Submitted:
21 May 2026
Posted:
28 May 2026
You are already at the latest version
Abstract
Bovine viral diarrhea (BVD) is a globally prevalent infectious disease in cattle caused by bovine viral diarrhea virus (BVDV) and poses a serious threat to the livestock industry. Clinical manifestations of BVD suggest that inflammatory responses are involved in the pathogenesis of BVDV, whereas the underlying molecular mechanisms remain unclear. As a key signaling hub of innate immunity, inflammasomes activate Caspase-1, thereby facilitating the cleavage of Gasdermin D (GSDMD), promoting membrane pore formation and triggering pyroptosis, thereby contributing critically to host antiviral immunity and viral pathogenesis. This study used Madin-Darby bovine kidney (MDBK) cells as a model to compare infection responses between the cytopathic BVDV NADL strain and the non-cytopathic TC strain. The results showed that both strains upregulate the expression of IL-1β and IL-18 and promote the assembly of the NLRP3 inflammasome, indicating activation of upstream inflammasome signaling; however, only the NADL strain markedly induced Gasdermin D (GSDMD) cleavage, membrane pore formation, and pyroptosis. Further mechanistic studies revealed that core protein C (from the NADL strain) interacts with the inflammasome adaptor protein ASC and promotes NLRP3 inflammasome activation, thereby enhancing the secretion of inflammatory cytokines. In summary, this study provides the molecular mechanism by which cytopathic BVDV activates NLRP3 inflammasome-mediated pyroptosis in an ASC-dependent manner through the core protein C, providing new insights into the inflammation-related pathogenic process of BVDV and the differences in infection outcomes of different viral biotypes.
Keywords:
bovine viral diarrhea virus
; NLRP3 inflammasome
; ASC
; pyroptosis
1. Introduction
Bovine viral diarrhea (BVD), as a major infectious disease affecting cattle worldwide, has long imposed substantial economic burdens on the cattle industry [1]. Its pathogen, bovine viral diarrhea virus (BVDV), has complex biological characteristics, including the existence of different biotypes, the capacity to cross the placental barrier, and the capacity to establish persistent infections, which result in sustained viral circulation in natural populations and hinder effective eradication by conventional control measures [2,3]. BVDV infection can cause the host to exhibit a series of clinical symptoms such as fever, diarrhea, reproductive disorders, and lethal mucosal disease, and the pathological process is often accompanied by marked inflammatory responses, such as leukopenia and tissue inflammatory infiltration [4]. This suggests that host immune responses, particularly inflammatory responses, play a critical role in BVDV pathogenesis [5,6]. However, the inflammatory responses and infection outcomes caused by different biotypes of BVDV show significant differences, and their underlying molecular mechanisms remain poorly understood.
BVDV belongs to the family Flaviviridae and the genus Pestivirus [7], When the virus enters host cells, its single-stranded positive-sense RNA genome is first translated to produce a large polyprotein precursor; which is then proteolytically processed into mature viral proteins by host signal peptidases and viral proteases [8]. These proteins are classified into structural proteins (C, Erns, E1, and E2) and non-structural proteins (Npro, p7, NS2/3, NS4A, NS4B, NS5A, and NS5B) [9,10,11], These viral proteins not only constitute key components of the viral replication complex, directly participating in genome replication and virion assembly, but also modulate host cellular signaling pathways and immune responses through diverse mechanisms, thereby contributing to the molecular basis of BVDV pathogenesis [12].
Based on whether it can cause significant cytopathic effects in cultured cells in vitro, BVDV is generally classified into two classical biotypes: cytopathogenic (CP) and non-cytopathogenic (NCP), This difference in biological characteristics is closely associated with infection outcomes in vivo [13,14]. Epidemiological evidence indicates that NCP BVDV is the predominant strain circulating in natural populations, and after infecting pregnant dams, it can cross the placental barrier to infect the fetus [15]. Owing to the immature state of the fetal immune system, it recognizes the virus as a self component, thereby forming immune tolerance, resulting in calves born as persistently infected (PI) animals that harbor and shed the virus throughout life [16]. PI animals are the primary reservoirs for the sustained transmission of BVDV within cattle herds. In contrast, CP BVDV usually cannot establish persistent infection, but when it subsequently infects PI animals, it is often a key factor in triggering fatal mucosal disease[17]. Although this macro-level pathogenic pattern has been recognized for many years, the molecular mechanisms underlying the differences in biological behavior between CP and NCP BVDV, especially their differential regulation of host inflammatory responses, still remain poorly understood.
The Madin-Darby bovine kidney (MDBK) cell is highly susceptible to different BVDV biotypes, and can stably support efficient viral replication [18,19], thus, it has become a well-established in vitro model for studying the biology and pathogenic mechanisms of BVDV infection. With the help of this model, previous studies have initially revealed the differences between CP and NCP BVDV in viral replication kinetics and protein accumulation, among other aspects [20]; However, whether these two biotypes determine infection outcomes toward lytic clearance or persistent survival by differentially regulating host inflammatory responses and cell death programs remains to be systematically elucidated at the mechanistic level.
Pyroptosis is an important critical innate immune defense mechanism of the host against viral infection, in which inflammasomes play a central regulatory role in this process [21]. The NLRP3 inflammasome consists of the sensor molecule NLRP3, the adaptor ASC, and the downstream effector pro-Caspase-1. Its activation can promote the self-cleavage activation of Caspase-1, thereby mediating GSDMD cleavage, forming membrane pores and inducing pyroptosis, while also driving the maturation and secretion of IL-1β and IL-18 [22,23]. Current research indicates that various viruses can activate the NLRP3 pathway by disturbing ion homeostasis, inducing mitochondrial damage, or directly acting on inflammasome components, thereby triggering GSDMD-dependent pyroptosis and further amplifying inflammatory responses and tissue damage [21,24,25]. However, there is still a lack of systematic research on whether BVDV infection of MDBK cells can activate inflammasomes, induce pyroptosis, and the differential effects of different BVDV biotypes in the above processes. More importantly, whether the BVDV-encoded proteins directly participate in the regulation of inflammasomes and their potential molecular mechanisms has not yet been elucidated.
This study employs MDBK cells as a model to compare the effects of the cytopathogenic BVDV strain NADL and the non-cytopathogenic strain TC on inflammasome activation and pyroptosis, and to analyze the key mechanisms by which BVDV-encoded viral proteins mediate inflammasome activation. Aiming to elucidate the pathogenic mechanism of BVDV from the perspective of inflammatory immunity and programmed cell death, thereby providing theoretical support for the development of prevention and control strategies targeting the inflammasome pathway.
2. Materials and Methods
2.1. Cells Culture and Virus Infection
Madin-Darby bovine kidney (MDBK, Bio-68289) cells were obtained from the Cell Bank of Type Culture Collection of Chinese Academy of Sciences (TCCCAS) and grown in Dulbecco’s Modified Eagle Medium (DMEM, Gibco, USA) supplemented with 10% fetal bovine serum (Gibco, USA), at 37 °C in a humidified incubator containing 5% CO₂. The BVDV NADL and TC strains were isolated and characterized in our laboratory.
2.2. Antibodies and Reagents
Goat anti-mouse IgG H&L (FITC) (ab6785, Abcam, UK); anti-F-actin antibody (bs-1571R, Bioss, China); Coralite488-conjugated Goat Anti-Rabbit IgG (H+L) (SA00013-2), and Coralite594-conjugated Goat Anti-Mouse IgG (H+L) (SA00013-3) were from Proteintech (USA); Caspase-1 Rabbit pAb (A0964), GAPDH Rabbit pAb (AC027), NLRP3 Rabbit mAb (A24294), ASC Rabbit mAb (A24165), and GSDMD (Full Length+N terminal) Rabbit pAb (A24476) were from ABclonal (China); Poly I:C (42-875-0) was from Gibco (USA).
2.3. Lentiviral Packaging
Mix the target gene plasmid, packaging plasmid psPAX2, and envelope plasmid pMD2.G at a 3:2:1 ratio, and use Opti-MEM (Gibco, USA) to adjust the volume to a final volume of 250 µL, and incubate at room temperature for 10 minutes. At the same time, mix 34.5 μL of PEI (Servicebio, China) with Opti-MEM to a total volume of 250 μL, and mix by vortexing, and allow to stand at room temperature for 5 minutes. Then slowly add the PEI solution to the plasmid mixture, mix gently, and incubate at room temperature for 15 minutes to form the transfection complex. The complex was slowly added dropwise to the cell culture, gently mixed, and subsequently returned to the incubator for further cultivation. At 18 h after transfection, the culture medium was replaced with fresh complete medium. At 48 hours post-transfection, the culture supernatants were collected, centrifuged to remove cell debris, filtered through a 0.45 μm filter, aliquoted, and stored at -80℃ for future use.
2.4. Immunofluorescence
MDBK cells were cultured on coverslips in a 48-well plate at a density of 2 × 10⁵ cells/well, and then infected with BVDV NADL and TC strains at an MOI of 1, respectively. After 36 h of incubation, the culture medium was aspirated, cells were washed thoroughly, fixed with 4% paraformaldehyde (PFA) for 10 minutes, and then permeabilize with 0.5% Triton X-100 in PBS for 8 minutes; Subsequently, cells were blocked with 3% bovine serum albumin (BSA, Source Leaf, China), followed by incubation with primary antibodies targeting NLRP3, ASC, or Caspase-1 (1:100), overnight at 4 °C. After washing with PBS, secondary antibodies conjugated with Coralite488 and 594 (1:200) were added and incubated for 2 hours in the dark, then cell nuclei were stained with DAPI (4′,6-diamidino-2-phenylindole) and slides were mounted using FluoroGuard Antifade Mounting Medium (Invitrogen). Finally, images were observed and acquired using a laser confocal microscope (Leica STELLARIS 5, Germany).
2.5. RNA Extraction and Quantitative PCR (qPCR)
Total RNA was extracted from BVDV-infected MDBK cells at 36 h post-infection using an RNA extraction kit (TaKaRa, Japan) following the manufacturer’s instructions; Following cell lysis and centrifugation, the supernatant was collected, mixed with 70% ethanol, and then transferred to a purification column for sequential washing and centrifugation, and then DEPC water was added to the column, following incubation at room temperature for 5 min, the sample was centrifuged at 12,000 rpm for 2 min to elute RNA. The purified RNA was quantified and stored at −80 °C.
Genomic DNA was subsequently removed using the PrimeScript RT Reagent Kit with gDNA Eraser (TaKaRa, Japan), reverse transcription was performed following the manufacturer’s protocol.: The protocol involved gDNA removal at 42℃ for 5 min, cDNA was synthesized at 37℃ for 15 min, and the enzyme was inactived by incubating at 85℃ for 5 s. cDNA as a template, the real-time quantitative PCR mixture was prepared using TB Green™ Premix Ex Taq™ II (TaKaRa, Japan), The 2⁻ΔΔCt method was used to calculate the relative expression of mRNA. Primers used for real-time PCR are mentioned (Table 1).
2.6. TME
MDBK cells were seeded onto coverslips in 6-well plates at a density of 1 × 10⁵ cells per well, and infect them with BVDV NADL strain and TC strain at an MOI of 1, respectively. After 36 hours, gently scrape with a precooled cell scraper, centrifuge at 1,000 rpm, and fix the cells overnight at 4 °C using 2.5% glutaraldehyde (Yuan Ye, China). Afterwards, following three washes with 0.1 M PBS, the samples were post-fixed in 1% osmium tetroxide for 1 h, and dehydrated through a graded series of ethanol and acetone. Subsequently, they were gradually infiltrated with a mixture of acetone and epoxy resin, embedded, and polymerized to fully cure the resin. Ultrathin sections were subjected to double staining with uranyl acetate and lead citrate, followed by examination of cellular ultrastructure using transmission electron microscopy.
2.7. ELISA
Bovine IL-1β and IL-18 concentrations in culture supernatants were quantified using ELISA. MDBK cells were seeded into 6-well plates and cultured until they reached approximately 80% confluence, then infected with BVDV NADL and TC strains at an MOI of 1 to establish infection models. After 36 hours of infection, the cell supernatant was collected. Standard solutions were prepared and standard curves were drawn according to the kit instructions (Panteze Bio, China). During detection, the sample was diluted proportionally and added to the enzyme-labeled plate, and the plate was incubated with the standard product at 37 °C for 1 hour. Following plate washing, the enzyme-labeled secondary antibody was added and incubated again for 1 hour, followed by repeating the washing steps. Subsequently, the chromogenic substrate solution was added, and the reaction was terminated after 15 minutes in the dark. Finally, the samples were measured using the Hidex Sense microplate reader (Synergy HTX, USA) at 450 nm, and the concentrations of IL-1β and IL-18 in the samples were calculated according to the standard curve.
2.8. Immunoblotting
After 36 h of treatment, the culture medium was aspirated, and MDBK cells were harvested for total protein extraction. Protein concentration was quantified using a bicinchoninic acid (BCA) assay kit (Beyotime, China); Equal amounts of protein were mixed with 4 × loading buffer and denatured by heating at 100 °C for 10 min; Protein samples were separated on sodium dodecylsulfate-polyacrylamide gel electtophoresis (SDS-PAGE) (Seven, China) and transferred to a polyvinyl difluoride (PVDF) membranes, membranes were blocked with 5% skim milk in TBST for 2 h at room temperature, and incubated overnight at 4 °C with antibodies diluted as indicaed: GAPDH (1:2000), Caspase-1 (1:3000), NLRP3 (1:1500), and GSDMD (1:3500). Following three washes with TBST, the membranes were incubated with HRP-conjugated secondary antibodies (goat anti-mouse IgG (H+L) (1:10000) or goat anti-rabbit IgG (H+L) (1:20000)) at 37 °C for 2 hours and visualized using an ultra-sensitive ECL kit, and protein band signals were captured using an imaging system.
2.9. Co-Immunoprecipitation assay (Co-IP)
Upon reaching 80% confluency, MDBK cells were transfected with a plasmid expressing Flag-tagged BVDV C protein, with an empty vector as a negative control. After 36 hours of transfection, the cells were washed with pre-cooled PBS and then lysed on ice with IP lysis buffer containing protease inhibitors (Beyotime, China) on ice. The lysates were centrifuged at 12,000 g at 4 °C for 15 minutes to collect the supernatant, and the protein concentration was measured. An equal amount of protein was incubated with anti-Flag M2 affinity gel or magnetic beads (Beyotime, China) at 4 °C overnight. Following incubation, the beads/gel were washed, and 5 × SDS loading buffer was added to boil and elute the immunoprecipitated proteins. Immunoprecipitated products and input samples were separated by SDS-PAGE and transferred to a PVDF membrane. After blocking, primary antibodies against NLRP3, ASC, Caspase-1, and Flag were incubated at 4 °C overnight. The membrane was incubated with HRP-conjugated secondary antibodies, and visualized with ECL chemiluminescence reagents. The results were analyzed by image software.
2.10. Statistical Analysis
All data are presented as the mean ± SD. Statistical analysis was performed using one-way ANOVA, followed by Bonferroni post hoc tests for multiple comparisons. Statistical analysis and graphing were performed using GraphPad Prism 8.0.2 software (GraphPad Software Inc, San Diego, CA, USA). Symbols in figures indicate statistical significance levels: * denotes P < 0.05, ** denotes P < 0.01, *** denotes P < 0.001.
3. Results
3.1. BVDV Infection Induce NLRP3 Inflammasomes-Meidated Inflammatory Responses
To investigate whether BVDV infection of MDBK cells induces an inflammatory response, ELISA assays were performed to measure IL-1β and IL-18 secretion levels following infection with the cytopathic NADL strain and the non-cytopathogenic TC strain. Results demonstrated that both BVDV NADL and TC strains significantly induced proinflammatory cytokine secretion (Figure 1a). Given that the NLRP3 inflammasome comprises NLRP3, ASC, and Caspase-1, we further employed laser confocal microscopy to detect the expression and activation status of its key components, thereby assessing whether the NLRP3 inflammasome was activated. Results showed that both the BVDV NADL and TC strains exhibited colocalization of ASC and Caspase-1 with NLRP3 in the cytoplasm (Figure 1b). Collectively, these results indicate that BVDV infection activates the NLRP3 inflammasome and contributes to the inflammatory response.
3.2. BVDV NADL Induces NLRP3 Inflammasome Activation not TC Strain
To further confirm the occurrence of pyroptosis, the ultrastructure of MDBK cells was examined by transmission electron microscopy. The results showed that infection with the cytopathic NADL strain disrupted cell membrane integrity, resulting in membrane rupture and cleft formation, as well as chromatin condensation within the nuclei. These morphological features were consistent with pyroptosis, whereas no significant changes were observed in cells infected with the non-cytopathogenic TC strain (Figure 2a). Subsequently, cellular damage was assessed using a lactate dehydrogenase (LDH) release assay, which revealed significantly elevated LDH levels in NADL strain-infected cells (Figure 2b). Immunofluorescence analysis of the pyroptosis effector protein GSDMD expression showed red fluorescence in the cytoplasm of NADL strain-infected cells, whereas no significant red fluorescence was observed in TC strain-infected cells (Figure 2c1,c2).
qPCR analysis of mRNA expression levels of pyroptosis-related genes revealed significant upregulation of the NLRP3 inflammasome complex (NLRP3, ASC, and Caspase-1), the pyroptosis executor protein (GSDMD), and downstream inflammatory cytokines (IL-1β and IL-18) (Figure 2d). Given that qPCR only reflects gene transcription levels and cannot assess the processing and functional activation status of key pyroptosis proteins, Western blot analysis was further employed in this study to examine the expression and cleavage patterns of key proteins in the NLRP3 inflammasome pathway. Results demonstrated that NADL strain infection induced a time-dependent upregulation of NLRP3 and ASC protein levels in cells. Simultaneously, NADL strain infection significantly induced the functional cleavage of Caspase-1 and GSDMD, yielding Caspase-1 p20 and GSDMD-N fragments, respectively (Figure 2e1,e2). In contrast, no significant cleavage fragments of Caspase-1 or GSDMD were detected in cells infected with the TC strain (Figure 2e1,e2). These findings indicate that the BVDV NADL strain can effectively activate the NLRP3 inflammasome downstream effector pathway, thereby inducing Caspase-1 and GSDMD-mediated pyroptosis.
3.4. BVDV Core Protein C Induces Pyroptosis in MDBK Cells
Infection of MDBK cells with the BVDV NADL strain can further induce pyroptosis by activating the NLRP3 inflammasome. Previous studies generally suggest that virus-induced pyroptosis typically relies on viral-encoded proteins to mediate host inflammasome activation and programmed cell death processes. Therefore, this section of the study employed ELISA to detect IL-1β and IL-18 secretion in the supernatants following infection of MDBK cells with a recombinant lentivirus encoding components of the BVDV NADL strain Results indicate that IL-1β and IL-18 secretion levels were highest in cells expressing the C protein (Figure 3a). Immunofluorescence assay (IFA) analysis revealed significant expression levels of the inflammasome component proteins NLRP3, ASC, and GSDMD in the C protein group as well (Figure 3b1–b3).
3.5. BVDV Core Protein Binds with ASC to Induce Inflammasome Assemble and Pyroptosis
Given that the NLRP3 inflammasome is composed of three key components: NLRP3, ASC, and Caspase-1; its complete assembly and functional activation depend on precise interactions among these components. Therefore, systematically elucidating the interactions between the core protein C of the BVDV NADL strain and the various components of the NLRP3 inflammasome is crucial for gaining a deeper understanding of the molecular mechanisms mediating pyroptosis. Co-immunoprecipitation (Co-IP) was used to analyze the relationship between viral protein C and inflammasome complex components, which revealed that protein C interacts exclusively with ASC in MDBK cells, but not with NLRP3 or Caspase-1 (Figure 4a). This indicates that BVDV core protein C interacts with ASC of the NLRP3 inflammasome. To further validate the interaction between BVDV core protein C and ASC, which subsequently activates the NLRP3 inflammasome, this study targeted ASC with synthetic siRNA. qPCR results indicated that si-525 exhibited the most potent inhibitory effect (Figure 4b). Using an ELISA kit, we assessed proinflammatory cytokine secretion in cell supernatants after BVDV core protein C lentiviral infection in ASC-interfered cells. Results demonstrated that ASC gene interference significantly reduced IL-1β and IL-18 secretion (Figure 4c). Concurrently, immunofluorescence analysis revealed that ASC interference effectively suppressed the expression of pyroptosis effector protein GSDMD (Figure 4d).
To further investigate the effects of BVDV core protein C on pyroptosis in MDBK cells following ASC knockdown, qPCR was employed to detect mRNA expression levels of pyroptosis-related genes after ASC knockdown and subsequent infection with a lentivirus expressing BVDV core protein C. Results demonstrated that interfering with ASC protein expression significantly reduced mRNA expression levels of Caspase-1, ASC, GSDMD, IL-1β, and IL-18 mRNA expression, while NLRP3 mRNA levels remained unaffected (Figure 4e). Western blot analysis detected the protein expression levels of pyroptosis-related genes after reinfection with the BVDV core protein C recombinant lentivirus following ASC knockdown, yielding results consistent with qPCR findings (Figure 4f). These findings indicate that interfering with ASC protein expression effectively suppressed BVDV core protein C-induced secretion of IL-1β and IL-18 in MDBK cells, as well as the cleavage and activation of pro-Caspase-1 and GSDMD, thereby inhibiting pyroptosis.
4. Discussion
Bovine viral diarrhea virus (BVDV) is an economically important pathogen that substantially affects the global cattle industry, can significantly reduce animal productivity, resulting in an economic loss of approximately £46.50 per cow per year [26,27]. Given its widespread prevalence and the severe impact it causes, It has been classified by the World Organisation for Animal Health (WOAH) as a Category B infectious animal disease [28]. Currently, BVDV continues to be prevalent in 88 countries and regions worldwide, underscoring its efficient transmission capability and the unignorable epidemiological risk [29]. Therefore, systematically analyzing the molecular characteristics and regulatory networks of BVDV-induced pyroptosis in MDBK cells is of crucial scientific value for elucidating the pathogenic mechanisms of the virus and revealing the regulatory mechanisms of the host inflammatory response.
The concept of cell pyroptosis was first proposed by the American scholar Brad T. Cookson in 2001 based on observations of macrophages infected by pathogens, revealing the molecular basis by which innate immune cells achieve an active defense through programmed inflammatory death [30], Recent studies have further expanded this understanding, confirming that various non-immune cells, including epithelial cells [31,32], endothelial cells [33,34] and organ parenchymal cells [35,36], also possess the complete molecular mechanism for pyroptosis execution, It has established the key regulatory role of this mode of death in the occurrence and progression of infectious diseases. However, in the process of bovine viral diarrhea virus (BVDV) infecting bovine kidney epithelial cells (MDBK), whether pyroptosis is activated and its regulatory mechanism still lacks systematic research. To our knowledge, this study is the first to demonstrate that BVDV can induce pyroptosis in MDBK cells; ELISA and IFA results showed that both the NADL strain and the TC strain can upregulate the expression of inflammatory factors such as IL-1β and IL-18, and promote the co-localization of NLRP3, ASC, and Caspase-1. This finding is in line with the results reported by Pandey et al. in their study on pathogen-induced inflammasome assembly. It indicates that both biotypes of BVDV have the ability to activate the upstream signals of the NLRP3 inflammasome [37]. However, further ultrastructural and functional analyses revealed the fundamental differences between the two strains in inducing pyroptosis: Only the NADL strain can induce the formation of typical pyroptotic pores in the cell membrane and significantly exacerbate cell damage, whereas the TC strain, although able to activate some inflammatory signals, fails to cause obvious membrane rupture. This phenomenon is highly consistent with the research findings of Li and Weir [38,39], These results indicate that inflammasome signaling activation and transcriptional upregulation of inflammatory mediators alone are insufficient to trigger pyroptosis. The actual execution of pyroptosis must rely on the effective cleavage of the GSDMD protein and the transmembrane pore formation it mediates. Only when the integrity of the cell membrane is compromised can the pyroptosis program be fully activated [40,41]. It is worth noting that although both strains can upregulate the expression of NLRP3 inflammasome-related genes at the transcriptional level, only the CP-type NADL strain can effectively induce GSDMD cleavage and initiate downstream pyroptosis programs. This difference indicates that CP-type BVDV tends to fully activate the inflammasome effector axis, triggering an acute inflammatory response through the induction of lytic cell death, whereas NCP-type BVDV is blocked at the execution phase of pyroptosis. This phenomenon may help the virus avoid rapid lysis of host cells, thereby creating favorable conditions for the establishment of persistent infection (PI) [42,43].
Previous studies have shown that various viral structural proteins, in addition to completing virus assembly and replication, can also act as immune regulatory factors directly involved in the regulation of host inflammasome signaling pathways [44], For example, the influenza virus M2 protein activates the NLRP3 inflammasome by disrupting intracellular ion homeostasis [45], the hepatitis B virus core protein can trigger downstream inflammatory responses [46], The Zika virus non-structural protein NS5 has been shown to target inflammasome components to amplify inflammatory signals [47]. These findings collectively reveal that the precise regulation of inflammasome pathways by virus-encoded proteins is often the key factor determining whether an infection progresses toward acute inflammation or immune evasion. However, as an important pathogen of the cattle family, the direct role of BVDV-encoded proteins in inflammasome activation and pyroptosis regulation has previously lacked systematic evidence. This study found that the core protein C of the BVDV NADL strain can significantly promote the secretion of IL-1β and IL-18 and upregulate the expression of NLRP3, ASC, and GSDMD, confirming its ability to activate the inflammasome and drive pyroptosis, thereby providing new evidence for understanding the molecular mechanism of acute inflammation induced by CP BVDV. Further mechanistic analysis showed that the core protein C does not induce an inflammatory response through nonspecific cellular damage, but rather promotes the effective assembly of the NLRP3 inflammasome through specific interaction with ASC, thereby activating downstream pyroptotic processes in an ASC-dependent manner. Specifically, after interfering with ASC expression, both the secretion of inflammatory factors induced by the core protein C and the upregulation of molecules related to pyroptosis were significantly weakened, confirming the pivotal role of ASC in this process. It is worth noting that the regulation mode of the core protein C targeting ASC is not an isolated case. Zhang et al. [48] reported that the Pseudorabies virus UL4 protein also activates NLRP3 and AIM2 inflammasomes by targeting ASC, suggesting that ASC may be an inflammasome regulatory node commonly targeted by multiple viral proteins. As a key adaptor protein, ASC plays a central role in inflammasome assembly and signal amplification, and has both PYD and CARD domains, allowing it to bridge upstream sensors and downstream effector protein Caspase-1 [49,50]. Viral proteins can effectively modulate inflammatory signaling pathways by targeting ASC, which can either amplify inflammation to promote viral spread or suppress inflammation to establish persistent infection.
This study reveals the key molecular differences in the pathogenic mechanisms between cytopathogenic and non-cytopathogenic BVDV from the perspective of inflammasome-mediated pyroptosis, However, certain limitations persist. Firstly, this study primarily utilized an in vitro MDBK cell model, which may not fully recapitulate the dynamic regulation of inflammasome signals in the complex immune microenvironment in vivo; Secondly, beyond the NLRP3 inflammasome, the potential roles of other types of inflammasomes and their cross-regulatory effects in BVDV infection warrant further investigation. Additionally, the specific structural basis of the interaction between BVDV core protein C and ASC and its precise regulatory mechanism, likewise require verification through more in-depth structural biology studies and in vivo functional experiments. Moving forward research could leverage animal infection models and gene editing technology to systematically evaluate the dual roles of the inflammasome-pyroptosis pathway in BVDV pathogenesis and host antiviral immunity, as well as explore intervention strategies targeting this pathway. In conclusion, this study not only deepens our understanding of the regulatory mechanisms of inflammatory responses during BVDV infection, but also provides important theoretical insights for the development of new antiviral control measures.
5. Conclusions
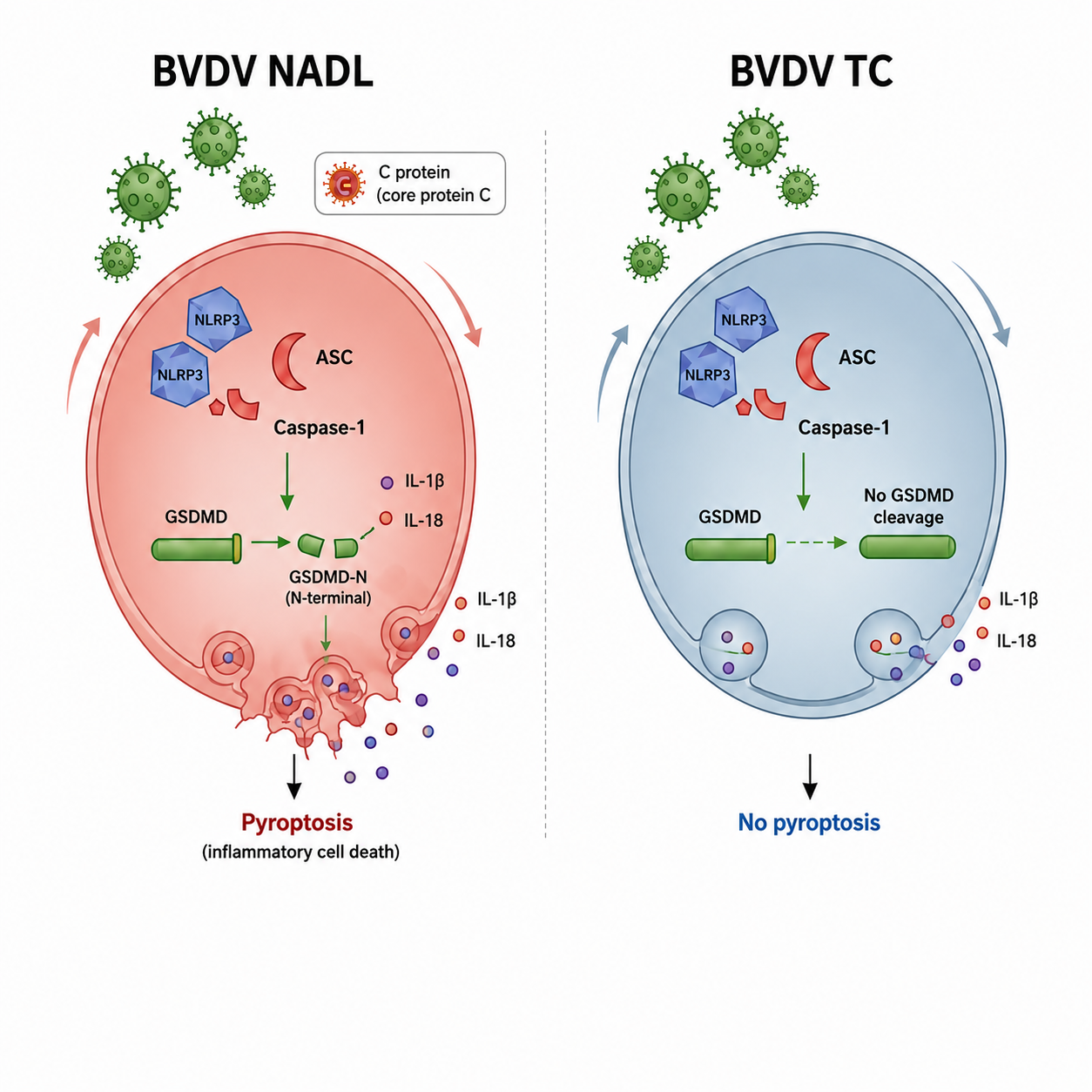
This study investigates mechanisms of pyroptosis mediated by inflammasomes and systematically analyzes the pathogenic differences of different BVDV biotypes. Our findings demonstrate that the cytopathic variant NADL strain specifically interacts with the adaptor protein ASC via its core protein C, leading to NLRP3 inflammasome assembly and Caspase-1 activation, This subsequently mediates GSDMD cleavage, membrane pore formation, and ultimately, pyroptosis. Non-cytopathic TC strains can activate upstream signals of the inflammasome, but are insufficient to drive the complete pyroptosis pathway. These findings collectively suggest that the BVDV core protein C-ASC axis serves as a key molecular switch regulating inflammasome activation and pyroptosis, thereby dictating the distinct outcomes of infection with different biotypes of the virus. This study elucidates the key molecular of BVDV-induced host inflammatory damage at the molecular level and provides a theoretical basis for intervention strategies targeting the inflammasome pathway (Figure 5).
Author Contributions
Conceptualization, H.Z.; Data curation, J.Y. (Jiaming Yang); Formal analysis, N.H.; Funding acquisition, K.S. and R.D.; Investigation, N.H. and Y.L.; Methodology, Q.W. and Z.J.; Project administration, J.L. and J.Y. (Jiying Yin); Resources, N.D. and F.Z.; Supervision, K.S. and R.D.; Writing—original draft, N.H. and Y.K.; Writing—review & editing, N.H. and K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Jilin Provincial Department of science and technology (20230101255JC) and the National Natural Science Foundation of China (32573352).
References
- Rana, E.A. Molecular Epidemiology and Control Strategies for BVDV: A Global Systematic Review From 2000 to 2025. Vet. Med. Int. 2025, 6732453. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K. Bovine viral diarrhea virus eradication in Germany: A never-ending success story or just the last 46 PI animals? Vet. Microbiol. 2025. 309, 110697. [Google Scholar] [CrossRef]
- Hu, X. Bovine Viral Diarrhea Virus and Vaccine Protection Strategies. Vet. Sci. 2026, 13(2). [Google Scholar] [CrossRef] [PubMed]
- Li, Y. Vitamin C combined with anti-PD-1 antibody alleviates peripheral lymphopenia and enhances CD4(+) T-cell antiviral immunity during BVDV infection. Vet. Microbiol. 2026, 312, 110825. [Google Scholar] [CrossRef]
- Tesfaye Melkamsew, A.; Tessema, T. Sisay; Paeshuyse, J. Host Immune Response to Bovine Viral Diarrhea Virus (BVDV): Insights and Strategies for Effective Vaccine Design. Vaccines 2025, 13(5). [Google Scholar] [CrossRef] [PubMed]
- Hou, Z. A Systematic Study of Bovine Viral Diarrhoea Virus Co-Infection with Other Pathogens. Viruses 2025, 17(5). [Google Scholar] [CrossRef]
- Yeşilbağ, K.; Alpay, G.; Becher, P. Variability and Global Distribution of Subgenotypes of Bovine Viral Diarrhea Virus. Viruses 2017, 9(6). [Google Scholar] [CrossRef]
- Tsu, B.V. Running With Scissors: Evolutionary Conflicts Between Viral Proteases and the Host Immune System. Front Immunol. 2021, 12, 769543. [Google Scholar] [CrossRef]
- Pang, F.; Long, Q.; Wei, M. Immune evasion strategies of bovine viral diarrhea virus. Front Cell Infect. Microbiol. 2023, 13, 1282526. [Google Scholar] [CrossRef]
- Zhang, Y. Identification of three novel B cell epitopes targeting the bovine viral diarrhea virus NS3 protein for use in diagnostics and vaccine development. Int. J. Biol. Macromol. 2025. 308, Pt 4, 142767. [Google Scholar] [CrossRef]
- Dong, Q. Screening and Characterization of TAT-Fused Nanobodies Targeting Bovine Viral Diarrhea Virus NS3/NS5A for Antiviral Application. Biomolecules 2025, 15(11). [Google Scholar] [CrossRef] [PubMed]
- Zhang, K. Integrative Transcriptomics and Proteomics Analysis Reveals Immune Response Process in Bovine Viral Diarrhea Virus-1-Infected Peripheral Blood Mononuclear Cells. Vet. Sci. 2023, 10(10). [Google Scholar] [CrossRef] [PubMed]
- Peterhans, E. Cytopathic bovine viral diarrhea viruses (BVDV): emerging pestiviruses doomed to extinction. Vet. Res. 2010, 41(6), 44. [Google Scholar] [CrossRef]
- La Polla, R. Identification of differentially expressed gene pathways between cytopathogenic and non-cytopathogenic BVDV-1 strains by analysis of the transcriptome of infected primary bovine cells. Virology 2022, 567, 34–46. [Google Scholar] [CrossRef]
- Georges, H.M. Epigenomic and Proteomic Changes in Fetal Spleens Persistently Infected with Bovine Viral Diarrhea Virus: Repercussions for the Developing Immune System, Bone, Brain, and Heart. Viruses 2022, 14(3). [Google Scholar] [CrossRef]
- Holthausen, D.J. Bovine viral diarrhea virus 2 strains generate deletion viral genomes primarily in the NS2 region of the viral genome. Front Vet. Sci. 2025. 12, 1686098. [Google Scholar] [CrossRef]
- Zhong, J. Molecular Characteristics and Pathogenicity Analysis of Bovine Viral Diarrhea Virus Strain Isolated from Persistently Infected Cattle. Animals 2026, 16(1). [Google Scholar] [CrossRef]
- Li, Z. Non-cytopathic bovine viral diarrhea virus (BVDV) inhibits innate immune responses via induction of mitophagy. Vet. Res. 2024, 55(1), 27. [Google Scholar] [CrossRef]
- Cui, Y. The activation of liver X receptors in Madin-Darby bovine kidney cells and mice restricts infection by bovine viral diarrhea virus. Vet. Microbiol. 2024, 288, 109948. [Google Scholar] [CrossRef]
- Mirosław, P. Transcriptomic Analysis of MDBK Cells Infected with Cytopathic and Non-Cytopathic Strains of Bovine Viral Diarrhea Virus (BVDV). Viruses 2022, 14(6). [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q. The NLRP3 inflammasome in viral infection (Review). Mol. Med. Rep. 2023, 28(3). [Google Scholar] [CrossRef]
- Vande Walle, L.; Lamkanfi, M. Drugging the NLRP3 inflammasome: from signalling mechanisms to therapeutic targets. Nat. Rev. Drug Discov. 2024, 23(1), 43–66. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, L. Mechanistic and therapeutic insights into the function of NLRP3 inflammasome in sterile arthritis. Front Immunol. 2023, 14, 1273174. [Google Scholar] [CrossRef] [PubMed]
- Vora, S.M.; Lieberman, J.; Wu, H. Inflammasome activation at the crux of severe COVID-19. Nat. Rev. Immunol. 2021, 21(11), 694–703. [Google Scholar] [CrossRef]
- Shi, W. Inflammasome activation by viral infection: mechanisms of activation and regulation. Front Microbiol. 2023, 14, 1247377. [Google Scholar] [CrossRef]
- Su, N. Prevalence of bovine viral diarrhea virus in cattle between 2010 and 2021: A global systematic review and meta-analysis. Front Vet. Sci. 2022, 9, 1086180. [Google Scholar] [CrossRef]
- Zirra-Shallangwa, B. The Epidemiology of Bovine Viral Diarrhea Virus in Low- and Middle-Income Countries: A Systematic Review and Meta-Analysis. Front Vet. Sci. 2022, 9, 947515. [Google Scholar] [CrossRef]
- Rana, E.A. A Critical Review of Bovine Viral Diarrhea Virus: Spotlights on Host Plasticity and Potential Spillover Events. Viruses 2025, 17(9). [Google Scholar] [CrossRef]
- Yin, J. Tunneling nanotubes provide a new route for bovine viral diarrhea virus spreading. Front Vet. Sci. 2025, 12, 1667394. [Google Scholar] [CrossRef] [PubMed]
- Cookson, B.T.; Brennan, M.A. Pro-inflammatory programmed cell death. Trends Microbiol. 2001, 9(3), 113–4. [Google Scholar] [CrossRef]
- Ma, X. LPS Mediates Bovine Endometrial Epithelial Cell Pyroptosis Directly Through Both NLRP3 Classical and Non-Classical Inflammasome Pathways. Front Immunol. 2021, 12, 676088. [Google Scholar] [CrossRef]
- Fan, H.; Sun, M.; Zhu, J.H. S-nitrosoglutathione inhibits pyroptosis of kidney tubular epithelial cells in sepsis via the SIRT3/SOD2/mtROS signaling pathway. Ren. Fail 2025, 47(1), 2472987. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.T. Caspase-11-mediated endothelial pyroptosis underlies endotoxemia-induced lung injury. J. Clin. Invest 2017, 127(11), 4124–4135. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y. Nano-selenium alleviates the pyroptosis of cardiovascular endothelial cells in chicken induced by decabromodiphenyl ether through ERS-TXNIP-NLRP3 pathway. Sci. Total Env. 2024. 915, 170129. [Google Scholar] [CrossRef]
- Ma, Y. Effects of Adipose-Derived Mesenchymal Stem Cell-Secretome on Pyroptosis of Laparoscopic Hepatic Ischemia Reperfusion Injury in a Porcine Model. Cells 2025, 14(10). [Google Scholar] [CrossRef]
- Li, C. Dynamic changes in pyroptosis following spinal cord injury and the identification of crucial molecular signatures through machine learning and single-cell sequencing. J. Pharm. BioMed Anal. 2024, 251, 116449. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.P.; Zhou, Y. Influenza A Virus Infection Activates NLRP3 Inflammasome through Trans-Golgi Network Dispersion. Viruses 2022, 14(1). [Google Scholar] [CrossRef]
- Li, Y.; Jiang, Q. Uncoupled pyroptosis and IL-1β secretion downstream of inflammasome signaling. Front Immunol. 2023, 14, 1128358. [Google Scholar] [CrossRef]
- Weir, A.; Vince, J.E. No longer married to inflammasome signaling: the diverse interacting pathways leading to pyroptotic cell death. Biochem J. 2022, 479(10), 1083–1102. [Google Scholar] [CrossRef]
- Zhang, N.; Xu, D. Controlling pyroptosis through post-translational modifications of gasdermin D. Dev. Cell 2025, 60(7), 994–1007. [Google Scholar] [CrossRef] [PubMed]
- Schiffelers, L.D.J. Antagonistic nanobodies implicate mechanism of GSDMD pore formation and potential therapeutic application. Nat. Commun. 2024, 15(1), 8266. [Google Scholar] [CrossRef] [PubMed]
- Charleston, B. Establishment of persistent infection with non-cytopathic bovine viral diarrhoea virus in cattle is associated with a failure to induce type I interferon. J. Gen. Virol. 2001, 82 Pt 8, 1893–1897. [Google Scholar] [CrossRef]
- Yamane, D. The double-stranded RNA-induced apoptosis pathway is involved in the cytopathogenicity of cytopathogenic Bovine viral diarrhea virus. J. Gen. Virol. 2006, 87 Pt 10, 2961–2970. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.; Borg, N.A. The multifaceted roles of NLRP3-modulating proteins in virus infection. Front Immunol. 2022, 13, 987453. [Google Scholar] [CrossRef]
- Ichinohe, T.; Pang, I.K.; Iwasaki, A. Influenza virus activates inflammasomes via its intracellular M2 ion channel. Nat. Immunol. 2010, 11(5), 404–10. [Google Scholar] [CrossRef]
- Ding, X. Hepatitis B core antigen can regulate NLRP3 inflammasome pathway in HepG2 cells. J. Med. Virol. 2019, 91(8), 1528–1536. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. Zika virus infection induces host inflammatory responses by facilitating NLRP3 inflammasome assembly and interleukin-1β secretion. Nat. Commun. 2018, 9(1), 106. [Google Scholar] [CrossRef]
- Zhang, X. Pseudorabies Virus UL4 protein promotes the ASC-dependent inflammasome activation and pyroptosis to exacerbate inflammation. PLoS Pathog. 2024, 20(9), e1012546. [Google Scholar] [CrossRef]
- Xue, D. Atomic mechanisms of full-length ASC-mediated inflammasome assembly. Nat. Commun. 2025, 16(1), 10564. [Google Scholar] [CrossRef]
- Hoss, F.; Rodriguez-Alcazar, J.F.; Latz, E. Assembly and regulation of ASC specks. Cell Mol. Life Sci. 2017, 74(7), 1211–1229. [Google Scholar] [CrossRef]
Figure 1.
BVDV infection induce NLRP3 inflammasomes-meidated inflammatory responses. (a) Cells were infected with 1 MOI of the cytopathic BVDV strain NADL and the non-cytopathogenic TC strain. Pro-inflammatory factor secretion levels were measured at 12 h, 24 h, 36 h, and 48 h post-infection. (b) Co-localization of ASC and Caspase-1 with NLRP3 after 36 hours of IFA detection. Scale bar: 50 μm. Data are expressed as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 1.
BVDV infection induce NLRP3 inflammasomes-meidated inflammatory responses. (a) Cells were infected with 1 MOI of the cytopathic BVDV strain NADL and the non-cytopathogenic TC strain. Pro-inflammatory factor secretion levels were measured at 12 h, 24 h, 36 h, and 48 h post-infection. (b) Co-localization of ASC and Caspase-1 with NLRP3 after 36 hours of IFA detection. Scale bar: 50 μm. Data are expressed as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001.

Figure 2.
BVDV NADL induces NLRP3 inflammasome activation not TC strain; (a) Cells were infected at an MOI of 1 with the cytopathic BVDV strain NADL and the non-cytopathogenic TC strain. Transmission electron microscopy was performed 36 hours post-infection. A. Mock group; B. BVDV NADL group; C. BVDV TC group; D. Poly I:C group M: Mitochondria; N: Nucleus; Blue arrow: Site of membrane rupture; Scale bar: 50 μm; (b) Cell viability was assessed by LDH assay after 36 h; (c) Fluorescent expression of pyroptosis effector GSDMD detected by IFA. Scale bar: 50 μm. (d) mRNA expression levels of pyroptosis-related genes were detected using qPCR.; (e) Expression levels of pyroptosis-related proteins were detected using Western blot. Data are expressed as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 2.
BVDV NADL induces NLRP3 inflammasome activation not TC strain; (a) Cells were infected at an MOI of 1 with the cytopathic BVDV strain NADL and the non-cytopathogenic TC strain. Transmission electron microscopy was performed 36 hours post-infection. A. Mock group; B. BVDV NADL group; C. BVDV TC group; D. Poly I:C group M: Mitochondria; N: Nucleus; Blue arrow: Site of membrane rupture; Scale bar: 50 μm; (b) Cell viability was assessed by LDH assay after 36 h; (c) Fluorescent expression of pyroptosis effector GSDMD detected by IFA. Scale bar: 50 μm. (d) mRNA expression levels of pyroptosis-related genes were detected using qPCR.; (e) Expression levels of pyroptosis-related proteins were detected using Western blot. Data are expressed as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001.

Figure 3.
BVDV core protein C induces pyroptosis in MDBK cells; (a) Infection of MDBK cells with a lentivirus encoding the NADL strain protein at an MOI of 5 for 36 h, followed by detection of proinflammatory factor secretion; (b) Fluorescent expression of NLRP3, ASC, and GSDMD detected by IFA 36 h after lentiviral infection. Scale bar: 50 μm. Data are presented as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 3.
BVDV core protein C induces pyroptosis in MDBK cells; (a) Infection of MDBK cells with a lentivirus encoding the NADL strain protein at an MOI of 5 for 36 h, followed by detection of proinflammatory factor secretion; (b) Fluorescent expression of NLRP3, ASC, and GSDMD detected by IFA 36 h after lentiviral infection. Scale bar: 50 μm. Data are presented as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001.

Figure 4.
BVDV Core Protein binds with ASC to induce Inflammasome assemble and Pyroptosis; (a) Flag-tagged C protein was overexpressed in MDBK cells, where NLRP3, ASC, and Caspase-1 were labeled with specific antibodies. Co-immunoprecipitation (Co-IP) analysis was performed to investigate the interaction between viral protein C and components of the inflammasome complex. (b) Screening the interference efficiency of the four synthesized siRNAs (si-232, si-350, si-525, and si-570) by qPCR; (c) ELISA assay to detect proinflammatory cytokine secretion in the cell supernatants following ASC knockdown; (d) IFA assay to detect GSDMD protein expression in cells after ASC knockdown, Scale bar: 50 μm; (e) Detection of mRNA expression levels of pyroptosis-related genes following ASC knockdown using qPCR; (f) Detection of protein expression levels of pyroptosis-related genes following ASC knockdown using Western blot analysis. Data are presented as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 4.
BVDV Core Protein binds with ASC to induce Inflammasome assemble and Pyroptosis; (a) Flag-tagged C protein was overexpressed in MDBK cells, where NLRP3, ASC, and Caspase-1 were labeled with specific antibodies. Co-immunoprecipitation (Co-IP) analysis was performed to investigate the interaction between viral protein C and components of the inflammasome complex. (b) Screening the interference efficiency of the four synthesized siRNAs (si-232, si-350, si-525, and si-570) by qPCR; (c) ELISA assay to detect proinflammatory cytokine secretion in the cell supernatants following ASC knockdown; (d) IFA assay to detect GSDMD protein expression in cells after ASC knockdown, Scale bar: 50 μm; (e) Detection of mRNA expression levels of pyroptosis-related genes following ASC knockdown using qPCR; (f) Detection of protein expression levels of pyroptosis-related genes following ASC knockdown using Western blot analysis. Data are presented as mean ± SD. * p < 0.05; ** p < 0.01; *** p < 0.001.

Figure 5.
Simplified model of BVDV-induced pyroptosis via ASC-dependent NLRP3 inflammasome activation.
Figure 5.
Simplified model of BVDV-induced pyroptosis via ASC-dependent NLRP3 inflammasome activation.


Table 1.
Primers sequence used in real-time PCR.
| gene | Primer sequence (5′→3′) |
|---|---|
| NLRP3 | GATATFFCAGAGAATACGGAGAAGATG |
| AAFCACAGCACTACACAGAGAAG | |
| Caspase-1 | ATGGTCCTTCAACTTGGTGAATGTG |
| CTGACCACTTCGATAAGAACTGTATC | |
| ASC | GCTCTACTGTATGCACGAGA |
| TCTGCTFAAGACACTCGCT | |
| GSDMD | AGTGGTGACACCCTTGCCTT |
| GGGTCCTTGAGTCGTGTGAA | |
| IL-1β | AGATGAAGAGCGTGAGGAGC |
| CGAGCATCACTATGTGGGCC | |
| IL-18 | ACCGAGACTTCAGAGAGAATTGGAGAC |
| CCTTACCTGGCTATGGCTGTCTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



