Submitted:
30 May 2026
Posted:
02 June 2026
You are already at the latest version
Abstract
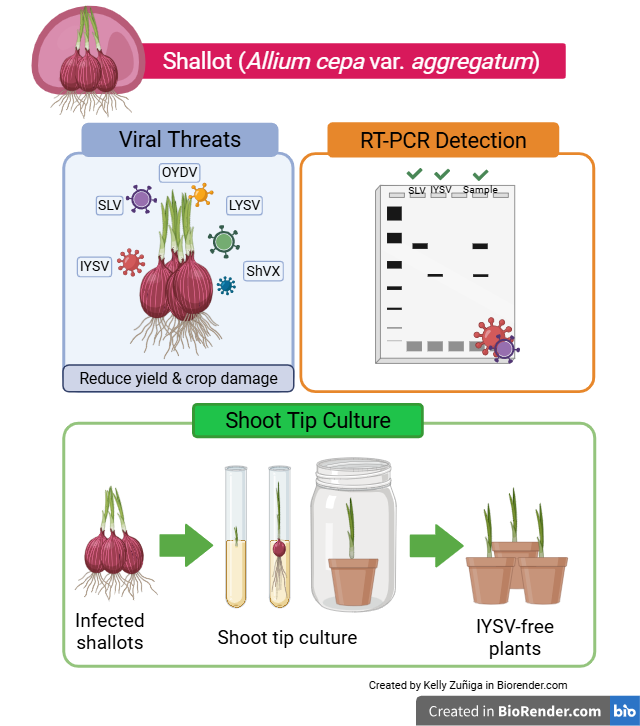
Shallot (Allium cepa var. aggregatum) is an annual herbaceous plant of the Amaryllidaceae family cultivated worldwide for its gastronomic importance, unique flavor, and nutri-tional properties. In Ecuador, shallot is considered an emerging crop; nevertheless, the lack of effective detection and treatment options for phytoviruses remains a challenge for farmers. These viruses negatively impact crop yield, leading to substantial economic losses and declining crop quality. Simultaneous infections can synergistically exacerbate crop damage. In this study, the molecular detection of the five most common shallot vi-ruses was explored: Shallot latent virus (SLV), Onion yellow dwarf virus (OYDV), Shallot virus X (ShVX), Leek yellow stripe virus (LYSV), and Iris yellow spot virus (IYSV) in shallots from different suppliers of markets in Quito, Ecuador. We identified two viruses using RT-PCR: SLV (latent) and IYSV (responsible for severe disease symptoms). Shoot tip culture produced shallot plants completely free of IYSV, while SLV persisted in the regenerated plants. This study constitutes the first report of IYSV in shallots in the coun-try and shows that shoot tip culture can serve as a partial cleaning method to improve crop quality and support more effective pathogen management and control.
Keywords:
Allium cepa var. aggregatum
; shallot latent virus
; iris yellow spot virus
; molecular detection
; shoot tips
; in vitro culture
1. Introduction
Shallot (Allium cepa var. aggregatum), a distinct variety within the Allium cepa species commonly known as onions, is an annual herbaceous plant of the family Amaryllidaceae [1]. It is widely cultivated across the Americas, Europe, and Asia [2]. This variety, known for its vegetative reproduction via bulbs, is distinguished by its extended storage life, color, and bioactive compounds [3,4]. Shallots are well known in the culinary world as an essential ingredient in various dishes due to their unique flavor and nutritional properties [5]. They are a rich source of valuable phytochemicals, including organosulfur compounds, fructans and flavonoids [1,6]. Nevertheless, they are highly susceptible to pests and diseases, with viral infections being one of the most severe threats to yield and quality [7,8]. Viruses account for nearly half of emerging infectious diseases in various crops and are responsible for approximately 40% of total crop losses [9]. Several economically important viral pathogens belong to the Allexivirus, Carlavirus, and Potyvirus genera, which frequently lead to simultaneous infections that can further exacerbate yield reduction and crop losses [7]. The development and severity of these viral diseases depend on several factors, including the plant’s age, environmental conditions, and the vector dynamics [10].
Viral pathogens can usually be detected using serological tests, bioassays, or DNA- and RNA-based molecular techniques [11]. Bioassays involve grafting plant material or mechanical inoculation onto a viroid indicator host, which typically develops symptoms over a prolonged period of time [12]. Serological methods are based on the specific interaction between viral antigens and the plant’s antibodies. Nevertheless, the production of highly specific antisera against Allium plants is troublesome due to coinfections [13,14]. Monoclonal antibodies have been utilized to overcome this problem and differentiate between strains of potyviruses and carlaviruses by using the variability of the N-terminal viral coat protein (CP) reaction [15,16]. Although serology has been the main method for routine testing, cross-reactions can occur, and many host plants contain phenolic substances that precipitate proteins, complicating the isolation of active viruses [15,17]. In this way, molecular methods for viral nucleic acids have been optimized and developed for large-scale virus testing of shallot leaves and bulbs. Specific and accessible molecular assay development, such as Polymerase Chain Reaction (PCR) and Reverse Transcription PCR (RT-PCR), allows a specific, versatile, and sensitive diagnosis of viruses [18,19]. This methodology involves primer design based on specific conserved sequences of viral genetic material, which leads to a wide range of detection levels and specificity [7,20].
While such diagnostic tools are essential for early detection, controlling viral infection in plants requires a comprehensive approach. One widely use strategy for propagating virus-free plants in Allium crops is through meristem or shoot tip culture [21,22,23]. This method is particularly effective due to its high genetic stability and regeneration potential [7,11,21,24,25,26,27]. Shoot tip culture is a type of meristem culture in which the explant consists of the shoot tip, including the apical meristem and a small number of surrounding leaf primordia [28]. Notably, this method has been reported as an efficient tool for obtaining virus-free plants while maintaining a high regeneration rate [28,29]. For example, in Allium species, virus elimination efficiencies of approximately 50–70% and even higher have been reported depending on the virus [21,30,31]. Beyond in vitro techniques, other virus management strategies are also considered, such as the use of certified virus-free seeds, control of arthropod vectors, use of insecticides, proper field management, removal of infected plants, usage of resistant varieties, virus management during post-harvest, crop rotation, surveillance, and monitoring [7,32,33,34,35].
In Ecuador, shallots are considered an emerging crop [36]. However, the lack of effective detection and treatment options for viral infections remains a significant challenge for farmers [9]. The viral detection and its elimination are essential for sustainable crop production through vegetative propagation strategies [9,11]. The objectives of this study are (1) to molecularly detect the following five most common viruses that infect shallots: shallot latent virus (SLV), onion yellow dwarf virus (OYDV), shallot virus X (ShVX), leek yellow streak virus (LYSV), and iris yellow spot virus (IYSV) [7,37], and (2) to evaluate the effectiveness of shoot tip culture in eliminating viruses present in shallots (Allium cepa var. aggregatum) and thus propagating virus-free plants.
2. Materials and Methods
2.1. Design and Selection of Primers
Five of the most common viruses affecting shallots were selected for molecular detection by RT-PCR: OYDV, SLV, ShVX, IYSV, and LYSV. To begin the process, nucleotide sequences of the target genes were obtained from the NCBI. The target sites selected were the coat protein for SLV, OYDV, and LYSV, and the replicase and nucleocapsid genes for ShVX and IYSV, respectively. Sequences were downloaded in FASTA format and aligned in UGENE. Primers were designed using Primer3web [38] and validated with PrimerBLAST [39] and MFEprimer [40]. Because sequence variations existed, degenerate primers were created. Two types of primers were tested: primers designed in this study and those previously reported in the literature (Table S1). Based on the analysis of the PCR results from pilot shallot samples, literature-based primers were selected for SLV and OYDV, while designed primers were chosen for ShVX, IYSV, and LYSV for molecular analysis (Table S2 and Figure S1-S2).
2.2. Synthetic DNA-Based Positive Controls for RT-PCR Detection of Plant Viruses
The target sequences for each virus (nucleocapsid for IYSV, replicase for ShVX, and coat protein for SLV, OYDV, and LYSV) were obtained from NCBI. Small fragments between 269 and 625 bp of the target sequences were selected for cloning into an ampicillin plasmid vector (pMG-Amp) and evaluated through an in silico analysis. The expected sizes of PCR products were SLV (308 bp), OYDV (625 bp), ShVX (160 bp), IYSV (189 bp), and LYSV (191 bp) (Figure S1).
2.3. Plant Material for Viral Detection and In Vitro Culture
A total of 99 shallot plants (Allium cepa var. aggregatum) were obtained from three different suppliers based on their availability: 23 from the Experimental Farm of Universidad San Francisco de Quito (EF), 26 from the Organic market (OM), and 25 from each supplier at the Iñaquito market in Quito (Iq1 and Iq2) (Figure 1). The plants were transported to the Plant Biotechnology Laboratory at USFQ. Bulbs were separated from the leaves and stored at room temperature (18 °C) for in vitro culture. Meanwhile, the leaves were stored at –20 °C for 24 hours and then at –80 °C for viral detection.
2.4. RNA Extraction from Shallot Leaves Pre- and Post-Culture
Virus detection was carried out both before and after in vitro culture. Pre-culture detection aimed to identify the viruses present in the original material, while post- in vitro culture detection assessed the effectiveness of shoot tip culture in eliminating them. Before in vitro culture, RNA was extracted from all the 99 samples of shallot leaves. After the shoot tip culture, only 31 plantlets were available for molecular analysis: 11 from EF (Assay 1), 8 from Iq1 (Assay 2), and 12 from Iq2 (Assay 3). Total RNA was extracted from leaves using the TRIzol-chloroform protocol as described by Rio et al. [41] with approximately 200–300 mg of shallot leaf powder per sample. RNA quantity and purity, as well as integrity, were checked by Nanodrop (Thermo Scientific) and Qubit (Invitrogen). The RNA was left at 4 °C for 24 hours and then stored at -20 °C.
2.5. RT-PCR
Complementary DNA (cDNA) was synthesized using the SuperScript III Reverse Transcriptase (Invitrogen) and random hexamer primers (Thermo Fisher Scientific), following the manufacturer’s instructions. PCR was then performed using virus-specific primers, along with synthetic positive and negative controls. The PCR cycling conditions were 94 °C for 2 min, followed by 30 cycles of 94 °C for 30 s, 58 °C (OYDV, LYSV), 47 °C (SLV), 61.5 °C (ShVX), and 62 °C (IYSV) for 30 s, 72 °C for 1 min, and finally an extension of 72 °C for 3 min. RT-PCR products were visualized in a 1.5% agarose gel, stained with SYBR Safe, and examined under UV light using a photo-documenter (BIO-RAD).
2.6. Bulb Sterilization
The protocol of in vitro culture used in this study was standardized and taken from previously developed protocols [36,42], with some modifications introduced as detailed in Table S3 and Figure S3. A total of 73 fresh shallot bulb samples were analyzed: 23 from EF (Assay 1), 25 from Eq1 (Assay 2), and 25 from Eq2 (Assay 3). The OM samples were excluded from the in vitro assays due to fungal contamination. The disinfection protocol began with a 10-minute pre-wash of the bulbs in tap water to remove soil residues and eliminate dry outer layers. The bulbs were then immersed in Mertect fungicide (Syngenta; 3.5 g/L) and agitated at 420 rpm for 15 minutes.
Afterwards, in a laminar flow chamber (LABCONCO Purifier Clean Bench), bulbs were immersed in ethanol (70%) for 10 minutes. Then the bulbs were submerged in sodium hypochlorite (4.5%) with three drops of Tween 20 and agitated for 35 minutes. The bulbs were then rinsed with sterile distilled water. One-third of the length of the bulb was excised with a sterile scalpel, retaining the basal portion and removing external cataphylls to obtain a basal disc explant of about 1.5 cm in diameter and 1 cm in height. Each explant was cultured on M1 initial medium in a glass bottle (Table S4).
2.7. Shoot Tip Culture
After 15 days in culture, plantlets regenerated from basal discs and shoot tips (6 to 10 mm long) were excised. Shoot tip refers to the apical portion of the plant shoot, which includes the apical meristem (0.1 to 0.5 mm) and surrounding leaf primordia [28,43,44]. The excised shoot tips were cultured on M1 regeneration medium to induce the formation of plantlets (Figure 2). After one month of shoot tip culture, young leaves were collected from each plantlet for the post-culture viral detection. All plantlets were subcultured on M2 rooting medium (Table S4). After four weeks, when plantlets developed roots, they were transferred to M3 medium and then subcultured every 30 days (Table S4). All plant material was kept at 25 ± 1 °C with a photoperiod of 16 hours in white light.
2.8. Acclimatization
When plants showed roots and bulbs, acclimatization took place. First, shallot plants were carefully removed from the 25 mL culture tubes, keeping the plants and their roots intact, and then rinsed with distilled water to remove the residual agar. They were then transferred to clay plots with autoclaved black soil inside 1,900-mL glass jars. Each jar was covered with plastic wrap and maintained at 25 °C with a 16-hour photoperiod. Twice a week, plants were watered with sterile distilled water, and two holes were made in the plastic wrap. After one month, the plastic was completely removed. All plants received Hoagland fertilizer [45] every 15 days.
2.9. Data Collection and Statistical Analysis
The efficiency of the disinfection protocol was assessed by measuring the percentage of sterility after 15 days of culture. We also determined the sprouting rate of basal discs after 15 days of culture, as well as the percentage of shoot tip explants that successfully regenerated into plants. The efficiency of the in vitro culture system was evaluated over a 360-day period based on sprouting response, plantlet development, rooting capacity, and bulb formation. Differences in sterility, plantlet growth, rooting, and bulb formation were analyzed using Chi-squared test (χ2). One-way Analysis of Variance (ANOVA) was used to determine differences in the number of shoot tips excised per assay. Two-way ANOVA was applied to evaluate differences in rooting and bulb formation over the time and by assay. All statistical analyses were conducted using GraphPad Prism version 8.0.1 (GraphPad Software, San Diego, CA, USA), and statistical significance was established at p < 0.05. Additionally, the effectiveness of the elimination of viruses was determined by the presence or absence of them after shoot tip culture.
3. Results
3.1. Viral Detection
3.1.1. RNA Extraction Before and After Shoot Tip Culture
Total RNA extracted from the 99 shallot leaf samples before in vitro culture showed concentration averages that varied between 261.4 and 552.2 ng/µL (Table S5). After in vitro culture, the average RNA concentration from the 31 leaf samples of regenerated shallot plantlets ranged from 691.3 to 910.5 ng/µL (Table S6).
3.1.2. Virus Detection Before Shoot Tip Culture
From the 99 plants evaluated before in vitro culture, a total of 87 samples were positive for SLV, 29 samples were positive for IYSV, 27 samples presented both viruses (co-infection), and 10 samples were negative for both viruses. Of the 27 samples coinfected, 72% belonged to Iq2 and 16% to Iq1 (Table 1). All samples analyzed tested negative for OYDV, ShVX, and LYSV viruses (Table 1).
3.2. Disinfection of Plant Material
The disinfection protocol of basal discs resulted in a sterility rate of 47.8% for Assay 1, 32% for Assays 2 and 48% for Assay 3 (Table 2). Statistical analysis showed no significant difference in the sterility of the basal discs among the three assays (Chi-square test, p = 0.7899), indicating the reproducibility of the sterilization process.
3.3. Shoot Regeneration from Basal Discs
Sprouting of basal discs was 100% in all assays and controls after 15 days of culture.
3.4. Shoot Tip In Vitro Culture
The mean of shoot tips obtained per basal disc was 6.5 for Assay 1, 5.5 for Assay 2, and 4.8 for Assay 3 (Figure S4). These differences were not statistically significant (one-way ANOVA, p = 0.2466). The percentage of plantlet development from shoot tips was highest in Assay 1 (88.7%) and lowest in Assay 2 (59.1%) (Table 3), with the difference being statistically significant (Chi-square test, p = 0.0084). In all assays, the regenerated plantlets developed into complete plants and successfully rooted (Figure 3A). Regarding bulb formation, it was faster in Assay 1, in which plants began to develop bulbs after 90 days of culture. In contrast, bulb development in Assays 2 and 3 was observed only after 210 days. By 300 days of culture, 100% of the plants in Assay 1 formed bulbs, whereas plants in Assays 2 and 3 required an additional 60 days to reach full bulb formation (Figure 3B). These differences were statistically significant (Two-way ANOVA, p = 0.0004).
3.5. Detection of Plant Viruses After In Vitro Culture
After shoot tip culture, IYSV was not detected in any sample, indicating its successful elimination. However, SLV remained present in all shallot samples that had previously tested positive (Table 4, Table 5 and Table 6). Moreover, four samples that tested negative for SLV before in vitro culture were found to be positive afterward (Table 5 and Table 6).
4. Discussion
4.1. Effectiveness of Viral Detection
The results of this study revealed that our virus detection protocol using RT-PCR, with positive controls and a combination of specific and degenerate primers, was effective. The primers amplified the expected target region of the virus, enabling positive detection of SLV and IYSV in shallot samples. The analyzed shallots tested negative for OYDV, ShVX, and LYSV. OYDV has previously been reported in shallots and garlic in Ecuador [42,46], whereas LYSV and ShVX have been detected exclusively in garlic [47,48]. The absence of these viruses in our samples may be influenced by seasonal factors [49] or may reflect that they remain restricted to other Allium species, such as garlic, and have not yet spread to shallots. Future studies could consider the use of more sensitive techniques, such as nested RT-PCR [50] or digital PCR (ddPCR) [51], which can detect very low levels of viral nucleic acids and help to confirm the presence and distribution of these viruses in shallot crops across Ecuador. Moreover, techniques such as multiplex RT-qPCR or high-throughput sequencing (including RNA-seq) [52,53,54,55] could be employed for quantitative detection and characterization of known and unknown viral variants.
The presence of SLV in 87.9% of the analyzed shallot plants indicates that this virus is widely distributed within the sampled population. IYSV was present in 29.3% of shallot samples, with 27.3% showing co-infection with both viruses. SLV, a member of the Carlavirus genus and Betaflexiviridae family, is easily transmitted through sap and by aphid vectors such as Myzus ascalonicus and possibly Aphis fabae in a non-persistent manner [7]. Its presence is well-documented, having been detected in shallots [42] and garlic (Allium sativum) [48] in Ecuador, with reported SLV detection rates of 100% in shallots and approximately 88.7% in garlic. This virus has also been reported in other American countries such as Brazil, Mexico, and Argentina [56,57,58], as well as in Europe and Asia, including Hungary, India, and China [59,60,61].
Interestingly, 10.1% of samples tested negative for SLV, suggesting that although SLV is widespread in shallot plants, it is not always present in all samples. These negative cases were detected in the Iq1 and Iq2 markets, with Iq2 also showing a higher incidence of IYSV infection. This variation in viral presence could be related to differences in shallot crop management practices, including water management, crop rotation, fertilization, and pest and disease control measures, such as the use of chemical pesticides [62,63,64]. Other studies have reported different viral profiles in shallots, including cases where SLV was less dominant than other viruses. For example, a study from Indonesia reported SLV infection rates ranging from 60% to 78%, indicating that 22% to 40% of the samples were free of SLV [65]. Furthermore, another study in France reported some shallot samples negative for SLV but positive for ShVX and newly identified carlaviruses and potyviruses [66]. Collectively, these findings suggest that shallots are susceptible to diverse viral infections and that SLV is not consistently present in all populations.
The other virus detected in our samples was IYSV, which belongs to the Tospovirus genus and Bunyaviridae family and represents an emerging threat to onion bulb and seed production [67]. Symptoms produced by IYSV include diamond-shaped lesions of 1–5 cm limited by a chlorotic zone resulting from chloroplast malformation [68,69,70,71]. These symptoms were observed on some leaves of shallot plants from OM, Iq1, and Iq2, but not in EF. Subsequent molecular analyses revealed a correlation between the observed symptoms and the presence of the viruses. Although IYSV had not been previously reported in shallots or garlic in Ecuador, it has been detected in onions [72]. To our knowledge, this is the first report of IYSV infecting shallots in Ecuador. Given that IYSV infects a wide range of Allium species worldwide—including onion, shallot, leek, garlic, and chive—in Europe (the Netherlands, Germany, Spain, France, and Austria), the Americas (the United States, Mexico, Peru, Brazil, and Uruguay), Africa (South Africa), Asia (Japan and India), Oceania (Australia), and the Mediterranean region [7,73,74,75,76,77,78,79,80,81,82,83,84,85,86], these findings are particularly noteworthy. Considering the agricultural importance of Allium crops in Ecuador, the implementation of early preventive measures against IYSV could be highly beneficial in the long term.
The transmission of IYSV is due to vectors and virus reservoirs in plants [87]. The main vector, Thrips tabaci Lindeman, commonly known as “onion trips”, transmits the virus in a persistent and propagative manner [88,89]. The detection of IYSV in shallots and onions in Ecuador probably indicates the presence of the vector. A recent study confirmed the presence of Thrips palmi and Thrips tabaci in Ecuador [90]. Although there is limited evidence regarding the crops affected by T. tabaci in the country, this warrants further investigation, as it is the main vector of IYSV. Moreover, this virus can infect other Allium species, such as garlic and onion, which are also widely cultivated in Ecuador [47,72,91]. The infection with IYSV also makes the plant more vulnerable to environmental stresses such as drought, high temperatures, and excessive irrigation [7,89]. Moreover, some secondary pathogens, such as Alternaria spp. and Stemphylum spp., can colonize those necrotic viral lesions [73], complicating the diagnosis. Therefore, early IYSV detection is critical and could help to implement suitable control and prevention methods to limit viral spread and reduce potential economic crop losses in the future. In addition, accurate identification of the vector and study of its biology could provide valuable insights to help growers manage this disease, particularly in countries like Ecuador, where Allium crops are commonly used and cultivated.
Several viruses are capable of simultaneously infecting shallots, resulting in mixed infections within a single plant [66]. Such coinfections were observed in samples OM, Iq1, and Iq2. Notably, infections of IYSV with Carlavirus and Allexivirus have been documented, and their concurrent presence often produces a synergistic effect that intensifies symptoms and causes severe yield losses [92]. This poses particular challenges for farmers in rural areas, who may lack the necessary information and resources to manage these complex infections effectively. In addition, virus monitoring in shallots and other Allium crops within the country remains limited and is not as extensively documented as in other countries. These findings underscore the importance of government agricultural agencies establishing routine screening programs, not only for the viruses identified in this study, but also for additional viruses that may threaten crop production.
4.2. In Vitro Shoot Tip Culture of Shallot
Our standardized disinfection protocol achieved sterility in 32% to 48% of shallot basal disc assays. Although these percentages were lower than expected, they provided us enough sterile plant material for subsequent in vitro assays. Fungi are the most common pests that attack shallots, especially Fusarium oxysporum, Spodoptera exigua, Alternaria porri; each causing a different disease in the plant [93]. Therefore, selecting an effective fungicide to obtain sterile basal discs was essential. We used Mertect, which contains thiabendazole, a compound known for its effectiveness and broad spectrum of activity against various pathogenic fungi [94]. Benomyl is often recommended and has been widely reported to inhibit most fungal taxa [95,96], and to achieve 100% sterility in shallots [42]; however it is banned in Ecuador [97] due to its teratogenic and toxic effects [98].
In all assays, 100% basal disc sprouting was achieved after the sterilization process. This should be highlighted because, unlike previous studies by Vega et al. [36] and Ramírez [42], we did not use a drying or cooling treatment for the shallot bulbs [99,100]; all the bulbs were fresh. Our results showed that all discs sprouted after only 15 days of culture, demonstrating the efficiency of our protocol.
More than 59.1% of shoot tips developed into plantlets, 100% of shallots developed roots, and over 83.1% of plantlets produced bulbs after up to 300 days of culture. Rapid root system development is a key strategy for plant survival and growth [101], and these regeneration rates are notably higher than those reported in previous studies by Vega et al. [36] and Ramírez [42], which achieved only 30% to 40% and 68%, respectively. In this study, adding ancymidol (an inhibitor of gibberellin biosynthesis) to the M3 medium significantly increased the percentage of bulb formation. Ancymidol promotes bulbing and rhizogenesis by reducing sucrose levels at the leaf bases and increasing the contents of glucose, fructan, and fructose [102]. The observed bulb formation rate surpassed the 77% previously reported [36], underscoring the effectiveness of this approach.
All rooted and bulb-forming plants were successfully acclimatized, representing a clear improvement over previous studies in which acclimatization was either not achieved [42] or only partially successful [36]. Further research is needed to clarify all the factors that influence shallot development, particularly bulb formation, and to enhance the production and selection of high-quality plants for in vitro propagation. Overall, shoot tip culture proved effective, yielding high regeneration rates and efficient bulb formation, making it a promising tool for in vitro research on shallots.
To explore the virus elimination potential of shoot tip culture, we evaluated its effectiveness against both viruses. The shoot tip culture effectively removed IYSV, resulting in 100% virus-free plants, regardless of the source (Iq1, Iq2). Since 1999, IYSV has been included in the European and Mediterranean Plant Protection Organization (EPPO) alert list due to its potential hazard to onion crops [103,104]. It is an industry concern due to the limited availability of effective treatment options to control the virus and its vector [105]. Unfortunately, until 2023, no chemical and biological methods were available to eliminate the virus within the plant [67]. Our study demonstrates the effectiveness of shoot tip culture in eliminating IYSV, revealing its potential for future applications.
Coinfections are more challenging to eliminate than infections caused by a single virus [106,107], and virus-host interactions likely affect eradication success [108,109]. However, the precise reasons behind this remained unclear due to its complexity [108,110]. SLV is generally asymptomatic and, on its own, is not considered to cause significant economic losses in shallot crops. However, coinfections with other viruses, such as IYSV, may intensify the disease symptoms and reduce plant vigor and yield [7,66]. In this case, although SLV could not be eliminated, the elimination of IYSV may benefit co-infected plants by reducing the overall viral load and improving plant health, thereby allowing continued growth and development.
A factor that may contribute to SLV persistence is explant size, which is critical for successful virus eradication, particularly for latent viruses [109]. In our study, we selected shoot tips measuring 6 to 10 mm, as smaller explants failed to regenerate in pilot experiments. While shoot tips contain the meristem, they also include leaf primordia tissue, and some viruses are known to migrate through the vascular elements [111]. Meristem culture using explants of 0.5–0.8 mm has produced 25% to 50% virus-free garlic and shallot plants for SLV and OYDV [112], while other studies report higher eradication rates even with smaller meristem sizes [30,108,113]. For example, Verbeek et al. [30] obtained 91 to 100% OYDV-G-free garlic plants using meristems of 0.21 to 0.33 mm, while cultures of 0.21 mm meristems produced 62-65% virus-free garlic plants for OYDV, LYSV, and GCLV [113]. In shallots, Wang et al. [108] found that 0.5 mm meristem culture resulted in only 10% OYDV and 15% SLV-free plants. Conversely, larger explants such as shoot tips produce plants with a viral elimination rate of 96.2% for SLV and OYDV by the third generation [114]. Generally, smaller explants improve virus elimination but reduce survival and regeneration rates [28,115,116]. Since one of the main objectives of in vitro culture is to generate new plants, selecting an appropriate explant size requires balancing these factors. The size of shoot tips used in our study allowed both a high rate of regeneration and viral elimination of more symptomatic viruses.
SLV is a complex virus that is difficult to eliminate fully with a single treatment. Bradamante et al. [117] suggest that some viruses can also evade the antiviral barriers of the meristem. Supporting this, Magyar-Tábori [118] found that only 20% to 27% of the 0.5 mm shallot meristematic region was free of SLV, demonstrating that combined methods may be necessary for higher or complete elimination. This study serves as a preliminary screening before testing other or integrated treatments. Therefore, future efforts should focus on integrated approaches, combining methods like thermotherapy, chemotherapy, and shoot tip culture for SLV higher eradication rates [21,108].
Unexpectedly, four shallot samples from assays 2 and 3 tested negative for SLV before in vitro culture and positive after shoot tip culture. This result may be explained by low initial viral loads below RT-PCR detection thresholds. SLV levels appeared to fluctuate across plant developmental stages [119]. Velásquez-Valle [120] also showed that virus concentrations tend to increase after an ineffective eradication treatment, suggesting that the virus may have replicated and accumulated during shoot tip culture, making it detectable later. Studies on SLV mechanism are limited, but in a few cases latent virus titers may vary under certain conditions, such as those induced by environmental stress (produced by the in vitro culture process itself) or during specific developmental stages of the host plant [121,122,123].
5. Conclusions
This study reports an efficient protocol for the molecular detection of two of the most common viruses in shallot crops, SLV and IYSV. To our knowledge, this is the first report of the presence of IYSV in shallots in Ecuador. This detection may support the implementation of control, monitoring, and prevention methods to limit the spread of the virus and reduce significant economic and crop losses in shallot or other Allium crops. The presence of the virus implies the presence of its vector [88,91]. Now that Thrips tabaci has been reported in Ecuador [90], understanding its life cycle and interactions with IYSV could help growers to control the disease. The results of our in vitro shoot tip culture proved to be effective in eliminating IYSV, which in other countries has caused significant economic losses by impacting crop quality and yield, and no chemical and biological methods were available to eliminate the virus within the plant [67]. Although our study focused on the detection of five viruses, other viruses may be present in shallots, so it would be interesting in further studies to use third-generation sequencing to understand the complete shallot virome. The in vitro culture was efficient and allowed a high percentage of plant development. Therefore, in vitro shoot tip culture could be a valuable tool for future research and an effective method to eradicate symptomatic viruses in shallots. Performing multiple rounds of shoot tip culture could further ensure the production of truly virus-free plants. Good-quality plants (free of IYSV and effectively acclimatized) obtained in this study were provided to local farmers, thereby enabling them to obtain high-quality plant material for propagation in the field. Research of this kind not only advances scientific knowledge but also provides valuable inputs for the improvement of agricultural practices.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Amplification of PCR products of positive controls with designed and literature-based primers; Figure S2: Pilot evaluation of PCR products of shallot samples with designed and literature-based primers; Figure S3: Scheme of shallot shoot regeneration from shoot tips; Figure S4: The average number of shoots tips extracted per basal disc. Table S1: Primers preliminary tested in the study; Table S2: Primers and target genes used in the study.; Table S3: Disinfection protocol for shallot bulbs; Table S4: Media composition obtained from Vega et al [36]; Table S5: Quantification of total RNA extracted from shallot leaves and absorbance index before in vitro culture; Table S6: Quantification of total RNA extracted from shallot leaves and absorbance index after in vitro culture.
Author Contributions
Conceptualization, Maria de Lourdes Torres (M.L.T.), Miguel Orellana (M.O.), Diana Calderón (D.C.), and Carlos Ruales (C.R.); methodology, Kelly Zúñiga-Vera (K.Z.V.), M.O., Martina Albuja-Quintana (M.A.Q.), D.C.; investigation, K.Z.V., D.C., M.A.Q.; data curation, K.Z.V.; formal analysis, K.Z.V.; resources, M.O., K.Z.V, and C.R.; validation, K.Z.V. and D.C., M.A.Q.; visualization, K.Z.V.; writing—original draft preparation, K.Z.V., M.L.T.; writing—review and editing, K.Z.V., M.O., M.A.Q., D.C. and M.L.T.; supervision, M.L.T.; project administration, M.L.T.; funding acquisition, M.L.T, and D.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the COCIBA grant number 20170 (2018, 2019) provided by the Colegio de Ciencias Biológicas y Ambientales COCIBA– USFQ to María de Lourdes Torres.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All original contributions are included in the article and Supplementary Material; further inquiries should be directed to the corresponding authors.
Acknowledgments
We thank the USFQ Laboratory of Plant Biotechnology for providing access to equipment, laboratory space, and other facilities. Special thanks to Carlos Ruales and the USFQ Experimental Farm for their support with shallot samples, Darío Cueva for his assistance with primer design, and Lizeth Larreátegui for her support during in vitro culture work. Our work was possible thanks to COCIBA Grants provided by the Colegio de Ciencias Biológicas y Ambientales COCIBA–USFQ.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pareek, S.; Sagar, N.A.; Sharma, S.; Kumar, V. Onion ( Allium cepa L.). In Fruit and Vegetable Phytochemicals, 1st ed.; Yahia, E.M., Ed.; Wiley, 2017; pp. 1145–1162. [Google Scholar] [CrossRef]
- Fritsch, R.M.; Friesen, N. Evolution, domestication and taxonomy. In Allium crop science: recent advances, 1st ed. 1st ed; Rabinowitch, H.D., Currah, L., Eds.; CABI Publishing: UK, 2002; pp. 5–30. [Google Scholar] [CrossRef]
- Perković, J.; Major, N.; Ban, D.; Cvitan, D.; Ban, S.G. Shallot Species and Subtypes Discrimination Based on Morphology Descriptors. Plants 2020, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Major, N.; Perković, J.; Palčić, I.; Bažon, I.; Horvat, I.; Ban, D.; et al. The Phytochemical and Nutritional Composition of Shallot Species (Allium × cornutum, Allium × proliferum and A. cepa Aggregatum) Is Genetically and Environmentally Dependent. Antioxidants 2022, 11, 1547. [Google Scholar] [CrossRef]
- Nhung, T.; Quoc, L. Efficacy of Black Shallot Extract in Analgesic and Antipyretic Activities in Experimental Mice. TJNPR 2024, 8. [Google Scholar] [CrossRef]
- Gargi, B.; Singh, P.; Painuli, S.; Rai, N.; Semwal, P.; Cruz-Martins, N.; et al. Literature-based screening and bibliometric analysis of the chemical composition, antioxidant and antimicrobial potential of essential oils isolated from Allium genus: 23 years of investigation. Pharmacol. Res.-Mod. Chin. Med. 2024, 10, 100354. [Google Scholar] [CrossRef]
- Katis, N.I.; Maliogka, V.I.; Dovas, C.I. Viruses of the Genus Allium in the Mediterranean Region. In Advances in Virus Research; Elsevier, 2012; pp. 163–208. [Google Scholar] [CrossRef]
- Rabinowitch, H.D.; Kamenetsky, R. Shallot ( Allium cepa, Aggregatum group). In Allium crop science: recent advances, 1st ed. 1st ed; Rabinowitch, H.D., Currah, L., Eds.; CABI Publishing: UK, 2002; pp. 409–430. [Google Scholar] [CrossRef]
- Mehetre, G.T.; Leo, V.V.; Singh, G.; Sorokan, A.; Maksimov, I.; Yadav, M.K.; et al. Current Developments and Challenges in Plant Viral Diagnostics: A Systematic Review. Viruses 2021, 13, 412. [Google Scholar] [CrossRef] [PubMed]
- Gunaeni, N.; Adiyoga, W.; Rosliani, R.; Sulastrini, I. The Effect of Plant Growth Regulators and Planting Density against Viral Infection and the Production from Bulbs of True Shallot Seed in the Highlands. IOP Conf. Ser. Earth Env. Sci. 2021, 752, 012033. [Google Scholar] [CrossRef]
- Varveri, C.; Maliogka, V.I.; Kapari-Isaia, T. Principles for Supplying Virus-Tested Material. In Advances in Virus Research; Elsevier, 2015; pp. 1–32. [Google Scholar] [CrossRef]
- Nie, X.; Singh, R.P. Viroid Detection and Identification by Bioassay. In Viroids and Satellites; Elsevier, 2017; pp. 347–356. [Google Scholar] [CrossRef]
- Turina, M.; Tavella, L.; Ciuffo, M. Tospoviruses in the Mediterranean Area. In Advances in Virus Research; Elsevier, 2012; pp. 403–437. [Google Scholar] [CrossRef]
- Van Der Vlugt, R.A.A.; Steffens, P.; Cuperus, C.; Barg, E.; Lesemann, D.-E.; Bos, L.; et al. Further Evidence that Shallot Yellow Stripe Virus (SYSV) Is a Distinct Potyvirus and Reidentification of Welsh Onion Yellow Stripe Virus as a SYSV Strain. Phytopathol. 1999, 89, 148–155. [Google Scholar] [CrossRef]
- Lima, J.A.; Q. Nascimento, A.K.; Radaelli, P.E. D. Serology Applied to Plant Virology. In Serological Diagnosis of Certain Human, Animal and Plant Diseases. InTech; Al-Moslih, M., Ed.; 2012. [Google Scholar] [CrossRef]
- Jordan, R.; Hammond, J. Comparison and Differentiation of Potyvirus Isolates and Identification of Strain-, Virus-, Subgroup-specific and Potyvirus Group-common Epitopes Using Monoclonal Antibodies. J. General. Virol. 1991, 72, 25–36. [Google Scholar] [CrossRef]
- Matthews, R.E.F. 5 - Serological Techniques for Plant Viruses. In Methods in Virology; MARAMOROSCH, K., KOPROWSKI, H., Eds.; Elsevier, 1967; pp. 199–241. [Google Scholar] [CrossRef]
- Cassedy, A.; Parle-McDermott, A.; O’Kennedy, R. Virus Detection: A Review of the Current and Emerging Molecular and Immunological Methods. Front Mol. Biosci. 2021, 8. [Google Scholar] [CrossRef]
- Rubio, L.; Galipienso, L.; Ferriol, I. Detection of Plant Viruses and Disease Management: Relevance of Genetic Diversity and Evolution. Front Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.D.; Ramasamy, S.; Obbineni, J.M. An evaluation of nucleic acid-based molecular methods for the detection of plant viruses: a systematic review. VirusDis 2024, 35, 357–376. [Google Scholar] [CrossRef]
- Benke, A.P.; Krishna, R.; Khandagale, K.; Gawande, S.; Shelke, P.; Dukare, S.; et al. Efficient Elimination of Viruses from Garlic Using a Combination of Shoot Meristem Culture, Thermotherapy, and Chemical Treatment. Pathogens 2023, 12, 129. [Google Scholar] [CrossRef]
- Karjadi, A.K.; Aswani, N.; Gunaeni, N. Elimination of systemic viral disease in shallot (Allium ascolonicum L) var. Bima Brebes through unconventional approach. IOP Conf. Ser. Earth Env. Sci. 2023, 1230, 012099. [Google Scholar] [CrossRef]
- Bhat, A.I.; Rao, G.P. Virus Elimination by Meristem-Tip Culture. In Characterization of Plant Viruses : Methods and Protocols; Bhat, A.I., Rao, G.P., Eds.; Springer US: New York, NY, 2020; pp. 465–477. [Google Scholar] [CrossRef]
- Krishna, R.; Ansari, W.A.; Khandagale, K.; Benke, A.P.; Soumia, P.S.; Manjunathagowda, D.C.; et al. Chapter 14 - Meristem culture: A potential technique for in vitro virus-free plants production in vegetatively propagated crops. In Advances in Plant Tissue Culture; Chandra Rai, A., Kumar, A., Modi, A., Singh, M., Eds.; Academic Press, 2022; pp. 325–343. [Google Scholar] [CrossRef]
- Grout, B.W.W. Meristem-Tip Culture for Propagation and Virus Elimination. In Plant Cell Culture Protocols; Hall, R.D., Ed.; Humana Press: Totowa, NJ, 1999; pp. 115–125. [Google Scholar] [CrossRef]
- Vivek, M.; Modgil, M. Elimination of viruses through thermotherapy and meristem culture in apple cultivar ‘Oregon Spur-II. VirusDis 2018, 29, 75–82. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Su, Y.; Li, X.Y.; Zhang, C.Y.; Ma, J.F.; Liu, J.Q.; et al. First Report of Shallot Virus X and Shallot Latent Virus on Shallot ( Allium cepa var. aggregatum ) in China. Plant Dis. 2019, 103, 2972–2972. [Google Scholar] [CrossRef]
- Maliogka, V.I.; Martelli, G.P.; Fuchs, M.; Katis, N.I. Control of Viruses Infecting Grapevine. In Advances in Virus Research; Elsevier, 2015; pp. 175–227. [Google Scholar] [CrossRef]
- Vieira, R.L.; Da Silva, A.L.; Zaffari, G.R.; Steinmacher, D.A.; De Freitas Fraga, H.P.; Guerra, M.P. Efficient elimination of virus complex from garlic (Allium sativum L.) by cryotherapy of shoot tips. Acta Physiol. Plant. 2015, 37, 1733. [Google Scholar] [CrossRef]
- Verbeek, M.; Van Dijk, P.; Van Well, P.M.A. Efficiency of eradication of four viruses from garlic (Allium sativum) by meristem-tip culture. Eur. J. Plant Pathol. 1995, 101, 231–239. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, H.-L.; Zhang, C.-J.; Kang, Y.-Q. High rate of virus-free plantlet regeneration via garlic scape-tip culture. Plant Cell Rep. 1994, 14. [Google Scholar] [CrossRef]
- Wyckhuys, K.A.G.; Zou, Y.; Crowder, D.W.; Adriani, E.; Albaytar, A.B.; Beltran, M.J.B.; et al. Biological control mitigates spread of vector-borne plant pathogens. Agric. Ecosyst. Environ. 2025, 388, 109683. [Google Scholar] [CrossRef]
- Anikina, I.; Kamarova, A.; Issayeva, K.; Issakhanova, S.; Mustafayeva, N.; Insebayeva, M.; et al. Plant protection from virus: a review of different approaches. Front Plant Sci. 2023, 14, 1163270. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.C. Using epidemiological information to develop effective integrated virus disease management strategies. Virus Res. 2004, 100, 5–30. [Google Scholar] [CrossRef]
- Montagnini, F. (Ed.) Integrating Landscapes: Agroforestry for Biodiversity Conservation and Food Sovereignty; Springer International Publishing: Cham, 2017. [Google Scholar] [CrossRef]
- Vega, J.; Arahana, B. V.S.; Torres, M.D.L. Estandarización de un protocolo de regeneración de cebolla chalote (Allium cepa var. aggregatum) a partir de meristemas apicales. Av. Cienc. Ing. 2015, 7. [Google Scholar] [CrossRef]
- Ward, L.I.; Perez-Egusquiza, Z.; Fletcher, J.D.; Clover, G.R.G. A survey of viral diseases of Allium crops in New Zealand. Austral Plant Pathol. 2009, 38, 533. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; et al. Primer3—new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115–e115. [Google Scholar] [CrossRef]
- NCBI. 2024. Available online: https://www.ncbi.nlm.nih.gov/tools/primer-blast/.
- Qu, W.; Zhang, C. Selecting Specific PCR Primers with MFEprimer. In PCR Primer Design; Basu, C., Ed.; Springer New York: New York, NY, 2015; pp. 201–213. [Google Scholar] [CrossRef]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA Using TRIzol (TRI Reagent). In Cold Spring Harb Protoc; 2010; Volume 2010, p. pdb.prot5439. [Google Scholar] [CrossRef]
- Ramírez, D. Regeneración de plántulas de chalote (Allium cepa var. aggregatum) libres de los virus Latente del Chalote y del Enanismo Amarillo de la Cebolla por medio de cultivo de meristema apical y quimioterapia; Universidad San Francisco de Quito, 2012. [Google Scholar]
- Nehra, N.S.; Kartha, K.K. Meristem and Shoot Tip Culture: Requirements and Applications. In Plant Cell and Tissue Culture; Vasil, I.K., Thorpe, T.A., Eds.; Springer Netherlands: Dordrecht, 1994; pp. 37–70. [Google Scholar] [CrossRef]
- Wang, M.-R.; Zhang, Z.; Zámečník, J.; Bilavčík, A.; Blystad, D.-R.; Haugslien, S.; et al. Droplet-vitrification for shoot tip cryopreservation of shallot (Allium cepa var. aggregatum): effects of PVS3 and PVS2 on shoot regrowth. Plant Cell Tiss. Organ Cult. 2020, 140, 185–195. [Google Scholar] [CrossRef]
- Rajan, P.; Lada, R.R.; MacDonald, M.T. Advancement in Indoor Vertical Farming for Microgreen Production. AJPS 2019, 10, 1397–1408. [Google Scholar] [CrossRef]
- Heredia, S. Erradicación de Onion Yellow Dwarf Virus (OYDV) en cebolla Shallot (Allium cepa var. Aggregatum), mediante el Cultivo de meristemas, quimioterapia y termoterapia para la producción de bulbos libres de virus. 2016. [Google Scholar]
- Granda, R.; Landázuri, G.; Arkhipov, A.V. First Report of Shallot virus X in Garlic in Ecuador. Plant Dis. 2017, 101, 1066. [Google Scholar] [CrossRef]
- Oleas, A.; Arahana, V. First report of leek yellow stripe virus, shallot latent virus, and onion yellow dwarf virus in Garlic from Ecuador. Plant Dis. 2016, 100, 232. [Google Scholar] [CrossRef]
- Flasco, M.; Hoyle, V.; Powell, G.; Seiter, J.; Wise, A.; Cieniewicz, E.J.; et al. Seasonal Variation in Grapevine Red Blotch Virus Titer in Relation to Disease Symptom Expression in Vineyards. Phytobiomes J. 2024, 8, 192–200. [Google Scholar] [CrossRef]
- Zambrana-Echevarría, C.; Roth, M.G.; Dasgupta, R.; German, T.L.; Groves, C.L.; Smith, D.L. Sensitive and Specific qPCR and Nested RT-PCR Assays for the Detection of Tobacco Streak Virus in Soybean. PhytoFrontiersTM 2021, 1, 291–300. [Google Scholar] [CrossRef]
- Wu, H.; Liu, M.; Li, W.; Wang, M.; Xiu, J.; Peng, B.; et al. Development and Application of Droplet Digital PCR Assay for the Detection of Watermelon Silver Mottle Virus and Melon Yellow Spot Virus. Horticulturae 2024, 10, 199. [Google Scholar] [CrossRef]
- Dantes, W.; Boatwright, L.; Cieniewicz, E.J. Comparing RT-PCR of Individual Samples with High-Throughput Sequencing of Pooled Plant Samples for Field-Level Surveillance of Viruses in Blackberry and Wild Rubus. Plant Dis. 2024, 108, 2435–2446. [Google Scholar] [CrossRef]
- Fei, S.; Yu, J.; Zhou, Y.; Xie, Y.; Xie, L.; Fu, S.; et al. Discovery and characterization of a novel carlavirus in Ligularia jaluensis plants. Virol. Sin. 2025, 40, 71–79. [Google Scholar] [CrossRef]
- Dias, N.P.; Hu, R.; Hensley, D.D.; Hansen, Z.R.; Domier, L.L.; Hajimorad, M.R. A Survey for Viruses and Viroids of Peach in Tennessee Orchards by RNA Sequencing. Plant Health Progress. 2022, 23, 265–268. [Google Scholar] [CrossRef]
- Dong, J.; Chen, Y.; Xie, Y.; Cao, M.; Fu, S.; Wu, J. The Identification of Viral Pathogens in a Physostegia virginiana Plant Using High-Throughput RNA Sequencing. Viruses 2023, 15, 1972. [Google Scholar] [CrossRef]
- Velásquez-Valle, R.; Zacatecas, C.; Chew-Madinaveitia, I. Presencia de Virus en el Cultivo de Ajo (Allium sativum L.) en Zacatecas, México Virus Presence in the Cultivation of Garlic (Allium sativum L.) in Zacatecas, México. Rev. Mex. DE Fitopatol. 2010, 28, 135–X. [Google Scholar]
- Mituti, T.; Marubayashi, J.M.; Moura, M.F.; Krause-Sakate, R.; Pavan, M.A. First Report of Shallot latent virus in Garlic in Brazil. Plant Dis. 2011, 95, 227–227. [Google Scholar] [CrossRef]
- Torrico, A.K.; Cafrune, E.E.; Conci, V.C. First Report of Shallot latent virus in Garlic in Argentina. Plant Dis. 2010, 94, 915–915. [Google Scholar] [CrossRef]
- Hu, X.; Lei, Y.; Wang, P.; Tang, L.; He, C.; Song, Y.; et al. Development of a multiplex reverse transcription-PCR assay for simultaneous detection of garlic viruses. J. Integr. Agric. 2015, 14, 900–908. [Google Scholar] [CrossRef]
- Koczor, Á.; Ádám, J.; Ágoston, J.; Salánki, K.; Palkovics, L. Investigation of viral diseases of garlic (Allium sativum L.), new primers for RT-PCR detection and diversity of garlic viruses in Hungary. Physiol. Mol. Plant Pathol. 2024, 134, 102394. [Google Scholar] [CrossRef]
- Majumder, S.; Baranwal, V.K.; Joshi, S. Simultaneous detection of onion yellow dwarf virus and shallot latent virus in infected leaves and cloves of garlic by duplex RT-PCR. J. Plant Pathol. 2008, 90, 371–374. [Google Scholar]
- Rovicky, * A.; Widowati, W.; Astutik, A. Pest and Disease Control Strategies to Increase the Productivity of Shallot Plants (Allium ascalonium L.). REJHH 2024, 7, 1253–1260. [Google Scholar] [CrossRef]
- Sopha, G.A. Influence of Plant Density, Compost and Biofertilizer on True Shallot Seed Growth in Alluvial Soil. Indones. J. Agric. Sci. 2020, 21, 70. [Google Scholar] [CrossRef]
- Sutardi; Kristamtini; Purwaningsih, H.; Widyayanti, S.; Arianti, F.D.; Pertiwi, M.D.; et al. Nutrient Management of Shallot Farming in Sandy Loam Soil in Tegalrejo, Gunungkidul, Indonesia. Sustainability 2022, 14, 11862. [Google Scholar] [CrossRef]
- Sulistio, M.; Sulistyaningsih, E.; Subandiyah, S. Elimination of shallot bulb viruses through heat treatment. IJBiotech 2017, 20, 133. [Google Scholar] [CrossRef]
- Marais, A.; Faure, C.; Theil, S.; Candresse, T. Characterization of the virome of shallots affected by the shallot mild yellow stripe disease in France. In PLoS ONE; Tatineni, S., Ed.; 2019; Volume 14. [Google Scholar] [CrossRef]
- Sharma, S.; Cramer, C.S. Reduced Iris yellow spot symptom expression in the selected onion germplasm. V 2023, 3, 0–0. [Google Scholar] [CrossRef]
- Colnago, P.; Achigar, R.; González, P.H.; Peluffo, S.; González Idiarte, H.; Pianzzola, M.J.; et al. First Report of Iris yellow spot virus on Onion in Uruguay. Plant Dis. 2010, 94, 786–786. [Google Scholar] [CrossRef]
- Bhattacharyya, D.; Chakraborty, S. Chloroplast: the Trojan horse in plant–virus interaction. Mol. Plant Pathol. 2018, 19, 504–518. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, J.; Chen, B.; Cui, W.; Zhou, Z.; Song, X.; et al. Characterization of Proteins Involved in Chloroplast Targeting Disturbed by Rice Stripe Virus by Novel Protoplast–Chloroplast Proteomics. IJMS 2019, 20, 253. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Qari, S.H.; Hafez, E. Iris yellow spot virus–induced chloroplast malformation results in male sterility. J. Biosci. 2019, 44, 142. [Google Scholar] [CrossRef]
- Sivaprasad, Y.; Garrido, P.; Mendez, K.; Garrido, A.; Ramos, L. First report of Iris yellow spot virus infecting onion in the Pichincha and Tungurahua provinces of Ecuador. New Dis. Rep. 2016, 33, 16–16. [Google Scholar] [CrossRef]
- Colnago, P.; Achigar, R.; Ravelino, P.G.; Peluffo, S.; Idiarte, H.G.; Pianzzola, M.J.; et al. PRESENCIA DEL VIRUS IRIS YELLOW SPOT VIRUS (IYSV) EN SEMILLEROS DE CEBOLLA. INIA Las Brujas – Estación Experimental “Wilson Ferreira Aldunate.”; 2010; pp. 53–57. [Google Scholar]
- Cortês, I.; Livieratos, I.C.; Derks, A.; Peters, D.; Kormelink, R. Molecular and Serological Characterization of Iris Yellow Spot Virus, a New and Distinct Tospovirus Species. Phytopathol. 1998, 88, 1276–1282. [Google Scholar] [CrossRef]
- Coutts, B.A.; McMichael, L.A.; Tesoriero, L.; Rodoni, B.C.; Wilson, C.R.; Wilson, A.J.; et al. Iris yellow spot virus found infecting onions in three Australian states. Austral Plant Pathol. 2003, 32, 555. [Google Scholar] [CrossRef]
- Creamer, R.; Sanogo, S.; Moya, A.; Romero, J.; Molina-Bravo, R.; Cramer, C. Iris yellow spot virus on Onion in New Mexico. Plant Dis. 2004, 88, 1049–1049. [Google Scholar] [CrossRef] [PubMed]
- Gawande, S.; Gurav, V.; Ingle, A.; Gopal, J. First Report of Iris yellow spot virus Infecting Allium tuberosum in India. Plant Dis. 2014, 98, 1161. [Google Scholar] [CrossRef]
- Gent, D.H.; Schwartz, H.F.; Khosla, R. Distribution and Incidence of Iris yellow spot virus in Colorado and Its Relation to Onion Plant Population and Yield. Plant Dis. 2004, 88, 446–452. [Google Scholar] [CrossRef]
- Leinhos, G.; Müller, J.; Heupel, M.; Krauthausen, H.-J. Iris Yellow Spot Virus an Bund- und Speisezwiebeln – erster Nachweis in Deutschland. Nachrichtenbl Deut Pflanzenschutzd. 2007, 59, 310–312. [Google Scholar]
- Mullis, S.W.; Gitaitis, R.D.; Nischwitz, C.; Csinos, A.S.; Rafael Mallaupoma, Z.C.; Inguil Rojas, E.H. First Report of Onion ( Allium cepa ) Naturally Infected with Iris yellow spot virus in Peru. Plant Dis. 2006, 90, 377–377. [Google Scholar] [CrossRef]
- Pozzer, L.; Bezerra, I.C.; Kormelink, R.; Prins, M.; Peters, D.; Resende, R.D.O.; et al. Characterization of a Tospovirus Isolate of Iris Yellow Spot Virus Associated with a Disease in Onion Fields in Brazil. Plant Dis. 1999, 83, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, H.F.; Brown, W.M.; Blunt, T.; Gent, D.H. Iris yellow spot virus on Onion in Colorado. Plant Dis. 2002, 86, 560–560. [Google Scholar] [CrossRef] [PubMed]
- Toit, L.J.; du; Burger, J.T.; McLeod, A.; Engelbrecht, M.; Viljoen, A. Iris yellow spot virus in Onion Seed Crops in South Africa. Plant Dis. 2007, 91, 1203–1203. [Google Scholar] [CrossRef] [PubMed]
- Weilner, S.; Bedlan, G. Detection of Iris yellow spot virus (IYSV) in selected Allium species and overwintering hosts in Austrian onion-producing areas. J. Fur Kultur. 2013, 65, 60–67. [Google Scholar] [CrossRef]
- Córdoba-Sellés, C.; Martínez-Priego, L.; Muńoz-Gómez, R.; Jordá-Gutiérrez, C. Iris yellow spot virus: A New Onion Disease in Spain. Plant Dis. 2007, 89, 1243. [Google Scholar] [CrossRef]
- Huchette, O.; Bellamy, C.; Filomenko, R.; Pouleau, B.; Seddas, S.; Pappu, H.R. Iris yellow spot virus on Shallot and Onion in France. Plant Health Progress. 2008, 9, 46. [Google Scholar] [CrossRef]
- Bohdan, M.M.; Kyrychenko, A.M.; Shcherbatenko, I.S.; Kraeva, H.V. Weed Plants of the Asteraceae and Malvaceae Families as Reservoirs of Harmful Viruses of Vegetable Crops in Ukraine and the World. Mikrobiol. Z. 2023, 85, 66–76. [Google Scholar] [CrossRef]
- Ávila-Alistac, N. Patosistema de Iris yellow spot orthotospovirus, hospederos del virus y el vector (Thrips tabaci). Vol. 2024;42.
- Ramírez-Rojas, S.; Ornelas-Ocampo, K.; Osuna-Canizalez, F.D.J.; Bartolo-Reyes, J.C.; Varela-Loza, V.; Hernández-Romano, J.; et al. Detección de Iris yellow spot virus en cebolla cultivada en Tepalcingo, Morelos, México. RMF 2016, 34. [Google Scholar] [CrossRef]
- Sunidhi; Singla, P.; Kaur, R.; Sharma, S. Eavesdropping the pivotal defensive representatives of plant-thrips interaction. Physiol. Mol. Biol. Plants 2025, 31, 173–197. [Google Scholar] [CrossRef]
- Pandi, A.; Perumal, R.; John Samuel, K.; Subramanian, J.; Malaichamy, K. Orthotospovirus iridimaculaflavi (iris yellow spot virus): An emerging threat to onion cultivation and its transmission by Thrips tabaci in India. Microb. Pathog. 2024, 193, 106716. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, O.; Snihur, H.; Shevchenko, T.; Budzanivska, I. Onion and Garlic. Viral Diseases of Field and Horticultural Crops; Elsevier, 2024; pp. 505–512. [Google Scholar] [CrossRef]
- Utari, S.R.P.; Purnamasari, A.; Khairi, A. A major pest and diseases of shallot (Allium cepa L. Aggregatum group) in Bima Regency. 2024.
- Papadopoulou, E.S.; Genitsaris, S.; Omirou, M.; Perruchon, C.; Stamatopoulou, A.; Ioannides, I.; et al. Bioaugmentation of thiabendazole-contaminated soils from a wastewater disposal site: Factors driving the efficacy of this strategy and the diversity of the indigenous soil bacterial community. Environ. Pollut. 2018, 233, 16–25. [Google Scholar] [CrossRef]
- Coronel, A.C.; Parraguirre Lezama, C.; Pacheco Hernández, Y.; Santiago Trinidad, O.; Rivera Tapia, A.; Romero-Arenas, O. Efficacy of Four In Vitro Fungicides for Control of Wilting of Strawberry Crops in Puebla-Mexico. Appl. Sci. 2022, 12, 3213. [Google Scholar] [CrossRef]
- Summerbell, R.C. The benomyl test as a fundamental diagnostic method for medical mycology. J. Clin. Microbiol. 1993, 31, 572–577. [Google Scholar] [CrossRef]
- Agrocalidad. Plaguicidas Prohibidos en el Ecuador. 2020. Available online: chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://www.agrocalidad.gob.ec/wp-content/uploads/2020/05/Plaguicidas-prohibidos-en-Ecuador-1.pdf.
- Kara, M.; Oztas, E.; Ramazanoğulları, R.; Kouretas, D.; Nepka, C.; Tsatsakis, A.M.; et al. Benomyl, a benzimidazole fungicide, induces oxidative stress and apoptosis in neural cells. Toxicol. Rep. 2020, 7, 501–509. [Google Scholar] [CrossRef]
- Swamy, K.R.M.; Veere Gowda, R. 22 - Leek and shallot. In Handbook of Herbs and Spices; Peter, K.V., Ed.; Woodhead Publishing, 2006; pp. 365–389. [Google Scholar] [CrossRef]
- Bufler, G. Exogenous ethylene inhibits sprout growth in onion bulbs. Ann. Bot. 2009, 103, 23–28. [Google Scholar] [CrossRef]
- Gallegos-Cedillo, V.M.; Diánez, F.; Nájera, C.; Santos, M. Plant Agronomic Features Can Predict Quality and Field Performance: A Bibliometric Analysis. Agronomy 2021, 11, 2305. [Google Scholar] [CrossRef]
- Saos, F.L.G.-L. In vitro Bulb Development in Shallot (Allium cepa L. Aggregatum Group): Effects of Anti-gibberellins, Sucrose and Light. Ann. Bot. 2002, 89, 419–425. [Google Scholar] [CrossRef]
- Cordoba-Sellés, C.; Martínez-Priego, L.; Muńoz-Gómez, R.M.; Jordá-Gutiérrez, C.; Tobarra, M. Iris yellow spot virus: A New Onion Disease in Spain. Plant Dis. 2007, 89, 1243. [Google Scholar] [CrossRef] [PubMed]
- Bulajić, A.; Djekić, I.; Jović, J.; Krnjajić, S.; Vučurović, A.; Krstić, B. Incidence and Distribution of Iris yellow spot virus on Onion in Serbia. Plant Dis. 2009, 93, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.; Raikhy, G.; Goodin, M.M.; Dietzgen, R.G.; Pappu, H.R. In Vivo Localization of Iris yellow spot Tospovirus (Bunyaviridae)-Encoded Proteins and Identification of Interacting Regions of Nucleocapsid and Movement Proteins. In PLoS ONE; Rao, A.L.N., Ed.; 2015; Volume 10. [Google Scholar] [CrossRef]
- Sánchez-Tovar, M.R.; Rivera-Bustamante, R.F.; Saavedra-Trejo, D.L.; Guevara-González, R.G.; Torres-Pacheco, I. Mixed Plant Viral Infections: Complementation, Interference and Their Effects, a Review. Agronomy 2025, 15, 620. [Google Scholar] [CrossRef]
- Pramesh, D.; Baranwal, V.K. Production of virus-free garlic (Allium sativumL.) through meristem tip culture after solar or hot air treatment of cloves. J. Hortic. Sci. Biotechnol. 2015, 90, 180–186. [Google Scholar] [CrossRef]
- Wang, M.; Hamborg, Z.; Blystad, D.; Wang, Q. Combining thermotherapy with meristem culture for improved eradication of onion yellow dwarf virus and shallot latent virus from infected in vitro-cultured shallot shoots. Ann. Appl. Biol. 2021, 178, 442–449. [Google Scholar] [CrossRef]
- Wang, M.-R.; Cui, Z.-H.; Li, J.-W.; Hao, X.-Y.; Zhao, L.; Wang, Q.-C. In vitro thermotherapy-based methods for plant virus eradication. Plant Methods 2018, 14, 87. [Google Scholar] [CrossRef]
- McLaughlin, A.A.; Hanley-Bowdoin, L.; Kennedy, G.G.; Jacobson, A.L. Vector acquisition and co-inoculation of two plant viruses influences transmission, infection, and replication in new hosts. Sci. Rep. 2022, 12, 20355. [Google Scholar] [CrossRef]
- Hipper, C.; Brault, V.; Ziegler-Graff, V.; Revers, F. Viral and Cellular Factors Involved in Phloem Transport of Plant Viruses. Front Plant Sci. 2013, 4. [Google Scholar] [CrossRef]
- Walkey, D.G.A.; Webb, M.J.W.; Bolland, C.J.; Miller, A. Production of virus-free garlic ( Allium sativum L.) and shallot ( A. ascalonicum L.) by meristem-tip culture. J. Hortic. Sci. 1987, 62, 211–220. [Google Scholar] [CrossRef]
- Shiboleth, Y.M.; Gal-On, A.; Koch, M.; Rabinowitch, H.D.; Salomon, R. Molecular characterisation of Onion yellow dwarf virus (OYDV) infecting garlic (Allium sativum L.) in Israel: Thermotherapy inhibits virus elimination by meristem tip culture. Ann. Appl. Biol. 2001, 138, 187–195. [Google Scholar] [CrossRef]
- Lou, H.; Liu, Z.; Xu, Q. Detection and elimination of shallot latent virus and onion yellow dwarf virus for potato onion (Allium cepa L. var. aggregatum Don). Ann. Appl. Biol. 2022, 182. [Google Scholar] [CrossRef]
- Kahane, R.; Rancillac, M.; De La Serve, B.T. Long-term multiplication of onion (Allium cepa L.) by cyclic shoot regeneration in vitro. Plant Cell Tiss. Organ Cult. 1992, 28, 281–288. [Google Scholar] [CrossRef]
- Panattoni, A.; Luvisi, A.; Triolo, E. Review. Elimination of viruses in plants: twenty years of progress. Span. J. Agric. res. 2013, 11, 173–188. [Google Scholar] [CrossRef]
- Bradamante, G.; Mittelsten Scheid, O.; Incarbone, M. Under siege: virus control in plant meristems and progeny. Plant Cell. 2021, 33, 2523–2537. [Google Scholar] [CrossRef]
- Magyar-Tábori, K.; Mendler-Drienyovszki, N.; Hanász, A.; Zsombik, L.; Dobránszki, J. Phytotoxicity and Other Adverse Effects on the In Vitro Shoot Cultures Caused by Virus Elimination Treatments: Reasons and Solutions. Plants 2021, 10, 670. [Google Scholar] [CrossRef] [PubMed]
- Pauzi, Y.S.; Lestari, S.M.; Hidayat, S.H. Variations of Garlic Common Latent Virus and Shallot Latent Virus Concentration on Shallot and Garlic. IOP Conf. Ser. Earth Env. Sci. 2018, 197, 012045. [Google Scholar] [CrossRef]
- Velásquez-Valle, R.; Reveles-Hernández, M.; Ileana, Y.; Medinaveitia, C.-; Reveles-Torres, L.R. Efecto del tratamiento térmico sobre la presencia de virus en bulbos de ajo (Allium sativum L.). Rev. FCA UNCUYO 2017, 49(1), 157–165. [Google Scholar]
- Takahashi, H.; Fukuhara, T.; Kitazawa, H.; Kormelink, R. Virus Latency and the Impact on Plants. Front Microbiol. 2019, 10, 2764. [Google Scholar] [CrossRef] [PubMed]
- De Klerk, G.-J. Stress in plants cultured in vitro. Propag. Ornam. Plants 2007, 7, 129–137. [Google Scholar]
- Richert-Pöggeler, K.R.; Minarovits, J. Diversity of latent plant–virus interactions and their impact on the virosphere. In Plant Virus–Host Interaction; Elsevier, 2014; pp. 263–275. [Google Scholar] [CrossRef]
Figure 1.
Plant material. A) Experimental farm. B) Organic market. C) Iñaquito market 1. D) Iñaquito market 2.
Figure 1.
Plant material. A) Experimental farm. B) Organic market. C) Iñaquito market 1. D) Iñaquito market 2.

Figure 2.
Shoot tip with leaf primordia used in shoot tip culture.

Figure 3.
Percentage of rooting and bulb formation. (A). Percentage of rooting throughout 250 days of in vitro culture. (B). Percentage of bulb formation throughout 360 days of in vitro culture. Data represent mean ± SD (n = X). Statistical significance was assessed using two-way ANOVA (rooting: p = 0.0419; bulb formation: p = 0.0004). Abbreviation: EF, Experimental Farm; Iq1, Iñaquito 1; Iq2, Iñaquito 2.
Figure 3.
Percentage of rooting and bulb formation. (A). Percentage of rooting throughout 250 days of in vitro culture. (B). Percentage of bulb formation throughout 360 days of in vitro culture. Data represent mean ± SD (n = X). Statistical significance was assessed using two-way ANOVA (rooting: p = 0.0419; bulb formation: p = 0.0004). Abbreviation: EF, Experimental Farm; Iq1, Iñaquito 1; Iq2, Iñaquito 2.


Table 1.
Viral detection in shallot plants before in vitro culture.
| Virus detection | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source | Plants analyzed (n) | One virus | Coinfection | SLV and IYSV Negative | Total SLV positive | Total IYSV positive | ||||||||
| SLV | IYSV | SLV and IYSV | ||||||||||||
| n | % | n | % | n | % | n | % | n | % | n | % | |||
| EF | 23 | 23 | 100.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 23 | 100.00 | 0 | 0.00 | |
| OM | 26 | 21 | 80.77 | 0 | 0.00 | 5 | 19.23 | 0 | 0.00 | 26 | 100.00 | 5 | 19.23 | |
| Iq1 | 25 | 10 | 40.00 | 2 | 8.00 | 4 | 16.00 | 9 | 36.00 | 14 | 56.00 | 6 | 24.00 | |
| Iq2 | 25 | 6 | 24.00 | 0 | 0.00 | 18 | 72.00 | 1 | 4.00 | 24 | 96.00 | 18 | 72.00 | |
| Total | 99 | 60 | 60.61 | 2 | 2.02 | 27 | 27.27% | 10 | 10.10 | 87 | 87.88 | 29 | 29.29 | |
Table 2.
Sterility and sprouting rates of basal discs of shallots after 15 days of in vitro culture.
Table 2.
Sterility and sprouting rates of basal discs of shallots after 15 days of in vitro culture.
| Assay | Source | Sterility1 (n, %) | Sprouting (n,%) | ||
|---|---|---|---|---|---|
| 1 | EF | 11 of 23 | 47.83% | 23 of 23 | 100% |
| 2 | Iq1 | 8 of 25 | 32% | 25 of 25 | 100% |
| 3 | Iq2 | 12 of 25 | 48% | 25 of 25 | 100% |
| Mean | 31 of 73 | 42.47% | 73 of 73 | 100% | |
1Statistical significance was assessed using Chi-square (sterility: p= 0.7899).
Table 3.
Percentages of plantlet development, rooting, and bulb formation.
| Assay | Source | Shoot tips | Plantlets1 | ||
|---|---|---|---|---|---|
| Plant development (n; %) |
Rooting (n; %) |
Bulb formation2 (n; %) |
|||
| 300 days | 360 days | ||||
| 1 | EF | 63 of 71; 88.7% |
74 of 74; 100% | 74 of 74; 100% |
74 of 74; 100% |
| 2 | Iq1 | 26 of 44; 59.1% |
36 of 36; 100% | 33 of 36; 91.7% | 36 of 36; 100% |
| 3 | Iq2 | 45 of 58; 77.6% |
65 of 65; 100% | 54 of 65; 83.1% | 65 of 65; 100% |
1Plantlet number increased after subculture; subsequent percentages were calculated based on total plantlets. 2Bulb formation data up to 360 days of culture. Statistical significance was assessed using Chi-square (plant development: p = 0.0084; bulb formation by 300 days: p = 0.0092).
Table 4.
Virus detection results in shallots from the experimental farm (Assay 1), before and after in vitro culture.
Table 4.
Virus detection results in shallots from the experimental farm (Assay 1), before and after in vitro culture.
| Sample | Virus | |||
|---|---|---|---|---|
| SLV | IYSV | |||
| Before | After | Before | After | |
| 1 | + | + | - | - |
| 2 | + | + | - | - |
| 3 | + | + | - | - |
| 4 | + | + | - | - |
| 5 | + | + | - | - |
| 6 | + | + | - | - |
| 7 | + | + | - | - |
| 8 | + | + | - | - |
| 9 | + | + | - | - |
| 10 | + | + | - | - |
| 11 | + | + | - | - |
Abbreviations: IND, Individual; SLV, Shallot Latent Virus; IYSV, Iris Yellow Spot Virus.
Table 5.
Virus detection results in shallots from Iñaquito 1 (Assay 2), before and after in vitro culture.
Table 5.
Virus detection results in shallots from Iñaquito 1 (Assay 2), before and after in vitro culture.
| Sample | Virus | |||
|---|---|---|---|---|
| SLV | IYSV | |||
| Before | After | Before | After | |
| 1 | + | + | + | - |
| 2 | - | +1 | - | - |
| 3 | + | + | - | - |
| 4 | - | +1 | + | - |
| 5 | + | + | - | - |
| 6 | - | +1 | - | - |
| 7 | + | + | + | - |
| 8 | + | + | - | - |
Abbreviations: IND, Individual; SLV, Shallot Latent Virus; IYSV, Iris Yellow Spot Virus. 1Samples that tested negative before and positive after the disinfection protocol.
Table 6.
Virus detection results in shallots from Iñaquito 2 (Assay 3), before and after in vitro culture.
Table 6.
Virus detection results in shallots from Iñaquito 2 (Assay 3), before and after in vitro culture.
| Sample | Virus | |||
|---|---|---|---|---|
| SLV | IYSV | |||
| Before | After | Before | After | |
| 1 | + | + | - | - |
| 2 | + | + | + | - |
| 3 | + | + | + | - |
| 4 | + | + | + | - |
| 5 | + | + | + | - |
| 6 | + | + | + | - |
| 7 | + | - | - | - |
| 8 | + | + | + | - |
| 9 | - | +1 | - | - |
| 10 | + | + | + | - |
| 11 | + | + | + | - |
| 12 | + | + | + | - |
Abbreviations: IND, Individual; SLV, Shallot Latent Virus; IYSV, Iris Yellow Spot Virus. 1Samples that tested negative before and positive after the disinfection protocol.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



