Submitted:
02 May 2026
Posted:
04 May 2026
You are already at the latest version
Abstract
Brucellosis and tuberculosis (TB) are chronic infectious diseases of international public health importance, with developing countries being most affected. Co-infection in spite of low prevalence raises a diagnostic challenge due to the common symptoms of infection and immune response. Recently, it has been shown that chronic infections affect cell stress pathways such as oxidative stress and telomere function.
The current literature review provides an overview of the relationship between brucellosis and tuberculosis at a molecular level, focusing on telomere biology, oxidative stress and the mechanisms of antimicrobial resistance. Due to chronic immune response in brucellosis and TB patients, an increase in ROS level is observed leading to DNA damage and subsequent telomere shortening and alteration of telomerase activity.
These alterations might be responsible for immune senescence, weakened defense response and persistent infection. In addition, different methods of drug resistance have been discovered among brucellae and mycobacteria, such as mutation in target sites, efflux systems and intracellular persistence, making their eradication difficult. Finally, the potential role of telomere-related genes and biomarkers of oxidative stress in diagnosis and prognosis is also highlighted.
Insight into these interrelated pathways would allow us to have a better understanding of host-pathogen interaction, and hence offer a possible means of developing new strategies in the fight against co-infection.
Keywords:
brucellosis
; tuberculosis
; telomere biology
; oxidative stress
; drug resistance
; co-infection
; telomerase
; ROS
; DNA damage
; immune senescence
1. Introduction
Brucellosis and tuberculosis (TB) remain formidable challenges to global public health, particularly in low- and middle-income countries where they are endemic [1]. In combination, all these bacterial diseases affect a huge number of people each year, causing morbidity, economic losses, and even death. Brucellosis, a zoonosis infection, which is caused by bacteria from the Brucella genus, especially B. melitensis, B. abortus, and B. suis, is contracted in humans from infected animals, contaminated unpasteurized milk, or inhalation of aerosols [2]. Tuberculosis, caused by Mycobacterium tuberculosis, spreads via airborne droplets and primarily affects the lungs, though it can disseminate to extrapulmonary sites. Despite their different etiological origins, both pathogens share a critical pathogenic feature: they are facultative intracellular organisms that have evolved sophisticated mechanisms to survive and replicate within host immune cells, particularly macrophages [2]. This intracellular lifestyle enables them to evade host immune responses, establish persistent infections, and cause chronic disease that can last for years if untreated.
There is significant geographic association between brucellosis and tuberculosis, particularly within the Mediterranean region, the Middle East, certain parts of Asia, Central/South America, and sub-Saharan Africa. Within this geographical area, livestock rearing is practiced, tuberculosis rates are high, and health care facilities can be few and far between. Coinfections with both Brucella spp. and M. tuberculosis occur quite commonly, but are greatly underrecognized and under-researched [3]. The main reason for this diagnostic discrepancy is the striking resemblance in the presentation of both disorders. Both conditions usually manifest themselves through a range of symptoms such as fever, sweating at night, extreme tiredness, unexpected weight loss, joint pains, and loss of appetite [4]. As far as brucellosis is concerned, hepatosplenomegaly and lymphadenopathy are also some symptoms of this infection, whereas TB can cause symptoms such as chronic cough, hemoptysis, and chest pain if it affects the lungs of a patient. When both these infections happen simultaneously, they create further confusion regarding the proper diagnosis of patients, since they show many overlapping symptoms. A patient may be diagnosed and treated with the wrong type of disease and antibiotics due to this problem [5]. In endemic regions, clinical algorithms that rely solely on symptom assessment frequently fail to distinguish between these entities, underscoring the urgent need for better molecular and biomarker-based diagnostic tools [6].
On a cellular level, both brucellosis and tuberculosis are marked by sustained immune response. Following the infection, the host cells, including macrophages and dendritic cells, recognize the PAMPs with the help of the pattern recognition receptors (PRRs) like Toll-like receptors (TLRs) [7]. It leads to a series of pro-inflammatory signal transduction pathways that involve the nuclear factor kappa B pathway (NF-kB) and mitogen-activated protein kinases (MAPKs). This results in the secretion of various cytokines, including tumour necrosis factor alpha (TNF-α), and interleukin 1beta (IL-1β) [8]. In addition, ROS and RNS are produced by the activation of phagocytes through the activity of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase complex and iNOS enzymes, respectively. These chemicals are designed to eliminate or inhibit the survival of pathogens within the cell [9]. In chronic infection cases, however, this defense strategy can be viewed as a two-sided weapon. The prolonged activity of the immune response results in constant ROS and RNS generation, outpacing the body’s natural defenses against antioxidants. This situation is called oxidative stress, which results in collateral damage to the body’s own components, such as protein (carbonylation, nitration), lipids (lipid peroxidation), and DNA (strand breakage, base alterations) [10]. Among the most susceptible segments of genomic DNA are telomeres, composed of TTAGGG nucleotide repeats that form the chromosome ends. Telomeres are highly reactive towards oxidants due to their large amount of guanine residues, which can be easily oxidized to form 8-oxo-7,8-dihydroguanine (8-oxo-dG). Oxidative damage to telomeres causes telomere shortening and activates DNA damage response leading to genomic instability [11].
The biology of telomeres is a new but highly important area that needs to be addressed for chronic infections. Telomeres help guard chromosomes against fusing together, being degraded, and engaging in illegitimate recombination. Telomeres are largely stabilized by telomerase, which is a ribonucleoprotein enzyme with a TERT subunit and TERC RNA component [12]. The telomere length decreases with each replication process in healthy subjects and acts as a cellular replicative life span meter. However, in case of oxidative stress and inflammation, this process becomes much faster. In relation to tuberculosis infection, there is evidence that shows that in PBMCs of the infected subjects, the telomeres are shorter than those in the healthy subjects [13]. While some studies show that successful therapy against TB disease could help recover telomere length, brucellosis remains poorly understood in this regard.
Notably, no specific study has evaluated the role of telomeres in brucellosis infection in humans, nor has there been any investigation of telomeres in patients with concurrent infections by Brucella and M. tuberculosis [14]. This knowledge gap is striking, given the chronic nature of brucellosis, its capacity to induce prolonged oxidative stress, and its epidemiological overlap with TB.
Yet another dimension of complexity is posed by the increasing issue of drug resistance. MDR-TB refers to tuberculosis that is resistant to multiple drugs, such as isoniazid and rifampicin. MDR-TB poses a huge challenge to global health, with half a million new cases occurring each year [15]. XDR-TB involves the addition of fluoroquinolones and second-line injectable antibiotics. With regard to brucellosis, although acquired resistance is less likely to occur, there is frequent failure and relapses of the disease due to the persistence of bacteria intracellularly, the failure of antibiotics to penetrate into the bacteria, or insufficient duration of the treatment course. Relapse rates with brucellosis treatment can range from 5% to 15% despite combination therapy with appropriate drugs such as doxycycline in conjunction with either rifampicin or an aminoglycoside. Notably, rifampicin, which is commonly used in the treatment of TB, is also used in the treatment of brucellosis. The extensive use of rifampicin in treating TB in endemic regions is likely to lead to the selection of rifampicin-resistant Brucella species, although this aspect has not been systematically investigated [16]. Understanding the molecular mechanisms that drive drug resistance in both pathogens, and how these mechanisms intersect with oxidative stress and telomere dysfunction, is therefore an urgent research priority.
This review aims to compile the current knowledge on the molecular interplay between brucellosis and TB, with a specific focus on three interconnected domains based on the role of oxidative stress in disease pathogenesis and host damage; the impact of chronic infection on telomere biology and cellular ageing; and the mechanisms of drug resistance in each infection and how co-infection may exacerbate treatment challenges. Through this combination of themes, it is hoped that a complete theoretical framework will be developed that will guide future research, suggest possible biomarkers for diagnostic purposes, and indicate new treatment options for patients afflicted with these debilitating chronic infections.

Table 1.
Comparative overview of brucellosis and tuberculosis pathogenesis, oxidative stress, telomere biology, drug resistance mechanisms, and clinical features, with emphasis on the impact of co-infection.
Table 1.
Comparative overview of brucellosis and tuberculosis pathogenesis, oxidative stress, telomere biology, drug resistance mechanisms, and clinical features, with emphasis on the impact of co-infection.
| Feature | Brucellosis | Tuberculosis | Impact of Co-Infection | References |
|---|---|---|---|---|
| Causative Agent | Brucella melitensis, B. abortus, B. suis (Gram-negative coccobacillus) | Mycobacterium tuberculosis (acid-fast bacillus) | Dual intracellular infection: potential competition for macrophage niches | [16] |
| Primary Reservoir | Domestic animals (goats, cattle, swine); zoonotic | Humans (airborne transmission) | Overlap in endemic rural/agricultural regions | [17] |
| Intracellular Niche | Brucella-containing vacuole (BCV) that acquires ER markers | M. tuberculosis-containing phagosome that inhibits lysosomal fusion | Both evade phagolysosome fusion; may compete for host resources | [18] |
| Immune Response | Th1-mediated (IFN-γ, TNF-α) but with anti-inflammatory IL-10 modulation | Th1-dominated (IFN-γ, IL-12, TNF-α); granuloma formation | Potential immune exhaustion and dysregulation | [19] |
| Oxidative Stress Markers | ↑ MDA, ↓ SOD, ↓ GPx; limited human studies | ↑ MDA, ↑ 8-oxo-dG, ↓ GSH, ↓ total antioxidant capacity | Likely amplified oxidative stress; more severe DNA damage | [20] |
| Telomere Biology | Telomere shortening | Shorter telomere length in PBMCs; altered telomerase activity | Expected accelerated telomere attrition and immune senescence | [21] |
| Primary Drug Resistance Mechanisms | rpoB mutations (rifampicin); Efflux pumps (TetA/B for doxycycline); Intracellular persistence/relapse | rpoB mutations (rifampicin); katG / inhA mutations (isoniazid); gyrA/B (fluoroquinolones); Efflux pumps | Overlapping rifampicin use; risk of cross-resistance; drug drug interactions (e.g., rifampicin reduces doxycycline levels) | [22] |
| Standard Treatment | Doxycycline + rifampicin OR doxycycline + gentamicin/streptomycin | RHZE (rifampicin, isoniazid, pyrazinamide, ethambutol) | Prolonged rifampicin exposure; complex regimens; higher relapse risk | [23] |
| Common Clinical Overlap | Fever, night sweats, weight loss, fatigue, arthralgia, hepatosplenomegaly | Fever, night sweats, weight loss, fatigue, chronic cough, hemoptysis | High risk of misdiagnosis or delayed treatment | [24] |
| Pathognomonic Finding | Non-caseating granulomas (liver, spleen, bone marrow) | Caseating granulomas with Langhans giant cells (lungs, lymph nodes) | Histology alone cannot distinguish; requires culture or molecular diagnostics | [25] |
2. Pathogenesis of Brucellosis and Tuberculosis
2.1. Brucellosis Pathogenesis
The genus Brucella consists of Gram-negative bacteria with the ability of being a facultative intracellular organism with an extremely evolved mechanism of subverting the host immune system. Entry usually occurs through the mucosa of the oropharynx, gastrointestinal tract, and respiratory tract. As illustrated in Figure 1, the organs involved in brucellosis [26]. Including the skin, lungs, mouth, lymph nodes, spleen, liver, gastrointestinal tract, bone marrow, and genital tract. The next step involves the uptake of the Brucella organisms by the professional phagocytic cells, such as macrophages and dendritic cells. Unlike other intracellular bacteria [27]. The bacteria release T4SS proteins, which are coded for by virB operon. The bacterial effector proteins that are released by the bacterium target the vesicle traffic, thus blocking the development of the phagosome into a lysosome. Rather than developing into a lysosome, the BCV acquires the membrane markers of the endoplasmic reticulum (ER) and later converts into an ER-like niche [28]. In this enclosed environment, the Brucella species multiplies without causing inflammatory stimuli.
The pathogen also suppresses the host’s immune system through inhibiting TLR signaling, decreased levels of TNF-α production, and induction of an anti-inflammatory state where IL-10 prevails [29]. This mode of immune escape makes Brucella capable of developing chronic and long-lasting infections. One of the most prominent pathological features of chronic brucellosis is the development of granulomas. These are defined as organized collections of macrophages, lymphocytes, and multinucleated giant cells. Nevertheless, the granulomas formed in brucellosis cases are not caseating granulomas like those seen in tuberculosis cases and can be dissolved independently [30].
2.2. Tuberculosis Pathogenesis
Mycobacterium tuberculosis is an acid-fast, slow-growing bacillus with a unique lipid-rich cell wall that contributes to its virulence and drug resistance. Infection begins when aerosolized droplets containing the bacteria are inhaled and reach the alveolar spaces. Alveolar macrophages are the first line of defence, engulfing the bacilli through phagocytosis. However, M. tuberculosis has evolved multiple strategies to survive within these hostile cells. The bacterium produces a variety of factors that inhibit phagosome-lysosome fusion, including sulfolipids, trehalose dimycolate (cord factor), and the secreted protein kinase G (PknG) [31]. It also prevents phagosomal acidification by excluding the vacuolar ATPase proton pump. The M. tuberculosis-containing phagosome retains an early endosomal phenotype, allowing the bacterium to access nutrients and avoid degradation [32]. Like Brucella, M. tuberculosis manipulates host cell death pathways, inhibiting apoptosis while promoting necrotic cell death, which facilitates bacterial dissemination. The hallmark of TB pathology is the granuloma, a highly organized immune structure that contains the infection but also serves as a site of bacterial persistence [33]. Within the granuloma, M. tuberculosis can enter a dormant, non-replicating state characterized by altered metabolism, reduced cell wall synthesis, and increased tolerance to antibiotics. Reactivation of dormant bacteria is a major cause of active TB in latently infected individuals, especially when immunity is compromised by conditions such as HIV co-infection, malnutrition, or ageing [34].
2.3. Co-Infection Dynamics
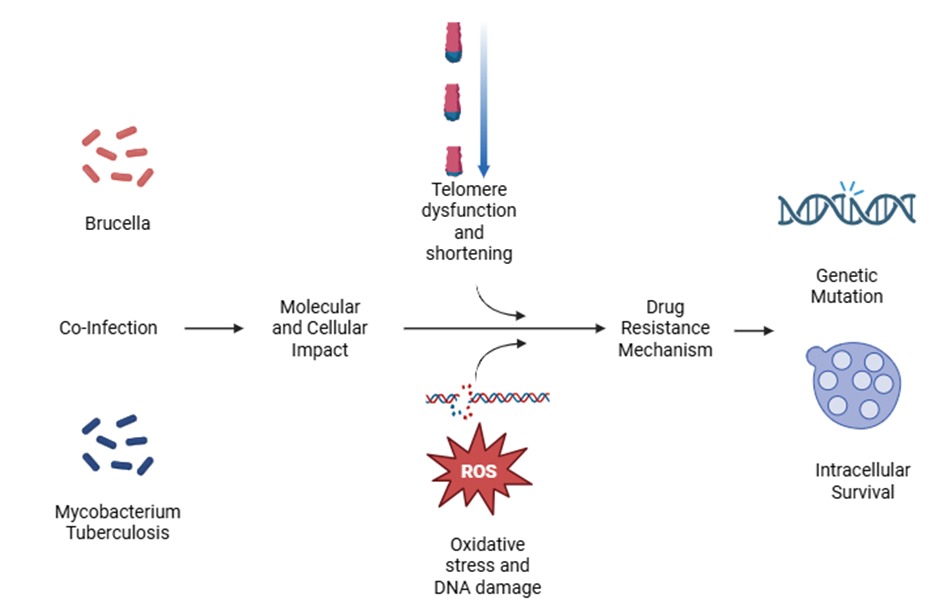
The presence of both Brucella and M. tuberculosis in the same organism is not just additive but synergistic for causing an immune imbalance. Both microorganisms have a tendency to infect similar intracellular locations such as macrophages, and cause immune reactions that produce IFN-γ, activate macrophages, and form granulomas.[35]. T-cell exhaustion due to prolonged antigenic stimulation can occur for patients suffering from both infections; this may be accompanied by increased expression of inhibitory receptors such as PD-1, CTLA-4, and TIM-3, as well as reduced production of effector cytokines. This will prevent the body from clearing the infection [36]. Furthermore, the continuous state of inflammation results in higher levels of oxidative stress compared to cases of isolated infection Figure 2. It is still unknown what kind of competition for nutrients might occur or whether bacteria can interact directly or indirectly.
2.4. Oxidative Stress in Brucellosis and TB
In oxidative stress, which is an essential pathophysiologic component of brucellosis and TB, there is an interaction between chronic inflammatory response, tissue injury, immunological malfunctioning, and genetic instability. In healthy individuals, there is an equilibrium of free radical generation and neutralization. However, this balance becomes unbalanced in chronic infections [37].
In TB, there is ample evidence of increased oxidative stress biomarkers. The plasma concentration of malondialdehyde (MDA), which results from lipid peroxidation, is consistently higher among TB-infected subjects relative to healthy individuals. It also falls following effective therapy of the disease [38].
In the same way, high levels of 8-hydroxy-2′-deoxyguanosine (8-oxo-dG) from urine and plasma, an indicator of DNA oxidative damage, have been found in TB cases, and these levels correlate positively with disease severity and bacterial burden. TB patients have decreased antioxidative status in the form of decreased levels of GSH, TAS, and decreased enzyme activities like SOD, CAT, and GPx. In some studies, the ratio of GSSG to GSH was found to be significantly higher [39]. It should be noted that the bacteria itself is subjected to reactive oxygen species produced by the host and, hence, has developed its own antioxidant mechanisms comprising catalase-peroxidase (KatG), superoxide dismutase (SodA, SodC), and alkyl hydroperoxide reductase (AhpC). The role of these antioxidant enzymes in protecting the bacteria is supplemented by their ability to regulate the redox state of the host. Catalase-peroxidase (KatG), for instance, is involved in the activation of the prodrug isoniazid; katG gene mutations have been identified as the principal cause of isoniazid resistance [40].
However, the pattern of oxidative stress in cases of brucellosis is not that well-researched. It was found out that in patients with both acute and chronic forms of the disease, serum MDA levels are elevated, and antioxidant enzyme activity (SOD, GPx) is diminished [41]. Nitric oxide (NO) overproduction has been observed in some other studies, evidenced by nitrite/nitrate accumulation. The genus Brucella is endowed with several antioxidants, such as superoxide dismutase (SodA, SodC), catalase (KatE), and an unusual periplasmic cytochrome c peroxidase (Ccp). This helps Brucella to survive oxidative damage during the burst response of activated macrophages, leading to chronic infection. Notably, the genus Brucella does not have a KatG orthologue; therefore, it is intrinsically more sensitive than Mycobacterium tuberculosis to oxidative conditions [42].
It would seem logical to postulate that there should be an enhancement of oxidative stress in co-infected patients because the burden of two organisms within the same cell will stimulate increased ROS and RNS formation along with depleting the body's supply of antioxidants. As a result, there would be faster degradation of DNA, loss of telomeres, and premature immune senescence. However, there is still no data on co-infections available [43].
2.5. Telomere Biology and Chronic Infection
Telomeres are made up of tandem repeats of TTAGGG nucleotides along with shelterin protein complex TRF1, TRF2, POT1, TIN2, TPP1, and RAP1. Their main biological role is to recognize the true chromosome ends and protect them from being repaired as broken DNA molecules [44].
Telomere length decreases during each round of cell division owing to the end replication problem and oxidative damage to DNA. Oxidized guanines that accumulate at the telomere sequence are not efficiently removed via base excision repair pathways. Instead, these lesions result in DNA breaks, which accelerate the loss of telomere length. Short telomeres result in senescence and apoptosis of immune cells [45].
Telomere shortening has been shown to occur due to chronic infections such as TB. TB patients were found to have shorter telomere length in PBMCs compared to control individuals, and telomere shortening was positively related to high bacterial load and radiologic disease severity [46]. Other research discovered that the level of telomerase expression was decreased in T-cells from patients with tuberculosis, and thus telomeres were not maintained properly. However, once patients were cured from their diseases, there was an evidence that there is partial restoration of telomere length. In cases of co-infection of HIV and tuberculosis, telomere shortening became even more obvious [47].
For brucellosis, no direct studies of telomere length or telomerase activity have been published to date. However, given that chronic brucellosis induces sustained inflammation and oxidative stress, it is highly plausible that telomere attrition occurs [46]. In co-infected patients, the combined effect of two chronic bacterial infections, each driving immune cell proliferation and oxidative damage, could lead to profound telomere shortening, accelerating immune senescence. This, in turn, would impair the host's ability to control both infections, creating a vicious cycle of persistent inflammation, immune exhaustion, and increasing pathogen burden [48].
Telomere-associated proteins also warrant attention. TRF2, for example, protects telomeres from ATM kinase-mediated DNA damage responses and is highly sensitive to oxidative modification. Chronic oxidative stress could impair TRF2 function, leading to telomere deprotection and genomic instability even in the absence of extreme telomere shortening [49]. Similarly, POT1, which binds single-stranded telomeric overhangs, can be inactivated by ROS. Future studies should investigate not only telomere length but also the integrity of the shelterin complex in TB and brucellosis patients [50].
3. Drug Resistance Mechanisms
3.1. Drug Resistance in Tuberculosis
Resistance to tuberculosis drugs occurs genetically, as a result of spontaneous mutations in genes on the chromosome. In the case of Rifampicin, the strongest antitubercular drug, the cause for resistance occurs due to mutations in the gene rpoB, responsible for encoding the β subunit of RNA polymerase [51]. More than 95% of rifampicin-resistant isolates have mutations within an 81-base pair segment in rpoB known as the rifampicin resistance-determining region (RRDR). Such mutations include Ser531Leu and His526Asp, which decrease rifampicin binding [52]. Isoniazid resistance requires several genetic modifications and is complicated because it is mediated by multiple mutations. The most common type of katG mutation leads to elimination or reduced catalase-peroxidase activity, which results in the failure of isoniazid activation. Mutation in the promoter region of inhA leads to overproduction of its protein target (enoyl-ACP reductase) and consequently results in isoniazid resistance. Other rare mutations occur in ahpC, kasA, and ndh [53]. Fluoroquinolone resistance, seen in MDR TB treatment, results from mutations in the QRDR (quinolone resistance determining region) in gyrA and gyrB genes encoding DNA gyrase enzyme. The second line injectables such as amikacin, kanamycin, and capreomycin get deactivated due to mutation in rrs genes (encoding for 16S rRNA) and eis promoters. Apart from genetic mutations in TB bacteria, there are efflux pumps (like Rv1258c, Rv1410c), which pump out the drug and help in achieving resistance to these drugs at low levels while developing high-level resistance. Unlike biofilms in other bacteria, biofilms in M. tuberculosis are not very common [54].
3.2. Drug Resistance in Brucellosis
The Brucella species exhibit natural resistance to multiple drugs like colistin, bacitracin, and various β-lactams, owing to their non-permeability and use of efflux pumps. However, for the combination therapy using doxycycline and rifampicin or doxycycline and an aminoglycoside like streptomycin or gentamicin, the occurrence of resistance is very rare but still exists [55]. Brucella's resistance to rifampicin involves mutations within rpoB, just like TB bacteria, thereby creating the potential that antibiotic-resistance programs against TB could lead to rifampicin-resistant strains of Brucella [56]. Resistance to doxycycline is uncommon but can be due to efflux pumps like TetA and TetB or mutations in the 16S rRNA genes. Resistance to aminoglycosides can be achieved by aminoglycoside-modifying enzymes like AAC(6')-I and APH(3')-I or mutations in the ribosomal proteins.
The biggest threat faced by clinicians is not resistance but relapse [57]. Brucella is capable of surviving intracellularly for extended periods, avoiding any contact with antibiotics, only to resurface when treatment is discontinued. Such a phenomenon does not represent resistance, but persistence or tolerance, potentially associated with the existence of non-dividing persister cells, characterized by metabolic dormancy and higher survival rates in the presence of antibiotics. The formation of such cells can potentially be stimulated by oxidative stress in both Brucella and M. tuberculosis [58].
3.3. Impact of Co-Infection on Drug Resistance
Co-infection with Brucella and M. tuberculosis complicates drug therapy in several ways. First, treatment regimens overlap: rifampicin is used in both standard TB therapy (Rifampicin, Isoniazid, Pyrazinamide, Ethambutol; RHZE) and standard brucellosis therapy (doxycycline plus rifampicin) [5]. A co-infected patient may therefore receive prolonged rifampicin therapy, increasing the risk of rifampicin resistance in either pathogen. Second, drug-drug interactions are possible, particularly involving rifampicin, a potent inducer of cytochrome P450 enzymes. Rifampicin can reduce serum levels of doxycycline, potentially compromising brucellosis treatment [59]. Third, the immune dysregulation and oxidative stress in co-infection may impair the efficacy of antibiotics that require host immune cooperation (e.g., isoniazid requires KatG activation, which may be affected by redox status). Fourth, co-infection may promote the selection of multidrug-resistant strains by creating a high-burden, prolonged, and suboptimally treated environment. Systematic studies of drug resistance patterns in co-infected patients are urgently needed [60].
3.4. Interconnection Between Telomere Biology, Oxidative Stress, and Drug Resistance
The three topics discussed above have an intrinsic relationship and are interconnected. First, oxidative stress leads to direct damage to the structure of telomeres, resulting in their rapid degradation. Secondly, dysfunctional and short telomeres contribute to cellular senescence and exhaustion of the host's immune response, preventing the elimination of intracellular pathogenic microorganisms. Finally, persistent infection causes oxidative stress [61]. In this cycle, drug resistance can occur or be enhanced. Mutations rates can be increased by DNA damage caused by oxidative stress. ROS in M. tuberculosis can induce oxidative DNA lesions which can facilitate faster development of drug resistance-conferring mutations [62]. On the contrary, successful pharmacotherapy decreases bacterial load and inflammation, leading to reduced oxidative stress and telomere repair.
Thus, telomere length as well as oxidative stress parameters can be used as biomarkers of response to pharmacotherapy [63]. Antioxidants like N-acetylcysteine (NAC) or vitamin C, decreasing oxidative stress, may increase efficiency of antibiotics and prevent telomere shortening; however, no studies on their efficacy in clinical practice have been conducted so far. Finally, modulation of telomerase activity (such as TERT activators) represents an innovative strategy to promote immunity [64].
4. Clinical Symptoms and Implications with Future Perspectives
Both brucellosis and TB have overlapping symptoms, which are depicted in Figure 3, such as fever, malaise, and weight loss, which can be difficult to distinguish from each other. In areas where TB is endemic, health care professionals need to be on the lookout for cases of brucellosis in patients who have been in contact with animals or consumed unpasteurized milk [65]. The presence of persistent fever, arthralgia, or hepatosplenomegaly in patients undergoing anti-tuberculosis therapy should prompt consideration of concurrent Brucella infection and appropriate serological testing [66].
From a clinical perspective, new molecular biomarkers like telomere length, telomerase expression, and oxidative stress, including MDA, 8-oxo-dG, and the GSH/GSSG ratio, have promising potentials in disease staging, risk of relapse prediction, and drug efficacy evaluation [13]. However, their application remains largely investigational, and robust validation through large-scale, longitudinal studies is required before routine clinical implementation.
Adjunct therapy that focuses on oxidative stress could be beneficial from a therapeutic point of view. Antioxidants like N-acetylcysteine and vitamin C have been suggested for reducing oxidative stress, which may improve immune response and increase antimicrobial effectiveness [67]. However, there is little proof to support this, and clinical studies are required to determine their effectiveness. Future work must focus on conducting exhaustive epidemiological studies that will help determine the true prevalence of the disease. At the same time, efforts should be made to conduct mechanistic studies that can help shed light on the involvement of oxidative stress and telomere dynamics in the disease process. It is just as important to conduct clinical trials focusing on the effects of treatment in co-infected patients, as well as issues of drug resistance and host–pathogen interactions.
5. Conclusions
Brucellosis and tuberculosis co-infection represents a complex, understudied, and clinically challenging intersection of two chronic intracellular bacterial infections. Chronic inflammation and sustained production of reactive oxygen and nitrogen species lead to oxidative stress, causing collateral damage to host tissues, including accelerated telomere shortening and genomic instability. Telomere attrition, in turn, promotes immune senescence and reduces the host's capacity to eliminate pathogens, perpetuating a vicious cycle of infection, inflammation, and tissue damage. Drug resistance, already a major problem in TB and an emerging concern in brucellosis, is likely exacerbated by co-infection through overlapping treatment regimens, drug–drug interactions, and oxidative stress-induced mutagenesis.
Understanding the molecular interplay among oxidative stress, telomere biology, and drug resistance is not only scientifically fascinating but also clinically urgent. It offers the promise of novel biomarkers for diagnosis and prognosis, as well as new therapeutic avenues that target not only the pathogens themselves but also the host's redox and telomere maintenance pathways. Future research should prioritize well-designed clinical studies of co-infected patients, animal models that recapitulate dual infection, and mechanistic investigations at the host-pathogen interface. Only through such integrated efforts can we hope to improve outcomes for the millions of people living in regions where brucellosis and tuberculosis continue to exact a heavy toll.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Haileamlak. Tuberculosis Continued as Global Challenge Though the Burden Remained High in Low-Income and High-Income Countries. In NLM (Medline); 01 Sep 2018. [Google Scholar] [CrossRef]
- Khairullah, R.; et al. Brucellosis: Unveiling the complexities of a pervasive zoonotic disease and its global impacts. In Faculty of Veterinary Medicine, University of Tripoli; 2024. [Google Scholar] [CrossRef]
- Rubach, M.P.; Halliday, J.E.B.; Cleaveland, S.; Crump, J.A. Brucellosis in low-income and middle-income countries. 2013. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, N.R.; Szostek, J.H. 52-year-old man with arthralgias, fever, and fatigue. Mayo Clin. Proc. 2010, vol. 85(no. 6), 568–571. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.; Zhou, J.; Jiang, X. Diagnosis and management of spinal tuberculosis combined with brucellosis: A case report and literature review. In Spandidos Publications; 01 Apr 2018. [Google Scholar] [CrossRef]
- Assudani, P.J.; et al. Artificial intelligence and machine learning in infectious disease diagnostics: a comprehensive review of applications, challenges, and future directions; Elsevier Inc, 01 Nov 2025. [Google Scholar] [CrossRef]
- Yu, H.; Gu, X.; Wang, D.; Wang, Z. Brucella infection and Toll-like receptors. In Frontiers Media SA; 2024. [Google Scholar] [CrossRef]
- Moens, U.; Kostenko, S.; Sveinbjørnsson, B. The role of mitogen-activated protein kinase-activated protein kinases (MAPKAPKs) in inflammation. 2013. [Google Scholar] [CrossRef]
- Ushio-Fukai, M.; Ash, D.; Nagarkoti, S.; De Chantemèle, E.J.B.; Fulton, D.J.R.; Fukai, T. Interplay between Reactive Oxygen/Reactive Nitrogen Species and Metabolism in Vascular Biology and Disease; Mary Ann Liebert Inc, 01 Jun 2021. [Google Scholar] [CrossRef]
- Manful, F.; Fordjour, E.; Subramaniam, D.; Sey, A.A.; Abbey, L.; Thomas, R. “Antioxidants and Reactive Oxygen Species: Shaping Human Health and Disease Outcomes,” Aug. 01. In Multidisciplinary Digital Publishing Institute (MDPI); 2025. [Google Scholar] [CrossRef]
- Barnes, R.P.; Fouquerel, E.; Opresko, P.L. The impact of oxidative DNA damage and stress on telomere homeostasis. In Elsevier Ireland Ltd; 01 Jan 2019. [Google Scholar] [CrossRef]
- Kam, M.L.W.; Nguyen, T.T.T.; Ngeow, J.Y.Y. Telomere biology disorders. In Nature Research; 01 Dec 2021. [Google Scholar] [CrossRef]
- Gavia-García, G.; Rosado-Pérez, J.; Arista-Ugalde, T.L.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. Telomere length and oxidative stress and its relation with metabolic syndrome components in the aging. In MDPI AG; 01 Apr 2021. [Google Scholar] [CrossRef]
- Katoto, P.D.M.C.; et al. Telomere length and outcome of treatment for pulmonary tuberculosis in a gold mining community. Sci. Rep. 2021, vol. 11(no. 1). [Google Scholar] [CrossRef]
- Seung, K.J.; Keshavjee, S.; Rich, M.L. “Multidrug-resistant tuberculosis and extensively drug-resistant tuberculosis,” Cold Spring Harb. Perspect. Med. 2015, vol. 5(no. 9). [Google Scholar] [CrossRef]
- Batt, J.; Khan, K. Responsible use of rifampin for the treatment of latent tuberculosis infection. In Canadian Medical Association; 2019. [Google Scholar] [CrossRef]
- Zhang, T.; et al. The impacts of animal agriculture on One Health—Bacterial zoonosis, antimicrobial resistance, and beyond; Elsevier B.V., 01 Jun 2024. [Google Scholar] [CrossRef]
- Olowu, I.; Zakariya, M.E.; Abdulkareem, A.A.; Okewale, O.T.; Idris, M.H.; Olayiwola, H.O. Drug Design and Delivery for Intracellular Bacteria: Emerging Paradigms. In John Wiley and Sons Inc; 01 Dec 2025. [Google Scholar] [CrossRef]
- Cavalcanti, Y.V.N.; Brelaz, M.C.A.; Neves, J.K.D.A.L.; Ferraz, J.C.; Pereira, V.R.A. Role of TNF-alpha, IFN-gamma, and IL-10 in the development of pulmonary tuberculosis. 2012. [Google Scholar] [CrossRef] [PubMed]
- Borrego, S.; et al. Oxidative stress and DNA damage in human gastric carcinoma: 8-Oxo-7’8-dihydro-2’-deoxyguanosine (8-oxo-dG) as a possible tumor marker. Int. J. Mol. Sci. 2013, vol. 14(no. 2), 3467–3486. [Google Scholar] [CrossRef]
- Lin, J.; et al. Systematic and Cell Type-Specific Telomere Length Changes in Subsets of Lymphocytes. J. Immunol. Res. 2016, vol. 2016. [Google Scholar] [CrossRef]
- Meng, F.; et al. Efflux pumps positively contribute to rifampin resistance in rpoB mutant Mycobacterium tuberculosis. Ann. Clin. Microbiol. Antimicrob. 2025, vol. 24(no. 1). [Google Scholar] [CrossRef]
- Alavi, S.M.; Alavi, L. Systematic Review Treatment of brucellosis: a systematic review of studies in recent twenty years.
- Levy, L.; Nasereddin, A.; Rav-Acha, M.; Kedmi, M.; Rund, D.; Gatt, M.E. Prolonged fever, hepatosplenomegaly, and pancytopenia in a 46-year-old woman. 2009. [Google Scholar] [CrossRef]
- Kumar, S.N.; Prasad, T.S.; Narayan, P.A.; Muruganandhan, J. Granuloma with langhans giant cells: An overview. J. Oral. Maxillofac. Pathol. 2013, vol. 17(no. 3), 420–423. [Google Scholar] [CrossRef]
- Guimarães, S.; Gomes, M.T.R.; de Araujo, A.C.V.S.C.; Ramos, K.K.S.; Oliveira, S.C. Pathogenicity and virulence of Brucella: Strategies for metabolic adaptation and immune evasion; Taylor and Francis Ltd, 2025. [Google Scholar] [CrossRef]
- Huy, T.X.N.; Nguyen, T.T.; Kim, H.; Reyes, A.W.B.; Kim, S. Brucella Phagocytosis Mediated by Pathogen-Host Interactions and Their Intracellular Survival. In MDPI; 01 Oct 2022. [Google Scholar] [CrossRef]
- Ke, Y.; Wang, Y.; Li, W.; Chen, Z. Type IV secretion system of Brucella spp. and its effectors; Frontiers Media S.A, 2015. [Google Scholar] [CrossRef]
- Pellegrini, J.M.; Gorvel, J.P.; Mémet, S. Immunosuppressive mechanisms in brucellosis in light of chronic bacterial diseases. Microorganisms 2022, vol. 10(no. 7). [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Zhang, L.; Liu, R.; Sa, R.; Zheng, L. Spatiotemporal dynamics of Brucella immune evasion across infection stages. Front. Immunol. 2026, vol. 16. [Google Scholar] [CrossRef]
- Podinovskaia, M.; Lee, W.; Caldwell, S.; Russell, D.G. Infection of macrophages with Mycobacterium tuberculosis induces global modifications to phagosomal function. Cell. Microbiol. 2013, vol. 15(no. 6), 843–859. [Google Scholar] [CrossRef]
- Queval, J. Mycobacterium tuberculosis Controls Phagosomal Acidification by Targeting CISH-Mediated Signaling. Cell Rep. 2017, vol. 20(no. 13), 3188–3198. [Google Scholar] [CrossRef] [PubMed]
- Nisa, A.; Kipper, F.C.; Panigrahy, D.; Tiwari, S.; Kupz, A.; Subbian, S. Different modalities of host cell death and their impact on Mycobacterium tuberculosis infection. In American Physiological Society; 01 Nov 2022. [Google Scholar] [CrossRef]
- Salina, G.; Makarov, V. Mycobacterium tuberculosis Dormancy: How to Fight a Hidden Danger. In MDPI; 01 Dec 2022. [Google Scholar] [CrossRef]
- Masoudian, M.; Derakhshandeh; Seno, G. Brucella melitensis and Mycobacterium tuberculosis depict overlapping gene expression patterns induced in infected THP-1 macrophages. 2015. Available online: http://rest.gene-.
- Apol, Á.D.; Winckelmann, A.A.; Duus, R.B.; Bukh, J.; Weis, N. The Role of CTLA-4 in T Cell Exhaustion in Chronic Hepatitis B Virus Infection. In MDPI; 01 May 2023. [Google Scholar] [CrossRef]
- Chaudhary, P.; et al. Oxidative stress, free radicals and antioxidants: potential crosstalk in the pathophysiology of human diseases; Frontiers Media S.A, 2023. [Google Scholar] [CrossRef]
- Kulkarni, R.; Deshpande, A.; Saxena, R.; Saxena, K. A study of serum malondialdehyde and cytokine in tuberculosis patients. J. Clin. Diagn. Res. 2013, vol. 7(no. 10), 2140–2142. [Google Scholar] [CrossRef]
- Graille, M.; Wild, P.; Sauvain, J.J.; Hemmendinger, M.; Canu, I.G.; Hopf, N.B. Urinary 8-OHDG as a biomarker for oxidative stress: A systematic literature review and meta-analysis. In MDPI AG; 01 Jun 2020. [Google Scholar] [CrossRef]
- Tyagi, P.; Dharmaraja, A.T.; Bhaskar, A.; Chakrapani, H.; Singh, A. Mycobacterium tuberculosis has diminished capacity to counteract redox stress induced by elevated levels of endogenous superoxide. Free Radic. Biol. Med. 2015, vol. 84, 344–354. [Google Scholar] [CrossRef]
- Gholami, M.; Roushan, M.H.; Mahjoub, S.; Bijani, A. How is total antioxidant status in plasma of Patients with brucellosis?
- Gusti, A.M.T.; Qusti, S.Y.; Alshammari, E.M.; Toraih, E.A.; Fawzy, M.S. Antioxidants-related superoxide dismutase (Sod), catalase (cat), glutathione peroxidase (gpx), glutathione-s-transferase (gst), and nitric oxide synthase (nos) gene variants analysis in an obese population: A preliminary case-control study. Antioxidants 2021, vol. 10(no. 4). [Google Scholar] [CrossRef] [PubMed]
- Kell, L.; Simon, A.K.; Alsaleh, G.; Cox, L.S. The central role of DNA damage in immunosenescence. In Frontiers Media SA; 2023. [Google Scholar] [CrossRef]
- Galati, A.; et al. TRF1 and TRF2 binding to telomeres is modulated by nucleosomal organization. Nucleic Acids Res. 2015, vol. 43(no. 12), 5824–5837. [Google Scholar] [CrossRef]
- Assalve; et al. Exploring the Link Between Telomeres and Mitochondria: Mechanisms and Implications in Different Cell Types. In Multidisciplinary Digital Publishing Institute (MDPI); 01 Feb 2025. [Google Scholar] [CrossRef]
- Kordinas, V.; Ioannidis, A.; Chatzipanagiotou, S. The telomere/telomerase system in chronic inflammatory diseases. Cause or effect? In MDPI AG; 03 Sep 2016. [Google Scholar] [CrossRef]
- Choi, J.; Fauce, S.R.; Effros, R.B. Reduced telomerase activity in human T lymphocytes exposed to cortisol. Brain Behav. Immun. 2008, vol. 22(no. 4), 600–605. [Google Scholar] [CrossRef]
- Johnson, A.; Rought, T.; Aronov, J.; Pokharel, P.; Chiu, A.; Nasuhidehnavi, A. The impacts of chronic infections on shaping cellular senescence. In BioMed Central Ltd; 01 Dec 2025. [Google Scholar] [CrossRef]
- Karlseder, J.; et al. The telomeric protein TRF2 binds the ATM Kinase and Can Inhibit the ATM-dependent DNA damage response. PLoS Biol. 2004, vol. 2(no. 8). [Google Scholar] [CrossRef] [PubMed]
- Aramburu, T.; Plucinsky, S.; Skordalakes, E. POT1-TPP1 telomere length regulation and disease; Elsevier B.V, 01 Jan 2020. [Google Scholar] [CrossRef]
- Maleki, M.R. Genetic Analysis of Molecular Mechanisms of Drug Resistance in Mycobacterium tuberculosis Against Four Major First-Line Anti-Tuberculosis Drugs (Isoniazid, Rifampin, Ethambutol, and Pyrazinamide). In Dove Medical Press Ltd; 2025. [Google Scholar] [CrossRef]
- Mboowa; Namaganda, C.; Ssengooba, W. “Rifampicin resistance mutations in the 81 bp RRDR of rpoB gene in Mycobacterium tuberculosis clinical isolates using Xpert® MTB/RIF in Kampala, Uganda: a retrospective study,” 2014. Available online: http://www.biomedcentral.com/1471-2334/14/481.
- Hazbón, M.H.; et al. Population genetics study of isoniazid resistance mutations and evolution of multidrug-resistant Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2006, vol. 50(no. 8), 2640–2649. [Google Scholar] [CrossRef]
- Liu, J.; et al. Mutations in efflux pump Rv1258c (Tap) cause resistance to pyrazinamide, isoniazid, and streptomycin in M. tuberculosis. Front. Microbiol. 2019, vol. 10, no. FEB. [Google Scholar] [CrossRef] [PubMed]
- Alamian, S.; Dadar, M.; Etemadi, A.; Afshar, D.; Alamian, M.M. “Antimicrobial susceptibility of Brucella spp. isolated from Iranian patients during 2016 to 2018,” 2019. Available online: http://www.oie.int/en/animal-health-in-the-.
- Marianelli, C.; Ciuchini, F.; Tarantino, M.; Pasquali, P.; Adone, R. Genetic bases of the rifampin resistance phenotype in Brucella spp. J. Clin. Microbiol. 2004, vol. 42(no. 12), 5439–5443. [Google Scholar] [CrossRef] [PubMed]
- Wachino, J.I.; Doi, Y.; Arakawa, Y. Aminoglycoside Resistance: Updates with a Focus on Acquired 16S Ribosomal RNA Methyltransferases. In W.B. Saunders; 01 Dec 2020. [Google Scholar] [CrossRef]
- Mode, S.; Ketterer, M.; Québatte, M.; Dehio, C. Antibiotic persistence of intracellular Brucella abortus. PLoS Negl. Trop. Dis. 2022, vol. 16(no. 7). [Google Scholar] [CrossRef]
- Chen, J.; Raymond, K. Roles of rifampicin in drug-drug interactions: Underlying molecular mechanisms involving the nuclear pregnane X receptor; 15 Feb 2006. [Google Scholar] [CrossRef]
- Pattanaik, K.P.; Sengupta, S.; Jit, B.P.; Kotak, R.; Sonawane, A. Host-mycobacteria conflict: Immune responses of the host vs. the mycobacteria TLR2 and TLR4 ligands and concomitant host-directed therapy. In Elsevier GmbH; 01 Nov 2022. [Google Scholar] [CrossRef]
- Sienkiewicz, M.; Sroka, K.; Binienda, A.; Jurk, D.; Fichna, J. A new face of old cells: An overview about the role of senescence and telomeres in inflammatory bowel diseases. In Elsevier Ireland Ltd; 01 Nov 2023. [Google Scholar] [CrossRef]
- Liebenberg, D.; Gordhan, B.G.; Kana, B.D. Drug resistant tuberculosis: Implications for transmission, diagnosis, and disease management; Frontiers Media S.A, 23 Sep 2022. [Google Scholar] [CrossRef]
- Akter, R.; Bishakha, A.C.; Rajib, R.; Raj, A.; Roja, M.S.; Noor, F. Targeting telomere dynamics with plant-derived compounds: Molecular strategies against aging; Elsevier B.V., 01 Jan 2025. [Google Scholar] [CrossRef]
- Voghel, G.; et al. Chronic treatment with N-acetyl-cystein delays cellular senescence in endothelial cells isolated from a subgroup of atherosclerotic patients. Mech. Ageing Dev. 2008, vol. 129(no. 5), 261–270. [Google Scholar] [CrossRef]
- Lamba, A.S.; et al. Brucellosis Masquerading as Disseminated Tuberculosis: a Clinical Case report. 2023. [Google Scholar] [CrossRef]
- Xu, N.; et al. Evaluating the efficacy of serological testing of clinical specimens collected from patients with suspected brucellosis. PLoS Negl. Trop. Dis. 2023, vol. 17(no. 2). [Google Scholar] [CrossRef]
- Vecchio; Jamil, T.; Khamesipour, F.; Heidary, M. Prevalence, diagnosis, and manifestations of brucellosis: A systematic review and meta-analysis.
Figure 1.
The schematic diagram showing the organs that are primarily affected by brucellosis. It can be noted from the schematic illustration that brucellosis affects various body organs. Among them are bone marrow, lungs, liver, and spleen. Infection of the bone marrow is linked to the occurrence of such disorders as spondylitis, which primarily targets the spinal cord. The lungs constitute a relatively uncommon site for the infection but are still considered clinically important in cases of brucellosis. The liver and spleen infections may present as hepatosplenomegaly.
Figure 1.
The schematic diagram showing the organs that are primarily affected by brucellosis. It can be noted from the schematic illustration that brucellosis affects various body organs. Among them are bone marrow, lungs, liver, and spleen. Infection of the bone marrow is linked to the occurrence of such disorders as spondylitis, which primarily targets the spinal cord. The lungs constitute a relatively uncommon site for the infection but are still considered clinically important in cases of brucellosis. The liver and spleen infections may present as hepatosplenomegaly.

Figure 2.
Figure demonstrating the interaction between chronic infections, oxidative stress, and telomere dysfunction. Chronic infection causes prolonged production of pro-inflammatory cytokines, such as IL-6, TNF-α, and IFN-γ, which contribute to the production of ROS and RNS, causing oxidative stress. The increased oxidative stress is a challenge for the cellular antioxidant defense mechanism, such as GSH, SOD, and catalase. Oxidative stress causes shortening of telomeres and damage to DNA, leading to compromised cell structure. Cell damage and telomere shortening aid in the establishment of biofilm or resistant bacteria, leading to chronic infection and a vicious cycle of pathology.
Figure 2.
Figure demonstrating the interaction between chronic infections, oxidative stress, and telomere dysfunction. Chronic infection causes prolonged production of pro-inflammatory cytokines, such as IL-6, TNF-α, and IFN-γ, which contribute to the production of ROS and RNS, causing oxidative stress. The increased oxidative stress is a challenge for the cellular antioxidant defense mechanism, such as GSH, SOD, and catalase. Oxidative stress causes shortening of telomeres and damage to DNA, leading to compromised cell structure. Cell damage and telomere shortening aid in the establishment of biofilm or resistant bacteria, leading to chronic infection and a vicious cycle of pathology.

Figure 3.
Venn diagram illustrating the overlap and distinction in clinical manifestations of brucellosis and tuberculosis, along with the associated risk of misdiagnosis. Brucellosis (orange circle) is characterized by arthralgia, hepatosplenomegaly, and undulant fever, whereas tuberculosis (grey circle) presents with chronic cough, hemoptysis, and apical infiltrates. The overlapping region highlights shared symptoms, including fever, night sweats, weight loss, and fatigue, which contribute significantly to diagnostic challenges. The upper intersecting area emphasizes the increased risk of misdiagnosis due to these nonspecific and overlapping clinical features, particularly in endemic settings.
Figure 3.
Venn diagram illustrating the overlap and distinction in clinical manifestations of brucellosis and tuberculosis, along with the associated risk of misdiagnosis. Brucellosis (orange circle) is characterized by arthralgia, hepatosplenomegaly, and undulant fever, whereas tuberculosis (grey circle) presents with chronic cough, hemoptysis, and apical infiltrates. The overlapping region highlights shared symptoms, including fever, night sweats, weight loss, and fatigue, which contribute significantly to diagnostic challenges. The upper intersecting area emphasizes the increased risk of misdiagnosis due to these nonspecific and overlapping clinical features, particularly in endemic settings.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



