Submitted:
15 May 2026
Posted:
15 May 2026
You are already at the latest version
Abstract
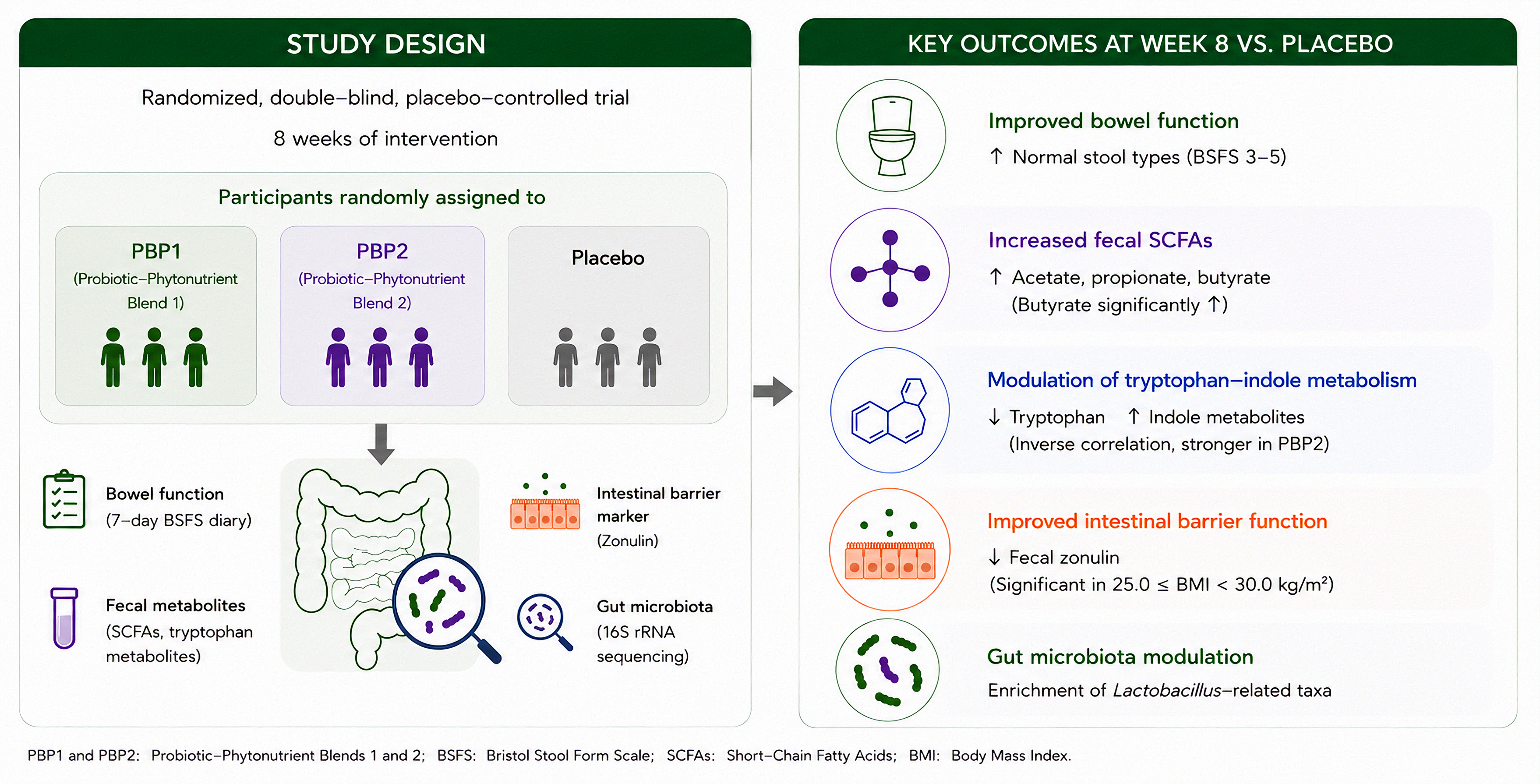
Background/Objectives: Probiotic interventions are widely used to improve intestinal health; however, comparative evidence on multi-strain formulations with different po-tencies, particularly when combined with plant-based complexes, remains limited. eval-uated the effects of two probiotic blends containing phytonutrients (PBP1 and PBP2) on bowel function, microbial metabolites, and gut barrier-related markers. Methods: In this randomized, double-blind, placebo-controlled trial, participants received PBP1, PBP2, or placebo for 8 weeks. Stool patterns (7-day Bristol Stool Form Scale (BSFS) diary), fecal short-chain fatty acids (SCFAs), tryptophan metabolites, zonulin, and gut microbiota were assessed at baseline and Week 8. Efficacy was evaluated by comparing each inter-vention group with the placebo group. Results: Both PBP1 and PBP2 significantly in-creased the proportion of normal stool types (BSFS types 3–5) compared with placebo (p < 0.05). Fecal SCFA levels, including acetate, propionate, and butyrate, were significantly increased in both intervention groups. Notably, butyrate levels were significantly ele-vated compared with placebo. Fecal tryptophan levels decreased, while indole metabo-lites showed increasing trends, with an inverse correlation observed between tryptophan and indole, particularly in the PBP2 group. Fecal zonulin showed a decreasing trend, with significant reductions in participants with 25.0≤BMI<30.0 kg/m². Microbiome analysis revealed preserved alpha diversity with selective compositional shifts, including en-richment of Lactobacillus-related taxa. Conclusions: Supplementation with PBP1 and PBP2 improved bowel function and was associated with changes in microbiome-derived metabolites, including SCFAs and tryptophan–indole metabolism, with BMI-dependent changes in barrier markers. These findings suggest a potential role of microbi-ome-mediated metabolic modulation in intestinal health.
Keywords:
probiotic
; phytonutrients
; SCFAs
; butyrate
; tryptophan
; gut barrier function
; gut microbiome
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



