Submitted:
01 June 2026
Posted:
03 June 2026
You are already at the latest version
Abstract
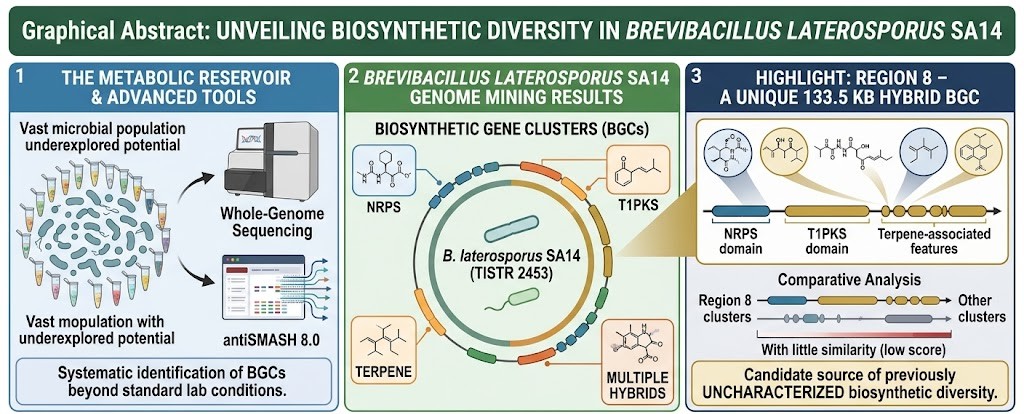
Microbial genomes represent a vast and still underexplored reservoir of biosynthetic diversity with significant pharmaceutical and biotechnological potential. Advances in whole-genome sequencing and computational genome mining have enabled the systematic identification of biosynthetic gene clusters (BGCs), providing insights into microbial secondary metabolic capabilities that may not be apparent under standard laboratory conditions.In this study, we performed genome mining of Brevibacillus laterosporus SA14 (TISTR 2453) using antiSMASH 8.0 to investigate its biosynthetic potential. The analysis identified a diverse repertoire of biosynthetic gene clusters, including non-ribosomal peptide synthetase (NRPS), type I polyketide synthase (T1PKS), terpene-associated, and multiple hybrid biosynthetic architectures.Notably, a large 133.5 kb hybrid biosynthetic gene cluster (Region 8) containing NRPS, T1PKS, and terpene-associated biosynthetic features was identified within a single contiguous genomic locus. The genomic organization of this region suggests a complex hybrid biosynthetic architecture with the potential to encode structurally diverse secondary metabolites. Comparative analysis indicated limited similarity to currently characterized biosynthetic gene clusters, highlighting Region 8 as a candidate source of previously uncharacterized biosynthetic diversity.The coexistence of peptide-, polyketide-, and terpene-associated biosynthetic features within a single genomic region highlights the evolutionary plasticity of microbial secondary metabolism and the potential for generating chemically diverse metabolites through hybrid biosynthetic systems. Such architectures are increasingly recognized as important contributors to microbial biosynthetic diversity and natural product innovation.Overall, the genome of B. laterosporus SA14 (TISTR 2453) contains a rich repertoire of biosynthetic gene clusters and represents a promising resource for future functional characterization, metabolomic investigation, and synthetic biology-based exploration of microbial secondary metabolism.
Keywords:
genome mining
; antiSMASH
; biosynthetic gene cluster
; NRPS
; PKS
; terpene
; hybrid biosynthesis
; Brevibacillus laterosporus SA14 (TISTR 2453)
1. Introduction
Microbial secondary metabolites represent one of the most important sources of bioactive compounds used in medicine, agriculture, and biotechnology. Numerous antibiotics, anticancer agents, immunosuppressants, and antifungal compounds have been discovered from microbial secondary metabolism, particularly from biosynthetic pathways encoded by biosynthetic gene clusters (BGCs) (Fischbach & Walsh, 2006; Covington et al., 2021; Cane, Walsh, & Khosla, 1998). Advances in whole-genome sequencing technologies have revealed that many microorganisms possess a substantially greater biosynthetic capacity than previously recognized through conventional cultivation-based approaches, leading to the emergence of genome mining as a powerful strategy for natural-product discovery (Medema & Fischbach, 2015; Bentley, Chater, Cerdeño-Tárraga, et al., 2002; Medema et al., 2011).
Members of the genus Brevibacillus are widely distributed in soil, water, and plant-associated environments and are recognized for their ability to produce diverse bioactive compounds with antimicrobial, insecticidal, and plant growth-promoting properties (Jiao, Takishita, Zhou, & Smith, 2016; Ruiu, 2013). Among them, Brevibacillus laterosporus has attracted considerable attention because of its capacity to synthesize antimicrobial peptides, extracellular enzymes, and biologically active metabolites that contribute to ecological fitness and biotechnological applications (Favret & Yousten, 1989; Ruiu, 2013).
The identification of biosynthetic gene clusters has become increasingly efficient through bioinformatics platforms such as antiSMASH (antibiotics and Secondary Metabolite Analysis Shell), which integrates multiple analytical modules for the prediction and annotation of secondary metabolite biosynthetic pathways in microbial genomes (Blin, Shaw, Kloosterman, Charlop-Powers, van Wezel, Medema, & Weber, 2021; Kautsar et al., 2020). AntiSMASH enables the detection of non-ribosomal peptide synthetase (NRPS), polyketide synthase (PKS), terpene, ribosomally synthesized and post-translationally modified peptide (RiPP), and hybrid biosynthetic systems, facilitating the discovery of potentially novel natural products (Medema et al., 2011; Navarro-Muñoz et al., 2020).
Particularly interesting are hybrid biosynthetic gene clusters containing NRPS, type I polyketide synthase (T1PKS), and terpene biosynthetic modules. Such hybrid systems can generate structurally complex metabolites through the integration of peptide, polyketide, and terpenoid biosynthetic pathways, frequently resulting in enhanced chemical diversity and biological activity (Fischbach & Walsh, 2006; Yamada et al., 2015; Chevrette et al., 2020; Rutledge & Challis, 2015).
Therefore, the present study aimed to investigate the biosynthetic potential of Brevibacillus laterosporus SA14 through whole-genome mining and identification of secondary metabolite biosynthetic gene clusters. Particular emphasis was placed on the characterization of a putative hybrid NRPS–T1PKS–terpene biosynthetic gene cluster that may represent a promising source of novel bioactive metabolites (Rudolf, Chang, Ma, & Shen, 2021; Helfrich et al., 2019).
2. Methodology
2.1. Bacterial Strain and Genome Sequence
The bacterial strain used in this study was Brevibacillus laterosporus SA14 (TISTR 2453). Whole-genome sequencing (WGS) of strain SA14 was performed, and a draft genome assembly was generated and used for downstream genome mining analyses.
The assembled genome was annotated and analyzed for biosynthetic gene clusters (BGCs) using antiSMASH version 6.0 (Blin et al., 2021) and online weblink under default bacterial parameters. The predicted BGCs were further used to explore the biosynthetic potential of strain SA14 and to identify putative secondary metabolite pathways (Medema et al., 2011).
Comparative sequence analysis indicated that the SA14 draft genome is consistent with publicly available genomes of Brevibacillus laterosporus, reflecting conserved genomic features within the species. However, strain-specific variations were observed in biosynthetic gene cluster regions, which were the primary focus of this study.
2.2. Genome Mining and Biosynthetic Gene Cluster Identification
Secondary metabolite biosynthetic gene clusters (BGCs) were identified using antiSMASH version 8.0, one of the most widely used platforms for microbial secondary metabolite genome mining (Blin, Shaw, Kloosterman, Charlop-Powers, van Wezel, Medema, & Weber, 2021; Medema et al., 2011). The assembled genome sequence was uploaded to the antiSMASH web server and analyzed using default parameters with relaxed detection strictness to maximize the detection of putative biosynthetic regions.
The analysis included prediction of biosynthetic pathways associated with non-ribosomal peptide synthetases (NRPSs), type I polyketide synthases (T1PKSs), terpenes, ribosomally synthesized and post-translationally modified peptides (RiPPs), and hybrid biosynthetic systems. Cluster annotations were compared against reference biosynthetic gene clusters available in the Minimum Information about a Biosynthetic Gene Cluster (MIBiG) database integrated within antiSMASH (Kautsar et al., 2020; Navarro-Muñoz et al., 2020).
2.3. Identification of Hybrid Biosynthetic Gene Clusters
Hybrid biosynthetic gene clusters were identified based on antiSMASH annotations indicating the presence of multiple biosynthetic systems within a single genomic region. Particular attention was given to clusters containing NRPS, T1PKS, and terpene-associated biosynthetic genes because hybrid pathways frequently generate structurally diverse natural products with enhanced biological activities (Fischbach & Walsh, 2006; Yamada et al., 2015; Chevrette et al., 2020).
The genomic coordinates, cluster size, gene composition, and predicted biosynthetic modules were extracted and analyzed for further characterization.
2.4. Functional Annotation of Biosynthetic Genes
Genes located within identified biosynthetic regions were functionally annotated using antiSMASH-integrated domain prediction algorithms. Conserved catalytic domains including adenylation (A), condensation (C), thiolation (T), ketosynthase (KS), acyltransferase (AT), ketoreductase (KR), and terpene synthase domains were examined to infer their potential biosynthetic functions (Medema et al., 2011; Fischbach & Walsh, 2006).
Predicted biosynthetic enzymes and accessory proteins were analyzed based on domain architecture and genomic organization to evaluate their potential contribution to secondary metabolite biosynthesis.
2.5. Analysis of the Hybrid NRPS–T1PKS–Terpene Cluster
A putative hybrid NRPS–T1PKS–terpene biosynthetic gene cluster identified within the genome of B. laterosporus SA14 was selected for detailed investigation. Cluster organization, biosynthetic modules, and neighboring genes were examined to identify core biosynthetic enzymes and accessory proteins potentially involved in metabolite assembly and modification (Rutledge & Challis, 2015; Rudolf, Chang, Ma, & Shen, 2021).
The cluster length, gene arrangement, and biosynthetic architecture were visualized using antiSMASH-generated outputs and manually inspected for evidence of integrated peptide, polyketide, and terpene biosynthetic systems.
2.6. Data Visualization
Graphical representations of biosynthetic gene clusters were generated using antiSMASH visualization tools. Gene organization, domain architecture, and biosynthetic features were interpreted according to established genome mining methodologies for microbial secondary metabolites (Medema & Fischbach, 2015; Helfrich et al., 2019).
3. Results
3.1. Genome Mining Revealed Extensive Secondary Metabolite Biosynthetic Potential in Brevibacillus laterosporus SA14
Genome mining analysis of Brevibacillus laterosporus SA14 using antiSMASH identified a substantial repertoire of biosynthetic gene clusters (BGCs), highlighting the metabolic versatility of this strain. A total of 20 biosynthetic regions comprising 29 predicted protoclusters were detected throughout the genome, indicating a rich reservoir of secondary metabolite biosynthetic pathways.
To obtain an overview of the biosynthetic capacity of strain SA14, the identified BGCs were categorized according to their predicted biosynthetic classes. As shown in Figure 1, NRPS-containing clusters represented the most abundant category, followed by hybrid biosynthetic systems, RiPP-associated clusters, and PKS-containing pathways. The diversity of BGC classes suggests that strain SA14 possesses extensive genetic resources for the biosynthesis of structurally diverse secondary metabolites.
The identified BGCs included diverse classes of natural product pathways, such as non-ribosomal peptide synthetases (NRPSs), type I polyketide synthases (T1PKSs), terpene-associated clusters, and several hybrid biosynthetic systems. The presence of multiple biosynthetic categories suggests that strain SA14 possesses considerable genetic capacity for the production of structurally diverse bioactive metabolites.
A detailed summary of all biosynthetic regions identified by antiSMASH is presented in Table 1. The detected clusters varied considerably in both biosynthetic class and genomic size, ranging from small RiPP-associated loci to large hybrid biosynthetic systems exceeding 100 kb. Notably, Region 8 was identified as the largest and most complex cluster, containing integrated NRPS, T1PKS, and terpene biosynthetic modules.
Among the detected clusters, several exhibited complex biosynthetic architectures characterized by the co-occurrence of multiple catalytic systems within a single genomic locus. Such hybrid clusters are frequently associated with the biosynthesis of chemically unique metabolites and are considered promising targets for natural-product discovery.
3.2. Identification of a Putative Hybrid NRPS–T1PKS–Terpene Biosynthetic Gene Cluster
Genome mining analysis of Brevibacillus laterosporus SA14 using antiSMASH 8.0 identified 20 putative biosynthetic gene clusters (BGCs) associated with secondary metabolite production. These included NRPS, terpene, RiPP, and hybrid biosynthetic systems. A genome-wide overview of all predicted BGCs is presented in Supplementary Figure S1.
Among the predicted BGCs, Region 8 was identified as one of the most notable biosynthetic loci. This region is located between nucleotide positions 2,467,611 and 2,601,073 and spans approximately 133.5 kb. AntiSMASH annotated Region 8 as a hybrid biosynthetic gene cluster containing NRPS, T1PKS, and terpene-associated biosynthetic features. Owing to its large size, complex domain architecture, and low similarity to previously characterized biosynthetic gene clusters, Region 8 was selected for further investigation.
The cluster contained more than 60 predicted coding sequences, including genes designated BRLA_RS11305 through BRLA_RS11605 together with several neighboring accessory genes. AntiSMASH further classified the region as an interleaved hybrid biosynthetic system, indicating the co-localization of genes associated with multiple biosynthetic pathways within a single genomic locus.
To further examine the genetic architecture of this biosynthetic region, the organization of Region 8 was visualized based on antiSMASH annotation outputs. As illustrated in Figure 2, the cluster contains NRPS-, T1PKS-, and terpene-associated biosynthetic genes together with putative transporter and regulatory elements distributed throughout the locus. The arrangement of these biosynthetic components supports its classification as a large hybrid biosynthetic gene cluster.
Comparison against the MIBiG reference database integrated within antiSMASH did not identify any closely related characterized biosynthetic gene cluster, suggesting that Region 8 may represent a previously uncharacterized biosynthetic architecture. Nevertheless, functional relationships to known biosynthetic systems cannot be excluded and will require further comparative genomic and experimental validation.
The large size of the cluster and the coexistence of peptide-, polyketide-, and terpene-associated biosynthetic genes suggest the potential for the biosynthesis of structurally diverse secondary metabolites. Similar hybrid biosynthetic architectures have been reported in microorganisms possessing complex secondary metabolic repertoires. However, the chemical structures and biological functions of any metabolites associated with Region 8 cannot be determined solely from genomic information and require experimental validation.
Key structural characteristics of Region 8 are summarized in Table 2. Among the biosynthetic regions identified in the genome, Region 8 represents the largest predicted hybrid biosynthetic locus and is distinguished by the coexistence of three major biosynthetic systems within a single genomic region. These features identify Region 8 as a candidate hybrid biosynthetic gene cluster warranting further functional, metabolomic, and biochemical investigation.
3.3. Biosynthetic Organization of the Hybrid Cluster
The hybrid biosynthetic region identified in Brevibacillus laterosporus SA14 exhibited a complex genetic organization spanning approximately 133.5 kb. AntiSMASH classified this region as an interleaved hybrid biosynthetic gene cluster containing NRPS-, T1PKS-, and terpene-associated biosynthetic features. More than 60 predicted coding sequences were distributed throughout the cluster, including genes designated BRLA_RS11305 to BRLA_RS11605. The extensive size of the region and the coexistence of multiple biosynthetic categories suggest a highly integrated secondary metabolite biosynthetic locus.
To provide a detailed overview of the functional composition within Region 8, the predicted genes were categorized based on their putative roles. Table 3 summarizes the occurrence of core biosynthetic genes, accessory enzymes, regulatory elements, transporters, and hypothetical proteins
Figure 3 illustrates the proposed biosynthetic model of Region 8. The diagram represents a predicted pathway integrating NRPS-mediated peptide assembly, T1PKS-mediated polyketide chain elongation, and terpene modification processes, potentially contributing to the biosynthesis of structurally diverse secondary metabolites. This conceptual model complements the functional overview provided in Table 3, highlighting the coordinated interactions between core and accessory biosynthetic components.
3.4. Candidate Core Biosynthetic Genes
Several genes located within the central portion of Region 8 were identified as candidate core biosynthetic genes based on antiSMASH annotations and conserved biosynthetic signatures. These genes were primarily associated with NRPS, T1PKS, and terpene-associated biosynthetic functions and are predicted to contribute to the biosynthetic capacity of the cluster.
The identified genes are predicted to constitute major biosynthetic components of the hybrid cluster. Representative candidate genes together with their predicted biosynthetic functions are summarized in Table 4.
The identified genes collectively represent the enzymatic core of the hybrid biosynthetic pathway. Notably, multiple NRPS-associated genes were distributed throughout the central region of the cluster, together with a type I polyketide synthase and a terpene-associated cyclase. This arrangement is consistent with the interleaved hybrid architecture predicted by antiSMASH and supports the classification of Region 8 as a complex hybrid biosynthetic gene cluster containing NRPS-, T1PKS-, and terpene-associated features.
Furthermore, several transporter proteins and regulatory elements were identified in proximity to the core biosynthetic genes, including MFS transporters, ABC transporters, and a Crp/Fnr-family transcriptional regulator. These accessory components may contribute to pathway regulation, metabolite export, and self-resistance mechanisms associated with secondary metabolite production.
To further illustrate the enzymatic complexity of Region 8, representative biosynthetic enzymes were organized according to their predicted catalytic domains. As shown in Figure 4, the identified enzymes contain characteristic NRPS, T1PKS, and terpene-associated domains that collectively support the hybrid biosynthetic architecture predicted by antiSMASH.
3.5. Distribution of Biosynthetic Classes
To further evaluate the biosynthetic diversity of B. laterosporus SA14, the identified biosynthetic regions were grouped according to their predicted biosynthetic classes. NRPS-containing clusters represented the most abundant category, followed by hybrid biosynthetic systems and RiPP-associated clusters. The predominance of NRPS-related pathways suggests that peptide-based secondary metabolism constitutes a major component of the biosynthetic repertoire of strain SA14.
4. Discussion
The genome mining of Brevibacillus laterosporus SA14 reveals a remarkably rich biosynthetic landscape, characterized by 20 biosynthetic regions comprising 29 predicted protoclusters, indicating extensive metabolic potential for secondary metabolite production. This level of biosynthetic density is consistent with the emerging view that environmental bacteria often encode substantially more biosynthetic capacity than is expressed under standard laboratory conditions (Medema & Fischbach, 2015; Blin et al., 2021). The dominance of NRPS-containing clusters further supports the established role of non-ribosomal peptide systems as major contributors to bioactive chemical diversity (Fischbach & Walsh, 2006).
Beyond overall abundance, SA14 exhibits a marked diversification in biosynthetic architecture. In addition to canonical single-class clusters, the genome contains multiple hybrid NRPS–PKS and NRPS–T1PKS–terpene systems. This indicates a shift from simple modular biosynthesis toward more integrated enzymatic architectures, distinguishing SA14 from many previously characterized Brevibacillus strains that typically display less frequent hybridization of biosynthetic systems.
Importantly, this architectural complexity suggests that SA14 does not merely expand biosynthetic quantity, but reorganizes biosynthetic logic at the genomic level. Such a pattern implies increased potential for chemical diversification through the coupling of peptide, polyketide, and terpene biosynthetic pathways within shared genetic loci.
Beyond the overall abundance of biosynthetic gene clusters, the genome of Brevibacillus laterosporus SA14 exhibits substantial diversity in biosynthetic architecture. In addition to canonical NRPS, PKS, and terpene-associated clusters, multiple hybrid biosynthetic regions were identified, including NRPS–PKS and NRPS–T1PKS–terpene-associated systems. The coexistence of multiple biosynthetic classes within single genomic loci suggests a complex organization of secondary metabolism and highlights the potential for the generation of structurally diverse metabolites.
Among the identified biosynthetic regions, Region 8 represents the largest predicted biosynthetic locus, spanning approximately 133.5 kb and containing more than 60 coding sequences. AntiSMASH classified this region as an interleaved hybrid biosynthetic gene cluster containing NRPS-, T1PKS-, and terpene-associated biosynthetic features. Such hybrid architectures have increasingly been recognized as important contributors to microbial biosynthetic diversity because they combine distinct biosynthetic logics within a single genetic framework (Fischbach & Walsh, 2006; Rutledge & Challis, 2015; Chevrette et al., 2020).
Comparison against the MIBiG database did not identify a closely matching characterized biosynthetic gene cluster, suggesting that Region 8 may represent a previously uncharacterized biosynthetic architecture. However, genomic predictions alone are insufficient to determine metabolite structures, biological activities, or pathway functionality. Consequently, experimental validation will be necessary to establish the biochemical products encoded by this region.
Within the species context, this positions SA14 as a distinct biosynthetic outlier characterized by increased modular integration. Rather than existing as isolated biosynthetic units, NRPS, PKS, and terpene-associated pathways appear to be more frequently co-localized and interwoven, suggesting an evolutionary trajectory favoring biosynthetic connectivity rather than independent pathway expansion.
Such genomic organization is consistent with evolutionary mechanisms including horizontal gene transfer, domain shuffling, and recombination-driven assembly of biosynthetic modules, all of which are recognized drivers of microbial secondary metabolite diversification (Fischbach et al., 2008; Chevrette et al., 2020).
In addition to this structural integration, several biosynthetic gene clusters identified in SA14 exceed 80–100 kb in length and show low similarity to entries in the MIBiG repository. This suggests that a substantial fraction of the biosynthetic repertoire may represent uncharacterized or novel natural product scaffolds, expanding the chemical space associated with the genus Brevibacillus (Kautsar et al., 2020; Navarro-Muñoz et al., 2020).
Among all identified loci, Region 8 represents the most prominent biosynthetic feature. This 133.5 kb cluster contains more than 60 predicted coding sequences and integrates NRPS, T1PKS, and terpene-associated genes within a single contiguous genomic region. Unlike typical single-pathway clusters, Region 8 exhibits a composite architecture in which multiple biosynthetic systems are physically co-localized, suggesting coordinated metabolic potential.
The internal structure of Region 8 further supports this interpretation. Core biosynthetic genes are accompanied by regulatory elements, transport systems, and accessory enzymes, indicating that the cluster functions as an integrated biosynthetic unit rather than a fragmented assembly of genes. The presence of multiple NRPS modules alongside a type I PKS and terpene cyclase supports the possibility of coordinated biosynthetic processing across chemically distinct pathways.
A notable feature of this locus is the integration of terpene biosynthetic capacity within a predominantly NRPS–PKS framework. Although bacterial terpene biosynthesis was historically underappreciated, recent genomic studies have demonstrated that terpene synthases are widely distributed and functionally diverse in bacterial systems (Yamada et al., 2015). In this context, terpene-derived moieties may contribute structural rigidity, hydrophobic interactions, or ecological signaling functions that enhance the overall chemical performance of hybrid metabolites.
From a functional perspective, Region 8 appears to represent a tightly coordinated biosynthetic system. The presence of regulatory proteins, ABC/MFS transporters, and multiple tailoring enzymes suggests that biosynthesis is coupled with transcriptional regulation and metabolite export, ensuring metabolic efficiency and cellular protection (Medema et al., 2011; Kautsar et al., 2020).
Nevertheless, it is important to emphasize that all biosynthetic predictions remain computational. While antiSMASH provides high-confidence identification of biosynthetic gene clusters, it does not confirm metabolite production or chemical structures (Blin et al., 2021). Furthermore, many biosynthetic gene clusters remain transcriptionally silent under standard laboratory conditions, limiting direct correlation between genomic potential and metabolomic output (Rutledge & Challis, 2015; Covington et al., 2021). Therefore, Region 8 should be regarded as a high-confidence candidate locus requiring experimental validation.
To bridge this gap between prediction and function, integrative experimental strategies such as transcriptomics and metabolomics will be essential. These approaches may reveal environmental conditions that activate silent biosynthetic clusters and enable detection of corresponding metabolites. In addition, structural prediction tools such as AlphaFold provide opportunities to refine enzymatic function hypotheses at near-atomic resolution, particularly for modular NRPS and PKS systems (Jumper et al., 2021; Varadi et al., 2022).
From an evolutionary perspective, the biosynthetic architecture observed in SA14 is consistent with models of microbial secondary metabolite diversification driven by gene duplication, horizontal gene transfer, and domain recombination. These processes collectively enable the continuous generation of novel biosynthetic configurations, expanding microbial chemical space (Fischbach et al., 2008; Chevrette et al., 2020). In this framework, Region 8 may represent a long-term evolutionary fusion event in which distinct biosynthetic systems have become integrated into a unified functional platform.
Overall, B. laterosporus SA14 represents a metabolically versatile and biosynthetically enriched organism with substantial potential for natural product discovery. The presence of a large hybrid NRPS–T1PKS–terpene cluster expands the known biosynthetic repertoire of the genus and highlights the utility of genome mining for uncovering previously inaccessible chemical space. While functional and structural validation remains necessary, the genomic evidence strongly supports SA14 as a promising source of structurally novel and biologically relevant secondary metabolites.
4.1. Hypothetical Biosynthetic Model of the Hybrid NRPS–T1PKS–Terpene Cluster
Region 8 of B. laterosporus SA14 spans 133.5 kb and contains more than 60 predicted coding sequences. Based on antiSMASH annotation, the region is predicted to contain NRPS, type I PKS (T1PKS), and terpene-associated biosynthetic features. The genomic organization of these biosynthetic genes suggests potential interactions among multiple biosynthetic systems within a single contiguous locus. However, the proposed biosynthetic model should be regarded as a computational prediction and requires experimental validation through transcriptomic, proteomic, and metabolomic analyses.
Candidate NRPS genes (e.g., BRLA_RS11395, BRLA_RS11410, BRLA_RS11525–11535) encode canonical adenylation (A), condensation (C), and thiolation (T) domains that are predicted to initiate biosynthesis through sequential amino acid activation and peptide chain assembly. This early peptide scaffold likely serves as the structural backbone onto which additional chemical complexity is introduced. The interspersed organization of NRPS modules further supports a modular, iterative assembly-line mechanism consistent with classical non-ribosomal peptide biosynthesis (Fischbach & Walsh, 2006; Medema et al., 2011).
Downstream of the NRPS core, the T1PKS gene BRLA_RS11400 encodes a multidomain enzyme containing ketosynthase (KS), acyltransferase (AT), and ketoreductase (KR) domains. We hypothesize that polyketide chain extension occurs either through direct interaction with the NRPS-derived intermediate or via a hybrid NRPS–PKS interface, supported by the presence of NRPS/PKS fusion features in BRLA_RS11475. This suggests a metabolically coupled elongation process in which peptide and polyketide chemistries are integrated rather than sequentially separated (Cane et al., 1998; Fischbach & Walsh, 2006).
In parallel, the terpene-associated gene BRLA_RS11365, encoding a squalene–hopene cyclase, is predicted to generate cyclic terpenoid scaffolds. Although traditionally considered an independent pathway, in the context of Region 8 it is plausible that terpene-derived moieties function as structural appendages or hydrophobic anchors, either introduced during early prenylation events or appended during late-stage tailoring reactions (Yamada et al., 2015). Collectively, these observations suggest that peptide, polyketide, and terpene biosynthesis may converge into a single hybrid molecular architecture.
- Stepwise Mechanistic Model of Region 8 Biosynthesis
- Step 1: Peptide initiation by NRPS modules
NRPS enzymes are predicted to initiate biosynthesis by activating amino acid substrates through adenylation domains and assembling peptide intermediates through condensation and thiolation cycles (Fischbach & Walsh, 2006).
- Step 2: Polyketide incorporation via T1PKS machinery
The T1PKS enzyme BRLA_RS11400 is predicted to participate in chain extension or modification reactions through KS-, AT-, and KR-associated catalytic activities. The presence of a putative NRPS/PKS hybrid enzyme (BRLA_RS11475) suggests possible interactions between peptide- and polyketide-associated biosynthetic modules. The presence of a hybrid NRPS/PKS enzyme (BRLA_RS11475) suggests direct crosstalk between peptide and polyketide modules, enabling formation of chimeric peptide–polyketide backbones (Cane et al., 1998).
- Step 3: Terpene-derived structural diversification
BRLA_RS11365 is annotated as a terpene-associated cyclase and may participate in the biosynthesis of terpene-derived structural components within the cluster. The precise relationship between terpene biosynthesis and the NRPS–PKS-associated modules remains to be experimentally determined. This step introduces hydrophobic or rigid ring structures, potentially enhancing membrane interaction or biological activity (Yamada et al., 2015).
- Step 4: Tailoring modifications
Accessory enzymes including oxidoreductases, methyltransferases, and cyclases (Table 3) are expected to introduce post-assembly modifications such as oxidation, methylation, and cyclization. These reactions likely diversify the final metabolite pool and enhance structural complexity (Rutledge & Challis, 2015).
- Step 5: Transport and regulatory coordination
The presence of MFS and ABC transporters, together with Crp/Fnr-family regulators, indicates that biosynthesis is coupled to export and transcriptional regulation. This ensures metabolite secretion and prevents intracellular toxicity, consistent with fully functional BGC architectures (Kautsar et al., 2020).
- Step 6: Final metabolite assembly
The integrated activity of NRPS, T1PKS, and terpene modules, followed by tailoring reactions, is hypothesized to produce a structurally complex hybrid metabolite featuring peptide, polyketide, and terpenoid elements. Such molecular architectures are typically associated with enhanced chemical novelty and multifunctional bioactivity (Chevrette et al., 2020; Rudolf et al., 2021).
Although this mechanistic model is supported by genomic evidence derived from cluster organization, functional gene annotation, and domain architecture analyses (Table 2, Table 3 and Table 4 and Figure 2, Figure 3 and Figure 4), it remains a predictive framework requiring experimental validation, as genome mining alone cannot establish metabolite production, pathway activity, or molecular structure (Rutledge & Challis, 2015; Blin et al., 2021). Integrative approaches combining transcriptomics, proteomics, and metabolomics will therefore be necessary to verify pathway expression and identify the associated metabolites. Furthermore, structural prediction tools such as AlphaFold may improve our understanding of enzyme architecture and substrate specificity, thereby facilitating rational pathway engineering and heterologous expression studies (Jumper et al., 2021; Varadi et al., 2022).
4.2. Biosynthetic Significance of Region 8
The identification of Region 8 as a 133.5-kb hybrid NRPS–T1PKS–terpene biosynthetic gene cluster highlights the remarkable biosynthetic complexity encoded within the genome of Brevibacillus laterosporus SA14. Among the 20 biosynthetic regions identified, Region 8 represents the largest predicted biosynthetic locus and is distinguished by the coexistence of NRPS-, T1PKS-, and terpene-associated biosynthetic features within a single contiguous genomic region. Such integrated architectures are increasingly recognized as important drivers of natural-product diversification because they combine distinct biosynthetic mechanisms capable of generating structurally complex metabolites (Chevrette et al., 2020; Rudolf et al., 2021).
The organization of Region 8 further suggests a high degree of biosynthetic integration. The presence of multiple core biosynthetic genes together with regulatory elements, transport-associated proteins, and accessory enzymes indicates that the cluster may function as a coordinated biosynthetic unit rather than a collection of independent pathways. Similar large hybrid biosynthetic systems have been associated with expanded chemical diversity and increased opportunities for evolutionary innovation through the recombination of peptide, polyketide, and terpenoid biosynthetic strategies (Fischbach & Walsh, 2006; Chevrette et al., 2020).
Notably, comparison against the MIBiG database did not reveal a closely matching characterized biosynthetic gene cluster, suggesting that Region 8 may represent a previously uncharacterized biosynthetic architecture. Although the metabolite(s) encoded by this region remain unknown, its genomic organization, large size, and hybrid composition collectively identify Region 8 as a promising target for future transcriptomic, metabolomic, and functional investigations aimed at discovering novel secondary metabolites and elucidating their biosynthetic pathways (Kautsar et al., 2020; Blin et al., 2021).
4.3. Ecological and Functional Implications
The extensive repertoire of biosynthetic gene clusters identified in B. laterosporus SA14 may reflect adaptation to complex microbial environments where competition, resource acquisition, and ecological interactions influence microbial fitness. The predominance of NRPS-containing clusters together with multiple hybrid biosynthetic systems suggests substantial genetic capacity for the production of structurally diverse secondary metabolites, many of which may contribute to ecological functions such as microbial interactions and environmental adaptation (Fischbach & Walsh, 2006; Chevrette et al., 2020).
Region 8 is of particular interest because it integrates NRPS-, T1PKS-, and terpene-associated biosynthetic features within a single genomic locus. The coexistence of these biosynthetic systems may provide opportunities for the generation of structurally complex metabolites through coordinated biosynthetic processes. Similar hybrid biosynthetic architectures have been proposed to contribute to chemical diversification and ecological versatility in microorganisms (Rudolf et al., 2021; Chevrette et al., 2020).
The presence of transporter genes, regulatory elements, and accessory enzymes within Region 8 further suggests that the cluster may function as an integrated biosynthetic unit. Such auxiliary components are commonly associated with pathway regulation, metabolite export, and cellular protection mechanisms in secondary metabolite biosynthetic gene clusters (Kautsar et al., 2020; Medema et al., 2011). However, the biological roles and ecological significance of metabolites potentially encoded by Region 8 remain speculative and require experimental verification.
Taken together, these observations suggest that the hybrid biosynthetic organization observed in SA14 may represent an evolutionary strategy for expanding biosynthetic diversity and enhancing ecological adaptability. Future transcriptomic, metabolomic, and functional studies will be necessary to determine the ecological roles and biological activities of metabolites associated with this biosynthetic region.
4.4. Biotechnological and Drug Discovery Potential
From an applied perspective, the identification of the hybrid NRPS–T1PKS–terpene biosynthetic gene cluster in B. laterosporus SA14 highlights the potential value of this strain as a source of biosynthetic diversity for natural product research. Hybrid biosynthetic systems are increasingly recognized as important reservoirs of chemical innovation because they combine distinct biosynthetic mechanisms capable of generating structurally diverse secondary metabolites (Rutledge & Challis, 2015; Rudolf et al., 2021). The integration of multiple biosynthetic features within Region 8, together with associated regulatory and transporter elements, identifies this locus as a promising target for future functional characterization, heterologous expression, and synthetic biology-based investigations (Medema & Fischbach, 2015).
In addition, the combination of genome mining, biosynthetic pathway reconstruction, and structural prediction approaches provides a useful framework for exploring enzyme function and biosynthetic organization. Structural modeling tools such as AlphaFold may contribute to improved understanding of enzyme architecture and domain organization, thereby supporting future studies of pathway engineering and heterologous production systems (Jumper et al., 2021; Varadi et al., 2022).
The apparent novelty of Region 8, reflected by the absence of a closely matching characterized biosynthetic gene cluster in the MIBiG database, further increases its relevance for natural product discovery. Although the chemical products encoded by this region remain unknown, its large size, hybrid organization, and extensive biosynthetic repertoire suggest that it may represent a valuable target for future metabolomic and biochemical investigations. Collectively, these findings highlight the utility of genome mining as a strategy for identifying previously unexplored biosynthetic resources and prioritizing candidate gene clusters for downstream experimental validation (Medema & Fischbach, 2015; Rudolf et al., 2021).
- Study Limitations
This study was based exclusively on computational genome mining and biosynthetic gene cluster prediction. Consequently, the presence of a biosynthetic gene cluster does not necessarily indicate active expression, metabolite production, or biological activity under laboratory conditions. Furthermore, biosynthetic functions were inferred from domain architecture and sequence-based annotations generated by antiSMASH and therefore remain predictive. Experimental validation using transcriptomic, proteomic, metabolomic, biochemical, and genetic approaches will be necessary to confirm pathway activity and characterize the metabolites associated with Region 8.
5. Conclusion
Genome mining of Brevibacillus laterosporus SA14 (TISTR 2453) revealed a diverse biosynthetic landscape comprising 20 biosynthetic regions and 29 predicted protoclusters, indicating substantial genetic potential for secondary metabolite biosynthesis. Among these regions, Region 8 was identified as the largest biosynthetic locus and was predicted to encode a hybrid NRPS–T1PKS–terpene biosynthetic gene cluster spanning approximately 133.5 kb and containing more than 60 coding sequences.
The coexistence of peptide-, polyketide-, and terpene-associated biosynthetic features within a single genomic region, together with transporter and regulatory elements, suggests a highly integrated biosynthetic organization. Comparative analysis against the MIBiG database did not identify a closely matching characterized biosynthetic gene cluster, indicating that Region 8 may represent a previously uncharacterized biosynthetic architecture.
Although the biological products and functional roles associated with this cluster remain to be experimentally verified, its genomic organization and predicted biosynthetic composition identify Region 8 as a promising candidate for future transcriptomic, metabolomic, biochemical, and synthetic biology investigations. Collectively, these findings demonstrate the utility of genome mining for prioritizing candidate biosynthetic gene clusters and provide a foundation for future exploration of secondary metabolism in B. laterosporus SA14.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, methodology, formal analysis, data interpretation, visualization, and writing—original draft preparation were performed by Monthon Lertcanawanichakul. Husna Madoromae contributed to genome mining analysis, antiSMASH annotation, and figure preparation. Komkrit Auesukool provided conceptual support and critical revision of the bioinformatics analysis. Nuttapon Songnaka contributed to genomic data processing and database comparison. All authors contributed to manuscript review, provided intellectual input, and approved the final version of the manuscript.
Funding
This research was supported by the Plant Genetic Conservation Project under the Royal Initiative of Her Royal Highness Princess Maha Chakri Sirindhorn, Thailand. (RSPG-WU-12/2567, RSPG-WU-20/2569).
Ethics Approval and Consent to Participate
Not applicable. This study was based entirely on in silico genome mining and publicly or previously obtained genomic sequence data. No human participants, animal experiments, or clinical samples were involved in this research.
Consent for Publication
Not applicable. No identifiable human data, images, or clinical information are included in this manuscript.
AI Declaration
The authors acknowledge the use of artificial intelligence tools for language refinement, logical consistency checking, and manuscript structuring. The AI tool was not used to generate or modify experimental data, results, or scientific conclusions. All analyses, interpretations, and final decisions were performed and verified by the authors.
Availability of Data and Materials
The genome sequence data and antiSMASH analysis outputs generated and analyzed during the current study are available from the corresponding author upon reasonable request. All computational analyses were performed using publicly accessible bioinformatics resources, including the antiSMASH platform and the MIBiG database. Nuttapon Songnaka contributed to genomic data processing and database-supported comparative analyses.
Acknowledgments
The authors gratefully acknowledge the antiSMASH online platform for facilitating genome mining and biosynthetic gene cluster analyses. The authors also appreciate the availability of public genomic resources and biosynthetic gene cluster repositories, including MIBiG, which supported comparative analyses and functional annotation of the identified biosynthetic gene clusters.
Competing Interests
The authors declare that they have no competing interests.
Abbreviations
| NRPS | Non-ribosomal peptide synthetase |
| T1PKS | Type I polyketide synthase |
| PKS | Polyketide synthase |
| RiPP | Ribosomally synthesized and post-translationally modified peptide |
| BGC | Biosynthetic gene cluster |
| antiSMASH | antibiotics & Secondary Metabolite Analysis Shell |
| MIBiG | Minimum Information about a Biosynthetic Gene Cluster |
| CDS | Coding sequence |
| KS | Ketosynthase |
| AT | Acyltransferase |
| KR | Ketoreductase |
| MFS | Major facilitator superfamily |
| ABC transporter | ATP-binding cassette transporter |
References
- Alanjary, M.; Kronmiller, B.; Adamek, M.; Blin, K.; Weber, T.; Medema, M. H. ARTS: Antibiotic resistant target seeker. Nucleic Acids Res. 2017, 45(W1), W42–W48. [Google Scholar] [CrossRef]
- Bentley, S. D.; Chater, K. F.; Cerdeño-Tárraga, A. M.; Thomson, N. R.; James, K. D.; Harris, D. E.; Quail, M. A.; Kieser, H.; Harper, D.; Bateman, A.; Brown, S.; Chandra, G.; Chen, C. W.; Collins, M.; Cronin, A.; Fraser, A.; Goble, A.; Hidalgo, J.; Hornsby, T.; Howarth, S.; Huang, C. H.; Kieser, T.; Larke, L.; Murphy, L.; Oliver, K.; O’Neil, S.; Rabbinowitsch, E.; Rajandream, M. A.; Rutherford, K.; Rutter, S.; Seeger, K.; Saunders, D.; Sharp, S.; Squares, R.; Squares, S.; Taylor, K.; Warren, T.; Wietzorrek, A.; Woodward, J.; Barrell, B. G.; Parkhill, J.; Hopwood, D. A. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417(6885), 141–147. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A. M.; Charlop-Powers, Z.; van Wezel, G. P.; Medema, M. H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49(W1), W29–W35. [Google Scholar] [CrossRef]
- Cane, D. E.; Walsh, C. T.; Khosla, C. Harnessing the biosynthetic code: Combinations, permutations, and mutations. Science 1998, 282(5386), 63–68. [Google Scholar] [CrossRef]
- Chevrette, M. G.; Gutiérrez-García, K.; Selem-Mojica, N.; et al. Evolutionary dynamics of natural product biosynthesis in bacteria. Nat. Commun. 2020, 11, 2452. [Google Scholar] [CrossRef] [PubMed]
- Covington, B. C.; et al. Biosynthetic logic of microbial secondary metabolism. Annu. Rev. Biochem. 2021, 90, 763–788. [Google Scholar] [CrossRef] [PubMed]
- Favret, M. E.; Yousten, A. A. Characteristics of a mosquito-active toxin produced by Bacillus laterosporus. J. Invertebr. Pathol. 1989, 53(2), 206–216. [Google Scholar] [CrossRef]
- Fischbach, M. A.; Walsh, C. T. Assembly-line enzymology for polyketide and nonribosomal peptide antibiotics. Chem. Rev. 2006, 106(8), 3468–3496. [Google Scholar] [CrossRef]
- Fischbach, M. A.; Rasmussen, S. A.; Clardy, J. A conserved gene cluster for secondary metabolite biosynthesis in bacteria. Proc. Natl. Acad. Sci. 2008, 105(14), 4601–4606. [Google Scholar] [CrossRef]
- Helfrich, E. J. N.; Lin, G.-M.; Voigt, C. A.; Clardy, J. Natural product biosynthesis. Beilstein J. Org. Chem. 15 2019, 2889–2906. [Google Scholar] [CrossRef]
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. 2009, 48(26), 4688–4716. [Google Scholar] [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D. L. Plant-associated rhizobacteria for biocontrol and plant growth enhancement. Front. Plant Sci. 2016, 7, 1237. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; Bridgland, A.; Meyer, C.; Kohl, S. A. A.; Ballard, A. J.; Cowie, A.; Romera-Paredes, B.; Nikolov, S.; Jain, R.; Adler, J.; Hassabis, D. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596(7873), 583–589. [Google Scholar] [CrossRef] [PubMed]
- Kautsar, S. A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J. C.; Terlouw, B. R.; van der Hooft, J. J. J.; van Santen, J. A.; Tracanna, V.; Suárez Duran, H. G.; Pascal Andreu, V.; Selem-Mojica, N.; Alanjary, M.; Robinson, S. L.; van Wezel, G. P.; Medema, M. H. MIBiG 2.0: A repository for biosynthetic gene clusters of known function. Nucleic Acids Res. 2020, 48(D1), D454–D458. [Google Scholar] [CrossRef]
- Medema, M. H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M. A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters. Nucleic Acids Res. 2011, 39 (suppl_2), W339–W346. [Google Scholar] [CrossRef] [PubMed]
- Medema, M. H.; Fischbach, M. A. Computational approaches to natural product discovery. Nat. Chem. Biol. 2015, 11(9), 639–648. [Google Scholar] [CrossRef]
- Navarro-Muñoz, J. C.; Selem-Mojica, N.; Mullowney, M. W.; et al. A computational framework for systematic exploration of biosynthetic diversity. Nat. Chem. Biol. 2020, 16, 60–68. [Google Scholar] [CrossRef]
- Rudolf, J. D.; Chang, C. Y.; Ma, M.; Shen, B. Natural product biosynthesis. Nat. Product. Rep. 2021, 38(5), 905–980. [Google Scholar] [CrossRef]
- Ruiu, L. Brevibacillus laterosporus, a pathogen of invertebrates and a broad-spectrum antimicrobial species. Insects 2013, 4(3), 476–492. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, P. J.; Challis, G. L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13(8), 509–523. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; Žídek, A.; Green, T.; Tunyasuvunakool, K.; Petersen, S.; Jumper, J.; Clancy, E.; Green, R.; Vora, A.; Lutfi, M.; Figurnov, M.; Cowie, A.; Hobbs, N.; Kohli, P.; Kleywegt, G. J.; Birney, E.; Hassabis, D.; Velankar, S. AlphaFold protein structure database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50(D1), D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Kuzuyama, T.; Komatsu, M.; Shin-Ya, K.; Omura, S.; Cane, D. E.; Ikeda, H. Terpene synthases are widely distributed in bacteria. Proceedings of the National Academy of Sciences of the United States of America 2015, 112(3), 857–862. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Distribution of biosynthetic gene cluster (BGC) classes identified in the genome of Brevibacillus laterosporus SA14 using antiSMASH analysis. NRPS-containing clusters represented the most abundant biosynthetic category, indicating substantial potential for secondary metabolite production.
Figure 1.
Distribution of biosynthetic gene cluster (BGC) classes identified in the genome of Brevibacillus laterosporus SA14 using antiSMASH analysis. NRPS-containing clusters represented the most abundant biosynthetic category, indicating substantial potential for secondary metabolite production.

Figure 2.
Genomic organization of Region 8 identified in B. laterosporus SA14. The 133.5-kb hybrid biosynthetic gene cluster contains NRPS-, T1PKS-, and terpene-associated biosynthetic features together with accessory transporters and regulatory genes.
Figure 2.
Genomic organization of Region 8 identified in B. laterosporus SA14. The 133.5-kb hybrid biosynthetic gene cluster contains NRPS-, T1PKS-, and terpene-associated biosynthetic features together with accessory transporters and regulatory genes.

Figure 3.
Hypothetical biosynthetic model for the hybrid NRPS–T1PKS–terpene gene cluster identified in B. laterosporus SA14.
Figure 3.
Hypothetical biosynthetic model for the hybrid NRPS–T1PKS–terpene gene cluster identified in B. laterosporus SA14.

Figure 4.
Predicted domain architecture of representative core biosynthetic enzymes encoded within Region 8 of *Brevibacillus laterosporus* SA14. Representative terpene-associated, NRPS, T1PKS, and hybrid NRPS–PKS enzymes are shown with their predicted catalytic domains based on antiSMASH annotation. The coexistence of peptide, polyketide, and terpene biosynthetic modules supports the hypothesis that Region 8 encodes a structurally complex hybrid secondary metabolite biosynthetic pathway.
Figure 4.
Predicted domain architecture of representative core biosynthetic enzymes encoded within Region 8 of *Brevibacillus laterosporus* SA14. Representative terpene-associated, NRPS, T1PKS, and hybrid NRPS–PKS enzymes are shown with their predicted catalytic domains based on antiSMASH annotation. The coexistence of peptide, polyketide, and terpene biosynthetic modules supports the hypothesis that Region 8 encodes a structurally complex hybrid secondary metabolite biosynthetic pathway.


Table 1.
Summary of Biosynthetic Gene Clusters Identified in B. laterosporus SA14.
| Region | Product Type | Size (kb) |
| 1 | RRE-containing | 21.2 |
| 2 | NI-siderophore | 31.7 |
| 3 | NRPS-like, NRPS | 94.9 |
| 4 | Terpene precursor, NRPS-like | 53.5 |
| 5 | NRPS | 51.1 |
| 6 | transAT-PKS | 66.5 |
| 7 | Ranthipeptide | 21.6 |
| 8 | NRPS, Terpene, T1PKS | 133.5 |
| 9 | NRPS, transAT-PKS | 71.6 |
| 10 | NRPS | 81.6 |
| 11 | NRPS, RiPP-like | 97.7 |
| 12 | NRPS, transAT-PKS | 97.7 |
| 13 | T3PKS | 41.1 |
| 14 | NRPS | 47.0 |
| 15 | Cyclic lactone autoinducer | 20.5 |
| 16 | Phosphonate | 13.2 |
| 17 | NRPS, NRP-metallophore | 98.6 |
| 18 | NRPS | 47.6 |
| 19 | RiPP-like | 10.8 |
| 20 | Azole-containing RiPP | 23.5 |
Table 2.
Characteristics of Region 8.
| Feature | Value |
| Cluster type | Hybrid NRPS–T1PKS–Terpene |
| Start position | 2,467,610 |
| End position | 2,601,073 |
| Length | 133.5 kb |
| Number of CDS | >60 |
| Biosynthetic systems | NRPS, T1PKS, Terpene |
| MIBiG similarity | No closely matching characterized cluster identified |
Table 3.
Predicted Functional Composition of the Hybrid NRPS–T1PKS–Terpene Biosynthetic Gene Cluster.
Table 3.
Predicted Functional Composition of the Hybrid NRPS–T1PKS–Terpene Biosynthetic Gene Cluster.
| Gene Category | Predicted Function | Occurrence in Region 8 |
| NRPS biosynthetic genes | Peptide assembly | Present |
| T1PKS biosynthetic genes | Polyketide chain elongation | Present |
| Terpene-associated genes | Terpene biosynthesis and modification | Present |
| Transporter genes | Export of metabolites | Present |
| Regulatory genes | Pathway regulation | Present |
| Hypothetical proteins | Unknown functions | Present |
| Accessory enzymes | Tailoring and modification reactions | Present |
Table 4.
Candidate core biosynthetic genes identified within Region 8 of Brevibacillus laterosporus SA14.
Table 4.
Candidate core biosynthetic genes identified within Region 8 of Brevibacillus laterosporus SA14.
| Gene ID | Predicted Product | Biosynthetic Category | Putative Function |
| BRLA_RS11365 | Squalene–hopene cyclase | Terpene | Terpene cyclization |
| BRLA_RS11395 | Non-ribosomal peptide synthetase | NRPS | Peptide assembly |
| BRLA_RS11400 | Type I polyketide synthase | T1PKS | Polyketide chain elongation |
| BRLA_RS11410 | Non-ribosomal peptide synthetase | NRPS | Peptide assembly |
| BRLA_RS11475 | NRPS/PKS hybrid enzyme | Hybrid NRPS–PKS | Integrated biosynthesis |
| BRLA_RS11525 | Non-ribosomal peptide synthetase | NRPS | Peptide assembly |
| BRLA_RS11530 | Non-ribosomal peptide synthetase | NRPS | Peptide assembly |
| BRLA_RS11535 | Non-ribosomal peptide synthetase | NRPS | Peptide assembly |
NRPS, non-ribosomal peptide synthetase; T1PKS, type I polyketide synthase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



