Submitted:
26 May 2026
Posted:
27 May 2026
You are already at the latest version
Abstract
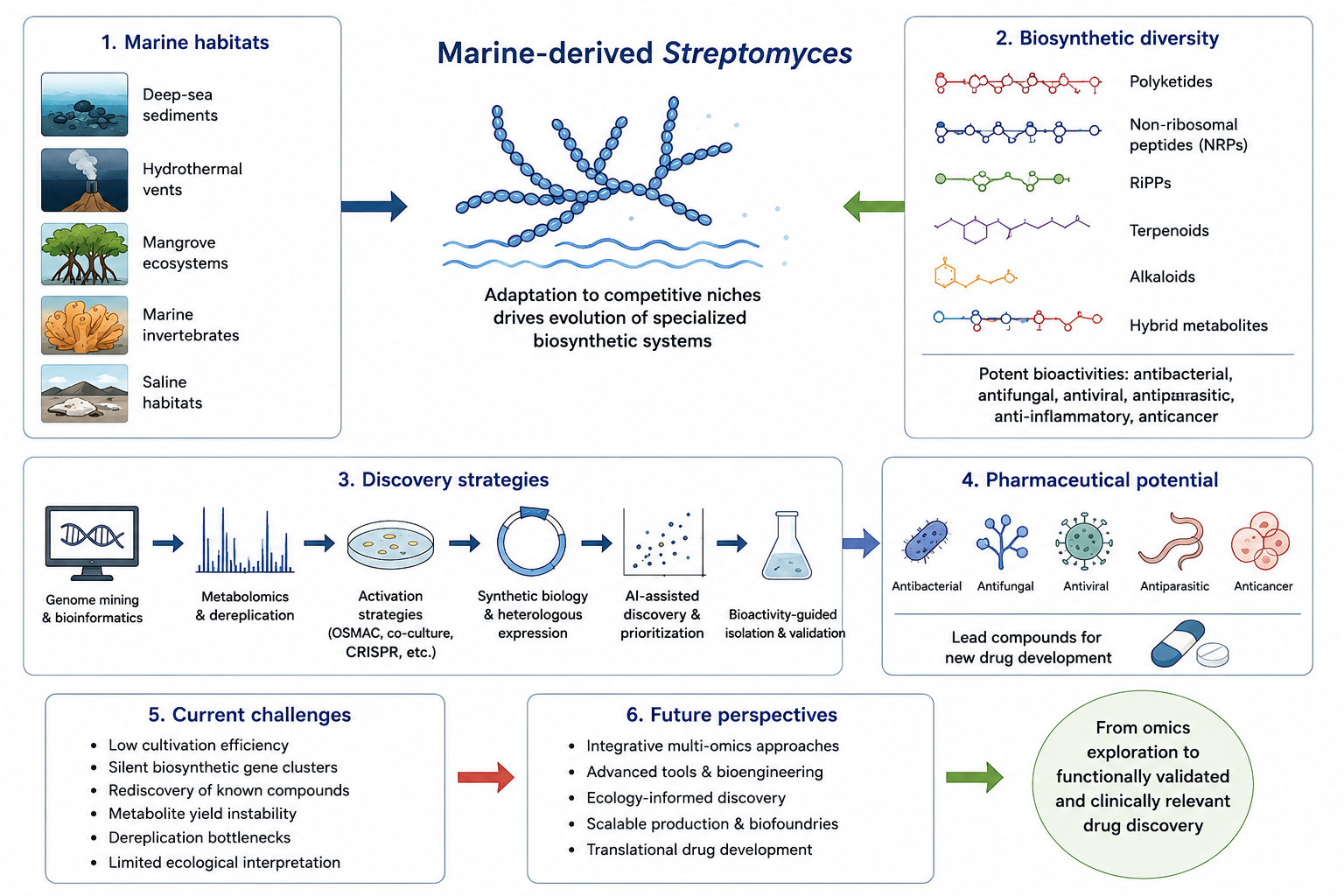
Marine-derived Streptomyces have emerged as one of the most prolific microbial resources for structurally diverse and pharmacologically significant natural products. Adaptation to highly competitive marine ecological niches, including deep-sea sediments, hydrothermal vents, mangrove ecosystems, marine invertebrates, and saline habitats, has driven the evolution of specialized biosynthetic systems capable of generating chemically complex secondary metabolites. Recent advances in genome sequencing, bioinformatics, metabolomics, synthetic biology, and artificial intelligence-assisted discovery pipelines have significantly expanded understanding of cryptic biosynthetic gene clusters (BGCs) encoded within marine actinomycete genomes. These BGCs produce structurally diverse metabolites including polyketides, non-ribosomal peptides, ribosomally synthesized and post-translationally modified peptides (RiPPs), terpenoids, alkaloids, and hybrid metabolites with potent antibacterial, antifungal, antiviral, antiparasitic, anti-inflammatory, and anticancer activities. Nevertheless, despite rapid technological progress, major translational limitations remain, including low cultivation efficiency, silent pathway expression, rediscovery of known compounds, metabolite yield instability, dereplication bottlenecks, and insufficient ecological interpretation. This critical review comprehensively discusses the biosynthetic diversity of marine Streptomyces-derived metabolites with emphasis on biosynthetic pathways, enzymatic tailoring reactions, genome mining strategies, metabolomics-guided discovery, synthetic biology approaches, and pharmaceutical applications. In addition, current limitations and future perspectives in marine natural product research are critically evaluated to highlight the transition from descriptive omics-driven exploration toward functionally validated and translationally relevant drug discovery.
Keywords:
marine Streptomyces
; biosynthetic gene clusters
; marine natural products
; genome mining
; polyketides
; non-ribosomal peptides
; synthetic biology
; metabolomics
; drug discovery
1. Introduction
Marine ecosystems represent one of the largest reservoirs of microbial diversity on Earth and harbor a vast number of microorganisms capable of producing structurally unprecedented secondary metabolites [1,2]. Among marine microorganisms, members of the genus Streptomyces have received exceptional scientific attention because of their remarkable biosynthetic capabilities and their historical contribution to antibiotic discovery [3,4]. Streptomyces species are Gram-positive filamentous actinomycetes characterized by large linear chromosomes enriched with biosynthetic gene clusters (BGCs) responsible for the production of structurally diverse natural products with important pharmaceutical relevance [4,5]. Historically, terrestrial Streptomyces contributed substantially to modern medicine through the discovery of clinically important compounds including streptomycin, chloramphenicol, tetracycline, and avermectin [5]. However, conventional terrestrial screening approaches gradually became less productive because of the frequent rediscovery of structurally known metabolites and declining rates of chemical novelty [6]. Consequently, marine-derived Streptomyces have emerged as alternative and highly promising resources for the discovery of chemically unique bioactive compounds [7,8,9].
Marine environments impose highly distinctive physicochemical pressures including elevated salinity, fluctuating hydrostatic pressure, nutrient limitation, oxidative stress, and intense microbial competition [8,9]. These ecological constraints are believed to drive the evolution of specialized biosynthetic pathways and adaptive metabolic systems. As a result, marine Streptomyces frequently produce structurally distinctive metabolites containing unusual halogenated moieties, sulfur-containing functional groups, rare amino acids, macrocyclic architectures, and highly modified polyketide scaffolds rarely encountered in terrestrial microorganisms [10,11]. Many marine-derived metabolites exhibit potent antibacterial, antifungal, antiviral, antiparasitic, anti-inflammatory, and anticancer activities, thereby attracting increasing attention for pharmaceutical development [2,10,11]. Compared with terrestrial actinomycetes, marine-derived Streptomyces are exposed to unique ecological pressures including high salinity, hydrostatic pressure, nutrient limitation, interspecies competition, and host-associated symbiosis, which collectively contribute to the evolution of distinct biosynthetic pathways and structurally diverse secondary metabolites.
The rapid expansion of next-generation sequencing technologies and genome mining approaches has fundamentally transformed marine natural product research [12,13]. Bioinformatic tools such as antiSMASH, PRISM, BAGEL, DeepBGC, and BiG-SCAPE have demonstrated that marine Streptomyces genomes harbor substantially greater biosynthetic potential than previously recognized [13,14,15,16,17]. Importantly, many biosynthetic gene clusters remain transcriptionally silent under standard laboratory conditions, leading to the concept of cryptic or silent metabolic pathways [14,15]. Consequently, innovative activation strategies including co-cultivation systems, OSMAC cultivation approaches, ribosome engineering, CRISPR-Cas-mediated activation, epigenetic modulation, and heterologous expression platforms have become central strategies for unlocking previously inaccessible metabolites [14,15].
Simultaneously, integrated metabolomics and molecular networking technologies have accelerated dereplication and enabled rapid prioritization of chemically novel compounds [16]. Artificial intelligence-assisted metabolite prediction and machine learning-based biosynthetic analysis are further transforming natural product discovery into a highly data-driven discipline [17]. Nevertheless, despite major technological advances, substantial conceptual and translational limitations remain unresolved. Many studies continue to emphasize metabolite discovery without sufficiently addressing ecological function, biosynthetic regulation, pharmacological feasibility, or industrial scalability. Therefore, future progress in marine Streptomyces research will require stronger integration of genomics, metabolomics, systems biology, synthetic biology, and translational pharmacology.
Despite rapid advances in marine natural product discovery, several critical limitations remain unresolved, including low cultivation efficiency, silent biosynthetic gene cluster expression, frequent rediscovery of known metabolites, unstable metabolite production, and insufficient ecological interpretation of biosynthetic functions. Moreover, many recent studies remain heavily dependent on predictive omics and computational genome mining without adequate functional validation of biosynthetic pathways and metabolite bioactivities. Although numerous reviews have summarized marine-derived natural products, a comprehensive synthesis integrating ecological adaptation, biosynthetic logic, genome-guided discovery, enzymatic diversification, synthetic biology, and translational pharmaceutical perspectives specifically in marine-derived Streptomyces remains limited. This review therefore aims to critically evaluate the biosynthetic diversity and translational potential of marine Streptomyces-derived metabolites in the context of modern omics-driven drug discovery.
2. Methodology
This review was conducted using a narrative-critical review approach focusing on the biosynthetic diversity, enzymatic mechanisms, genomic potential, metabolomics-guided discovery, and pharmaceutical applications of marine-derived Streptomyces. Relevant literature was systematically collected from PubMed, Scopus, Web of Science, ScienceDirect, and Google Scholar databases. The literature search primarily covered publications from 1990 to 2026 using combinations of keywords including “marine Streptomyces”, “marine actinomycetes”, “biosynthetic gene clusters”, “marine natural products”, “genome mining”, “polyketides”, “non-ribosomal peptides”, “RiPPs”, “metabolomics”, “secondary metabolites”, “synthetic biology”, and “drug discovery”.
Only peer-reviewed articles published in English were included. Studies focusing exclusively on terrestrial actinomycetes without direct relevance to marine biosynthesis were excluded unless they provided essential comparative biosynthetic insights. Emphasis was placed on studies involving experimentally validated biosynthetic pathways, metabolite characterization, enzymatic tailoring reactions, genome-guided discovery strategies, synthetic biology approaches, and translational pharmaceutical applications.
The selected literature was critically analyzed to evaluate not only technological advances but also current limitations and conceptual gaps within the field. Particular attention was given to challenges involving silent biosynthetic gene clusters, metabolite rediscovery, annotation overinterpretation, low cultivation efficiency, dereplication bottlenecks, scalability limitations, and insufficient ecological interpretation. References were organized according to Vancouver style and aligned sequentially with their first citation in the text.
Unlike traditional narrative surveys focused predominantly on compound cataloging, this review emphasizes critical evaluation of current methodological limitations, translational barriers, and conceptual gaps within marine Streptomyces research. Particular attention is given to discrepancies between computational biosynthetic prediction and experimental validation, as well as the emerging need for integrative and functionally validated biosynthetic research frameworks.
3. Marine Streptomyces Diversity and Ecological Distribution
Marine-derived Streptomyces have been isolated from highly diverse ecological niches including marine sediments, deep-sea environments, hydrothermal vents, mangrove ecosystems, marine sponges, algae, seawater, and saline habitats [7,8,9]. These ecosystems represent chemically dynamic and highly competitive environments that appear to stimulate the evolution of specialized secondary metabolism. Marine sediments remain the most extensively investigated source of marine Streptomyces. Sediment-associated strains frequently produce structurally diverse polyketides, alkaloids, peptides, and hybrid metabolites exhibiting potent antibacterial and anticancer activities.
Deep-sea Streptomyces isolated from abyssal sediments and hydrothermal vents possess particularly distinctive biosynthetic capabilities due to adaptation to oligotrophic conditions, elevated hydrostatic pressure, and limited nutrient availability [18,19,20]. Comparative genomic studies suggest that these environmental pressures contribute substantially to biosynthetic diversification and may promote the accumulation of cryptic biosynthetic pathways [19].
Marine sponge-associated Streptomyces constitute another important ecological group because marine sponges harbor dense microbial consortia involved in chemical defense and symbiotic interactions. Numerous sponge-associated actinomycetes produce cytotoxic metabolites, quorum sensing inhibitors, antibiofilm compounds, and antimicrobial agents believed to contribute to host defense mechanisms. Similarly, mangrove-derived Streptomyces have attracted considerable interest because mangrove ecosystems represent highly fluctuating interfaces between terrestrial and marine environments characterized by changing salinity, oxygen gradients, and redox conditions [8,9]. Such environmental variability appears to promote remarkable metabolic flexibility and chemical diversity.
Comparative genomic analyses indicate that marine Streptomyces often harbor larger numbers of biosynthetic gene clusters than many terrestrial relatives [12,19]. Horizontal gene transfer, ecological adaptation, selective environmental pressures, and interspecies competition are believed to contribute significantly to this biosynthetic expansion. However, despite growing ecological and genomic evidence supporting the uniqueness of marine Streptomyces, many studies remain descriptive and insufficiently address the ecological roles of secondary metabolites in natural marine ecosystems.
4. Biosynthetic Gene Clusters in Marine Streptomyces
Secondary metabolite biosynthesis in marine Streptomyces is primarily encoded by biosynthetic gene clusters comprising structural biosynthetic genes, regulatory proteins, resistance determinants, transport systems, and diverse tailoring enzymes [21,22]. These clusters collectively orchestrate highly coordinated biosynthetic pathways responsible for the production of structurally complex secondary metabolites.
The most abundant biosynthetic systems identified in marine Streptomyces include type I and type II polyketide synthases (PKSs), non-ribosomal peptide synthetases (NRPSs), PKS-NRPS hybrid systems, ribosomally synthesized and post-translationally modified peptide (RiPP) pathways, terpene biosynthetic clusters, and alkaloid-associated pathways [12,13,14,15,16,17]. These systems collectively contribute to the remarkable chemical diversity observed among marine actinomycetes.
Polyketide synthases represent one of the largest biosynthetic systems in marine microorganisms because they generate structurally diverse metabolites through modular enzymatic assembly lines [23,24]. Type I PKS systems catalyze successive Claisen condensation reactions using malonyl-CoA-derived extender units, resulting in highly complex macrocyclic and polyene-containing metabolites. Marine-derived polyketides such as marinomycins and abyssomicins exhibit highly complex structures and potent biological activities [25,26]. Structural diversification is further amplified by extensive tailoring reactions including methylation, hydroxylation, glycosylation, oxidation, and halogenation.
Non-ribosomal peptide synthetases synthesize peptides independently of the ribosome through thiotemplate biosynthesis involving adenylation, thiolation, and condensation domains [23,24]. Marine NRPS-derived metabolites frequently contain unusual amino acids, D-amino acids, heterocyclic residues, and N-methylated structures that contribute substantially to biological activity and pharmacological specificity. Cyclomarins and related marine peptides have demonstrated potent antitubercular and anticancer properties [27].
Hybrid PKS-NRPS systems are particularly abundant in marine actinomycetes and generate structurally sophisticated metabolites integrating both polyketide and peptide biosynthetic features [24,27]. These hybrid metabolites often display potent anticancer, antibacterial, and antiparasitic properties because of their highly complex stereochemical architectures.
RiPP biosynthetic pathways constitute another important class of marine biosynthetic systems. These pathways encode precursor peptides that subsequently undergo extensive post-translational modifications to generate mature bioactive metabolites [16,17]. Marine RiPPs including thiopeptides and lantipeptides frequently exhibit potent antibacterial activities against Gram-positive pathogens.
Despite major advances in biosynthetic annotation, a substantial disconnect remains between computational biosynthetic prediction and experimental validation. Many predicted clusters remain functionally uncharacterized, and annotation accuracy is frequently limited by incomplete understanding of marine biosynthetic enzymology. Consequently, excessive reliance on predictive bioinformatics without biochemical confirmation may contribute to annotation overinterpretation and inaccurate functional assignment.
The remarkable chemical diversity observed in marine Streptomyces is associated with several major classes of biosynthetic systems, each contributing distinct structural scaffolds and pharmacological activities. Representative biosynthetic classes, characteristic metabolites, and their major biological functions are summarized in Table 1.
5. Genome Mining Approaches
The emergence of affordable next-generation sequencing technologies has dramatically transformed marine natural product discovery by enabling systematic exploration of biosynthetic gene clusters at the genomic level [12,13]. Genome mining refers to the computational identification and prediction of secondary metabolite pathways based on conserved enzymatic motifs, domain organization, regulatory signatures, and comparative genomic architectures. This approach has become particularly important in marine Streptomyces research because conventional cultivation-based screening frequently fails to activate the majority of biosynthetic pathways encoded within microbial genomes.
Several bioinformatic platforms have become central to marine biosynthetic discovery. antiSMASH is currently one of the most widely utilized tools for automated identification and annotation of biosynthetic gene clusters because it enables rapid recognition of PKS, NRPS, RiPP, terpene, and hybrid biosynthetic systems [13]. PRISM further enables predictive structural analysis of secondary metabolites based on biosynthetic domain organization and substrate specificity [12]. BAGEL specializes in the identification of RiPP-associated pathways and bacteriocin biosynthesis [16], whereas DeepBGC utilizes machine learning-assisted algorithms for improved biosynthetic cluster recognition [17]. BiG-SCAPE enables comparative clustering and evolutionary analysis of biosynthetic pathways across large genomic datasets, thereby facilitating biosynthetic family classification and diversity analysis [17].
Genome mining studies have demonstrated that many marine Streptomyces strains contain between 20 and 50 biosynthetic gene clusters, although only a minority are actively expressed under standard laboratory conditions [12,13,14,15]. This discrepancy between biosynthetic potential and observable metabolite production has become one of the central conceptual challenges in marine natural product research. Consequently, activation of silent or cryptic biosynthetic pathways has become a major research priority.
To overcome transcriptional silencing, several activation strategies have been developed including OSMAC cultivation systems, co-cultivation approaches, ribosome engineering, epigenetic modulation, CRISPR-Cas-mediated activation, and heterologous expression platforms [14,15]. OSMAC strategies manipulate environmental parameters such as salinity, nutrient composition, pH, aeration, trace elements, and temperature in order to induce alternative metabolic states. Co-cultivation systems exploit microbial interactions and chemical signaling mechanisms to stimulate cryptic biosynthesis that may not occur in monoculture conditions. Ribosome engineering introduces mutations affecting translational regulation and secondary metabolism, whereas CRISPR-based approaches allow direct transcriptional activation of silent gene clusters through targeted promoter engineering and regulatory rewiring [15].
Although these technologies have substantially expanded access to previously inaccessible metabolites, important translational limitations remain unresolved. Activated pathways frequently produce metabolites only at trace concentrations, metabolic instability complicates downstream purification, and heterologous hosts often fail to reproduce native regulatory conditions required for complete biosynthetic expression. Furthermore, computational prediction alone frequently overestimates actual metabolite production capacity because many predicted clusters remain nonfunctional or incompletely assembled under laboratory conditions. Consequently, future genome mining strategies will likely require stronger integration with systems biology, metabolomics, synthetic biology, and artificial intelligence-assisted pathway optimization to improve both biosynthetic prediction accuracy and experimental validation.
The rapid expansion of genome mining technologies has substantially accelerated biosynthetic pathway prediction and natural product prioritization in marine actinomycetes. Major bioinformatic platforms currently used in marine natural product discovery are summarized in Table 2.
6. Biosynthetic Enzymology and Tailoring Reactions
One of the defining characteristics of marine Streptomyces-derived natural products is the extraordinary diversity of enzymatic tailoring reactions that transform core biosynthetic scaffolds into structurally complex metabolites. Tailoring enzymes contribute substantially to chemical diversification, ecological adaptation, pharmacological specificity, and bioactivity enhancement. In many cases, the biological activity of marine metabolites depends not only on the core biosynthetic scaffold but also on extensive post-assembly enzymatic modifications that alter stereochemistry, polarity, membrane permeability, and target selectivity.
Halogenation represents one of the most distinctive biosynthetic modifications in marine microorganisms because marine environments contain high concentrations of chloride and bromide ions that facilitate enzymatic halogen incorporation [28,29,30,31]. Marine Streptomyces frequently encode flavin-dependent halogenases, non-heme iron halogenases, and vanadium-dependent haloperoxidases capable of catalyzing highly regioselective chlorination and bromination reactions [28,29,30,31]. Halogenated marine metabolites including marinopyrroles and napyradiomycins often exhibit enhanced membrane permeability, improved stability, and increased antibiotic potency [32,33]. The discovery of highly selective marine halogenases has therefore attracted substantial interest in biocatalysis and synthetic biology because these enzymes may provide environmentally sustainable alternatives to conventional chemical halogenation.
Glycosylation represents another major tailoring strategy in marine biosynthesis. Glycosyltransferases attach deoxysugars and modified carbohydrate residues to aglycone scaffolds, thereby altering solubility, pharmacokinetic behavior, target recognition, and toxicity profiles [34,35]. Marine metabolites such as lobophorins and kijanimicin analogs contain highly complex glycosylated structures generated through specialized sugar biosynthetic cassettes embedded within biosynthetic gene clusters [27,36]. Glycosylation diversity contributes substantially to pharmacological specificity and may significantly influence therapeutic efficacy.
Cytochrome P450 monooxygenases are also abundant in marine Streptomyces genomes and catalyze oxidation, hydroxylation, epoxidation, and oxidative cyclization reactions that substantially increase molecular complexity and stereochemical diversity [37,38]. Sequential oxidative tailoring frequently determines the final biological activity of marine metabolites and contributes significantly to scaffold diversification.
Macrocyclization further contributes to conformational rigidity and receptor specificity in marine natural products. Thioesterases, cyclases, and Diels–Alderases catalyze ring closure reactions generating macrocycles, polyethers, and spirotetronate architectures [39]. The discovery of enzymatic Diels–Alder reactions in marine biosynthesis has attracted considerable attention because these reactions generate highly complex polycyclic frameworks with exceptional stereoselectivity under biologically mild conditions.
Despite major advances in biosynthetic enzymology, many marine tailoring reactions remain poorly characterized at both structural and mechanistic levels. Consequently, a substantial gap persists between genomic annotation and functional enzymology. Many biosynthetic enzymes are predicted solely through sequence homology without direct biochemical confirmation, increasing the risk of functional overinterpretation. Future research will therefore require stronger integration of structural biology, enzymology, metabolomics, and synthetic biology to achieve experimentally validated understanding of marine biosynthetic systems.
7. Polyketide Diversity in Marine Streptomyces
Polyketides constitute one of the most chemically diverse classes of marine microbial natural products and are biosynthesized through successive decarboxylative Claisen condensation reactions catalyzed by polyketide synthases (PKSs) [40,41]. Marine-derived polyketides frequently display exceptional structural complexity characterized by highly unsaturated macrocycles, polyenes, aromatic frameworks, spirotetronates, and macrodiolide architectures. The remarkable diversity of marine polyketides reflects both biosynthetic modularity and adaptation to chemically competitive marine ecosystems.
Type I PKSs are large multifunctional modular enzymes in which each module catalyzes one elongation cycle during chain assembly [40]. These assembly-line biosynthetic systems allow extraordinary structural flexibility through variation in ketosynthase, acyltransferase, ketoreductase, dehydratase, and enoylreductase domains. Marine-derived type I polyketides frequently possess extensive oxidation patterns and unusual stereochemical complexity rarely observed in terrestrial metabolites.
Representative marine polyketides include saliniketals, marinomycins, arenicolides, abyssomicins, and lobophorins [41,42]. Marinomycins exhibit potent anticancer activity against melanoma cells and possess highly unsaturated macrodiolide structures generated through complex PKS-mediated biosynthesis [42]. Abyssomicin C inhibits para-aminobenzoic acid biosynthesis and demonstrates potent antibacterial activity against Gram-positive pathogens. Lobophorins represent glycosylated spirotetronate antibiotics with antibacterial and cytotoxic activities mediated by highly modified macrocyclic scaffolds.
Type II PKSs differ substantially from modular type I systems because they utilize dissociated multienzyme complexes that repeatedly catalyze iterative chain elongation reactions [40,41]. These systems primarily generate aromatic polyketides including anthracyclines, angucyclines, tetracenomycins, and related aromatic metabolites. Many marine aromatic polyketides exhibit antibacterial, anticancer, kinase inhibitory, and anti-inflammatory activities. Structural diversification is further amplified through oxidative tailoring, glycosylation, halogenation, and cyclization reactions.
Spirotetronates represent another particularly important class of marine actinomycete polyketides characterized by tetronic acid-containing macrocycles with highly constrained stereochemical architectures. Representative marine spirotetronates include abyssomicins, chlorothricins, and lobophorins [41,42]. These metabolites frequently display potent antibacterial or cytotoxic properties because their rigid macrocyclic conformations enable highly selective molecular interactions with biological targets.
The remarkable structural diversity of marine polyketides is strongly associated with extensive tailoring reactions occurring after core scaffold assembly. Glycosylation, methylation, oxidation, epoxidation, and halogenation collectively generate highly modified metabolites with enhanced pharmacological specificity. Marine environments appear to favor particularly extensive halogenation patterns because of elevated chloride and bromide availability in seawater [28,29,30,31].
Despite substantial chemical diversity and pharmacological potential, translation of marine polyketides into clinically applicable therapeutics remains limited by several major challenges. Many metabolites are produced only at extremely low concentrations under laboratory cultivation conditions, and biosynthetic instability frequently complicates metabolite purification and scale-up production. Furthermore, numerous marine polyketides possess highly complex stereochemistry that complicates total synthesis and medicinal chemistry optimization. Consequently, future development of marine polyketide therapeutics will require improved biosynthetic engineering, scalable fermentation strategies, and integrated synthetic biology approaches.
The major classes of marine-derived polyketides and their biological activities are summarized in Table 3.
8. Non-Ribosomal Peptides and Lipopeptides
Non-ribosomal peptides (NRPs) are synthesized by large modular non-ribosomal peptide synthetase (NRPS) megaenzymes through thiotemplate mechanisms independent of the ribosome [40]. Each NRPS module typically contains adenylation, thiolation, and condensation domains responsible for amino acid activation, carrier attachment, and peptide bond formation. Unlike ribosomal biosynthesis, NRPS systems are capable of incorporating highly unusual amino acids, hydroxy acids, D-amino acids, heterocyclic residues, and N-methylated building blocks, thereby generating exceptional structural diversity and pharmacological complexity.
Marine Streptomyces produce structurally distinctive non-ribosomal peptides enriched with rare amino acid residues and highly modified peptide architectures. These metabolites frequently exhibit potent antibacterial, antifungal, anticancer, antiparasitic, and anti-inflammatory activities because their conformational rigidity and chemical diversity enable highly selective interactions with biological targets.
Cyclomarins represent cyclic heptapeptides isolated from marine actinomycetes exhibiting anti-inflammatory and antitubercular activities [36,42]. Cyclomarin A specifically targets the ClpC1 ATPase of Mycobacterium tuberculosis, highlighting its significance as a promising lead compound for tuberculosis drug discovery. The unique binding mechanism of cyclomarin A has attracted considerable pharmaceutical interest because it differs from conventional antitubercular agents and may therefore reduce cross-resistance development.
Ohmyungsamycins are cyclic depsipeptides displaying potent antibacterial and anticancer properties mediated through disruption of cellular signaling and membrane-associated processes. Marine lipopeptides frequently exhibit membrane-disrupting activity because hydrophobic lipid tails facilitate membrane insertion and target selectivity. Such amphiphilic properties often enhance antibacterial potency against multidrug-resistant pathogens.
Marine NRPS-derived metabolites also display substantial structural diversity through hybridization with PKS systems. PKS-NRPS hybrid pathways integrate peptide and polyketide biosynthetic modules, generating highly sophisticated metabolites with exceptional stereochemical complexity [40,41]. These hybrid compounds frequently demonstrate potent cytotoxic and antiparasitic activities.
Despite remarkable pharmacological potential, translational development of marine non-ribosomal peptides remains constrained by several major limitations. Many compounds exhibit poor stability, limited oral bioavailability, high production costs, and complex biosynthetic regulation. Furthermore, large-scale industrial production remains difficult because marine NRPS pathways are frequently transcriptionally silent or poorly expressed under laboratory conditions. Consequently, future progress will likely require stronger integration of synthetic biology, metabolic engineering, and heterologous expression systems to improve metabolite accessibility and scalability.
The major classes of marine non-ribosomal peptides and lipopeptides are summarized in Table 4.
9. Ribosomally Synthesized and Post-Translationally Modified Peptides (RiPPs)
Ribosomally synthesized and post-translationally modified peptides (RiPPs) represent an increasingly important class of marine microbial natural products characterized by extensive enzymatic modification of genetically encoded precursor peptides [16,17]. Unlike NRPS-derived metabolites, RiPP biosynthesis begins with ribosomal peptide synthesis followed by highly specialized post-translational tailoring reactions including cyclization, dehydration, methylation, epimerization, heterocyclization, and proteolytic processing. These modifications collectively generate structurally complex metabolites with remarkable biological activities.
Marine Streptomyces genomes contain numerous cryptic RiPP biosynthetic gene clusters encoding lantipeptides, lasso peptides, thiopeptides, cyanobactin-like compounds, and other peptide-derived metabolites [16,17]. Genome mining studies have demonstrated that RiPP biosynthetic capacity in marine actinomycetes is substantially greater than previously recognized, suggesting that marine ecosystems remain an underexplored reservoir of peptide diversity.
One major advantage of RiPP biosynthesis is its genetic simplicity relative to PKS and NRPS systems. Because precursor peptides are directly encoded by structural genes, RiPP pathways are often more amenable to genetic engineering, combinatorial biosynthesis, and synthetic biology-based diversification. This modularity has attracted increasing interest for rational peptide engineering and next-generation antimicrobial development.
Thiopeptides isolated from marine actinomycetes exhibit exceptionally potent antibacterial activity against Gram-positive pathogens and have emerged as promising candidates for antimicrobial drug discovery [16]. These sulfur-rich macrocyclic peptides frequently target ribosomal machinery or essential translational processes, thereby providing highly selective antibacterial activity. Similarly, lantipeptides possess unusual lanthionine bridges generated through post-translational dehydration and cyclization reactions, contributing to enhanced structural rigidity and proteolytic stability.
Lasso peptides represent another distinctive RiPP class characterized by threaded peptide topologies resembling molecular knots. Their conformational stability frequently confers resistance to thermal degradation and proteolysis, making them particularly attractive for pharmaceutical applications. Marine-derived RiPPs may therefore represent valuable scaffolds for development of peptide-based therapeutics with improved stability and target specificity.
Despite increasing genomic evidence for RiPP diversity in marine microorganisms, many predicted RiPP pathways remain experimentally uncharacterized [16,17]. In numerous cases, computational prediction has substantially outpaced biochemical validation. Consequently, a major conceptual gap remains between genomic biosynthetic prediction and experimentally verified metabolite production. Furthermore, low expression levels, cryptic regulation, and insufficient cultivation strategies continue to limit access to many marine RiPP metabolites.
Another important limitation involves annotation accuracy. Automated genome mining algorithms frequently predict RiPP clusters based solely on conserved motifs and precursor peptide signatures, which may result in false-positive predictions or incomplete pathway annotation. Therefore, future progress in marine RiPP research will require stronger integration of comparative genomics, structural biology, metabolomics, and functional enzymology to achieve experimentally validated understanding of marine peptide biosynthesis.
Major classes of marine RiPPs and their biosynthetic features are summarized in Table 5.
10. Terpenoids and Isoprenoid Metabolites
Although marine actinomycete research has historically focused predominantly on polyketides and non-ribosomal peptides, terpenoid biosynthesis is increasingly recognized as an important metabolic capability in marine Streptomyces [18,20]. Terpenoids constitute one of the largest classes of natural products in nature and are biosynthesized through condensation of isoprene-derived precursors generated via either the mevalonate pathway or the methylerythritol phosphate (MEP) pathway.
Marine-derived terpenoids frequently exhibit cytotoxic, antimicrobial, antioxidant, antiviral, and anti-inflammatory activities. In contrast to many terrestrial terpenoids, marine actinomycete-derived terpenoids often possess halogenated substituents, rearranged carbon skeletons, or highly oxidized frameworks reflecting adaptation to chemically unique marine environments [28,29,30,31]. These structural features contribute significantly to biological activity and pharmacological specificity.
Marine Streptomyces genomes encode diverse terpene cyclases and prenyltransferases capable of generating structurally complex mono-, sesqui-, di-, and triterpenoid metabolites. Biosynthetic diversification is further enhanced through oxidative tailoring reactions mediated by cytochrome P450 monooxygenases and related oxidative enzymes [37,38]. Such enzymatic modifications generate highly modified terpenoid scaffolds rarely observed in terrestrial microorganisms.
Several marine terpenoids possess unusual carbocyclic frameworks resulting from extensive rearrangement reactions during biosynthesis. These structural rearrangements frequently produce metabolites with enhanced molecular rigidity and receptor-binding specificity. Additionally, marine terpenoids may contribute ecologically to chemical defense, microbial competition, signaling interactions, and environmental adaptation within marine microbial communities.
Despite growing recognition of marine terpenoid diversity, this biosynthetic field remains comparatively underexplored relative to PKS- and NRPS-derived metabolites. One major reason is that terpene biosynthetic pathways are frequently smaller and more difficult to identify computationally using conventional genome mining algorithms. Furthermore, many marine terpenoids are produced at low concentrations, complicating metabolite isolation and structural characterization.
Another important limitation involves insufficient functional characterization of marine terpene cyclases and tailoring enzymes. Many predicted terpene biosynthetic clusters remain experimentally unvalidated, and mechanistic understanding of marine terpene biosynthesis remains incomplete. Consequently, future research integrating genome mining, structural biology, enzymology, and metabolomics will be essential for expanding understanding of marine terpenoid biosynthesis and pharmaceutical potential.
The major classes of marine terpenoids and their biosynthetic characteristics are summarized in Table 6.
11. Metabolomics and Dereplication Strategies
Modern marine natural product discovery increasingly relies on metabolomics-guided workflows integrating LC-MS/MS, UHPLC-QTOF, NMR metabolomics, imaging mass spectrometry, and molecular networking approaches [16,17]. These technologies have fundamentally transformed marine biosynthetic research by enabling high-throughput chemical profiling and rapid prioritization of structurally novel metabolites from highly complex microbial extracts.
Dereplication refers to the rapid identification of previously known compounds during early stages of natural product screening in order to minimize repetitive rediscovery of structurally characterized metabolites [16]. Historically, marine natural product discovery suffered from substantial inefficiency because many studies repeatedly isolated already known antibiotics or cytotoxic compounds. Metabolomics-guided dereplication strategies now allow researchers to identify known metabolites at early analytical stages, thereby improving discovery efficiency and prioritization of chemically unique compounds.
GNPS molecular networking has emerged as one of the most valuable platforms in contemporary natural product metabolomics because it enables clustering of structurally related metabolites based on MS/MS fragmentation similarity [16]. This approach facilitates visualization of chemical families and allows rapid identification of analog series, biosynthetic relationships, and potentially novel metabolites within complex datasets. Molecular networking has proven particularly valuable for marine Streptomyces extracts because these organisms frequently produce diverse families of structurally related secondary metabolites.
Untargeted metabolomics further enables comparative analysis of metabolic responses under different cultivation conditions including OSMAC systems, co-cultivation strategies, salinity variation, and nutrient modulation. Such approaches can reveal cryptic metabolites that remain undetected under conventional cultivation conditions. Imaging mass spectrometry additionally enables spatial visualization of metabolite distribution during microbial interactions, thereby providing insight into ecological functions of marine secondary metabolites.
NMR-based metabolomics also contributes significantly to marine natural product discovery because it enables structural characterization of metabolites without complete purification. Combined LC-MS/MS and NMR approaches therefore provide complementary analytical platforms for structural elucidation and biosynthetic interpretation.
Despite substantial technological advances, metabolomics-guided discovery remains constrained by several important limitations. Spectral databases remain incomplete for many marine natural products, resulting in substantial annotation uncertainty [16,17]. Low-abundance metabolites frequently escape detection, while ion suppression effects and extraction biases may distort metabolic profiles. Furthermore, excessive dependence on automated annotation pipelines may increase the risk of metabolite misidentification and biosynthetic overinterpretation.
Another major limitation involves the growing disparity between data generation capacity and experimental validation. Modern metabolomics platforms can generate enormous datasets containing thousands of spectral features, yet only a small proportion are structurally characterized or biologically validated. Consequently, future marine metabolomics research will require improved spectral databases, artificial intelligence-assisted annotation systems, integrated multi-omics platforms, and experimentally validated metabolite characterization pipelines.
Although genome mining, metabolomics, and artificial intelligence-assisted prediction platforms have dramatically accelerated biosynthetic gene cluster identification, the field remains disproportionately biased toward computational prediction rather than experimental validation. Future studies should increasingly integrate transcriptomics, heterologous expression, gene knockout strategies, metabolite isolation, and ecological functional assays to validate predicted biosynthetic pathways and biological activities.
marin
The major metabolomics technologies currently applied in marine natural product discovery are summarized in Table 7.
12. Synthetic Biology and Drug Development
Synthetic biology has emerged as a transformative strategy for engineering marine natural product biosynthesis and overcoming many limitations associated with conventional cultivation-based metabolite discovery [14,15]. Advances in genome engineering, pathway refactoring, heterologous expression, chassis optimization, and CRISPR-Cas technologies now enable targeted manipulation of marine biosynthetic pathways for enhanced metabolite production and diversification.
One of the major applications of synthetic biology in marine actinomycete research involves activation of silent or cryptic biosynthetic gene clusters [14,15]. Many marine Streptomyces genomes contain numerous biosynthetic pathways that remain transcriptionally inactive under standard laboratory conditions. Synthetic biology approaches allow targeted activation of these pathways through promoter engineering, transcriptional rewiring, regulatory manipulation, and heterologous pathway reconstruction.
Engineered hosts such as Streptomyces coelicolor and Streptomyces albus are widely utilized as heterologous expression platforms because they possess well-characterized genetic systems and relatively low endogenous metabolite backgrounds [4,15]. Heterologous expression enables production of marine metabolites in genetically tractable hosts capable of improved fermentation performance and biosynthetic accessibility.
CRISPR-Cas systems have further revolutionized marine biosynthetic engineering by enabling precise genome editing and transcriptional regulation [15]. CRISPR-based activation systems can directly stimulate silent biosynthetic clusters, whereas multiplex editing approaches enable simultaneous manipulation of multiple biosynthetic pathways. Such technologies provide unprecedented opportunities for combinatorial biosynthesis and generation of structurally diversified metabolite libraries.
Synthetic biology also enables pathway refactoring through modular reconstruction of biosynthetic systems independent of native regulatory constraints. Refactored pathways can be optimized for improved precursor supply, enhanced metabolic flux, and increased production yield. Additionally, artificial intelligence-assisted pathway prediction and machine learning-guided metabolite prioritization are increasingly integrated into synthetic biology workflows to accelerate rational natural product engineering.
Marine Streptomyces-derived metabolites have demonstrated substantial pharmaceutical potential in oncology, infectious disease treatment, inflammation, neurodegeneration, and metabolic disorders [1,11,42]. Salinosporamide A represents one of the most prominent examples of successful marine-derived drug development because it advanced into clinical development as a proteasome inhibitor for cancer therapy [1,11]. This compound illustrates the enormous translational potential of marine actinomycete-derived metabolites when supported by integrated biosynthetic and pharmacological development strategies.
Nevertheless, technological capability alone does not guarantee successful pharmaceutical translation. Many promising marine metabolites fail during preclinical or clinical development because of toxicity, instability, poor pharmacokinetic behavior, limited bioavailability, or manufacturing constraints. Furthermore, large-scale industrial production frequently remains economically challenging because marine biosynthetic systems are often metabolically unstable or poorly expressed.
Another important limitation involves excessive emphasis on chemical novelty without sufficient consideration of translational feasibility. Many structurally complex marine metabolites are difficult to synthesize, optimize, or formulate as clinically viable therapeutics. Consequently, future marine drug discovery efforts must prioritize not only structural diversity but also pharmacological tractability, scalability, and therapeutic applicability.
The major synthetic biology strategies currently applied in marine natural product engineering are summarized in Table 8.
13. Challenges and Future Perspectives
Despite substantial technological progress, marine natural product discovery continues to face several major scientific, methodological, and translational limitations that collectively restrict efficient pharmaceutical development [12,13,14,15,16,17]. Although genome mining, metabolomics, synthetic biology, and artificial intelligence-assisted discovery platforms have dramatically accelerated biosynthetic exploration, many fundamental bottlenecks remain unresolved.
One of the most persistent limitations involves the low cultivation efficiency of marine microorganisms. A large proportion of marine microbial diversity remains uncultivable under conventional laboratory conditions because native marine ecosystems possess highly specific physicochemical and ecological parameters that are difficult to reproduce experimentally [18,19,20]. Consequently, many biosynthetic pathways encoded within marine microbial genomes remain inaccessible for experimental characterization. Even cultivable strains frequently display unstable metabolite production due to environmental sensitivity, nutritional dependence, or complex microbial interactions.
Silent biosynthetic gene clusters represent another major challenge in marine actinomycete research [14,15]. Genome sequencing studies have consistently demonstrated that marine Streptomyces harbor significantly greater biosynthetic capacity than observable metabolite production suggests. However, activation of cryptic pathways remains highly unpredictable, and many activation strategies produce only trace quantities of metabolites insufficient for structural elucidation or pharmacological testing. Furthermore, heterologous expression systems often fail to reproduce native regulatory networks required for complete pathway functionality.
Rediscovery of structurally known metabolites also remains a substantial obstacle despite major advances in dereplication technologies [16]. Marine microorganisms frequently produce common biosynthetic scaffolds shared across multiple taxa, thereby reducing discovery efficiency. Although metabolomics-guided prioritization and GNPS molecular networking have improved dereplication workflows, repetitive isolation of previously characterized compounds continues to consume considerable research resources.
Another important challenge involves annotation overinterpretation arising from excessive dependence on computational prediction [12,17]. Artificial intelligence-assisted genome mining and machine learning-based biosynthetic analysis have greatly expanded biosynthetic prediction capacity, but computational annotations frequently lack biochemical validation. Many predicted biosynthetic clusters are assigned hypothetical functions solely through sequence homology without experimental confirmation. Consequently, the field increasingly faces risks associated with annotation inflation, metabolite misidentification, and overestimation of biosynthetic novelty.
Industrial scalability also remains a major translational limitation. Numerous marine metabolites exhibit highly promising biological activities but cannot be efficiently produced at industrial scale because of low biosynthetic yield, metabolic instability, or highly complex structural features. In addition, many marine-derived compounds possess poor pharmacokinetic profiles, limited aqueous solubility, or unacceptable toxicity, thereby restricting clinical applicability despite impressive in vitro bioactivity.
Another conceptual limitation involves insufficient ecological interpretation of marine secondary metabolism. Many current studies emphasize metabolite identification and biosynthetic prediction without adequately investigating ecological function, microbial interactions, evolutionary significance, or environmental regulation. Secondary metabolites likely play important ecological roles involving competition, signaling, symbiosis, defense, and environmental adaptation within marine microbial communities. However, these ecological dimensions remain comparatively underexplored relative to purely chemical characterization.
Future progress in marine Streptomyces research will likely depend on stronger integration of long-read sequencing, single-cell genomics, spatial metabolomics, systems biology, artificial intelligence-assisted genome mining, synthetic ecology, automated biofoundry systems, and high-throughput functional screening [12,13,14,15,16,17]. Long-read sequencing technologies may substantially improve biosynthetic gene cluster assembly and structural accuracy, whereas single-cell genomics may provide access to previously uncultivable microbial populations.
Spatial metabolomics and imaging mass spectrometry may further improve understanding of ecological interactions and metabolite localization within marine microbial communities. Simultaneously, synthetic ecology approaches involving controlled multispecies cultivation systems may better reproduce natural microbial signaling interactions responsible for cryptic metabolite activation.
Artificial intelligence-assisted biosynthetic prediction will likely continue to accelerate marine natural product discovery, but future research must balance computational prediction with rigorous experimental validation. Functional genomics, structural enzymology, metabolomics, and translational pharmacology should therefore become more tightly integrated to ensure biologically meaningful interpretation of biosynthetic datasets.
Importantly, the field must gradually transition beyond descriptive omics-driven exploration toward experimentally validated and translationally relevant biosynthetic biology. Future marine natural product research should prioritize not only chemical novelty but also pharmacological feasibility, biosynthetic scalability, ecological relevance, and therapeutic applicability. Such an integrated strategy will likely define the next generation of marine-derived drug discovery and biosynthetic innovation.
A major challenge in marine Streptomyces research is that many biosynthetic gene clusters remain transcriptionally silent under conventional laboratory cultivation conditions. In natural marine environments, biosynthetic pathway activation is often tightly regulated by ecological interactions, nutrient limitation, osmotic stress, quorum sensing signals, and host-associated microbial communication. Consequently, standard monoculture conditions frequently fail to reproduce the environmental cues necessary for metabolite biosynthesis.
Major translational challenges and potential future solutions in marine Streptomyces drug discovery are summarized in Table 9.
14. Conclusion
Marine-derived Streptomyces represent one of the most prolific and biosynthetically versatile microbial resources for discovery of structurally diverse and pharmacologically important natural products. Adaptation to chemically dynamic marine environments has driven the evolution of highly specialized biosynthetic systems capable of generating diverse polyketides, non-ribosomal peptides, RiPPs, terpenoids, alkaloids, and hybrid metabolites with broad biological activities including antibacterial, antifungal, antiviral, anti-inflammatory, antiparasitic, and anticancer effects [1,2,3].
Recent advances in genome sequencing, genome mining, metabolomics, molecular networking, synthetic biology, and artificial intelligence-assisted discovery pipelines have substantially expanded understanding of marine biosynthetic diversity [12,13,14,15,16,17]. These technologies have revealed that marine Streptomyces genomes contain far greater biosynthetic capacity than previously recognized, including numerous cryptic biosynthetic gene clusters inaccessible through conventional cultivation approaches.
Simultaneously, progress in biosynthetic enzymology has highlighted the importance of tailoring reactions including halogenation, glycosylation, oxidative cyclization, and macrocyclization in generating structurally complex marine metabolites [28,29,30,31,32,33,34,35,36,37,38,39]. Such enzymatic diversification contributes significantly to pharmacological specificity and ecological adaptation.
Despite remarkable technological progress, major conceptual and translational limitations remain unresolved. Silent pathway expression, metabolite rediscovery, annotation overinterpretation, low cultivation efficiency, insufficient metabolite yield, and industrial scalability limitations continue to restrict efficient pharmaceutical development [12,13,14,15,16,17]. Furthermore, many studies remain predominantly descriptive and insufficiently address ecological function, biosynthetic regulation, or translational feasibility.
Future progress in marine natural product discovery will therefore require stronger integration of genomics, metabolomics, systems biology, synthetic biology, functional enzymology, ecological microbiology, and translational pharmacology. Importantly, the field must move beyond purely omics-driven descriptive exploration toward experimentally validated and clinically relevant biosynthetic research.
Marine Streptomyces remain a largely untapped reservoir of biosynthetic diversity with enormous pharmaceutical potential. Continued interdisciplinary integration will likely define the next generation of marine-derived therapeutic discovery and biosynthetic innovation.
The major strength of this review lies in its integrative and critical perspective that bridges marine microbial ecology, biosynthetic genomics, metabolomics, synthetic biology, and translational pharmacology within a single conceptual framework. Rather than functioning solely as a descriptive survey of marine-derived metabolites, this review highlights the urgent need to transition from omics-driven biosynthetic prediction toward experimentally validated and clinically relevant marine natural product research. Such an integrated strategy will likely define the next generation of marine-derived therapeutic discovery and biosynthetic innovation.
Author Contributions
Monthon Lertcanawanichakul (ML) led the conceptualization and overall design of the study, performed formal analysis, and contributed to investigation, data curation, and patent-related analysis. He was responsible for drafting the original manuscript as well as critical revision, editing, and supervision of the research work. Phuangthip Bhoopong (PB), Tuanhawanti Sahabuddeen (TS), Patchara Pedpradab (PD), Husna Madoromae (HS), Sueptrakool Wisessombat (SW), Attarat Pattanawongsa (AP), and Nuttapon Songnaka (NS) contributed to validation of results, including investigation, data collection, analysis, visualization, and validation, and participated in manuscript review and editing. All co-authors contributed to manuscript revision and approved the final version for submission.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Ethics Statement/Declarations
This study did not involve human participants or live vertebrate animals. All microbial isolates were obtained from commercially available or traditionally fermented Thai foods, and no endangered or protected species were used. Therefore, formal ethics approval was not required.
Clinical Trial
Not applicable.
Consent for Publication
Not applicable.
AI Usage Statement
During the preparation of this manuscript, AI-assisted tools were used for language editing, refinement of graphical elements, and development of preliminary conceptual figure layouts. All AI-generated outputs were subsequently reviewed, critically edited, and approved by the authors to ensure scientific accuracy, coherence, and consistency with the manuscript content. No AI tools were used for data fabrication, statistical analysis, interpretation of experimental results, or generation of references. The authors take full responsibility for the integrity and validity of the final manuscript.
Final Approval Statement
All authors have read and approved the final version of the manuscript and agree to be accountable for all aspects of the work.
Acknowledgments
The authors thank the Walailak University Science and Technology Park (WUSTP) for patent support, as well as the faculty and staff of the Clinical Microbiology Program and the School of Allied Health Sciences for laboratory and teaching support. The authors also acknowledge students and collaborators for their valuable contributions to discussions and experimental assistance.
Conflicts of Interest
The authors declare that they have no competing interests.
Abbreviations
| antiSMASH | antibiotics & Secondary Metabolite Analysis Shell |
| BAGEL | BActeriocin GEnome mining tooL |
| BGC | Biosynthetic Gene Cluster |
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| GNPS | Global Natural Products Social Molecular Networking |
| HPLC | High-Performance Liquid Chromatography |
| LC-MS/MS | Liquid Chromatography–Tandem Mass Spectrometry |
| MEP | Methylerythritol Phosphate pathway |
| MS/MS | Tandem Mass Spectrometry |
| NMR | Nuclear Magnetic Resonance |
| NRP(S) | Non-Ribosomal Peptide (Synthetase) |
| NRPS | Non-Ribosomal Peptide Synthetase |
| OSMAC | One Strain Many Compounds |
| PKS | Polyketide Synthase |
| PRISM | Prediction Informatics for Secondary Metabolomes |
| RiPPs | Ribosomally synthesized and Post-translationally modified Peptides |
| UHPLC-QTOF | Ultra-High Performance Liquid Chromatography–Quadrupole Time-of-Flight |
| WGS | Whole Genome Sequencing |
References
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2(12), 666–673. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2024, 41(1), 1–180. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; et al. Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80(1), 1–43. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.M.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417(6885), 141–147. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L.; Sanchez, S. Microbial drug discovery: 80 years of progress. J. Antibiot. 2009, 62(1), 5–16. [Google Scholar] [CrossRef]
- Berdy, J. Thoughts and facts about antibiotics: where we are now and where we are heading. J. Antibiot. 2012, 65(8), 385–395. [Google Scholar] [CrossRef]
- Jensen, P.R.; Dwight, R.; Fenical, W. Distribution of actinomycetes in near-shore tropical marine sediments. Appl. Env. Microbiol. 1991, 57(4), 1102–1108. [Google Scholar] [CrossRef]
- Zhang, X.; Li, S.J.; Li, J.J.; Liang, Z.Z.; Zhao, C.Q.; Cai, W.J. Marine microbial natural products: structure, biosynthesis, and ecology. Microbiol. Mol. Biol. Rev. 2021, 85(4), e00026-21. [Google Scholar] [CrossRef]
- Bull, A.T.; Stach, J.E. Marine actinobacteria: new opportunities for natural product search and discovery. Trends Microbiol. 2007, 15(11), 491–499. [Google Scholar] [CrossRef] [PubMed]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2011, 28(2), 290–344. [Google Scholar] [CrossRef]
- Hughes, C.C.; Fenical, W. Antibacterials from the sea. Chem. Eur. J. 2010, 16(42), 12512–12525. [Google Scholar] [CrossRef]
- Kautsar, S.A.; van der Hooft, J.J.J.; de Ridder, D.; Medema, M.H. BiG-SLiCE and the next generation of biosynthetic gene cluster mining. Curr. Opin. Microbiol. 2023, 69, 102199. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; et al. antiSMASH 5.0: updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47(W1), W81–W87. [Google Scholar] [CrossRef]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13(8), 509–523. [Google Scholar] [CrossRef]
- Zhao, H.; Medema, M.H. Synthetic biology approaches for natural product discovery. Nat. Rev. Microbiol. 2024. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; et al. Sharing and community curation of mass spectrometry data with GNPS. Nat. Biotechnol. 2016, 34(8), 828–837. [Google Scholar] [CrossRef] [PubMed]
- Kautsar, S.A.; Blin, K.; Shaw, S.; et al. BiG-SLiCE 2 and scaling biosynthetic diversity analysis. Nat. Chem. Biol. 2023, 19(12), 1520–1529. [Google Scholar] [CrossRef]
- Skropeta, D. Deep-sea natural products. Nat. Prod. Rep. 2008, 25(6), 1131–1166. [Google Scholar] [CrossRef]
- Jensen, P.R. Natural products and the gene cluster revolution. Trends Microbiol. 2016, 24(12), 968–977. [Google Scholar] [CrossRef]
- Sugimoto, Y.; Camacho, F.R.; Wang, S.; et al. A metagenomic strategy for harnessing uncultured microbes as sources of bioactive compounds. Nat. Rev. Microbiol. 2024. [Google Scholar] [CrossRef]
- Cane, D.E.; Walsh, C.T.; Khosla, C. Harnessing the biosynthetic code: combinations, permutations, and mutations. Science 1998, 282(5386), 63–68. [Google Scholar] [CrossRef]
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. Engl. 2009, 48(26), 4688–4716. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, M.; Wang, X.; Chen, C.; Deng, Z.; Zhang, Q. Marine actinomycetes-derived angucyclines and angucyclinones with biosynthesis and activity. Eur. J. Med. Chem. 2024, 271, 117161. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, W.; Hu, Z.; Wang, W.; Zhang, H. Marine Streptomyces-derived novel alkaloids. Mar. Drugs 2024, 22(1), 51. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.C.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Marinomycins A–D. J. Am. Chem. Soc. 2006, 128(5), 1622–1632. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.C.; Prieto-Davo, A.; Jensen, P.R.; Fenical, W. Marinopyrroles. Org. Lett. 2008, 10(4), 629–631. [Google Scholar] [CrossRef] [PubMed]
- Olano, C.; Méndez, C.; Salas, J.A. Antitumor compounds from marine actinomycetes. Mar. Drugs 2009, 7(2), 210–248. [Google Scholar] [CrossRef]
- Gribble, G.W. Naturally occurring organohalogen compounds. Acc. Chem. Res. 1998, 31(3), 141–152. [Google Scholar] [CrossRef]
- Winter, J.M.; Moore, B.S. Vanadium-dependent haloperoxidases. J. Biol. Chem. 2009, 284(28), 18577–18581. [Google Scholar] [CrossRef]
- Wagner, C.; El Omari, M.; König, G.M. Biohalogenation. J. Nat. Prod. 2009, 72(3), 540–553. [Google Scholar] [CrossRef]
- Neumann, C.S.; Fujimori, D.G.; Walsh, C.T. Halogenation strategies. Chem. Biol. 2008, 15(2), 99–109. [Google Scholar] [CrossRef]
- Shigeno, S.; Kadowaki, M.; Nagai, K.; Takahashi, Y.; Iwai, Y.; Omura, S. New polycyclic tetramate macrolactams. J. Antibiot. 2024, 77, 265–271. [Google Scholar] [CrossRef]
- Shiomi, K.; Iinuma, H.; Hamada, M.; Naganawa, H.; Manabe, M.; Matsuki, C.; et al. Napyradiomycins. J. Antibiot. 1986, 39(4), 487–493. [Google Scholar] [CrossRef]
- Thibodeaux, C.J.; Melançon, C.E., 3rd; Liu, H.W. Natural-product sugar biosynthesis. Nature 2007, 446(7139), 1008–1016. [Google Scholar] [CrossRef]
- Weymouth-Wilson, A.C. Carbohydrates in natural products. Nat. Prod. Rep. 1997, 14(2), 99–110. [Google Scholar] [CrossRef] [PubMed]
- Salas, J.A.; Méndez, C. Glycosylation engineering. Trends Microbiol. 2007, 15(5), 219–232. [Google Scholar] [CrossRef]
- Bernhardt, R. Cytochrome P450 biocatalysts. J. Biotechnol. 2006, 124(1), 128–145. [Google Scholar] [CrossRef] [PubMed]
- Podust, L.M.; Sherman, D.H. P450 diversity. Nat. Prod. Rep. 2012, 29(10), 1251–1266. [Google Scholar] [CrossRef]
- Kim, H.J.; Ruszczycky, M.W.; Choi, S.H.; Liu, Y.N.; Liu, H.W. Enzyme-catalysed [4+2] cycloaddition. Nature 2011, 473(7346), 109–112. [Google Scholar] [CrossRef]
- Cane, D.E.; Walsh, C.T.; Khosla, C. Biosynthetic code. Science 1998, 282(5386), 63–68. [Google Scholar] [CrossRef] [PubMed]
- Hertweck, C. Polyketide biosynthetic logic. Angew. Chem. Int. Ed. Engl. 2009, 48(26), 4688–4716. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Wang, J.; Chen, Q.; Zhao, P. Marine Streptomyces sp. PGC 39. Food Biosci. 2024, 57, 103600. [Google Scholar] [CrossRef]

Table 1.
Major Biosynthetic Classes Identified in Marine Streptomyces.
| Biosynthetic class | Representative metabolites | Major biological activities | Key biosynthetic features | References |
|---|---|---|---|---|
| Type I polyketides | Marinomycins, salinosporamides | Anticancer, antibacterial | Modular PKS assembly lines | [23,24,25,26] |
| Type II polyketides | Angucyclines, anthracyclines | Antibacterial, kinase inhibition | Aromatic PKS pathways | [23,24] |
| Non-ribosomal peptides | Cyclomarins, ohmyungsamycins | Antitubercular, anticancer | NRPS thiotemplate synthesis | [24,27] |
| PKS-NRPS hybrids | Lobophorins, abyssomicins | Cytotoxic, antibacterial | Hybrid modular pathways | [24,27] |
| RiPPs | Thiopeptides, lantipeptides | Gram-positive antibacterial | Post-translational modification | [16,17] |
| Terpenoids | Marine isoprenoids | Antioxidant, antimicrobial | MEP/mevalonate pathways | [18,19,20] |
Table 2.
Major Genome Mining and Bioinformatic Platforms Used in Marine Natural Product Discovery.
| Platform | Primary application | Major strengths | Current limitations | References |
| antiSMASH | Automated BGC annotation | Comprehensive detection | Limited novel enzyme prediction | [13] |
| PRISM | Metabolite structure prediction | Predictive biosynthetic modeling | Dependent on known pathways | [12] |
| BAGEL | RiPP identification | Specialized RiPP analysis | Limited non-RiPP analysis | [16] |
| DeepBGC | Machine learning-assisted prediction | Improved sensitivity | Training dataset dependency | [17] |
| BiG-SCAPE | Comparative BGC clustering | Evolutionary analysis | Functional validation required | [17] |
Table 3.
Representative Marine Streptomyces-Derived Polyketides and Their Biological Activities.
| Polyketide class | Representative metabolites | Major biological activities | Key structural features | References |
|---|---|---|---|---|
| Macrodiolides | Marinomycins | Anticancer, antibacterial | Highly unsaturated macrocycles | [42] |
| Spirotetronates | Abyssomicins, lobophorins | Antibacterial, cytotoxic | Tetronic acid-containing macrocycles | [41,42] |
| Aromatic polyketides | Anthracyclines, angucyclines | Anticancer, kinase inhibition | Aromatic ring systems | [40,41] |
| Glycosylated polyketides | Lobophorins | Antibacterial, cytotoxic | Deoxysugar-containing scaffolds | [34,35,36] |
Table 4.
Representative Marine Streptomyces-Derived Non-Ribosomal Peptides and Lipopeptides.
| Metabolite class | Representative compounds | Major biological activities | Key structural features | References |
| Cyclic peptides | Cyclomarins | Antitubercular, anti-inflammatory | Cyclic heptapeptides | [36,42] |
| Cyclic depsipeptides | Ohmyungsamycins | Antibacterial, anticancer | Ester-linked peptide macrocycles | [36] |
| Lipopeptides | Marine lipopeptides | Membrane-disrupting antibacterial activity | Lipid tail-containing amphiphiles | [40] |
| PKS-NRPS hybrids | Lobophorins | Cytotoxic, antibacterial | Hybrid peptide-polyketide scaffolds | [41] |
Table 5.
Major Classes of Marine RiPPs Identified in Marine Streptomyces.
| RiPP class | Representative features | Major biological activities | Key biosynthetic modifications | References |
| Thiopeptides | Sulfur-rich macrocycles | Potent antibacterial activity | Cyclodehydration and thiazole formation | [16,17] |
| Lantipeptides | Lanthionine-containing peptides | Antimicrobial activity | Dehydration and thioether bridge formation | [16] |
| Lasso peptides | Threaded peptide topology | Proteolytic stability, antibacterial activity | Macrolactam ring threading | [17] |
| Cyanobactin-like peptides | Cyclic modified peptides | Cytotoxic and antimicrobial activity | Cyclization and heterocyclization | [16,17] |
Table 6.
Major Classes of Marine Streptomyces-Derived Terpenoids.
| Terpenoid class | Major biological activities | Key biosynthetic features | Structural characteristics | References |
| Monoterpenoids | Antimicrobial activity | Isoprene condensation | Small volatile structures | [18,20] |
| Sesquiterpenoids | Cytotoxic, anti-inflammatory | Farnesyl pyrophosphate-derived | Rearranged carbon skeletons | [20] |
| Diterpenoids | Antioxidant, antimicrobial | Geranylgeranyl pyrophosphate-derived | Polycyclic frameworks | [18] |
| Halogenated terpenoids | Antibacterial, antiviral | Halogenase-associated pathways | Brominated/chlorinated scaffolds | [28,29,30,31] |
Table 7.
Major Metabolomics and Dereplication Technologies Used in Marine Natural Product Discovery.
Table 7.
Major Metabolomics and Dereplication Technologies Used in Marine Natural Product Discovery.
| Technology | Primary application | Major strengths | Current limitations | References |
| LC-MS/MS | Metabolite profiling | High sensitivity | Ion suppression effects | [16] |
| UHPLC-QTOF | High-resolution metabolomics | Accurate mass detection | Complex data interpretation | [16,17] |
| GNPS molecular networking | Metabolite clustering | Rapid dereplication | Database dependency | [16] |
| NMR metabolomics | Structural elucidation | Non-destructive analysis | Lower sensitivity | [17] |
| Imaging mass spectrometry | Spatial metabolite visualization | Ecological interaction analysis | Expensive instrumentation | [16,17] |
Table 8.
Synthetic Biology Approaches in Marine Streptomyces Drug Discovery.
| Synthetic biology strategy | Major application | Key advantages | Current limitations | References |
| CRISPR-Cas activation | Silent BGC activation | Precise genome regulation | Off-target effects | [15] |
| Heterologous expression | Metabolite production | Improved biosynthetic accessibility | Host compatibility issues | [14,15] |
| Pathway refactoring | Biosynthetic optimization | Regulatory independence | Complex engineering requirements | [15] |
| Chassis engineering | Industrial metabolite production | Enhanced scalability | Metabolic burden | [14] |
| AI-assisted pathway prediction | Metabolite prioritization | Accelerated discovery | Annotation overprediction | [17] |
Table 9.
Major Translational Challenges in Marine Streptomyces Drug Discovery.
| Challenge | Current impact | Potential future solutions | References |
| Silent biosynthetic gene clusters | Limited metabolite accessibility | CRISPR activation, synthetic biology | [14,15] |
| Low cultivation efficiency | Reduced strain recovery | Advanced cultivation systems | [18,19,20] |
| Rediscovery of known metabolites | Reduced novelty | Metabolomics-guided dereplication | [16] |
| Low metabolite yield | Limited scalability | Chassis engineering and pathway optimization | [14,15] |
| Annotation overinterpretation | Misleading biosynthetic prediction | Functional validation and enzymology | [12,17] |
| Industrial scalability limitations | Delayed pharmaceutical translation | Biofoundries and systems biology | [12,17] |
| Poor pharmacokinetic properties | Limited clinical applicability | Medicinal chemistry optimization | [1,11] |
| Insufficient ecological interpretation | Incomplete functional understanding | Spatial metabolomics and synthetic ecology | [18,19,20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



