Submitted:
25 April 2026
Posted:
28 April 2026
You are already at the latest version
Abstract
The mechanosensitive ion channel Piezo1 acts as a crucial molecule for cellular mechanical signal sensing and transduction. It transforms physical mechanical cues in the microenvironment, including matrix stiffness, fluid shear stress, and tissue tension, into intracellular biochemical signals through Ca2⁺ influx-mediated mechanisms. Consequently, it regulates the activation, proliferation, differentiation, migration, and effector functions of blood cells. Herein, we review the research progress of Piezo1 in various white blood cells, with a particular emphasis on its functional regulatory mechanisms in neutrophils, macrophages, platelets, and T and B lymphocytes. We briefly summarize its current functional status in natural killer cells, dendritic cells, plasma cells, platelets, and other cell types. We analyze the integrated effects and multi-cellular cooperative interactions of Piezo1-mediated blood cell mechanotransduction across physiological and pathological contexts. We discuss the potential value of Piezo1 as a mechano-immunotherapeutic target, therapeutic strategies, and challenges facing clinical translation. Finally, we provide perspectives on future research directions, offering theoretical references for deepening the understanding of the molecular mechanisms by which mechanical microenvironments regulate white blood cell function and disease progression and for developing novel therapeutic strategies.
Keywords:
Piezo1
; blood cells
; mechanosensing
; immune regulation
; signal transduction
1. Introduction
Tissue microenvironment remodeling in disease states is a multidimensional process [1]. Beyond classical biochemical signals such as inflammatory cytokines and chemokines, biophysical cues have emerged as another critical dimension for understanding disease mechanisms [2]. These cues notably include matrix stiffness, fluid shear stress, and interstitial pressure [3]. Such as in solid tumors, uncontrolled proliferation of cancer cells and resident fibroblasts drives extensive extracellular matrix deposition and crosslinking [4]. In fibrotic diseases, persistent fibroblast activation drives excessive collagen deposition. This leads to a marked increase in extracellular matrix stiffness [5]. And in cardiovascular disease, altered hemodynamics generate aberrant shear stress along the vessel wall [6]. These mechanical cues profoundly influence immune cell migration, activation, polarization, and effector functions [7]. Among the mechanosensitive molecules, Piezo1 has emerged as a key ion channel that directly senses mechanical stimuli and transduces them into intracellular Ca2+ signals, thereby regulating a wide range of immune cell behaviors [8].
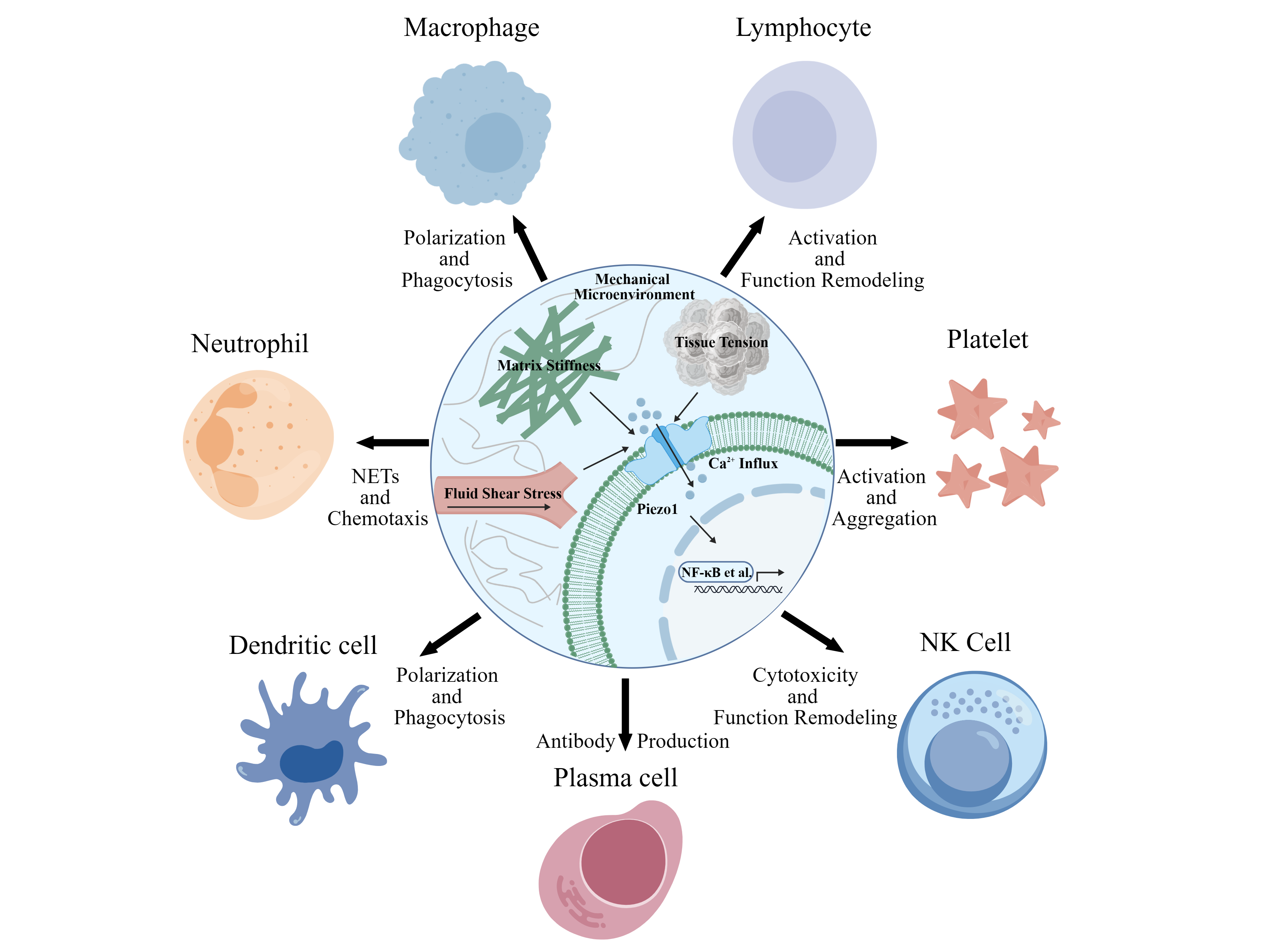
Piezo1 is widely expressed in diverse blood cell lineages [9]. In neutrophils, Piezo1 contributes to shear-induced neutrophil extracellular trap formation (NETosis) and facilitates the process of trans-endothelial migration [10]. Within macrophages, this mechanosensitive channel drives the polarization toward M1 or M2 phenotypes under different matrix stiffness conditions [11]. In platelets Piezo1 was shown to regulate platelet rolling and adhesion to the growing thrombus [12], also it modulates Ca2+-dependent activation and thrombosis [13]. Besides, Piezo1 regulate megakaryocyte mechanosensing, thus influencing platelet production [14]. Piezo1 pathological activation was recently shown to cause thromboembolic events in patients with hereditary xerocytosis [15], thus predicting its implication in pathological thrombosis. In both T and B lymphocytes, it shapes cellular activation, differentiation programs, and antibody class switching processes [16,17]. Moreover, emerging evidence indicates that Piezo1 also functions in natural killer cells (NKs), dendritic cells (DCs), and plasma cells, suggesting conserved mechanotransduction mechanisms across both innate and adaptive immunity (Figure 1) [18,19].
This review offers a systematic summary of current knowledge regarding Piezo1-mediated mechanotransduction across various blood cell types. We place particular emphasis on neutrophils, macrophages, platelets, and T and B lymphocytes. Available findings on NK cells, dendritic cells, and plasma cells are also briefly consolidated. Beyond describing individual cell responses, we examine how Piezo1 helps coordinate interactions among different immune populations within the mechanical microenvironment. This coordination gives rise to a multi-cellular mechano-immunological network, one that can either restrain or accelerate tumor progression depending on context. We evaluate the potential of Piezo1 as a novel mechano immunotherapeutic target, highlight the challenges for clinical translation, and propose future research directions. This review aims to provide a theoretical framework for understanding how the mechanical factors regulate influence disease via blood cell modulation and to inspire new therapeutic strategies.
2. Mechanoresponsive and Functional Regulatory Roles of Piezo1 in Major White Blood Cells
2.1. Piezo1-Mediated Regulation of Neutrophil Activation and Functional Reprogramming
Neutrophils, as core constituents of innate immunity, represent the earliest infiltrating immune cells within the mechanical microenvironment, and their activation status, functional output, and survival duration directly influence inflammatory homeostasis and tumor progression dynamics [20,21]. Mechanical microenvironment such as tumor microenvironment contains multiple mechanical cues. These cues include interstitial fluid shear stress, blood flow shear stress, abnormally increased matrix stiffness, changed tissue tension, and intercellular mechanical contacts. Neutrophils can accurately detect these mechanical cues through the mechanosensitive ion channel Piezo1. This detection process further triggers a series of intracellular signal transduction cascades. These cascades can comprehensively regulate multiple biological processes of neutrophils. The processes include neutrophil activation, adhesion, migration, effector functions, and apoptosis [17,22]. While Piezo1-mediated Ca2+ influx serves as the central hub for mechanical force-regulated neutrophil functional modulation [8], Piezo1 also exhibits multifaceted regulatory mechanisms in this process. While Piezo1-mediated Ca2+ influx serves as the central hub for mechanical force-regulated neutrophil functional modulation, this mechanosensitive ion channel also operates through multifaceted regulatory mechanisms [23]. Under varying levels of shear stress (4–80 dyne/cm2), Ca2+ influx through Piezo1 activates the calcium-dependent calpain protease pathway, triggering cytoskeletal reorganization and thereby promoting canonical neutrophil extracellular trap formation (NETosis). The resulting NETs function as a scaffold for platelet adhesion and aggregation, generating a self-amplifying feedback loop of NET-mediated platelet recruitment and platelet-driven NETosis that propagates immunothrombosis [10,24,25]. In an atherosclerosis model, low shear stress (5 dyne/cm2) at arterial bifurcations and other sites leads to downregulation of Piezo1 expression. This, in turn, induces NETs through the histone deacetylase 2 (HDAC2)–reactive oxygen species (ROS) axis, promoting endothelial cell injury and plaque formation [26]. Furthermore, in an influenza virus infection model, mechanical signals were shown to integrate magnesium ion signaling via Piezo1, thereby promoting neutrophil NETs formation through the Sirtuin 2 (SIRT2)-hypoxia-inducible factor-1α (HIF-1α) pathway. Subsequent uptake of NET-derived DNA by macrophages drives classical macrophage activation (M1 polarization) through the Toll-like receptor 9 (TLR9) pathway. This process also involves the cyclic GMP-AMP synthase and stimulator of interferon genes (cGAS-STING) pathway, further amplifying local inflammatory responses [22].
Piezo1 is also deeply involved in the regulation of neutrophil migration, adhesion, and functional reprogramming [27,28]. During transendothelial migration, plasma membrane tension activates Piezo1 as neutrophils traverse interendothelial gaps, eliciting transient Ca2+ influx. This calcium signal subsequently stabilizes hypoxia-inducible factor-1α (HIF-1α) and upregulates nicotinamide adenine dinucleotide phosphate oxidase 4 (NOX4), thereby markedly enhancing reactive oxygen species (ROS) production and bactericidal capacity [29,30]. This mechanically driven functional activation mechanism indicates that neutrophil effector functions are already enhanced during the process of transendothelial migration, representing an efficient pre-adaptive strategy [23]. In pulmonary microvessels, capillaries with diameters less than 5 μm impose physical constraints on neutrophils, inducing repetitive cellular deformation that activates Piezo1 and reprograms their transcriptome via the extracellular signal-regulated kinase (ERK)/nuclear factor-kappa B (NF-κB) pathway. This mechanotransduction event confers pro-angiogenic functionality upon neutrophils, upregulating effector molecules including vascular endothelial growth factor A (VEGFA), interleukin-1 beta (IL-1β), and C-X-C motif chemokine ligand 1/2 (CXCL1/2), while simultaneously modulating their retention time and migratory behavior within pulmonary vasculature [31].
Under pathological mechanical microenvironments, Piezo1 exerts more significant regulatory effects on neutrophils. In an ankylosing spondylitis model, mechanical strain initiates a multistep cascade by first acting upon syndecan-1–positive (SDC1+) sheath fibroblasts, driving their differentiation through SOX5-mediated transcriptional regulation. These differentiated fibroblasts subsequently secrete high levels of C-X-C motif chemokine ligand 5 (CXCL5), which recruits C-X-C chemokine receptor type 4–positive (CXCR4+) neutrophils and promotes their NETosis, ultimately exacerbating enthesitis and new bone formation. This reveals that mechanical forces can remotely regulate neutrophil function through an indirect crosstalk mechanism between stromal cells and neutrophils [32]. From the hematopoietic origin, upon inflammation-induced mobilization of hematopoietic stem cells (HSCs) into circulation, blood flow shear stress activates two parallel pathways through Piezo1. The junctional adhesion molecule 3 (JAM3) pathway regulating cell homing and transendothelial migration, and the calpain 2 (CAPN2) pathway driving myeloid-biased differentiation (Figure 2). This dual-pathway activation promotes neutrophil generation at the source level and accelerates inflammatory aging processes [33]. Collectively, these findings demonstrate that therapeutic strategies targeting Piezo1 hold promise for disrupting this mechanically-immunologically coupled vicious cycle, offering novel targets for tumor immunotherapy [34].
2.2. Piezo1-Dependent Mechanical Signal Regulation of Macrophage Polarization and Inflammatory Phenotype Specialization
Macrophages are key immune cells that participate in immune regulation in the tissue mechanical microenvironment.Their polarization direction and inflammatory phenotype directly affect immune surveillance.They also directly affect the remodeling of the mechanical microenvironment. Extensive studies have confirmed that mechanical cues in the mechanical microenvironment, including matrix stiffness, fluid shear stress, and tissue tension are transduced into intracellular biochemical signals via the mechanosensitive ion channel Piezo1, thereby regulating macrophage polarization toward either the M1 phenotype or the alternatively activated macrophage (M2) phenotype, ultimately determining their inflammatory and functional outputs [11,35,36]. Under matrix stiffness stimulation, Piezo1 mediates Ca2+ influx and activates Yes-associated protein/transcriptional coactivator with PDZ-binding motif (YAP/TAZ) nuclear translocation, thereby directly regulating the expression of macrophage polarization-related genes and completing phenotypic switching [37]. Simultaneously, under conditions of appropriate mechanical stretching intensity, Piezo1-mediated M2 polarization further promotes the secretion and activation of transforming growth factor-beta 1 (TGF-β1), thereby promoting osteogenic differentiation and bone formation of bone marrow mesenchymal stem cells (BMSCs) [38]. In a sepsis-induced myocardial injury model, macrophage-specific Piezo1 knockout activates the phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT) signaling pathway and upregulates secreted phosphoprotein 1 (SPP1) expression, driving macrophage polarization toward the anti-inflammatory M2 phenotype, significantly reducing myocardial inflammation levels and improving cardiac function [39]. Additionally, fluid shear stress can directly activate the Piezo1 channel, promoting inflammasome activation through the Ca2+–NOD-like receptor family pyrin domain-containing 3 (NLRP3)–cysteine-aspartic acid protease-1 (Caspase-1) signaling axis, thereby enhancing the inflammatory response capacity of macrophages [40].
In pathological mechanical microenvironments, Piezo1 plays a particularly critical regulatory role in macrophages.Fibrotic tissues can produce sustained mechanical tension.This tension activates Piezo1 in macrophages.Activated Piezo1 further promotes the activation of hepatic stellate cells.This process relies on inflammatory responses and paracrine signaling mediated by macrophages.It ultimately aggravates the progression of tissue fibrosis.In contrast, specific knockout of Piezo1 in macrophages can significantly alleviate tissue damage and inflammatory reactions [41]. The elastic mechanical signaling of nanomaterials can similarly reprogram the phenotype of tumor-associated macrophages via Piezo1, thereby ameliorating the local immunosuppressive state within the mechanical microenvironment [42]. Intercellular mechanical contact can couple and activate Piezo1 through αvβ3 integrin, thereby amplifying profibrotic signaling [43]. Extrinsic mechanical stimuli, such as ultrasound and mechanical vibration, can also directionally regulate macrophage polarization via Piezo1, participating in tissue repair and osteogenic activities [44,45]. During atherosclerotic plaque progression, the abnormally stiffened mechanical microenvironment resulting from significantly increased vascular wall matrix stiffness can activate macrophage Piezo1, promoting inflammasome activation through the Ca2+-NF-κB signaling axis and driving macrophage polarization toward the pro-inflammatory M1 phenotype, thereby accelerating plaque destabilization and rupture [11]. Furthermore, during the ischemia/reperfusion pathological process of acute kidney injury (AKI), aberrant fluid shear stress resulting from hemodynamic disturbances in peritubular capillaries and localized tissue hypoxia can activate macrophage Piezo1, driving macrophage polarization toward the pro-inflammatory M1 phenotype through the Ca2+-calpain-HIF-1α/Neurogenic locus notch homolog protein (Notch) signaling axis, thereby exacerbating renal inflammatory injury and the fibrotic process (Figure 3) [46].
In summary, Piezo1 serves as a critical mechanosensor in macrophages for perceiving diverse pathological mechanical signals. By mediating distinct intracellular signaling axes, it orchestrates macrophage polarization and functional phenotypes, playing a central regulatory role in various pathological processes including tumorigenesis, fibrosis, atherosclerosis, and acute kidney injury. Piezo1 thus represents a pivotal molecular hub linking the mechanical microenvironment to macrophage-mediated inflammation and tissue injury.
2.3. Piezo1-Dependent Mechanical Signal Regulation of Platelet Mechanosensing and Coagulation Function
Platelets are a vital type of blood cell. They are involved in coagulation, thrombosis, and immune regulation. When carrying out these functions, platelets rely heavily on the precise sensing of mechanical cues. These cues come from the tissue mechanical microenvironment, including tumor microenvironment. Starting with the study of Ilkan et al. in 2017, which demonstrated Piezo1 functioning in platelets, as well as in the megakaryocytes [12], it was proposed that Piezo1 serves as the core platelet mechanosenser [13]. However, a platelet’s capabilities of direct membrane tension sensing are limited due to its small size and discoid shape. The recent review of Mammadova-Bach et al. [47] proposes platelet main glycoproteins, GPIb and GPIIbIIIa (integrin αIIbβ3) as the shear stress sensors. On the other hand, plasma membrane swelling was reported to activate Piezo1 [48], therefore it is possible, that rolling platelets “sense” shear stress by their swelling. For platelets and red blood cells the impact of Piezo1 is elevated by its direct coupling with the scramblase transmembrane protein 16F (TMEM16F) [49,50], thus promoting phosphatidylserine (PS) exposure by these cells, followed by the induction of blood coagulation [51].
Mechanical stimuli prevalent in the tumor microenvironment, including elevated matrix stiffness [52], aberrant blood flow shear stress [53], and mechanical contact triggered by vascular endothelial injury [54], can directly activate the Piezo1 channel on the platelet surface, thereby mediating Ca2+ ion influx [12]. This calcium signal triggers a cascade amplification reaction intracellularly. On one hand, it activates downstream signaling pathways including the Calpain-Talin1 axis [13,55] and Ras homolog family member A (RhoA)/rho-associated coiled-coil containing protein kinase ROCK [56]. thereby driving cytoskeletal reorganization and enhancing integrin αIIbβ3 activation, which significantly augments platelet adhesion to vascular endothelium, tumor cells, and the extracellular matrix, thus establishing the foundation for platelet aggregation and microthrombus formation [57]. Additionally, Piezo1-mediated Ca2+ influx facilitates platelet degranulation, promoting the release of bioactive molecules including platelet-derived growth factor (PDGF), TGF-β, and platelet factor 4 (PF4). These molecules promote tumor cell proliferation, invasion, and angiogenesis, while concurrently recruiting immune cells such as macrophages and neutrophils, thereby fostering a pro-tumorigenic microenvironment [58]. Concurrently, Piezo1 inhibition diminishes heterotypic adhesion between tumor cells and platelets, suggesting that Piezo1 serves as a pivotal regulatory molecule linking platelet mechanosensation, coagulation function, and tumor progression [59].
In the pathological progression of tumors, Piezo1-mediated mechanosensation and functional dysregulation of platelets constitute critical regulatory factors for cancer-associated thrombosis and metastasis. Elevated shear stress within the tumor stroma activates Piezo1, thereby facilitating platelet hyperactivation and inducing aberrant platelet aggregation to form tumor-associated microthrombi. These microthrombi may occlude tumor vasculature, resulting in hypoxia and metabolic dysregulation that can further drive malignant progression [60,61]. Simultaneously, activated platelets and their secreted factors can coat circulating tumor cells (CTCs), thereby attenuating immune surveillance and establishing a protective shield for tumor cell distant colonization (Figure 4) [62].
The process of platelet formation in the bone marrow, called megakaryopoiesis, is known to be crucially dependent on the mechanistic stimuli, and thus could be Piezo1-dependent. Indeed, in the early steps Piezo1 was shown to have an inhibitory effect [63]. While it was recently demonstrated, that pharmacological Piezo1 stimulation promotes pro-platelet formation at the later stages of megakaryopoiesis [14]. Upon induction of Piezo1 in this model a phenotype resembling myelofibrosis occurred, and knockout of janus kinase 2 (JAK2) prevented this, thus suggesting possible role of Piezo1 in hematological malignancies.
In addition to its direct effects, Piezo1 serves as an important mediator in the intercellular communication between platelets and neutrophils [10]. Activated platelets significantly promote the recruitment and local adhesion of neutrophils within the microenvironment through the release of soluble mediators and surface interactions [64]. Upon adhesion to platelets and damaged extracellular matrix via integrins, neutrophils undergo cytoskeletal remodeling and generate intracellular mechanical tension. These mechanical signals are sensed by mechanosensitive ion channels such as Piezo1, leading to Ca2+ ion influx. Elevated intracellular Ca2+ levels facilitate neutrophil activation and promote the formation of NETs. As a reciprocal regulatory mechanism, the reticulated NETs are capable of capturing and intercepting circulating platelets, providing a natural scaffold for the physical reinforcement of microthrombi [65]. This pathological positive feedback loop of platelet activation, NETs release, and thrombus stabilization continuously amplifies local pro-tumorigenic inflammatory signals. It also establishes a protective barrier for the survival and targeted colonization of circulating tumor cells, ultimately exacerbating the risk of tumor metastasis [66].
2.4. Piezo1-Dependent Mechanical Signal Regulation of Lymphocyte Immune Activation and Differentiation
Lymphocytes, as the core components of adaptive immunity, undergo regulation of their activation, proliferation, differentiation, and functional execution by mechanical signals within the mechanical microenvironment. Piezo1 serves as a critical mechanotransducer. It plays a central role in lymphocyte mechanosignaling. It directly participates in their immune activation and phenotypic differentiation. Through these actions, Piezo1 modulates the magnitude and direction of anti-tumor immune responses [17]. Piezo1 can precisely sense mechanical signals within the tissue remoding microenvironment and regulate lymphocyte functions by mediating Ca2+ influx and activating downstream signaling pathways. In T lymphocytes, upon activation by mechanical stimuli, Piezo1 mediates early and rapid Ca2+ influx, primarily modulating rapid cytoskeletal responses such as microtubule-organizing center (MTOC) reorientation [67]. In addition, chronic mechanical stress induces T cell exhaustion through Piezo1 [68], whereas inhibition of Piezo1 enhances their cytotoxic capacity [69]. The activation and antibody secretion of B lymphocytes are likewise regulated by Piezo1. The response of B cells to membrane-bound antigens is entirely dependent on the mechanotransduction function of Piezo1. When Piezo1 is inhibited or knocked down, B cells fail to effectively form immune synapses, undergo Ca2+ influx and execute proliferation and antibody secretion [16]. The Piezo1 signaling pathway exhibits specificity in regulating B cell antibody class switching. Studies have revealed that Piezo1 activation selectively enhances TGF-β1–induced Smad3 phosphorylation, thereby specifically promoting IgA class switching and secretion, which holds significant implications for mucosal immunity [70]. During chemokine-induced T cell migration, Piezo1 serves as a critical component of the “outside-in” signal transduction machinery. After chemokine receptor activation and, closely following the activation of focal adhesion kinase, Piezo1 senses increased membrane tension and mediates the recruitment of the integrin lymphocyte function-associated antigen-1 (LFA-1) at the leading edge of the cell, thereby driving efficient chemotactic movement [71].
MTOC, microtubule organizing center; TIM-3,T-cell immunoglobulin and mucin domain-containing protein 3; PD-1,Programmed cell death protein 1; CD4+ T-cell, cluster of differentiation 4-positive T cell; Treg, regulatory T cell; Th1, T helper type 1 cell; Th17, T helper type 17 cell; IFN-γ, interferon-gamma; IL-17, interleukin-17; BCR, B cell receptor; TGF-β1, Transforming Growth Factor-beta 1;SMAD3, Mothers Against Decapentaplegic Homolog 3;IgA, Immunoglobulin A.
Different lymphocyte subtypes exhibit distinct responses to mechanical signals, and Piezo1 maintains immune homeostasis through precise regulation of these differential responses. In CD4+ T cells, Piezo1 demonstrates differential regulatory effects on helper T cell subset differentiation. Studies have shown that Piezo1-deficient CD4+ T cells exhibit significantly increased production of interferon-gamma (IFN-γ) and interleukin-17 (IL-17) under in vitro T helper 1 (Th1) and T helper 17 (Th17) polarization conditions, indicating an intrinsic function of Piezo1 in suppressing excessive inflammatory responses [72]. In regulatory T cells (Tregs), Piezo1-mediated mechanical signals influence their suppressive function. Although Piezo1 deficiency does not affect Treg suppressive capacity in vitro, its absence leads to significant expansion of the Treg cell population in chronic inflammatory models such as experimental autoimmune encephalomyelitis, thereby ameliorating disease severity. This suggests that Piezo1 specifically upregulates Treg responses in vivo to maintain immune tolerance [73]. Aberrant Piezo1 activation may disrupt the balance between Treg cells and effector T cells, facilitating tumor immune evasion (Figure 5). Piezo1 functional deficiency directly compromises humoral immune responses, particularly at mucosal barrier defense sites [74]. Both genetic ablation and persistent overactivation of Piezo1 disrupt the coupling between mechanical cues and metabolic rewiring in CAR-T cells, leading to impaired cytotoxic function; in contrast, transient, moderate Piezo1 signaling coordinates synaptic mechanics, calcium transients, and ATP production to maintain peak effector activity [75]. Collectively, these findings establish Piezo1 as a core mediator of lymphocyte mechanotransduction and a critical target for regulating tumor immunity and intervening in immune evasion.
2.5. Piezo1 and Other White Blood Cells
In addition to macrophages, NKs, DCs, and plasma cells can also sense mechanical cues in the mechanical microenvironment via the Piezo1 channel, thereby completing functional specialization and immune effector output. As critical cytotoxic cells of innate immunity, NKs recognize matrix stiffness and target cell mechanical properties through Piezo1, which mediates Ca2+ influx and regulates immunological synapse assembly. Increased matrix stiffness significantly enhances NK cytotoxic activity, while Piezo1 inhibition markedly impairs their migration and killing capacity within solid tissues [18]. Dendritic cell function is similarly regulated by mechanical signals. Studies have shown that increased matrix stiffness has a specific effect. It promotes the polarization of dendritic cells toward a pro-inflammatory phenotype. This shift enhances their maturation. It also boosts their antigen presentation and phagocytic capabilities [76]. Matrix stiffness and inflammatory signals activate Piezo1 in DCs, triggering Ca2+ influx and influencing cytokine secretion patterns through the Piezo1–Ca2+–nuclear factor of activated T cells (NFAT) pathway, thereby regulating the Th1/Treg balance and participating in the construction of the tumor immune microenvironment [19]. Under non-tumor, ous or acute infection conditions, Piezo1 activation may enhance DC type I interferon production, thereby activating NKs and cytotoxic T lymphocytes (CTLs), demonstrating the context-dependent nature of its function [77]. B cells and plasma cells can sense plasma membrane tension and matrix mechanical alterations via Piezo1, regulating TGF-β1-mediated IgA class switching and influencing antibody synthesis and secretion efficiency. Piezo1 functional deficiency directly compromises systemic humoral immune response levels [70,77]. Beyond direct regulation of immune cell function through calcium ion signaling, Piezo1 can also synergize with integrin signaling pathways to transduce extracellular matrix mechanical stimuli into intracellular biochemical signals [78]. Studies have shown that Piezo1 activation or deficiency in DCs affects their interaction with the extracellular matrix (ECM), thereby modulating immune cell migration and antigen-specific T cell activation [19]. This synergistic signaling mechanism plays a critical role in mediating inflammatory responses during both physiological and pathological processes [79]. As a central hub, Piezo1 integrates mechanical signals to coordinate immune cell migration, synapse formation, and effector molecule release, thereby playing a pivotal role in the coordinated response between innate and adaptive immunity [80].
3. Integrated Effects of Piezo1-Mediated Blood Cell Mechanotransduction in Disease Mechanical Microenvironments
3.1. Interactive Regulation Between Disease Mechanical Microenvironment Mechanical Signals and Blood Cell Functions
The mechanical characteristics of the mechanical microenvironment are critical physical signals. They regulate blood cell function. Solid tumors universally show mechanical alterations. These alterations include elevated matrix stiffness. They also include enhanced interstitial fluid shear stress. In addition, they include dysregulated tissue tension [81,82], These mechanical cues are sensed by tumor-infiltrating immune cells via the mechanosensitive ion channel Piezo1 and subsequently transduced into intracellular biochemical signals, thereby regulating immune cell activation, migration, and effector functions [23]. Upon activation by mechanical cues, Piezo1 primarily mediates Ca2+ influx [83], initiating downstream calcium-dependent signaling cascades, including activation of the nuclear factor of activated T cells (NFAT) and Yes-associated protein (YAP) transcriptional axes that broadly regulate the functional phenotypes of neutrophils, platelets, NKs, dendritic cells, and plasma cells, driving immune cell reprogramming toward pro-tumorigenic phenotypes [84]. Concurrently, activated blood cells can further exacerbate matrix stiffening and collagen cross-linking through the release of inflammatory factors, matrix metalloproteinases, and pro-coagulant molecules [85], establishing a positive feedback loop of mechanical abnormality–blood cell activation–tumor progression. Throughout this process, Piezo1 functions as the principal mechanotransducer, serving as the critical molecular nexus that couples mechanical microenvironmental cues with immune regulatory programs.
3.2. Mechanical Synergism Among Different Blood Cells Mediated by Piezo1
The diverse blood cells in the TME do not function as independent mechanical signaling units.Instead, these blood cells form a complex multi-cellular mechano-immunological synergistic network.This network centers on the Piezo1 channel [86]. Individual cells first perform local primary mechanical sensing.This sensing process is gradually amplified into systematic cascading responses.These responses cover both innate immunity and adaptive immunity.
During the early phase of tumor infiltration, mechanical interplay between neutrophils and platelets constitutes the initiating step for pro-inflammatory and pro-metastatic responses, with both cell types acutely sensing aberrant shear stress and vascular matrix stiffness. Upon mechanical stimulation, neutrophils are robustly activated via the Piezo1–Ca2+–calpain signaling pathway, resulting in massive release of NETs [10]. These reticular structures provide a robust physical scaffold for platelet adhesion and aggregation. They also further amplify platelet activation levels, establishing a positive feedback loop of mechanical stimulation, NET release, and the deterioration of the thrombotic microenvironment, which synergistically constructs a pro-inflammatory and pro-metastatic environment. NKs synchronously participating in immune surveillance are similarly highly dependent on this channel [18]. NKs can perceive three-dimensional matrix stiffness and target cell rigidity alterations through Piezo1, transducing these mechanical cues into intrinsic signals that regulate immunological synapse formation and granule exocytosis. Aberrant Piezo1 function or excessive matrix stiffening significantly impairs NK cell infiltration capacity and cytotoxic killing efficiency, enabling tumors to readily breach innate immune surveillance defenses [87].
At the adaptive immunity level, DCs, as critical antigen-presenting cells, exhibit highly multifaceted responses to microenvironmental mechanical cues. On one hand, DCs can perceive matrix stiffness through activation of intracellular YAP/TAZ signaling pathways, thereby upregulating glycolytic metabolism and promoting the secretion of pro-inflammatory factors such as tumor necrosis factor (TNF) and interleukin-6 (IL-6) [76]. However, within the complex mechanical microenvironment, the direction of T cell polarization guided by DCs is likewise under the surveillance of the core mechanosensory channel Piezo1. High-stiffness microenvironmental stimulation of DCs triggers Piezo1-mediated Ca2+ inducing the differentiation ofinflux, which not only activates the Ca2+–NFAT pathway but also integrates the Sirtuin 1 (SIRT1)–HIF-1α metabolic reprogramming pathway, thereby modulating the cytokine secretion profile (e.g., suppressing interleukin-12 (IL-12) while promoting TGF-β1 secretion), ultimately priming DCs to promote the differentiation of immunosuppressive Tregs [19]. This interplay and signal integration between distinct mechanosensors (YAP/TAZ and Piezo1) ultimately determines the trajectory of local adaptive immunity within tumors. Furthermore, when B cells and plasma cells sense antigen-presenting cell membrane tension and lymph node matrix mechanical signals, Piezo1 may participates in regulating their antibody class switching and secretion efficiency, constituting an important complement to the mechanical regulation of humoral immunity [70].
Piezo1 on the macrophage surface acts as a critical mechano-immunological switch that determines phenotypic polarization, determining phenotypic polarization. Under general mechanoresponsive conditions, high-stiffness matrices typically activate the Piezo1–YAP axis and drive macrophage polarization toward the classically activated M1 phenotype [37]. However, during tumor progression, microenvironmental signals may induce downregulation or functional suppression or functional suppression of Piezo1 expression on tumor-associated macrophage (TAM) surfaces to evade immune surveillance. This inactivation of the core mechanosensor blocks normal pro-inflammatory signal transduction, thereby promoting macrophage polarization toward the pro-tumorigenic alternatively activated M2 phenotype [39,88]. These M2-polarized TAMs subsequently secrete substantial anti-inflammatory factors and exosomes, directly suppressing local CD8+ T cell cytotoxic function, thereby constructing a deeply immunosuppressive microenvironment [89,90].
Piezo1 is not merely the molecular basis for individual blood cells to perceive mechanical signals but also serves as an important hub mediating collaborative interactions among multiple blood cell types. By transducing mechanical signals across cells and amplifying immune effects, it regulates tumor inflammation, immune suppression, and malignant progression, highlighting its potential as a theoretical foundations for immunotherapeutic strategies targeting the tumor mechanical microenvironment.
4. Potential and Challenges of Piezo1 as a Therapeutic Target in Diseases Associated with Tissue Stiffening
4.1. Therapeutic Strategies Targeting Piezo1-Mediated Blood Cell Functional Regulation
Piezo1 plays a critical mediating role between the mechanical microenvironment and immune cell function. Based on this role, targeted Piezo1 modulation has emerged as a potential therapeutic strategy. This strategy aims to reshape the mechanical microenvironment. It also aims to block immune suppression mediated by tissue abnormalities. Small-molecule agonists or inhibitors can precisely regulate Piezo1 activity. This precise regulation can reverse the aberrant shaping of immune cells by mechanical cues in various mechanical microenvironments. In macrophage-targeted interventions, Piezo1 inhibition can block matrix stiffness-induced alternatively activated M2 polarization, reduce pro-inflammatory factor release, and simultaneously enhance pro-inflammatory macrophage antigen presentation and inflammatory clearance capacities, thereby dismantling the immunosuppressive environment caused by tissue matrix alterations [91]. For NKs and cytotoxic T lymphocytes, moderate Piezo1 activation can enhance their infiltration efficiency and cytotoxic activity within dense tumor matrices, strengthening immune surveillance against solid tumors [18]. Concurrently, Piezo1 targeting can also regulate aberrant neutrophil and platelet activation, reduce NETosis and thrombosis-associated microenvironment establishment, and decrease tumor invasion and distant metastasis risk. Piezo1-targeted therapy has a core advantage. It can precisely interface with the mechanical characteristics of the mechanical microenvironment. Through this interface, it achieves synergistic regulation of multiple blood cell functions. Conventional immunotherapeutic approaches cannot offer this unique value. Combination of immune checkpoint inhibitors with Piezo1 modulators holds promise for further enhancing immunotherapeutic response rates and overcoming mechanically-mediated therapeutic resistance.
4.2. Challenges and Optimization Directions in Clinical Translation
Piezo1 plays a key role in regulating the mechanical microenvironment and modulating blood cell function, making it a promising target for diseases associated with tissue stiffening and fibrosis. However, its clinical application presents several challenges. Piezo1-targeted therapy offers a unique advantage by interacting with the mechanical properties of various diseased tissues and regulating multiple blood cell functions. This approach differs from traditional therapies that typically target a single cell type. Piezo1 modulators can influence various blood cells, including macrophages, neutrophils, lymphocytes, and platelets, enhancing immune activation and inhibiting factors such as M2 polarization and platelet activation. This creates a multi-dimensional therapeutic effect by addressing inflammation, immune activation, and tissue remodeling.
However, Piezo1’s widespread presence in normal tissues, such as vascular endothelium and neural cells, raises safety concerns, as non-specific inhibition can lead to off-target effects in these systems. Additionally, Piezo1’s function varies across different cell types, with pro-tumorigenic effects in some immune cells and enhanced anti-tumor responses in others. Therefore, precise regulation including optimal dosing, timing, and combination strategies is necessary to avoid therapeutic failure.
The heterogeneity of mechanical microenvironments, such as variations in matrix stiffness, hydrostatic pressure, and shear stress across different diseases, complicates the prediction of Piezo1-mediated effects. Future research should focus on developing immune cell-specific delivery systems, individualized dosing protocols, and allosteric modulators with reduced side effects. Combining mechanical microenvironment interventions with other therapeutic approaches, such as immunotherapy or anti-fibrotic treatments, may further enhance efficacy.
In conclusion, Piezo1 offers a novel approach to mechano-immunotherapy in diseases associated with tissue stiffening and fibrosis. However, challenges related to specificity, safety, and tissue heterogeneity must be addressed for effective clinical translation. Intensive investigation into these issues is crucial for realizing Piezo1’s potential as a therapeutic target.
5. Summary and Perspectives
This review summarizes the current understanding of Piezo1, a mechanosensitive ion channel crucial for mechanotransduction in blood cells. It focuses on its role in various immune cells such as neutrophils, macrophages, platelets, lymphocytes, NK cells, dendritic cells, and plasma cells, examining how Piezo1 regulates their functions within the mechanical microenvironment. Piezo1 plays a pivotal role in converting mechanical signals (e.g., matrix stiffness, shear stress, and tissue tension) into biochemical signals via ca2+ influx, which influences cell behavior including activation, proliferation, differentiation, migration, and effector functions. This allows Piezo1 to link the mechanical microenvironment to systemic immune responses, acting as a core molecular hub.
In neutrophils, Piezo1 triggers NETs formation, pro-inflammatory factor release, and migration via calpain protease and NF-κB signaling, aggravating inflammation and metastasis. In macrophages, it regulates M1/M2 polarization and inflammatory phenotypes through YAP/TAZ, NLRP3-Caspase-1 pathways, contributing to an immunosuppressive tumor microenvironment. In platelets, Piezo1 modulates adhesion, aggregation, and thrombosis, promoting tumor-associated thrombogenesis. In lymphocytes, it affects T cell activation, B cell antibody secretion, and Treg function, helping maintain immune homeostasis and regulating tumor immune evasion. Piezo1’s role extends to NKs, DCs, and plasma cells, where it integrates mechanical cues to fine tune innate and adaptive immunity.
Piezo1-mediated mechanotransduction in the tissue mechanical microenvironment operates within a multicellular network, where blood cells interact to form feedback loops that accelerate tumor growth and immune modulation. These interactions reshape the mechanical and immune landscapes of tumors, enhancing malignant progression. Thus, Piezo1 emerges as a promising therapeutic target for mechano-immunotherapy, where small-molecule modulators could restore anti-tumor immune responses and complement immune checkpoint inhibitors for improved treatment outcomes.
However, Piezo1’s clinical translation faces challenges, primarily due to its widespread expression in normal tissues, which may lead to off-target effects in the cardiovascular, hematopoietic, and nervous systems. Furthermore, its role in immune cells and tumors can be paradoxical, suppressing tumor-promoting activity in immune cells while supporting tumor growth. Variability in tumor mechanical properties across types and stages complicates Piezo1’s immunoregulatory effects. Future research should focus on targeted delivery systems for immune cells, selective modulators to reduce off-target effects, and tailoring treatments based on tumor type and disease stage. Additionally, understanding how Piezo1 functions across different cell types and its synergy with other therapies will be essential for its clinical application.
As research deepens, breakthroughs in Piezo1-targeted interventions may open new avenues for the treatment of various diseases. These interventions hold promising mechano immunological strategies, particularly with significant potential in addressing tissue stiffness and fibrosis related diseases.
Author Contributions
Conceptualization, J.L.; methodology, J.L.; investigation, J.H.; data curation, L.L. and D.L.; writing-original draft preparation, J.L. and J.H.; writing, review and editing, Panteleev MA., Sveshnikova AN. and Q.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (31870928 and 32271360), the Natural Science Foundation of Guangdong Province, China (2021A1515010040 and 2023A1515010829).
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
Thank all collaborators for their contributions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mayorca-Guiliani, A. E.; et al. ECM formation and degradation during fibrosis, repair, and regeneration of the article. Npj Metab. Health Dis. 2025, 3, 25. [Google Scholar] [CrossRef]
- Tiskratok, W.; et al. Extracellular matrix stiffness: mechanotransduction and mechanobiological response-driven strategies for biomedical applications targeting fibroblast inflammation of the article. Polymers 2025, 17, 822. [Google Scholar] [CrossRef]
- Ai, J.; et al. Mechanical microenvironment in tumor immune evasion: bidirectional regulation between matrix stiffness and immune cells and its therapeutic implications of the article. Int. J. Biol. Sci. 2026, 22, 280–307. [Google Scholar] [CrossRef]
- Levental, K. R.; et al. Matrix crosslinking forces tumor progression by enhancing integrin signaling of the article. Cell 2009, 139, 891–906. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T. A.; et al. Mechanisms of fibrosis: therapeutic translation for fibrotic disease of the article. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef]
- Paliwal, N.; et al. Presence of left atrial fibrosis may contribute to aberrant hemodynamics and increased risk of stroke in atrial fibrillation patients of the article. Front. Physiol. 2021, 12, 657452. [Google Scholar] [CrossRef]
- Yan, R.; et al. Mechanotransduction in shaping immunity: pathways, crosstalk, and pathophysiological relevance of the article. Adv. Sci. 2025, 12, e12164. [Google Scholar] [CrossRef]
- Pirri, C. PIEZO channels in mechano-inflammation: gatekeepers of neuroimmune crosstalk of the article. Diseases 2025, 13, 263. [Google Scholar] [CrossRef] [PubMed]
- Karkempetzaki, A. I.; et al. Piezo1 and its function in different blood cell lineages of the article. Cells 2024, 13, 482. [Google Scholar] [CrossRef]
- Baratchi, S.; et al. Piezo1 expression in neutrophils regulates shear-induced NETosis of the article. Nat. Commun. 2024, 15, 7023. [Google Scholar] [CrossRef] [PubMed]
- Atcha, H.; et al. Mechanically activated ion channel Piezo1 modulates macrophage polarization and stiffness sensing of the article. Nat. Commun. 2021, 12, 3256. [Google Scholar] [CrossRef]
- Ilkan, Z.; et al. Evidence for shear-mediated Ca2+ entry through mechanosensitive cation channels in human platelets and a megakaryocytic cell line of the article. J. Biol. Chem. 2017, 292, 9204–9217. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; et al. Involvement of Piezo 1 in inhibition of shear-induced platelet activation and arterial thrombosis by ginsenoside Rb1 of the article. Br. J. Pharmacol. 2025, 182, 1957–1974. [Google Scholar] [CrossRef] [PubMed]
- Demagny, J.; et al. Role of the mechanotransductor PIEZO1 in megakaryocyte differentiation of the article. J. Cell. Mol. Med. 2024, 28, e70055. [Google Scholar] [CrossRef] [PubMed]
- Bachelot-Loza, C.; et al. Piezo1 gain of function induces a platelet preactivation state of the article. J. Thromb. Haemost. JTH 2026, 24, 1118–1129. [Google Scholar] [CrossRef]
- Kwak, K.; et al. B cell responses to membrane-presented antigens require the function of the mechanosensitive cation channel Piezo1 of the article. Sci. Signal. 2023, 16, eabq5096. [Google Scholar] [CrossRef]
- Qu, P.; et al. The dual role of Piezo1 in tumor cells and immune cells: a new target for cancer therapy of the article. Front. Immunol. 2025, 16, 1635388. [Google Scholar] [CrossRef]
- Yanamandra, A. K.; et al. PIEZO1-mediated mechanosensing governs NK-cell killing efficiency and infiltration in three-dimensional matrices of the article. Eur. J. Immunol. 2024, 54, e2350693. [Google Scholar] [CrossRef]
- Wang, Y.; et al. Dendritic cell Piezo1 directs the differentiation of TH1 and Treg cells in cancer of the article. eLife 2022, 11, e79957. [Google Scholar] [CrossRef]
- Wu, L.; et al. Neutrophils in the tumor microenvironment of the article. Adv. Exp. Med. Biol. 2020, 1224, 1–20. [Google Scholar]
- Hou, R.; et al. Tumor-associated neutrophils: critical regulators in cancer progression and therapeutic resistance (Review) of the article. Int. J. Oncol. 2025, 66, 28. [Google Scholar] [CrossRef]
- Wang, Y.; et al. Piezo1-directed neutrophil extracellular traps regulate macrophage differentiation during influenza virus infection of the article. Cell. Death Dis. 2025, 16, 60. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, A.; et al. Trans-endothelial neutrophil migration activates bactericidal function via Piezo1 mechanosensing of the article. Immunity 2024, 57, 52–67.e10. [Google Scholar] [CrossRef] [PubMed]
- Knaus, U. G.; et al. Challenging the role of a NOX4-Piezo1 axis in neutrophil bactericidal function of the article. Immunity 2024, 57, 1716–1718. [Google Scholar] [CrossRef]
- Lightsey, S.; et al. Natural killer cell mechanosensing in solid tumors of the article. Bioengineering 2024, 11, 328. [Google Scholar] [CrossRef]
- Zhu, Y.; et al. Low shear stress exacerbates atherosclerosis by inducing the generation of neutrophil extracellular traps via Piezo1-mediated mechanosensation of the article. Atherosclerosis 2024, 391, 117473. [Google Scholar] [CrossRef]
- Zhao, W.; et al. Mechanotransduction in neutrophil: mechanosensing and immune function regulation of the article. Mechanobiol. Med. 2025, 3, 100157. [Google Scholar] [CrossRef]
- Wang, S.; et al. Mechanosensation by endothelial PIEZO1 is required for leukocyte diapedesis of the article. Blood 2022, 140, 171–183. [Google Scholar] [CrossRef]
- Kim, O. H.; et al. Fluid shear stress facilitates prostate cancer metastasis through Piezo1-Src-YAP axis of the article. Life Sci. 2022, 308, 120936. [Google Scholar] [CrossRef]
- Zhao, F.; et al. Mechanosensitive ion channel PIEZO1 signaling in the hall-marks of cancer: structure and functions of the article. Cancers 2022, 14, 4955. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; et al. PIEZO1 mediates mechanical reprogramming of neutrophils for proangiogenic specialization in the lung of the article. J. Clin. Investig. 2025, 135. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; et al. Mechanical strain-programmed SDC1+ sheath fibroblasts trigger CXCR4hi neutrophil-mediated enthesitis in ankylosing spondylitis of the article. Adv. Sci. 2026, 13, e20617. [Google Scholar] [CrossRef]
- Shang, T.; et al. Shear stress governs hematopoietic stem cell fate to promote inflammation-induced aging of the article. Nat. Aging 2026, 6, 88–107. [Google Scholar] [CrossRef]
- Aykut, B.; et al. Targeting Piezo1 unleashes innate immunity against cancer and infectious disease of the article. Sci. Immunol. 2020, 5, eabb5168. [Google Scholar] [CrossRef]
- Murray, P. J.; et al. Protective and pathogenic functions of macrophage subsets of the article. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Xiong, J.; et al. Matrix stiffness affects tumor-associated macrophage functional polarization and its potential in tumor therapy of the article. J. Transl. Med. 2024, 22, 85. [Google Scholar] [CrossRef]
- Mei, F.; et al. Matrix stiffness regulates macrophage polarisation via the Piezo1-YAP signalling axis of the article. Cell. Prolif. 2024, 57, e13640. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; et al. Piezo1-mediated M2 macrophage mechanotransduction enhances bone formation through secretion and activation of transforming growth factor-β1 of the article. Cell. Prolif. 2023, 56, e13440. [Google Scholar] [CrossRef]
- Zhang, Y.; et al. Piezo1 knockdown activates PI3K/AKT and enhances SPP1 to drive M2 macrophage polarization and reduce cardiac inflammation of the article. Sci. Rep. 2026, 16, 4879. [Google Scholar] [CrossRef]
- Fish, A.; et al. Flow-induced shear stress primes NLRP3 inflammasome activation in macrophages via Piezo1 of the article. ACS Appl. Mater. Interfaces 2024, 16, 4505–4518. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; et al. Piezo1 specific deletion in macrophage protects the progression of liver fibrosis in mice of the article. Theranostics 2023, 13, 5418–5434. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; et al. Nano-mechanical immunoengineering: nanoparticle elasticity reprograms tumor-associated macrophages via Piezo1 of the article. ACS Nano 2024, 18, 21221–21235. [Google Scholar] [CrossRef]
- Ezzo, M.; et al. Acute contact with profibrotic macrophages mechanically activates fibroblasts via αvβ3 integrin-mediated engagement of Piezo1 of the article. Sci. Adv. 2024, 10, eadp4726. [Google Scholar] [CrossRef]
- Jiang, J.; et al. Mobile mechanical signal generator for macrophage polarization of the article. Exploration 2023, 3, 20220147. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; et al. Synergistic effect of ultrasound and reinforced electrical environment by bioinspired periosteum for enhanced osteogenesis via immunomodulation of macrophage polarization through Piezo1 of the article. Mater. Today. Bio 2024, 27, 101147. [Google Scholar] [CrossRef]
- Chen, X.; et al. Piezo1 aggravates ischemia/reperfusion-induced acute kidney injury by Ca2+-dependent calpain/HIF-1α/Notch signaling of the article. Ren. Fail. 2025, 47, 2447801. [Google Scholar] [CrossRef]
- Mammadova-Bach, E.; et al. Platelet mechanotransduction: Regulatory cross talk between mechanosensitive receptors and calcium channels of the article. Arterioscler. Thromb. Vasc. Biol. 2023, 43, 1339–1348. [Google Scholar] [CrossRef]
- Sforna, L.; et al. Piezo1 controls cell volume and migration by modulating swelling-activated chloride current through Ca2+ influx of the article. J. Cell. Physiol. 2022, 237, 1857–1870. [Google Scholar] [CrossRef]
- Liang, P.; et al. Targeting PIEZO1-TMEM16F Coupling to Mitigate Sickle Cell Disease Complications of the article. Am. J. Hematol. 2025, 100, 2261–2275. [Google Scholar] [CrossRef]
- Zhu, W.; et al. PIEZO1 mediates a mechanothrombotic pathway in diabetes of the article. Sci. Transl. Med. 2022, 14, eabk1707. [Google Scholar] [CrossRef] [PubMed]
- Podoplelova, N. A.; et al. Coagulation factors bound to procoagulant platelets concentrate in cap structures to promote clotting of the article. Blood 2016, 128, 1745–1755. [Google Scholar] [CrossRef]
- Zhang, M.; et al. Extracellular matrix stiffness: mechanisms in tumor progression and therapeutic potential in cancer of the article. Exp. Hematol. Oncol. 2025, 14, 54. [Google Scholar] [CrossRef]
- Zhang, C.; et al. Mechanoadaptation via myosin cytoplasmic redistribution protects circulating tumor cells from shear-induced death during hematogenous dissemination of the article. Adv. Sci. 2026, e23112. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C. K.; et al. Biophysical and biochemical roles of shear stress on endothelium: a revisit and new insights of the article. Circ. Res. 2025, 136, 752–772. [Google Scholar] [CrossRef]
- Zhao, W.; et al. Piezo1 initiates platelet hyperreactivity and accelerates thrombosis in hypertension of the article. J. Thromb. Haemost. JTH 2021, 19, 3113–3125. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, M.; et al. Cell surface flip-flop of phosphatidylserine is critical for PIEZO1-mediated myotube formation of the article. Nat. Commun. 2018, 9, 2049. [Google Scholar] [CrossRef] [PubMed]
- Leng, L.; et al. RhoA and the function of platelet integrin alphaIIbbeta3 of the article. Blood 1998, 91, 4206–4215. [Google Scholar] [CrossRef]
- Rab, S. O.; et al. Targeting platelet-tumor cell interactions: a novel approach to cancer therapy of the article. Med. Oncol. 2025, 42, 232. [Google Scholar] [CrossRef]
- Li, Y.; et al. Effects of the interactions between platelets with other cells in tumor growth and progression of the article. Front. Immunol. 2023, 14, 1165989. [Google Scholar]
- Palumbo, J. S.; et al. Platelets and fibrin(ogen) increase metastatic potential by impeding natural killer cell-mediated elimination of tumor cells of the article. Blood 2005, 105, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; et al. Hypoxia stimulates CTC-platelet cluster formation to promote breast cancer metastasis of the article. iScience 2024, 27, 109547. [Google Scholar] [CrossRef]
- Schmied, L.; et al. Platelet-mediated protection of cancer cells from immune surveillance - possible implications for cancer immunotherapy of the article. Front. Immunol. 2021, 12, 640578. [Google Scholar] [CrossRef]
- Abbonante, V.; et al. Newly identified roles for PIEZO1 mechanosensor in controlling normal megakaryocyte development and in primary myelofibrosis of the article. Am. J. Hematol. 2024, 99, 336–349. [Google Scholar] [CrossRef]
- Kim, S. J.; et al. Role of platelets in neutrophil extracellular trap (NET) production and tissue injury of the article. Semin. Immunol. 2016, 28, 546–554. [Google Scholar] [CrossRef]
- Caudrillier, A.; et al. Platelets induce neutrophil extracellular traps in transfusion-related acute lung injury of the article. J. Clin. Investig. 2012, 122, 2661–2671. [Google Scholar] [CrossRef]
- Li, J.; et al. Pathological roles of NETs-platelet synergy in thrombotic diseases: from molecular mechanisms to therapeutic targeting of the article. Int. Immunopharmacol. 2025, 159, 114934. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; et al. T cell polarization and NFAT activation are stiffness dependent and differentially regulated by the channels PIEZO1 and ORAI1 of the article. Sci. Signal. 2026, 19, eadt9566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; et al. Osr2 functions as a biomechanical checkpoint to aggravate CD8+ T cell exhaustion in tumor of the article. Cell 2024, 187, 3409–3426.e3424. [Google Scholar] [CrossRef]
- Pang, R.; et al. PIEZO1 mechanically regulates the antitumour cytotoxicity of T lymphocytes of the article. Nat. Biomed. Eng. 2024, 8, 1162–1176. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; et al. Piezo1 selectively enhances TGF-β1-induced IgA class switching by B cells of the article. Cell. Mol. Life Sci. CMLS 2025, 82, 243. [Google Scholar] [CrossRef]
- Liu, C. S. C.; et al. Piezo1 mechanosensing regulates integrin-dependent chemotactic migration in human T cells of the article. eLife 2024, 12, RP91903. [Google Scholar] [CrossRef] [PubMed]
- Choi, S. H.; et al. Piezo1 restrains proinflammatory response but is essential in T-cell-mediated immunopathology of the article. J. Leukoc. Biol. 2025, 117, qiae242. [Google Scholar] [CrossRef]
- Jairaman, A.; et al. Piezo1 channels restrain regulatory T cells but are dispensable for effector CD4+ T cell responses of the article. Sci. Adv. 2021, 7, eabg5859. [Google Scholar] [CrossRef]
- Chang, J. E.; et al. Mechanosensing by Peyer’s patch stroma regulates lymphocyte migration and mucosal antibody responses of the article. Nat. Immunol. 2019, 20, 1506–1516. [Google Scholar] [CrossRef]
- Abiff, M.; et al. Piezo1 facilitates optimal T cell activation during tumor challenge of the article. Oncoimmunology 2023, 12, 2281179. [Google Scholar] [CrossRef]
- Chakraborty, M.; et al. Mechanical stiffness controls dendritic cell metabolism and function of the article. Cell. Rep. 2021, 34, 108609. [Google Scholar] [CrossRef]
- Pattanayak, S.; et al. Spatiotemporal regulation of ligand trafficking and TLR9 activation by PIEZO1 in human plasmacytoid dendritic cells of the article. Research 2026, 9, 1067. [Google Scholar]
- Friedrich, E. E.; et al. Endothelial cell Piezo1 mediates pressure-induced lung vascular hyperpermeability via disruption of adherens junctions of the article. Proceedings of the National Academy of Sciences of the United States of America 2019, 116, 12980–12985. [Google Scholar] [CrossRef]
- Albarrán-Juárez, J.; et al. Piezo1 and Gq/G11 promote endothelial inflammation depending on flow pattern and integrin activation of the article. J. Exp. Med. 2018, 215, 2655–2672. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; et al. Multimodal regulation of dendritic cells via mineralized vaccines for postsurgical tumor relapse prevention of the article. ACS Nano 2025, 19, 19901–19917. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; et al. Role of the mechanical microenvironment in cancer development and progression of the article. Cancer Biol. Med. 2020, 17, 282–292. [Google Scholar] [CrossRef]
- Bi, Y.; et al. Mechanical models and measurement methods of solid stress in tumors of the article. Appl. Microbiol. Biotechnol. 2024, 108, 363. [Google Scholar] [CrossRef]
- Liu, Y.; et al. Mechanosensitive channel Piezo1 in calcium dynamics: structure, function, and emerging therapeutic strategies of the article. Front. Mol. Biosci. 2025, 12, 1693456. [Google Scholar]
- Deng, J.; et al. Piezo1 regulates actin cytoskeleton remodeling to drive EMT in cervical cancer through the RhoA/ROCK1/PIP2 signaling pathway of the article. Discov. Oncol. 2025, 16, 787. [Google Scholar] [CrossRef] [PubMed]
- Obermann, W. M. J.; et al. Platelets, constant and cooperative companions of sessile and disseminating tumor cells, crucially contribute to the tumor microenvironment of the article. Front. Cell. Dev. Biol. 2021, 9, 674553. [Google Scholar] [CrossRef]
- Cui, X.; et al. Piezo1 as a mechanical checkpoint in T cell immunotherapy for solid tumors of the article. Front. Pharmacol. 2026, 17, 1722027. [Google Scholar] [CrossRef]
- Cui, X.; et al. Mechanotransduction of Piezo1 in the cancer microenvironment: implications for NK cell-based immunotherapy of the article. Front. Oncol. 2025, 15, 1729588. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; et al. Tumor mechanobiology-inspired nanomechanics: Piezo1 mediated tumor-associated macrophages reprogramming and exosome-driven immune amplification of the article. ACS Nano 2025, 19, 41746–41764. [Google Scholar] [CrossRef]
- Xu, J.; et al. Dual roles and therapeutic targeting of tumor-associated macrophages in tumor microenvironments of the article. Signal Transduct. Target. Ther. 2025, 10, 268. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; et al. Tumor-associated macrophages remodel the suppressive tumor immune microenvironment and targeted therapy for immunotherapy of the article. J. Exp. Clin. Cancer Res. CR 2025, 44, 145. [Google Scholar]
- Wang, Y.; et al. Immunoregulatory role of the mechanosensitive ion channel Piezo1 in inflammation and cancer of the article. Molecules 2022, 28, 213. [Google Scholar] [CrossRef]
Figure 1.
Schematic Diagram of Piezo1-Mediated Mechanosensation in Major Blood Cell Types. NETs, neutrophil extracellular traps; NK cell, natural killer cell.
Figure 1.
Schematic Diagram of Piezo1-Mediated Mechanosensation in Major Blood Cell Types. NETs, neutrophil extracellular traps; NK cell, natural killer cell.

Figure 2.
Mechanistic Diagram of Piezo1-Induced Neutrophil Activation and Functional Reprogramming. NETs, neutrophil extracellular traps; NETosis, neutrophil extracellular trap cell death; Calpain, calcium-dependent cysteine protease. ERK, extracellular signal-regulated kinase; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; HIF-1α, hypoxia-inducible factor 1-alpha; VEGFA, vascular endothelial growth factor A; IL-1β, interleukin-1 beta; CXCL1/2, C-X-C motif chemokine ligand 1/2. Mg2+, magnesium ion; SIRT2, sirtuin 2; DNA, deoxyribonucleic acid; TLR9, toll-like receptor 9; cGAS, cyclic GMP-AMP synthase; STING, stimulator of interferon genes; M1, classically activated macrophage.SDC1, syndecan-1; CXCL5, C-X-C motif chemokine ligand 5; CXCR4, C-X-C chemokine receptor type 4; CAPN2, calpain 2; JAM3, junctional adhesion molecule 3.
Figure 2.
Mechanistic Diagram of Piezo1-Induced Neutrophil Activation and Functional Reprogramming. NETs, neutrophil extracellular traps; NETosis, neutrophil extracellular trap cell death; Calpain, calcium-dependent cysteine protease. ERK, extracellular signal-regulated kinase; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; HIF-1α, hypoxia-inducible factor 1-alpha; VEGFA, vascular endothelial growth factor A; IL-1β, interleukin-1 beta; CXCL1/2, C-X-C motif chemokine ligand 1/2. Mg2+, magnesium ion; SIRT2, sirtuin 2; DNA, deoxyribonucleic acid; TLR9, toll-like receptor 9; cGAS, cyclic GMP-AMP synthase; STING, stimulator of interferon genes; M1, classically activated macrophage.SDC1, syndecan-1; CXCL5, C-X-C motif chemokine ligand 5; CXCR4, C-X-C chemokine receptor type 4; CAPN2, calpain 2; JAM3, junctional adhesion molecule 3.

Figure 3.
Mechanistic Diagram of Piezo1-Induced Macrophage Polarization. NF-κB, Nuclear factor kappa-light-chain-enhancer of activated B cells; Calpain, Calcium-dependent cysteine protease; HIF-1α, Hypoxia-inducible factor 1-alpha; TNF-α, Tumor necrosis factor-alpha; IL-6, Interleukin-6; M1, Classically activated macrophage; PI3K/AKT, Phosphatidylinositol 3-kinase/Protein kinase B. YAP, Yes-associated protein; p53, Tumor protein p53; SPP1, Secreted phosphoprotein 1; IL-10, Interleukin-10; Interleukin-10; M2 Alternatively activated macrophage; TGF-β1, Transforming growth factor-beta 1.
Figure 3.
Mechanistic Diagram of Piezo1-Induced Macrophage Polarization. NF-κB, Nuclear factor kappa-light-chain-enhancer of activated B cells; Calpain, Calcium-dependent cysteine protease; HIF-1α, Hypoxia-inducible factor 1-alpha; TNF-α, Tumor necrosis factor-alpha; IL-6, Interleukin-6; M1, Classically activated macrophage; PI3K/AKT, Phosphatidylinositol 3-kinase/Protein kinase B. YAP, Yes-associated protein; p53, Tumor protein p53; SPP1, Secreted phosphoprotein 1; IL-10, Interleukin-10; Interleukin-10; M2 Alternatively activated macrophage; TGF-β1, Transforming growth factor-beta 1.

Figure 4.
Mechanistic Diagram of Piezo1-Induced Platelets Mechanosensation and Coagulation Function. NETs, neutrophil extracellular traps; PDGF, platelet-derived growth factor; PF4, platelet factor 4; RhoA, Ras homolog family member A; ROCK, Rho-associated protein kinase; TGF-β, transforming growth factor beta.
Figure 4.
Mechanistic Diagram of Piezo1-Induced Platelets Mechanosensation and Coagulation Function. NETs, neutrophil extracellular traps; PDGF, platelet-derived growth factor; PF4, platelet factor 4; RhoA, Ras homolog family member A; ROCK, Rho-associated protein kinase; TGF-β, transforming growth factor beta.

Figure 5.
Mechanistic Diagram of Piezo1-Induced Lymphocytes Phenotypic Alterations.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



