Submitted:
08 April 2026
Posted:
09 April 2026
You are already at the latest version
Abstract
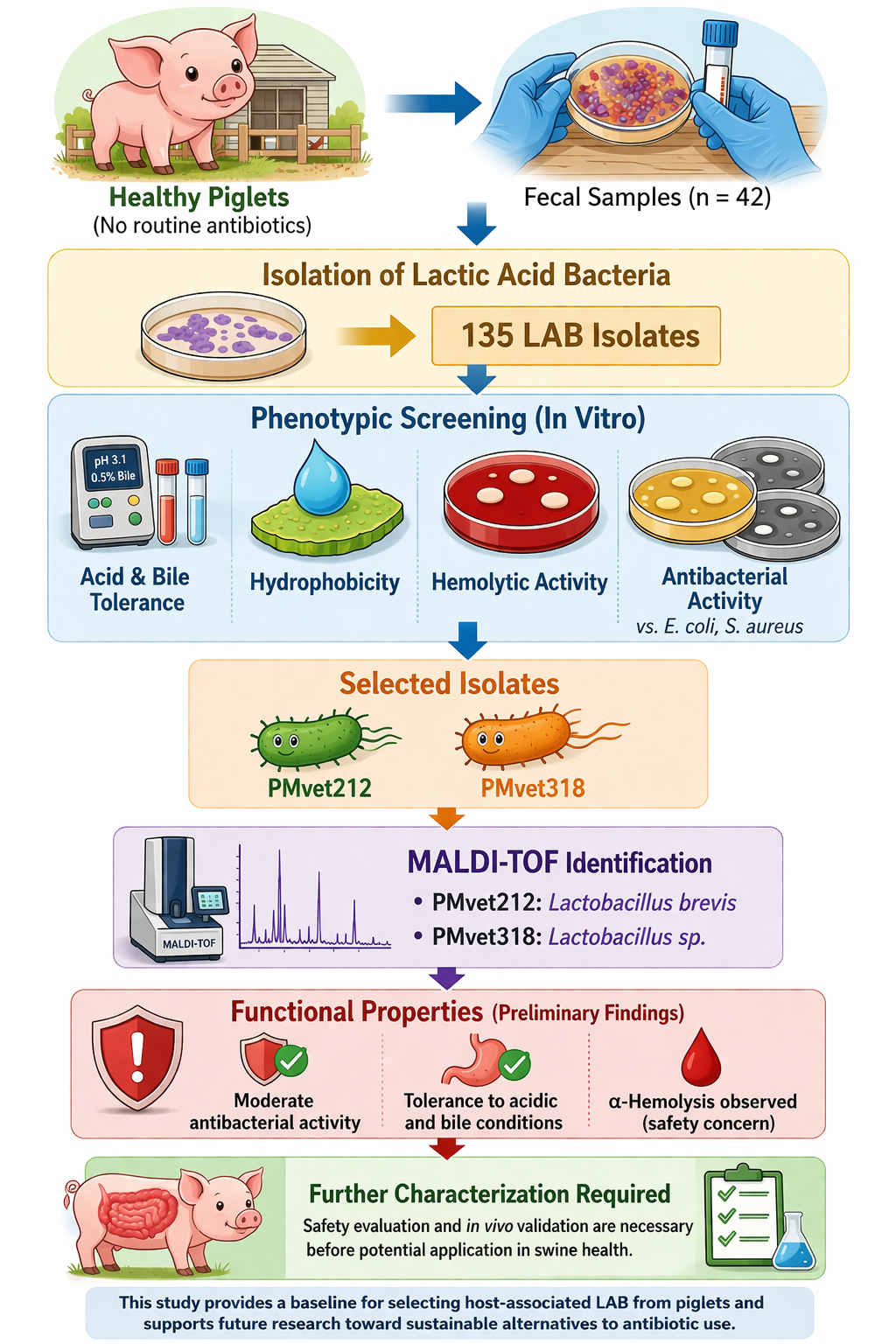
Lactic acid bacteria are important components of the early gut microbiota in piglets and may contribute to gastrointestinal stability and control of enteric pathogens, particularly under increasing restrictions on antibiotic use in livestock production. This study aimed to perform in vitro phenotypic screening and characterization of lactic acid bacteria isolated from feces of suckling piglets aged 7–28 days. A total of 42 fecal samples were collected and cultured on selective media, yielding 318 colonies, of which 135 Gram-positive, rod-shaped, catalase-negative isolates were retained for further evaluation. These isolates were assessed for tolerance to acidic conditions (pH 2.0–3.1), bile salts (0.3–0.5%), cell surface hydrophobicity, hemolytic activity, and antibacterial activ-ity against Escherichia coli and Staphylococcus aureus. Among them, only two isolates (PMvet212 and PMvet318) showed detectable tolerance to acidic and bile conditions and exhibited moderate antibacterial activity, with inhibition zones of approximately 10-12 mm. Identification using matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry indicated that PMvet212 was closely related to Lactobacillus brevis, whereas PMvet318 was identified at the genus level as Lactobacillus sp. Both isolates displayed α-hemolytic activity, indicating the need for further safety evaluation. Overall, the findings provide preliminary evidence of selected functional properties of piglet-derived lactic acid bacteria; however, additional molecular characterization and in vivo studies are required before any practical application can be considered.
Keywords:
piglets
; lactic acid bacteria
; in vitro screening
; MALDI-TOF
; antibacterial activity
1. Introduction
Escherichia coli (E. coli) is one of the most important pathogens causing diarrhea in neonatal and weaned piglets. Piglet diarrhea leads to substantial economic losses in the swine industry due to increased morbidity and mortality, reduced growth performance, and higher veterinary costs [1]. Prevention is generally more effective than treatment; however, traditional control strategies have relied heavily on antibiotics [2]. This dependence has raised serious concerns regarding antibiotic residues in animal products and the global spread of antimicrobial resistance (AMR) [3]. As regulatory authorities and consumers increasingly demand reductions in antibiotic use, there is a growing need for alternative approaches that can maintain animal health and productivity without contributing to AMR [4]. Among the available strategies, probiotics have received considerable attention due to their potential to improve intestinal health, reduce enteric infections, and support growth performance in pigs [5]. In addition, the use of probiotics is consistent with the One Health approach for mitigating AMR across animal, human, and environmental interfaces [6].
At birth, the gastrointestinal tract of piglets is rapidly colonized by microorganisms derived from the sow and the surrounding environment. This early colonization plays a critical role in the development of digestive, metabolic, and immune functions [7]. The composition of the gut microbiota is influenced by several factors, including maternal microbiota, diet, and environmental exposure [8]. In particular, the suckling period (approximately 7-28 days of age) represents a crucial stage in microbial succession, during which beneficial bacteria such as lactic acid bacteria (LAB) become established and contribute to gut stability and resistance to pathogens [9]. Therefore, isolating LAB from piglets at this early stage may provide valuable insight into host-adapted bacterial populations with potential functional relevance.
Probiotics are defined as live microorganisms that, when administered in adequate amounts, confer health benefits to the host (FAO/WHO, 2002) [10]. To be considered suitable, probiotic microorganisms must meet several minimum criteria, including tolerance to gastric acid and bile salts, ability to adhere to intestinal surfaces, absence of harmful activities, and antagonistic effects against pathogens [10,11]. LAB are among the most widely studied probiotic groups because of their generally recognized safety status, ability to survive gastrointestinal conditions, and production of antimicrobial compounds such as organic acids and bacteriocins [12]. However, these functional properties are highly strain-specific, and not all LAB isolates meet the required criteria for probiotic application.
Recent advances in microbial identification techniques, such as matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry, have enabled rapid and reliable identification of bacterial isolates at the genus and species levels [13]. This approach provides an important advantage over traditional phenotypic methods by improving the accuracy of taxonomic classification and facilitating comparison with previously characterized strains. Incorporating such identification methods is essential for interpreting the functional relevance of LAB isolates and for aligning experimental findings with existing microbiological knowledge.
Therefore, the aim of the present study was to perform in vitro phenotypic screening and characterization of LAB isolated from fecal samples of piglets raised without routine antibiotic or probiotic supplementation. The study focused on identifying isolates that exhibit tolerance to simulated gastrointestinal conditions, antibacterial activity against E. coli, and selected functional properties relevant to probiotic evaluation. In addition, selected isolates were identified using MALDI-TOF mass spectrometry to provide preliminary taxonomic classification. The findings of this study contribute to the understanding of piglet-associated LAB and provide a basis for further molecular characterization and functional validation.
2. Materials and Methods
2.1. Ethical Approval
Fecal samples were collected non-invasively from freshly voided feces without handling or restraining the animals. Therefore, specific animal use approval was not required according to institutional and national animal welfare guidelines. However, all procedures involving microbial isolates were reviewed and approved by the Institutional Biosafety Committee (IBC) of Chiang Mai University (Approval No. CMUIBC A-0763004).
2.2. Sample Collection
The study population comprised suckling piglets aged 7-28 days. A total of 42 fecal samples were collected from 10 commercial swine farms in Lamphun Province, Thailand, with samples obtained from multiple farrowing pens (approximately 1-5 samples per pen). The selected farms did not use antibiotics or probiotics routinely, except for therapeutic purposes when necessary. Freshly voided feces with normal consistency and yellow to brown coloration were collected aseptically from the pen floor, placed in sterile 5 mL microcentrifuge tubes (≥2 g per sample), and transported to the laboratory at 4 °C. All samples were processed within 6 h after collection to minimize changes in microbial composition.
2.3. Isolation of Lactic Acid Bacteria (LAB)
One gram of each fecal sample was homogenized in 9 mL of sterile 0.85% saline solution and serially diluted (10−4 to 10−8). Aliquots (100 µL) were spread onto de Man, Rogosa and Sharpe (MRS) agar supplemented with 0.5% CaCO3. Plates were incubated anaerobically at 37 °C for 48 h using an anaerobic jar system with gas-generating sachets (Oxoid, UK). Colonies producing clear zones were selected and repeatedly streaked on MRS agar to obtain pure cultures. Gram-positive, non-spore-forming isolates were further tested for catalase activity, and only catalase-negative isolates were preserved at -20 °C in MRS broth containing 20% (v/v) sterile glycerol [14].
2.4. Evaluation of Probiotic Properties
2.4.1. Acid Tolerance
Bacterial suspensions were prepared in phosphate-buffered saline (PBS; pH 7.2) and adjusted to 0.5 McFarland standard (~1.5 × 108 CFU/mL). One milliliter of suspension was mixed with 9 mL PBS adjusted to pH 2.0 or pH 3.1 using 1 N HCl. The mixtures were incubated at 37 °C for 3 h. Viable cell counts before and after incubation were determined by plate counting on MRS agar. Acid tolerance was expressed as the reduction in viable counts (log CFU/mL) compared with the initial value [15].
2.4.2. Bile Salt Tolerance
Bacterial suspensions were inoculated into MRS broth supplemented with 0.3%, 0.5%, or 1% (w/v) bile salts (oxgall; Sigma-Aldrich, USA). Control cultures without bile salts were included. Cultures were incubated anaerobically at 37 °C for 24 h. Viable cell counts were determined at 0 and 24 h, and tolerance was evaluated based on survival relative to controls [15].
2.4.3. Adhesion Ability to Intestinal Epithelium (Surface Hydrophobicity)
Cell surface hydrophobicity was evaluated using the microbial adhesion to hydrocarbons (MATH) method with xylene as the hydrophobic solvent. Bacterial suspensions were adjusted to an optical density (OD600) of 0.6 in PBS. After mixing with xylene and phase separation, OD600 values were measured, and hydrophobicity (%) was calculated.
%H = [(OD600 before mixing – OD600 after mixing)/OD600 before mixing] × 100
Xylene was selected as a standard nonpolar solvent commonly used for evaluating bacterial surface hydrophobicity. Control tubes containing PBS and xylene without bacteria were included. All assays were performed in duplicate [16]
2.4.4. Hemolytic Activity
Hemolytic activity was evaluated on tryptic soy agar (TSA) supplemented with 5% (v/v) defibrinated sheep blood (HiMedia, India). Plates were incubated at 37 °C for 48 h. Hemolysis patterns were classified as β-, α-, or γ-hemolysis based on visual inspection. Staphylococcus aureus ATCC 6538 was used as a positive control [17].
2.4.5. Antimicrobial Activity Against E. coli and S. aureus ATCC 6538
Antibacterial activity was evaluated using the agar well diffusion method. Cell-free supernatants (CFS) were obtained by centrifugation and filtration (0.45 µm). Indicator strains included field-isolated Escherichia coli and Staphylococcus aureus ATCC 6538. Wells were filled with 80 µL CFS, and plates were incubated at 37 °C for 24 h. Lactic acid (2% v/v) was used as a positive control. Inhibition zones were measured in millimeters [18]. The pH of the cell-free supernatants was not neutralized prior to testing.
2.5. Data Interpretation Probiotic Properties
2.5.1. Acid Tolerance
Survival at acidic pH was assessed by comparing bacterial counts (log CFU/mL) before and after incubation at pH 2.0 and pH 3.1. A reduction of less than 1 log CFU/mL at pH 3.1 was considered acceptable for acid tolerance [19].
2.5.2. Bile Salt Tolerance
Bacterial counts (log CFU/mL) at 0 h and 24 h in MRS broth containing bile salts were compared. Strains were classified as tolerant if no significant reduction in viable counts was observed over the incubation period.
2.5.3. Surface Adhesion Ability
Adhesion potential was expressed as surface hydrophobicity (%H) calculated by the MATH assay. Higher %H values indicated stronger cell surface adhesion capabilities.
2.5.4. Hemolytic Activity
Hemolysis was classified into three types: β-hemolysis, indicating complete lysis of red blood cells and the presence of a clear zone around the colony; α-hemolysis, representing partial lysis with a green or brown discoloration surrounding the colony; and γ-hemolysis, showing no lysis or visible change in the medium. According to safety criteria for probiotic use, LAB strains exhibiting α- or β-hemolysis were considered un-suitable, whereas only γ-hemolytic strains were deemed acceptable [20].
2.5.5. Antimicrobial Activity Against E. coli and S. aureus ATCC 6538
The antimicrobial potential of the cell-free supernatants (CFS) was assessed by measuring the diameter of the inhibition zones against Escherichia coli and Staphylococcus aureus ATCC 6538. The results were interpreted according to criteria modified from Sirichokchatchawan et al. (2018) [18], as summarized in Table 1. This approach allowed for a comparative evaluation of the inhibitory effects of different CFS preparations on the selected pathogenic strains.
2.6. Molecular Identification of Selected Isolates
Selected isolates showing tolerance to acidic and bile conditions were identified using matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry (Bruker Daltonics, Germany). Bacterial colonies were prepared according to the manufacturer’s instructions and analyzed using the reference database. Identification scores were interpreted as follows: ≥2.0 indicated reliable identification at the species level, 1.7-1.99 indicated genus-level identification, and <1.7 was considered unreliable. The closest matching species and corresponding score values were recorded [13].
2.7. Statistical Analysis
All experiments were performed in triplicate using the same bacterial isolate (technical replicates). Data were expressed as mean ± standard deviation (SD). Statistical analysis was conducted using SPSS software (IBM SPSS Statistics, version 29). As this study was designed as a preliminary in vitro screening, no inferential statistical comparisons among isolates were performed. No hypothesis testing was performed due to the exploratory screening design.
3. Results
3.1. Isolation of Lactic Acid Bacteria (LAB) from Piglet Feces
From 42 fecal samples collected from suckling piglets, a total of 318 colonies producing clear zones on MRS agar supplemented with CaCO3 were initially obtained after anaerobic incubation at 37 °C for 48 h. These colonies were considered presumptive lactic acid bacteria based on their acid-producing ability. Gram staining revealed that 296 isolates were Gram-positive, including 146 rod-shaped (bacilli), 136 cocci, and 14 coccobacilli. Catalase testing showed that 277 isolates were catalase-negative, while 19 were catalase-positive and excluded. Based on morphological and biochemical characteristics, 135 Gram-positive, rod-shaped, catalase-negative isolates were retained for further evaluation.
Nineteen isolates exhibited budding and pleomorphic morphology consistent with yeast and were excluded from subsequent analysis. Additionally, three isolates could not be maintained during subculturing. Among isolates with ambiguous Gram-staining results, PMvet120, PMvet151, and PMvet183 were confirmed to be yeast-like and excluded (Figure 1).
3.2. Functional Properties of Selected Isolates
3.2.1. Acid Tolerance
Among the 135 isolates screened, only two isolates (PMvet212 and PMvet318) demonstrated tolerance to acidic conditions. These isolates showed a reduction in viable counts of less than 1 log CFU/mL after incubation at pH 3.1 for 3 h. No isolates survived exposure to pH 2.0 under the same conditions. The survival profiles are summarized in Table 2.
3.2.2. Bile Salt Tolerance
The two acid-tolerant isolates were further evaluated for their ability to survive in the presence of bile salts. Both isolates maintained viable counts above 6 log CFU/mL after incubation in 0.3% bile salts. At 0.5% bile salts, only PMvet212 retained substantial viability, whereas PMvet318 decreased below the detection limit. At 1.0% bile salts, no survival was observed for either isolate. The survival data are presented in Table 3.
3.2.3. Cell Surface Hydrophobicity (MATH Assay)
Cell surface hydrophobicity of the selected isolates showed variation between isolates. PMvet318 exhibited the highest hydrophobicity value (12.38 ± 0.03%), followed by PMvet212 (7.85 ± 0.02%). These values indicate low to moderate adhesion potential based on the MATH assay. The results are summarized in Table 4.
3.2.4. Hemolytic Activity (Safety Evaluation)
Hemolytic activity of the selected isolates was assessed on blood agar. Both PMvet212 and PMvet318 exhibited α-hemolysis, characterized by greenish discoloration surrounding the colonies, indicating partial hemolysis (Figure 2). No β-hemolytic activity was observed.
3.2.5. Antimicrobial Activity Against Escherichia coli and Staphylococcus aureus
The antibacterial activity of cell-free supernatants from the selected isolates was evaluated against Escherichia coli and Staphylococcus aureus. Both PMvet212 and PMvet318 demonstrated measurable inhibitory activity against the tested pathogens, with inhibition zones ranging from approximately 10 to 12 mm. The results are presented in Table 5.
3.3. Molecular Identification of Selected Isolates
The two selected isolates were further identified using MALDI-TOF mass spectrometry. Isolate PMvet212 showed a high-confidence match with Lactobacillus brevis (score ≥2.0), indicating reliable identification at the species level. In contrast, PMvet318 yielded a lower score (<2.0) and was identified at the genus level as Lactobacillus sp (Table 6). These results confirm that both isolates belong to the genus Lactobacillus, with one isolate identified at species level.
4. Discussion
The present study identified lactic acid bacteria (LAB) isolates from fecal samples of suckling piglets (7–28 days old) through a stepwise phenotypic screening approach. From an initial pool of 318 colonies, 135 Gram-positive, rod-shaped, catalase-negative isolates were retained for further evaluation. The detection of LAB in piglets at this early developmental stage is consistent with previous reports indicating that the gastrointestinal tract of piglets is rapidly colonized by Lactobacillus spp. prior to weaning [9,19]. This early colonization is influenced by maternal and environmental sources, including sow feces and milk, and plays a critical role in shaping gut microbial stability and disease resistance [5,8,21]. The present findings support the concept that fecal samples from suckling piglets represent a relevant source of host-adapted LAB for preliminary screening.
Although yeast-like isolates were detected during the initial screening process, these were excluded from further analysis to maintain consistency in the evaluation of bacterial isolates. Previous studies have demonstrated that yeast species may exhibit probiotic-like properties, including resistance to gastrointestinal conditions and pathogen inhibition [22,23]. However, the present study was designed to focus specifically on LAB, and therefore yeast isolates were not included in subsequent analyses. Future studies may investigate these yeast isolates separately to assess their potential role in swine gut health.
The acid and bile tolerance assays revealed substantial variability among isolates. Of the 135 LAB isolates, only two (PMvet212 and PMvet318) demonstrated detectable tolerance to acidic conditions (pH 3.1), while none survived at pH 2.0. This finding is consistent with previous studies indicating that tolerance to simulated gastric conditions is highly strain-dependent [24]. Both isolates also showed measurable tolerance to bile salts, with PMvet212 demonstrating greater resistance at 0.5% bile concentration. Such tolerance is important for bacterial survival in the small intestine, where bile salts exert antimicrobial effects [25]. However, given that the present study was based on a preliminary in vitro screening design using technical replicates, these findings should be interpreted as indicative rather than conclusive evidence of gastrointestinal survival.
Molecular identification using MALDI-TOF mass spectrometry provided additional insight into the taxonomic identity of the selected isolates. PMvet212 was identified as Lactobacillus brevis with high confidence, whereas PMvet318 was identified at the genus level as Lactobacillus sp. The identification of L. brevis is consistent with previous reports describing its presence in the gastrointestinal tract of pigs and its potential role in producing antimicrobial metabolites and modulating gut microbiota [26]. The relatively lower identification score for PMvet318 suggests limitations in database matching or strain variability, highlighting the need for further molecular confirmation, such as 16S rRNA gene sequencing, to achieve precise taxonomic classification. The integration of MALDI-TOF identification strengthens the reliability of the present findings by linking phenotypic characteristics with taxonomic information [13].
In addition to phenotypic characterization, the present study incorporated MALDI-TOF mass spectrometry for the identification of selected isolates. MALDI-TOF offers a rapid and cost-effective approach for bacterial identification based on protein fingerprinting, allowing routine classification at the genus and species levels within a short turnaround time [13]. In contrast, 16S rRNA gene sequencing remains the reference standard for bacterial taxonomy, particularly for resolving closely related species and confirming phylogenetic relationships. However, 16S sequencing is generally more time-consuming and resource-intensive compared to MALDI-TOF. Therefore, MALDI-TOF is increasingly used as a practical alternative for preliminary identification in microbiological studies, while molecular methods such as 16S rRNA sequencing are often recommended for confirmatory analysis [27].
Previous studies have demonstrated that MALDI-TOF can reliably identify members of the genus Lactobacillus, including species commonly associated with the gastrointestinal tract of animals [13,28]. In swine, several LAB species have been frequently reported, such as Lactobacillus reuteri, Lactobacillus salivarius, and Lactobacillus plantarum, which are known to contribute to gut microbial balance and host health [9,29]. These species have been associated with beneficial effects including inhibition of enteric pathogens, enhancement of intestinal barrier function, and modulation of immune responses.
Although the present study identified one isolate as Lactobacillus brevis and another at the genus level, these findings are consistent with the diversity of LAB populations in piglet gastrointestinal systems. Previous in vivo studies have shown that certain Lactobacillus species can improve growth performance, reduce diarrhea incidence, and enhance gut health in piglets [30,31]. However, it is important to emphasize that such effects are strain-specific and require comprehensive evaluation, including safety assessment and controlled in vivo trials. Therefore, while MALDI-TOF identification provides valuable preliminary taxonomic insight, further molecular confirmation and functional validation are necessary before translating these findings into practical applications.
Additional functional properties, including cell surface hydrophobicity and hemolytic activity, were also evaluated. The hydrophobicity values observed in this study were relatively low (approximately 7–12%), suggesting limited adhesion potential based on the MATH assay alone. However, bacterial adhesion is a multifactorial process involving surface proteins, extracellular polysaccharides, and host interactions [32]. Regarding safety, both PMvet212 and PMvet318 exhibited α-hemolysis, indicating partial hemolytic activity. Although α-hemolysis is less severe than β-hemolysis, current probiotic safety guidelines recommend the selection of non-hemolytic strains [10,33,34]. Therefore, despite their functional properties, these isolates require further safety evaluation before any consideration of application.
The antibacterial activity observed in PMvet212 and PMvet318 against Escherichia coli and Staphylococcus aureus suggests the production of inhibitory metabolites, such as organic acids or bacteriocin-like compounds [18,24]. The inhibition zones (10–12 mm) are consistent with values reported for LAB isolated from piglets [18], indicating moderate antibacterial activity. However, as the cell-free supernatants were not neutralized prior to testing, the observed inhibition is likely influenced by acid-mediated effects. Future studies should include pH-neutralized assays to distinguish between organic acid and bacteriocin-associated antimicrobial activity. Overall, the present study provides preliminary screening data on LAB isolates from piglets and highlights the need for further molecular characterization, safety assessment, and in vivo validation to fully evaluate their functional potential.
5. Conclusions
This study provides a structured in vitro phenotypic screening of lactic acid bacteria isolated from feces of suckling piglets. From 318 initial colonies, 135 Gram-positive, rod-shaped, catalase-negative isolates were retained for evaluation, among which only two isolates (PMvet212 and PMvet318) demonstrated detectable tolerance to acidic and bile conditions, along with moderate antibacterial activity. Identification using MALDI-TOF mass spectrometry revealed that PMvet212 was closely related to Lactobacillus brevis, while PMvet318 was identified at the genus level as Lactobacillus sp. Despite these functional properties, both isolates exhibited α-hemolytic activity, indicating that further safety assessment is required. The findings of this study should be interpreted as preliminary evidence derived from an exploratory screening approach rather than confirmation of probiotic suitability. Although selected isolates demonstrated characteristics associated with gastrointestinal stress tolerance and antimicrobial activity, additional investigations including molecular confirmation, genome-based safety evaluation, and controlled in vivo studies are necessary before any practical application can be considered. Overall, this study provides a useful baseline for the identification and selection of host-associated LAB from piglets and supports future research aimed at developing evidence-based microbial strategies for improving gut health in swine production systems.
Author Contributions
Writing-original draft, Conceptualization, Data curation, Formal analysis, Methodology, Writing-review and editing, Panuwat Yamsakul; Sample collection, Investigation, Data curation, Duangporn Pichpol; Methodology, Laboratory, Montira Intanon and Nattakarn Awaiwanont. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Faculty of veterinary medicine, Chiang Mai university, Thailand and Fundamental Fund 2026 (4828004) Chiang Mai university, Thailand.
Institutional Review Board Statement
All microbiological work was reviewed and approved by the Institutional Biosafety Committee (IBC) of Chiang Mai University (Approval No. CMUIBC A-0763004).
Informed Consent Statement
Not applicable.
Data Availability Statement
Relevant information is included in the article.
Acknowledgments
The authors also thank the faculty of veterinary medicine and Fundamental Fund 2026 (4828004) Chiang Mai university, Thailand for supporting the budget.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fairbrother, J.M.; Nadeau, E.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6(1), 17–39. [Google Scholar] [CrossRef]
- Luppi, A. Swine enteric colibacillosis: Diagnosis, therapy, and antimicrobial resistance. Porcine Health Manag. 2017, 3, 16. [Google Scholar] [CrossRef]
- Davis, M.E.; Maxwell, C.V.; Brown, D.C.; de Rodas, B.Z.; Johnson, Z.B.; Kegley, E.B.; Hellwig, D.H.; Dvorak, R.A. Effect of dietary mannan oligosaccharides and/or copper sulfate on growth and immunocompetence of pigs. J. Anim. Sci. 2002, 80, 2887–2894. [Google Scholar] [CrossRef]
- Moredo, F.A.; Pineyro, P.E.; Marquez, G.C.; Sanz, M.; Colello, R.; Etcheverria, A.; Padola, N.L.; Quiroga, M.A.; Perfumo, C.J.; Galli, L.; Leotta, G.A. Enterotoxigenic Escherichia coli subclinical infection in pigs: Bacteriological and genotypic characterization and antimicrobial resistance profiles. Foodborne Pathog. Dis. 2015, 12, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Bogere, P.; Choi, Y.J.; Heo, J. Probiotics as alternatives to antibiotics in treating post-weaning diarrhoea in pigs. S. Afr. J. Anim. Sci. 2019, 49, 403–416. [Google Scholar] [CrossRef]
- Laird, T.J.; Abraham, S.; Jordan, D.; Pluske, J.R.; Hampson, D.J.; Trott, D.J.; Dea, M.O. Porcine enterotoxigenic Escherichia coli: Antimicrobial resistance and development of microbial-based alternative control strategies. Vet. Microbiol. 2021, 258, 109117. [Google Scholar] [CrossRef] [PubMed]
- Fouhse, J.M.; Zijlstra, R.T.; Willing, B.P. The role of gut microbiota in the health and disease of pigs. Anim. Front. 2016, 6(3), 30–36. [Google Scholar] [CrossRef]
- Petri, D.; Hill, J.E.; Van Kessel, A.G. Microbial succession in the gastrointestinal tract of pre-weaned pigs. Livest. Sci. 2010, 133, 107–109. [Google Scholar] [CrossRef]
- Dowarah, R.; Verma, A.K.; Agarwal, N.; Singh, P.; Singh, B.R. Selection and characterization of probiotic lactic acid bacteria and its impact on growth, nutrient digestibility, and health in weaned piglets. PLoS ONE 2018, 13, e0192978. [Google Scholar] [CrossRef]
- FAO/WHO. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; World Health Organization: Rome, Italy, 2006. [Google Scholar]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Guidance on the assessment of the safety of feed additives for the target species. EFSA J. 2018, 16, e05206.
- Ozugul, F.; Hamed, I. The importance of lactic acid bacteria for prevention of bacterial growth and biogenic amine formation: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1660–1671. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- Guo, X.H.; Kim, J.M.; Nam, H.M.; Park, S.Y.; Kim, J.M. Screening lactic acid bacteria from swine origins for multistrain probiotics based on in vitro functional properties. Anaerobe 2010, 16, 321–326. [Google Scholar] [CrossRef]
- Ren, D.; Li, C.; Qin, Y.; Yin, R.; Du, S.; Ye, F.; Liu, C.; Liu, H.; Wang, M.; Li, Y.; Sun, Y.; Li, X.; Tian, M.; Jin, N. In vitro evalua-tion of the probiotic and functional potential of Lactobacillus strains isolated from fermented food and human intestine. Anaerobe 2014, 30, 1–10. [Google Scholar] [CrossRef]
- Ekmekci, H. Characterization of vaginal lactobacilli coaggregation ability with Escherichia coli. Microbiol. Immunol. 2009, 53, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Adimpong, D.B.; Nielsen, D.S.; Sørensen, K.I.; Derkx, P.M.F.; Jespersen, L. Genotypic characterization and safety assessment of lactic acid bacteria from indigenous African fermented food products. BMC Microbiol. 2012, 12, 75. [Google Scholar] [CrossRef] [PubMed]
- Sirichokchatchawan, W.; Pupa, P.; Praechansri, P.; Amin, N.; Tanasupawat, S.; Sonthayanon, P.; Prapasarakul, N. Autochthonous lactic acid bacteria isolated from pig feces in Thailand show probiotic properties and antibacterial activity against enteric pathogenic bacteria. Microb. Pathog. 2018, 119, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Petsuriyawong, B.; Khunajakr, S. Screening of probiotic lactic acid bacteria from piglet feces. Kasetsart J. Nat. Sci. 2011, 45, 245–253. [Google Scholar]
- Buxton, R. Blood agar plates and hemolysis protocols. American Society for Microbiology. 2005. Available online: https://asm.org/getattachment/7ec0de2b-bb16-4f6e-ba07-2aea25a43e76/protocol-2885.pdf.
- Mustakim, M.; Sinawat, S.; Salleh, S.N.; Purwati, E.; Alias, R.; Mohamad, S.A.; Mat Issa, Z. Human milk as a potential source for isolation of probiotic lactic acid bacteria: A mini review. Food Res. 2019, 4, 274–285. [Google Scholar] [CrossRef]
- Czerucka, D.; Piche, T.; Rampal, P. Yeast as probiotics-Saccharomyces boulardii. Aliment. Pharmacol. Ther. 2007, 26, 767–778. [Google Scholar] [CrossRef]
- Elghandour, M.M.Y.; Tan, Z.L.; Abu Hafsa, S.H.; Adegbeye, M.J.; Greiner, R.; Ugbogu, E.A.; Cedillo Monroy, J.; Salem, A.Z.M. Saccharomyces cerevisiae as a probiotic feed additive to non- and pseudo-ruminants: A review. J. Appl. Microbiol. 2020, 128, 658–674. [Google Scholar] [CrossRef]
- Yeo, S.; Lee, S.; Park, H.; Shin, H.; Holzapfel, W.; Huh, C.S. Development of putative probiotics as feed additives: Validation in a porcine-specific gastrointestinal tract model. Appl. Microbiol. Biotechnol. 2016, 100, 10043–10054. [Google Scholar] [CrossRef]
- Gotcheva, V.; Hristozova, E.; Hristozova, T.; Guo, M.; Roshkova, Z.; Angelov, A. Assessment of potential probiotic properties of lactic acid bacteria and yeast strains. Food Biotechnol. 2002, 16, 211–225. [Google Scholar] [CrossRef]
- Zhang, L.; Li, N.; Caicedo, R.; Neu, J. Probiotic Lactobacillus species and their role in gastrointestinal health. Front. Microbiol. 2022, 13, 845678. [Google Scholar]
- Clark, A.E.; Kaleta, E.J.; Arora, A.; Wolk, D.M. Matrix-assisted laser desorption ionization–time of flight mass spectrometry: A fundamental shift in the routine practice of clinical microbiology. Clin. Microbiol. Rev. 2013, 26, 547–603. [Google Scholar] [CrossRef] [PubMed]
- Dušková, M.; Šedo, O.; Kšicová, K.; Zdráhal, Z.; Karpíšková, R. Identification of lactobacilli isolated from food by MALDI-TOF MS. Int. J. Food Microbiol. 2012, 159, 107–114. [Google Scholar] [CrossRef]
- Wang, H.; Xu, R.; Zhang, H.; Su, Y.; Zhu, W. Swine gut microbiota and its interaction with host nutrient metabolism. Anim. Nutr. 2020, 6, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhou, X.; Birch Hansen, L.H.; Sheng, Y.; Yu, B.; He, J.; Yu, J.; Zheng, P. Complex probiotics reduce diarrhea by enhancing immunity and balancing gut microbiota in weaned piglets. Front. Immunol. 2025, 16, 1629044. [Google Scholar] [CrossRef]
- Qiao, J.; Li, H.; Wang, Z.; Wang, W. Effects of Lactobacillus acidophilus dietary supplementation on performance, intestinal barrier function, rectal microflora, and serum immune function in weaned piglets challenged with Escherichia coli lipopolysaccharide. Antonie Van Leeuwenhoek 2015, 107, 883–891. [Google Scholar] [CrossRef]
- Garcia-Cayuela, T.; Korany, A.; Bustos, I.; Cadinanos, L.; Requena, T.; Pelaez, C.; Martinez-Cuesta, M.C. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- Anadón, A.; Martínez-Larrañaga, M.R.; Martínez, M.A. Probiotics for animal nutrition in the European Union: Regulation and safety assessment. Regul. Toxicol. Pharmacol. 2006, 45, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Zendeboodi, F.; Khorshidian, N.; Mortazavian, A.M. Probiotic: Conceptualization from a new approach. Curr. Opin. Food Sci. 2020, 32, 103–123. [Google Scholar] [CrossRef]
Figure 1.
Microscopic images of representative isolates after Gram staining, observed under a light microscope at 1,000 × magnification: (a) PMvet120; (b) PMvet151; (c) PMvet183; (d) PMvet212; and (e) PMvet318.
Figure 1.
Microscopic images of representative isolates after Gram staining, observed under a light microscope at 1,000 × magnification: (a) PMvet120; (b) PMvet151; (c) PMvet183; (d) PMvet212; and (e) PMvet318.

Figure 2.
Hemolytic patterns of the five LAB isolates on tryptic soy agar supplemented with 5% sheep blood after 48 h of incubation at 37 °C. Isolates PMvet120 (b), PMvet151 (c), and PMvet183 (d) exhibited γ-hemolysis (no lysis), whereas PMvet212 (e) and PMvet318 (f)showed α-hemolysis (partial lysis, greenish discoloration). Staphylococcus aureus ATCC 6538 (a) was included as a positive control, displaying β-hemolysis (complete lysis with a clear zone).
Figure 2.
Hemolytic patterns of the five LAB isolates on tryptic soy agar supplemented with 5% sheep blood after 48 h of incubation at 37 °C. Isolates PMvet120 (b), PMvet151 (c), and PMvet183 (d) exhibited γ-hemolysis (no lysis), whereas PMvet212 (e) and PMvet318 (f)showed α-hemolysis (partial lysis, greenish discoloration). Staphylococcus aureus ATCC 6538 (a) was included as a positive control, displaying β-hemolysis (complete lysis with a clear zone).


Table 1.
Interpretation of antimicrobial activity based on inhibition zone diameter.
| Symbol | Interpretation | Inhibition Zone Diameter (mm) |
|---|---|---|
| - | Non-inhibition | <5 |
| + | Weak inhibition | >5 |
| ++ | Intermediate inhibition | >10 |
| +++ | Strong inhibition | >15 |
| ++++ | Very strong inhibition | >20 |
Table 2.
Survival of selected LAB isolates at pH 3.1 after 3 h of incubation at 37 °C. Values are expressed as mean ± standard deviation (n = 3).
Table 2.
Survival of selected LAB isolates at pH 3.1 after 3 h of incubation at 37 °C. Values are expressed as mean ± standard deviation (n = 3).
| Isolate ID | log CFU/mL | |
| Initial Count (average) | pH 3.1 | |
| 212 | 6.37±0.5 | 5.60±0.5 |
| 318 | 5.56±0.6 | |
Table 3.
Survival of LAB isolates in MRS broth containing different bile salt concentrations after 24 h of incubation at 37 °C. Values are expressed as mean ± standard deviation (n = 3).
Table 3.
Survival of LAB isolates in MRS broth containing different bile salt concentrations after 24 h of incubation at 37 °C. Values are expressed as mean ± standard deviation (n = 3).
| Isolate ID | 0% (0 h) | 0% (24 h, Control) | 0.3% (24 h) | 0.5% (24 h) |
|---|---|---|---|---|
| 212 | 6.81 ± 0.4 | 9.66 ± 0.5 | 7.81 ± 0.3 | 6.56 ± 0.4 |
| 318 | 6.00 ± 0.6 | 9.08 ± 0.6 | 7.38 ± 0.5 | <3.00 |
Table 4.
Cell surface hydrophobicity (%) of selected LAB isolates as determined by the MATH assay using xylene. Values are expressed as mean ± standard deviation (n = 3).
Table 4.
Cell surface hydrophobicity (%) of selected LAB isolates as determined by the MATH assay using xylene. Values are expressed as mean ± standard deviation (n = 3).
| Isolate ID | OD600 (before mixing with xylene) | OD600 after mixing (0 min) | OD600 after phase separation (30 min) | %Hydrophobicity (mean±SD) |
|---|---|---|---|---|
| 212 | 0.871 ± 0.02 | 1.014 ± 0.02 | 0.924 ± 0.02 | 7.85 ± 0.02 |
| 318 | 0.935 ± 0.02 | 1.171 ± 0.03 | 1.010 ± 0.02 | 12.38 ± 0.03 |
Table 5.
Antimicrobial activity of cell-free supernatants (CFS) from selected LAB isolates against Escherichia coli (field isolate) and Staphylococcus aureus ATCC 6538, determined by the agar well diffusion assay. Values are expressed as mean ± standard deviation (n = 3).
Table 5.
Antimicrobial activity of cell-free supernatants (CFS) from selected LAB isolates against Escherichia coli (field isolate) and Staphylococcus aureus ATCC 6538, determined by the agar well diffusion assay. Values are expressed as mean ± standard deviation (n = 3).
| Isolate ID | Clear zone (mm) - E. coli | Clear zone (mm) - S. aureus |
|---|---|---|
| PMvet212 | 12.50 ± 0.4 | 11.75 ± 0.5 |
| PMvet318 | 11.50 ± 0.3 | 10.50 ± 0.4 |
| Control (2% lactic acid) | 18.50 ± 0.5 | 15.25 ± 0.4 |
Table 6.
Identification of selected LAB isolates using MALDI-TOF.
| Isolate ID | Closest match | Score value | Identification level |
|---|---|---|---|
| PMvet212 | Lactobacillus brevis | 2.1–2.3 | Species level |
| PMvet318 | Lactobacillus sp. | 1.7–1.9 | Genus level |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



