Submitted:
23 April 2026
Posted:
24 April 2026
You are already at the latest version
Abstract
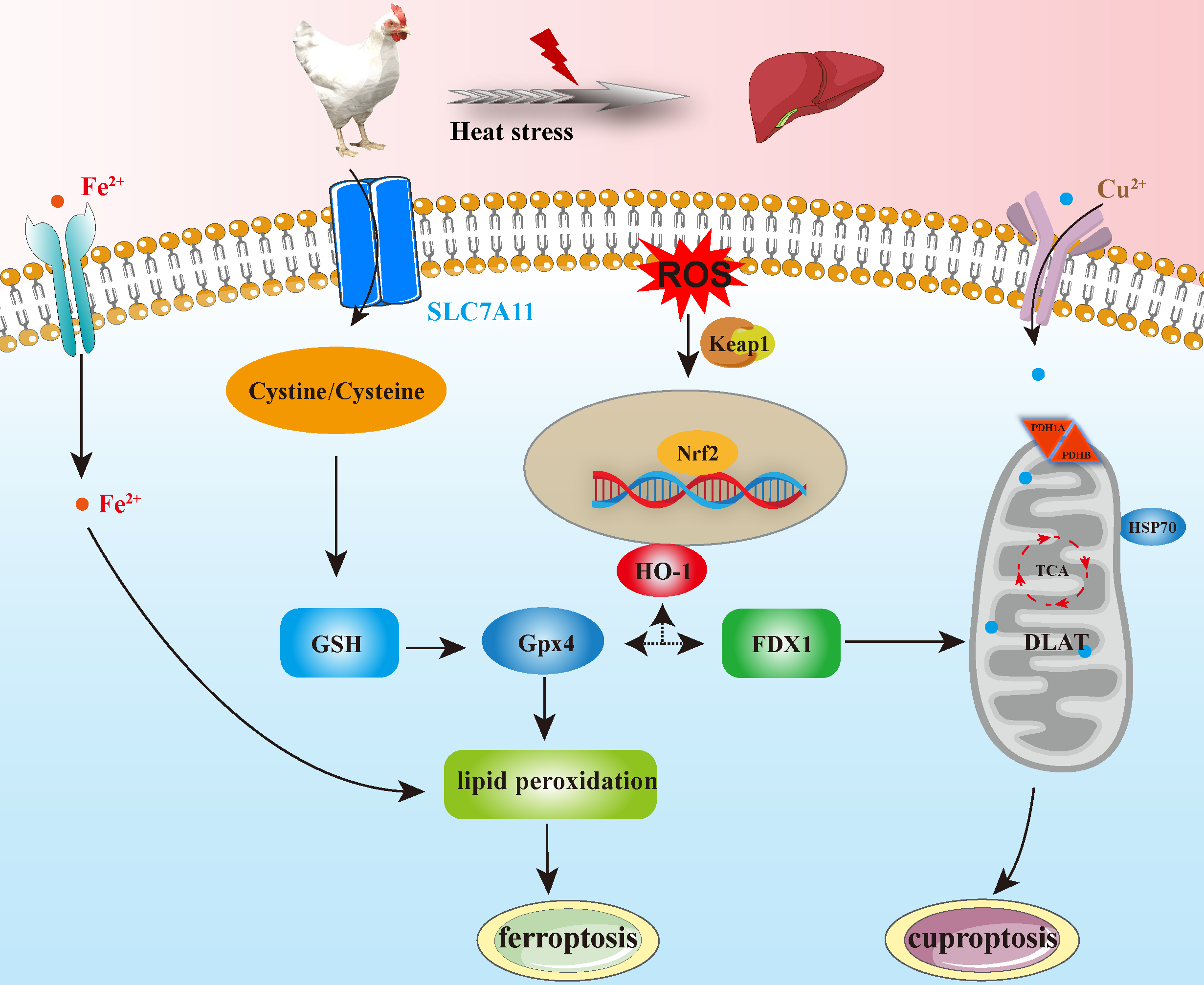
As the global average temperature increases, heat stress (HS) caused by high temperatures has become a key constraint to the development of the poultry industry. As the primary metabolic organ, HS can induce liver injury in chickens, thereby compromising food safety. However, the precise mechanisms underlying HS -induced liver injury remain to be elucidated. The objective of this study is to explore the impact of HS on liver damage, oxidative stress, the Keap1-Nrf2 pathway, ferroptosis and cuproptosis in chickens. A total of 70 chickens were selected for this experiment and divided into a CON group and HS group: the CON group was reared in a normal-temperature environment (24 ± 1 ℃), whilst the HS group was reared in a high-temperature environment (33 ± 1 ℃). The findings of the study suggested that HS has the potential to induce liver dysfunction, oxidative stress, and disruption of the Keap1-Nrf2 pathway. HS has been demonstrated to induce Fe2+ accumulation in chicken livers, inhibit the expression of FTH1, FSP1, SLC7A11 and Gpx4, and simultaneously upregulate the expression of CD71, PTGS2 and ACSL4, thereby promoting ferroptosis. Furthermore, Cu2+ accumulation in the liver upregulates HSP70, DLAT and Lip-DLAT levels and downregulates the expression of ATP7B, PDH1A, PDHB, PDK4, DLST and FDX1, thereby inducing cuproptosis. Subsequent correlation analysis revealed that HS can induce ferroptosis and cuproptosis via the HO-1/FDX1/Gpx4 pathway. This finding provides new insights into the mechanisms underlying HS-induced liver injury.
Keywords:
heat stress
; cuproptosis
; ferroptosis
; HO-1/FDX1/Gpx4 axis
; liver
1. Introduction
Elevated ambient temperatures can result in heat stress (HS) in animals, which has become a significant challenge for the global livestock industry [1]. HS can act as a significant environmental stressor, negatively affecting the performance of broiler chickens and resulting in economic losses for the poultry industry. Meanwhile, HS can cause a range of diseases in animals, including liver damage, heart damage, intestinal damage, and immune organ damage [2]. It is imperative to acknowledge that HS has been evidenced to induce liver damage, culminating in liver dysfunction. Furthermore, the results of research studies have revealed that HS has the capacity to initiate mitochondrial dysfunction in the liver. Moreover, the results of research undertaken to date have demonstrated that HS has the potential to induced mitochondrial dysfunction in the liver [3]. Nevertheless, further comprehensive investigation is required to illuminate the molecular mechanisms by which HS instigates liver injury.
Oxidative stress is defined as an equilibrium disturbance between oxidative and antioxidant processes within the body [4,5]. Excessive accumulation of oxidative stress has been proven to result in the damage of proteins, DNA and organelles. It has been revealed through further research that the presence of oxidative stress within cells has the capacity to result in the disruption of mitochondrial structure and function [6]. Concurrently, the Keap1-Nrf2 pathway functions as a pivotal regulatory mechanism for oxidative stress. In recent years, research has shown that cell damage can cause different types of cell death, including apoptosis, necrosis and pyroptosis. The recent research findings have indicated that cuproptosis plays a substantial role in the process of cell death [7,8]. Currently, cuproptosis currently plays a significant role in a wide range of diseases. Research indicates that HS exacerbates oxidative stress, increasing the demand for copper-dependent antioxidant enzymes (such as Cu/Zn-SOD), which leads to improved copper utilization in the body [9]. What's more, copper can stick to DLAT and encourage it to be lipoylated, which can trigger cuproptosis [10,11]. Research has shown that cuproptosis is involved in a variety of diseases, but we still don't know what its role is in HS.
Ferroptosis is defined as a form of programmed cell death that is dependent on iron and is defined by the progressive accumulation of lipid peroxides, which leads to the disruption of membrane structure by oxidation [12]. As a pivotal molecule in ferroptosis, glutathione peroxidase 4 (Gpx4) effectively maintains cellular lipid homeostasis and prevents the onset of ferroptosis. Concurrently, under the influence of copper ions, Gpx4 is more readily degraded via the autophagy pathway, thereby indirectly promoting iron-induced ferroptosis [13,14]. This finding underscores a pivotal nexus between cuproptosis and ferroptosis, thereby imparting novel insights into the mechanisms of cellular damage engendered by the disruption of metal ion homeostasis. As evidenced by Zhang et al. [15], ferroptosis has been shown to play a pivotal role in hepatocyte injury triggered by HS. Concurrently, copper-binding ferredoxin 1 (FDX1), a pivotal regulator of cuproptosis, functions in both iron metabolism and transport [16]. Despite the differences in their mechanisms, ferroptosis and cuproptosis are both characterized by dysregulation of metal ion metabolism and elevated oxidative stress. This finding indicated the potential for cross-regulatory interactions under specific physiological conditions. Consequently, ascertaining the mechanisms of HS-induced liver injury and the interplay between cuproptosis and ferroptosis is of significant importance.
In accordance with the aforementioned fundamental scientific inquiries, this study employed AA broiler chickens as experimental subjects for the purpose of establishing an HS model. The present study is chiefly concerned with the molecular mechanisms underlying HS-induced liver injury and with elucidating the potentially interactive relationship between cuproptosis and ferroptosis. This research offers significant scientific substantiation for the formulation of preventative and therapeutic measures for HS, thus facilitating the mitigation of economic losses.
2. Materials and Methods
2.1. Animals and Treatment
This animal experiment was approved by the Animal Experiment Management Committee of the Anhui Science and Technology University (Approval No. AK2025080). Seventy 1-day-old healthy Abbott's Acres (AA) broiler chicks were selected for the experiment and fed (Table S1) and watered ad libitum during the feeding period. After feeding to 28 days old, they were separated divided into the control group (CON) (environment temperature 24 ± 1 °C) and heat stress group (HS) (environment temperature 33 ± 1 ℃) [17,18]. Until 42 days, animals in each group were euthanized under ether anaesthesia, and serum, liver were collected.
2.2. Biochemical Analysis
After blood collection, the serum is separated. The concentrations of lactate dehydrogenase (LDH), aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP) in the serum are then analysed using an automated blood chemistry analyzer (Mindray BS-200, China).
2.3. Histopathological Analysis
Chicken liver was fixed in 4% paraformaldehyde solution, gradient xylene and alcohol treatment were performed as per the previous method. HE staining of liver was stained using hematoxylin-eosin staining. Liver Masson and PAS staining was performed according to the previous method, and finally the results were observed by microscopy.
2.4. Determination of Oxidative Stress Level
Weigh out 100 mg of liver tissue and add 900 µL of saline solution, then mix it all together. Centrifuge at 3,500 rpm for 15 minutes, then collect the supernatant. Determine total antioxidant capacity (T-AOC), glutathione peroxidase (GSH-Px), superoxide dismutase (SOD) and malondialdehyde (MDA) levels according to the kit instructions [18].
2.5. Determination of Cu and Iron Levels
Take an appropriate amount of serum and liver samples. The Cu2+, Fe2+ and total iron assays should be carried out in conformity with the kit instructions [19].
2.6. Immunohistochemical Assay
Liver sections were subjected to a deparaffinization process using xylene, following a standard protocol, followed by antigen retrieval using citrate buffer. Subsequently, the sections were immersed in a blocking solution comprising 10% horse serum. Subsequent to this, the incubation process was continued for a period of 16 hours, during which the following antibodies were utilized: CD71 (1:1000, Servicebio, China), PTGS2 (1:200, ABclonal, China) and ATP7B (1:200, Zenbio, China). The results obtained are presented in photographs taken following staining with DAB and hematoxylin.
2.7. Immunofluorescence Assay
The pre-treatment was analogous to immunohistochemistry, followed by cultivation with 8-OHdg, DLAT (1:50, ABclonal, China), HO-1 (1:100, Proteintech, China), Gpx4 (1:200, Proteintech, China), and FDX1 (1:200, ABclonal, China) antibodies, with a duration of 14 h. DAPI staining (Beyotime, China) was then conducted, followed by a secondary antibody incubation. The findings were meticulously recorded through the utilization of confocal microscopy (Leica, Germany).
2.8. Western Blot Analysis
2.9. Molecular Docking
The investigation of potential interactions between target proteins is facilitated by the utilization of molecular docking techniques. The protein structures were predicted using AlphaFold 3 and visualized using PyMOL. Subsequently, molecular docking was undertaken using AutoDock Vina and the results were rendered in PyMOL.
2.10. Statistical Analysis
The data were examined using SPSS version 24.0 (SPSS Inc., USA) and presented as the mean ± standard deviation (SEM) (n = 6). The generation of statistical graphs was facilitated by the GraphPad Prism 8.0 software program (GraphPad Inc., La Jolla, CA, USA). The confidence intervals were calculated as follows: * for P < 0.05, ** for P < 0.01, and *** for P < 0.001.
3. Results
3.1. Heat Stress Induced Liver Injury in Chicken
We investigated the effects of HS on the liver. Serum biochemical markers (LDH, AST, ALT, ALP) were utilized to evaluate liver function. The findings indicated that, in comparison with the CON group, HS led to a substantial augmentation in the levels of these markers (P < 0.05) (Figure 1A-D). In contrast, the hepatic lobules in the HS group exhibited a disorganized pattern, accompanied by the presence of inflammatory cell infiltration. It is also worthy of note that mild degenerative changes to hepatocytes were also identified in the liver of the HS group (Figure 1E). Concurrently, the use of Masson staining revealed increased collagen fibre deposition (blue) in the HS group in comparison with the CON group (Figure 1F). PAS staining revealed increased glycogen deposition (purple) in the HS group than CON group (Figure 1G). The results of the present study demonstrate that HS can induce liver damage in chickens.

3.2. Effect of Heat Stress on Oxidative Stress in Chicken Liver
This study examines the effect of HS on oxidative stress in the liver through the Keap1-Nrf2 pathway. The results of the protein imaging and analysis demonstrated that the levels of proteins Keap1, Nrf2, and HO-1 were dropping in the HS group, but these changes were not big enough to be considered important (Figure 2A-B). Subsequent analysis of antioxidant markers revealed no significant alteration in the levels of T-AOC in the liver (Figure 2C). Concurrently, a marked decline in the enzymatic activities of GSH-Px and SOD was observed in the HS group when compared with the CON group (P < 0.05) (Figure 2D-E). In addition, a significant increase in MDA levels was detected in the HS group in comparison with the CON group (P < 0.05) (Figure 2F). 8-ohdg, a significant biomarker of oxidative DNA damage, exhibited a substantial increase in the HS group, as evidenced by immunofluorescence analysis (Figure 2G). The above findings show that HS causes oxidative stress in the liver via the Keap1-Nrf2 pathway.

3.3. Effect of Heat Stress on Ferroptosis in Chicken Liver
The current study explores the effect of HS on ferroptosis in the liver. Serological analysis indicated that Fe2+ levels were heightened in the HS group (P < 0.05), while total iron levels remained unchanged (Figure 3A). Concurrently, levels of Fe2+ and total iron in the liver was observed to elevated in the HS group in comparison with the CON group (P < 0.05) (Figure 3B). The results for ferroptosis-related proteins are shown in Figure 3C. Protein analysis revealed that, compared with the CON group, the levels of CD71, PTGS2, and ACSL4 were higher in the HS group (P < 0.05) (Figure 3D, 3F-H). However, no significant alterations in the levels of FTH1 and SLC7A11 were observed (Figure 3E, 3I). Concurrently, the level of FSP1 and Gpx4 were found to be decreased in the HS group in compared to the CON group (P < 0.05) (Figure 3J). The results of the immunohistochemical tests showed increase in the levels of proteins CD71 and PTGS2 in the HS group than the CON group (P < 0.05) (Figure 3K-M). The findings outlined above suggest that HS can induce ferroptosis in chicken livers.

3.4. Effect of Heat Stress on Cuproptosis in Chicken Liver
The serological analysis demonstrated that the HS group exhibited reduced levels of Cu2+ in comparison with the CON group (Figure 4A). Concurrently, a significant elevation in liver Cu2+ levels was observed in the HS group in comparison with the CON group (P < 0.05) (Figure 3B). The results of the immunohistochemical analysis demonstrated that, in comparison with the CON group, the level of ATP7B-brown positivity was diminished in the HS group (Figure 4C). The results pertaining to cuproptosis-related proteins are illustrated in Figure 4D. The analysis of the protein results indicates a downward trend in the levels of proteins ATP7B, PDH1A, PDHB, PDK4, DLST and FDX1 in the HS group (Figure 4E-F). Concurrently, protein HSP70, DLAT and Lip-DLAT exhibited a marked increase in the HS group in comparison with the CON group (P < 0.05). The immunofluorescence results indicated a marked increase in the fluorescence intensity of DLAT in the HS group compared to the CON group (Figure 4G). The findings outlined above suggested that HS can induced cuproptosis in chicken livers.

3.5. Correlation Analysis of HO-1, FDX1 and Gpx4
In light of the aforementioned findings, it is hypothesized that there may be a correlation between the Keap1-Nrf2 signalling pathway, and the processes of iron-mediated ferroptosis and copper-mediated cuproptosis (Figure 5A). STING predictions revealed a relationship between HO-1, FDX1 and Gpx4 (Figure 5B). Furthermore, to investigate this issue, we first used AlphaFold3 to predict the three-dimensional structures of chicken HO-1, FDX1 and GPX4. Molecular docking revealed interactions between them (Figure 5C-E). To gain a more comprehensive and intuitive insight into these alterations, the fluorescent localization of HO-1, FDX1 and GPX4 was examined further using immunofluorescence confocal microscopy (Figure 5F-H). The results indicated that, under the influence of HS, the number of fluorescent levels for HO-1, FDX1 and GPX4 was significantly reduced.

4. Discussion
In the context of global environmental changes, HS has emerged as a substantial health hazard for animals, particularly given its capacity to induce organ damage in poultry [20]. HS has been demonstrated to cause a disruption in liver function and metabolism, with the potential to impair multi-organ function, thus constituting a grave threat to animal and human health [21]. HS makes organisms' tissues injury and messes up the body's oxidative balance. Moreover, the prevailing consensus among numerous scholars is that the mechanisms underlying HS injury may encompass a multitude of cell death phenomena. However, the relationship between ferroptosis and cuproptosis in HS-induced hepatic injury remains incompletely elucidated. The present study employed histopathological observation with a view to identifying alterations in hepatic function and pathology. Subsequent research has indicated that the disruption of copper and iron metabolism in the livers of HS broiler chickens can be facilitated by the Keap1-Nrf2 pathway, resulting in ferroptosis and cuproptosis.
It is widely acknowledged that the deleterious effects of HS are associated with the generation of ROS, which in turn leads to oxidative damage to organisms [22]. The Keap1-Nrf2 pathway has been identified as a pivotal regulatory mechanism in cellular responses to oxidative stress [23]. In typical circumstances, Nrf2 is bound by its repressor protein, Keap1, and targeted for ubiquitination and degradation. This process ensures the maintenance of stable expression levels [24]. During periods of oxidative stress, this pathway activates the transcription of multiple antioxidant enzyme genes. Superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) are essential antioxidant enzymes that protect cells by eliminating reactive oxygen species and breaking down peroxides, while total antioxidant capacity (T-AOC) reflects the body’s overall ability to counteract oxidative stress [25]. Conversely, malondialdehyde (MDA), a by-product of lipid peroxidation, functions as a pivotal indicator of oxidative damage. The findings of this study are corroborated by those of preceding research [26,27,28], suggesting that HS functions as a mediator for oxidative stress in the liver via the Keap1-Nrf2 pathway. Meanwhile, 8-hydroxy-deoxyguanosine (8-ohdg) is an oxidation product of deoxyguanosine and an established biomarker of oxidative DNA damage [29]. This study found that immunofluorescence results showed increased levels of 8-OHdG fluorescence in the cytoplasm of hepatocytes in the HS group, indicating mitochondrial DNA damage. Furthermore, as the cell's powerhouse, mitochondria fulfil a number of functions. These include the regulation of cell proliferation, apoptosis and signal transduction [30]. The findings outlined above indicate that HS has the capacity to induce Keap1-Nrf2-mediated oxidative stress and mitochondrial damage within the liver.
HS can induce oxidative stress and mitochondrial damage in hepatocytes, thereby exacerbating metabolic dysfunction. At the same time, the imbalance in trace elements within the body caused by HS has been extensively studied, and can lead to disturbances in hepatic iron metabolism [31]. Consequently, the absorption and transport of iron in the liver is disrupted by HS, which may lead to pathological iron accumulation. HS has been demonstrated to disrupt iron metabolism in the liver, with this disruption potentially inducing ferroptosis [32]. Ferroptosis is characterized by a dynamic equilibrium between iron uptake, facilitated by CD71, and iron storage, driven by FTH1 (ferritin heavy chain 1) [33]. Disruption of this balance exerts a substantial influence on cellular fate. In addition, as a glutathione-independent ferroptosis inhibitor, FSP1 functions as a redox enzyme, thereby inhibiting lipid peroxidation [34]. Furthermore, ACSL4 has been proven to instigate ferroptosis by promoting the transfer of polyunsaturated fatty acids to membrane phospholipids, thereby rendering the membrane phospholipids more susceptible to peroxidation. PTGS2 has been demonstrated to be intimately associated with the ferroptosis process and is extensively utilized as a molecular marker for ferroptosis [35]. Gpx4 primarily exerts its protective effects by scavenging lipid peroxides, whilst SLC7A11 facilitates cystine uptake to support glutathione synthesis, thereby providing the functional basis for Gpx4 [36]. It has been demonstrated by related studies that exposure of the livers of broiler chickens to HS can result in the induction of ferroptosis [37,38]. This study found that HS increased total iron and Fe2+ levels in serum and the liver, while reducing the liver's FTH1 protein expression levels. Furthermore, HS resulted in significant upregulation of CD71, PTGS2 and ACSL4 protein levels, and downregulation of FSP1, SLC7A11 and Gpx4 protein levels, in the liver. These results imply that HS induces ferroptosis in the liver due to iron overload.
Cuproptosis, a recently identified form of cell death, is initiated by the excessive accumulation of copper ions. This results in the aberrant accumulation of thiol-modified proteins, the disruption of iron-sulphur proteins involved in mitochondrial respiration, and the subsequent occurrence of protein-induced toxic stress-mediated cell death [10]. The findings of the research suggested that copper accumulation has the potential to induced cuproptosis [19]. The present study suggests that, in HS chickens, serum copper levels were reduced, while liver copper levels were significantly elevated. This phenomenon may be attributed to the elevated levels of inflammatory cytokines and acute-phase proteins, which are known to induce a heightened transfer of copper from the blood to the liver [9]. Concurrently, the interaction of acetylated DLAT with copper has been demonstrated to result in its aberrant aggregation, thereby instigating copper-induced cuproptosis. This is similar to other studies that have shown that, in heart failure, an imbalance in copper metabolism can trigger a process called cuproptosis via the SIRT1/HMGB1 pathway [39]. The experimental results obtained in this study suggest that HS can induce cuproptosis by regulating key proteins, including ATP7B, PDHA1, PDHB, PDK4, HSP70, DLAT, Lip-DLAT and FDX1. The current study's findings indicate that HS can lead to the accumulation of copper within the liver and the induction of cuproptosis.
Cuproptosis is characterised by a distinct molecular mechanism whilst also exhibiting certain similarities to the molecular mechanism of ferroptosis [40]. As a key regulator of ferroptosis, Gpx4 has been demonstrated to bind directly with copper ions, inducing their aggregation and subsequent degradation via the autophagic pathway, thereby diminishing their capacity to inhibit cuproptosis [41]. FDX1 is a small-molecule protein that comprises an iron-sulphur centre and functions in a variety of essential biological processes. Moreover, any disruption to the biosynthesis of iron-sulphur clusters has the potential to disrupt intracellular iron homeostasis and mitochondrial function, thereby exacerbating the occurrence of ferroptosis [42]. In this study, it was observed that HS upregulates the acetylation levels of DLAT. This finding suggests the potential for a degree of mutual regulation and an underlying mechanism linking cuproptosis and ferroptosis. The experimental data obtained in this present analysis suggest a considerable decline in the levels of FDX1 and Gpx4, accompanied by a decrease in their fluorescent colocalization, in response to HS. In addition, the prediction of the protein configurations of FDX1 and Gpx4 has revealed an interaction between these proteins, at least to a certain extent, with respect to their structural configuration. The findings of this study indicate that the interaction between HS-induced cuproptosis death and ferroptosis may be facilitated by the interplay between FDX1 and Gpx4. Furthermore, STING results and molecular docking studies indicate an interaction of HO-1, FDX1 and Gpx4. Research findings have indicated a correlation between HO-1 and cuproptosis [43,44]. Concurrently, HO-1 exerts a substantial influence on the process of ferroptosis [45,46]. The present study indicates that the HO-1/FDX1/Gpx4 axis serves as a pivotal link between HS-induced cuproptosis and ferroptosis.
5. Conclusions
In conclusion, the results of this study indicate that exposure of chicken liver to HS leads to an imbalance in trace elements, which in turn promotes cuproptosis and ferroptosis induces iron-mediated apoptosis via the HO-1/FDX1/Gpx4 axis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, G.Q.X. and D.Q.Y.; methodology, G.Q.X.; software, G.Q.X.; validation, G.Q.X., D.Q.Y. and Z.K.W.; formal analysis, G.Q.X. and D.Q.Y.; investigation, J.X.D. and F.Y.M.; resources, F.Y.M.; data curation, G.Q.X.; writing—original draft preparation, G.Q.X.; writing—review and editing, G.Q.X., F.Y.M. and S.J.H.; visualization, G.Q.X. and F.Y.M.; supervision, F.Y.M. and S.J.H.; project administration: F.Y.M. and S.J.H.; funding acquisition, F.Y.M. and S.J.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Talent Introduction Project of Anhui Science and Technology University (No. DKYJ202302), Veterinary Science Peak Discipline Project of Anhui Science and Technology University (No. XK-XJGF002) and Major projects supported by Department of Education Anhui Province (No. 2023AH040282).
Conflicts of Interest
We declare that there is no conflict of interest in this work.
References
- Hossain, M.M.; Ahn, J.; Choi, S.Y.; Hur, S.P.; Lim, D.; Shin, D.; Lee, S.; Park, J.E. Thermal stress responses and heat stress resilience genes in chickens are revealed through genomic and transcriptomic insights. Journal of animal science and biotechnology 2026, 17, 42. [CrossRef]
- Aryal, B.; Kwakye, J.; Ariyo, O.W.; Ghareeb, A.F.A.; Milfort, M.C.; Fuller, A.L.; Khatiwada, S.; Rekaya, R.; Aggrey, S.E. Major oxidative and antioxidant mechanisms during heat stress-induced oxidative stress in chickens. Antioxidants (Basel) 2025, 14, 471. [CrossRef]
- Chen, S.; Liu, H.; Zhang, J.; Zhou, B.; He, X.; Wang, T.; Wang, C. Dietary rutin improves breast meat quality in heat-stressed broilers and protects mitochondria from oxidative attack via the AMPK/PINK1-Parkin pathway. J. Sci. Food Agric. 2023, 103, 2367-2377. [CrossRef]
- Ma, F.; Wang, M.; Zhong, G.; Liao, J.; Huo, Y.; Wang, Z.; He, S. The impact of copper-induced oxidative damage on the endoplasmic reticulum quality control system in broiler kidneys. BioMetals 2025, 38, 1131-1143. [CrossRef]
- Alqhtani, H.A.; Elbaz, A.M.; Hegazy, S.A.; Abdelhady, A.Y.; Safhi, F.A.; Marzok, M.; Rizk, M.A.; Al-Rasheed, M.; Mohamed, M.H.; Abdel-Raheem, S.M.; et al. Dietary Combined Thyme Meal and Bacillus subtilis to Promote Growth Performance, Immune Function, Gene Expression, Antioxidant Defense, and Cecal Microbiota in Growing Rabbits Under Heat Stress Conditions. Veterinary Sciences 2026, 13. [CrossRef]
- Ji, L.; Yu, H.; Wang, R.; Yan, H.; Yin, X.; Guo, S.; Wang, R. Targeting magnesium homeostasis: a novel therapeutic strategy for liver diseases. Frontiers in nutrition 2026, 13, 1709477. [CrossRef]
- Ma, F.; Huo, Y.; Liao, J.; Xu, G.; Wang, Z.; He, S. The role of lipid metabolism imbalance in copper-induced PANoptosis in broiler kidney. Poult. Sci. 2025, 104, 105549. [CrossRef]
- Qin, W.; Sheng, H.; Hu, X.; Burden, R.E.; Martin, S.L.; Wu, H. Cuproptosis in cancer: emerging mechanism and therapeutic opportunities. Trends Pharmacol. Sci. 2026, 47, 386-402. [CrossRef]
- Siquier-Coll, J.; Bartolomé, I.; Perez-Quintero, M.; Grijota, F.J.; Arroyo, J.; Muñoz, D.; Maynar-Mariño, M. Serum, erythrocyte and urinary concentrations of iron, copper, selenium and zinc do not change during an incremental test to exhaustion in either normothermic or hyperthermic conditions. J. Therm. Biol. 2019, 86, 102425. [CrossRef]
- Tsvetkov, P.; Coy, S.; Petrova, B.; Dreishpoon, M.; Verma, A.; Abdusamad, M.; Rossen, J.; Joesch-Cohen, L.; Humeidi, R.; Spangler, R.D.; et al. Copper induces cell death by targeting lipoylated TCA cycle proteins. Science 2022, 375, 1254-1261. [CrossRef]
- Xiong, W.; Luo, Z.; Wang, H.; Su, Q.; Wang, H.; Xu, J.; Zheng, W. Amyloid beta 42 disrupts cardiac function in Alzheimer's disease mice via SLC31A1 upregulation-mediated cuproptosis. Basic Res. Cardiol. 2026, 121, 285-299. [CrossRef]
- Cui, S.; Ye, J. Ferroptosis: the demise of cells through phospholipid peroxidation. Advanced science (Weinheim, Baden-Wurttemberg, Germany) 2026, 20, e24387. [CrossRef]
- Wu, Y.; Fu, L.; Xu, X.; Chan, P.S.; Wu, R. Systematic and quantitative investigation of newly synthesized proteins reveals distinct ion homeostasis and mitochondrial changes between cuproptosis and ferroptosis in human cells. Anal. Chem. 2026, 98, 8309-8320. [CrossRef]
- Hao, L.; Yu, P.; Yan, R.; Li, K.; Luo, Z.; Xiang, S.; Wang, Y.; Fang, C.; Wang, G.; Ma, S.; et al. Transdermal siRNA delivery via biomineralized nanoparticle-incorporated microneedles modulates cuproptosis-ferroptosis interaction for psoriasis therapy. Materials today. Bio 2026, 37, 102991. [CrossRef]
- Zhang, Y.; Li, Y.; Wang, R.; Wang, S.; Sun, B.; Cao, D.; Sun, Z.; Lv, W.; Ma, B.; Zhang, Y. Chronic heat stress caused lipid metabolism disorder and tissue injury in the lliver of Huso dauricus via oxidative-stress-mediated ferroptosis. Antioxidants (Basel) 2025, 14, 92. [CrossRef]
- Huang, X.; Yu, S.; Wei, W.; Tao, W.; Cai, T.; Wen, L.; Ye, J.; Zhang, C.; Feng, H.; Cao, S.; et al. Fe-S protein FDX1 triggers tumor-intrinsic innate immunity via mitochondrial nucleic acids release to orchestrate ferroptosis in CCRCC. Advanced science (Weinheim, Baden-Wurttemberg, Germany) 2026, 13, e18323. [CrossRef]
- Ding, J.; Miao, J.; Zhang, X.; Xiong, Y.; Ma, F.; He, S. Total saponins of Gynostemma pentaphyllum mitigate chronic heat stress-induced thymus and spleen inflammation in broilers via NF-κB pathway activation. Poult. Sci. 2026, 105, 106797. [CrossRef]
- Xu, G.; Ma, F.; Li, Y.; Miao, J.; Zhang, X.; Xiong, Y.; He, S. Puerarin alleviates liver injury induced by heat-stress through regulating oxidative stress and apoptosis in broilers. J. Therm. Biol. 2025, 134, 104317. [CrossRef]
- Zhang, T.; Dai, X.; Wang, H.; Hu, Y.; Gao, X.; Shu, X.; Cao, H.; Hu, G.; Xing, C.; Yang, F. Copper-induced duodenal injury: Unveiling the dual role of cuproptosis and ferroptosis via the FDX1/GPX4 axis. J. Nutr. Biochem. 2026, 149, 110181. [CrossRef]
- Tolba, S.A.; Amer, S.A.; Abdel-Warith, A.A.; Younis, E.M.; Gouda, A.; Elshopakey, G.E.; Abd-Allah, N.A.; Reda, R.; Hamed, R.I.; Saleh, G.K.; et al. Fennel essential oil as a natural feed additive to mitigate heat stress in broilers: Effects on growth, physiological and immunological responses, and intestinal morphology. J. Therm. Biol. 2026, 137, 104437. [CrossRef]
- Zendeli, D.; Colaninno, N.; van Esch, M.; Eldesoky, A.H.; Morello, E.; van Timmeren, A. Urban heat stress and health: A systematic literature review of dimensions and indicators for planning and design. Health Place 2026, 99, 103643. [CrossRef]
- Pradhan, P.; Foresti, R.; Motterlini, R.; Immenschuh, S. Regulatory interplay between nitric oxide and heme in redox signaling and inflammation. Redox Biol 2026, 92, 104126. [CrossRef]
- García-Yagüe Á, J.; Cazalla, E.; Cuadrado, A. Therapeutic potential of isothiocyanates by targeting the NRF2 pathway. Free Radic. Biol. Med. 2026, 248, 554-596. [CrossRef]
- Lan, T.; Zhou, K.; Duan, G.; Zhang, Y.; Ma, Y.; Fan, G.; Qi, Y. Lycium L. flavonoids: extraction, purification, signal transduction pathways, and interactions with intestinal microbiota. NPJ science of food 2026, 10.1038/s41538-026-00784-w. [CrossRef]
- Durço, A.O.; da Silva Alcântara, F.; Rocha, C.S.G.; Mota, K.O.; Conceição, L.S.R.; Orts, D.; Sousa, G.M.; Santos, M.R.V.; Souza, D.S.; Roman-Campos, D. Pyrethroid-induced cardiac dysfunction: A systematic review and meta-analysis of preclinical evidence. Chem. Biol. Interact. 2026, 432, 112030. [CrossRef]
- Ding, K.N.; Lu, M.H.; Guo, Y.N.; Liang, S.S.; Mou, R.W.; He, Y.M.; Tang, L.P. Resveratrol relieves chronic heat stress-induced liver oxidative damage in broilers by activating the Nrf2-Keap1 signaling pathway. Ecotoxicol. Environ. Saf. 2023, 249, 114411. [CrossRef]
- Yuan, J.; Li, Y.; Miao, J.; Zhang, X.; Xiong, Y.; Ma, F.; Ding, J.; He, S. Bamboo leaf flavonoids ameliorate cyclic heat stress-induced oxidative damage in broiler liver through activation of Keap1-Nrf2 signaling pathway. Poult. Sci. 2025, 104, 104952. [CrossRef]
- Lei, Y.; Liu, H.; Xiang, Q.; Liu, Y.; Wu, D.; Zhang, J.; Lin, Y. Dietary lycopene mitigates reproductive impairment in heat-stressed rongchang boars: roles of antioxidant, anti-inflammatory and Nrf2 pathway. Antioxidants (Basel) 2026, 15, 245. [CrossRef]
- Kuninaka, Y.; Ishida, Y.; Grimaldi, F.; Irmici, M.; Nosaka, M.; Munekawa, M.; Pelotti, S.; Fais, P.; Kondo, T. 8-OHdG and NT-proBNP as complementary biomarkers in the postmortem diagnosis of acute ischemic heart disease. Sci. Rep. 2026, 16, 6154. [CrossRef]
- Liu, Y.; Chang, Y.Z. The regulation of mitochondrial ferritin on mitochondrial redox balance is essential to cell fate decision. Redox Biol 2026, 92, 104124. [CrossRef]
- McKay, A.K.A.; McCormick, R.; Tee, N.; Peeling, P. Exercise and heat stress: inflammation and the iron regulatory response. Int. J. Sport Nutr. Exerc. Metab. 2021, 31, 460-465. [CrossRef]
- An, Q.; Wei, R.; Huang, Z.; Tang, Y.; Wang, M.; He, S.; Huang, K.; Liu, Z.; Zhang, M.; Li, R.; et al. Palmitoylation of Tfr1 enhances platelet ferroptosis and liver injury in heat stroke. Acta pharmaceutica Sinica. B 2026, 16, 270-286. [CrossRef]
- Caverzan, M.D.; Ibarra, L.E. Advancing glioblastoma treatment through iron metabolism: A focus on TfR1 and Ferroptosis innovations. Int. J. Biol. Macromol. 2024, 278, 134777. [CrossRef]
- Wang, J.; Guo, D.; Jiang, S.; Wu, W.; Gao, X. Targeting ferroptosis in cancer: from mechanistic insights to therapeutic approaches. Molecular biomedicine 2026, 7, 20. [CrossRef]
- Jiang, M.; Zhang, L.; Lin, Q.; Guo, D.; Jin, Q.; Xu, J.; Guo, M.; Peng, S.; Sun, Q.; Liu, N.; et al. Endoplasmic reticulum-targeting prodrug nanoassemblies for potent ferroptosis therapy via concurrent glutathione depletion and arachidonic acid accumulation. Acta Biomater. 2026, 214, 542-556. [CrossRef]
- Chen, H.; Dai, X.; Xiong, Z.; Cao, H.; Xing, C.; Li, H.; Gao, X.; Hu, M.; Yang, F. Dual-pathway mechanism of vanadium-induced hepatotoxicity in ducks: Synergistic crosstalk between glucose homeostasis disruption and NADH/FSP1/COQ10 axis-driven ferroptosis. Int. J. Biol. Sci. 2026, 22, 43-59. [CrossRef]
- Liang, Q.H.; Liu, Q.Q.; Tian, S.Z.; Yao, Q.H.; Ye, X.Q.; Liu, W.C. Dietary fucoidan supplementation ameliorates heat stress-induced liver injury in broilers via modulating peroxidation, lipid metabolism, and ferroptosis. Poult. Sci. 2026, 105, 106489. [CrossRef]
- Zhao, Z.X.; Yuan, Y.M.; Zhao, Z.H.; Yao, Q.H.; Ye, X.Q.; Wang, Y.Y.; Liu, H.M.; Jha, R.; Balasubramanian, B.; Liu, W.C. Phlorotannin alleviates liver injury by regulating redox balance, apoptosis, and ferroptosis of broilers under heat stress. Antioxidants (Basel) 2024, 13, 1048. [CrossRef]
- Yan, L.; Zheng, H.; Chen, K.; Fang, M.; Ren, F.; Huang, T.; Cai, W.; Huang, K.; Wu, X.; Zheng, F. The copper transporter protein SLC31A1-mediated copper metabolism imbalance drives cuproptosis through the SIRT1/HMGB1 pathway in heart failure. Chem. Biol. Interact. 2026, 431, 112006. [CrossRef]
- Ma, G.; Li, Y.; Li, X.; Yu, D.; Chao, M.; Li, J.; Guo, J.; Wang, K.; Liu, Z.; Sun, R.; et al. Multi-ion channel nanomedicines targeting zinc transporter 1/ATPase copper transporters disrupt copper/iron homeostasis to enhance tumor immunotherapy. ACS nano 2026, 10.1021/acsnano.6c00358. [CrossRef]
- Xu, L.; Feng, J.; Xu, S.; Jin, M.; Bai, X.; Zhang, H.; Zhu, M.; Lin, S.; Song, J.; Lu, C. GSH-responsive triple-action photosensitizer nanoplatforms orchestrate cuproptosis-ferroptosis synergy to potentiate antitumor PDT efficacy. Journal of nanobiotechnology 2026, 10.1186/s12951-026-04331-3. [CrossRef]
- Lu, Y.; Wu, Y.; Yang, C.; Zhou, Y.; Ren, X.; Li, H.; Huang, H.; Pan, F.; Deng, A.; Lu, Y.; et al. Ferredoxins: master regulators in mitochondrial redox homeostasis and programmed cell death. Redox Biol 2025, 88, 103930. [CrossRef]
- Ding, S.; Hu, D.; Pan, Y.; Wang, J.; Wang, L. Vitisin A inhibits liver fibrosis by promoting Nrf2/HO-1 pathway and inhibiting Cuproptosis. Sci. Rep. 2025, 15, 44186. [CrossRef]
- Zha, Y.; Li, H.; Jiang, S.; Wang, Z.; Liu, Y.; Wei, Z.; Yang, Y. Research on the mechanism of Zuogui Wan in ameliorating oligoasthenozoospermia in rats via modulating cuproptosis-related pathways. J. Ethnopharmacol. 2026, 359, 120990. [CrossRef]
- Xu, F.; Xie, L.; Zhang, Z.; Yang, X.; Xu, X. Nicotine suppresses ferroptosis in colon cancer cells via HMOX1/NF-κB pathway to reduce oxaliplatin sensitivity. Apoptosis 2026, 31, 102. [CrossRef]
- Jin, C.; Li, J.; Han, S.; Bai, C.; Yang, J.; Wang, Z. HELLS inhibits autophagy-dependent ferroptosis in nasopharyngeal carcinoma by modulating the Nrf2/HO-1/GPX4 pathway. Int. J. Mol. Med. 2026, 57. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



