved, 21 mg EDC (Sigma Aldrich, St. Louis, MO) and 9.66 mg of NHS (Sigma Aldrich, St. Louis, MO) were added. Rat tail collagen I (11.61 mg/mL; BD Corning, Corning, NY) was diluted to a concentration of 3 mg/mL in 1X phosphate-buffered saline (PBS; Cytiva, South Logan, Utah). In a 24-well plate, 1.47 mL of the crosslinked-gelatin solution was combined with 630 μL of the collagen solution and 17.5 μL of LM-111 solution (6 mg/mL; R&D Systems, Minneapolis, MN). The final concentrations of the components were 21 mg/mL of gelatin, 0.9 mg/mL of collagen, 50 μg/mL of LM-111, 14.6 mM of EDC, and 5.6 mM of NHS. The solution was polymerized in the refrigerator (4 °C) for 1 hour to form a hydrogel and then froze overnight at -8 °C. The well plate was then moved to -80 °C for a minimum of 72 hours before lyophilization for 12-24 hours. The lyophilized scaffolds were frozen at -20 °C until needed.
4.2. GelGroover Design
To create longitudinal channels through the bioscaffold, a custom device was designed. As channel formation is a highly mechanical process, it was necessary to create a 3D printed device that would ensure accuracy and reproducibility of channels for experimentation purposes. The GelGroover device was made in collaboration with SLU Center for Additive Manufacturing (Saint Louis University, St. Louis, MO), following specifications in diameter, probe height, and probe width (Figure 8). The device was designed to be easily sterilized for stamping channels throughout the bioscaffold height.
4.3. Channel Formation and Scaffold Sterilization
Bioscaffolds were thawed at room temperature (RT) for 30 minutes, then sterilized with 70% ethanol and biopsy punched into a cylindrical geometry with a diameter of 10 mm. Bioscaffolds were moved to a 48-well plate for channel formation. The custom-made GelGroover device was sterilized with 70% ethanol and rinsed with 1X PBS. The GelGroover device was used to stamp 5 evenly spaced, longitudinal channels through ~70% of the height of the bioscaffold to form the BioGrooved scaffolds. Afterward, the BioGrooved scaffolds were rinsed twice with 1X PBS for 5 minutes and were allowed to dry at RT in the cell culture hood. Freshly lyophilized BioGrooved scaffolds were characterized using SEM (Saint Louis University, St. Louis, MO). Select bioscaffolds were cut transversely to observe channel depth. The BioGrooved scaffolds were sputter coated for 3 minutes (SCD 005, Bal-Tec, Canonsburg, PA). SEM imaging was performed at 2 kV to avoid sample burning. Channel diameter and depth were approximated using the accompanying SEM software and used to verify successful channel formation prior to cellular seeding.
4.4. Formation of Chemotactic Gradient
To form the crosslinking solution, 19.17 mg of EDC and 5.75 mg of NHS were added to 20 mL of sterile 1X PBS. Aliquots of MIP-1α (0.01mg/mL, PeproTech, Cranbury, NJ) were thawed on ice. In two microcentrifuge tubes, 1 mL of EDC/NHS crosslinking solution was added. In one of the tubes, 5 μL of MIP-1α was added to form a 50 ng/mL MIP-1α/EDC/NHS conjugate crosslinking solution. In a new 48-well plate, 100-200 μL of the select crosslinking solutions were added (n=5 wells MIP-1α/EDC/NHS solution, n=5 wells EDC/NHS solution). Thoroughly dried, sterilized BioGroove scaffolds were placed channel-side down in the wells containing crosslinking solutions and were allowed to soak in the incubator for 30 minutes to form the chemotactic gradient through absorption. The bioscaffolds were then flipped to be channel-side up and rinsed with 1X PBS for 5 minutes to remove unreacted EDC and NHS.
4.5. Spheroid Formation
Schwann cells (SCs; RT4-D6P2T murine Schwannoma Cells, ATCC, Manassas, VA) were cultured at 37 °C (5% CO₂) in a T75 flask with warm proliferation media until 80% confluent. Prior to spheroid formation, a small amount of SCs were cultured on a 24-well plate, then stained with s100β antibody and DAPI to verify cell identity and ensure homogeneity.
An Aggrewell-400™ well plate (STEMCELL Technologies, Vancouver, BC) was prepared via manufacturers protocol with anti-adherence solution (STEMCELL Technologies; Vancouver, BC) and rinsed twice with warm proliferation media. Then, 1 mL of warm proliferation media was added to each prepared well and placed in the incubator until usage. SCs were trypsinized (Trypsin; Gibco, Waltham, MA) and resuspended to a density of 720,000 cells/100 µL. Directly following resuspension, the prepared Aggrewell-400™ plate was removed from the incubator and 100 µL of cell suspension was added to each prepared well to achieve a density of 600 cells/spheroid. Another 0.9 mL of proliferation media was added to each well to achieve a total volume of 2 mL, and the plate was centrifuged at 100 g for 5 minutes to form micro-aggregates. The well plate was then transferred to the incubator on a rotary rocker and allowed culture for three days to form compact cellular spheroids. Spheroid formation was checked using brightfield microscopy to verify successful formation prior to cellular seeding.
4.6. Scaffold Culture
SC spheroids were removed from the microwells via vigorous pipetting and media washing per manufacturer’s protocol. The spheroids were collected in a 50 mL conical tube following each media wash and centrifuged at 200 g for 5 minutes. Then, SC spheroids were gently resuspended in proliferation media to a density of 1200 spheroids/50 µL of suspension. Each channel of the chemotactic and control BioGroove scaffolds were seeded directly with 10 µL of spheroid cell suspension to achieve a density of 1200 spheroids/bioscaffold. The spheroids were allowed to attach to the BioGroove scaffolds in the incubator for 15-20 minutes, then an additional 500 µL of proliferation media was added to supply nutrients and the plate was placed into the incubator for culture. After 24 hours, supernatant was collected in labeled microcentrifuge tubes, then the seeded bioscaffolds were moved to new wells and flipped horizontally to further promote cell migration. New proliferation media was added to all wells (former and current) and the bioscaffolds continued to culture horizontally. Thereafter, supernatant was collected every other day during feedings from each well and placed in labeled microcentrifuge tubes for later analysis. The seeded BioGroove scaffolds were cultured for a total of nine days, then cryopreserved in histomolds using optimal cutting temperature compound (OCT; Scigen, Houston, TX).
4.7. BioGroove Scaffold Histological Analysis Techniques
The preserved bioscaffolds were cryosectioned (Leica CM1850; Lecia Biosystems, St. Louis, MO) on charged slides. Cell-seeded bioscaffold cryosections were stained and cover slipped in a single step using Vectashield with DAPI (Vector Laboratories, Newark, CA). The slides were imaged using fluorescent microscopy (Axiovert 200M; Zeiss, Oberkochen, Germany) at 5X, 10X, and 20X magnification, ensuring no cellular overlap between images for analysis. DAPI Area fraction was determined for each 10X magnification images using the Area Fraction Analysis and a thresholding between 30-40 units on ImageJ (FIJI, GitHub). Spheroid quantification and characterization were determined for each 5X magnification image using the Particle Analysis (2000-Infinity, pixel units) and a thresholding between 30-40 units on ImageJ. DAPI Area Fraction Percent (%) and Spheroid Count were plotted in GraphPad PRISM version 10.0.0 (GraphPad, Boston, MA) for statistical analysis.
For cellular distribution analysis, the 5X magnification channel images were stitched together to form complete representations of channel cross-sections. These images were transformed into 16-bit representations and processed using the Oval Profile Plugin (Bill O’Connell, 2015, attbi.com) in ImageJ by encapsulating the spheroid channels with the oval tool. This allows the alignment of spheroids within the channels to be represented by relative intensity peaks (Gray Value) through Fast Fourier Transform analysis (FFT). The relative intensity FFT data was plotted using Excel for comparison.
A select group of SC spheroids cultured for four days were stained with Live/Dead™ staining (Life Technologies Corporation, Eugene, OR) and imaged at 5X magnification using fluorescent microscopy. Spheroids were quantified using ImageJ (Particle Analysis, 2000-Infinity) and the number of dead cells was also quantified (Particle Analysis, 0-Infinity). The total number of cells per field of view (PFV) was determined by multiplying spheroid count by the number of cells per spheroid (600 cells/spheroid). The percentage of dead cells and living cells was manually determined using Excel for each image and averaged to determine spheroid viability. Spheroid diameter was quantified from the Live/Dead stain images using scaling on ImageJ. Spheroid Viability and Spheroid Diameter were plotted in GraphPad PRISM version 10.0 for statistical analysis.
4.8. BioGroove Scaffold Biochemical Analysis Techniques
Cell culture supernatants collected on days 3, 5, 7, and 9 of culture were subjected to interleukin-1β (IL-1β) ELISA analysis (Kit 900-K91; PeproTech, Cranbury, NJ) and VEGF ELISA analysis (Kit 900-K436K; PeproTech, Cranbury, NJ) to determine expression throughout culture. ELISA data was measured using a spectrophotometer (SpectraMax i3, Molecular Devices, San Jose, CA) and its corresponding software (SoftMax Pro 6.3). Data collected from the software was processed through a customized MATLAB program to determine the desired standard fit, then was plotted in GraphPad PRISM version 10.0.0 for statistical analysis and visualization. A cytokine array on the SC spheroids (Proteome Profiler ™ Array, ARY008, R&D Systems, Minneapolis, MN) was performed on day 7 collected supernatant, as this represents the peak early inflammatory time point for cellular secretion. The intensity of positive expression demonstrated by the antibody dots on the membranes was quantified using the gel analysis feature on ImageJ. The dot intensities were plotted into peaks, and the area under each curve was measured to obtain relative values. A representative Log₂ Fold Change was calculated from the intensity values using Excel, which was plotted in GraphPad PRISM version 10.0.0 for further analysis.
4.9. NSC-34 Extension Assay
Neural stem cells (Mouse Motor Neuron-Like Hybrid Cell Line (NSC-34; RRID:CVCL_D356; CLU140, Cedarlane Cellutions Biosystems Inc.) were differentiated in response to collected experiment secretome. NSC-34 cells were cultured in a T75 flask with proliferation media (DMEM-F12 with 10% FBS and 1% P/S) to 60% confluency. Cells were trypsinized and resuspended to a density of 1x10⁶ cells/mL for plating. In a 96-well plate, 1 µL of cell suspension (~ 1000 cells) was added to 100 µL of proliferation media (n=5 wells) for each of the three experimental groups. NSC-34s were allowed to culture in proliferation media for 24 hours, then were differentiated using differentiation media based on experimental groupings. The positive control group was differentiated in regular differentiation media containing DMEM-F12, 1% FBS, 1% P/S, 1% Minimum Essential Medium Non-essential Amino Acids (MEM NEAA; 100X; Gibco, Brooklyn, NY), and 1 µM retinoic acid (RA; 50 mM in dimethyl sulfoxide). The non-MIP-1α and MIP-1α groups were cultured with a mix of regular differentiation media and collected D9 supernatant (no conjugation vs. conjugation) in a 1:1 ratio. The cells were differentiated for 72 hours with 100 µL of new differentiation media exchanged each day of culture. Brightfield microscopy was used to image the wells each day of differentiation for quantification in ImageJ of neurite extension length. Extension data was plotted in GraphPad PRISM version 10.0.0 for statistical analysis.
4.10. Data Analysis and Processing
All statistical analysis was performed in the GraphPad Prism version 10.0.0 program. For ELISA data, two-way ANOVA analysis was performed to determine statistical significance. To determine statistical significance of DAPI Area Fraction % and Spheroid Count, two-tailed Welch’s T-Test analysis was performed. To analyze Spheroid viability data, unpaired t-test analysis was performed. Outlier identification (ROUT) was performed on PRISM for all data sets.
Author Contributions
Conceptualization, K.G. and K.P.; methodology, K.G., K.P., A.J. and A.R.; software, K.P.; validation, K.P., A.R. and A.J.; formal analysis, K.G. and K.P.; investigation, K.P.; resources, K.G.; data curation, K.P.; writing—original draft preparation, K.P.; writing—review and editing, K.G., K.P., A.J. and A.R.; project administration, K.G.; funding acquisition, K.G.
Funding
This research was funded by National Institute of General Medical Sciences, grant number (NIGMS)-1R15GM129731- 02.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data is included in the manuscript. Additional information is available upon request.
Acknowledgments
None.
Conflicts of Interest
GenAssist, Inc. is developing products related to the research described in this paper. KG has an equity interest in GenAssist, Inc. and serves on the company’s scientific advisory board. The terms of this arrangement have been reviewed and approved by Saint Louis University, in accordance with its conflict-of-interest policies. The authors declare no other competing financial or non-financial interests.
Abbreviations
The following abbreviations are used in this manuscript:
| PNS | Peripheral Nervous System |
| PNI | Peripheral Nerve Injury |
| LPNI | Laceration Peripheral Nerve Injury |
| SC | Schwann Cells |
| MIP-1α | Macrophage Inflammatory Protein-1α |
| BNC | Biomaterial Nerve Conduit |
| ECM | Extracellular Matrix |
| EDC | 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide |
| NHS | N-hydroxysuccinimide |
| SEM | Scanning Electron Microscopy |
| PFV | Per Field of View |
| FFT | Fast Fourier Transform |
| VEGF | Vascular Endothelial Growth Factor |
| IL-1β | Interleukin-1β |
| RANTES | Regulated on Activation Normal T-Cell Expressed and Secreted |
| IL-6 | Interleukin-6 |
| GM-CSF | Granulocyte-Macrophage Colony-Stimulating Factor |
| CINC-1 | Cytokine-induced Neutrophil Chemoattract 1 |
| IL-13 | Interleukin-13 |
| IL-17 | Interleukin-17 |
| RT | Room Temperature |
| NSC-34 | Nerve Stem Cells-34 |
References
- Salome Stierli, V.I., Alison C. Lloyd, Schwann cell plasticity-roles in tissue homeostasis, regeneration, and disease. Wiley Online Library, 2019. 67(11): p. 2203-2215. [CrossRef]
- Bing Chen, Q.C., David B. Parkinson, Xin-peng Dun, Analysis of Schwann Cell Migration and Axon Regeneration Following Nerve Injury in the Sciatic Nerve Bridge. Frontiers in Molecular Neuroscience, 2019. 12. [CrossRef]
- Jue Ling, C.H., Shuxuan Zhang, Yahong Zhao, Meifeng Zhu, Xiaoxuan Tang, Qiaoyuan Li, Liming Xu, Yumin Yang, Progress in methods for evaluating Schwann cell myelination and axonal growth in peripheral nerve regeneration via scaffolds. Frontiers in Bioengineering and Biotechnology, 2023. 11. [CrossRef]
- Cecilia M M Motta, K.J.E., Chrys Wesdemiotis, Rebecca K Willits, Matthew L Becker, Enhancing Schwann cell migration using concentration gradients of laminin-derived peptides. Biomaterials, 2019. 218: p. 119335. [CrossRef]
- Georgios N. Panagopoulos, M., Panayiotis D. Megaloikonomos, MD, and Andreas F. Mavrogenis, MD, The Present and Future for Peripheral Nerve Regeneration. Orthopedics, 2017. 40: p. e141-e156. [CrossRef]
- Faranak Mankavi, R.I.a.H.W., Advances in Biomimetic Nerve Guidance Conduits for Peripheral Nerve Regeneration. Nanomaterials, 2023. 13(18): p. 2528. [CrossRef]
- Chun Liu, J.K., Christina Chan, Schwann Cells Enhance Penetration of Regenerated Axons into Three-Dimensional Microchannels. Tissue Engineering Regenerative Medicine, 2018. 15: p. 351-361. [CrossRef]
- Shih-Heng Chen, H.-W.W., Pei-Ching Yang, Shih-Shien Chen, Chia-Hsin Ho, Pei-Ching Yang, Ying-Chi Kao, Shao-Wen Liu, Han Chiu, Yu-Jie Lin, Er-Yuan Chuang, Jen-Huang Huang, Huang-Kai Kao, Chieh-Cheng Huang, Schwann cells acquire a repair phenotype after assembling into spheroids and show enhanced in vivo therapeutic potential for promoting peripheral nerve repair. Bioengineering and Translational Medicine, 2023. 9(2). [CrossRef]
- Alessandro Faroni, S.A.M., Paul J. Kingham, Adam J. Reid, Peripheral nerve regeneration: Experimental strategies and future perspectives. Advanced Delivery Drug Reviews, 2015. 82-83: p. 160-167. [CrossRef]
- Gianluca Ciardelli, V.C., Materials for Peripheral Nerve Regeneration. Macromolecular Bioscience, 2005. 6(1): p. 13-26. [CrossRef]
- Haesun Kim, T.M., David B. Parkinson, Plastic Fantastic: Schwann Cells and Repair of the Peripheral Nervous System. Stem Cells Translational Medicine, 2013. 2(8): p. 553-557. [CrossRef]
- Taveggia, C., Schwann cells–axon interaction in myelination. Current Opinion in Neruobiology, 2016. 39: p. 24-29. [CrossRef]
- Anne-Laure Cattin, A.C.L., The multicellular complexity of peripheral nerve regeneration. Current Opinion in Neruobiology 2016. 39: p. 38-46. [CrossRef]
- Jessica Schira, A.H., Gereon Poschmann, Brigida Ziegler, Hans-Peter Hartung, Kai Stühler, Patrick Küry, Secretome analysis of nerve repair mediating Schwann cells reveals Smad-dependent trophism. FASEB Publications, 2018. 33(4): p. 4703-4715. [CrossRef]
- Lucie Van Emmenis, G.M.-C., Elizabeth Harford-Wright, Alex Power, Anne-Laure Cattin, Ian J. White, Giulia Casal, Inês Boal-Carvalho, Clare L. Bennett, Alison C. Lloyd, Identification of CCL3 as a Schwann cell chemotactic factor essential for nerve regeneration. Cell Reports, 2025. 44(2). [CrossRef]
- Adriana Panayi, D.O., Current Use of Biological Scaffolds in Plastic Surgery. Journal of the American Society of Plastic Surgeons, 2018. 143(1): p. 209-220. [CrossRef]
- Le Yu, C.J.B., Chung-Hsun Lin, Su Yan and Jian Yang, Scaffold design considerations for peripheral nerve regeneration. Journal of Neural Engineering, 2024. 21(4). [CrossRef]
- Federica Cerri, L.S., Danish Memon, Filippo Martinelli Boneschi, Marta Madaghiele, Paola Brambilla, Ubaldo Del Carro, Carla Taveggia, Nilo Riva, Amelia Trimarco, Ignazio D. Lopez, Giancarlo Comi, Stefano Pluchino, Gianvito Martino, Alessandro Sannino, Angelo Quattrini, Peripheral nerve morphogenesis induced by scaffold micropatterning. Biomaterials, 2014. 35(13): p. 4035-4045. [CrossRef]
- Leila Lotfi, M.K., Mehrdad Moosazadeh Moghaddam, Shahin Bonakdar, A biomaterials approach to Schwann cell development in neural tissue engineering. Journal of Biomedical Materials Research Part A, 2019. 107(11): p. 2425-2446. [CrossRef]
- Raju Poongodi, Y.-L.C., Tao-Hsiang Yang, Ya-Hsien Huang, Kuender D. Yang, Hsin-Chieh Lin and Jen-Kun Cheng, Bio-Scaffolds as Cell or Exosome Carriers for Nerve Injury Repair. International Journal of Molecular Sciences, 2021. 22(24). [CrossRef]
- Gabriel J Haas, A.J.D., Madison Marcinczyk, Muhamed Talovic, Mark Schwartz, Robert Scheidt, Anjali D Patel, Katherine R Hixon, Hady Elmashhday, Sarah H McBride-Gagyi, Scott A Sell, Koyal Garg, Biomimetic sponges for regeneration of skeletal muscle following trauma. Journal of Biomedical Materials Part A, 2018. 107(1): p. 92-103. [CrossRef]
- Jiani Cao, C.S., Hui Zhao, Zhifeng Xiao, Bing Chen, Jian Gao, Tiezheng Zheng, Wei Wu, Shuang Wu, Jingyu Wang, Jianwu Dai, The use of laminin modified linear ordered collagen scaffolds loaded with laminin-binding ciliary neurotrophic factor for sciatic nerve regeneration in rats. Biomaterials, 2011. 32(16): p. 3939-48. [CrossRef]
- Xianhao Dong, S.L., Yueyue Yang, Shan Gao, Wenlei Li, Jiasong Cao, Ye Wan, Ziqi Huang, Guanwei Fan, Quan Chen, Hongjun Wang, Meifeng Zhu, Deling Kong, Aligned microfiber-induced macrophage polarization to guide schwann-cell-enabled peripheral nerve regeneration. Biomaterials, 2021. 272. [CrossRef]
- Victor T Ribeiro-Resende, B.K., Susanne Nichterwitz, Sven Oberhoffner, Burkhard Schlosshauer, Strategies for inducing the formation of bands of Büngner in peripheral nerve regeneration. Biomaterials, 2009. 30(29): p. 5251-9. [CrossRef]
- Namgung, U., The Role of Schwann Cell-Axon Interaction in Peripheral Nerve Regeneration. Cells Tissues Organs, 2015. 200(1): p. 6-12. [CrossRef]
- Christopher Rathnam, L.Y., Sofia Castro-Pedrido, Jeffrey Luo, Li Cai, Ki-Bum Lee Hybrid SMART spheroids to enhance stem cell therapy for CNS injuries. Cell Biology, 2021. 7(40). [CrossRef]
- K Ming Chan, T.G., Douglas W Zochodne, Hollie A Power, Improving peripheral nerve regeneration: from molecular mechanisms to potential therapeutic targets. Experimental Neurology, 2014. 261: p. 826-835. [CrossRef]
- Wesley S. Warner, M.R., Stewart Yeoh, Whitney E. Muhlestein, Sama Noroozi Gilandehi, Mark A. Mahan, The neuroimmune microenvironment of peripheral nerve injury: mechanisms, pathophysiology, and therapeutic implications. Fronteirs in Surgery, 2026. 13. [CrossRef]
- Randall S. Carpenter, M.K.L., George S. Karagiannis, Maria Maryanovich Neural regulation of the thymus: past, current, and future perspectives. Fronteirs in Immunology, 2025. 16. [CrossRef]
- Fangyuan Wang, C.Z., Zhou Jing, Qingyi Wang, Minghe Li, Bingqi Lu, Ao Huo, Wulong Liang, Weihua Hu, Xudong Fu, The dual roles of chemokines in peripheral nerve injury and repair. Inflammation and Regeneration, 2025. 45. [CrossRef]
- Yue Shen, M.L., ShengBo Jin, ZiYang Yu, Qian Liu, HuiNi Yao, YuXin Jiang, JiaQing Fu, and N. Li, The role of interleukins in peripheral nerve injury and the current status of treatment. Fronteirs in Immunology, 2026. 16. [CrossRef]
- María Erta, A.Q., Juan Hidalgo, Interleukin-6, a Major Cytokine in the Central Nervous System. International Journal of Biological Sciences, 2012. 8(9): p. 1254-1266. [CrossRef]
- Triet M Bui, H.L.W., Ronen Sumagin ICAM-1: A master regulator of cellular responses in inflammation, injury resolution, and tumorigenesis. Journal of Leukocyte Biology, 2020. 108(3): p. 787-799. [CrossRef]
- Akshay Chaudhari, J.P., Sanjay Awathale, Sameer Goyal, Kartik Nakhate, Abdulla Sherikar, From glial cells to pain pathways: ICAM-1 as a central player in neuroinflammation and neuropathy. Discover Neurosciences, 2025. 20(5). [CrossRef]
- Takahiro Katayama, H.T., Tadashi Yoshida, Takashi Uehara, Masabumi Minami, Neuronal Injury Induces Cytokine-Induced Neutrophil Chemoattractant-1 (CINC-1) Production in Astrocytes. Journal of Pharmacological Sciences, 2009. 109(1): p. 88-93. [CrossRef]
- Sylvain Nadeau, M.F., Ji Zhang, Bradley J Kerr, Serge Rivest, Denis Soulet, Yoichiro Iwakura, Juan Pablo de Rivero Vaccari, Robert W Keane, Steve Lacroix, Functional Recovery after Peripheral Nerve Injury is Dependent on the Pro-Inflammatory Cytokines IL-1β and TNF: Implications for Neuropathic Pain. The Journal of Neuroscience, 2011. 31(35): p. 12533-12542. [CrossRef]
- Yongkun Zhang, H.Z., Yun Su, Min Nuo, Wenguang Wu, Haochen Jiang, Xiangjun Meng The role of immune regulation in peripheral nerve regeneration: functions of inflammatory cells and cytokines. Fronteirs in Pharmacology, 2026. 17. [CrossRef]
- Anca-Laura Amati, A.Z., Robin Siebers, Sigrid Wilker, Sarah Heldmann, Dariusz Zakrzewicz, Andreas Hecker, J. Michael McIntosh, Winfried Padberg, Veronika Grau, Chemokines (CCL3, CCL4, and CCL5) Inhibit ATP-Induced Release of IL-1β by Monocytic Cells. Mediators of Inflammation, 2017. 2017(1). [CrossRef]
- Nevena Tzekova, A.H., Patrick Küry Molecules Involved in the Crosstalk Between Immune- and Peripheral Nerve Schwann Cells. Journal of Clinical Immunology, 2014. 34: p. 86-104. [CrossRef]
Figure 1.
SEM of lyophilized BioGroove Scaffolds at (A) 35X magnification, (B) 75X magnification, and (C) a transverse cross section at 45X magnification to demonstrate channel diameter and depth.
Figure 1.
SEM of lyophilized BioGroove Scaffolds at (A) 35X magnification, (B) 75X magnification, and (C) a transverse cross section at 45X magnification to demonstrate channel diameter and depth.

Figure 2.
Live/Dead assay analysis of SC spheroids at day 4 of culture to assess spheroid viability. (A) 5X magnification images of stained spheroids with the Live/Dead assay kit, with the green regions representing the alive cells and the red regions representing the dead cells; scale bar = 200 µm. (B) Spheroid viability (%) per field of view (PFV) in the 5X images. (C) Histogram of spheroid diameter measured from the Live/Dead images. **** indicates significance (p < 0.0001).
Figure 2.
Live/Dead assay analysis of SC spheroids at day 4 of culture to assess spheroid viability. (A) 5X magnification images of stained spheroids with the Live/Dead assay kit, with the green regions representing the alive cells and the red regions representing the dead cells; scale bar = 200 µm. (B) Spheroid viability (%) per field of view (PFV) in the 5X images. (C) Histogram of spheroid diameter measured from the Live/Dead images. **** indicates significance (p < 0.0001).

Figure 3.
DAPI staining and quantification of BioGroove scaffolds seeded with SC spheroids (D9). (A) 5X and 20X images, yellow outline represents the 20X region of interest, orange arrow represents the upward direction of the seeded channels; scale bars – 200 µm. (B) DAPI Area Fraction (%) per field of view of 10X DAPI images. (C) Number of spheroids quantified per field of view of the 5X images. ** indicates significance (p < 0.01).
Figure 3.
DAPI staining and quantification of BioGroove scaffolds seeded with SC spheroids (D9). (A) 5X and 20X images, yellow outline represents the 20X region of interest, orange arrow represents the upward direction of the seeded channels; scale bars – 200 µm. (B) DAPI Area Fraction (%) per field of view of 10X DAPI images. (C) Number of spheroids quantified per field of view of the 5X images. ** indicates significance (p < 0.01).

Figure 4.
(A) Stitched 5X images showing cellular distribution and migration in MIP-1α and non-MIP-1α conjugated channels; scale bar = 200 µm. (B) FFT analysis of non-MIP-1α conjugated channel distribution. (C) FFT analysis of MIP-1α-conjugated channel distribution.
Figure 4.
(A) Stitched 5X images showing cellular distribution and migration in MIP-1α and non-MIP-1α conjugated channels; scale bar = 200 µm. (B) FFT analysis of non-MIP-1α conjugated channel distribution. (C) FFT analysis of MIP-1α-conjugated channel distribution.

Figure 5.
ELISA quantification of collected supernatants from seeded BioGroove scaffold culture. (A) IL-1β ELISA quantification and (B) VEGF ELISA quantification. * indicates significance (* p < 0.05, *** p < 0.001).
Figure 5.
ELISA quantification of collected supernatants from seeded BioGroove scaffold culture. (A) IL-1β ELISA quantification and (B) VEGF ELISA quantification. * indicates significance (* p < 0.05, *** p < 0.001).

Figure 6.
Cytokine array of collected day 1 (D1) and day 7 (D7) supernatants from seeded BioGroove scaffold culture to compare early inflammatory time points. (A) D1 MIP-1α conjugated membrane and non-MIP-1α conjugated membrane. (B) D7 MIP-1α conjugated membrane and non-MIP-1α conjugated membrane. Corresponding color boxes around the specific secretion factors in (C) are located on the membranes (A, B) to demonstrate antibody location
Figure 6.
Cytokine array of collected day 1 (D1) and day 7 (D7) supernatants from seeded BioGroove scaffold culture to compare early inflammatory time points. (A) D1 MIP-1α conjugated membrane and non-MIP-1α conjugated membrane. (B) D7 MIP-1α conjugated membrane and non-MIP-1α conjugated membrane. Corresponding color boxes around the specific secretion factors in (C) are located on the membranes (A, B) to demonstrate antibody location

Figure 7.
(A) NSC-34 cells were differentiated in three groups (Control: regular differentiation media; No MIP-1α: 50/50 differentiation media with non-conjugated supernatant; MIP-1α: 50/50 differentiation media with conjugated supernatant) to examine elongation in response to D9 SC secretome; scale bar = 100 µm. (B) Elongation throughout entirety of culture duration and a (C-E) size-bin extension breakdown of each individual day. * indicates significance (p < 0.05).
Figure 7.
(A) NSC-34 cells were differentiated in three groups (Control: regular differentiation media; No MIP-1α: 50/50 differentiation media with non-conjugated supernatant; MIP-1α: 50/50 differentiation media with conjugated supernatant) to examine elongation in response to D9 SC secretome; scale bar = 100 µm. (B) Elongation throughout entirety of culture duration and a (C-E) size-bin extension breakdown of each individual day. * indicates significance (p < 0.05).

Figure 8.
(A) Schematic of GelGroover device design and (B) the GelGroover device.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



