Submitted:
27 May 2026
Posted:
27 May 2026
You are already at the latest version
Abstract
The Drosophila midgut serves as a powerful model system for studying the mechanisms under-lying stem cell hyperproliferation during aging and tissue regeneration. Two distinct pathways, the nutrient-sensing Insulin signaling axis and the growth-regulating Hippo pathway, are known to mediate intestinal stem cell (ISC) hyperproliferation in response to stress; however, the coor-dination between these two pathways remains unclear. Here, we show that Homeodo-main-interacting protein kinase (Hipk), recently identified as a nutrient transducer in progenitors, links Insulin signaling to Yorkie (Yki) activation in progenitor cells. Progeni-tor-specific hipk knockdown or knockout abolished gut hyperplasia induced by aging, dextran sulfate sodium (DSS)-mediated injury, and Yki overexpression. Mechanistically, Hipk promotes Yki protein stability and nuclear accumulation in progenitors. This regulation is specific to wild-type Yki, as the constitutively active YkiS168A mutant, which escapes Warts-mediated inhi-bition, is insensitive to Hipk depletion. Together, these findings identify Hipk as an integrator that couples Insulin signaling to the Hippo pathway to drive stress-induced ISC hyperprolifera-tion and intestinal regeneration.
Keywords:
hipk
; yki
; hippo
; insulin
; regeneration
; intestinal stem cells
; proliferation
; Drosophila
1. Introduction
The Drosophila midgut serves as an invaluable model for studying the plasticity of adult stem cells during homeostasis, aging, and regeneration [1,2,3,4,5]. Intestinal stem cells (ISCs) are located basally in the gut epithelium, where they maintain extensive contact with the basement membrane [6,7]. This membrane is apposed to visceral muscle cells, which form a specialized niche that supports ISC proliferation, stemness, and differentiation [8,9,10,11]. Under homeostatic conditions, ISCs undergo asymmetric cell division to produce a self-renewed ISC and a transient, post-mitotic progenitor cell, either an enteroblast (EB) or an enteroendocrine (EE) precursor (pre-EE) [12,13,14]. EBs further differentiate into enterocytes (ECs), the absorptive cells that constitute the epithelial monolayer, while pre-EEs differentiate into hormone-producing EEs [13]. In this binary lineage decision, most ISCs adopt an EB fate to supply the major EC population, while a subset adopts a pre-EE fate [12,13,15]. ISC proliferation is tightly regulated to maintain gut homeostasis.
However, homeostatic ISC proliferation is disrupted by aging and injury, such as exposure to dextran sulfate sodium (DSS), which induces hyper-proliferation and leads to a rapid expansion of ISCs and EBs, collectively termed progenitors [4,6,16,17,18]. Intriguingly, aging- and DSS-induced intestinal hyperplasia are attenuated by progenitor-specific knockdown of the insulin signaling pathway [16,19]. Although insulin signaling is a well-known nutrient-sensing pathway, how it integrates with aging- and DSS-induced progenitor expansion remains poorly understood.
The Hippo (Mst1 and Mst2 in mammals) pathway is an evolutionarily conserved regulator of cell growth and organ size that functions by restricting the oncoprotein Yorkie (Yki) (YAP and TAZ in mammals), a transcriptional co-activator [20,21,22,23]. The Hippo pathway is a critical regulator of intestinal homeostasis and regeneration [24,25,26,27,28,29,30]. The Ste20-like kinases Misshapen (Msn) and Hippo (Hpo) regulate the Warts-Yorkie (Wts-Yki) axis in progenitors and ECs, respectively [24,28,31,32]. Msn and Yki are enriched in progenitors, where Yki activity is tightly regulated under normal homeostatic conditions [27,28,29,32]. Gut damage by DSS relieves the suppression of Yki by the upstream kinase cascade [27,28]. Once dephosphorylated, active Yki translocates to the nucleus to drive the expression of genes that promote cell division, underlying the ISC hyperproliferation observed during regeneration. Correspondingly, DSS-stimulated ISC hyperproliferation is significantly reduced by progenitor-specific yki knockdown [27,28]. Conversely, progenitor-specific activation of Yki either through loss of upstream signaling or direct Yki overexpression is sufficient to phenocopy the ISC hyperproliferation induced by chemical injury [27,28].
Although insulin and Hippo signaling are both implicated in gut regeneration, their integration remains poorly understood. Recent findings characterize the kinase Hipk as a transducer of nutrient and insulin signaling that regulates homeostatic progenitor proliferation [33]. Here, we show that Hipk is essential for age-, DSS-, and Yki-induced proliferation, establishing it as a key integrator of insulin and Hippo signaling during intestinal regeneration.
2. Materials and Methods
2.1. Drosophila Stocks and Husbandry
Animals were maintained on a standard cornmeal diet (68 g dry yeast, 90 g sugar, 43 g cornmeal, 9 g agar, 4.5 mL propionic acid, 1 g methyl-4-hydroxybenaoate per 1-liter water) at 25 ℃ and 40% relative humidity under 12-hour light/dark cycle conditions. Midguts of virgin females were used for experiments unless otherwise stated. W1118 was used as control for experiments. Temporal control of transgene expression using tub-Gal80ts in adult flies was achieved by raising flies at 20 ℃ throughout development until three days after eclosion followed by shifting to 29 ℃ to induce Gal4-mediated transgene expression. The duration of UAS transgene induction is stated in each figure. Flies were transferred to fresh medium every second day. The following lines were generous gifts from the colleagues in the fly community: esgts refers to esg-Gal4, UAS-GFP/Cyo; tub-Gal80ts (a gift from Perrimon). esgTS refers to esg-Gal4; tub-Gal80ts (this study). UAS-ykiS168A:GFP, UAS-yki:GFP (a gift from Irvine) and UAS-HA-hipk (a gift from Verheyen). esgts/CyO;UAS.cas9.P2/TM6B and HD_CFD00782 (UAS-hipk sgRNA) (a gift from Boutros). UAS-Hipk RNAi (III) (BDSC #35363). UAS-Hipk RNAi (II) (BDSC #60084) were obtained from the Bloomington Drosophila Stock Center (BDSC). UAS-Wts RNAi (III) (VDRC #9928) were acquired from Vienna Drosophila Resource Center (VDRC).
2.2. Gut Regeneration Assay
Female flies (1 day old) were raised at 20 ℃ for three days and then shifted to 29 ℃ for seven days. Then, the flies were fed for another two days at 29 ℃ with sucrose (5%), DSS (5%), or Pe, after which midgut dissection and immunohistochemical analysis were performed. In detail, flies were put in a vial containing a disc of filter paper (Fisher catalog number 11111) wet with 5% sucrose solution or 5% dextran sulfate sodium (DSS, MB Biomedicals) dissolved in 5% sucrose, resting on fly food. For Pe infection, a bacterial pellet from overnight culture in LB broth at 30 ℃ was resuspended in 5% sucrose solution to OD600 = 200. One hundred l of the resuspension was smeared onto a filter paper disc. To ensure efficient intake of DSS or Pe, flies were starved for six hours before transfer to vials containing DSS or Pe. Flies were transferred to fresh vials with newly prepared DSS or Pe every day.
2.3. Immunohistochemistry
Adult guts were dissected out into 1x phosphate-buffered saline (PBS) and fixed in 4% paraformaldehyde for 30 min at room temperature. Samples were then washed 2x15 min in 1x PBS with 0.3% Triton X-100 (PBST) and incubated in blocking solution (PBST with 5% normal goat serum) for 30 min at room temperature. Samples were incubated with primary antibodies diluted in blocking solution overnight at 4 ℃, washed 2x15 min at room temperature in PBST, incubated with secondary antibodies diluted in blocking solution at room temperature for 2 h, and washed again 2x15 min with PBST. Finally, guts were stained with DAPI in blocking solution for 30 min at room temperature and mounted in Fluoromount-G® (SouthernBiotech) on a glass slide. Primary antibodies were used with the following dilutions: rabbit anti-phospho-Histone H3 Ser10 (pH3) (06-570, 1:1000 Merck Millipore), mouse anti-β-Galactosidase (Z3781, 1:400 Promega), mouse anti-Delta (C594.9B, 1:50 DSHB), and rabbit anti-eGFP (CAB4211, 1:400 Invitrogen). Secondary antibodies were used with the following dilutions: Alexa Fluor 488-conjugated goat anti-rabbit (A11008, 1:800 Invitrogen), Alexa Fluor 488-conjugated goat anti-mouse (A11001, 1:800 Invitrogen), Alexa Fluor 555-conjugated donkey anti-mouse (A31570, 1:800 Invitrogen) and Alexa Fluor 555-conjugated goat anti-rabbit (A21429, 1:800 Invitrogen). Images were taken from the posterior R4 region of the midgut with a Leica Application Suite X confocal microscope system. Images were processed using ImageJ/Fiji software.
2.4. Statistical Analysis
For quantitative analysis of cell populations, z-stacks were acquired from 20000 μm² square areas in the R4 region of each mounted midgut. Z-stacks were converted to maximum intensity projections, and cell numbers were manually counted using the Fiji Cell Counter plugin. For pH3-positive cell quantification, tile z-stacks covering the entire posterior midgut were acquired. The number of pH3-positive cells were determined by manual counting following confocal imaging. Statistical analyses were performed using GraphPad Prism 6 software. Statistical significance was denoted as follows: ns (not significant) >0.05, *P<0.05, **P<0.01, ***P<0.001, and ****P<0.0001. All representative images were independently replicated at least three times with consistent results.
3. Results
3.1. Hipk Is Required for Age-Related Hyperplasia
To elucidate the role of Hipk in age-associated intestinal hyperplasia, we generated hipk knockout flies using CRISPR/Cas9-mediated genome engineering, a technique that induces targeted small insertions or deletions (indels) [34,35,36]. Specifically, we utilized the esgts driver (esg-Gal4, UAS-GFP; tub-Gal80ts) to express UAS-Cas9 and UAS-hipk-sgRNA, thereby achieving progenitor-specific hipk deletion. The esgts system allows for temporal control of transgene expression in progenitors upon shifting the temperature from 22 °C to 29 °C. In control flies (esgts > UAS-Cas9), the progenitor population expanded progressively at 29 °C (Figure 1A, C,E), consistent with previous reports [17,18,19]. In contrast, hipk knockout flies (esgts > UAS-Cas9,UAS-hipk-sgRNA) maintained low levels of progenitors throughout the 20-day period (Figure 1B, D, F), indicating that Hipk is required for age-associated intestinal hyperplasia.
3.2. Hipk Is Required to Mediate DSS-Induced Hyperplasia
To evaluate the role of Hipk in DSS-induced hyperplasia, we employed esgts-driven UAS-hipk-RNAi, a line previously validated for hipk knockdown in progenitors [33]. We used the Su(H)Gbe-lacZ; esgts (esg-Gal4, UAS-GFP; tub-Gal80ts) reporter line to distinguish between ISCs (GFP+, lacZ-) and EBs (GFP+, lacZ+). In control midguts, the progenitor population (GFP+ cells) increased following two days of DSS treatment (compare Figure 2C with Figure 2A), accompanied by the formation of large progenitor clusters (averaging ~5 cells per cluster; Figure 2C), which is indicative of accelerated ISC proliferation. Notably, the EB population showed a dramatic increase (from 10% to 50%), whereas the ISC population increased more modestly (from 20% to 30%) (Figure 2G, H). Progenitor-specific hipk knockdown significantly suppressed this DSS-induced expansion and abolished the formation of progenitor clusters (compare Figure 2D with Figure 2C). In particular, hipk depletion resulted in a marked decrease in the EB population (from 50% to 2%) and a moderate decrease in the ISC population (from 30% to 10%) (Figure 2G, H). Consistent with these findings, mitotic activity, as assessed by phospho-histone H3 (pH3) staining, was significantly reduced by hipk knockdown (Figure 3). In contrast, hyperplasia induced by Pseudomonas entomophila (Pe) infection was not significantly attenuated by hipk knockdown (Figure 2E–H), suggesting that Hipk-mediated regulation is specific to DSS-induced injury rather than Pe-stimulated damage.
3.3. Hipk Is Essential for Yki-Driven Hyperplasia
Yki activation in progenitor cells promotes hyperproliferation in ISCs, resulting in intestinal hyperplasia [27,28]. We next tested whether Hipk is required for Yki-induced ISC hyperproliferation. Consistent with previous studies, Yki activation via wts knockdown in progenitors (esgts > wts RNAi) increased the progenitor population and induced tumor-like clusters (compare Figure 4D with Figure 4A), and ISC proliferation (Figure 5). Concurrent expression of wts RNAi and hipk RNAi in progenitors (esgts > wts RNAi, hipk RNAi) reduced the progenitor population and eliminated these clusters (compare Figure 4E with Figure 4D) and mitotic cells (Figure 5), indicating that the hyperplasia and ISC hyperproliferation induced by wts knockdown requires Hipk.
Similarly, hyperplasia and ISC hyperproliferation induced by progenitor-specific Yki overexpression (esgts > UAS-yki:GFP) were suppressed by hipk knockdown (compare Figure 6C with Figure 6A, Figure 7). The YkiS168A mutant is a constitutively active form refractory to Wts-mediated inhibition [37,38,39]. Progenitor-specific overexpression of YkiS168A (esgts > UAS-ykiS168A:GFP) increased the progenitor population (compare Figure 6D with Figure 4A). Strikingly, YkiS168A-mediated hyperplasia was not inhibited by concomitant hipk knockdown (esgts > UAS-ykiS168A:GFP, UAS-hipk RNAi) (compare Figure 6F with Figure 6D), nor was mitotic activity reduced (Figure 8).
3.4. Hipk Regulates Both Yki Protein Stability and Nuclear Localization
The differential effects of Hipk on Yki and YkiS168A suggest that Hipk regulates the stability or localization of wild-type Yki, but not that of the constitutively active mutant. To test this hypothesis, we utilized the esgTS driver (esg-Gal4, tub-GAL80ts; lacking UAS-GFP) to express UAS-Yki:GFP or UAS-YkiS168A:GFP, where the GFP signal serves as a proxy for Yki:GFP or YkiS168A:GFP protein levels in progenitors. In control progenitors, Yki:GFP signals were predominantly detectable in the cytoplasm (Figure 9A). Hipk overexpression elevated Yki:GFP levels and promoted its nuclear localization (compare Figure 9B with Figure 9A), whereas hipk knockdown rendered the Yki:GFP signal virtually undetectable (compare Figure 9C with Figure 9A).
In contrast, the signals from YkiS168A:GFP were localized primarily to the nucleus and remained robust, showing no response to Hipk manipulation (Figure 9D–F). These results demonstrate that Hipk is essential for the stability and nuclear accumulation of wild-type Yki, but is dispensable for YkiS168A.
4. Discussion
Hipk has recently been identified as a downstream effector of the insulin/Akt/TOR pathways in regulating ISC proliferation and progenitor populations [33]. Hipk expression is directly governed by insulin signaling; specifically, it is abolished by defects in insulin signaling within progenitors [33]. Here, we show that Hipk is required for age- and DSS-activated ISC proliferation and progenitor expansion. Thus, our findings suggest that the requirement for insulin signaling in age- and DSS-stimulated ISC proliferation and progenitor expansion is mediated by Hipk.
Dextran sulfate sodium (DSS)-activated ISC proliferation and progenitor expansion require Yki in progenitors [27,28]. Consistently, progenitor-specific Yki activation phenocopies the hyperplasia observed after DSS injury [27,28]. Here, we demonstrate that Hipk is required for DSS- and Yki-induced ISC hyperproliferation and gut hyperplasia. Strikingly, Hipk is dispensable for YkiS168A-induced ISC hyperproliferation and gut hyperplasia. When Yki:GFP is overexpressed using the esgts driver (esgts > UAS-Yki:GFP), the protein is weakly detectable in progenitors and predominantly sequestered in the cytoplasm, confirming active Hippo signaling under homeostatic conditions, in agreement with previous reports. Yki:GFP levels are further reduced, becoming essentially undetectable, upon hipk knockdown, whereas Hipk overexpression increases Yki:GFP protein levels and promotes its nuclear localization. These findings indicate that Yki protein stability and nuclear accumulation are Hipk-dependent in progenitors. In contrast, when the constitutively active YkiS168A:GFP is overexpressed (esgts > UAS-YkiS168A:GFP), protein levels remain high and predominantly nuclear, indicating resistance to Hippo signaling. Notably, Hipk manipulation does not affect the abundance or localization of YkiS168A, demonstrating that Hipk is dispensable for the activation of this mutant.
YkiS168A is a mutant allele in which Serine 168 is replaced by alanine, a non-phosphorylatable amino acid. Ser168 is the critical Wts phosphorylation site; its phosphorylation by Wts triggers Yki cytoplasmic sequestration and degradation [37,38,39,40,41]. Recent evidence suggests that Hipk phosphorylates Yki at Ser169 and Ser172, residues proximal to the Ser168 target site [42]. Our data suggest that Hipk-dependent phosphorylation at Ser169 and Ser172 may sterically or chemically inhibit Wts-mediated phosphorylation at Ser168, thereby relieving Wts-dependent suppression, stabilizing Yki, and promoting nuclear translocation. This model accounts for why Hipk is dispensable for YkiS168A, which is inherently refractory to Ser168 phosphorylation. Previous studies have established that phosphorylation at Ser169 and Ser172 modulates Yki activity. While phosphomimetic mutations (S169D/S172D) reduce Ser168 phosphorylation in S2 cells and enhance Yki activity in vivo, alanine substitutions (S169A/S172A), mimicking a lack of Hipk phosphorylation, increase Ser168 phosphorylation in S2 cells and suppress Yki activity in vivo [39].
The conservation of serine residues at positions +1 and +4 relative to Ser168 (e.g., Ser169 and Ser172 in Drosophila Yki) across Yki and YAP proteins suggests that mammalian Hipk may regulate YAP through a similar mechanism. Indeed, mammalian Hipk2 increases YAP abundance in cultured cells, suggesting an evolutionarily conserved regulatory axis [43]. Given the role of YAP in mammalian intestinal biology [44,45,46], our findings suggest that mammalian Hipk2 may similarly regulate intestinal hyperplasia, a process frequently associated with tumorigenesis.
Our data further show that hipk knockdown reduces DSS-stimulated progenitor expansion but fails to suppress expansion following infection by the bacterium Pseudomonas entomophila (Pe). This differential requirement for Hipk parallels that of Yki [28]. DSS activates Yki, as evidenced by nuclear localization, whereas Pe infection activates MAPK signaling in progenitors, indicated by elevated dpERK [27,47]. Specifically, DSS-induced ISC hyperproliferation is abolished by yki knockdown, whereas Pe-induced hyperproliferation is abolished by egfr knockdown [27,28,47]. Thus, Hipk, which integrates insulin and Hippo signaling (this study), appears to be decoupled from the EGFR signaling pathway. Consistent with this, hipk expression is regulated by insulin signaling but does not require EGFR signaling, nor does forced EGFR activation affect hipk expression [33]. This explains why Hipk is required for DSS-stimulated, but not Pe-stimulated, ISC proliferation (this study).
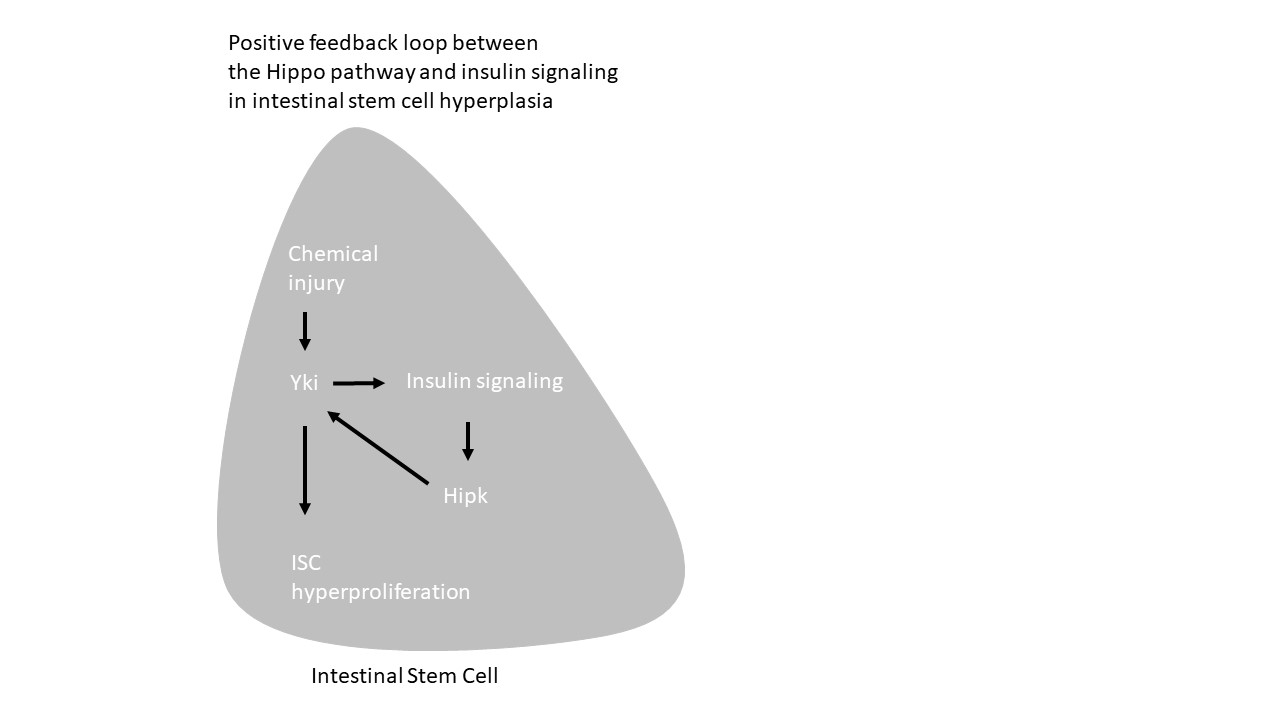
It is worth noting that previous studies demonstrated that progenitor-specific overexpression of active Yki increases levels of dilp3, InR and Akt, all of thes are components of insuling signaling, in the midgut, indicating that Yki activation strengthening insulin signaling in the midgut [48]. It is recently shown that insulin signaling induces expression of Hipk in progenitors [33]. Here, we show that Hipk directly regulates Yki protein activity. Thus our studies reveal a positive feedback loop between insulin signaling and hippo signaling in progenitors via Hipk, which integrates insuling signaling to Hippo pathway during regeneration. Future studies should determine whether this positive feedback loop is conserved in mammalian tissues.
Author Contributions
X.W. performed the experiments and analyzed the data. H.C.L. and C.K. conceived the study and wrote the manuscript.
Funding
This research was supported by a grant from the National Research Foundation of Korea (NRF) grants (NRF-2021R1A2C1010334 to C.K.) and NRF funded by the Korea government (MSIT) (RS-2026-25479096 to H.C.L.).
Institutional Review Board Statement
NA.
Informed Consent Statement
NA.
Data Availability Statement
Data supporting the results of this study are available from the corresponding author on reasonable request.
Acknowledgments
We thank the Bloomington
Drosophila
Stock Center (BDSC) and the Vienna
Drosophila
RNAi Center (VDRC) for providing fly stocks. We are grateful to Drs N. Perrimon, B. Edgar, E. Verheyen, M. Boutros, K. Irvine, S. Cohen for generously sharing reagents.
Conflicts of Interest
The authors have no conflicts of interest to disclose.
References
- Yu, Z.; Zhu, Y.; Chen, Y.; Feng, C.; Zhang, Z.; Guo, X.; Chen, H.; Liu, X.; Yuan, Y.; Chen, H. Nutrient-sensing alteration leads to age-associated distortion of intestinal stem cell differentiating direction. Nat. Commun. 2024, 15(1), 9243. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Lucchetta, E.; Rafel, N.; Ohlstein, B. Maintenance of the adult Drosophila intestine: all roads lead to homeostasis. Curr. Opin. Genet Dev. 2016, 40, 81–86. [Google Scholar] [CrossRef]
- Jiang, H.; Tian, A.; Jiang, J. Intestinal stem cell response to injury: lessons from Drosophila. Cell Mol. Life Sci. 2016, 73(17), 3337–3349. [Google Scholar] [CrossRef]
- Jiang, H.; Edgar, B.A. Intestinal stem cells in the adult Drosophila midgut. Exp. Cell Res. 2011, 317(19), 2780–2788. [Google Scholar] [CrossRef]
- Zhou, J.; Boutros, M. Intestinal stem cells and their niches in homeostasis and disease. Cells Dev. 2023, 175, 203862. [Google Scholar] [CrossRef] [PubMed]
- Micchelli, C.A. Perrimon N: Evidence that stem cells reside in the adult Drosophila midgut epithelium. Nature 2006, 439(7075), 475–479. [Google Scholar] [CrossRef]
- Ohlstein, B. Spradling A: The adult Drosophila posterior midgut is maintained by pluripotent stem cells. Nature 2006, 439(7075), 470–474. [Google Scholar] [CrossRef]
- O’Brien, L.E.; Soliman, S.S.; Li, X.; Bilder, D. Altered modes of stem cell division drive adaptive intestinal growth. Cell 2011, 147(3), 603–614. [Google Scholar] [CrossRef]
- Lin, G.; Xi, R. Intestinal stem cell, muscular niche and Wingless signaling. Fly 2008, 2(6), 310–312. [Google Scholar] [CrossRef]
- Joly, A.; Rousset, R. Tissue Adaptation to Environmental Cues by Symmetric and Asymmetric Division Modes of Intestinal Stem Cells. Int. J. Mol. Sci. 2020, 21(17). [Google Scholar] [CrossRef] [PubMed]
- Biteau, B.; Jasper, H. EGF signaling regulates the proliferation of intestinal stem cells in Drosophila. Development 2011, 138(6), 1045–1055. [Google Scholar] [CrossRef]
- Ohlstein B, Spradling A: Multipotent Drosophila intestinal stem cells specify daughter cell fates by differential notch signaling. Science 2007, 315(5814), 988–992. [CrossRef] [PubMed]
- Zeng, X.; Hou, S.X. Enteroendocrine cells are generated from stem cells through a distinct progenitor in the adult Drosophila posterior midgut. Development 2015, 142(4), 644–653. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Tian, A.; Jiang, J. Numb provides a fail-safe mechanism for intestinal stem cell self-renewal in adult Drosophila midgut. bioRxiv 2024. [Google Scholar]
- de Navascues, J.; Perdigoto, C.N.; Bian, Y.; Schneider, M.H.; Bardin, A.J.; Martinez-Arias, A.; Simons, B.D. Drosophila midgut homeostasis involves neutral competition between symmetrically dividing intestinal stem cells. EMBO J. 2012, 31(11), 2473–2485. [Google Scholar] [CrossRef] [PubMed]
- Amcheslavsky, A.; Jiang, J. Ip YT: Tissue damage-induced intestinal stem cell division in Drosophila. Cell Stem Cell 2009, 4(1), 49–61. [Google Scholar] [CrossRef]
- Biteau, B.; Hochmuth, C.E.; Jasper, H. JNK activity in somatic stem cells causes loss of tissue homeostasis in the aging Drosophila gut. Cell Stem Cell 2008, 3(4), 442–455. [Google Scholar] [CrossRef]
- Choi, N.H.; Kim, J.G.; Yang, D.J.; Kim, Y.S.; Yoo, M.A. Age-related changes in Drosophila midgut are associated with PVF2, a PDGF/VEGF-like growth factor. Aging Cell 2008, 7(3), 318–334. [Google Scholar] [CrossRef]
- Biteau, B.; Karpac, J.; Supoyo, S.; Degennaro, M.; Lehmann, R.; Jasper, H. Lifespan extension by preserving proliferative homeostasis in Drosophila. PLoS Genet 2010, 6(10), e1001159. [Google Scholar] [CrossRef]
- Huang, J.; Wu, S.; Barrera, J.; Matthews, K.; Pan, D. The Hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating Yorkie, the Drosophila Homolog of YAP. Cell 2005, 122(3), 421–434. [Google Scholar] [CrossRef]
- Pan D: The hippo signaling pathway in development and cancer. Dev. Cell 2010, 19(4), 491–505. [CrossRef]
- Staley, B.K.; Irvine, K.D. Hippo signaling in Drosophila: recent advances and insights. Dev. Dyn. 2012, 241(1), 3–15. [Google Scholar] [CrossRef] [PubMed]
- Misra, J.R. Irvine KD: The Hippo Signaling Network and Its Biological Functions. Annu Rev. Genet 2018, 52, 65–87. [Google Scholar] [CrossRef]
- Ma, X.; Guo, X.; Richardson, H.E.; Xu, T.; Xue, L. POSH regulates Hippo signaling through ubiquitin-mediated expanded degradation. Proc. Natl. Acad. Sci. U S A 2018, 115(9), 2150–2155. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Li, P.; Lu, Y.; Tao, J.; Hao, X.; Wang, X.; Wu, W.; Xu, J.; Zhang, H.; Li, X. A feedback loop between Paxillin and Yorkie sustains Drosophila intestinal homeostasis and regeneration. Nat. Commun. 2025, 16(1), 570. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Li, Q.; Nirala, N.K.; Ip, Y.T. The Snakeskin-Mesh Complex of Smooth Septate Junction Restricts Yorkie to Regulate Intestinal Homeostasis in Drosophila. Stem Cell Rep. 2020, 14(5), 828–844. [Google Scholar] [CrossRef]
- Karpowicz, P.; Perez, J.; Perrimon, N. The Hippo tumor suppressor pathway regulates intestinal stem cell regeneration. Development 2010, 137(24), 4135–4145. [Google Scholar] [CrossRef]
- Ren, F.; Wang, B.; Yue, T.; Yun, E.Y.; Ip, Y.T.; Jiang, J. Hippo signaling regulates Drosophila intestine stem cell proliferation through multiple pathways. Proc. Natl. Acad. Sci. U S A 2010, 107(49), 21064–21069. [Google Scholar] [CrossRef]
- Shaw, R.L.; Kohlmaier, A.; Polesello, C.; Veelken, C.; Edgar, B.A.; Tapon, N. The Hippo pathway regulates intestinal stem cell proliferation during Drosophila adult midgut regeneration. Development 2010, 137(24), 4147–4158. [Google Scholar] [CrossRef]
- Staley, B.K.; Irvine, K.D. Warts and Yorkie mediate intestinal regeneration by influencing stem cell proliferation. Curr. Biol. 2010, 20(17), 1580–1587. [Google Scholar] [CrossRef]
- Li, Q.; Li, S.; Mana-Capelli, S.; Roth Flach, R.J.; Danai, L.V.; Amcheslavsky, A.; Nie, Y.; Kaneko, S.; Yao, X.; Chen, X. The conserved misshapen-warts-Yorkie pathway acts in enteroblasts to regulate intestinal stem cells in Drosophila. Dev. Cell 2014, 31(3), 291–304. [Google Scholar] [CrossRef]
- Li, Q.; Nirala, N.K.; Nie, Y.; Chen, H.J.; Ostroff, G.; Mao, J.; Wang, Q.; Xu, L.; Ip. YT: Ingestion of Food Particles Regulates the Mechanosensing Misshapen-Yorkie Pathway in Drosophila Intestinal Growth. Dev. Cell 2018, 45(4), 433–449 e436. [Google Scholar] [CrossRef]
- Wu, X.; Kim, H.; Jang, W.; Kim, C. Hipk transduces nutrient signals to control intestinal stem cell proliferation and fate in Drosophila. Sci. Rep. 2026. [Google Scholar] [CrossRef]
- Port, F.; Boutros, M. Tissue-Specific CRISPR-Cas9 Screening in Drosophila. Methods Mol. Biol. 2022, 2540, 157–176. [Google Scholar]
- Bahuguna, S.; Redhai, S.; Zhou, J.; Wang, T.; Port, F.; Boutros, M. Conditional CRISPR-Cas Genome Editing in Drosophila to Generate Intestinal Tumors. Cells 2021, 10(11). [Google Scholar] [CrossRef] [PubMed]
- Port, F.; Strein, C.; Stricker, M.; Rauscher, B.; Heigwer, F.; Zhou, J.; Beyersdorffer, C.; Frei, J.; Hess, A.; Kern, K. A large-scale resource for tissue-specific CRISPR mutagenesis in Drosophila. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Feldmann, G.; Huang, J.; Wu, S.; Zhang, N.; Comerford, S.A.; Gayyed, M.F.; Anders, R.A.; Maitra, A.; Pan, D. Elucidation of a universal size-control mechanism in Drosophila and mammals. Cell 2007, 130(6), 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Irvine, K.D. In vivo regulation of Yorkie phosphorylation and localization. Development 2008, 135(6), 1081–1088. [Google Scholar] [CrossRef]
- Oh, H.; Irvine, K.D. In vivo analysis of Yorkie phosphorylation sites. Oncogene 2009, 28(17), 1916–1927. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, F.; Zhang, Q.; Chen, Y.; Wang, B.; Jiang, J. The TEAD/TEF family of transcription factor Scalloped mediates Hippo signaling in organ size control. Dev. Cell 2008, 14(3), 377–387. [Google Scholar] [CrossRef]
- Ren, F.; Zhang, L.; Jiang, J. Hippo signaling regulates Yorkie nuclear localization and activity through 14-3-3 dependent and independent mechanisms. Dev. Biol. 2010, 337(2), 303–312. [Google Scholar] [CrossRef]
- Steinmetz, E.L.; Dewald, D.N.; Walldorf, U. Drosophila Homeodomain-Interacting Protein Kinase (Hipk) Phosphorylates the Hippo/Warts Signalling Effector Yorkie. Int. J. Mol. Sci. 2021, 22(4). [Google Scholar] [CrossRef]
- Poon, C.L.; Zhang, X.; Lin, J.I.; Manning, S.A.; Harvey, K.F. Homeodomain-interacting protein kinase regulates Hippo pathway-dependent tissue growth. Curr. Biol. 2012, 22(17), 1587–1594. [Google Scholar] [CrossRef]
- Kaur, M.; Mungurere, R.F.; Mitinje, N.; Sethi, G.K.; Kaur, A.S.; Mishra, A. Hippo-YAP/TAZ signaling in gastric cancer: molecular pathogenesis and emerging therapeutic horizons. Med. Oncol. 2026, 43(3), 147. [Google Scholar] [CrossRef]
- Camargo, F.D.; Gokhale, S.; Johnnidis, J.B.; Fu, D.; Bell, G.W.; Jaenisch, R.; Brummelkamp, T.R. YAP1 increases organ size and expands undifferentiated progenitor cells. Curr. Biol. 2007, 17(23), 2054–2060. [Google Scholar] [CrossRef]
- Cao, H.; Huang, X.; Jiang, X.; Deng, J.; Wang, J.; Wu, C.; Hu, M.; Zeng, B.; Hu, Z.; Pan, H. The WNK-OXSR1 osmosensing pathway mediates intestinal regeneration via Hippo-YAP signaling. EMBO J. 2026. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Grenley, M.O.; Bravo, M.J.; Blumhagen, R.Z.; Edgar, B.A. EGFR/Ras/MAPK signaling mediates adult midgut epithelial homeostasis and regeneration in Drosophila. Cell Stem Cell 2011, 8(1), 84–95. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.; Song, W.; Droujinine, I.A.; Hu, Y.; Asara, J.M. Perrimon N: Systemic organ wasting induced by localized expression of the secreted insulin/IGF antagonist ImpL2. Dev. Cell 2015, 33(1), 36–46. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
hipk is required for the age-dependent expansion of the progenitor population. (A–F) Representative confocal images of the posterior midgut (R4 region) from flies expressing in progenitors: cas9 (esgts > UAS-cas9) (control; A, C, E) or cas9 and hipk-sgRNA (esgts > UAS-cas9 + UAS-hipk-sgRNA) (hipk knockout in progenitors; B, D, F) aged for 2, 15, and 20 days at 29 °C. The esgts driver (esg-Gal4, tub-Gal80ts, UAS-GFP) marks progenitor cells and drives expression of UAS-transgenes upon shift to 29 °C. GFP labels progenitors (green); DAPI labels nuclei (blue). The progenitor population expands to occupy the majority of the gut epithelium with age in control guts (E), but this expansion is significantly attenuated in hipk knockout guts (F). Scale bar: 20 µm. (G) Schematic diagram showing the superficial view of progenitors (green, small diploid cells) and enterocytes (ECs, blue, large polyploid cells) in the wild-type midgut. (H) Schematic diagram of the Drosophila midgut illustrating distinct regions. The R4 region was analyzed for all confocal images presented throughout this manuscript.
Figure 1.
hipk is required for the age-dependent expansion of the progenitor population. (A–F) Representative confocal images of the posterior midgut (R4 region) from flies expressing in progenitors: cas9 (esgts > UAS-cas9) (control; A, C, E) or cas9 and hipk-sgRNA (esgts > UAS-cas9 + UAS-hipk-sgRNA) (hipk knockout in progenitors; B, D, F) aged for 2, 15, and 20 days at 29 °C. The esgts driver (esg-Gal4, tub-Gal80ts, UAS-GFP) marks progenitor cells and drives expression of UAS-transgenes upon shift to 29 °C. GFP labels progenitors (green); DAPI labels nuclei (blue). The progenitor population expands to occupy the majority of the gut epithelium with age in control guts (E), but this expansion is significantly attenuated in hipk knockout guts (F). Scale bar: 20 µm. (G) Schematic diagram showing the superficial view of progenitors (green, small diploid cells) and enterocytes (ECs, blue, large polyploid cells) in the wild-type midgut. (H) Schematic diagram of the Drosophila midgut illustrating distinct regions. The R4 region was analyzed for all confocal images presented throughout this manuscript.

Figure 2.
Hipk is required for DSS-induced progenitor cell expansion. (A–F) Representative confocal images of posterior midguts from control (Gbe-lacZ; esgts) (A, C, E) and hipk-knockdown (Gbe-lacZ; esgts, hipk-RNAi) (B, D, F) flies treated with sucrose (A, B), DSS (C, D), or Pseudomonas entomophila (Pe) (E, F) for two days at 29 °C. esgts refers to esg-Gal4, UAS-GFP, tub-Gal80ts; hipk-RNAi denotes UAS-hipk-RNAi. GFP labels progenitor cells (esg-expressing cells); β-Gal labels EBs (Gbe-lacZ, a reporter for EBs); DAPI labels nuclei. Scale bars: 10 µm. Note that the progenitor population in DSS-treated midguts is significantly reduced upon hipk knockdown (compare C and D). (G, H) Quantification of the ratios of ISCs to total cells (G) and EBs to total cells (H) in the midguts of the indicated genotypes (mean ± SEM, n = 5). p-values were determined by two-tailed unpaired t-test: ns (not significant), p > 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001.
Figure 2.
Hipk is required for DSS-induced progenitor cell expansion. (A–F) Representative confocal images of posterior midguts from control (Gbe-lacZ; esgts) (A, C, E) and hipk-knockdown (Gbe-lacZ; esgts, hipk-RNAi) (B, D, F) flies treated with sucrose (A, B), DSS (C, D), or Pseudomonas entomophila (Pe) (E, F) for two days at 29 °C. esgts refers to esg-Gal4, UAS-GFP, tub-Gal80ts; hipk-RNAi denotes UAS-hipk-RNAi. GFP labels progenitor cells (esg-expressing cells); β-Gal labels EBs (Gbe-lacZ, a reporter for EBs); DAPI labels nuclei. Scale bars: 10 µm. Note that the progenitor population in DSS-treated midguts is significantly reduced upon hipk knockdown (compare C and D). (G, H) Quantification of the ratios of ISCs to total cells (G) and EBs to total cells (H) in the midguts of the indicated genotypes (mean ± SEM, n = 5). p-values were determined by two-tailed unpaired t-test: ns (not significant), p > 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001.

Figure 3.
hipk is required for DSS-stimulated ISC hyperproliferation. (A–D) Representative confocal images of midguts from control (esgts > GFP; A, C) and progenitor-specific hipk knockdown (esgts > GFP, hipk-RNAi; B, D) flies treated with either 5% sucrose (control; A, B) or 5% DSS (C, D) for 2 days at 29 °C. Anti-Delta marks ISCs (green); pH3 marks mitotic cells (red); DAPI marks nuclei (blue). Scale bars: 20 µm. (E) Quantification of pH3+ cells per midgut in adults of the indicated genotypes (mean ± SEM, n = 9). p-values were determined by two-tailed unpaired t-test: ** p < 0.01, **** p < 0.0001.
Figure 3.
hipk is required for DSS-stimulated ISC hyperproliferation. (A–D) Representative confocal images of midguts from control (esgts > GFP; A, C) and progenitor-specific hipk knockdown (esgts > GFP, hipk-RNAi; B, D) flies treated with either 5% sucrose (control; A, B) or 5% DSS (C, D) for 2 days at 29 °C. Anti-Delta marks ISCs (green); pH3 marks mitotic cells (red); DAPI marks nuclei (blue). Scale bars: 20 µm. (E) Quantification of pH3+ cells per midgut in adults of the indicated genotypes (mean ± SEM, n = 9). p-values were determined by two-tailed unpaired t-test: ** p < 0.01, **** p < 0.0001.

Figure 4.
Hipk is required for wts-knockdown-stimulated progenitor expansion. (A–E) Representative confocal images of posterior midguts from control (esgts > GFP) (A), hipk-overexpression (esgts > hipk) (B), hipk-knockdown (esgts > hipk RNAi) (C), wts-knockdown (esgts > wts RNAi) (D), and wts and hipk co-knockdown (esgts > wts RNAi + hipk RNAi) (E). Flies were treated for 7 days at 29 °C. The esgts driver denotes esg-Gal4, UAS-GFP, tub-Gal80ts. Progenitor cells (esg-expressing cells) are marked by GFP expression; nuclei are stained with DAPI. Note that the progenitor expansion induced by wts knockdown is suppressed by the co-depletion of hipk (compare D and E). wts knockdown induces clusters of cells comprising ~5 cells, which are encircled (D). Scale bars: 20 µm.
Figure 4.
Hipk is required for wts-knockdown-stimulated progenitor expansion. (A–E) Representative confocal images of posterior midguts from control (esgts > GFP) (A), hipk-overexpression (esgts > hipk) (B), hipk-knockdown (esgts > hipk RNAi) (C), wts-knockdown (esgts > wts RNAi) (D), and wts and hipk co-knockdown (esgts > wts RNAi + hipk RNAi) (E). Flies were treated for 7 days at 29 °C. The esgts driver denotes esg-Gal4, UAS-GFP, tub-Gal80ts. Progenitor cells (esg-expressing cells) are marked by GFP expression; nuclei are stained with DAPI. Note that the progenitor expansion induced by wts knockdown is suppressed by the co-depletion of hipk (compare D and E). wts knockdown induces clusters of cells comprising ~5 cells, which are encircled (D). Scale bars: 20 µm.

Figure 5.
Hipk is required for wts-knockdown-induced ISC hyperproliferation. (A–E) Representative confocal images of the posterior midgut from adult flies expressing in progenitors: control (esgts > GFP) (A), hipk (esgts > hipk) (B), hipk-RNAi (esgts > hipk-RNAi) (C), wts-RNAi (esgts > wts-RNAi) (D), and wts-RNAi + hipk-RNAi (esgts > wts-RNAi + hipk-RNAi) (E). Transgenes were expressed using the esgts driver for 7 days at 29 °C. The esgts driver denotes esg-Gal4, UAS-GFP, tub-Gal80ts. Anti-Delta labels ISCs (green); pH3 labels mitotic cells (red); DAPI marks nuclei (blue). Note that the increase in mitotic cells induced by wts knockdown is suppressed by the co-depletion of hipk (compare D and E). Scale bars: 10 µm. (F) Quantification of pH3+ cells in midguts of the indicated genotypes (mean ± SEM; n = 15, 9, 12, 10, and 15, respectively). p-values were determined by two-tailed unpaired t-test: * p < 0.05; ** p < 0.01; **** p < 0.0001.
Figure 5.
Hipk is required for wts-knockdown-induced ISC hyperproliferation. (A–E) Representative confocal images of the posterior midgut from adult flies expressing in progenitors: control (esgts > GFP) (A), hipk (esgts > hipk) (B), hipk-RNAi (esgts > hipk-RNAi) (C), wts-RNAi (esgts > wts-RNAi) (D), and wts-RNAi + hipk-RNAi (esgts > wts-RNAi + hipk-RNAi) (E). Transgenes were expressed using the esgts driver for 7 days at 29 °C. The esgts driver denotes esg-Gal4, UAS-GFP, tub-Gal80ts. Anti-Delta labels ISCs (green); pH3 labels mitotic cells (red); DAPI marks nuclei (blue). Note that the increase in mitotic cells induced by wts knockdown is suppressed by the co-depletion of hipk (compare D and E). Scale bars: 10 µm. (F) Quantification of pH3+ cells in midguts of the indicated genotypes (mean ± SEM; n = 15, 9, 12, 10, and 15, respectively). p-values were determined by two-tailed unpaired t-test: * p < 0.05; ** p < 0.01; **** p < 0.0001.

Figure 6.
Hipk is required for Yki-stimulated progenitor expansion but is dispensable for YkiS168A-induced hyperplasia. (A–F) Representative confocal images of posterior midguts expressing in progenitors: Yki (UAS-yki:GFP) (A), yki and hipk (UAS-yki:GFP + UAS-hipk) (B), yki and hipk-RNAi (UAS-yki:GFP + hipk-RNAi) (C), ykiS168A (UAS-ykiS168A:GFP) (D), ykiS168A and hipk (UAS- YkiS168A:GFP + UAS-hipk) (E), and ykiS168A and hipk-RNAi (UAS-ykiS168A:GFP + hipk-RNAi) (F). esgts refers to esg-Gal4, tub-Gal80ts, UAS-GFP. Flies were treated for 7 days at 29 °C. GFP marks progenitor cells (esg-expressing cells); DAPI marks nuclei. Note that the progenitor population induced by Yki:GFP is reduced by concomitant hipk knockdown (C), whereas the population induced by the constitutively active form, ykiS168A:GFP, remains unaffected by hipk knockdown (F). Scale bar: 20 µm.
Figure 6.
Hipk is required for Yki-stimulated progenitor expansion but is dispensable for YkiS168A-induced hyperplasia. (A–F) Representative confocal images of posterior midguts expressing in progenitors: Yki (UAS-yki:GFP) (A), yki and hipk (UAS-yki:GFP + UAS-hipk) (B), yki and hipk-RNAi (UAS-yki:GFP + hipk-RNAi) (C), ykiS168A (UAS-ykiS168A:GFP) (D), ykiS168A and hipk (UAS- YkiS168A:GFP + UAS-hipk) (E), and ykiS168A and hipk-RNAi (UAS-ykiS168A:GFP + hipk-RNAi) (F). esgts refers to esg-Gal4, tub-Gal80ts, UAS-GFP. Flies were treated for 7 days at 29 °C. GFP marks progenitor cells (esg-expressing cells); DAPI marks nuclei. Note that the progenitor population induced by Yki:GFP is reduced by concomitant hipk knockdown (C), whereas the population induced by the constitutively active form, ykiS168A:GFP, remains unaffected by hipk knockdown (F). Scale bar: 20 µm.

Figure 7.
hipk is required for wild-type Yki-stimulated ISC hyperproliferation. (A–C) Representative confocal images of the posterior midgut from adult flies expressing the indicated constructs in progenitor cells using the esgts driver for 7 days at 29 °C. (A) esgts > yki::GFP. (B) esgts > yki::GFP + hipk. (C) esgts > yki::GFP + hipk-RNAi (II). The esgts driver denotes esg-Gal4, UAS-GFP, tub-Gal80ts. Anti-Delta labels ISCs (green); pH3 labels mitotic cells (red); DAPI marks nuclei (blue). Scale bars: 10 µm. (D) Quantification of pH3+ cells in midguts of the indicated genotypes (mean ± SEM; n = 15). p-values were determined by two-tailed unpaired t-test: ns (not significant), p > 0.05; **** p < 0.0001.
Figure 7.
hipk is required for wild-type Yki-stimulated ISC hyperproliferation. (A–C) Representative confocal images of the posterior midgut from adult flies expressing the indicated constructs in progenitor cells using the esgts driver for 7 days at 29 °C. (A) esgts > yki::GFP. (B) esgts > yki::GFP + hipk. (C) esgts > yki::GFP + hipk-RNAi (II). The esgts driver denotes esg-Gal4, UAS-GFP, tub-Gal80ts. Anti-Delta labels ISCs (green); pH3 labels mitotic cells (red); DAPI marks nuclei (blue). Scale bars: 10 µm. (D) Quantification of pH3+ cells in midguts of the indicated genotypes (mean ± SEM; n = 15). p-values were determined by two-tailed unpaired t-test: ns (not significant), p > 0.05; **** p < 0.0001.

Figure 8.
Hipk is dispensable for constitutively active Yki-stimulated ISC hyperproliferation. (A–C) Representative confocal images of the posterior midgut from adult flies expressing the indicated constructs in progenitor cells using the esgts driver for 7 days at 29 °C. (A) esgts > ykiS168A::GFP. (B) esgts > ykiS168A::GFP + hipk. (C) esgts > ykiS168A::GFP + hipk-RNAi (III). The esgts driver denotes esg-Gal4, UAS-GFP, tub-Gal80ts. Anti-Delta (green) labels ISCs; pH3 (red) labels mitotic cells; DAPI (blue) marks nuclei. Scale bars: 10 µm. (D) Quantification of pH3+ cells in midguts of the indicated genotypes (mean ± SEM; n = 9, 12, and 9, respectively). p-values were determined by two-tailed unpaired t-test; ns (not significant), p > 0.05.
Figure 8.
Hipk is dispensable for constitutively active Yki-stimulated ISC hyperproliferation. (A–C) Representative confocal images of the posterior midgut from adult flies expressing the indicated constructs in progenitor cells using the esgts driver for 7 days at 29 °C. (A) esgts > ykiS168A::GFP. (B) esgts > ykiS168A::GFP + hipk. (C) esgts > ykiS168A::GFP + hipk-RNAi (III). The esgts driver denotes esg-Gal4, UAS-GFP, tub-Gal80ts. Anti-Delta (green) labels ISCs; pH3 (red) labels mitotic cells; DAPI (blue) marks nuclei. Scale bars: 10 µm. (D) Quantification of pH3+ cells in midguts of the indicated genotypes (mean ± SEM; n = 9, 12, and 9, respectively). p-values were determined by two-tailed unpaired t-test; ns (not significant), p > 0.05.

Figure 9.
Hipk regulates the stability of wild-type Yki protein but is dispensable for the constitutively active YkiS168A protein. (A–F) Representative confocal images of posterior midguts expressing in progenitors: yki (UAS-Yki:GFP) (A), yki and hipk (UAS-yki:GFP + UAS-hipk) (B), yki and hipk-RNAi (UAS-Yki:GFP + hipk-RNAi) (C), ykiS168A (UAS-ykiS168A:GFP) (D), ykiS168A and hipk (UAS-ykiS168A:GFP + UAS-hipk) (E), and ykiS168A and hipk-RNAi (UAS-ykiS168A:GFP + hipk-RNAi) (F). UAS-transgenes were expressed for 7 days at 29 °C. The esgts driver denotes esg-Gal4, tub-Gal80ts. GFP fluorescence signals label Yki:GFP and YkiS168A:GFP proteins. Note that Yki:GFP is predominantly cytoplasmic in control cells (A, arrows), elevated and nuclear-localized upon hipk overexpression (B, arrows), and, remarkably, is undetectable in hipk-knockdown progenitors (C, arrows). In contrast, YkiS168A:GFP is primarily nuclear-localized in control cells (D, arrow). Nuclear localization of YkiS168A:GFP is not altered by hipk overexpression (E, arrow) or hipk knockdown (F, arrow). DAPI marks nuclei. Scale bars: 10 µm.
Figure 9.
Hipk regulates the stability of wild-type Yki protein but is dispensable for the constitutively active YkiS168A protein. (A–F) Representative confocal images of posterior midguts expressing in progenitors: yki (UAS-Yki:GFP) (A), yki and hipk (UAS-yki:GFP + UAS-hipk) (B), yki and hipk-RNAi (UAS-Yki:GFP + hipk-RNAi) (C), ykiS168A (UAS-ykiS168A:GFP) (D), ykiS168A and hipk (UAS-ykiS168A:GFP + UAS-hipk) (E), and ykiS168A and hipk-RNAi (UAS-ykiS168A:GFP + hipk-RNAi) (F). UAS-transgenes were expressed for 7 days at 29 °C. The esgts driver denotes esg-Gal4, tub-Gal80ts. GFP fluorescence signals label Yki:GFP and YkiS168A:GFP proteins. Note that Yki:GFP is predominantly cytoplasmic in control cells (A, arrows), elevated and nuclear-localized upon hipk overexpression (B, arrows), and, remarkably, is undetectable in hipk-knockdown progenitors (C, arrows). In contrast, YkiS168A:GFP is primarily nuclear-localized in control cells (D, arrow). Nuclear localization of YkiS168A:GFP is not altered by hipk overexpression (E, arrow) or hipk knockdown (F, arrow). DAPI marks nuclei. Scale bars: 10 µm.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



