Submitted:
09 April 2026
Posted:
10 April 2026
You are already at the latest version
Abstract
Osmolytes, including proline, soluble sugars, and glycine betaine (GB), are essential for plant adaptation to environmental stress. They contribute to osmotic adjustment, membrane stabilization, and protection of cellular functions in arid and saline habitats. This study investigated major osmolytes in native Qatari plant species in natural field conditions and their physiological adaptation strategies. Significant interspecific variation indicated diverse mechanisms of stress acclimation. Although proline accumulation was common, it did not consistently correlate with salinity tolerance, which suggests that its accumulation may reflect stress-induced metabolic imbalance rather than being a reliable indicator of resistance. In contrast, the relative balance between soluble sugars and proline indicates coordinated carbon–nitrogen regulation that supports osmotic homeostasis and growth in fluctuating environmental conditions. Halophytic species exhibited distinct osmolyte profiles that highlight the potential role of additional compatible solutes (particularly GB) in stress adaptation. However, its occurrence and functional significance in these species have been insufficiently characterized. Given the predominance of C₃ photosynthesis in Qatari flora, GB may also help mitigate photorespiratory stress in extreme conditions. The findings expand the understanding of osmotic regulation in desert plants and highlight the potential of biotechnological approaches to enhance crop tolerance of harsh environments through manipulation of compatible solutes.
Keywords:
abiotic stress
; biochemical roles
; compatible solutes
; physiological roles
; salinity
; halophytes
; xerophytes
1. Introduction
In response to environmental stresses such as drought, salinity, and extreme temperatures, plants accumulate compatible solutes that help to stabilize cellular structures and maintain metabolic homeostasis. Once the stress is relieved, these solutes are either remobilized or catabolized, which leads to the release nitrogen and other nutrients back into the cellular pool. The reclaimed resources are subsequently redirected toward the synthesis of essential macromolecules, including proteins and nucleic acids, which supports cellular recovery and renewed growth.
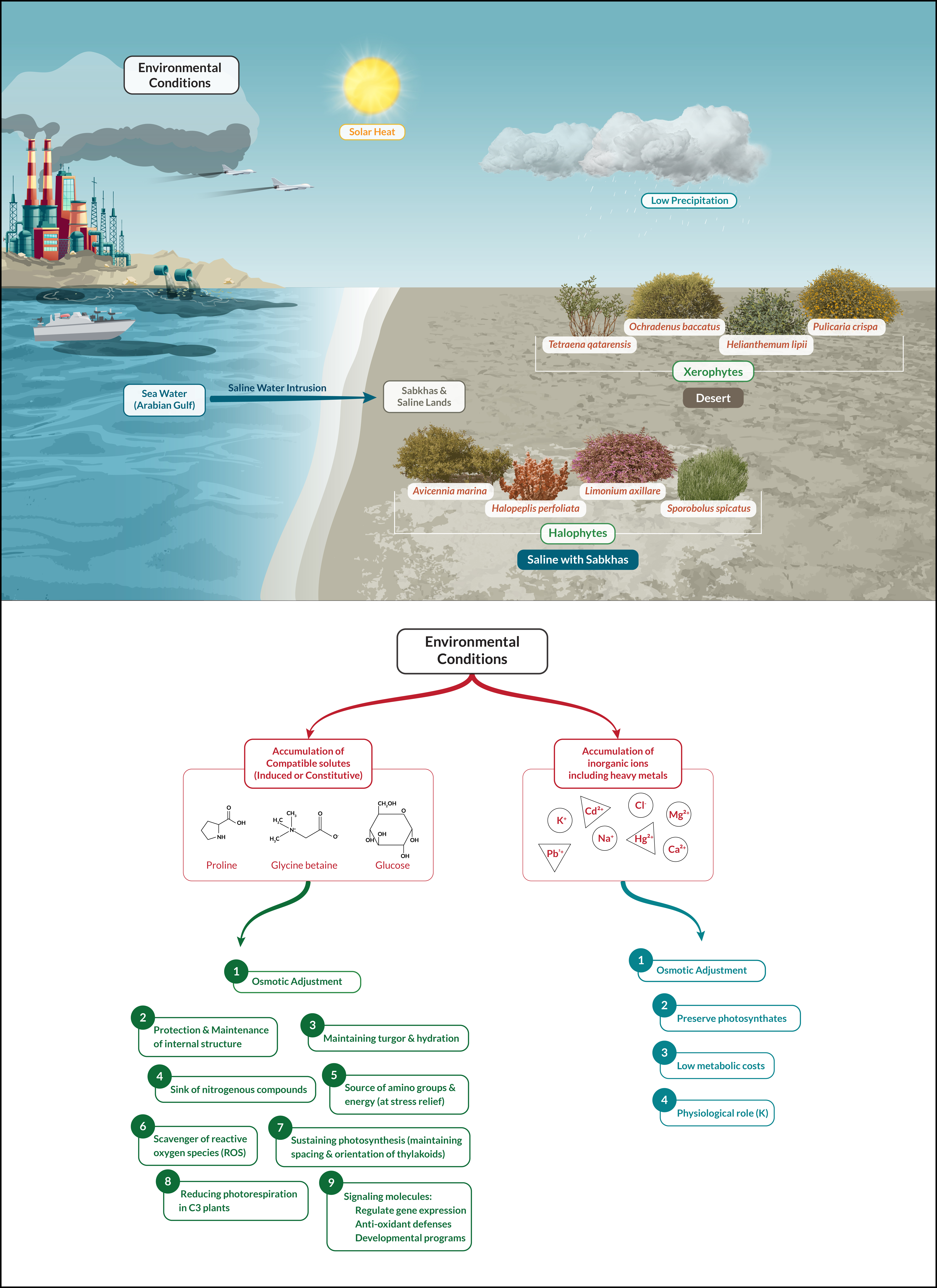
As part of these adaptive responses, plants synthesize and accumulate various organic compounds in their tissues to alleviate osmotic stress [1,2]. These compounds are typically grouped into three major categories: (1) organic acids, (2) nitrogenous compounds, and (3) soluble sugars [3,4,5]. Representative examples include organic acids such as malate, citrate, and succinate; nitrogenous compounds such as proline, glutamate, ornithine, and glycine betaine (GB); and soluble sugars such as sucrose, glucose, fructose, and trehalose. These metabolites play essential roles in maintaining osmotic homeostasis by regulating cellular water balance and stabilizing proteins and membranes in the context of stress-induced damage, which strengthens plant resilience and adaptive capacity in adverse environmental conditions [6,7]. Their accumulation significantly improves the plant’s tolerance of a broad spectrum of stresses, including salinity, drought, extreme and freezing temperatures, heavy metal toxicity, and anthropogenic and industrial pollution [1,8,9].
Studies have revealed that the organic solute content of plants vary according to the habitat conditions, and these solutes fulfil interchangeable physiological and biochemical roles during osmotic stress [10]. Osmoregulation in plant cells is achieved largely through the accumulation of compatible solutes such as proline, GB, soluble sugars, and essential inorganic ions such as potassium (K) [11]. These osmolytes also occur in associated microorganisms, including bacteria and fungi, where they perform similar protective and adaptive functions [1]. These mechanisms enable both plants and microbes to maintain cellular homeostasis and tolerate harsh environmental conditions [12]. This study examines the roles of compatible solutes in native Qatari plants—particularly xerophytes and halophytes—by exploring their types, functions, and patterns of accumulation across different plant tissues. The overall aim was to elucidate how these solutes contribute to stress tolerance in plants in diverse environmental conditions.
2. Adaptive Roles of Compatible Solutes in Native Plants
Among the major compatible solutes, proline, GB, and soluble sugars play central roles in plant responses to osmotic stress [13]. In both native plant species and their associated microorganisms, these solutes participate in several essential functions. One primary role is osmotic adjustment, which maintains osmotic homeostasis through the accumulation of organic and inorganic solutes that balance the osmotic potential between root tissues and the external environment [14]. Proline is particularly important in the event of drought and salinity stress because it decreases cellular osmotic potential, which enables continued water uptake and maintenance of turgor pressure [15,16]. By sustaining water absorption from dry or saline soils, proline supports long-term osmotic adjustment and prevents cellular dehydration [13]. GB and soluble sugars complement these effects by stabilizing proteins, membranes, and other cellular structures during stress.
A key component of effective osmotic regulation is the differential distribution of solutes across cellular compartments. Plant cells vary in their capacity to sequester solutes, which makes the regulation of their internal allocation central to achieving osmotic homeostasis [15,17]. GB shows a strong correlation with the osmotic potential of leaf-cell sap in many species, which emphasizes its role in cytoplasmic osmotic balance [18]. Histochemical and physiological studies demonstrate that GB accumulates primarily in the cytoplasm, which counterbalances the decreases in vacuolar osmotic potential that occur when inorganic ions (Na⁺, Cl⁻, Ca²⁺) accumulate [19]. This compartmentation prevents these ions from disrupting metabolic processes and preserves cytoplasmic integrity and turgor [20]. Recent studies have confirmed that higher GB concentrations are associated with reduced osmotic potential in stressed plant tissues and contribute to improved water retention and stress tolerance [11].
Inorganic ions predominantly accumulate in the vacuole, whereas compatible solutes—such as proline, GB, selected sugars, and K⁺—accumulate in the cytoplasm and counterbalance vacuolar ion loads without compromising metabolic functions. Compatible solutes do not interfere with enzyme activity even at high concentrations and safeguard cellular hydration and biochemical stability. This coordinated regulation of internal solute levels is widely recognized as osmoregulation, which is applicable to not only plant cells, but also animals and microorganisms [21,22].
Soluble sugars also contribute to osmotic adjustment, although their responses to stress vary between species and conditions. Sugars such as fructose, glucose, sucrose, and trehalose often increase in conditions of stress, and their contributions depend on the developmental stage, composition of the growth medium, and environmental factors. Research has shown that sugars account for 30–50% of the osmotic contribution in leaves of some glycophytes [3]. In contrast, halophytes often rely more on inorganic ions for osmotic balance, and organic solutes play a relatively small role [23,24].
The protection and maintenance of intracellular structures, including organelles and enzyme systems, are essential for cell survival [1,15]. Solutes such as proline play a key role in this process by safeguarding membranes and proteins from the detrimental effects of high concentrations of inorganic ions (e.g., Na, Cl, and certain heavy metals), as well as extreme temperatures. Rather than interacting directly with enzymes, proline acts primarily through preferential exclusion from the protein surface, which promotes proper hydration of proteins and stabilizes their native folded conformations. Proline also contributes to membrane integrity and helps to maintain overall cellular homeostasis under stress. Both proline and GB have been shown to effectively protect enzyme systems and organelles during heat stress [16]. These compatible solutes also act as multifunctional protectants during heat stress and preserve the structural and functional integrity of enzymes, membranes, and cell organelles, such as mitochondria, chloroplasts, the endoplasmic reticulum, and the cytoskeleton [26,27]. Their complementary biochemical properties make them some of the most effective natural solutes for protecting cells against temperature-induced damage.
During osmotic stress, proline accumulates markedly in most plant species, and recent research confirms that this amino acid functions as a key protective metabolite [25]. Its biosynthesis requires substantial metabolic investment in the form of ATP, which is stored in high-energy phosphate bonds, as well as NADPH, which provides reducing power. In the glutamate pathway, each proline molecule requires one ATP and two NADPH, which amount to roughly six ATP equivalents. In contrast, the ornithine pathway consumes only one NADPH and approximately 2.5 ATP equivalents per molecule of proline produced. Both routes also draw on central metabolites that are otherwise needed for tissue growth and essential physiological functions, including immune responses to biotic stress [28].
Although both pathways occur in most plants, their relative contributions depend on species, stress severity, and tissue nutrient status. Generally, the glutamate pathway predominates in conditions of osmotic stress when nitrogen is limited because it relies more directly on carbon skeletons rather than free nitrogen pools. Conversely, the ornithine pathway is favoured in nitrogen-rich tissues [29,30].
The of biosynthesis GB is metabolically and energetically costly. In terms of metabolite investment, its synthesis proceeds via the conversion of choline to GB through the following pathway: choline → betaine aldehyde → GB. This process which requires substantial energy input and the involvement of multiple metabolites. Notably, choline is also metabolically expensive, and its biosynthesis involves a series of reactions and the contribution of metabolites (serine → ethanolamine → phosphor-ethanolamine → choline).
Multiple SAM-dependent methylation reactions are involved, in which a methyl group (–CH₃) is transferred from S-adenosyl-L-methionine. Oxygen is required in some of these reactions. Moreover, reducing power is needed in the form of 2NAD+ or NADP+, and indirect use of ATP is required for the synthesis of choline and 3 SAM-dependent methylations, which each cost ~3 ATP equivalents. Altogether, about 12–15 ATP equivalents per GB molecule are required, and one nitrogen molecule is required to build one GB molecule [1,31,32].
Soluble sugars originate from photosynthetic carbon fixation and various reactions related to carbohydrate metabolism. The primary carbon skeletons of these compounds are generated through the Calvin cycle and other metabolic pathways, including the pentose phosphate pathway, which is also referred to as the phosphogluconate pathway or the hexose monophosphate (HMP) shunt. The roles and functions of soluble sugars have been discussed in detail [3,33].
The accumulation of compatible solutes to perform the roles discussed thus far imposes a substantial metabolic cost on plants under stress. Large amounts of energy and carbon metabolites are diverted toward the synthesis and maintenance of these solutes, which are essential for cellular protection, osmotic adjustment, and stress tolerance. Consequently, this metabolic investment can reduce the resources available for growth, development, and yield formation, which ultimately decreases plant productivity in conditions of prolonged or severe stress. Nevertheless, the considerable energy demand associated with the accumulation of compatible solutes such as proline, GB, and soluble sugars highlights their high adaptive value as these compounds contribute to osmotic balance, redox homeostasis, and the stabilization of proteins, membranes, and cellular structures in stress conditions.
Proline maintains turgor and hydration, and Paleg and Aspinall [34] proposed that it plays a key role in regulating cellular water balance in microstructures and plasma membranes. This mechanism is essential for sustaining cell turgor as proline interacts with hydrophobic protein surface residues and increases the overall hydrophilicity of associated molecules. These interactions improve the stability of cellular components in conditions of salinity and water-deficit stress [35,36].
Proline is a source of amino groups and energy, its concentration falls rapidly once the stress is relieved, and it can be readily utilized as a source of amino groups and energy [34]. One major pathway is the oxidation of proline in the mitochondria, where it is oxidized rapidly into glutamate as tissues become turgid. In this pathway, NADH is produced at the final stage of oxidation. The equation below summarizes this process:

Energy is released in the form of ATP when electrons are passed from NADH to O2 through the electron transport chain on the cristae of mitochondria. Glutamate is produced during proline oxidation and can be converted to α-ketoglutaric acid, which frees giving its amino group through a transamination process and allows it to enter the Krebs cycle. This is supported by the rapid evolution of CO2 when stressed plants regain access to water [33,37].
Proline accumulation under stress conditions can also function as a sink for nitrogenous compounds that are released during net protein loss. Stress increases protein degradation while inhibiting protein synthesis. This leads to increased availability of nitrogen, which is subsequently directed toward proline formation [38,39]. Compatible solutes such as proline act as an effective scavenger of reactive oxygen species (ROS), which helps to protect cells from oxidative stress.
Proline can interact directly with several types of ROS, including hydroxyl radicals (•OH), singlet oxygen (^1O₂), and superoxide anions (O₂•), which mitigates their damaging effects [40]. Through redox-mediated reactions, proline helps to neutralize reactive species and reduce their harmful effects, which protects key cellular macromolecules from oxidative damage, including lipids, proteins, nucleic acids, and membranes. In addition, proline influences protein solvation, acts as an efficient scavenger of hydroxyl radicals, and contributes to membrane stabilization through its interactions with phospholipids [15,41,42].
Some compatible solutes such as GB can sustain photosynthetic activity by maintaining chloroplast volume [43] or reducing photorespiration and increasing stomatal conductance [44], which stabilizes photosynthetic structures under stress [45]. Notably, GB supports photosynthesis in plants in the event of various types of environmental stresses by balancing the osmotic potential and preventing volume loss of cells and organelles. This is accomplished by maintaining optimal spacing and orientation of thylakoid membranes, which preserves efficient light reactions of photosynthesis.
C₄ plants are generally assumed to dominate saline and arid habitats due to their advantages in conditions of high temperature, intense light, drought, and salinity, but this trend does not apply to the Arabian Gulf. In the State of Qatar, C₃ species constitute the majority of the native flora among all documented and recognized plant species. As multifunctional signalling molecules, compatible solutes integrate metabolic, redox, and hormonal cues to regulate gene expression, antioxidant defences, and developmental programs. Their dynamic metabolism allows them to function as not only compatible solutes, but also as central regulators of cellular communication and stress adaptation [46].
2.1. Compatible Solutes as Indicators of Osmotic Stress Response
Some controversial evidence from numerous plant species, including species that are native to Qatar, suggests that the accumulation of compatible solutes may correlate with resistance, susceptibility, or visible injury when plants are exposed to harsh environmental conditions. This evidence is derived from histochemical, biochemical, and physiological studies that have been conducted on a wide range of plant taxa. The osmotic potential of vacuoles is primarily reduced by inorganic ions such as Na⁺ and Cl⁻, whereas organic compatible solutes predominantly accumulate in the cytoplasm [1,47]. Notably, the capacity to accumulate compatible solutes may represent a key parameter that distinguishes native species. Identifying the specific solutes involved and the genetic mechanisms underlying their accumulation in resilient native plants could provide valuable targets for improving stress tolerance in economically important crops. Such insights could have significant potential for developing cultivars that are capable of thriving in environments that are increasingly impacted by climate change and environmental degradation [47,48].
Proline has been extensively studied over the past five decades, but the results have often been divergent and sometimes controversial [39,49,50]. Studies suggest that proline accumulation in plants correlates with salt tolerance in halophytes [51]. This hypothesis is based on observations in the halophyte Triglochin maritima L., which exhibits low proline levels in non-saline conditions, but a progressive increase occurs as salinity increases. Furthermore, comparisons between inland and coastal populations of Armeria maritima Willd have demonstrated that the capacity to accumulate proline positively correlates with salt tolerance. Therefore, it was proposed that proline functions as an osmolyte in intracellular osmotic adjustment in saline conditions by reducing osmotic potential in plant tissues and thereby maintaining a favourable water-uptake gradient. This mechanism may represent an adaptive strategy that allows plants to withstand salinity and drought stress [52].
Other studies have supported the role of proline in resistance to environmental stresses. Dar et al. [53] provided evidence of a positive correlation between proline accumulation and plant stress tolerance. Proline plays important roles during stress conditions by acting as a metal chelator and as part of the antioxidative defence system. When applied exogenously at low concentrations, it improves stress tolerance. El-Moukhtari et al. [54] support the role of proline in improving salt tolerance in various plant species. Exogenous application of proline has also been shown to improve plant growth at different developmental stages, including germination, vegetative growth, and productivity. In saline conditions, proline treatment improves biomass accumulation, photosynthetic activity, and gas exchange.
Nguyen et al. [55] concluded that salt-resistant cultivars accumulate free proline as a strategy to cope with salinity stress by maintaining relative water content and cell-membrane integrity during exposure to salt stress. A recent study on cultivated eggplant and its wild relatives [56] demonstrated that salt-resistant plants accumulate high concentrations of proline under conditions of severe salinity stress. These findings may contribute to the development of salt-resistant crops as proline accumulation could be increased in salt-tolerant inbred lines and cultivars through interspecific hybridization.
On the other hand, several studies have reported that plants that are susceptible to osmotic stress accumulate higher levels of proline than resistant ones. For example, Yasseen [57] found that some Mexican wheat varieties that are sensitive to salt stress accumulate significantly greater amounts of proline compared to their more tolerant relatives (Table 1). In support of this finding, numerous studies and reviews have concluded that proline content does not consistently correlate with resistance to salinity and water stress.
The reported means of proline concentration were calculated from four experimental observations.
Although proline accumulation is a common plant response to stress, it is not always a reliable indicator or causal factor of osmotic stress resistance. In some cases, high proline levels have been associated with stress susceptibility rather than stress tolerance, and proline has been reported as an insensitive indicator of salt stress in certain plants, including soybean [58]. Data show that both a susceptible cultivar (Bragg) and a resistant cultivar (Ransom) have similar proline levels (0.4 μmol g⁻¹ fresh weight) under low salt stress (≤ 20 mM NaCl). However, at 40 and 60 mM NaCl, Bragg accumulates much higher proline concentrations (1.2 and 1.9 μmol g⁻¹ fresh weight, respectively), whereas proline levels in Ransom do not exceed 0.5 μmol g⁻¹ fresh weight, even at 100 mM NaCl. Proline accumulation occurred primarily in the susceptible cultivar when salt stress caused physiological injury. These results indicate that proline is not a sensitive or reliable indicator of salt stress tolerance in soybean plants.
Sundaresan and Sudhakaran [59] examined cassava plants and reported that drought resistance did not correlate with proline accumulation. Water stress induced a significant increase of proline content in leaves in both cultivars examined: the drought-susceptible M-4 and the drought-resistant S-1315. However, proline accumulation was substantially higher in the susceptible cultivar than in the resistant one: M-4 exhibited an approximately 25-fold increase in proline content, whereas the tolerant cultivar showed only about a 9-fold increase. The study also revealed cultivar-specific differences in the activities of enzymes involved in proline metabolism under water stress. These findings support the general conclusion that proline accumulation is primarily a consequence of stress rather than a direct cause of stress tolerance or salt resistance [60].
Although proline performs several protective functions, elevated proline levels are often associated with stress sensitivity rather than higher tolerance and acts as a symptom of cellular injury rather than an indicator of adaptive stress resistance [61,62]. Overall, proline accumulation represents a conserved yet largely non-specific metabolic response to diverse abiotic stresses, so its differential accumulation alone cannot be considered a reliable molecular marker for breeding stress-tolerant crops. This aligns with the concept that abiotic stress responses frequently converge in common metabolic and molecular pathways rather than reflecting stress-specific adaptations [63].
Notably, several studies have shown that salt-sensitive genotypes may accumulate higher levels of proline than tolerant ones, as discussed earlier, which indicates that proline concentration per se is not predictive of resistance. Although proline contributes to osmotic adjustment, ROS detoxification, and protein and membrane stabilization, its functional significance depends on the coordinated regulation of its biosynthesis, degradation, and cellular compartmentalization [64]. The contradictory outcomes reported in the literature over the past two decades likely arise from differences in experimental methodologies, growth and environmental conditions, species- and genotype-dependent responses, and limited integration of key regulatory variables, particularly the activities of enzymes controlling proline metabolism [49,65].
The roles of proline in various environmental conditions have been shown in many reports [15,66]. These stresses commonly induce proline accumulation via shared signal transduction pathways involving abscisic acid (ABA). Stress-induced ABA accumulation activates downstream signalling cascades that upregulate numerous genes, including those encoding key enzymes responsible for proline biosynthesis [38,67]. At the same time, ABA promotes stomatal closure [68] and triggers a range of physiological responses (some detrimental and others beneficial), which contribute to stress-related effects, as illustrated in Figure 1. These observations suggest that in some species, proline accumulation may be a consequence of stress-induced damage rather than a reliable indicator of salinity tolerance [61]. Furthermore, many halophytes accumulate only low levels of proline in their natural habitats compared with xerophytes and even xero-halophytes (Table 2).
Halophytes accumulate a variety of compatible solutes and inorganic ions that do not disrupt cytoplasmic metabolism, including proline, GB, soluble sugars, and potassium [69,70]. However, the synthesis of organic osmolytes is energetically expensive and depends on numerous metabolic intermediates, which helps to explain why many halophytes preferentially utilize abundant environmental ions (e.g., Na⁺, Cl⁻, Ca²⁺, and Mg²⁺) for bulk osmotic adjustment under saline conditions. Such plant species have evolved efficient systems for ion uptake, transport, compartmentation, and detoxification, which mainly occurs via vacuolar sequestration, whereas organic osmolytes function largely in cytosolic protection and stress mitigation. Because inorganic ions can accumulate in large quantities, they effectively support osmotic balance and help to maintain a favourable water-potential gradient between the plant and the environment [23,71,72].
Accordingly, the use of readily available salts instead of synthesizing organic osmolytes confers four principal advantages to these plants: (1) inorganic ions are readily available for uptake and do not require highly specialized structures or mechanisms, and (2) their uptake and translocation involve relatively low metabolic costs. (3) Dependence on inorganic ions for osmoregulation preserves photosynthates that would otherwise supply carbon skeletons and energy for growth-related biosynthesis, and (4) osmotic adjustment in roots through inorganic ions obviates the need to transport photosynthates from shoots to roots, thereby avoiding additional metabolic expenditure [73,74,75]. Nevertheless, halophytes also synthesize diverse organic solutes to satisfy physiological demands, which allows them to fulfil protective and regulatory roles in varying environmental conditions, as documented in numerous studies [13].
In halophytes, the accumulation of the organic solutes described thus far may limit plant productivity as these metabolites are largely directed toward osmotic regulation rather than growth, biomass accumulation, or yield formation. As a result, considerable energy and carbon are diverted to the production of protective compounds, which reduces their availability for core metabolic activities such as photosynthesis, cell proliferation, and structural development. In both halophytes and crop species cultivated on agricultural land, this internal redistribution of resources can ultimately limit productivity and yield, which poses a significant challenge to crop improvement in saline conditions [76]. Despite these physiological limitations, halophytes remain highly valuable as not only abundant sources of bioactive secondary metabolites [77,78,79], but also reservoirs of adaptive traits that can be harnessed to develop crop varieties that are suited for sustainable cultivation in saline soils [80].
Soluble sugars in plants such as glucose, fructose, sucrose, maltose, and trehalose also play essential roles in stress conditions [3]. They serve as energy sources and reserves, contribute to osmotic adjustment, and help stabilize cellular structures during stress. Their involvement in osmotic stress tolerance has been widely documented, particularly during two critical developmental stages: seed germination and vegetative growth. Increased levels of soluble sugar improve the ability of water-stressed plants to tolerate dehydration by contributing to osmotic adjustment [3]. As compatible solutes, these sugars help to maintain cellular turgor by facilitating water uptake from the soil, thereby alleviating the adverse effects of drought on growth and survival [81,82].
Consistently, Yasseen et al. [3] reported significant variation in compatible solute accumulation during seed germination under stress. Under mild osmotic stress, rapid degradation of storage reserves (starch, proteins, and lipids) increases metabolite availability, which supports cell division and seedling establishment. In contrast, high salinity suppresses reserve mobilization, limits metabolite supply, and constrains growth. Under such conditions, available resources are increasingly redirected toward the synthesis of compatible solutes, particularly proline, which may reduce the pool of soluble sugars available for germination and growth. Many of these protective functions overlap with those of proline. Therefore, the balance between sugar and proline accumulation reflects a coordinated carbon–nitrogen regulatory mechanism that allows plants to adjust osmotic regulation and growth in fluctuating stress conditions [36].
It is noteworthy that seeds with rich carbohydrate reserves, such as wheat and barley, accumulate proline by using carbon skeletons that are derived from soluble sugars and amino groups, which are supplied by the protein-rich aleurone layer. In contrast, seeds of fenugreek (Trigonella foenum-graecum L.) mobilize protein reserves to synthesize both sucrose and proline to supporting osmotic adjustment between the germinating seeds and the surrounding solution [83,84]. These observations suggest an exchange or shift in functional roles between proline and soluble sugars or nitrogen-containing compounds in key physiological and biochemical processes under osmotic stress [35,85]. In many plant systems, intracellular osmoregulation can be achieved primarily through the accumulation of either proline or soluble sugars. In such systems, particularly in cases of limited nitrogen availability, proline biosynthesis is largely supported by carbon derived from carbohydrate degradation [29].
During vegetative growth, plants depend on photosynthesis and associated metabolic pathways to generate the metabolites required for cellular maintenance. These processes produce soluble sugars and compatible solutes, such as proline, which contribute to osmotic adjustment in condition of salinity and drought stress [86]. When a local Yemeni cultivar of fenugreek is subjected to water deficit, monosaccharide and proline concentrations increase during vegetative growth, whereas sucrose levels remain relatively constant. This coordinated metabolic response helps to sustain cellular water status and protect intracellular structures [3,87]. Notably, the maintenance of stable sucrose concentrations likely supports continued carbon transport and allocation while contributing to osmotic balance, thereby reinforcing physiological stability in drought conditions.
GB is an osmo-protectant that accumulates in the cytoplasm of certain stress-tolerant crops and many native plant species, particularly halophytes, where it improves tolerance to abiotic stresses such as drought, salinity, and low temperature [26,45,88]. In contrast, crops including rice, potato, tomato, and tobacco synthesize little or no GB even under severe stress conditions, and genetically engineered crops generally accumulate significantly lower GB levels than halophytes [2,26,89,90]. Studies have suggested that GB and proline contribute to stress tolerance by stabilizing enzymes, membranes, and osmotic balance. Nevertheless, their adaptive roles are debated as some evidence indicates that their accumulation may be a consequence rather than a cause of stress tolerance [91].
The role of proline and GB has been examined in plants under extreme environmental conditions such as salinity, drought, extreme temperatures, UV radiation, and heavy metals, but their roles are still controversial. Both compounds have positive effects on enzyme and membrane integrity and adaptive roles in mediating osmotic adjustment in plants grown under stress conditions. Some studies have concluded that proline might play a role in helping plants to resist stress conditions, other studies have found opposite roles.
On the other hand, GB has consistently been reported as a compatible solute that accumulates in stress-resistant plants [40]. Supporting this view, exogenous GB application improves the growth of halophytes such as Suaeda salsa by improving osmotic stability and stimulating endogenous betaine synthesis, thereby significantly alleviating salt stress [92]. Accordingly, the application of GB to this species may contribute to the restoration of degraded S. salsa communities and serve as an adaptive strategy to cope with harsh environmental conditions, as reported in the Arabian Gulf region.
In summary, numerous studies have examined the roles of proline and GB in plants subjected to extreme environmental conditions, but their precise physiological significance remains a matter of debate. Both compounds are widely reported to improve stress tolerance by stabilizing proteins and membranes, protecting cellular structures, and contributing to osmotic adjustment under adverse conditions. However, while early research has emphasized a positive correlation between proline accumulation and stress resistance, subsequent findings have revealed inconsistencies, which suggest that proline build-up may in some cases reflect stress injury rather than adaptive capacity. In contrast, GB is more consistently recognized as an effective compatible solute in stress-tolerant species, where it plays a clear role in osmo-protection, maintenance of cellular homeostasis, and preservation of photosynthetic and metabolic functions. Overall, the accumulation of compatible solutes is a complex and context-dependent adaptive strategy that is influenced by species-specific traits, stress intensity, and developmental stage. Therefore, a balanced interpretation of solute accumulation is essential when assessing plant stress tolerance mechanisms [91,93,94].
3. Case Studies on Compatible Solutes in Native Plants in Qatar
The Arabian Gulf region in general and Qatar in particular are classified as arid to semi-arid. The region is among the warmest in the world, and its soils are typically saline, with ECe values (the electrical conductivity of the saturated soil extract) reaching up to 200 dSm⁻¹ [95,96]. However, some rawdahs exhibit low to moderate salinity levels that average around 4 dSm⁻¹ [35,97].
In a study on 23 native plant species, including both xerophytes and halophytes, Abdel-Bari et al. [35] reported clear differences in soil water content and ECe in the species’ natural habitats (Table 3). In general, Qatari soils are characteristically dry and saline. Soil moisture content ranges from 4% to 71% field capacity, while soil salinity (ECe) varies from 4 dS m⁻¹ to more than 200 dS m⁻¹. Rawdahs tend to have mildly saline soils with relatively high moisture content, whereas other parts of the country—particularly sabkhas and coastal areas—exhibit very high salinity levels. With these conditions, natural habitats in Qatar are predominantly arid and saline across most regions of the country.
The vegetation in Qatar is dominated by obligate halophytes and facultative xerophytes/halophytes that are well adapted to these extreme environmental conditions. This means that most of the plants in these areas are special types that can survive in very salty and very dry environments. Obligate halophytes are plants that must live in salty soils, are fully adapted to high salinity, and cannot survive in normal non-salty soils. Facultative xerophytes/halophytes are plants that can also grow in less harsh environments. These plants are well adapted to Qatar’s extreme conditions, such as high soil salinity and low water availability.
Variation in soil salinity and moisture across the country is mirrored in the diversity of native plant species. This variation influences their physiological and biochemical processes and leads to the build-up of different inorganic and organic solutes. Plants commonly accumulate soluble organic compounds in response to environmental stresses [1,3,34,71], yet Qatari native species vary in their ability to store particular solutes, including proline, GB, and soluble sugars [10,35,99,100,101]. This variability corresponds to their ecological strategies, which are shaped by characteristic morphological, anatomical, physiological, and biochemical traits [102,103].
The broader objective of research in this area is to identify traits that can be leveraged in modern biotechnology to improve plant adaptation—particularly in crops—to environmental challenges such as salinity, drought, and pollution [13]. Approaches such as genetic engineering and marker-assisted selection can be used to harness these naturally evolved adaptive mechanisms to strengthen crop performance and support global food security [104]. Studies and reports over the past two decades indicate that these solutes—particularly proline, soluble nitrogen compounds such as GB, and soluble sugars—can fulfil complementary or even interchangeable roles in a range of physiological and biochemical processes under osmotic stress [10]. On the other hand, differences in the concentrations of these solutes indicate that native plants differ in their capacity to accumulate organic solutes and to exchange functional roles (Table 4).
Plants under investigation exhibit varying capacities to accumulate proline. For example, Ochradenus baccatus has shown the highest proline accumulation among all species, followed by Tetraena qatarensis, Limonium axillare, Suaeda vermiculata, Salsola rosmarinus, Suaeda aegyptiaca, and Capparis spinosa. Lower proline levels have been recorded in Anabasis setifera, Cocculus pendulus, and Avicennia marina, while Pulicaria crispa exhibits the lowest concentration among all plants examined. Variability in proline accumulation in native plants can be attributed to several factors [105].
Differences in species-specific metabolic responses to their native environmental conditions help explain the observed variation [28]. Each species has unique metabolic adaptations that are shaped by its environment, which is why species do not all behave or function the same way [106]. In particular, plant species vary in their capacities for the synthesis, degradation, and transport of proline, which are processes that support functions such as osmoregulation, stress resistance, and energy acquisition [29]. These capacities are controlled by distinct sets of enzymes and transporter proteins, which result in substantial differences in proline accumulation even among species experiencing similar stress conditions, as shown in Table 4. Such variation reflects the unique biochemical machinery that each species uses for proline production, turnover, and intracellular transport. Because proline synthesis is energetically costly, stress-adapted species must balance this expense with their photosynthetic capacity and respiration rate, while the availability of nitrogen and carbon precursors further modulates accumulation.
The expression levels of key enzymes in these pathways also differ widely across plant taxa [63]. Additionally, species have evolved different levels of activation in proline-biosynthetic pathways in response to stress, with some lineages exhibiting markedly stronger induction under adverse conditions [107]. The types and strength of abiotic stress (such as water stress in xerophytes) decrease osmotic potential, which induces higher proline production. On the other hand, salinity causes both osmotic stress and ion toxicity, which lead to proline production [108]. Temperature extremes, light intensity, and soil nutrient levels also affect proline levels [66].
Energy limitations are another important factor since considerable metabolic expenditure is required for active ion transport, ion removal, and the build-up of organic osmolytes such as proline, GB, and various sugars [109]. As a result, although these compatible solutes are vital for stress tolerance, their synthesis and the related ion-regulating activities impose a significant energetic burden on the plant. This diversion of energy can restrict growth and reduce productivity in agricultural systems [1,110]. Additionally, the plants may possess inherent adaptations to drought and salinity stress that lessen their dependence on energetically costly proline accumulation [111].
The developmental stage of a plant also affects its proline accumulation. Seedlings, for instance, often exhibit higher proline levels during growth because they tend to be more sensitive to environmental stress, whereas mature tissues generally display lower or more stable concentrations. In natural ecosystems, however, accurately and consistently determining the developmental stage of each individual plant is challenging and complicates efforts to study patterns of proline accumulation [112]. Tissue-specific osmotic adjustment is common in halophytes, and leaves often accumulate more proline than roots because they are exposed to greater transpiration-driven stress. In many Qatari succulent halophytes, such as Limonium axillare, Suaeda species, Tetraena qatarensis, and possibly others (Table 4), proline and other compatible solutes accumulate and are stored in specialized water-storing tissues, which improves tolerance to salinity and drought [35,113].
Differences in morphological and osmotic adjustment strategies influence how plants manage osmotic stress. Not all species rely on proline for osmo-protection, and many halophytes synthesize other compatible solutes such as GB, soluble sugars, and polyols, which reduces their dependence on proline for osmoregulation. Halophytes may also accumulate inorganic ions to achieve osmotic adjustment in harsh saline conditions. In contrast, xerophytes often develop morphological and anatomical adaptations that lessen their need for biochemical osmo-protectants and reduce the demand for proline accumulation [114].
Proline levels in plants are strongly shaped by their ecological and evolutionary history, and species that evolved in harsh or variable environments often accumulate higher amounts as part of their stress-tolerance strategy. Even within the same species, native or locally adapted populations may show stronger proline responses because they regularly encounter environmental stress. Microhabitat differences can further influence proline accumulation, which means that even plants growing close to one another may display distinct proline levels due to subtle variations in their immediate surroundings [3,15,50,112].
According to compatible-solute analyses, species in the family Amaranthaceae generally accumulate substantially less proline than what is typically expected for plants that are exposed to pronounced drought or salinity stress. However, some members of this family are halophytes and exhibit xerophytic traits that enable them to accumulate appreciable amounts of proline. For example, Salsola rosmarinus and Suaeda spp. can accumulate proline levels of up to 400 μgg⁻¹ fresh weight. Species from other families, such as Limonium axillare (Plumbaginaceae), Ochradenus baccatus (Resedaceae), and Tetraena qatarensis (Zygophyllaceae), also accumulate high concentrations of proline, although among these species, only L. axillare is considered an obligate halophyte [35,51,115]. These observations indicate that proline accumulation is species-specific as some plants have regulatory mechanisms that are highly responsive to environmental stresses.
In contrast, several xerophytic species do not accumulate substantial amounts of proline in water-stress or saline conditions. For instance, Heliotropium bacciferum and Pulicaria spp. display minimal proline accumulation in arid or saline environments. Instead, these species may rely on other compatible solutes to maintain osmotic balance, such as soluble sugars or possibly GB [3,88]. These findings support earlier observations indicating that native plants differ in the types and amounts of organic solutes that they accumulate depending on habitat conditions. Furthermore, such solutes can fulfil interchangeable physiological and biochemical roles during osmotic stress [10].
Other native Qatari flora may accumulate compatible solutes as an adaptive strategy to tolerate salinity and arid environments. Yasseen and Al-Thani have documented several of these species [12], and Table 5 summarizes these plants and their potential roles in the remediation of soils contaminated by salinity, petroleum hydrocarbons, and heavy metals. The table also highlights the reported or putative presence of compatible solutes in these species (particularly GB).
3.1. Glycine Betaine Accumulation in Native Qatari Plants
GB is part of a group of compatible osmolytes that include proline betaine, β-alanine betaine, choline-O-sulphate, and the tertiary sulphonium compound 3-dimethylsulphoniopropionate (DMSP) [31]. Quaternary ammonium compounds play an important role in cytoplasmic osmotic adjustment in plants that are subjected to osmotic stress. These compounds are widely distributed in nature and occur at relatively high concentrations in marine invertebrates, fish, and certain plant species compared with microorganisms and terrestrial vertebrates [162,163]. GB accumulation has frequently been associated with halophytic and (to some extent) xerophytic traits in plants that inhabit saline and arid environments [164]. In many species, particularly those in families Amaranthaceae, Chenopodiaceae, Gramineae, and Poaceae, GB accumulates at substantial levels ranging from >5 to 100 µmol g⁻¹ dry weight (DW), and plants that are capable of synthesizing GB are generally better adapted to saline conditions. In most cases, GB accumulates at higher concentrations in shoots than in roots [165].
The biosynthesis of GB involves several enzymatic steps and metabolic intermediates, which make the process energetically demanding [158]. It begins with the sequential methylation of ethanolamine to produce choline, which is then oxidized to form GB. The primary precursors for GB synthesis include glycine, serine, and formate. Ethanolamine is derived from glycine via serine in a process where formate donates a one-carbon unit during the conversion of glycine to serine, followed by the decarboxylation of serine to produce ethanolamine [166]. These metabolites are also intermediates in the photorespiratory carbon cycle [33]. Although metabolically costly, this pathway allows plants to maintain osmotic balance and survive in conditions of salinity and drought stress [167].
The biosynthesis of GB in plants that are native to harsh environments may also be linked to broader metabolic processes associated with photosynthesis. Most native plant species in Qatar utilize the C3 photosynthetic pathway, with fewer species employing C4 or CAM pathways [98,168]. Therefore, investigating GB accumulation in these plants is particularly relevant as C3 species undergo photorespiration, which is a process inherent to C3 photosynthesis.
Although often considered energetically inefficient in terms of carbon assimilation, photorespiration performs several important physiological roles [33,169]. It contributes to the biosynthesis of amino acids and organic acids and helps to maintain cellular redox balance by dissipating excess ATP and reducing equivalents when carbon fixation is restricted. By functioning as a metabolic safety valve under stress conditions, photorespiration prevents over-reduction of the photosynthetic apparatus and limits the formation of ROS, such as superoxide (O₂⁻), thereby protecting cellular structures from oxidative damage [170]. Thus, assessing GB levels in native Qatari plants is important as this compatible solute has rarely been investigated in plants of the Arabian Gulf, particularly those employing the C3 photosynthetic pathway.
Relatively few studies in the Arabian Gulf region have examined GB accumulation in native plants, including both halophytes and xerophytes, to determine their capacity to synthesize this osmolyte and the mechanisms underlying its accumulation. As shown in Table 5, both C3 and C4 plants can accumulate GB in their tissues, although comprehensive evaluation of all native plant species in the region is still required. Notably, rather than proline, some plants accumulate GB because it provides particularly effective protection against environmental stress [2,91,171].
The concentration of GB in plant tissues often reflects the plant’s stress tolerance capacity and contributes to resilience in harsh conditions. GB mitigates oxidative damage by detoxifying ROS and supporting the recovery of photosynthesis. It also stabilizes cellular membranes, proteins, and key photosynthetic components under stress conditions, such as ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) and photosystem II. Even at relatively low concentrations, GB can activate stress-response genes that enhance plant defence mechanisms. As a result, GB accumulation improves tolerance to abiotic stresses such as salinity and drought and has been associated with improved crop yield and quality. These effects highlight the importance of understanding and potentially engineering GB biosynthesis in plants [158,172,173].
The native plants in Table 4 show substantial variations in their proline levels. Therefore, determining GB levels in these plants is essential to provide a more complete understanding of the role of these compatible solutes in the environmental conditions of the Arabian Gulf, particularly since soluble sugars may play only a limited role [2,6,91,136]. Table 5 shows that many plant species, including both C3 and C4 types, are capable of accumulating GB in their tissues. Several species Qatari flora have been identified as GB accumulators, which contributes to stress tolerance and improves resilience in native plants. These include Atriplex spp., Capparis spinosa, Cleome spp., Frankenia pulverulenta, Halopyrum mucronatum, Haloxylon salicornicum, Ochradenus baccatus, Pulicaria spp., Salicornia spp., Salsola spp., Sporobolus spicatus, Suaeda spp., Tamarix spp., and Tetraena qatarensis.
Accumulation of GB in these species may occur both constitutively and in response to environmental stimuli as various stress conditions can induce its synthesis, such as salinity, drought, high temperatures, and pollution from petroleum hydrocarbons and heavy metals [1,118]. Therefore, examining this compatible solute in native plants in Qatar and other Arabian Gulf countries is essential to better understand its distribution and potential roles in the regional flora. To the best of our knowledge, GB has only been investigated in a limited number of native plant species in Qatar, which highlights the need for further studies to elucidate its functional significance in these plants.
4. Concluding Remarks and Future Perspectives
Over the past five decades, numerous studies have reported substantial variation in osmolyte levels. However, their outcomes and conclusions have often been inconsistent or even contradictory, which has led to divergent interpretations. Despite these discrepancies, there is broad agreement that compatible solutes, such as proline, GB, and soluble sugars, play a vital role in plant adaptation and can improve tolerance to extreme environmental conditions. The accumulation of these osmolytes in plant tissues, particularly in species that are adapted to saline and arid environments, represents a key adaptive strategy.
Modern biotechnological approaches should be further explored to develop crops with greater capacity to accumulate these osmolytes, thereby improving resilience and sustaining growth under adverse conditions. Genetic and biological strategies are promising tools to achieve this objective, particularly in the face of escalating global challenges such as population growth, environmental pollution, climate change, desertification, and increasing soil salinity. Genetic manipulation of plants is widely regarded as a potential solution for mitigating the adverse impacts of environmental stressors. Biological approaches have emerged as an effective means of addressing major environmental issues, including salinity stress and the contamination of ecosystems by petroleum hydrocarbons and heavy metals [1,8,174,175]. Such approaches should concentrate on native plants that are able to accumulate GB either through induction or constitutively as this compatible solute provides effective protection against environmental stresses, including pollution resulting from human activities or wars.
Funding
This research received no external funding.
Institutional Review Board Statement
No ethical approval was required for this study as it relied solely on publicly available data.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, and further inquiries can be directed to the corresponding author.
Acknowledgments
The authors thank Qatar University for its continuous support and encouragement of scientific publications. They also express their gratitude to Nada Abbara for designing Figure 1 and the graphical abstract (GA).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Al-Thani, R.F.; Yasseen, B.T. Solutes in Native Plants in the Arabian Gulf Region and the Role of Microorganisms: Future Research. J. Plant Ecology 2018, 11, 671–684. [Google Scholar] [CrossRef]
- Basit, F.; Alyafei, M.; Hayat, F.; Al-Zayadneh, W.; El-Keblawy, A.; Sulieman, S.; Sheteiwy, M.S. Deciphering the role of glycine betaine in enhancing plant performance and defense mechanisms against environmental stresses. Front. Plant Sci. 2025, 16, 1582332. [Google Scholar] [CrossRef]
- Yasseen, B.T.; Al-Thani, R.F.; Alhadi, F.A.; Abbas, R.A.A. Soluble sugars in plants under stress at the Arabian Gulf region: Possible roles of microorganisms. J. Plant Biochem. Physiol. 2018, 6, 224. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Li, R.; Ge, Y.; Li, Y.; Li, R. Plants’ response to abiotic stress: Mechanisms and strategies. Int. J. Mol. Sci. 2023, 24, 10915. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Maurya, V.; Gupta, K.; Sharma, I.; Sharma, A.; Kumar, R. Salt stress and its eco-friendly management using biostimulants in grain legumes: a review. Discov. Agric. 2025, 3, 13. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. Journal of Experimental Botany 2012, 63, 1593–1608. [Google Scholar] [CrossRef]
- Raza, A.; Anas, M.; Bhardwaj, S.; Mir, R.A.; Charagh, S.; Elahi, M.; Zhang, X.; Mir, R.R.; Weckwerth, W.; Fernie, A.R.; Siddique, K.H.M.; Hu, Z.; Varshney, R.K. Harnessing metabolomics for enhanced crop drought tolerance. The Crop Journal 2025, 13, 311–327. [Google Scholar] [CrossRef]
- Al-Thani, R.F.; Yasseen, B.T. Phytoremediation of Polluted Soils and Waters by Native Qatari Plants: Future Perspectives. Environmental Pollution 2020, 259, 113694. [Google Scholar] [CrossRef]
- Al-Thani, R.F.; Yasseen, B.T. Possible Future Risks of Pollution Consequent to the Expansion of Oil and Gas Operations in Qatar. Environment and Pollution 2023, 12, 12–52. [Google Scholar] [CrossRef]
- Yasseen, B.T. Urban Development Threatening Wild Plants in Doha City-Qatar: Ecophysiology is a Prerequisite for Ecological Restoration. Journal of Plant Sciences 2011, 6, 113–123. [Google Scholar] [CrossRef]
- Kumar, V.; Khare, T.; Shaikh, S.; Wani, S.H. Compatible Solutes and Abiotic Stress Tolerance in Plants. In Metabolic Adaptations in Plants During Abiotic Stress; pp. 221–228. [CrossRef]
- Yasseen, B.T.; Al-Thani, R.F. Endophytes and Halophytes to Remediate Industrial Wastewater and Saline Soils: Perspectives from Qatar. PLANTS 2022, 11, 1497. [Google Scholar] [CrossRef]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Khan, M.A.R. Understanding the roles of osmolytes for acclimatizing plants to changing environment: a review of potential mechanism. Plant Signal. Behav. 2021, 16, 1913306. [Google Scholar] [CrossRef]
- Silveira, J.A.G.; Araujo, S.A.M.; Lima, J.P.M.S.; Viegas, R.A. Roots and leaves display contrasting osmotic adjustment mechanisms in response to NaCl-salinity in Atriplex nummularia. Environmental and Experimental Botany 2009, 66, 1–8. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: a review. Plant Signal Behav. 2012, 7, 1456–66. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, Y.K. Proline as a key player in heat stress tolerance: insights from maize. Discov. Agric. 2024, 2, 121. [Google Scholar] [CrossRef]
- Khan, P.; Abdelbacki, A.M.M.; Albaqami, M.; Jan, R.; Kim, K.-M. Proline promotes drought tolerance in Maize. Biology 2025, 14, 41. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Abbas, Z.; Seleiman, M.F.; Rizwan, M.; Yavas, İ.; Alhammad, B.A.; Shami, A.; Hasanuzzaman, M.; Kalderis, D. Glycine betaine accumulation, significance and interests for heavy metal tolerance in plants. Plants (Basel) 2020, 9, 896. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, R.; Liu, T.; Zheng, H-L. Solute accumulation and osmotic adjustment characteristics of the mangrove Avicennia marina under NaCl-induced salinity stress. Botanica Marina 2011, 54, 335–341. [Google Scholar] [CrossRef]
- Mehta, D.; Vyas, S. Comparative bio-accumulation of osmo-protectants in saline stress tolerating plants: A review. Plant Stress 2023, 9, 100177. [Google Scholar] [CrossRef]
- Munns, R.; Passioura, J.B.; Colmer, T.D.; Byrt, C.S. Osmotic adjustment and energy limitations to plant growth in saline soil. New Phytologist 2019, 225, 1091–1096. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, J-G. Osmotic adjustment and plant adaptation to environmental changes related to drought and salinity. Environmental Reviews 2025a, 18, 309–319. [Google Scholar] [CrossRef]
- Xu, C.; Tang, X.; Shao, H.; Wang, H. Salinity tolerance mechanism of economic halophytes from physiological to molecular hierarchy for improving food quality. Curr. Genomics 2016, 17, 207–14. [Google Scholar] [CrossRef]
- Sun, L.; Wang, J.; Lian, L.; Song, J.; Du, X.; Liu, W.; Zhao, W.; Yang, L.; Li, C.; Qin, Y.; Yang, R. Systematic analysis of the sugar accumulation mechanism in sucrose- and hexose- accumulating cherry tomato fruits. BMC Plant Biol. 2022, 22, 303. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Javid, M.G.; Soltani, E.; Wojtyla, L.; Małgorzata Garnczarska, M. Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants: A Review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Wani, S.H.; Singh, N.B.; Haribhushan, A.; Mir, J.I. Compatible solute engineering in plants for abiotic stress tolerance—role of glycine betaine. Curr. Genomics 2013, 14, 157–65. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S.; Devi, P.; Bhardwaj, A.; Jha, U.C.; Sharma, K.D.; Prasad, P.V.V.; Siddique, K.H.M.; Bindumadhava, H.; Kumar, S.; Nayyar, H. Identification and characterization of contrasting genotypes/cultivars for developing heat tolerance in agricultural crops: Current status and prospects. Front. Plant Sci. 2020, 11, 587264. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [PubMed]
- Kavi Kishor, P.B.; Sangam, S.; Amrutha, R.N.; Sri Laxmi, P.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, P.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Science 2005, 88, 424–438. [Google Scholar]
- Mushtaq, N.U.; Saleem, S.; Rasool, A.; Shah, W.H.; Tahir, I.; Seth, C.S.; Rehman, R.U. Proline tagging for stress tolerance in plants. Int. J. Genomics 2025, 2025, 9348557. [Google Scholar] [CrossRef]
- Yu, J.; Li, Y.; Tang, W.; Liu, J.; Lu, B-R.; Liu, Y. The Accumulation of Glycine Betaine Is Dependent on Choline Monooxygenase (OsCMO), Not on Phosphoethanolamine N-Methyltransferase (OsPEAMT1), in Rice (Oryza sativa L. ssp. japonica). Plant Molecular Biology Reporter 2014, 32, 916–922. [Google Scholar] [CrossRef]
- Gu, J.; Ma, X.; Liu, J.; Ma, M. Physiological and molecular mechanisms of glycine betaine in alleviating Na2SO4 stress in Glycyrrhiza uralensis. Front Plant Sci. 2025, 16, 1667006. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Paleg, L.G.; Aspinall, D. The Physiology and Biochemistry of Drought Resistance in Plants; Academic Press: Sydney, New York, London, 1981. [Google Scholar]
- Abdel Bari, E.M.; Yasseen, B.T.; Al-Thani, R.F. Halophytes in the State of Qatar (Sponsored by Environmental Studies Center); Qatar University: Qatar, 2007. [Google Scholar]
- Siddique, A.; Kandpal, G.; Kumar, P. Proline accumulation and its defensive role under diverse stress condition in plants: An overview. J. Pure Appl. Microbiol. 2018, 12, 1655–1659. [Google Scholar] [CrossRef]
- Pallag, G.; Nazarian, S.; Ravasz, D.; Bui, D.; Komlódi, T.; Doerrier, C.; Gnaiger, E.; Seyfried, T.N.; Chinopoulos, C. Proline Oxidation Supports Mitochondrial ATP Production When Complex I Is Inhibited. Int. J. Mol. Sci. 2022, 23, 5111. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Trovato, M.; Forlani, G.; Signorelli, S.; Funk, D. Proline metabolism and its functions in development and stress tolerance. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants; Springer Nature: Switzerland AG, 2019; pp. 41–72. [Google Scholar] [CrossRef]
- Renzetti, M.; Funck, D.; Trovato, M. Proline and ROS: A unified mechanism in plant development and stress response? Plants (Basel) 2024, 14, 2. [Google Scholar] [CrossRef]
- Krishnan, N.; Dickman, M.B.; Becker, D.F. Proline modulates the intracellular redox environment and protects mammalian cells against oxidative stress. Free Radic. Biol. Med. 2008, 44, 671–81. [Google Scholar] [CrossRef]
- Dutta, T.; Neelapu, N.R.R.; Wani, S.H.; Challa, S. Chapter 11—Compatible Solute Engineering of Crop Plants for Improved Tolerance Toward Abiotic Stresses. In Biochemical; Wani, S.H., Ed.; Physiological and Molecular Avenues for Combating Abiotic Stress Tolerance in Plants, 2018; pp. 221–254. [Google Scholar] [CrossRef]
- Rajasekaran, L.R.; Kriedemann, P.E.; Aspinall, D.; Paleg, L.R. Physiological significance of proline and Glycinebetaine: Maintaining photosynthesis during NaCL stress in wheat. Photosynthetica 1997, 34, 357–366. [Google Scholar] [CrossRef]
- Makela, P.; Kontturi, M.; Pehu, E.; Somersalo, S. Photosynthetic response of drought- and salt-stressed tomato and turnip rape plants to foliar-applied glycinebetaine. Physiol. Plant 1999, 105, 45–50. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef]
- Singhal, R.K.; Saha, D.; Skalicky, M.; Mishra, U.N.; Chauhan, J.; Behera, L.P.; Lenka, D.; Chand, S.; Kumar, V.; Dey, P.; Indu; Pandey, S.; Vachova, P.; Gupta, A.; Brestic, M.; El-Sabagh, A. Crucial signalling compounds crosstalk and integrative multi-omics techniques for salinity stress tolerance in plants. Front. Plant Sci. 2021, 12, 670369. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Cuin, T.A.; Zhou, M.; Twomey, A.; Naidu, B.P.; Shabala, S. Compatible solute accumulation and stress-mitigating effects in barley genotypes contrasting in their salt tolerance. Journal of Experimental Botany 2007, 58, 4245–4255. [Google Scholar] [CrossRef] [PubMed]
- Adzigbe, J.; Frimpong, F.; Danquah, A.; Danquah, E.Y.; Asante, I.K.; Abebrese, S.O.; Dormatey, R.; Afriyie-Debrah, C.; Ribeiro, P.F.; Danquah, E.O.; Agyeman, K.; Ralph Kwame Bam, R.K.; Asante, M.D. The responses and adaptations of rice (Oryza sativa L.) to drought stress: A review. Climate Smart Agriculture 2025, 2, 100080. [Google Scholar] [CrossRef]
- Yasseen, B.T. Further follow-up on proline accumulation in plants. Seminar presented at the Department of Biological & Environmental Sciences, Qatar University, Doha, Qatar, 2004. [Google Scholar]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: a review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Stewart, G.R.; Lee, J.A.T. The role of proline accumulation in halophytes. Planta 1974, 120, 279–289. [Google Scholar] [CrossRef]
- Trovato, M.; Mattioli, R.; Costantino, P. Multiple roles of proline in plant stress tolerance and development. Rend. Fis. Acc. Lincei 2008, 19, 325–346. [Google Scholar] [CrossRef]
- Dar, M.I.; Naikoo, M.I.; Rehman, F.; Naushin, F.; Khan, F.A. Proline Accumulation in Plants: Roles in Stress Tolerance and Plant Development. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Iqbal, N., Nazar, R., Khan, A.N., Eds.; Springer: New Delhi, 2016. [Google Scholar] [CrossRef]
- El-Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savoure, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Bhowmik, S.D.; Long, H.; Cheng, Y.; Mundree, S.; Hoang, L.T.M. Rapid accumulation of proline enhances salinity tolerance Australian wild rice Oryza australiensis Domin. Plants 2021, 10, 2044. [Google Scholar] [CrossRef]
- Cebeci, E.; Boyaci, H.F.; Kiran, S.; Elliaitioglu, S.S. Comprehensive assessment to reveal that salt tolerance potential of cultivated eggplants and their wild relatives. Fron. Plant. Sci. 2025, 16, 1483409. [Google Scholar] [CrossRef]
- Yasseen, B.T. An analysis of the effects of salinity on leaf growth in Mexican wheats. Ph.D. thesis, The University of Leeds, United Kingdom, 1983. [Google Scholar]
- Moftah, A.E.; Michel, B.E. The effect of sodium chloride on solute potential and proline accumulation in soybean leaves. Plant Physiol. 1987, 83, 238–40. [Google Scholar] [CrossRef]
- Sundaresan, S.; Sudhakaran, P.R. Water stress-induced alterations in the proline metabolism of drought-susceptible and -tolerant cassava (Manihot esculenta) cultivars. Physiol. Plant 1995, 94, 635–642. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, J.K. Proline accumulation and salt-stress-induced gene expression in a salt-hypersensitive mutant of Arabidopsis. Plant Physiol. 1997, 114, 591–596. [Google Scholar] [CrossRef]
- Lutts, S.; Majerus, V.; Kinet, J.M. NaCl effects on proline metabolism in rice (Oryza sativa) seedlings. Physiologia Plantarum 1999, 105, 450–458. [Google Scholar] [CrossRef]
- Yaish, M. Proline accumulation is a general response to abiotic stress in the date palm tree (Phoenix dactylifera L.). Genetics and Molecular Research 2015, 14, 9943–9950. [Google Scholar] [CrossRef]
- Kavi Kishor, P.B.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2015, 37, 300–11. [Google Scholar] [CrossRef] [PubMed]
- Spormann, S.; Nadais, P.; Sousa, F.; Pinto, M.; Martins, M.; Sousa, B.; Fidalgo, F.; Soares, C. Accumulation of proline in plants under contaminated soils-are we on the same page? Antioxidants (Basel) 2023, 12, 666. [Google Scholar] [CrossRef]
- Masheva, V.; Spasova-Apostolova, V.; Aziz, S.; Tomlekova, N. Variations in proline accumulation and relative water content under water stress characterize bean mutant lines (P. vulgaris L.). Bulg. J. Agric. Sci. 2022, 28, 430–436. [Google Scholar]
- Raza, A.; Charagh, S.; Abbas, S.; Hassan, M.U.; Saeed, F.; Haider, S.; Sharif, R.; Anand, A.; Corpas, F.J.; Jin, W.; Varshney, R.K. Assessment of proline function in higher plants under extreme temperatures. Plant Biol. (Stuttg) 2023, 25, 379–395. [Google Scholar] [CrossRef]
- Rai, G.K.; Khanday, D.M.; Choudhary, S.M.; Kumar, P.; Kumari, S.; Martinez-Andújar, C.; Martinez-Melgarejo, A.; Rai, P.K. Unlocking nature’s stress buster: Abscisic acid’s crucial role in defending plants against abiotic stress. Plant Stress 2024, 11, 100359. [Google Scholar] [CrossRef]
- Bharath, P.; Gahir, S.; Raghavendra, A.S. Abscisic acid-induced stomatal closure: an important component of plant defense against abiotic and biotic stress. Front. Plant Sci. 2021, 12, 615114. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Khalid, M.F.; Muhammad, S.; Anjum, M.A. Role of transporters in accumulating salt ions by halophytes. In Approaches of the Remediation of Inorganic Pollutants; Hasanuzzaman, M., Ed.; Springer: Singapore, 2021; pp. 11–40. [Google Scholar] [CrossRef]
- Rahman, M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, N.; Ansary, M.U.; Das, A.K.; Rahman, A.; Tran, L.S.-P. Adaptive mechanisms of halophytes and their potential in improving salinity tolerance in plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, E.T.; Orcutt, D.M. Physiology of Plants under Stress, Abiotic Factors; John Wiley & Sons, Inc.: New York, USA, 1996. [Google Scholar]
- Larcher, W. Physiological Plant Ecology. Eco-physiology and Stress Physiology of Functional Groups, 4th ed.; Springer: Berlin, 2003. [Google Scholar]
- Epstein, E. Crops tolerant of salinity and other mineral stresses. Ciba Foundation Symposium 97. Better crops for food; Pitman: London, 1983; pp. 61–82. [Google Scholar]
- Yasseen, B.T.; Abu-Al-Basal, M.A. Ecophysiology of Chenopodiaceae at the Coastline of Arabian Gulf-Qatar: Possible Destruction and Conservation Perspective. European Journal of Scientific Research 2010, 39, 90–104. [Google Scholar]
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ response mechanisms to salinity stress. Plants (Basel) 2023, 12, 2253. [Google Scholar] [CrossRef] [PubMed]
- Gul, B.; Hameed, A.; Ahmed, M.Z.; Hussain, T.; Rasool, S.G.; Nielsen, B.L. Thriving under salinity: Growth, ecophysiology and proteomic insights into the tolerance mechanisms of obligate halophyte Suaeda fruticosa. Plants (Basel) 2024, 13, 1529. [Google Scholar] [CrossRef]
- Al-Thani, R.F.; Yasseen, B.T. Cyanoremediation of polluted seawater in the Arabian Gulf: Risks and benefits to human health. Processes 2024, 12, 2733. [Google Scholar] [CrossRef]
- Al-Thani, R.F.; Yasseen, B.T. The Role of Phytoplankton in Phycoremediation of Polluted Seawater: Risks, Benefits to Human Health, and a Focus on Diatoms in the Arabian Gulf. Water 2025, 17, 920. [Google Scholar] [CrossRef]
- Atzori, G.; Garcia-Caparros, P.; Castagna, A.; Custodio, L. Salt-induced nutritional and metabolic shifts in halophytes: implications for food security. Journal of the Science of Food and Agriculture 2025. [Google Scholar] [CrossRef]
- Yasseen, B.T.; Al-Thani, R.F. Wild Plants in the Qatari Peninsula are Hidden Gene Bank for Future Research: Perspectives of Desirable Traits, Chapter 6. In Cutting Edge Research in Biology; 2023; Vol. 8. B P International pp, pp. 207–252. [Google Scholar] [CrossRef]
- Gurrieri, L.; Merico, M.; Trost, P.; Forlani, G.; Sparla, F. Impact of Drought on Soluble Sugars and Free Proline Content in Selected Arabidopsis Mutants. Biology (Basel) 2020, 9, 367. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, R.; Li, X.; Dong, D.; Wang, S. Sugar metabolism and transport in response to drought-rehydration in Poa pratensis. Agronomy 2025, 15, 320. [Google Scholar] [CrossRef]
- Alhadi, F.A.; Yasseen, B.T.; Al-Dubaie, A.S. Changes in carbohydrate and nitrogen fractions during germination of fenugreek (Trigonella foenum-graecum L.) seeds presoaked in GA3, growing under different Osmotic potentials. Qatar Univ. Sci. J. 1997, 17, 271–279. [Google Scholar]
- Gill, P.K.; Sharma, A.D.; Singh, P.; Bhullar, S.S. Changes in germination, growth and soluble sugar contents of Sorghum bicolor (L.) Moench seeds under various abiotic stresses. Plant Growth Regulation 2003, 40, 157–162. [Google Scholar] [CrossRef]
- Yasseen, B.T.; Almuhannady, A.; Al-Marri, F.R.; Al-Hemiary, H. Changes in soluble sugars and proline in seedlings of a local wheat cultivar (Doha) due to the salt stress and temperature. Qatar Univ. Sci. J. 2006, 26, 71–82. [Google Scholar]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Int. J. Genomics 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Alhadi, F.A.; Yasseen, B.T.; Gabr, M.M. Water stress and gibberellic acid effects on growth of fenugreek plants. Irrig. Sci. 1999, 18, 185–190. [Google Scholar] [CrossRef]
- Jarin, A.; Ghosh, U.K.; Hossain, M.S.; Mahmud, A. Glycine betaine in plant responses and tolerance to abiotic stresses. Discover Agriculture 2024, 2, 127. [Google Scholar] [CrossRef]
- Park, E-J.; Jeknic, Z.; Sakamoto, A.; DeNoma, J. Genetic engineering of glycine betaine synthesis in tomato protects seeds, plants, and flowers from chilling damage. The Plant Journal 2004, 40, 474–487. [Google Scholar] [CrossRef]
- Khan, M.S.; Yu, X.; Kikuchi, A.; Asahina, M.; Watanabe, K.N. Genetic engineering of glycine betaine biosynthesis to enhance abiotic stress tolerance in plants. Plant Biotechnology 2009, 26, 125–134. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental & Experimental Botany 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Dong, X.; Liu, Y.; Ma, X.; Wang, S.; Yang, H.; Gao, X.; Wang, G.; Wang, H. Disclosing the effect of exogenous betaine on growth of Suaeda salsa (L.) Pall in the Liaohe coastal wetland, North China. Marine Pollution Bulletin 2024, 198, 115852. [Google Scholar] [CrossRef]
- Dikilitas, M.; Simsek, E.; Roychoudhury, A. Role of proline and glycine betaine in overcoming abiotic stresses. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, A., Tripathi, D.K., Eds.; 2020. [Google Scholar] [CrossRef]
- Dhansu, P.; Kaushik, S.; Kumar, R.; Krishnapriya, V.; Appunu, C.; Shariff, A.; Kaur, G.; Singh, P.; Kumari, A.; Kumar, A. Intricate relationship of physiological, biochemical and molecular mechanisms in deciphering abiotic stress tolerance in sugarcane. South African Journal of Botany 2026, 188, 135–150. [Google Scholar] [CrossRef]
- Abulfatih, H.A.; Abdel-Bari, E.M.; Alsubaey, A.; Ibrahim, Y.M. Vegetation of Qatar; Scientific and Applied Research Centre (SARC), University of Qatar: Doha, Qatar, 2001. [Google Scholar]
- Yasseen, B.T.; Al-Thani, R.F. Ecophysiology of Wild Plants and Conservation Perspectives in the State of Qatar, Chapter 3. In Agricultural Chemistry; Stoytcheva, M., Zlatev, R., Eds.; 2013; pp. 37–70. [Google Scholar] [CrossRef]
- Al-Thani, R.F.; Yasseen, B.T. Microbial ecology of Qatar, the Arabian Gulf: Possible roles of microorganisms. Front. Mar. Sci. 2021, 8, 697269. [Google Scholar] [CrossRef]
- Abdel-Bari, E.M.M. The Floraof Qatar. Environmental Studies Center; University of Qatar: Doha, Qatar, 2012; Volume 1. [Google Scholar]
- Youssef, A.M.; Hassanein, R.A.; Hassanein, A.A.; Morsy, A.A. Changes in quaternary ammonium compounds, proline and protein profile of certain halophytic plants under different habitat conditions. Pak. J. Biol. Sci. 2003, 6, 867–882. [Google Scholar] [CrossRef]
- Yasseen, B.T.; Al-Thani, R.F. Halophytes and associated properties of natural soils in the Doha area, Qatar. AEHMS 2007, 10, 320–326. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes Under Abiotic Stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef]
- Rindyastuti, R.; Hapsari, L.; Byun, C. Comparison of ecophysiological and leaf anatomical traits of native and invasive plant species. J. Ecology & Environ. 2021, 45. [Google Scholar] [CrossRef]
- Leotta, L.; Toscano, S.; Ferrante, A.; Romano, D. The use of Mediterranean native shrubs for improving the sustainability of urban environments. Front. Hortic. 2025, 4, 1652517. [Google Scholar] [CrossRef]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. Biological parts for engineering abiotic stress tolerance in plants. BioDesign Research 2022, 2022, 9819314. [Google Scholar] [CrossRef] [PubMed]
- Kebert, M.; Stojnić, S.; Rašeta, M.; Kostić, S.; Vuksanović, V.; Ivanković, M.; Lanšćak, M.; Markić, A.G. Variations in proline content, polyamine profiles, and antioxidant capacities among different provenances of European beech (Fagus sylvatica L.). Antioxidants (Basel) 2024, 13, 227. [Google Scholar] [CrossRef]
- Jiang, Z.; van Zanten, M.; Sasidharan, R. Mechanisms of plant acclimation to multiple abiotic stresses. Commun. Biol. 2025, 8, 655. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline metabolism and its implications for plant-environment interaction. The Arabidopsis Book 2010, 2010, e0140. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef]
- Yeo, A.R. Salinity resistance: physiologies and prices. Physiol. Plant 1983, 58, 214–222. [Google Scholar] [CrossRef]
- Grigore, M-N.; Boscaiu, M.; Vicente, O. Assessment of the relevance of osmolyte biosynthesis for salt tolerance of halophytes under natural conditions. The European Journal of Plant Science and Biotechnology 2011, 5, 12–19. [Google Scholar]
- Khan, F.; Adnan, M.Y.; Aziz, I. Metabolic implications of salt induced osmolyte accumulation in Avicennia marina. Pak. J. Bot. 2016, 48, 29–36. [Google Scholar]
- Mattioli, R.; Costantino, P.; Trovato, M. Proline accumulation in plants: not only stress. Plant Signal Behav. 2009, 4, 1016–8. [Google Scholar] [CrossRef]
- Hmidi, D.; Abdelly, C.; Athar, H.U.; Ashraf, M.; Messedi, D. Effect of salinity on osmotic adjustment, proline accumulation and possible role of ornithine-δ-aminotransferase in proline biosynthesis in Cakile maritima. Physiol. Mol. Biol. Plants 2018, 24, 1017–1033. [Google Scholar] [CrossRef]
- Hualpa-Ramirez, E.; Carrasco-Lozano, E.C.; Madrid-Espinoza, J.; Tejos, R.; Ruiz-Lara, S.; Stange, C.; Norambuena, L. Stress salinity in plants: New strategies to cope with in the foreseeable scenario. Plant Physiology and Biochemistry 2024, 208, 108507. [Google Scholar] [CrossRef] [PubMed]
- Yasseen, B.T.; Abu-Al–Basal, M.A. Ecophysiology of Limonium axillare and Avicennia marina from the Coastline of Arabian Gulf-Qatar. Journal of Coastal Conservation: Planning and Management 2008, 12, 35–42. [Google Scholar] [CrossRef]
- Alikhani, F.; Saboora, A.; Razavi, K. Changes in osmolites contents, lipid peroxidation, and photosynthetic pigment of Aeluropus lagopoides under potassium deficiency and salinity. Journal of Stress Physiology & Biochemistry 2011, 7, 5–19. [Google Scholar]
- Moinuddin, M.; Gulzar, S.; Ahmed, M.Z.; Gul, B.; Koyro, H-W.; Ajmal Khan, M. Excreting and non-excreting grasses exhibit different salt resistance strategies. AoB Plants 2014, 6, plu038. [Google Scholar] [CrossRef]
- Ajmal Khan, M.; Ungar, I.A.; Showalter, A.M.; Dewald, H.D. NaCl-induced accumulation of glycinebetaine in four subtropical halophytes from Pakistan. Physiol. Plant 1998, 102, 487–492. [Google Scholar] [CrossRef]
- Alam, H.; Zamin, M.; Adnan, M.; Ahmad, N.; Nawaz, T.; Saud, S.; Basi, A.; Liu, L.; Harrison, M.T.; Hassan, S.; Alharby, H.F.; Alzahrani, Y.M.; Alghamdi, S.A.; Majrashi, A.; Alharbi, B.M.; Alabdallah, N.M.; Fahad, S. Evaluating the resistance mechanism of Atriplex leucoclada (Orache) to salt and water stress; A potential crop for bio-saline agriculture. Front. Plant Sci. 2022, 13, 948736. [Google Scholar] [CrossRef]
- Hayes, M.A.; Chapman, S.; Jesse, A.; O’Brien, E.; Langley, J.A.; Bardou, R.; Devaney, J.; Parker, J.D.; Cavanaugh, K.C. Foliar water uptake by coastal wetland plants: A novel water acquisition mechanism in arid and humid subtropical mangroves. J. Ecol. 2020, 108, 2625–2637. [Google Scholar] [CrossRef]
- Khatib, M. Quaternary ammonium compounds in roots and leaves of Capparis spinosa L. from Saudi Arabia and Italy: investigation by HPLC-MS and 1H NMR. Natural Product Research 2018, 33. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; AL-Khateeb, A.M.H.; Ashrafi, F.; Hafiz, M.I.A. Accumulation of compatible solutes (proline and glycinebetaine) as part of osmoregulation in Cleome species. Journal of Stress Physiology & Biochemistry 2012, 8. [Google Scholar]
- Bhattacharya, E.; Biswas, S.M. First report of the hyperaccumulating potential of cadmium and lead by Cleome rutidosperma DC. With brief insight into the chemical vocabulary of its roots. Fron. Environ. Sci. 2022, 10, 830087. [Google Scholar] [CrossRef]
- Masrahi, Y.S. Ecological significance of wood anatomy in two lianas from arid southwestern Saudi Arabia. Saudi J. Biol. Sci. 2014, 21, 334–41. [Google Scholar] [CrossRef]
- Sivasankaramoorthy, S.; Balasubramanian, T.; Amuthavalli, P.; Sivaraman, P. Effect of sodium chloride on the seedlings of Cressa cretica L. Journal of Ecobiotechnology 2010, 2, 01–03. Available online: http://journal-ecobiotechnology.com/.
- Ghafar, M.A.; Akram, N.A.; Saleem, M.H.; Wang, J.; Wijaya, L.; Alyemeni, M.N. Ecotypic morphological and physio-biochemical responses of two differentially adapted forage grasses, Cenchrus ciliaris L.; Cyperus arenarius Retz. to drought stress. Sustainability 2021, 13, 8069. [Google Scholar] [CrossRef]
- Chagah, S.; Chehrazi, M.; Albaji, M. Effects of drought stress on growth and development Frankenia plant (Frankenia leavis). Bulg. J. Agric. Sci. 2013, 19, 659–665. [Google Scholar]
- Kaur, G.; Sanwal, S.K.; Kumar, A.; Pundir, R.K.; Yadav, M.; Sehrawat, N. Role of osmolytes dynamics in plant metabolism to cope with salinity induced osmotic stress. Discov. Agric. 2024, 2, 59. [Google Scholar] [CrossRef]
- Zengin, F.K.; Munzuroğlu, Ö. Compatible solutes accumulation in Halocnemum strobilaceum (Pall.) MBieb. leaves in response to salinity. Journal of Soil Science and Plant Nutrition 2007, 13, 481–494. [Google Scholar]
- Briens, M.; Larher, F. Osmoregulation in halophytic higher plants: A comparative study of soluble carbohydrates, polyols, betaines and free proline. Plant, Cell & Environment 2006, 5, 287–292. [Google Scholar] [CrossRef]
- Al-Taisan, W. The effect of soil properties on characteristics of Halopeplis perfoliata community in the coastal environment in eastern Saudi Arabia. Fresenius Environmental Bulletin 2016, 25(12a), 5819–5829. [Google Scholar]
- Panda, A.; Rangani, J.; Parida, A.K. Unravelling salt responsive metabolites and metabolic pathways using non-targeted metabolomics approach and elucidation of salt tolerance mechanisms in the xero-halophyte Haloxylon salicornicum. Plant Physiology and Biochemistry 2021, 158, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Yasseen, B.T. Traits of Wild Plants in Qatar Peninsula and Research Perspectives. J. Biology & Nature 2016, 5, 52–66. [Google Scholar]
- Bhatt, A.; Bhat, N.R.; Khalil, M.; Al-Mansour, H. Prioritization of potential native plants from Arabian Peninsula based on economic and ecological values: Implication for restoration. Sustainability 2023, 15, 6139. [Google Scholar] [CrossRef]
- Muslu, A.S.; Kadıoğlu, A. Heliotropium hirsutissimum from geothermal areas: evidence of thermal adaptation. Protoplasma 2025, 262, 1543–1562. [Google Scholar] [CrossRef]
- Pardo-Domenech, L.L.; Tifrea, A.; Grigore, M.N.; Boscaiu, M. Proline and glycine betaine accumulation in two succulent halophytes under natural and experimental conditions. Plant Biosystem 2016, 150, 904–915. [Google Scholar] [CrossRef]
- Syranidou, E.; Christofilopoulos, S.; Kalogerakis, N. Juncus spp.—The helophyte for all (phyto) remediation purposes? New Biotech. 2017, 38, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Orenga, S.; Grigore, M-N.; Boscaiu, M.; Vicente, O. Constitutive and induced salt tolerance mechanisms and potential uses of Limonium Mill species. Agronomy 2021, 11, 413. [Google Scholar] [CrossRef]
- Loconsole, D.; Murillo-Amador, B.; Cristiano, G.; De Lucia, B. Halophytes common ice plants: A future solution to arable land salinization. Sustainability 2019, 11, 6076. [Google Scholar] [CrossRef]
- Kataoka, R.; Akashi, M.; Taniguchi, T.; Kinose, Y.; Yaprak, A.E.; Turgay, O.C. Metabolomics analyses reveal metabolites affected by plant growth-promoting endophytic bacteria in roots of the halophyte Mesembryanthemum crystallinum. Int. J. Mol. Sci. 2021, 22, 11813. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, A.; Smaoui, A.; Zouhaier, B.; Abdelly, C. Differential response to salinity and water deficit stress in Polypogon monspeliensis (L.) Desf. provenances during germination. Plant Biology 2011, 13, 541–545. [Google Scholar] [CrossRef]
- Farzamisepehr, M.; Nourozi, F. Physiological responses of Polypogon monspeliensis L. in petroleum-contaminated soils. Iranian J. Plant Physiol. 2018, 8, 2391–2401. [Google Scholar]
- García-Mercado, H.D.; Villagómez, G.F.; Garzón-Zúñiga, M.A.; Duran, M.C. Fate of mercury in a terrestrial biological lab process using Polypogon monspeliensis and Cyperus odoratus. Int. J. Phytoremediat. 2019, 21, 1–9. [Google Scholar] [CrossRef]
- Zeng, Y.; Wang, S.; Huang, F.; Luo, Q.; Ren, B.; Abo El-Maati, MF.; El-Sappah, A.H. Fate of polycyclic aromatic hydrocarbons in the phytoremediation of different hydrocarbon contaminated soils with cotton, ryegrass, tall fescue, and wheat. Front Plant Sci. 2025, 16, 1550234. [Google Scholar] [CrossRef] [PubMed]
- Moghaieb, R.E.A.; Saneoka, H.; Fujita, K. Effect of salinity on osmotic adjustment, glycine betaine accumulation and betaine aldehyde dehydrogenase expression in Salicornia europaea. Plant Science 2004, 166, 1345–1349. [Google Scholar] [CrossRef]
- Duan, H.; Tiika, R.J.; Tian, F.; Lu, Y.; Zhang, Q.; Hu, Y.; Cui, G.; Yang, H. Metabolomics analysis unveils important changes involved in the salt tolerance of Salicornia europaea. Front Plant Sci. 2023, 13, 1097076. [Google Scholar] [CrossRef]
- Homayouni, H.; Razi, H.; Izadi, M.; Alemzadeh, A.; Kazemeini, S.A.; Niazi, A.; Vicente, O. Temporal changes in biochemical responses to salt stress in three Salicornia. Plants 2024, 13, 979. [Google Scholar] [CrossRef]
- Mendis, C.L.; Padmathilake, R.E.; Attanayake, R.N.; Perera, D. Learning from Salicornia: Physiological, biochemical, molecular mechanisms of salinity tolerance. Int. J. Mol. Sci. 2025, 26, 5936. [Google Scholar] [CrossRef] [PubMed]
- Hadi, M.R. Biotechnological potentials of Seidlitzia rosmarinus: A mini review. Afr. J. Biotechnol. 2009, 8, 2429–2431. Available online: http://www.academicjournals.org/AJB.
- Dragovic, R.; Zlatkovic, B.; Dragovic, S.; Petrovic, J.; Mandic, L.T. Accumulation of heavy metals in different parts of Russian thistle (Salsola tragus, Chenopodiaceae), a potential hyperaccumulator plant species. Biol. Nyssana 2014, 5, 83–90. [Google Scholar]
- Centofanti, T.; Bañuelos, G. Evaluation of the halophyte Salsola soda as an alternative crop for saline soils high in selenium and boron. J. Environ. Mang. 2015, 157, 96–102. [Google Scholar] [CrossRef]
- Karakas, S.; Cullu, M.A.; Dikilitas, S. Comparison of two halophyte species (Salsola soda and Portulaca oleracea) for salt removal potential under different soil salinity conditions. Turkish Journal of Agriculture and Forestry 2017, 41, 183–190. [Google Scholar] [CrossRef]
- Amin, M.A.; Shahhat, I.; Ismail, M.A.; Ibrahim, A. Vicia faba overcomes drought stress by spraying with xerophytic Anabasis setifera extract. Edelweiss Applied Science and Technology 2024, 8, 8008–8022. [Google Scholar] [CrossRef]
- Hamilton, E.W.; McNaughton, S.J.; Coleman, J.S. Molecular, physiological, and growth responses to sodium stress in C4 grasses from a soil salinity gradient in the Serengeti ecosystem. Amer. J. Botany 2001, 88, 1258–1265. [Google Scholar] [CrossRef]
- Jesus, J.M.; Danko, A.S.; Fiúza, A.; Borges, M.T. Phytoremediation of salt-affected soils: a review of processes, applicability, and the impact of climate change. Environ. Sci. Pollut. Res. Int. 2015, 22, 6511–25. [Google Scholar] [CrossRef]
- Dawalibi, V.; Monteverdi, M.C.; Moscatello, S.; Battistelli, A.; Valentini, R. Effect of salt and drought on growth, physiological and biochemical responses of two Tamarix species. iForest—Biogeosciences and Forestry 2015, 8, 772–779. [Google Scholar] [CrossRef]
- Bali, A.S.; Singh Sidhu, G.P.; Grover, D.; Dahiya, B. Biological trace elements confer abiotic stress tolerance in plants. In Biostimulants for Crop Production and Sustainable Agriculture; Hasanuzzaman, M., Hawrylak-Nowak, B., Fujita, M., Islam, M.T., Eds.; CABI, 2022; pp. 306–322. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, J.; Liu, Y.; Zhang, K.; Zhu, F.; Xie, Y. Advances in the biosynthetic regulation and functional mechanisms of glycine betaine for enhancing plant stress resilience. Int. J. Mol. Sci. 2025, 26, 7971. [Google Scholar] [CrossRef]
- Yasseen, B.T.; Al-Thani, R.F. Adaptive Strategies to Biotic Stress in Qatar’s Native Flora. Life 2025, 15, 1645. [Google Scholar] [CrossRef]
- Lianopoulou, V.; Patakas, A.A.; Bosabalidis, A.M. Seasonal dimorphism and winter chilling stress in Thymus sibthorpii. Biologia Plantarum 2014, 58, 139–146. [Google Scholar] [CrossRef]
- Khater, A.K.; Eldanasoury, M.; El-lamey, T.; El-Khamissi, H.A.Z. Seasonal Variation in Photosynthetic Pigments, Phytohormones, and Phenols of Teucrium polium L. Growing in Wadi Halazien, Egypt. Al-Azhar Journal of Agricultural Research 2021, 47. [Google Scholar] [CrossRef]
- Rhodes, D.; Hanson, A.D. Quaternary Ammonium and Tertiary Sulfonium Compounds in Higher Plants. Annual Review of Plant Physiology and Plant Molecular Biology 2003, 44, 357–384. [Google Scholar] [CrossRef]
- Obłąk, E.; Futoma-Kołoch, B.; Wieczyńska, A. Biological activity of quaternary ammonium salts and resistance of microorganisms to these compounds. World J. Microbiol. Biotechnol. 2021, 37, 22. [Google Scholar] [CrossRef] [PubMed]
- Hassanuzzaman, M.; Banerjee, A.; Bhuyan, M.H.M.B.; Roychoudhury, A. Targeting glycine betaine for abiotic stress tolerance in crop plants: Physiological mechanism, molecular interaction and signaling. Phyton-International Journal of Experimental Botany 2019, 88, 185–221. [Google Scholar] [CrossRef]
- Agboma, P.C.; Jones, M.G.K.; Peltonen-Sainio, P.; Rita, H. Exogenous glycinebetaine enhances grain yield of maize, sorghum and wheat grown under two supplementary watering regimes. Journal of Agronomy and Crop Science 2008, 178, 29–37. [Google Scholar] [CrossRef]
- McNeil, S.D.; Nuccio, M.L.; Ziemak, M.J.; Hanson, A.D. Enhanced synthesis of choline and glycine betaine in transgenic tobacco plants that overexpress phosphoethanolamine N-methyltransferase. Proc. Natl. Acad. Sci. USA 2001, 98, 10001–10005. [Google Scholar] [CrossRef]
- Xu, Z.; Sun, M.; Jiang, X.; Sun, H.; Dang, X.; Cong, H.; Qiao, F. Glycinebetaine biosynthesis in response to osmotic stress depends on jasmonate signalling in watermelon suspension cells. Front. Plant Sci. 2018, 9, 1469. [Google Scholar] [CrossRef]
- Norton, J.; Abdul Majid, S.; Allan, D.; Al Safran, M.; Böer, B.; Richer, R. An Illustrated Checklist of the Flora of Qatar; United Nations Educational, Scientific and Cultural Organization, UNESCO Office in Doha: Doha; Qatar Foundation, MAERSK OIL QATAR AS, 2009. [Google Scholar]
- Eisenhut, M.; Roell, M-S.; Weber, A.P.M. Mechanistic understanding of photorespiration paves the way to a new green revolution. New Phytologist 2019, 223, 1762–1769. [Google Scholar] [CrossRef]
- Wingler, A.; Lea, P.J.; Quick, W.P.; Leegood, R.C. Photorespiration: metabolic pathways and their role in stress protection. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2000, 355, 1517–1529. [Google Scholar] [CrossRef]
- Chen, T.H.H.; Murata, N. Glycine betaine: An effective protectant against abiotic stress in plants. Trends in Plant Stress in Plant Science 2008, 13, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Niu, T.; Zhang, T.; Qiao, Y.; Wen, P.; Zhai, G.; Liu, E.; Al-Bakre, D.A.; Al-Harbi, M.S.; Gao, X.; Yang, X. Glycinebetaine mitigates drought stress-induced oxidative damage in pears. PLoS One 2021, 16, e0251389. [Google Scholar] [CrossRef] [PubMed]
- Alam, Kh. A.; Karim, S.; Sultana, S.; Das, A.K.; Mahmud, A.; Rahman, Md. A.; Hossain, Md. M.; Arafat, Y.; Parvin, S.; Lee, M.-S. Glycine betaine improves photosynthetic performance and antioxidant defense in barley under water deficit conditions. Biomolecules 2026, 16, 372. [Google Scholar] [CrossRef] [PubMed]
- Al-Thani, R.F.; Yasseen, B.T. Perspectives of Future Water Sources in Qatar by Phytoremediation: Biodiversity at Ponds and Modern Approach. International Journal of Phytoremediation 2021, 23, 866–889. [Google Scholar] [CrossRef]
- Tan, T.; Hu, W.; Fu, T.; Li, M.; Campbell, G.; Smith, P.; Yi, Z.; Xue, S.; Xu, Y. Long-term Miscanthus cultivation enhances soil quality in degraded acidic red soils: A multi-indicator assessment. Industrial Crops and Products 2026, 242, 123001. [Google Scholar] [CrossRef]
Figure 1.
Physiological and biochemical responses of plants to abiotic stresses (salinity, drought, and extreme temperatures-cold stress) mediated by ABA signalling.
Figure 1.
Physiological and biochemical responses of plants to abiotic stresses (salinity, drought, and extreme temperatures-cold stress) mediated by ABA signalling.


Table 1.
Proline content in leaves of Mexican wheat cultivars differ in their salt resistance [57].
Table 1.
Proline content in leaves of Mexican wheat cultivars differ in their salt resistance [57].
| NaCl treatment (dSm-1) | μmoles g-1 FW | |
| Cajeme (salt resistance) | Yecora (salt sensitive) | |
| 0 | 0.21 | 0.21 |
| 75 | 0.33 | 0.57 |
| 150 | 1.96 | 3.75 |
Table 2.
Proline content varies among five native Qatari plants with different ecological types [35].
Table 2.
Proline content varies among five native Qatari plants with different ecological types [35].
| Plant species | Ecological type | μgg-1 FW* |
| Avicennia marina | Halophyte: Mangroves | 11–37 |
| Halopeplis perfoliata | Halophyte: Sabkhas | 56–281 |
| Limonium axillare | Halophyte: Sabkhas & coastlines | 103–810 |
| Tetraena qatarensis | Xero-halophyte | 419–1136 |
| Ochradenus baccatus | Xerophyte | 1277–1347 |
* The ranges are from six observations.
Table 3.
Studied plant species, their corresponding ecological types, and selected physical characteristics in their natural habitats [35].
Table 3.
Studied plant species, their corresponding ecological types, and selected physical characteristics in their natural habitats [35].
| Plant species | Family | Ecological type | Soil water content (% FC) I,* | ECe of the soil (dSm-1)* |
| Aeluropus lagopoides (Monocot) | Poaceae | Xerophyte; a grass can live in saline habitats as well | 40–66 | 45–50 |
| Anabasis setifera | Chenopodiaceae | Facultative halophyteII | 12–43 | 71–198 |
| Arhrocenmum macrostachyum | Amaranthaceae | Halophyte; it grows in coastal and inland salt marshes, alkali flats, and other habitats with saline soils | 24–48 | 27–198 |
| Avicennia marina | Acanthaceae | Halophyte; mangrove, well adapted at Coastal intertidal zones | Saturated Soil | Seawater |
| Capparis spinosa | Capparaceae | Xerophyte; its morphological and physiological features reflect its ecological type | 9–12 | 4–5 |
| Caroxylon imbricatum (previously Salsola imbricata) | Amaranthaceae | Xerohalophyte; a plant adapted to both drought and salinity; found in disturbed areas, saline soils, and coastal areas | 11–16 | 150–195 |
| Cocculus pedulus | Menispermaceae | Xerophyte | 9–12 | 4–5 |
| Halocnemum strobilaceum | Amaranthaceae | Halophyte; lives in tidal zones and salt flats, grows in coastal and inland salt marshes, alkali flats, and other habitats with saline soils | 34–61 | 12–198 |
| Halopeplis perfoliata | Amaranthaceae | Halophyte; found in highly saline sabkhas with sandy shelly soil | 34–50 | 12–178 |
| Haloxylon salicornicum | Amaranthaceae | Xerophyte; found in stony soil overlain with wind-blown sand | 9–12 | 80–90 |
| Heliotropium bacciferum | Boraginaceae | Xerophyte with minor salt tolerance, not a halophyte | 9–10 | 5–72 |
| Limonium axillare | Plumbaginaceae | Halophyte; found on coastline with saline shelly soil | 32–43 | 12–198 |
| Ochradenus baccatus | Resedaceae | Xerophyte; found in disturbed areas | 4–12 | 7–8 |
| Pulicaria crispa | Asteraceae | Xerophyte; found in low depressions and water catchment areas in sandy slightly saline soil | 5–8 | 4–6 |
| Pulicaria gnaphalodes | Asteraceae | Xerophyte; found in shallow depressions and rain pools with sandy stony soil, extremely woolly with pleasant smell | 5–8 | 4–6 |
| Salsola soda | Amaranthaceae | Halophyte; it thrives in saline environments, found in salt marshes, coastal areas, and saline soils | 43–71 | > 200 |
| Salicornia spp.III | Amaranthaceae | Halophytes; found on flooded landward side of mangrove associated with Salsola soda, and intidal zone and sabkha depressions | 43–71 | > 200 |
| Salsola rosmarinus (Syn. Seidlitzia rosmarinus) | Amaranthaceae | Xerohalophyte; can be regarded as both a halophyte and a xerophyte because it thrives in harsh, arid–saline desert habitats | 14–30 | 52–81 |
| Sporobolus spicatus (Monocot) | Poaceae | Halophyte; commonly known as salt grass due to its ability to thrive in high-salinity environments, has salt gland structures to excrete extra salts, grows in arid regions and can tolerate drought conditions | 32–44 | 107–128 |
| Suaeda aegyptiaca | Amaranthaceae | Xerohalophyte; is highly adapted to environments with both high salinity and low water availability, found in depressions with high water table and coastal areas | 11–16 | 150–195 |
| Suaeda vermiculata | Amaranthaceae | Halophyte; a xerophyte as well (drought-tolerant plant), found in moist saline soil and sabkhas | 8–43 | 32–142 |
| Tetraena qatarensisIV | Zygophyllaceae | Xerohalophyte; adapted to environments that are both dry and salty; many features should be discussed | 10–46 | 12–187 |
| Ziziphus nummularia | Rhamnaceae | Xerophyte; is highly adapted to and primarily found in hot, arid, and dry regions (xeric habitats); found in rawdahs with depth and fine soil |
9–12 | 4–5 |
I FC: Field Capacity, II Facultative halophyte: a plant that can grow in both saline and non-saline environments, although it is well adapted to high salt conditions and does not require salt for its growth and survival, IIISalicornia spp.: include about three species; however, their classification has not been clarified with well-known species. Abdel-Bari [98] has reported three species: Salicornia macrostachya, S. perfoliate, S. strobilacea, and S. europaea was included among the halophytes studied by Abdel-Bari et al. [35]. IV There are many characteristics that characterize this plant as a xero-halophyte. *The ranges are from six observations.
Table 4.
Plant water content, proline, total soluble sugars (TSS), and total soluble nitrogen (TSN) measured in shoot systems of selected native plant species in Qatar in natural habitat conditions. [35].
Table 4.
Plant water content, proline, total soluble sugars (TSS), and total soluble nitrogen (TSN) measured in shoot systems of selected native plant species in Qatar in natural habitat conditions. [35].
| Plant species | Plant water content (%)* | Range of proline (μgg-1 fresh weight)* | Range of TSS (mgg -1) DW* | Range of TSN (μgg-1) DW* |
| Aeluropus lagopoides | 70–72 | 241–253 | 1.5–1.7 | 84–126 |
| Anabasis setiferaI (Syn. Salsola setifera) | 72–76 | 21–54 | 1.6–2.0 | 81–92 |
| Arhrocenmum macrostachyum | 81–89 | 10–112 | 4.0–14.1 | 79–165 |
| Avicennia marina | 66–69 | 11–37 | 2.2–6.0 | 40–63 |
| Capparis spinosa | 62–68 | 131–302 | No data | No data |
| Caroxylon imbricatum previously: Salsola imbricata | 80–81 | 38–82 | 1.6–3.1 | 77–128 |
| Cocculus pedulusII | 60–69 | 34–51 | No data | No data |
| Halocnemum strobilaceum | 71–80 | 19–73 | 1.8–2.5 | 48–146 |
| Halopeplis perfoliata | 78–88 | 56–281 | 2.9–3.5 | 46–88 |
| Haloxylon salicornicum | 70–78 | 91–119 | No data | No data |
| Heliotropium bacciferum | 69–76 | 151–192 | 10.6–16.6 | 73–140 |
| Limonium axillare | 64–72 | 103–810 | 4.7–8.0 | 67–169 |
| Ochradenus baccatus | 68–71 | 1277–1347 | 2.9–3.5 | 92–109 |
| Pulicaria crispa | 59–66 | 25–32 | 8.8–9.9 | 127–144 |
| Pulicaria gnaphalodes | 59–66 | 32–119 | 6.3–7.2 | 63–92 |
| Salicornia spp.III | 80–87 | 55–172 | No data | No data |
| Salsola rosmarinus (Syn. Seidlitzia rosmarinus) | 84–88 | 161–327 | 2.8–4.7 | 77–119 |
| Salsola soda | 83–89 | 132–185 | No data | No data |
| Sporopolus spicatus (Monocot) | 58–66 | 40–147 | 3.3–3.4 | 195–236 |
| Suaeda aegyptiaca | 86–88 | 80–317 | 3.9–4.3 | 100–119 |
| Suaeda vermiculata | 77–82 | 96–409 | 6.7–19.8 | 104–161 |
| Tetraena qatarensis | 81–87 | 419–1136 | 2.9–4.3 | 85–190 |
| Ziziphus nummularia | 62–67 | 87–97 | No data | No data |
I Facultative halophyte: a plant that can grow in both saline and non-saline environments, although it is well adapted to high salt conditions and does not require salt for its growth and survival; this species was re-named as Salsola setifera. II More clarifications are needed for proline accumulation, and other solutes. III See Table 3 for more clarifications. TSS: Total soluble sugars, TSN: Total soluble nitrogen. *The ranges are from six observations.
Table 5.
List of some native Qatari plants, including halophytes and xerophytes, their remediation role, and presence of compatible solutes.
Table 5.
List of some native Qatari plants, including halophytes and xerophytes, their remediation role, and presence of compatible solutes.
| Species/Ecotype/Family | Phytoremediation activity | Compatible solute accumulation# | Remarks and observations | References |
|
Aeluropus lagopoides* Fam.: Poaceae |
Petroleum hydrocarbons, heavy metals such as Cd, Pb | Possible accumulation | C4 grass, exhibits some xerophytic adaptations | [12,116,117] |
| Arhrocenmum spp.*, 4 species, Amaranthaceae | Remediates heavy metals; petroleum hydrocarbons require further investigation | Little proline was detected; higher levels of soluble sugars and nitrogen were observed, and GB accumulation is possible | C3 plant, some adaptations to desert habitats | [12,18] |
|
Atriplex spp.*** 2 species Amaranthaceae |
Heavy metals: Cd, Cu, Ni, Pb, Zn; petroleum hydrocarbons need further investigation | Accumulate proline, GB, and soluble sugars in their natural habitats, ABA increases under salt stress | C3, C4 plants, xerophytes and halophytes are recognized among this genus | [118,119] |
|
Avicennia marina* Acanthaceae |
Heavy metals: Cd, Co, Cr, Cu, Fe, Ni, Zn, and petroleum hydrocarbons |
Little proline was detected; higher levels of soluble sugars were observed; GB needs further confirmation | C3 plant, mangrove tree | [12,120] |
|
Capparis spinosa** Capparaceae |
Accumulates heavy metals and degrades petroleum hydrocarbon | Accumulates proline, and QAC; needs further investigation for GB and soluble sugars | C3 plant, might resist salinity to a certain level | [35,121] |
| Cleome spp.** Cleomaceae (formerly Capparaceae) | Remediate heavy metals such as Cd and Cu, needs confirmation | Accumulate proline, GB, and soluble sugars in their natural habitats | C3 or C4 plants, depending on species, accumulates some fatty acids for detoxification | [12,122,123] |
| Cocculus pendulus** Menispermaceae | No information, needs to be tested | Little proline was detected, might accumulate GB, needs further investigation for soluble sugars | C3 plant | [35,45,124] |
| Cressa cretica* Convolvulaceae | Remediates some heavy metals, possibly remediates petroleum hydrocarbons |
Accumulates proline, needs further investigation for GB and soluble sugars | C4 plant | [12,125] |
| Cyperus spp. *, **, $, 3 species Cyperaceae | Remediates many heavy metals such as Al, Cd, Co, Cr, Cu, Fe, Hg, Mn, Ni, Pb, and Zn (phyto-stabilization of Ni), petroleum hydrocarbons | Accumulates proline, possibly GB, and soluble sugars | Mainly C4, some C3, species in Qatar are salt tolerant, xerophytes, and mesophytes | [12,126] |
| Frankenia pulverulenta* Frankeniaceae | Remediates heavy metals such as Cd, Cr, Cu, Ni, Sr, and Zn, petroleum hydrocarbons | Accumulates compatible solutes: proline, GB, and soluble sugars | C3 plant, found in moist saline soil | [12,127,128] |
| Halocnemum strobilaceum* Amaranthaceae | Accumulates heavy metals, such as Cd, Cu, Fe, Mn, Ni, Pb, Zn; needs further studies for petroleum hydrocarbons | Accumulates compatible solutes such as proline and soluble sugars | C4 plant, obligate halophyte, might accumulate GB | [1,12,129] |
| Halodule uninervis* Hydrocharitaceae | Accumulates heavy metals such as Cu, Fe, Ni, Pb; phyto-remediates petroleum hydrocarbons | Possible accumulation of compatible solutes; needs to be tested | C3 plant, marine hydrophyte seagrass, halophyte | [12,77,130] |
| Halopeplis perfoliata* Amaranthaceae | Remediates some heavy metals and may help remediate petroleum hydrocarbons | Accumulates proline and possibly other compatible solutes such as GB and soluble sugars; needs confirmation | C4 plant, succulent | [1,35,131] |
| Halopyrum mucronatum* Poaceae | Some heavy metals are accumulated; considered as bioindicator for Cr, Fe, Pb, and Zn; needs to be tested for phytoremediation of petroleum hydrocarbons | Accumulates GB and possibly other compatible solutes | C4 plant, coastal halophytic grass | [1,3,117,118] |
| Haloxylon salicornicum** Amaranthaceae | Accumulates heavy metals such as Cu, Fe, Mn, Zn; possibly phyto-remediates petroleum hydrocarbons | Little proline was detected; accumulates GB | C4 plant, native to arid and semi-arid desert region | [12,100,118,132] |
| Heliotropium spp., **, 8 species, Boraginaceae | Possible role in phytoremdiation of organic pollutants and heavy metals | Little proline was detected; large amounts of soluble sugars accumulated, needs to be tested for GB | C3 plants, can be considered as semi-xerophyte, might be found in coastal saline soil | [3,35,133,134,135] |
| Juncus rigidus***, Juncaceae | Phyto-remediates organic compounds, heavy metals, and saline soil | Might accumulate compatible solutes, further investigation is needed | C3 plant, shows some xerophytic features | [136,137] |
| Limonium axillare* Plumbaginaceae | Remediates saline soil and heavy metals | Proline and soluble sugars accumulate, some relatively compatible solutes such as such as β-alanine betaine & choline-O-sulphate | C3 plant, found in coastal saline habitats, moist lands | [12,115,138] |
| Mesembryanthemum spp. ***, 2 species, Aizoaceae | Could be used for phytoremediation of contaminated soil, needs testing | Might accumulate compatible solutes under stress, needs testing | CAM plant | [139,140] |
| Ochradenus baccatus**, Resedaceae | Needs testing for petroleum hydrocarbon phytoremediation | Accumulates compatible solutes such as proline, soluble sugars, and possibly GB | C3 plant | [1,3,35] |
| Polypogon monspeliensis*, Poaceae | Accumulates heavy metals, phyto-remediates petroleum hydrocarbons and saline soil | Needs testing for compatible solutes | C3 plant, possible accumulator of GB | [1,12,14,142,143] |
| Pulicaria spp.** 4 species, Asteraceae | Possible candidates for petroleum hydrocarbons, need testing | Little proline was detected, accumulate significant amounts of soluble sugars, possible accumulation of GB; needs testing | C3 plants, tolerate salinity | [35,45,144] |
| Salicornia spp. *, 3 species, Amaranthaceae | Possible phytoremediation candidates for petroleum hydrocarbon and heavy metals, need further investigation | Very strong evidence that these species accumulate compatible solutes such as proline, soluble sugars and GB; further investigation is needed | C3 plants, obligate halophytes, biochemical adaptation; these species have several applications in the food, feed, pharmaceutical, cosmetics, and bioenergy sectors | [35,145,146,147,148] |
|
Salsola spp., *, **, 9 species, Amaranthaceae |
Possible role in phytoremediation, especially heavy metals and saline soils, some species remediate heavy metals such as B, Cu, Mn, and Se; further investigations are needed | Accumulates proline and soluble sugars and might accumulate other compatible solutes such as GB; needs testing | C4 plants, including halophytes and xerophytes, some bioactive uses were reported; industrial and nutritional values | [35,149,150,151,152,153] |
| Sporobolus spicatus* (Monocot), Poaceae | Phytoremediates petroleum hydrocarbons, heavy metals, and saline soils | Accumulates compatible solutes such as proline and soluble sugars; GB might have a role in salt tolerance | C4 plant; proven efficient in controlling polluted soils | [12,154,155] |
| Suaeda spp. * 7 species, Amaranthaceae | Phytoremediate heavy metals and possibly petroleum hydrocarbons; testing is needed | Accumulates proline, soluble sugars, and possibly GB, some species proved to accumulate GB | C3, C4 plants are found in the genus, application of GB might improve plant growth in saline conditions | [35,92] |
| Tamarix spp.***, 5 species, Tamaricaceae | Phytoremediate heavy metals and his polycyclic aromatic hydrocarbons |
Accumulate significant amounts of compatible solutes such as proline and soluble sugars; GB accumulation needs confirmation | C3 plants | [12,156,157] |
| Tetraena qatarensis***, Zygophyllaceae | Accumulates heavy metals, and possible phytoremediation of petroleum hydrocarbons | Accumulates significant amounts of proline and les soluble sugars; GB accumulation needs confirmation, | C4 plant | [35,45,158,159] |
| Teucrium polium**, Lamiaceae | Accumulates heavy metals such as Co, Ni; phytoremediates petroleum hydrocarbons | Accumulates compatible solutes such as proline and soluble sugars | C3 plant | [12,160,161] |
*Halophyte, **Xerophyte, ***Xerohalophyte, # See Table 4 for proline, and soluble sugars, $Mesophyte.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



