Submitted:
07 April 2026
Posted:
08 April 2026
You are already at the latest version
Abstract
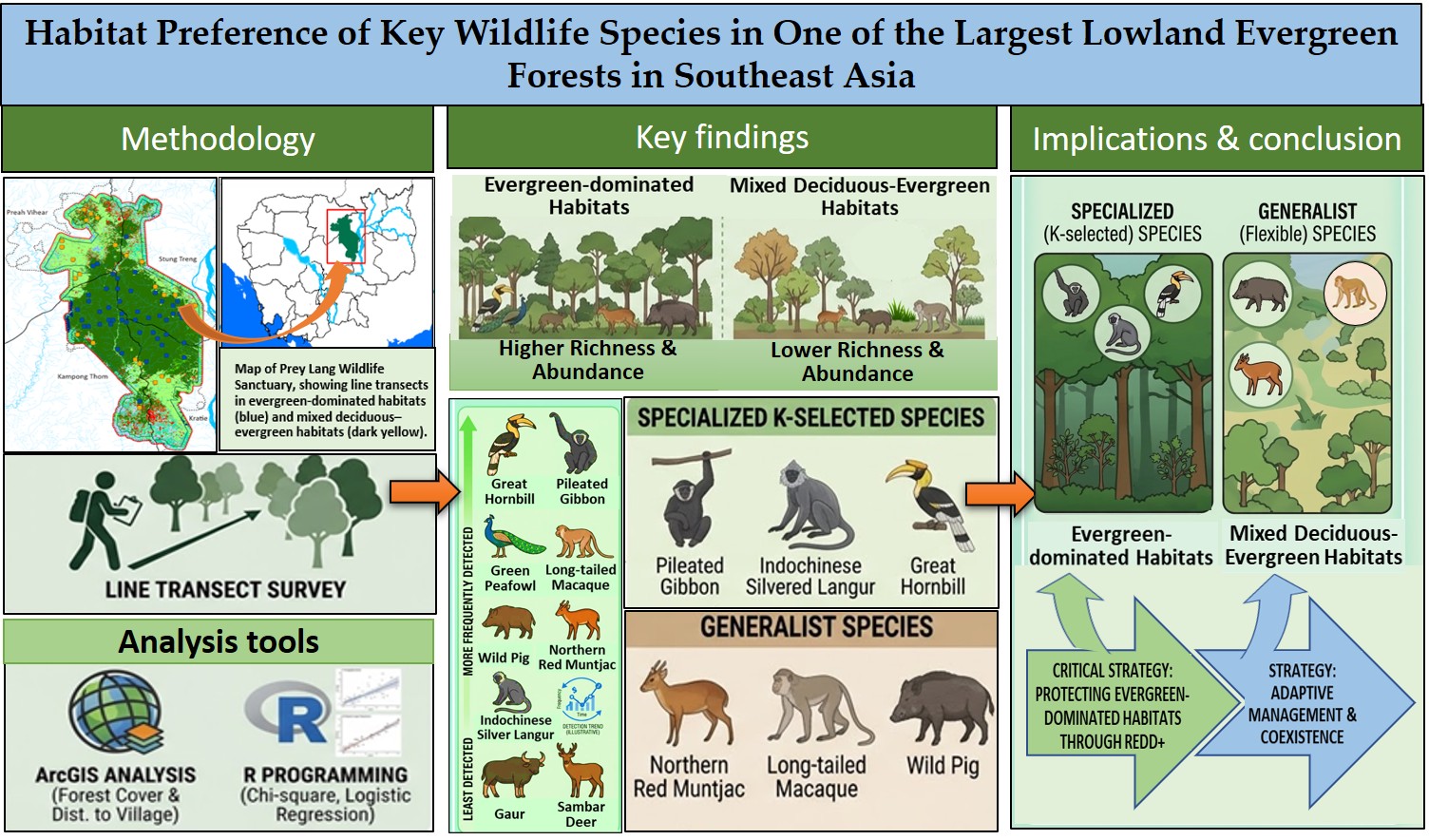
Wildlife plays a vital role in maintaining ecological balance and biodiversity, relying on habitats that provide shelter, food, and essential resources. This study investigated wildlife distribution and diversity across the REDD+ program area in Cambodia’s Prey Lang Wildlife Sanctuary, a lowland evergreen forest ecosystem, and assessed the effects of forest habitats and anthropogenic pressure on their distribution. We used square transects for sampling and ArcGIS to calculate forest cover and distance to the nearest village as a proxy for human impact. Overall, we recorded seven mammals and two birds, with the great hornbill most frequently detected, followed by pileated gibbon, wild pig, long-tailed macaque, green peafowl, northern red muntjac, and Indochinese silvered langur; gaur and sambar deer were least detected. Wildlife richness and abundance were higher in evergreen-dominated habitats than in mixed deciduous-evergreen forests. Certain K-selected species, including pileated gibbon, Indochinese silvered langur, and great hornbill, were highly specialized and preferred intact forests, whereas generalist species such as northern red muntjac, long-tailed macaque, and wild pig showed ecological flexibility in habitat use. These findings emphasize tailored conservation strategies: protecting intact evergreen forests via REDD+ supports specialized species, while adaptive management in mosaic landscapes benefits generalists, enhancing wildlife conservation and sustainable management of Prey Lang Wildlife Sanctuary.
Keywords:
distance sampling
; transect survey
; K-selected species
; generalist species
; mammals
; birds
; great hornbill
1. Introduction
Wildlife is an important component in natural ecosystems by contributing to ecological balance and biodiversity [1]. Their survival, reproduction and daily activities in the ecosystem require a suitable habitat that provides essential resources such as food, water, shelter, and space [2]. The required amount of these resources is varied from one to another taxonomic group and habitat types. For instance, some K-selected species such as the Asian elephant and gaur in lowland evergreen forest are more associated with dense forest cover and perennial water sources [3,4]. Bird species such as the great hornbill are influenced by the availability of large fruiting trees and nesting cavities [5], and others such as the giant ibis, lesser adjutant, and Asian woolly-necked stork have been mainly recorded from protected areas where deeper waterholes and wetlands are present [6].
Lowland evergreen forests are globally recognized for their exceptional biodiversity and complex ecological interactions, particularly species habitat relationships [7]. In mainland Southeast Asia, Cambodia’s Prey Lang Wildlife Sanctuary (PLWS) represents one of the largest remaining tracts of lowland evergreen forest, comprising a mosaic of evergreen and deciduous forest types [7]. PLWS supports a remarkable diversity of wildlife, including iconic species such as the Asian elephant, Malayan sun bear, pileated gibbon, long-tailed macaque, great hornbill, and green peafowl [8,9]. Notably, species richness recorded form this sanctuary are highest in the dense forest areas concentrated near the core zone, whereas wildlife abundance appears more randomly distributed across the broader landscape; for example, wild pig, Malayan sun bear, northern red muntjac, and long-tailed macaque [9,10].
In Cambodia, diverse forest ecosystems across protected areas continue to support rich wildlife biodiversity, yet research on species’ habitat preferences remains limited. Some exceptions include the study of Gray et al. [11] investigating the habitat requirements of Bengal florican and found that the species depends on grassland habitats maintained by traditional agricultural practices. Gray et al. [10] assessed the status and habitat preferences of wild cattle and large carnivores in eastern Cambodia, indicating that these species rely on mosaics of deciduous dipterocarp and mixed deciduous/semi-evergreen forest. Similarly, Gray and Phan [12] reported that large mammals in Phnom Prich Wildlife Sanctuary prefer mixed-deciduous and semi-evergreen forest. While these contributions provide valuable insights, the habitat preferences of other wildlife species in different protected areas, including the PLWS, remain poorly understood. Moreover, the influence of different forest cover classes (e.g., evergreen forest, semi-evergreen forest, dry dipterocarp forest) and anthropogenic pressures on wildlife distribution is scarcely documented, despite increasing threats from logging, land encroachment, and hunting.
To address this need and contribute to biodiversity research on habitat preferences of key species in PLWS, we (1) investigate the distribution patterns, species diversity, richness and abundance of ecologically important mammal and bird species across the sanctuary, and (2) assess the levels of impact of evergreen forest proportion and anthropogenic pressure on the faunal distribution. The results of this study also serve as a baseline survey using the transect sampling for long-term biodiversity monitoring in the area, and a preliminary assessment of the REDD+ project implementation. Previous records of the target species are also discussed and compared with their present records in this study to confirm their existence in the PLWS.
2. Methodology
2.1. Study Site
The study was conducted at PLWS, located in the central plains of Cambodia which comprises areas from four provinces, including Stung Treng, Kampong Thom, Preah Vihear, and Kratie. The total area of the PLWS is 4316.83 km2, mainly attributed to evergreen forest (57%) and de-ciduous forest (26%). PLWS experiences a tropical climate with distinct wet and dry seasons. The wet season typically spans from May to October, with the southwest monsoon bringing substantial rainfall, while the dry season lasts from November to April. During this period, the region receives approximately 1,300 to 1,800 mm of precipitation annually.
PLWS is essential to the well-being of over 250,000 indigenous and local people. The majority of local communities live in the western and northern parts of Prey Lang (particularly in Kampong Thom and Preah Vihear provinces) (Figure 1). The forest is deeply woven into their cultural and spiritual lives, and they rely heavily on the rich ecosystem services from the forest to meet their social and economic needs. Local villagers engage in fisheries and agriculture, and gather resin, building materials, medicine, and local food from the forest, with many depending on these resources for their livelihoods [13].
Our study site is located in the REDD+ project area (3752 km2) of the PLWS. As of the 2023, this REDD+ project area is mainly characterized by evergreen forests (69%), deciduous forests (24.35%), semi-evergreen forests (0.14%), cropland (0.33%), grassland (1.94%), and other planta-tions (0.043%). Forest clearance for agriculture is also occurring within the sanctuary, with an estimated rate of approximately 0.81% [14].
2.2. Target Species
Fifteen species of mammals and birds were targeted for the survey (Table 1). The selection of these is based on the fact that their occurrences are crucial for informing conservation strategies and facilitating the development of targeted management plans, particularly for species experiencing population declines. The fifteen species are also of high conservation value (i.e., Threatened category in the IUCN Red List) and their ecological and biological significance in the ecosystem. For instance, gibbons are critical for seed dispersal, while dholes serve as top predators, regulating prey population in the landscape [15,16]. These species play pivotal roles in maintaining ecological balance and are therefore essential components of conservation monitoring efforts [17]. For additional information on each species, we refer to Table 1 below.
2.3. Transect Designs and Development
In this study, we used a 1-km side square transect (i.e., 1-km2 grid) as the distance sampling method. This approach is an effective method to estimate key species density and population size and is a standard monitoring approach in two other protected landscapes in Cambodia [31].
A total of 74 1-km side squares were randomly selected across the REDD+ project area in PLWS (Figure 2) using Global Information System ArcMap 10.81. We initially randomly selected 400 grids 1 km2 grid. We removed grids that were adjacent or connected to each other to allow for independence and areas that had been previously deforested. This resulted in a reduced set of 149 grids. Due to time, resources, and logistic constraints, we randomly selected 74 grids from the 149 grids. The number of grids selected eventually corresponds to ~2.4% of the total area. This approach of using a subset of grids is commonly practiced in ecological studies, where sampling a smaller representative area is both efficient and effective for data collection and analysis [32].
Comprehensive training sessions of the field teams were conducted from July 26–28, 2022, to ensure that sampling and methodological protocols were highly adhered (Figure 2a). Transects were established and marked using a compass and paints to direct and mark the transect trails and corners. The transect trails along the four sides of each square were cleared using machetes, creating a trail width of ~ 1 m to minimize the impact on vegetation covers.
For each square transect, a trail of 4-km cumulative length (4 sides * 1-km length) required a collaborative effort of 8 individuals, with an average of 6- to 8-hour work. We spent approximately 4 months to complete the 74 square transects development at the study site, commencing on September 13, 2022, and concluding on December 29, 2022. The total length of all transect trails is 296 km.
2.4. Data Collection, Processing, and Preparation
The data collection period was from January 13 to May 21, 2023, corresponding to the second half of the dry season (which runs from around December through May). The order of square transects establishment was arranged for the data collection, meaning that the square transect that was first developed was chosen for the first data collection, and so forth. In this way, each square transect had about four months without any human presence. We adopted this process to minimize the effect of human-made trails on animal behaviors (e.g., foraging, courting, etc.).
Each square transect was walked twice per day, once in the morning from 06:00 – 11:30 and once from 13:00– 16:30. The morning team began their walk from right to left (4 km) at a starting point and ended their walk when arriving at the starting point. The afternoon team reversed their walk direction from left to right (4 km), beginning and ending also at the same starting point (Figure 2b). Each team consisted of two trained observers who walked along the transect line. One observer focused on the area directly along or near the transect, while the other observer focused on the areas to the left and right of the transect [31]. Only animals detected through direct visual sightings were recorded; other evidence such as calling or signs was not included in this study. During the survey, team members remained silent to avoid disturbing wildlife. They walked slowly along the transects, stopping whenever animals were observed. If no animals were detected, they paused every 50 meters to conduct a careful search.
A Global Positioning System (GPS) unit was used to record the relative location of detected animals. Some other field equipment was also used: a timer to record the time during the survey, a compass to measure the bearing from the observer to the detected animals, and a laser rangefinder to measure the distance from the observer to the detected animals. A measuring tape was used instead in case the laser rangefinder was not applicable, e.g., in a densely forested area.
The perpendicular distance (PD) from the transect line to the detected animals was computed using the formula: D = B * Sinα, where B is the distance from the observer to the detected animals and α is the bearing angle between the walking direction along the transect and the detected animals (Figure 2a). The perpendicular distance was also calculated for further analysis to estimate the density of target species.
2.5. Forest Cover Proportion Calculation and Anthropogenic Pressure
The estimation of forest cover proportion and anthropogenic pressure is used to assess the levels of their impact on the faunal distribution. The percentage of forest cover classes (e.g., evergreen forest, cropland, dry dipterocarp forest, semi-evergreen forest, shrub, other plantations, rice field, and grass) representing the forest habitat for each transect was calculated from an area of 9 km2, combining each surveyed transect located in the center and surrounding eight 1-km side squares. The process of combining the surrounding 8 km2 was the same across the 74 square transects. An area of 9 km2 is indicative of habitat preference for animals’ daily activities, especially foraging [34]. Based on the 2023 forest cover, deforestation and administrative data [14], we used ArcGIS version 3.4.0 to calculate the percentage of each forest cover class characterizing each surveyed transect, and the distances from each transect to the nearest village center and the nearest deforested areas. These distances are considered proxies for anthropogenic pressure on wildlife distribution.
From the calculated percentage of forest cover classes characterizing each transect, we further categorized them into two main habitat types: the forest habitat dominated by evergreen forest or the highly intact forest (hereafter refers as high-intact) habitat, which comprises ≥ 80% of evergreen forest, and the forest habitat dominated by deciduous and evergreen forest or the moderately intact forest (hereafter refers as moderate-intact) habitat, which comprises < 80% of evergreen forest. We used these thresholds because most of the global intact forest habitats comprise an approximately 75% of evergreen forest [35]. This selecting criterion (80% thresholds) is important to indicate landscape biodiversity hotspots or climate-sensitive regions for ensuring long-term sustainability of the study landscape, PLWS. The two categories of forest habitats were classified to analyze their association with wildlife diversity, species composition, and occurrences.
2.6. Data Analysis
2.6.1. Species Distribution, Detection, and Diversity
For each transect, the total number of species (richness), individuals (abundance), and the diversity index (Shannon index, the higher value indicates the higher diversity) were computed. We also calculated the encounter rate (per km) for every sighted animal in each square transect by taking the total number of sightings of each species divided by the length of the transect walked, as described by Pramod et al. [36]. ArcMap 10.81 was then used to project the number of species and their abundance recorded onto a map to illustrate their distribution patterns. To detect differences, species diversity metrics, such as richness, abundance, and Shannon diversity index (diversity H), were compared between the high- and moderate-intact forest habitats using an unpaired t-test, given that the two datasets had a normal distribution and homogeneity of variance; otherwise, the alternative Mann-Whitney U test was used.
2.6.2. Influences of Habitat Characteristics and Anthropogenic Pressure
Logistic regression analysis was employed to evaluate the influence of habitat characteristics (evergreen forest, cropland, dry dipterocarp forest, semi-evergreen forest, shrub, rice, other plantation, rice and grass) and anthropogenic pressure (distances from each transect to the nearest village center and the nearest deforested areas) on the presence or absence of detected wildlife species. The logistic regression model was used because it can estimate the probability of species occurrence at each site, revealing how different habitat characteristics and proximity to human activities and settlements influence habitat suitability of wildlife species [37,38]. In this model, the presence/absence of wildlife species was fitted using the standardized predictor variables, which were measured on different scales (e.g., kilometers and hectares) [39]. To identify which predictors mainly influence wildlife occurrence distribution, we performed a forward stepwise selection starting from a null model (no predictor included) to the final model that selected the most important predictors from the full model (all predictors included). This process resulted in a parsimonious logistic regression model, based on the lowest value of the Akaike’s Information Criterion (AIC), that included only the predictors that significantly contributed to explaining the variation in the presence/absence of the wildlife species. Moreover, to analyze whether any of the key species has a forest habitat preference, we used a chi-square test to identify a significant association between the presence-absence of each species and the two categorized forest habitat types (high- and moderate-intact habitats). For the analysis that yielded significant association results (p <0.05), we sorted the frequencies of only the presence data of the species and then compared their frequency between the two forest habitat types using also the Chi-Square test. This two-step chi-square analysis provides a sound and reliable result indicating which condition of the forest is the preferred habitat of sighted animals and thus informing conservation strategies in human-impacted landscapes. All statistical analysis was performed using the R statistical program, and the p-value <0.05 was considered significant for all analysis.
3. Results
3.1. Distribution Patterns and Diversity
Out of the fifteen species targeted, we detected only nine during the entire course of the survey. Seven species were mammals, and the remaining two species were birds. The most detected animal were great hornbills (B. bicornis, 76 sightings), followed by pileated gibbons (H. pileatus, 31), wild pigs (S. scrofa, 25), and long-tailed macaques (M. fascicularis, 23); whereas the least detected species, with only one sighting, were gaur (B. gaurus) and sambar deer (R. unicolor) (Table 2). We found no distinct spatial pattern emerged from the observational data across the sanctuary, but species counts were notably lowest in the northern sections of the park (Figure 3a) and abundance were highest around the central area of the park (Figure 3b). This pattern was similarly observed for the four most detected species (Figure 4a-d). For the least detected species, however, the small number of detections prevented a clear spatial inference (Figure A1).
Overall, we detected an average of 1.6±1.27 species, 7.6±9.75 individuals, and 0.4±0.46 of diversity H per transect. When computed based on each forest habitat type, the high-intact habitat supported an average of 1.9±1.4 species, 8.5±10.4 individuals, and 0.49±0.49 of diversity H; while the moderate-intact habitat supported an average of 1.0±0.89 species, 5.8±8.3 individuals, and 0.21±0.29 of diversity H per transect. Based on the Mann-Whitney U test, species richness (p = 0.008) and species diversity H (p = 0.015) were found to be significantly higher in the high-intact habitat compared to the moderate-intact habitat (Figure 5).
3.2. Habitat Preference and Anthropogenic Pressure
Based on the logistic regression model, evergreen forest was selected as the most important variable, appearing in the final models for three species – great hornbill, pileated gibbon, and northern red muntjac. It had a positive association with the occurrence of great hornbill (coefficient = 1.54) and pileated gibbon (0.69), but a negative association with northern red muntjac (–2.67). Dry dipterocarp forest was selected as the key variable negatively influencing the occurrence of green peafowl (–798.56) and long-tailed macaque (–1.01), while other plantations positively associated with the occurrence of the northern red muntjac (5.23) and long-tailed macaque (0.46). Cropland, rice field, and the distance to nearest village, were important variables for only one species; cropland exhibited a positive association (30.22) with green peafowl, rice field and distance to nearest village showed negative associations with Indochinese silvered langur (–506.7) and northern red muntjac (–3.6), respectively. No predictors were retained as the key predictors in the final model for wild pig (Table 3).
From the comparative analysis of species-habitat preference, we found that the presence-absence of great hornbill (Chi-squared = 6.87, p = 0.009), long-tailed macaque (Chi-squared = 6.89, p = 0.009), northern red muntjac (Chi-squared = 9.78, p = 0.002), and wild pig (Chi-squared = 4.47, p = 0.034), were significantly dependent on the conditions of the forest habitats (high- vs. moderate-intact habitats). The presence probability of great hornbill, long-tailed macaque, and wild pig is strongly dependent on the high-intact forest habitat, whereas that of northern red muntjac was entirely dependent on the moderate-intact habitat (Figure 6a-g).
Further analysis of the comparison of species presence data revealed a significantly high occurrence frequency of great hornbill, long-tailed macaques, pileated gibbon, and wild pig in the high-intact forest habitats (Figure 6h).
4. Discussion
We detected nine species out of the 15 targeted species that are important for ecosystem functions, e.g. Indochinese silvered langur regulates forest growth and structures, pileated gibbon distributes plant seeds across their habitat, while others play different important roles in their ecosystems such as seed dispersal, plant and insect population control, vegetation modification, and forest regeneration (see Table 1).
Average species richness and abundance were significantly higher in high-intact forest habitats, suggesting these areas are preferred by forest-dependent wildlife. This observation is consistent with Gibson et al. [40] who emphasized the critical and irreplaceable role of high-intact forests in sustaining tropical biodiversity. Compared to moderate-intact forests, high-intact forests generally support greater biodiversity due to increased food availability, numerous microhabitats, and diverse tree compositions [41]. These forests also provide essential cover from predators and protection from environmental and anthropogenic disturbances [42]. Furthermore, they demonstrate greater resistance to short-term climatic anomalies such as droughts and wildfires, and support high intraspecific genetic diversity, which enhances species’ adaptability to changing environmental conditions [43].
The analysis of habitat preference and anthropogenic pressure reveal a clear ecological gradient among the detected species in relation to forest integrity. Highly specialized species like great hornbill and pileated gibbon are strongly associated with high-intact forests, underscoring the need for strict protection of evergreen forest ecosystems to ensure their survival [44,45]. The Indochinese silvered langur, also a specialized species, showed a negative association with rice fields, indicating its sensitivity to human-modified landscapes and highlighting the detrimental impacts of agricultural expansion. These findings align with global studies demonstrating the vulnerability of K-selected species such as gibbons and langurs, and forest-dependent species (hornbills) to habitat loss and fragmentation [46,47].
On the other hand, species such as the northern red muntjac, long-tailed macaque, and wild pig exhibited greater adaptability. These species were found either in the high- or moderate-intact forests or human-modified landscapes including plantations, croplands, and areas near villages (Figure 6, Table 3). Their presence in these environments suggests either a lower sensitivity to disturbance or an ability to exploit available resources. While this adaptability may indicate resilience, it could also reflect shifts in ecological roles or increased reliance on human-provided resources, potentially leading to human-wildlife conflicts. For instance, the northern red muntjac is capable of inhabiting a wide range of habitats, including various forest types and dense vegetation [48]. Long-tailed macaques selectively use human-modified habitats when food and cover are sufficient [49]. Wild pigs in croplands can cause significant agricultural damage, a common issue across Southeast Asia [50]. The green peafowl showed a strong negative association with dry dipterocarp forest (DFF) and a positive association with cropland (CRL), suggesting a more nuanced habitat use. While pheasant species are typically associated with forests, this finding may indicate a preference for transitional zones, forest edges, or mosaic landscapes that include agricultural elements [51,52].
Despite previous documentation (Table 1), several targeted species including the Asian elephant, dhole, northern pig-tailed macaque, banteng, Malayan sun bear, and Asiatic black bear were not detected during this survey. However, signs such as footprints, scratches, and dung confirmed their presence. Their absence from direct sightings may be attributed to behavioral factors (e.g., nocturnal vs. diurnal activity), habitat types, and sensitivity to human disturbance. These species are often elusive and difficult to detect through line transect sampling, especially in dense vegetation. Elephants, for example, are known to be elusive and sensitive to human presence, particularly in areas with hunting pressure [53]. The dhole is considered one of the most obscure apex carnivores in Asia’s tropical forests [54]. Similarly, Malayan sun bears and Asiatic black bears exhibit elusive behaviors and are rarely observed directly, though they leave clear signs such as claw marks on trees [55].
This study, which employed transect surveys as a distance sampling method, represents the second biodiversity monitoring campaign in PLWS following the systematic camera trap survey conducted in 2021, which reported the presence of 54 species [8]. In our study, we selected 15 key fauna species to monitor wildlife biodiversity distribution and assess the impact of the REDD+ program in the sanctuary. Most species occurrences and individual detections were mainly found in high-intact forests. However, we were unable to estimate species density and population across the entire sanctuary. Future research should aim to address this gap, particularly for species detected above the threshold (≥60–80 detections), such as the great hornbill, to better assess the impact of forest conversion under the REDD+ project [56].
5. Implication and Conclusions
As the PLWS has been considered as one of the last and largest areas of lowland tropical forests left in mainland Southeast Asia, protecting the forest as the habitat for wildlife biodiversity is crucial. There have been conservation efforts operated in this area, such as law enforcement, community engagement, livelihood improvement, environmental safeguards, and biodiversity monitoring. Despite these significant protection efforts, ongoing threats continue to cause wildlife biodiversity loss due to wildlife trade, agricultural expansion, and especially habitat loss [57]. In our survey, we encountered hunting, habitat fragmentation, agricultural land clearance, and illegal logging, in particular along the edges in the northern and southern parts of the PLWS. These threats inevitably affect the distribution and population of the key wildlife biodiversity. Implementing REDD+ program is one of the crucial approaches in protecting habitats and wildlife biodiversity because it reduces deforestation and forest degradation. Protecting intact forests via REDD+ implementation means providing homes to a significant portion of the world's terrestrial biodiversity and protecting key wildlife species in particular ecosystems. One notable example of REDD+ conserving wildlife species is the Orangutan Conservation Project in Indonesia, which focuses on protecting the habitat of the critically endangered orangutans in the forests of Borneo and Sumatra by preventing deforestation and promoting sustainable forest management [58].
The findings of our study support the previous positive impact of REDD+ on wildlife conservation. We found a higher number of species and their abundance in the high-intact forest habitats of the REDD+ project in PLWS. This strategy serves as one of the sustainable conservation approaches for Cambodian wildlife biodiversity. Without significant enhancements in conservation management, key species in PLWS, like the endangered pileated gibbon, Indochinese silvered langur, banteng, etc., may follow the fate of kouprey, tiger, and wild water buffalo, leading to their complete absence from the sanctuary. The ecological ramifications of such a loss are profound because their absence not only disrupts the ecological equilibrium but also adversely affects the indigenous communities whose subsistence and genetic resources are inextricably linked to these forest resources. Therefore, effective and adaptive management of the sanctuary should be in place by incorporating wildlife monitoring, conservation planning, and sustainable carbon financing (e.g., via REDD+). The suggestion from our study is that conservation efforts need prioritize strict protection and restoration of core forest areas for species highly dependent on intact forests (e.g. great hornbill, pileated gibbon and silver langur). For more adaptable species, management strategies might focus on mitigating human-wildlife conflict and promoting coexistence in mosaic landscapes, while still ensuring the ecological integrity of their habitats. Effective conservation efforts and adaptive management of the sanctuary (both the high- and moderate-intact forest, and the edge/mosaic areas) will enhance biodiversity and maintain the system integrity, functions, and services (e.g., climate regulation, water and air purification, nutrient cycling, carbon sequestration, and erosion and flood control) [59] and consequently contribute to human-nature harmonization, employment, and socioeconomics of the people sharing this important protected area, the Prey Lang Wildlife Sanctuary.
Author Contributions
Kimnannara Khiev: Drafted, Reviewed and edited the manuscript, Visualization, Project administration, Methodology, Investigation, Formal analysis and result interpretation, Data curation. Ratha Sor: Conceptualization, Drafted, Reviewed and edited the manuscript, Visualization, Formal analysis and result interpretation, Data curation. Vanna Chea: Project administration, Investigation & Data entry, Data curation. Sophak Sett: Project administration, Investigation & Funding acquisition. Jackson Frechette: Research design, Reviewed and edited the manuscript, Funding acquisition.Naven Hon: Project administration, Research design, Reviewed and edited the manuscript, Data curation, Funding acquisition.
Funding
This research was funded by the United States Agency for International Development (USAID) Cambodia under the Greening Prey Lang (GPL) initiative, Mitsui & Co., Ltd., and Conservation International Cambodia (CI Cambodia). The APC was not funded.
Institutional Review Board Statement
Ethical review and approval were waived for this study because it involved non-invasive observational fieldwork using distance sampling (line transect methods), which caused minimal disturbance to wildlife. The study was conducted with permission from the Cambodian Ministry of Environment.
Acknowledgments
We extend our deepest gratitude to the individuals and organizations whose invaluable contributions made this line transect survey possible. This research was made possible through the support of USAID/Cambodia under the Greening Prey Lang (GPL) initiative, Mitsui & Co. Ltd., Conservation International Cambodia (CI Cambodia), the Royal Government of Cambodia’s Ministry of Environment (MoE), and the Provincial Ministries of Environment (PDoE). We also gratefully acknowledge the support and guidance of Mr. Nathan Conaboy, Cambodia Conservation lead at CI Cambodia. Our sincere thanks go to the Conservation Technology team (Socheat Kong, Samnang Hean, Veasna Mean and Vises Ung) for their exceptional work in creating the maps featured in this paper. We particularly appreciate the dynamic participation of all individuals who dedicated numerous days to fieldwork in the forest.
Conflicts of Interest
The authors declare that there are no financial conflicts of interest or personal relationships that could have influenced this work.
Abbreviations
The following abbreviations are used in this manuscript:
AIC – Akaike’s Information Criterion
CI – Conservation International
CRL – Cropland
DDF – Dry dipterocarp forest
DNV – Distance to Nearest Village
EVE – Evergreen Forest
GPS – Global Positioning System
MoE – Ministry of Environment
OTP – Other Plantations
PD – Perpendicular distance
PLWS – Prey Lang Wildlife Sanctuary
PDoE – Provincial Ministries of Environment
RIF – Rice Field
USAID – United States Agency for International Development
Appendix A
Figure A1.
Distribution pattern of the four least detected species (less than 20% of total transects): (a) green peafowl, (b) northern red muntjac, (c) Indochinese silvered langur, (d) guar, and (e) sambar deer.
Figure A1.
Distribution pattern of the four least detected species (less than 20% of total transects): (a) green peafowl, (b) northern red muntjac, (c) Indochinese silvered langur, (d) guar, and (e) sambar deer.

References
- Dirzo R, Young HS, Galetti M, Ceballos G, Isaac NJB, Collen B. Defaunation in the Anthropocene. Science (1979). 2014;345(6195):401–6.
- Yarrow G, others. Habitat requirements of wildlife: food, water, cover and space [Internet]. 2009. Available from: https://www.academia.edu/121106097/Habitat_Requirements_of_Wildlife_Food_Water_Cover_and_Space.
- Leimgruber P, Gagnon JB, Wemmer C, Kelly DS, Songer MA, Selig ER. Fragmentation of Asia’s remaining wildlands: implications for Asian elephant conservation. Anim Conserv. 2003;6(4):347–59.
- Duckworth JW, Steinmetz R, Timmins RJ, Pattanavibool A, Than Zaw DT, Hedges S. Bos gaurus. The IUCN Red List of Threatened Species 2008: e. T2891A9491805. 2008.
- Datta A, Rawat GS. Nest-site selection and nesting success of three hornbill species in Arunachal Pradesh, north-east India: Great Hornbill Buceros bicornis, Wreathed Hornbill Aceros undulatus and Oriental Pied Hornbill Anthracoceros albirostris. Bird Conserv Int. 2004;14(S1):S39–S52.
- Pin C, Ngoprasert D, Gray TNE, Savini T, Crouthers R, Gale GA. Utilization of waterholes by globally threatened species in deciduous dipterocarp forest of the Eastern Plains Landscape of Cambodia. Oryx. 2020;54(4):572–82.
- Theilade I, Phourin C, Schmidt L, Meilby H, van de Bult M, Friborg KG. Evergreen forest types of the central plains in Cambodia: floristic composition and ecological characteristics. Nord J Bot. 2022;2022(8):e03494.
- Auda E. Prey Lang Extended Landscape Camera Trap Survey. 2023.
- Olsson A, Emmett DE. A Floral and Faunal Biodiversity Assessment of Prey Long. 2007. Report.
- Gray TNE, Ou R, Huy K, Pin C, Maxwell AL. The status of large mammals in eastern Cambodia: a review of. Cambodian Journal of Natural History. 2012;42.
- Gray TNE, Chamnan H, Borey R, Collar NJ, Dolman PM. Habitat preferences of a globally threatened bustard provide support for community-based conservation in Cambodia. Biol Conserv. 2007;138(3–4):341–50.
- Gray TNE, Phan C. Habitat preferences and activity patterns of the larger mammal community in Phnom Prich Wildlife Sanctuary, Cambodia. Raffles Bull Zool. 2011;59(2):311–8.
- Theilade I, Schmidt L. REDD+ and Conservation of Prey Long Forest, Cambodia Summary of Scientific Findings 2007-2010 [Internet]. Forest & Landscape Working Papers no 66; 2011 [cited 2025 Feb 16]. Report. Available from: www.SL.life.ku.dk.
- SERVIR-Mekong. Biophysical M&E Dashboard Map Viewer [Internet]. 2025 [cited 2025 May 7]. Available from: https://cambodia-me-servir.adpc.net/mapviewer.
- Traeholt C, Bunthoen R, Rawson B, Samuth M, Vrak C, Vuthin S. Status Review of Pileated Gibbon, Hylobates Pileatus and Yellow-cheeked Crested Gibbon, Nomascus Gabriellae: In Cambodia. Fauna & Flora International Cambodia; 2005.
- Srivathsa A, Karanth KK, Jathanna D, Kumar NS, Karanth KU. On a dhole trail: Examining ecological and anthropogenic correlates of dhole habitat occupancy in the Western Ghats of India. PLoS One. 2014 Jun 3;9(6). https://doi.org/10.1371/journal.pone.0098803 PubMed PMID: 24893166.
- Ruikar S, Suryavanshi N, Nanaware J, Balasundaram H, Patil D, Pathade G. The Role of Wildlife in Ecosystem Health: Interactions and Impacts. REDVET-Revista electrónica de Veterinaria. 2024;25(1):275–95.
- Ong L, Tan WH, Davenport LC, McConkey KR, Mat Amin MKA bin, Campos-Arceiz A, et al. Asian elephants as ecological filters in Sundaic forests. Frontiers in Forests and Global Change. 2023;6. https://doi.org/10.3389/ffgc.2023.1143633.
- Hayes B, Khou EH, Neang T, Furey N, Chhin S, Holden J, et al. Biodiversity Assessment of Prey Lang–Kratie, Kampong Thom, Stung Treng and Preah Vihear Provinces. Conservation International, Winrock International, USAID, Phnom Penh, Cambodia. 2015.
- Chaichanathong S, Klinsawat W, Sukmak M, Sakulthai A, Wajjwalku W, Sripiboon S, et al. Genetic characterization of banteng (Bos javanicus) populations in Thailand for conservation. The Thai Journal of Veterinary Medicine. 2021 Dec 1;51(4):647–54. https://doi.org/10.56808/2985-1130.3162.
- Prayoon U, Suksavate W, Chaiyes A, Winitpornsawan S, Tunhikorn S, Angkaew C, et al. Importance of protected areas in conservation and recovery of gaur (Bos gaurus) in Thailand. 2022. Report.
- Gazagne E, Pitance JL, Savini T, Huynen MC, Poncin P, Brotcorne F, et al. Seed shadows of northern pigtailed macaques within a degraded forest fragment, Thailand. Forests. 2020 Nov 1;11(11):1–24. https://doi.org/10.3390/f11111184.
- Bunthoeun R, Rawson B, Samuth M, Virak C, Vuthin S. Status review of pileated gibbon, Hylobates pileatus and yellow-cheeked crested gibbon, Nomascus gabriellae, in Cambodia. 2005. Report.
- Hoang MD, Van BT, Le TH, Covert HH. Diet of Indochinese Silvered Langur (Trachypithecus germaini) in Kien Luong Karst Area, Vienam [Internet]. 2017. Report. Available from: https://www.researchgate.net/publication/282652802.
- Bernathirdin ANH, Wijayanto H, Kusindarta DL, Budipitojo T. Population Estimation and Identification of Long-Tailed Macaque (Macaca fascicularis) Group at Tlogo Muncar Tourism Area, Mount Merapi National Park, after Two Years Closure Caused by the Covid-19 Pandemic. In: IOP Conference Series: Earth and Environmental Science. Institute of Physics; 2023. https://doi.org/10.1088/1755-1315/1174/1/012028.
- Cuevas MF, Ojeda RA, Jaksic FM. Ecological strategies and impact of wild boar in phytogeographic provinces of Argentina with emphasis on aridlands. Mastozool Neotrop [Internet]. 2016;23(2):239–54. Available from: http://www.sarem.org.ar.
- Liang H, McConkey KR, Lim JY. From beak to fruit: An Asian hornbill database for frugivory and seed dispersal research. Glob Ecol Conserv. 2024 Jun 1;51. https://doi.org/10.1016/j.gecco.2024.e02879.
- HERNOWO JB, MARDIASTUTI ANI, ALIKODRA HS, KUSMANA CECEP. Behavior Ecology of the Javan Green Peafowl (Pavo muticus muticus Linnaeus 1758) in Baluran and Alas Purwo National Park, East Java. Hayati. 2011 Dec 1;18(4):164–76. https://doi.org/10.4308/hjb.18.4.164.
- Scotson L, Fredriksson G, Augeri D, Cheah C, Ngoprasert D, Wai- Ming W. IUCN Sun bear Helarctos malayaus Red list Assessment 2016 [Internet]. 2017 [cited 2025 Feb 6]. Available from: https://www.researchgate.net/publication/321682540.
- Steinmetz R, Garshelis DL, Chutipong W, Seuaturien N. Foraging ecology and coexistence of Asiatic black bears and sun bears in a seasonal tropical forest in Southeast Asia. J Mammal. 2013;94(1):1–18.
- Griffin O, Nuttall M. Status of Key Species in Keo Seima Wildlife Sanctuary 2010-2020 [Internet]. 2020 [cited 2025 Feb 8]. Report. Available from: https://doi.org/10.19121/2020.Report.38511.
- Nuñez-Penichet C, Cobos ME, Soberón J, Gueta T, Barve N, Barve V, et al. Selection of sampling sites for biodiversity inventory: Effects of environmental and geographical considerations. Methods Ecol Evol. 2022 Jul 1;13(7):1595–607. https://doi.org/10.1111/2041-210X.13869.
- Buckland ST, Rexstad EA, Marques TA, Oedekoven CS. Methods in Statistical Ecology Distance Sampling: Methods and Applications [Internet]. 2015. Report. Available from: http://www.springer.com/series/10235.
- Pavlacky DC, Green AW, George TL, Iovanna R, Bartuszevige AM, Correll MD, et al. Landscape-scale conservation mitigates the biodiversity loss of grassland birds. Ecological Applications. 2021. https://doi.org/10.5061/dryad.9zw3r22f3.
- Mittermeier RA, Mittermeier CG, Brooks TM, Pilgrim JD, Konstant WR, Da Fonseca GAB, et al. Wilderness and biodiversity conservation [Internet]. 2003. Report. Available from: www.pnas.org.
- Pramod AF, Kumara V, HT RG. Encounter rate of large mammals in Thanigebyle range of Bhadra wildlife sanctuary, Karnataka. Int Multidiscip Res J. 2012;2(9).
- Harrell FE. Regression Modeling Strategies: With Applications to Linear Models, Logistic and Ordinal Regression, and Survival Analysis [Internet]. 2nd ed. Springer; 2015. Available from: https://link.springer.com/book/10.1007/978-3-319-19425-7.
- Sor R, Park YS, Boets P, Goethals PLM, Lek S. Effects of species prevalence on the performance of predictive models. Ecol Modell. 2017;354:11–9.
- Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM, others. Mixed effects models and extensions in ecology with R. Vol. 574. Springer; 2009.
- Gibson L, Lee TM, Koh LP, Brook BW, Gardner TA, Barlow J, et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature. 2011;478(7369):378–81.
- Ghadiri Khanaposhtani M, Kaboli M, Karami M, Etemad V. Effect of habitat complexity on richness, abundance and distributional pattern of forest birds. Environ Manage. 2012;50(2):296–303.
- Spitzer HB, Meagher RK, Proudfoot KL. The impact of providing hiding spaces to farmed animals: A scoping review. PLoS One. 2022;17(11):e0277665.
- Miraldo A, Li S, Borregaard MK, Flórez-Rodríguez A, Gopalakrishnan S, Rizvanovic M, et al. An Anthropocene map of genetic diversity. Science (1979). 2016;353(6307):1532–5.
- Datta A. Hornbill abundance in unlogged forest, selectively logged forest and a forest plantation in Arunachal Pradesh, India. Oryx. 1998;32(4):285–94.
- Phoonjampa R, Koenig A, Brockelman WY, Borries C, Gale GA, Carroll JP, et al. Pileated gibbon density in relation to habitat characteristics and post-logging forest recovery. Biotropica. 2011;43(5):619–27.
- Fahrig L. Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol Syst. 2003;34(1):487–515.
- Gibbons MA, Harcourt AH. Biological correlates of extinction and persistence of primates in small forest fragments: a global analysis. Trop Conserv Sci. 2009;2(4):388–403.
- Sharma CM, Bhattacharjee S, Sharief A, Joshi BD, Thakur M, Sharma LK. A tapestry of habitats: exploring abundance and habitat preferences of the Northern Red Muntjac (Muntiacus vaginalis) across the Central Himalayan landscape. Mammalia. 2024;88(4):280–91.
- Brotcorne F, Maslarov C, Wandia IN, Fuentes A, Beudels-Jamar RC, Huynen MC. The role of anthropic, ecological, and social factors in sleeping site choice by long-tailed macaques (Macaca fascicularis). Am J Primatol. 2014;76(12):1140–50.
- Linkie M, Dinata Y, Nofrianto A, Leader-Williams N. Patterns and perceptions of wildlife crop raiding in and around Kerinci Seblat National Park, Sumatra. Anim Conserv. 2007;10(1):127–35.
- Delacour J. The pheasants of the world. (No Title). 1977.
- Saridnirun G, Meckvichai W, Dumrongrojwatthana P. Seasonal Distribution and Habitat Use of the Green Peafowl Pavo muticus, Linnaeus 1766, in Wiang Lor Wildlife Sanctuary, Phayao Province, Northern Thailand. Chiang Mai J Sci. 2017;44:824–38.
- Blake S, Hedges S. Sinking the flagship: the case of forest elephants in Asia and Africa. Conservation Biology. 2004;18(5):1191–202.
- Kamler JF. Extreme dogs: dholes [Internet]. 2013 [cited 2025 Apr 10]. Available from: https://www.researchgate.net/publication/277588881.
- Steinmetz R, Garshelis DL. Distinguishing Asiatic black bears and sun bears by claw marks on climbed trees. J Wildl Manage. 2008;72(3):814–21.
- Buckland ST, Rexstad EA, Marques TA, Oedekoven CS, others. Distance sampling: methods and applications. Vol. 431. Springer; 2015.
- Theilade I, Schmidt L. REDD+ and Conservation of Prey Long Forest, Cambodia. Forest & Landscape Denmark; 2011.
- Voet J. Conservation of Orangutans: reintroduction or habitat preservation? 2020.
- Svensson J, Bubnicki JW, Jonsson BG, Andersson J, Mikusiński G. Conservation significance of intact forest landscapes in the Scandinavian Mountains Green Belt. Landsc Ecol. 2020;35(9):2113–31.
Figure 1.
Map of Prey Lang Wildlife Sanctuary and the selected quadrat-transect lines (squares) inside the REDD+ area, with high-intact forest indicated in blue and moderate-intact forest in yellow.
Figure 1.
Map of Prey Lang Wildlife Sanctuary and the selected quadrat-transect lines (squares) inside the REDD+ area, with high-intact forest indicated in blue and moderate-intact forest in yellow.

Figure 2.
The distance sampling design. (a) The observers walk along the transect line following the walking direction, and (b) the opposite walking direction between the morning (blue short-dashed line) and the afternoon (orange long-dashed line) that begins and ends at the starting point [33].
Figure 2.
The distance sampling design. (a) The observers walk along the transect line following the walking direction, and (b) the opposite walking direction between the morning (blue short-dashed line) and the afternoon (orange long-dashed line) that begins and ends at the starting point [33].

Figure 3.
The distribution pattern of species richness (a) and abundance (b) of the target species recorded from the transect survey.
Figure 3.
The distribution pattern of species richness (a) and abundance (b) of the target species recorded from the transect survey.

Figure 4.
Abundance distribution pattern of the four most detected species: (a) great hornbill, (b) pileated gibbon, (c) wild pig, and (d) long-tailed macaque.
Figure 4.
Abundance distribution pattern of the four most detected species: (a) great hornbill, (b) pileated gibbon, (c) wild pig, and (d) long-tailed macaque.

Figure 5.
Box plots of species richness (counts), abundance, and diversity H between the high- and moderate-intact forest habitats.
Figure 5.
Box plots of species richness (counts), abundance, and diversity H between the high- and moderate-intact forest habitats.

Figure 6.
Results of habitat preference analysis. The significant association between the species presence-absence and the two types of forest habitats: high-intact and moderate-intact (a-g), and the significant differences in the proportion of occurrence frequency of the species between the two types of forest habitats (h).
Figure 6.
Results of habitat preference analysis. The significant association between the species presence-absence and the two types of forest habitats: high-intact and moderate-intact (a-g), and the significant differences in the proportion of occurrence frequency of the species between the two types of forest habitats (h).


Table 1.
Selected species for key biodiversity monitoring at PLWS, their IUCN status, ecological roles, and previous record in PLWS.
Table 1.
Selected species for key biodiversity monitoring at PLWS, their IUCN status, ecological roles, and previous record in PLWS.
| Common name | Scientific name | IUCN Status | Ecological roles | Previous records |
| Asian elephant | Elephas maximus | EN | Seed dispersal, habitat creation, and vegetation modification [18] | [8,9,19] |
| Banteng | Bos javanicus | EN | Seed dispersal [20] | [8,9,19] |
| Gaur | Bos gaurus | VU | Plant population control and main prey for large carnivores [21] | [9,19] |
| Dhole | Cuon alpinus | EN | Prey regulation [16] | [9,19] |
| Sambar deer | Rusa unicolor | VU | Seed dispersal and keystone herbivore (Quin, Morgan, & Murphy, 2023) | [8,9,19] |
| Northern pig-tailed macaque | Macaca leonina | VU | Seed dispersal and forest regeneration [22] | [9,19] |
| Pileated gibbon | Hylobates pileatus | EN | Seed dispersal [23] | [9,19] |
| Indochinese silvered langur | Trachypithecus germaini | EN | Regulate vegetation growth and forest structure [24] | [9,19] |
| Long-tailed macaque | Macaca fascicularis | VU | Seed dispersal and insect control through predation [25] | [8,9,19] |
| Northern red muntjac | Muntiacus vaginalis | LC | Herbivory, seed dispersal, and serving as prey species [50] | [8,9,19] |
| Wild pig | Sus scrofa | LC | Soil disturbance, seed dispersal, and maintaining ecological balance [26] | [8,9,19] |
| Great hornbill | Buceros bicornis | VU | Seed dispersal, forest regeneration, and ecological balance [27] | [9,19] |
| Green peafowl | Pavo muticus | EN | Feeding on plants and dispersing seeds [28] | [8,9,19] |
| Malayan sun bear | Helarctos malayanus | VU | Seed dispersal and maintenance of forest understory structure [29] | [8,9,19] |
| Asiatic black bear | Ursus thibetanus | VU | Seed dispersal and insect control [30] | [8,9,19] |
Table 2.
The detected species and their numbers of sighting, average cluster size, and encounter rate.
Table 2.
The detected species and their numbers of sighting, average cluster size, and encounter rate.
| No |
Common name |
Scientific name |
# Detections/ sightings |
Cluster Size (average) |
Encounter rate |
| 1 | Great hornbill | B. bicornis | 76 | 2.34 | 0.826 |
| 2 | Pileated gibbon | H. pileatus | 31 | 2.23 | 0.337 |
| 3 | Wild pig | S. scrofa | 25 | 2.88 | 0.272 |
| 4 | Long tailed macaque | M. fascicularis | 23 | 5.96 | 0.250 |
| 5 | Green peafowl | P. muticus | 13 | 1.62 | 0.141 |
| 6 | Northern red muntjac | M.vaginalis | 10 | 1 | 0.109 |
| 7 | Indochinese silvered langur | T. germaini | 8 | 8.13 | 0.087 |
| 8 | Gaur | B. gaurus | 1 | 1 | 0.011 |
| 9 | Sambar deer | R. unicolor | 1 | 1 | 0.011 |
Table 3.
The variables influencing the presence/absence of wildlife species based on results of multiple logistic regression analysis indicate selected variables (from most to least important) retained in two models based on the minimum AIC value.
Table 3.
The variables influencing the presence/absence of wildlife species based on results of multiple logistic regression analysis indicate selected variables (from most to least important) retained in two models based on the minimum AIC value.
| Wildlife Species | Important variables selected for the final model | AIC | ||||||
| EVE | DDF | OTP | CRL | RIF | DNV | Full model | Final model | |
| Great hornbill | 1.54 | - | - | - | - | - | 92.98 | 85.89 |
| Pileated gibbon | 0.69 | - | - | - | - | - | 96.74 | 87.64 |
| Indochinese silvered langur | - | - | - | - | -506.7 | - | 1103.3 | 49.62 |
| Northern red muntjac | -2.67 | - | 5.23 | - | - | -3.6 | 22 | 20.94 |
| Green peafowl | - | -798.56 | - | 30.22 | - | - | 48.41 | 34.66 |
| Long-tailed macaque | - | -1.01 | 0.46 | - | - | - | 91.42 | 80.32 |
| Wild pig | - | - | - | - | - | - | 107.7 | - |
Note: EVE = Evergreen Forest; DDF = Dry Dipterocarp Forest; OTP = Other Plantations; CRL = Cropland, RIF = Rice Field, DNV = Distance to Nearest Village; (+) indicates a positive association; and (–) indicates a negative association.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



