Submitted:
29 May 2026
Posted:
03 June 2026
You are already at the latest version
Abstract
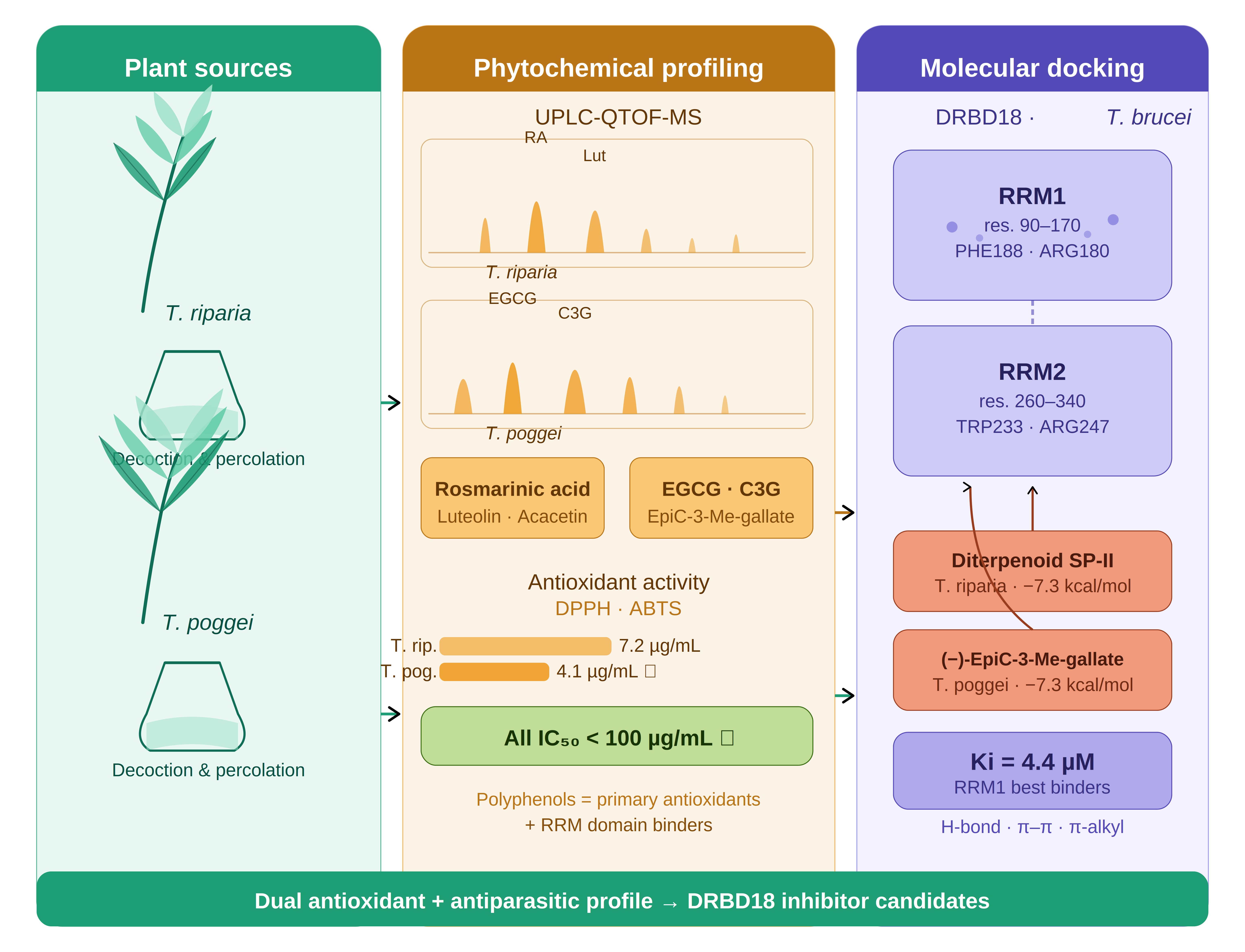
Oxidative stress plays a key role in the pathogenesis of African trypanosomiasis, and plant-derived antioxidants represent a promising source of dual-action compounds capable of both scavenging reactive oxygen species and interfering with parasite survival. The present study investigates the antioxidant activity and phytochemical composition of two Central African medicinal plants, Tetradenia riparia (Hochst.) Codd and Tetracera poggei (Gilg.) and evaluates the in silico trypanocidal potential of their major constituents against the mRNA-binding protein DRBD18 of Trypanosoma brucei. Aqueous and methanolic extracts were prepared by decoction and percolation. Qualitative phytochemical screening and quantitative spectrophotometry (Folin–Ciocalteu, AlCl3) revealed rich profiles of polyphenols, flavonoids, tannins, anthocyanins, and alkaloids in both species. UPLC-QTOF-MS in negative ESI mode identified eleven compounds in T. riparia, including rosmarinic acid, luteolin, acacetin, and diterpenoid SP-II and thirteen compounds in T. poggei, dominated by galloylated catechin derivatives (epi-gallocatechin 3-O-gallate, catechin 3-O-gallate, (−)-epicatechin-3-(3-O-methyl)-gallate). Antioxidant activity by DPPH and ABTS radical scavenging assays showed that percolated extracts consistently outperformed decocted extracts; T. poggei percolated extract showed the strongest DPPH activity (IC50 = 4.141 ± 0.175 µg/mL). Molecular docking against the RRM1 and RRM2 domains of DRBD18 (AlphaFold model AF-Q57XR9-F1) revealed that diterpenoid SP-II and (−)-epicatechin-3-(3-O-methyl)-gallate displayed the highest binding affinities (RRM1: −7.3 kcal/mol; Ki = 4.424 µM). The strong antioxidant capacity of the identified compounds, particularly galloylated catechins and rosmarinic acid, is mechanistically linked to their structural features that also favor RRM domain binding, highlighting a dual antioxidant–antiparasitic pharmacological profile. These findings provide a rational basis for the further development of plant-derived agents targeting oxidative stress and RNA-binding mechanisms in T. brucei.
Keywords:
Tetradenia riparia
; Tetracera poggei
; antioxidant activity
; DPPH
; ABTS
; UPLC-MS
; phytochemistry
; molecular docking
; DRBD18
; trypanosomiasis
1. Introduction
African Trypanosomiasis is a neglected tropical disease caused by protozoan flagellates of the genus Trypanosoma, transmitted by tsetse flies (Glossina spp.) across sub-Saharan Africa [1]. Human African trypanosomiasis (HAT) is caused by Trypanosoma brucei gambiense in West and Central Africa and by T. b. rhodesiense in East and Southern Africa, while Animal African trypanosomiasis (AAT) is caused by T. b. brucei, T. congolense, and T. vivax [1]. AAT alone threatens more than 55 million cattle and imposes critical barriers to food security and economic development [2]. The Democratic Republic of the Congo (DRC) remains one of the most affected endemic countries [3].
A well-established but underappreciated aspect of trypanosomiasis pathogenesis is the induction of oxidative stress in the mammalian host. T. brucei infection triggers a strong inflammatory response associated with excessive production of reactive oxygen species (ROS), including superoxide anions, hydrogen peroxide, and hydroxyl radicals, by activated macrophages and neutrophils at the site of infection [4]. This oxidative reaction overwhelms host antioxidant defenses, leading to lipid peroxidation, DNA damage, and tissue injury, contributing to the characteristic neurological deterioration observed in late-stage HAT [5]. Conversely, the parasite itself relies on a thiol-based antioxidant system, centered on trypanothione to neutralize ROS generated both by its own metabolism and by the host immune response. Disruption of this system is lethal to the parasite, reinforcing the relevance of redox biology as a therapeutic target [6]. Plant-derived antioxidants therefore hold dual promise; they may attenuate host oxidative damage while simultaneously interfering with parasite antioxidant defense or other essential biological functions.
Currently available antitrypanosomal drugs including pentamidine, suramin, melarsoprol, eflornithine, and fexinidazole are associated with significant limitations such as complex parenteral administration, severe toxic side effects, emerging drug resistance, and high costs inaccessible to the most vulnerable populations [7,8]. These constraints highlight the urgent need for novel, safe, and affordable therapeutic agents derived from renewable natural sources.
At the molecular level, trypanosomes regulate gene expression almost exclusively by post-transcriptional mechanisms. RNA-binding proteins (RBPs) are central regulators of mRNA stability, trans-splicing, polyadenylation, and translation [9,10]. DRBD18 (Double RRM-containing protein 18; UniProt ID: Q57XR9) is an essential T. brucei RBP that regulates mRNA trans-splicing and polyadenylation of transcripts containing polypyrimidine tracts in their 3’-UTR [11]. DRBD18 has also been identified as a transcript-specific RNA editing auxiliary factor [12]. Its depletion by RNA interference severely impairs parasite growth, making it a compelling drug target. Polyphenolic compounds, known for their strong antioxidant properties, have been proposed as candidates capable of disrupting RRM domain–RNA interactions through hydrogen bonding and aromatic stacking [13,14]
In this study, two Central African medicinal plants were selected, Tetradenia riparia (Hochst.) Codd (Lamiaceae), widely used for its insecticidal, antiparasitic, antimalarial, and anti-inflammatory properties [15,16] and Tetracera poggei Gilg (Dilleniaceae), a lesser-studied specie whose leaves are traditionally used for infectious diseases in Central Africa, with no prior UPLC-MS or computational study reported [17]. Previous work from our group identified rosmarinic acid as a major compound of T. riparia [18,19].
The present study aims to (i) characterize the phytochemical composition of T. riparia and T. poggei by TLC screening, quantitative spectrophotometry, and UPLC-QTOF-MS; (ii) evaluate the antioxidant activity of decocted and percolated extracts by DPPH and ABTS assays; and (iii) perform in silico molecular docking of the major identified compounds against DRBD18 RRM1 and RRM2 domains, to identify candidate inhibitors with dual antioxidant and antiparasitic potential.
2. Materials and Methods
2.1. Plant Collection and Preparation
Leaves of Tetradenia riparia were collected between March and April 2025 in the Ngansele district, Mont-Ngafula commune, Kinshasa, DRC (S 4°26’17.197”; E 15°17’32.812”; altitude 0 m). Leaves of Tetracera poggei (local names: Bojo, Monji) were collected in the Kimwenza area, Mont-Ngafula commune (S 4°27’4.251”; E 15°17’37.693”). Plant collection adhered to all ethical guidelines; permission was obtained from the Faculty of Sciences and Technologies, University of Kinshasa. Voucher specimens were deposited at the Herbarium of Kinshasa (IUK), INERA-RDC. Collected plant material was oven-dried (Raypa) at 40 °C for 4 days and ground using an electric grinder (Blinder Butterfly B-592). Geographic collection sites are shown in Figure 1.
2.2. Preparation of Extracts
Extracts were prepared by decoction and percolation following the protocol of Ngolo et al. (2025) [19] with minor modifications. For decoction, 10 g of powder were mixed with 100 mL of distilled water, heated at 100 °C for 10 min, cooled, and filtered through Whatman filter paper. For percolation, 10 g of powder were macerated cold in 50 mL of dichloromethane/methanol (1:1, v/v) for 48 h and the percolate collected dropwise. Both extracts were evaporated and stored as dry extracts at 4 °C. Extraction yield (R) was calculated as:
R (%) = (Mex / Mmv) × 100, where Mex is the dry residue mass and Mmv is the initial dry plant material mass.
2.3. Phytochemical Screening
Qualitative phytochemical screening was performed on the aqueous and organic extracts following the methods of Hamilton-Amachree et al. (2024) [20], Shaikh (2020) [21], and Tourabi et al. (2025) [22], detecting phenols, flavonoids, alkaloids, saponins, quinones, terpenoids, anthraquinones, tannins, anthocyanins, leucoanthocyanins, and steroids. Total polyphenol content was determined by the Folin–Ciocalteu method with gallic acid as standard [23]. Total flavonoid content was quantified by the AlCl3 colorimetric method [23]. All measurements were performed in triplicate, and results are expressed as mean ± SD.
2.4. UPLC-QTOF-MS Analysis
Compound identification was carried out following the protocol of Ngolo et al. (2025) [19]. Dry extract (1 mg) was re-dissolved in 1 mL MeOH/water (1:1) and filtered through a 0.22 µm nylon syringe filter. Analysis was performed on a Waters Acquity UPLC system (Waters BEH C18 column, 2.1 × 100 mm, 1.7 µm) coupled to a Waters Synapt G2 QTOF mass spectrometer in negative ESI mode, as described previously [19]. Chromatographic separation used a Waters BEH C18 column (2.1 mm × 100 mm, 1.7 µm) with a gradient of solvent A (water + 0.1% formic acid) and solvent B (methanol + 0.1% formic acid), applied as follow: 0 min, 3% B; 0.10 min, 3% B; 14 min, 100% B; 16 min, 100% B; 16.5 min, 3% B; 20 min, 3% B. The flow rate was set at 0.3 mL/min. The injection volume was 5 µL. The following ESI source parameters were used for negative mode: source temperature 120 °C; sampling cone 20 V; extraction cone 4.0 V; desolvation temperature 300 °C; cone gas 10.0 L/h; desolvation gas 600 L/h; capillary 2.6 kV; trap collision energy 28 V. Molecular formulae were generated with MassLynx V4, and tentative identifications were made by comparing fragmentation patterns with the KEGG and PubChem databases via MetFrag (https://msbi.ipb-halle.de/MetFrag/).
2.5. Antioxidant Activity
Radical scavenging activity was evaluated by DPPH (2,2-diphenyl-1-picrylhydrazyl) and ABTS (2,2’-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid)) assays as described by Bukatuka et al. [24] with minor modifications. For DPPH, 1980 µL of 0.1 mM DPPH in methanol was mixed with 20 µL of extract at varying concentrations, incubated in the dark for 30 min, and absorbance read at 517 nm. For ABTS, the radical cation was generated by reacting 20 mM ABTS with 10 mM K2S2O8 (1:1) for 16 h in the dark; the working solution was diluted to an absorbance of 0.8–1.0, mixed with extract, and read at 734 nm after 30 min. IC50 values were calculated from inhibition–concentration curves using GraphPad Prism 9.5.1.733. All measurements were performed in triplicate.
2.6. Molecular Docking Studies
2.6.1. Protein Preparation
T. brucei DRBD18 (UniProt: Q57XR9; gene: Tb927.11.14090) was selected as the target protein [12]. The AlphaFold-predicted model (AF-Q57XR9-F1, version 6) was retrieved from the AlphaFold Protein Structure Database [25] and visualized in PyMOL (v2.5) and NGL Viewer. The model comprises 480 residues with two RRM domains: RRM1 (residues 90–170) and RRM2 (residues 260–340). Low-confidence terminal regions (low pLDDT scores) were excluded; water molecules and non-standard atoms were removed. Binding pocket coordinates were as follows: RRM1 (x = 3, y = 12, z = −25) and RRM2 (x = −34, y = 3, z = 24).
2.6.2. Ligand Selection and Docking
Ligands were selected from UPLC-MS profiles based on peak abundance and structural diversity. For T. riparia, rosmarinic acid, astragalin, luteolin, acacetin, asiatic acid, diterpenoid SP-II, abieta-8,11,13-triene-7,15,18-triol, and trihydroxy octadecenoic acid were used. For T. poggei, epi-gallocatechin 3-O-gallate, catechin 3-O-gallate, (−)-epicatechin-3-(3-O-methyl)-gallate, isorhamnetin, 7-O-methylquercetin, 7-O-methylkaempferol, galloyl-hexose-sulfate, and 1-O-palmitoyl-3-O-(6-sulfo-α-D-quinovopyranosyl)-glycerol were considered. Three-dimensional structures were retrieved from PubChem and optimized with Open Babel. Molecular docking was performed with AutoDock Vina in PyRx 0.9.8 [26]. The lowest binding energy pose was retained. Ligand efficiency (LE), Fit Quality (FQ), Binding Efficiency Index (BEI), and estimated Ki values were calculated. Interactions were visualized using BIOVIA Discovery Studio Visualizer.
3. Results
3.1. Qualitative Phytochemical Screening
Table 1 summarizes the qualitative phytochemical composition of the aqueous and organic extracts of T. riparia and T. poggei. Polyphenols, flavonoids, anthocyanins, tannins, bound quinones, alkaloids, leucoanthocyanins, and saponins were detected in the aqueous extracts of both species. In the organic fractions, steroids were present in both species, while triterpenoids were identified exclusively in T. riparia and free quinones exclusively in T. poggei. Tannins were absent from the aqueous extract of T. poggei.
3.2. Total Phenolic and Flavonoid Content
T. poggei exhibited significantly higher polyphenol and flavonoid contents (polyphenols: 510.227 ± 74.608 mgGAE/gDW; flavonoids: 855.394 ± 5.292 mgQE/gDW) than T. riparia (polyphenols: 331.56 ± 89.22 mgGAE/gDW; flavonoids: 445.727 ± 1.8 mgQE/gDW) (Table 2).
3.3. UPLC-QTOF-MS Metabolite Profiling
UPLC-QTOF-MS in negative ESI mode tentatively identified eleven compounds in T. riparia (Figure 2, Table 3) and thirteen compounds in T. poggei (Figure 3, Table 4).
In Tetradenia riparia methanolic extracts (Figure 2), rosmarinic acid (m/z 359.0772, RT 6.85 min, error 1.4 ppm) was the dominant polar constituent, consistent with Ngolo et al. (2025) [19]. Caffeic acid (m/z 179.0337) and its biosynthetic precursor were also detected. Flavonoids were represented by astragalin (m/z 447.0926), luteolin (m/z 285.0400), acacetin (m/z 283.0617), and nepetoidin A or isomer (m/z 313.0707). Terpenoids included asiatic acid (m/z 487.3438), diterpenoid SP-II (m/z 335.2220), and abieta-8,11,13-triene-7,15,18-triol (m/z 317.2119). Trihydroxy octadecenoic acid (m/z 327.2168) and dibutyl succinate (m/z 229.1449) were also identified. This metabolic diversity is consistent with earlier reports for T. riparia [15,16]. The compounds tentatively identified in the extracts from Tetradenia riparia are listed in Table 3.
In Tetracera poggei (Figure 3), the profile is dominated by galloylated catechin derivatives including epi-gallocatechin 3-O-gallate (m/z 457.0765, error −1.3 ppm), catechin 3-O-gallate (m/z 441.0818), and (−)-epicatechin-3-(3-O-methyl)-gallate (m/z 455.0967), alongside simpler phenolic acids such as gallic acid (m/z 169.0134), protocatechuic acid (m/z 153.0184), and methyl 6-O-galloyl-β-D-glucopyranoside (m/z 345.0818).
Methylated flavonols included isorhamnetin or isomer (m/z 315.0512), 7-O-methylquercetin or isomer (m/z 315.0498), and 7-O-methylkaempferol (m/z 299.0554). Sulphated compounds galloyl-hexose-sulfate (m/z 415.2162) and 1-O-palmitoyl-3-O-(6-sulfo-α-D-quinovopyranosyl)-glycerol (m/z 555.2838) were detected at later retention times. This chemical profile has not previously been reported for T. poggei and represents a significant expansion of the known phytochemistry of the genus Tetracera [17]. The compounds tentatively identified in the extracts from Tetradenia poggei are listed in Table 4.
3.4. Antioxidant Activity
The antioxidant activities of decocted and percolated extracts are summarized in Table 5. Percolated extracts consistently showed higher radical scavenging activity than decocted extracts in both species. For DPPH, T. poggei percolated extract showed the strongest activity (IC50 = 4.141 ± 0.175 µg/mL), followed by T. riparia percolated extract (7.211 ± 0.222 µg/mL). In the ABTS assay, T. riparia percolated extract showed higher activity (IC50 = 10.79 ± 0.56 µg/mL) than T. poggei (13.99 ± 0.15 µg/mL). All IC50 values were below 100 µg/mL.
3.5. Molecular Docking Against DRBD18
3.5.1. T. poggei Compounds
Table 6 shows the docking results for T. poggei compounds. Among the eight compounds evaluated, (−)-epicatechin-3-(3-O-methyl)-gallate showed the highest RRM1 affinity (ΔG = −7.3 kcal/mol; Ki = 4.424 µM), followed by epi-gallocatechin 3-O-gallate (−7.2 kcal/mol; Ki = 4.645 µM) and catechin 3-O-gallate (−7.1 kcal/mol; Ki = 6.202 µM). In the RRM2 domain, catechin 3-O-gallate showed the highest affinity (−6.7 kcal/mol; Ki = 12.187 µM). Methylated flavonols showed moderate affinities (−5.7 to −6.6 kcal/mol for RRM1). The sulphated lipid derivatives showed the weakest binding (−4.5 to −6.1 kcal/mol).
The interaction profiles of epi-gallocatechin-3-O-gallate (EGCG) at RRM1 and catechin-3-O-gallate (C3G) at RRM2 reveal distinct but well-defined binding characteristics (Figure S1). Within the RRM1 binding pocket, EGCG forms conventional hydrogen bonds with residues GLY186, TYR177, ARG180, THR182, and GLU185, along with a π-sigma interaction with GLU181, π-alkyl contacts with LEU179 and ARG178, and a T-shaped π–π interaction with PHE188 (Figure 4). This pattern highlights the combined role of hydrogen bonding and aromatic contacts in RRM1 binding.
In contrast, the C3G interaction network at RRM2 is more complex and diverse; the compound forms a carbon hydrogen bond with THR232, conventional hydrogen bonds with ALA230, GLN243, and GLU231, a π-cation interaction with ARG247, and π–π stacking with TRP233, supplemented by π-alkyl interactions with ARG244 and LEU240 (Figure 5). This combination of electrostatic, aromatic, and hydrophobic interactions suggests a more balanced and versatile binding mode at RRM2.
Interaction distances generally fall in the range of approximately 1.6–4.8 Å, indicating favorable spatial positioning of the ligands within both binding pockets.
3.5.2. T. riparia Compounds
Table 7 shows the docking results for T. riparia compounds. Diterpenoid SP-II showed the highest RRM1 binding affinity (ΔG = −7.3 kcal/mol; Ki = 4.424 µM), followed by asiatic acid (−7.2 kcal/mol; Ki = 4.645 µM), astragalin (−6.8 kcal/mol), and abieta-8,11,13-triene-7,15,18-triol (−6.8 kcal/mol). Phenylpropanoids and flavonoids including rosmarinic acid (−6.7 kcal/mol), luteolin (−6.6 kcal/mol) and acacetin (−6.5 kcal/mol) also showed moderate to strong RRM1 binding. In RRM2, diterpenoid SP-II (−6.1 kcal/mol) and acacetin (−6.0 kcal/mol) formed the most stable complexes. Trihydroxy octadecenoic acid showed the lowest affinities (RRM1: −3.5 kcal/mol; RRM2: −2.9 kcal/mol; Ki > 2,700 µM).
The interaction profile of diterpenoid SP-II with RRM1 and RRM2 reveals limited but distinct contacts with both domains (Figure S2). Within the RRM1 binding site, the compound forms an alkyl interaction with LEU179 and conventional hydrogen bonds involving GLY186 and THR182, reflecting structural complementarity between the diterpenoid scaffold and the RRM1 pocket (Figure 6).
In the RRM2 binding region, a more restricted interaction pattern is observed: the compound establishes a π-alkyl interaction with TRP233, contributing to hydrophobic stabilization, and forms a conventional hydrogen bond with THR232 (Figure 7).
4. Discussion
The rich phytochemical diversity of both T. riparia and T. poggei revealed by qualitative screening is consistent with their traditional medicinal uses. The co-occurrence of flavonoids, polyphenols, tannins, and alkaloids suggests marked antioxidant, antimicrobial, and antiparasitic potential [27,28]. The exclusive detection of triterpenoids in T. riparia and free quinones in T. poggei indicates distinct metabolic specialization between the two species. Our results for T. riparia are consistent with Hamilton-Amachree et al. (2024) [20], with a minor divergence regarding anthraquinones.
T. poggei showed significantly higher polyphenol and flavonoid contents than T. riparia. These values exceed those reported by Chepng’etich et al. (2018) [29] and Ngolo et al. (2025) [19] for T. riparia. The variability may be attributed to harvest season, soil pH, drying conditions, geographic location, and extraction method [30]. The particularly high flavonoid content of T. poggei is consistent with the dense array of galloylated catechin derivatives subsequently identified by UPLC-MS. Phenolic compounds are known to modulate gene expression and exhibit antioxidant, antimutagenic, and antiparasitic activities [13].
UPLC-QTOF-MS analysis revealed complementary chemical profiles between the two species. In T. riparia, rosmarinic acid is the dominant polyphenol, already identified as a major constituent by Ngolo et al. [19]. Luteolin, acacetin, and astragalin are well-documented antioxidants. The terpenoid fraction containing diterpenoid SP-II, asiatic acid, and the abietane diterpene, represents structurally diverse scaffolds not previously described by UPLC-MS in this species. In T. poggei, the dominance of EGCG, catechin 3-O-gallate, and (−)-epicatechin-3-(3-O-methyl)-gallate is noteworthy; the galloyl ester moiety of these compounds is a well-established structural determinant of both antioxidant capacity and protein-binding affinity [31,32]. This chemical profile has not previously been reported for T. poggei [17].
Percolated extracts consistently outperformed decocted extracts in both DPPH and ABTS assays, likely reflecting the superior extraction efficiency of the dichloromethane/methanol solvent system for polyphenolic and terpenoid compounds. The higher DPPH activity of T. poggei (IC50 = 4.141 µg/mL) correlates with its greater polyphenol and flavonoid content, and with the strong hydrogen-donating capacity of its galloyl ester compounds [33]. The divergence between DPPH and ABTS rankings reflects the mechanistic differences between the two assays; DPPH primarily measures hydrogen atom transfer from hydrophilic compounds, while ABTS captures both hydrophilic and lipophilic antioxidants, explaining the relatively higher ABTS performance of T. riparia, which contains lipophilic diterpenoids [34]. The IC50 of T. riparia percolated extract by DPPH (7.211 ± 0.222 µg/mL) is superior to the value reported by Ngolo et al. (2025) [19], a difference attributable to seasonal, geographic, or protocol variations. Phenolic compounds particularly rosmarinic acid, gallic acid, caffeic acid, and the galloylated catechins are known to be the primary determinants of antioxidant capacity [35], consistent with our positive correlation between polyphenol/flavonoid content and IC50 values.
Importantly, many of the identified antioxidant compounds in both species are the same molecules that show the strongest binding affinities in the molecular docking analysis. This dual antioxidant–antiparasitic profile is mechanistically coherent; the structural features responsible for antioxidant activity including multiple hydroxyl groups, aromatic rings, and electron-donating substituents are the same features that drive hydrogen bonding and aromatic stacking interactions with RRM domain residues. EGCG and catechin 3-O-gallate are among the best-characterized RRM-binding polyphenols in the literature [31,32]. In T. poggei, (−)-epicatechin-3-(3-O-methyl)-gallate achieved the highest RRM1 binding energy (ΔG = −7.3 kcal/mol; Ki = 4.424 µM). Interaction analysis of EGCG at RRM1 reveals classical hydrogen bonds with GLY186, TYR177, ARG180, THR182, and GLU185, along with π-sigma (GLU181), π-alkyl (LEU179, ARG178), and T-shaped π–π (PHE188) interactions. The involvement of PHE188, a conserved aromatic residue in the RRM β-sheet that normally participates in RNA base stacking, strongly suggests direct competition with RNA substrates [31,32]. At RRM2, catechin 3-O-gallate forms an extensive network including a π-cation interaction with ARG247 and π–π stacking with TRP233.
In T. riparia, diterpenoid SP-II (ΔG = −7.3 kcal/mol) and asiatic acid (−7.2 kcal/mol) achieved RRM1 affinities comparable to the best polyphenols, through a different mechanistic basis: primarily hydrophobic and van der Waals contacts rather than hydrogen bonding. Diterpenoid SP-II at RRM1 forms alkyl interactions with LEU179 and hydrogen bonds with GLY186 and THR182; at RRM2, it engages π-alkyl with TRP233 and a hydrogen bond with THR232. This terpenoid binding mode is consistent with recent evidence that natural diterpenoids interact with RNA-binding proteins through hydrophobic surface complementarity [38]. The convergence of comparable RRM1 binding energies from structurally distinct chemical classes suggests that DRBD18 accommodates a broader range of ligand scaffolds than expected from a purely polar binding interface.
These in silico findings require experimental validation. Key limitations include the use of an AlphaFold structural model rather than an experimentally resolved crystal structure, the static nature of docking, which does not account for protein flexibility or solvation effects, and the absence of mammalian homologue selectivity data. Future studies should include in vitro RNA-binding inhibition assays, cellular anti-trypanosomal activity testing, and in vivo evaluation.
5. Conclusions
This study provides the first integrated antioxidant, phytochemical, and in silico characterization of Tetradenia riparia and Tetracera poggei as sources of dual-action compounds with antioxidant and antiparasitic potential. Both species displayed significant antioxidant activity (all IC50 < 100 µg/mL), with T. poggei percolated extracts achieving the highest DPPH activity (IC50 = 4.141 ± 0.175 µg/mL). UPLC-QTOF-MS identified complementary metabolite profiles including rosmarinic acid, diterpenoids, and flavonoids dominating in T. riparia, and galloylated catechin derivatives in T. poggei. Molecular docking against DRBD18 identified diterpenoid SP-II and (−)-epicatechin-3-(3-O-methyl)-gallate as the top candidates (RRM1: ΔG = −7.3 kcal/mol; Ki = 4.424 µM), with a consistent structure-activity relationship linking polyphenolic and terpenoid complexity with RRM binding affinity. The dual antioxidant–antiparasitic profile of the identified compounds provides a mechanistic basis for the traditional use of these plants and a rational foundation for the development of plant-derived trypanocidal agents targeting DRBD18. Experimental in vitro and in vivo validation remains an essential next step.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
WBB: Conceptualization, investigation, laboratory work and writing original draft. OMK: investigation and laboratory work. SMM: UPLC-MS analyses and methodology. ECA: Computational analyses and molecular docking. JPNKN and JIB: Supervision and critical review. TBT: Conceptualization, investigation, supervision, methodology, funding acquisition, writing, review and editing. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This publication was possible thanks to the Seed Grant for New African Principal Investigators (SG-NAPI) awarded to Tania Bishola Tshitenge (2024-2026), supported by the German Ministry of Education and Research (BMBF) through UNESCO-TWAS. We are indebted to the University of Kinshasa for institutional support and to all collaborators who contributed to data collection and analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Venturelli, A.; Tagliazucchi, L.; Lima, C.; et al. Current Treatments to Control African Trypanosomiasis and One Health Perspective. Microorganisms 2022, 10, 1–24. [Google Scholar] [CrossRef]
- Cecchi, Giuliano; Ngari, Nancy; Paone, Massimo; et al. Disease intelligence and modelling for progressive control of animal trypanosomosis in Africa. Trends Parasitol. 2025, 41, 734–745. [Google Scholar] [CrossRef] [PubMed]
- Hasker, E.; Lutumba, P.; Chappuis, F.; et al. Human African Trypanosomiasis in the Democratic Republic of the Congo: A Looming Emergency? PLoS Negl. Trop. Dis. 2012, 6, 1–3. [Google Scholar] [CrossRef]
- Kasozi, K.I.; MacLeod, E.T.; Ntulume, I.; et al. An Update on African Trypanocide Pharmaceutics and Resistance. Front. Vet. Sci. 2022, 9, 1–23. [Google Scholar] [CrossRef]
- Vincendeau, P.; Bouteille, B. Immunology and immunopathology of African trypanosomiasis. Ann. Braz. Acad. Sci. 2006, 78, 645–665. [Google Scholar] [CrossRef]
- Fairlamb, A.H.; Horn, D. Melarsoprol Resistance in African Trypanosomiasis. Trends Parasitol. 2018, 34, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Alvarado, M.; Basáñez, M.G.; et al. The Global Burden of Disease Study 2010: Interpretation and Implications for the Neglected Tropical Diseases. PLoS Negl. Trop. Dis. 2014, 8, 1–9. [Google Scholar] [CrossRef]
- Pereira, S.H.; Alves, F.P.; Teixeira, S.M.R. Animal Trypanosomiasis: Challenges and Prospects for New Vaccination Strategies. Microorganisms 2024, 12, 1–17. [Google Scholar] [CrossRef]
- Clayton, C.E. Networks of gene expression regulation in Trypanosoma brucei. Mol. Biochem Parasitol. 2014, 2, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Clayton, C. Regulation of gene expression in trypanosomatids: Living with polycistronic transcription. Open Biol. 2019, 9. [Google Scholar] [CrossRef]
- Tshitenge, T.B.; Clayton, C. The Trypanosoma brucei RNA-binding protein DRBD18 ensures correct mRNA trans splicing and polyadenylation patterns. RNA 2022, 28, 1239–1262. [Google Scholar] [CrossRef]
- Pandey, P.; Wackowski, K.; Dubey, A.P.; et al. DRBD18 acts as a transcript-specific RNA editing auxiliary factor in Trypanosoma brucei. RNA 2025, 31, 245–257. [Google Scholar] [CrossRef]
- Xu, L.; Wang, X. A Comprehensive Review of Phenolic Compounds in Horticultural Plants. Int. J. Mol. Sci. 2025, 26, 1–25. [Google Scholar] [CrossRef]
- Lešnik, S.; Jukić, M.; Bren, U. Unveiling polyphenol-protein interactions: a comprehensive computational analysis. J. Cheminform 2025, 17, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Gazim, Z.C.; Swain, S.S.; et al. Ethnomedicinal, Phytochemical and Pharmacological Investigations of Tetradenia riparia (Hochst.) Codd (Lamiaceae). Front Pharmacol. 2022, 13, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Sena J da, S.; Rodrigues, S.A.; Sakumoto, K.; et al. Antioxidant Activity, Antiproliferative Activity, Antiviral Activity, NO Production Inhibition, and Chemical Composition of Essential Oils and Crude Extracts of Leaves, Flower Buds, and Stems of Tetradenia riparia. Pharmaceuticals 2024, 17, 1–19. [Google Scholar] [CrossRef]
- Ogunlakin, A.D.; Sonibare, M.A. Phytochemistry and biological activities of Tetracera species. Trends Phytochem. Res. 2022, 6, 339–352. [Google Scholar]
- Konde, O.M.; Balela, W.B.; Tshitenge, T.B. Targeting RNA-binding proteins with small molecules: perspectives and challenges. Front. Chem. 2025, 13. [Google Scholar] [CrossRef]
- Ngolo, L.M.; Faraja, F.M.; Ngandu, O.K.; et al. Phytochemical screening, UPLC analysis, evaluation of synergistic antioxidant and antibacterial efficacy of three medicinal plants used in Kinshasa, D.R. Congo. Sci. Rep. Epub ahead of print 1 December 2025. 15. [CrossRef]
- Hamilton-Amachree, A.; A. Fadare, D.; Uzoekwe, N.M.; et al. Phytochemicals, Nutritional and Antibacterial Evaluation of a Domestic Plant –Tetradenia riparia. Al-Qadisiyah J. Pure Sci. 2024, 29, 94–98. [Google Scholar] [CrossRef]
- Shaikh, J.R.; Patil, M. Qualitative tests for preliminary phytochemical screening: An overview. Int. J. Chem. Stud. 2020, 8, 603–608. [Google Scholar] [CrossRef]
- Tourabi, M.; Faiz, K.; Ezzouggari, R.; et al. Optimization of extraction process and solvent polarities to enhance the recovery of phytochemical compounds, nutritional content, and biofunctional properties of Mentha longifolia L. extracts. Bioresour. Bioprocess Epub ahead of print. 2025, 12. [Google Scholar] [CrossRef]
- Mbadiko, C.M.; Ngbolua, K.-T.-N.; Mpiana, P.T.; et al. Antioxidant Potential and Anti-Sickling Activity of Different Organs of Curcuma longa: Correlation of the Antioxidant Capacity on Anti-Sickling Activity. South Asian Res. J. Nat. Prod. 2019, 2, 1–16. [Google Scholar]
- Bukatuka, F.; Ngombe, K.; Mutwale, K.; et al. Bioactivity and Nutritional Values of Some Dioscorea Species Traditionally Used as Medicinal Foods in Bandundu, DR Congo. Eur. J. Med. Plants 2016, 14, 1–11. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: an overview. Futur J. Pharm. Sci. 2021, 7, 1–13. [Google Scholar] [CrossRef]
- Caruso, G.; Godos, J.; Privitera, A.; et al. Phenolic Acids and Prevention of Cognitive Decline: Polyphenols with a Neuroprotective Role in Cognitive Disorders and Alzheimer’s Disease. Nutrients 2022, 14, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Chepng’etich, J.; Ngule, C.; Jepkorir, M.; et al. Total Phenolic Content and in vitro Antiproliferative Activity of Tragia brevipes (Pax) and Tetradenia riparia (Hochst) Leaves Extract. Eur. J. Med. Plants 2018, 22, 1–10. [Google Scholar] [CrossRef]
- Daniels, J.-P.; Gull, K.; Wickstead, B. Cell Biology of the Trypanosome Genome. Microbiol. Mol. Biol. Rev. 2010, 74, 552–569. [Google Scholar] [CrossRef]
- Saeki, K.; Hayakawa, S.; Nakano, S.; et al. In vitro and in silico studies of the molecular interactions of epigallocatechin-3-o-gallate (egcg) with proteins that explain the health benefits of green tea. Molecules 2018, 23, 1–24. [Google Scholar] [CrossRef]
- Hirci, J.; Škufca, S.; Kunej, T.; et al. Identification of potential human targets for epigallocatechin gallate through a novel protein binding site screening approach. J. Mol. Model 2025, 31, 1–2. [Google Scholar] [CrossRef]
- Al-Mudhafar, M.M.J.; Wadi, J.S. Synthesis and biological activity evaluation of new isatin-gallate hybrids as antioxidant and anticancer agents (in vitro) and in silico study as anticancer agents and coronavirus inhibitors. Pharmacia 2024, 71, 1–11. [Google Scholar] [CrossRef]
- Cano, A.; Maestre, A.B.; Hernández-Ruiz, J.; et al. ABTS/TAC Methodology: Main Milestones and Recent Applications. Processes 2023, 11, 1–10. [Google Scholar] [CrossRef]
- Kaviya, M.; Balamuralikrishnan, B.; Sangeetha, T.; et al. Evaluation of phytoconstituents of Triticum aestivum grass extracts on nutritional attributes, antioxidant, and antimicrobial activities against food pathogens with molecular in silico investigation. Food Front 2023, 4, 831–848. [Google Scholar] [CrossRef]
Figure 1.
Geographical location of the collection sites of the leaves of Tetradenia riparia and Tetracera poggei.
Figure 1.
Geographical location of the collection sites of the leaves of Tetradenia riparia and Tetracera poggei.

Figure 2.
ESI negative mode BPI chromatogram of the Tetradenia riparia methanolic extract.

Figure 3.
ESI negative mode BPI chromatogram of the Tetracera poggei methanolic extract.

Figure 4.
Molecular docking interaction profile of epi-gallocatechin-3-O-gallate with RRM1, showing (A) the 3D binding conformation in the active site and (B) a detailed 2D interaction map highlighting hydrogen bonds, van der Waals forces, and π-related interactions with key amino acid residues.
Figure 4.
Molecular docking interaction profile of epi-gallocatechin-3-O-gallate with RRM1, showing (A) the 3D binding conformation in the active site and (B) a detailed 2D interaction map highlighting hydrogen bonds, van der Waals forces, and π-related interactions with key amino acid residues.

Figure 5.
Molecular docking interaction profile of catechin-3-O-gallate with RRM2, illustrating (A) the 3D binding pose in the active site and (B) the 2D interaction diagram highlighting hydrogen bonds, π-cation, π–π stacking, and hydrophobic interactions with key amino acid residues.
Figure 5.
Molecular docking interaction profile of catechin-3-O-gallate with RRM2, illustrating (A) the 3D binding pose in the active site and (B) the 2D interaction diagram highlighting hydrogen bonds, π-cation, π–π stacking, and hydrophobic interactions with key amino acid residues.

Figure 6.
Molecular docking interaction profile of diterpenoid SP-II with RRM1, demonstrating (A) the 3D binding conformation in the active site and (B) the 2D interaction map highlighting hydrogen bonds, van der Waals forces, alkyl interactions, and donor–donor unfavorable contacts with key amino acid residues.
Figure 6.
Molecular docking interaction profile of diterpenoid SP-II with RRM1, demonstrating (A) the 3D binding conformation in the active site and (B) the 2D interaction map highlighting hydrogen bonds, van der Waals forces, alkyl interactions, and donor–donor unfavorable contacts with key amino acid residues.

Figure 7.
Molecular docking interaction profile of diterpenoid SP-II with RRM2, showing (A) the 3D binding pose in the active site and (B) the 2D interaction map highlighting hydrogen bonds, van der Waals forces, π-alkyl interactions, and unfavorable contacts with key amino acid residues.
Figure 7.
Molecular docking interaction profile of diterpenoid SP-II with RRM2, showing (A) the 3D binding pose in the active site and (B) the 2D interaction map highlighting hydrogen bonds, van der Waals forces, π-alkyl interactions, and unfavorable contacts with key amino acid residues.


Table 1.
Qualitative phytochemical screening of aqueous and organic extracts of T. riparia and T. poggei + indicates presence; − indicates absence.
Table 1.
Qualitative phytochemical screening of aqueous and organic extracts of T. riparia and T. poggei + indicates presence; − indicates absence.
| Phytochemical compounds |
T. riparia (Aqueous extract) |
T. riparia (Organic extract) |
T. poggei (Aqueous extract) |
T. poggei (Organic extract) |
|---|---|---|---|---|
| Polyphenols | + | — | + | — |
| Flavonoids | + | — | + | — |
| Anthocyanins | + | — | + | — |
| Tannins | + | — | − | — |
| Bound quinones | + | — | + | — |
| Alkaloids | + | — | + | — |
| Leucoanthocyanins | + | — | + | — |
| Saponins | + | — | + | — |
| Steroids | — | + | — | + |
| Free quinones | — | − | — | + |
| Triterpenoids | — | + | — | − |
Table 2.
Total polyphenol and flavonoid contents of the methanolic extracts (mean ± SD, n = 3).
| Species | Polyphenols (mgGAE/gDW) | Flavonoids (mgQE/gDW) |
|---|---|---|
| T. riparia | 331.56 ± 89.22 | 445.727 ± 1.8 |
| T. poggei | 510.227 ± 74.608 | 855.394 ± 5.292 |
GAE: Gallic Acid Equivalent; QE: Quercetin Equivalent; DW: Dry Weight.
Table 3.
Tentative identification of the major compounds in the methanolic extracts from Tetradenia riparia.
Table 3.
Tentative identification of the major compounds in the methanolic extracts from Tetradenia riparia.
| RT (min) | Exp. m/z [M-H]− | Mol. formula | Theor. m/z [M-H]− | Error (ppm) | Compound |
|---|---|---|---|---|---|
| 4.66 | 179.0337 | C9H8O4 | 179.0344 | -3.9 | Caffeic acid |
| 6.77 | 447.0926 | C21H20O11 | 447.0927 | -0.2 | Astragalin |
| 6.85 | 359.0772 | C18H16O8 | 359.0767 | 1.4 | Rosmarinic acid |
| 7.66 | 229.1449 | C12H22O4 | 229.1440 | 3.9 | Dibutyl succinate |
| 8.53 | 285.0400 | C15H10O6 | 285.0399 | 0.4 | Luteolin |
| 9.61 | 327.2168 | C18H32O5 | 327.2171 | -0.9 | Trihydroxy octadecenoic acid |
| 9.78 | 313.0707 | C17H14O6 | 313.0712 | -1.6 | Nepetoidin A or isomer |
| 10.60 | 283.0617 | C16H12O5 | 283.0606 | 3.9 | Acacetin |
| 11.66 | 487.3438 | C30H48O5 | 487.3423 | 3.1 | Asiatic acid |
| 12.18 | 335.2220 | C20H32O4 | 335.2222 | -0.6 | Diterpenoid SP-II |
| 12.82 | 317.2119 | C20H30O3 | 317.2117 | 0.6 | Abieta-8,11,13-triene-7,15,18-triol |
Table 4.
Tentative identification of the major compounds in the methanolic extracts from Tetradenia poggei.
Table 4.
Tentative identification of the major compounds in the methanolic extracts from Tetradenia poggei.
| RT (min) | Exp. m/z [M-H]− | Mol. formula | Theor. m/z [M-H]− | Error (ppm) | Compound |
|---|---|---|---|---|---|
| 0.90 | 191.0547 | C7H12O6 | 191.0556 | -4.7 | Quinic acid |
| 2.08 | 169.0134 | C7H6O5 | 169.0137 | -1.8 | Gallic acid |
| 2.90 | 345.0818 | C14H18O10 | 345.0822 | -1.2 | Methyl 6-O-galloyl-beta-D-glucopyranoside |
| 3.05 | 153.0184 | C7H6O4 | 153.0188 | -2.6 | Protocatechuic acid |
| 4.66 | 457.0765 | C22H18O11 | 457.0771 | -1.3 | epi-Gallocatechin 3-O-gallate (EGCG) |
| 5.50 | 441.0818 | C22H18O10 | 441.0822 | -0.9 | Catechin 3-O-gallate |
| 6.17 | 455.0967 | C23H20O10 | 455.0978 | -2.4 | (-)-Epicatechin-3-(3-O-methyl)-gallate |
| 7.70 | 315.0512 | C16H12O7 | 315.0505 | 2.2 | Isorhamnetin or isomer |
| 8.21 | 379.0121 | C16H12O9S | 379.0124 | -0.8 | Rhamnocitrin 3-O-sulfate |
| 9.73 | 315.0498 | C16H12O7 | 315.0505 | -2.2 | 7-O-Methylquercetin or isomer |
| 10.47 | 299.0554 | C16H12O6 | 299.0556 | -0.7 | 7-O-Methylkaempferol |
| 12.08 | 415.2162 | C21H36O6S | 415.2154 | 1.9 | Galloyl-hexose-sulfate |
| 13.64 | 555.2838 | C25H48O11S | 555.2839 | -0.2 | 1-O-Palmitoyl-3-O-(6-sulfo-alpha-D-quinovopyranosyl)-glycerol |
Table 5.
Antioxidant activities (IC50, µg/mL) of decocted and percolated extracts by DPPH and ABTS assays (mean ± SD, n = 3).
Table 5.
Antioxidant activities (IC50, µg/mL) of decocted and percolated extracts by DPPH and ABTS assays (mean ± SD, n = 3).
| Species | DPPH Decoction | DPPH Percolation | ABTS Decoction | ABTS Percolation |
|---|---|---|---|---|
| T. riparia | 14.826 ± 1.307 | 7.211 ± 0.222 | 18.14 ± 3.62 | 10.79 ± 0.56 |
| T. poggei | 8.713 ± 0.327 | 4.141 ± 0.175 | 7.044 ± 0.338 | 13.99 ± 0.15 |
Table 6.
Molecular docking results for T. poggei compounds against DRBD18 RRM1 and RRM2 domains.
| Compound | Domain | ΔG (kcal/mol) | LE | FQ | BEI | Ki (µM) |
|---|---|---|---|---|---|---|
| epi-Gallocatechin 3-O-gallate | RRM1 | -7.2 | 0.257 | 0.707 | 0.016 | 4.645 |
| RRM2 | -5.9 | 0.211 | 0.580 | 0.013 | 87.072 | |
| Catechin 3-O-gallate | RRM1 | -7.1 | 0.222 | 0.647 | 0.016 | 6.202 |
| RRM2 | -6.7 | 0.243 | 0.669 | 0.015 | 12.187 | |
| (-)-Epicatechin-3-(3-O-methyl)-gallate | RRM1 | -7.3 | 0.221 | 0.663 | 0.016 | 4.424 |
| RRM2 | -5.9 | 0.179 | 0.536 | 0.013 | 47.060 | |
| Isorhamnetin | RRM1 | -6.1 | 0.254 | 0.725 | 0.019 | 33.308 |
| RRM2 | -5.7 | 0.237 | 0.676 | 0.018 | 68.330 | |
| 7-O-Methylquercetin | RRM1 | -6.6 | 0.275 | 0.785 | 0.021 | 14.573 |
| RRM2 | -5.8 | 0.242 | 0.691 | 0.018 | 63.000 | |
| 7-O-Methylkaempferol | RRM1 | -6.2 | 0.282 | 0.563 | 0.021 | 28.354 |
| RRM2 | -5.7 | 0.259 | 0.517 | 0.019 | 65.969 | |
| Galloyl-hexose-sulfate | RRM1 | -6.1 | 0.227 | 0.517 | 0.015 | 33.308 |
| RRM2 | -5.2 | 0.194 | 0.441 | 0.013 | 162.041 | |
| 1-O-Palmitoyl-3-O-(6-sulfo-alpha-D-quinovopyranosyl)-glycerol | RRM1 | -4.8 | 0.136 | 0.377 | 0.009 | 388.063 |
| RRM2 | -4.5 | 0.127 | 0.352 | 0.008 | 802.955 |
BEI: Binding Efficiency Index; FQ: Fit Quality; Ki: estimated inhibition constant; LE: Ligand Efficiency.
Table 7.
Molecular docking results for T. riparia compounds against DRBD18 RRM1 and RRM2 domains.
| Compound | Domain | ΔG (kcal/mol) | LE | FQ | BEI | Ki (µM) |
|---|---|---|---|---|---|---|
| Rosmarinic acid | RRM1 | -6.7 | 0.258 | 0.614 | 0.019 | 12.187 |
| RRM2 | -5.1 | 0.196 | 0.467 | 0.014 | 181.722 | |
| Astragalin | RRM1 | -6.8 | 0.243 | 0.575 | 0.015 | 10.476 |
| RRM2 | -5.7 | 0.204 | 0.482 | 0.013 | 67.829 | |
| Luteolin | RRM1 | -6.6 | 0.275 | 0.734 | 0.023 | 14.573 |
| RRM2 | -5.9 | 0.246 | 0.657 | 0.021 | 87.082 | |
| Acacetin | RRM1 | -6.5 | 0.310 | 0.587 | 0.023 | 17.084 |
| RRM2 | -6.0 | 0.286 | 0.542 | 0.021 | 39.747 | |
| Asiatic acid | RRM1 | -7.2 | 0.207 | 0.621 | 0.015 | 4.645 |
| RRM2 | -5.8 | 0.167 | 0.501 | 0.012 | 63.000 | |
| Abieta-8,11,13-triene-7,15,18-triol | RRM1 | -6.8 | 0.243 | 0.619 | 0.021 | 10.476 |
| RRM2 | -5.7 | 0.203 | 0.518 | 0.018 | 67.829 | |
| Diterpenoid SP-II | RRM1 | -7.3 | 0.304 | 0.667 | 0.022 | 4.424 |
| RRM2 | -6.1 | 0.254 | 0.557 | 0.018 | 33.571 | |
| Trihydroxy octadecenoic acid | RRM1 | -3.5 | 0.152 | 0.392 | 0.011 | 2720.533 |
| RRM2 | -2.9 | 0.126 | 0.325 | 0.009 | 7456.653 |
BEI: Binding Efficiency Index; FQ: Fit Quality; Ki: estimated inhibition constant; LE: Ligand Efficiency.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



