Submitted:
30 March 2026
Posted:
31 March 2026
You are already at the latest version
Abstract
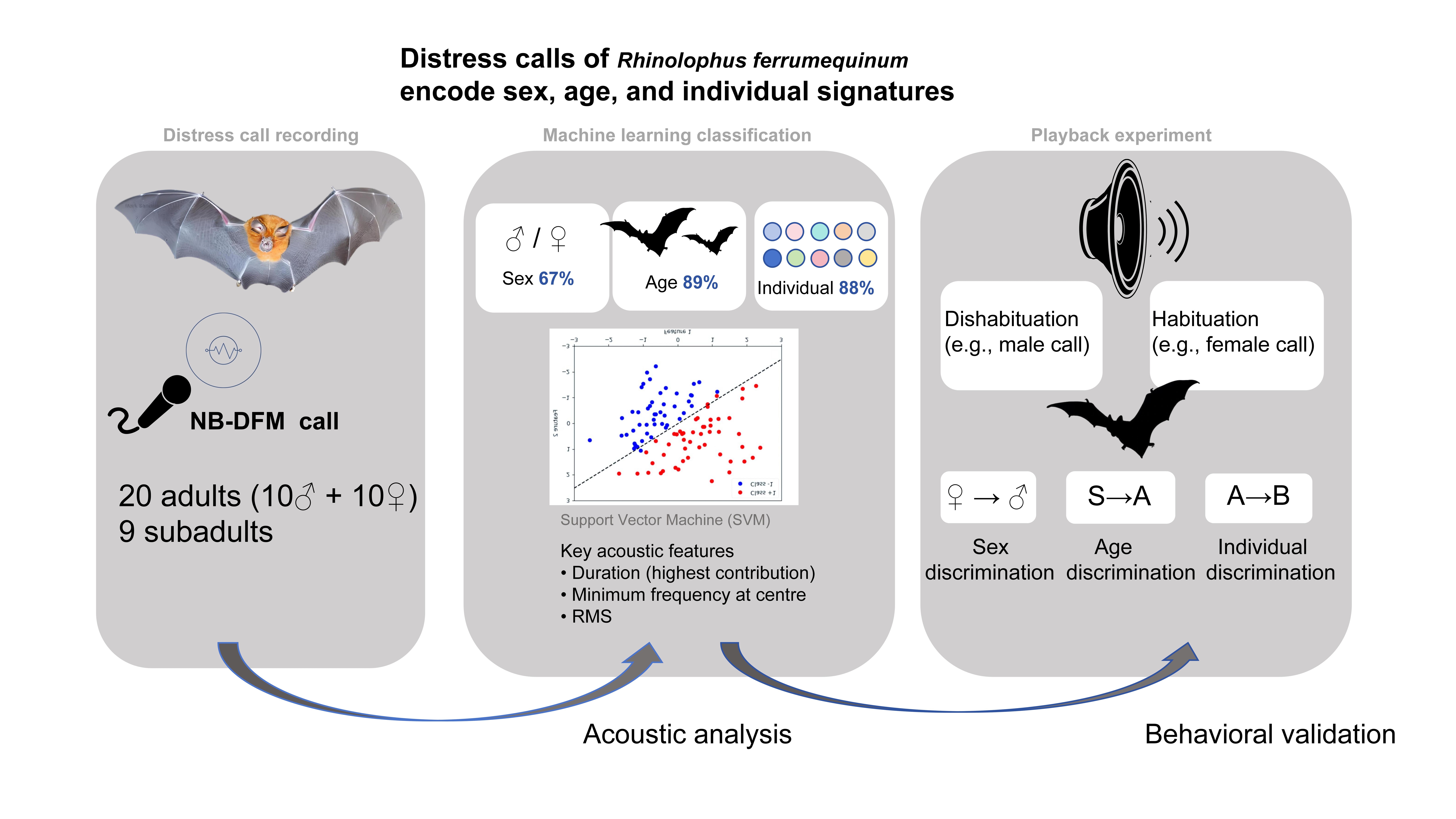
Vocal signals are the primary medium of intraspecific communication in bats, yet the encoding features and recognition mechanisms of distress calls remain largely unclear. This study aimed to examine sex, age, and individual signatures in noise burst to downward frequency modulation (NB-DFM) distress calls of the Rhinolophus ferrumequinum and verify conspecific recognition ability. We recorded NB-DFM calls from 20 adult and 9 subadult bats in Jilin Province, extracted 18 acoustic parameters, built classification models with support vector machines (SVMs), evaluated feature importance using random forest, and performed habituation-dishabituation playback experiments. SVM yielded classification accuracies of 67%, 89%, and 88% for sex, age, and individual identity, respectively, all significantly above chance levels. Call duration, central minimum frequency, and root mean square (RMS) were the most diagnostic parameters, and key acoustic variables differed significantly among classification levels. Playback tests elicited distinct behavioral responses to calls of different sexes, ages, and individuals, confirming discrimination ability. These findings reveal individual, age, and sex specific acoustic markers in bat distress calls, support the utility of machine learning for bioacoustic classification, and provide empirical insight into stress related acoustic communication in nocturnal mammals.
Keywords:
Rhinolophus ferrumequinum
; distress calls
; machine learning
; Support Vector Machine (SVM)
; acoustic identification
; playback experiment
1. Introduction
Acoustic signals represent a crucial behavioral phenotype in group-living vocal animals, playing a key role in individual survival, reproduction, and the maintenance of group stability [1]. Studies have shown that acoustic signals can encode multiple types of information, including emotional states, environmental cues, body condition, and individual signatures. When environmental constraints—such as darkness—preclude animals from obtaining visual information, individual signatures encoded in acoustic signals become crucial for conspecific recognition [2]. Birds frequently employ song to deter rivals and attract mates [3,4]. Although vocal repertoires vary significantly across species, songbirds utilize acoustic signatures for individual recognition, which in turn elicits context-specific behavioral responses. A classic example is a male exhibiting territorial defense upon detecting the song of a conspecific intruder from a different social group [5]. Social vocalizations are among the most prevalent acoustic signals in mammals. Numerous species generate context-specific vocal repertoires that vary according to their social system [6]. Research indicates that social vocalizations often carry acoustic signatures [7]. For instance, Microcebus murinus can recognize the trill calls produced by males and use this information to decide whether to mate with them [8]; Vocalizations produced by Hylobates agilis serve in territory defense and mate choice [9,10].
As nocturnal mammals, bats typically form colonies ranging in size from dozens to tens of thousands. Their acoustic repertoire includes echolocation signals used for navigation and foraging, as well as communication signals employed in both intraspecific and interspecific social contexts [11]. The social calls of bats comprise a variety of syllable types, including pure tones, frequency modulations (FM) of varying degrees, noise bursts (NB), and composite signals that integrate these fundamental acoustic elements [11]. Owing to their structural complexity, these calls are capable of encoding information such as sex, age, and individual identity [12,13]. Bat social calls are utilized in diverse behavioral contexts, including courtship and mating, cooperative foraging, territorial defense, anti-predator cooperation, and mother-offspring recognition [14]. Nearly all bats produce isolation calls during the pup stage, which emerge within minutes after birth. When separated from their mothers, bat pups frequently emit individually distinctive isolation calls, allowing mothers to identify their offspring among hundreds of pups and facilitate mother-pup reunions [15]. Research indicates that male Saccopteryx bilineata produce vocalizations with distinct syllable compositions within their territories to attract females [16]. Studies on female Phyllostomus hastatus reveal that they recognize the unique screech calls of conspecific females, thereby forming foraging organizations that facilitate social cooperation within the community [17].
The social calls of bats are complex and diverse across different behavioral contexts, comprising multiple distinct syllable types. For example, the R. ferrumequinum produces social calls that include 23 simple and 20 compound syllables. These syllables can be combined into various phrases and sequences. Both the specific syllables emitted and their combinatorial patterns vary with behavioral context [18,19]. Distress contexts represent an important behavioral context in the life history of bats, referring primarily to situations in which bats are under predation threat. For bats that fly and forage in darkness, timely distress or alarm calls upon encountering predators (e.g., birds of prey) can increase escape opportunities and enhance survival. Animals emit distress calls when their safety is threatened, such as during predator attacks or when handled by researchers. Distress calls may serve functions including warning companions, signaling for help from group members, and intimidating predators. They call play a vital role in survival and communication, making them an ideal model system for the study of acoustic recognition [20,21,22,23]. Additionally, the distress calls emitted by bats can also encode information about an individual's emotional state, allowing other members of the population to assess their own risk of predation by predators [24,25]. When bats encounter predators, they also emit distress calls to summon companions, cooperating to resist capture—a behavior analogous to the mobbing defense exhibited by birds [26,27]. Distress calls typically consist of a series of high-intensity, multi-harmonic syllables with lower frequencies, wider syllable bandwidths, and noise components [20,28]. However, whether bats exhibit sex, age, and individual characteristics in their distress calls—and whether these can be identified through acoustic features—remains to be verified through playback experiments.
In previous experiments identifying bat acoustic characteristics, traditional call classification methods were commonly used to determine sex, age, or individual traits. However, for communication sounds—which exhibit diverse types and complex spectrograms—traditional methods are time-consuming and ineffective for accurate differentiation. Machine learning approaches enable rapid and precise categorization of animal acoustic signals by establishing analytical programs. Machine learning employs statistical methods and techniques to enable computers to learn from data [29]. Leveraging its speed and accuracy in classifying and predicting large datasets, machine learning methods are widely applied across numerous fields [30], including species identification and call classification in animal acoustics research. For example, when classifying five frog species based on their calls, researchers achieved relatively accurate results using support vector machines (SVMs), a machine learning method [31]. In vocal recognition tasks for other taxa such as birds, cetaceans, and bats, machine learning methods including SVM have also been widely used [32,33,34,35].
R. ferrumequinum live in year-round colonies, with most roosts housing hundreds of individuals. Consequently, they emit a large volume of social calls to communicate with one another, making them an ideal bat species for acoustic recognition research. The vocalizations of R. ferrumequinum are more diverse than those of other birds, primates, and mammals, featuring greater structural complexity and a larger number of vocalizations. Therefore, this chapter focuses on R. ferrumequinum as the research subject. Machine learning methods are employed to determine whether the social calls of R. ferrumequinum exhibit sex, age, and individual characteristics under distress contexts. Subsequently, through sound playback experiments, we aim to verify whether R. ferrumequinum can discriminate among individuals based on social call features.
2. Materials and Methods
2.1. Bat Sample Collection and Acoustic Recording
From September to October 2023, 20 adult R. ferrumequinums (10 males, 10 females) and 9 subadult individuals (subadults distinguished from adults based on fur color and the degree of epiphyseal fusion of the metacarpals) were captured using mist nets at Dalazi Cave in Ji'an Village, Jilin Province, bats were captured and placed in breathable cloth bags for transport to the field laboratory for playback experiments. All procedures are in accordance with the regulations of Wildlife Conservation of the People's Republic of China (Chairman Decree (2016) No. 47). All animal experimental procedures were approved by the Forestry Bureau of Jilin Province, China (approval number: [2006] 178). Upon completion of the experiments, all bats were released back into their habitats.
Distress calls of R. ferrumequinum were recorded using an UltrasoundGate 116 unit (Avisoft Bioacoustics, Berlin, Germany) connected to a laptop. A condenser microphone was mounted on a tripod and positioned directly facing the bat's head at a distance of 1 meter. The microphone had a flat frequency response from 10 Hz to 200 kHz (±3 dB). Each bat was held manually by an experimenter to simulate predation threat, and gentle stimulation was applied to its back to elicit vocalizations [21]. Recordings were saved as 60 second files, with a total recording duration of 5 minutes per individual, at a sampling rate of 250 kHz with a 16 bits per sample.
2.2. Feature Selection for Machine Learning Models
Based on the classification of social call syllable types in R. ferrumequinum by Ying Liu, Kangkang Zhang, et al. [18,36], the social call recordings from Section 2.1 were categorized and counted by syllable type. The results revealed that noise burst to downward-frequency modulation (NB-DFM) syllables were emitted most frequently (Figure 1). Subsequent analyses and playback experiments were therefore conducted using NB-DFM syllables (Figure 2). All NB-DFM vocalizations recorded under distress contexts were included for machine learning classification model. A temporal parameter (duration, unit: seconds, s) and four spectral parameters (unit: kilohertz, kHz) were measured from the harmonic with the maximum energy: dominant frequency, minimum frequency, maximum frequency, and bandwidth. These parameters were commonly used in bat vocalization analysis to determine whether they possessed individual acoustic signatures [31,37,38]. For these four spectral parameters, measurements were taken at four specific points: the start, center, end, and point of maximum amplitude [39,40,41]. Consequently, the following 18 features were extracted for the machine learning classification model: Duration (seconds), Root-Mean-Square (RMS), Peak frequency in start position, Minimum frequency in start position, Maximum frequency in start position, Bandwidth in start position, Peak frequency at end position, Minimum frequency in end position, Maximum frequency in end position, Bandwidth in end position, Peak frequency in center position, Minimum frequency in center position, Maximum frequency in center position, Bandwidth in center position, Peak frequency in maximum amplitude position, Maximum frequency in maximum amplitude position, Minimum frequency in maximum amplitude position, Bandwidth in maximum amplitude position .
2.3. Machine Learning Model Selection
SVM is a precise class of kernel-based machine learning algorithms used for nonlinear classification. SVM makes no assumptions about the distribution of the original data, imposing minimal distributional prerequisites and thus offering high applicability. Furthermore, due to their combination of effectiveness and accuracy, SVMs are widely applied across various fields. In essence, a support vector machine is a classifier that, given two labeled sets of vectors, finds an optimal separating hyperplane to divide these vectors into two groups. This hyperplane maximizes the distance between the closest vectors from each group (known as support vectors) and the hyperplane itself. In animal acoustic research, support vector machines are primarily applied to acoustic classification and species identification tasks [42,43]. This study utilized Scikit-learn for model construction and analysis. Scikit-learn is a Python module that integrates various classical machine learning algorithms [44]. All classification algorithms in this chapter are implemented using Scikit-learn (version 1.0.2), specifically the method class sklearn.svm.SVC with the linear kernel function kernel = 'linear' and the hyperparameter C = 1.0.
2.4. Machine Learning Model Training and Evaluation
70% of the entire sound file dataset was randomly selected for training, with the remaining 30% reserved for model evaluation. The data were standardized to facilitate weight learning by the machine learning model. Model parameters were adjusted using cross-validation to prevent overfitting or underfitting. The performance of each machine learning model was evaluated using the area under the curve (AUC), with results presented in receiver operating characteristic (ROC) curves. ROC curves provide an effective evaluation of the model by plotting the true positive rate against the false positive rate. Based on SVM, a linear classifier was applied to the calls, and a binomial test was conducted to compare the discrimination rate against the chance-level classification percentage. Additionally, random forest was used to obtain the contribution of each feature value, comparing sex, age, and individual differences in social calls under distress conditions in R. ferrumequinum.
2.5. Sound Wave Editing and Playback Experiment
The editing of playback files and the specific playback experimental procedures followed the methodology outlined in the previous chapter's study on the echolocation sound recognition capabilities of R. ferrumequinum. Using the habituation-dishabituation method, habituation sound files were played to the experimental subjects. When the bats ceased activity and remained motionless (no body or head rotation, no crawling, no wing or leg stretching, and no echolocation calls) for 30 seconds, they were considered habituated to the auditory stimulus. At this point, the stimulus was immediately switched to the dehabituation file. If the bat resumed any behavior (head nodding, ear movements, body turning, or emitting echolocation calls) after the switch, it was defined as having discriminated between the two playback files.
Pearson's chi-square test was used to statistical analysis to determine whether there were significant differences in the proportion of bats exhibiting any behavioral response following the switch to auditory stimuli across different playback combinations. If significant differences were detected, pairwise comparisons were performed using Fisher's exact test. Additionally, Kolmogorov-Smirnov tests were conducted to test for normality of behavioral data. For normally distributed data, t-tests were applied; for non-normally distributed data, Kruskal-Wallis H tests were used to compare differences in nodding, ear movements, body rotations, and echolocation sound waves counts across different test combinations. All statistical analyses were performed using SPSS 22.0 (IBM Corp., Armonk, NY, USA).
3. Results
3.1. Comparison of Machine Learning Classification Results and Key Features in Acoustic Signal Analysis
3.1.1. Comparison of Classification Results by Sex and Key Features
In this study, the spectral characteristics of NB-DFM calls of R. ferrumequinum between sexes are presented in Table 1. The support vector machine model achieved an accuracy of 67% in the gender classification task for NB-DFM calls produced in distress contexts (Figure 3a). The ROC curve indicates that the area under the SVM model curve exceeds 0.5, demonstrating that the model quality is suitable for subsequent classification tasks. The linear classification accuracy of NB-DFM vocalizations based on SVM reached 63.9% (Figure 3b), significantly higher than the chance classification rate (50.0%, Binomial test: p < 0.01). This indicates that NB-DFM vocalization acoustic parameters exhibit significant gender differences.
A random forest model was used to measure the contribution of each feature. The results indicated that Duration has the highest contribution at 20.5%, followed by Minimum frequency in center position and Root-Mean-Square (RMS) at 7.4% and 7.3%, respectively. The fourth most important feature is the maximum frequency in the maximum amplitude position, with a contribution of 6.1% (Figure 3c).
3.1.2. Comparison of Age Classification Results with Key Characteristics
In this study, the spectral characteristics of NB-DFM calls in adult and subadult R. ferrumequinum are shown in Table 2. The SVM model achieved an accuracy of 89% in the age classification task for NB-DFM vocalizations under distress contexts (Figure 4a). The ROC curve indicates that the area under the SVM model curve approaches 1.0, demonstrating excellent model quality suitable for subsequent classification tasks. The linear classification accuracy of NB-DFM vocalizations based on SVM reached 82.4% (Figure 4b), significantly higher than the chance classification rate (50.0%, Binomial test: p < 0.01). This indicates that acoustic parameters of NB-DFM vocalizations exhibit significant age-related differences.
A random forest model was used to measure the contribution of each feature. The results indicated the highest contribution was from the minimum frequency at the center position, accounting for 12.1%; followed by the peak frequency at the maximum amplitude position and the peak frequency at the center position, contributing 9.2% and 7.1% respectively; The fourth most important feature was the maximum frequency at the maximum amplitude position, with a contribution of 6.8% (Figure 4c).
3.1.3. Comparison of Individual Classification Results with Key Features
In this study, the spectral characteristics of NB-DFM calls of R. ferrumequinum across different individuals are presented in Table 3. The support vector machine model achieved an accuracy of 88% in the individual classification task for NB-DFM vocalizations under distress contexts (Figure 5a). The ROC curve showed that the area under the curve (AUC) of the SVM model was close to 1.0, indicating excellent model performance for further classification. The linear classification accuracy of NB-DFM vocalizations at the individual level based on SVM was 51.7% (Figure 5b), significantly higher than the chance level (1/10 = 10.0%, Binomial test: p < 0.01). This indicates that acoustic parameters of NB-DFM vocalizations exhibit significant individual variation.
A random forest model was used to evaluate the relative contribution of each acoustic feature. The results showed that duration contributed the most (16.3%), followed by the minimum frequency at the center position (12.0%) and the minimum frequency at the maximum amplitude position (8.7%). Root-mean-square (RMS) amplitude ranked fourth, with a contribution of 8.3% (Figure 5c).
3.2. Habituation Discrimination Experiment
3.2.1. Differences in Behavioral Responses to Acoustic Gender Recognition
When the habitual sound was a ♀ and the dishabituated sound was a ♂, 18 bats resumed body and head nodding movements and emitted echolocation calls. In contrast, when the habitual sound was a ♀ and the dishabituated sound was a ♀, 7 bats resumed body and head nodding movements and emitted echolocation calls (Figure 6a). When the habituation sound was ♂ and the dishabituation sound was ♂, 4 out of 20 bats resumed body and head nodding movements and emitted echolocation calls; when the habituation sound was ♂ and the dishabituation sound was ♀, 18 out of 20 resumed body and head nodding movements and emitted echolocation calls (Figure 6b). Among the four combinations of playback types, the proportion of bat responses showed significant differences (Pearson chi-square test: λ = 32.56, df = 3, p < 0.01; Fisher's exact test: p < 0.05). Following the auditory stimulus switch, significant differences were observed in head nodding, ear movements, body rotation, and echolocation sound waves counts (Head nodding: pF = 0.008, pM = 0.036; Ear movements: pF < 0.001, pM = 0.009; Body rotation: pF < 0.001, pM = 0.007; Echolocation: pF < 0.001, pM < 0.001; Figure 6a, b). These results indicate that R. ferrumequinum can discriminate between male and female NB-DFM calls.
3.2.2. Differences in Behavioral Responses to Age Recognition via Acoustic Waves
When the habitual sound wave was a subadult sound wave and the dehabituated sound wave was a subadult sound wave, 7 out of 20 R. ferrumequinum bats resumed body and nodding movements while emitting echolocation sound waves. When the habitual sound wave was a subadult sound wave and the dehabituated sound wave was an adult sound wave, 17 bats resumed body and nodding movements while emitting echolocation sound waves (Figure 7a). When the habitual sound wave was an adult sound wave and the dehabituated sound wave was a subadult sound wave, 18 subjects resumed body and nodding movements while emitting echolocation sound waves. When the habitual sound wave is an adult sound wave and the dehabituated sound wave is an adult sound wave, five subjects restart body and nodding movements while emitting echolocation sound waves (Figure 7b). Among the four combinations of playback types, the proportion of bat responses showed significant differences (Pearson chi-square test: λ = 31.97, df = 3, p < 0.01; Fisher's exact test: p < 0.05). Following transformation of acoustic stimuli, significant differences were observed in nodding, ear movements, body rotation, and echolocation sound waves counts among R. ferrumequinum bats (nodding: pS = 0.008, pA = 0.036; ear movements: pS < 0.001, pA = 0.041; body movements: pS = 0.006, pA = 0.0024; echolocation: pS < 0.001, pA < 0.001, as shown in Figure 7a, b). This indicates that the Horseshoe Bat can distinguish between NB-DFM vocalizations of different ages.
3.2.3. Differences in Behavioral Responses to Individual Recognition via Acoustic Signals
When the habitual sound waves were A and the stimulus sound waves were A', 20 R. ferrumequinums resumed body and head movements and emitted echolocation sound waves. When habitual echolocation sound waves were designated as A and stimulus sound waves as B, 17 out of 20 R. ferrumequinum resumed body and head movements while emitting echolocation sound waves. The proportion of bat responses showed a significant difference between the two playback combinations (Pearson chi-square test: λ = 22.56, df = 1, p < 0.01). Following conversion of acoustic stimuli, significant differences were observed in the nodding, ear movements, body rotation, and echolocation sound waves counts of R. ferrumequinum (p nodding = 0.008; p ear movements < 0.001; p body movements = 0.009; p echolocation < 0.001, as shown in Figure 8). This indicates that the horseshoe bat can recognize NB-DFM calls from different individuals.
4. Discussion
4.1. Classifying Bat Calls Using Machine Learning Methods
This study employed the Support Vector Machine model within machine learning to classify the sex, age, and individuals of NB-DFM calls under distress contexts in R. ferrumequinum. Additionally, 18 acoustic parameters were extracted for feature analysis. The support vector machine model achieved an accuracy rate exceeding 85% for age and individual classification. Although the accuracy rate for sex classification was only 67%, it was significantly higher than 50%, indicating that the model was of reasonable quality. The relatively low accuracy may be attributed to the limited sample size of bats used to train the sex classification model (six individuals per sex). Additionally, the SVM-based model showed significant discrepancies between its sex, age, and individual-level classifications of NB-DFM calls and the probabilities derived from chance classification. Consistent with other studies that classify bat sonic signals using traditional methods [45,46,47]. Analysis of the contribution of each acoustic parameter to gender, age, and individual classification in the random forest model indicates that duration, minimum frequency, and peak frequency contribute most significantly to call classification. The differences in echolocation sounds produced by R. ferrumequinum bats across different sexes, ages, and individuals primarily manifested in these acoustic parameters. The distinction lies in the different parameter positions. For instance, sex differences in acoustic signals and individual labels primarily stem from Duration and Minimum frequency center; whereas the Minimum frequency start contributes most significantly to age characteristics in acoustic signals. The model's strong performance in acoustic signal classification and feature contribution analysis, coupled with the high discriminative power of acoustic signals, demonstrates that machine learning can serve as an effective tool for studying acoustic signal classification.
Machine learning methods are widely applied across numerous research fields and have become an efficient and convenient tool in animal acoustics research. Previously, complex problems have been resolved through machine learning methods, such as the classification and recognition of vocalizations from multiple experimental subjects, as well as the categorization of large-scale experimental datasets [48,49,50]. Research by Larrañaga et al. found that machine learning methods can accurately classify the barking sounds of domestic dogs [51]. Using machine learning methods to classify the vocalizations of Rousettus aegyptiacus, it was found that vocalizations vary depending on the individual producing them and the intensity of the calls [52]. Machine learning also serves as a highly efficient and ideal tool for classifying and identifying the parametric acoustic characteristics of bat echolocation sound waves [53,54,55]. Compared to conventional discriminant analysis methods, machine learning approaches can handle a greater number of features simultaneously [56,57,58]. Additionally, compared to traditional discriminant analysis, support vector machine models make no assumptions about the distribution of the original data and impose fewer requirements on data distribution, thereby offering greater applicability .
Additionally, the ease of operation during experiments is also a key consideration in selecting methodologies. The support vector machine model adopted in this study features an algorithm that is straightforward to master and widely applicable. The 18 acoustic parameters used for feature segmentation in this study were all measured on individual syllables. Therefore, the model is applicable to vocalizing animals that produce sounds in syllable units, making it universally applicable to the species studied [49]. Another major advantage of machine learning is its ability to integrate multiple types of parameter features, including acoustic, individual, and behavioral characteristics [59].
4.2. Recognition of Distress Calls
Distress calls are vocalizations emitted by animals under adverse conditions, such as when attacked by predators [22,60], trapped [28,61], or held by experimenters [62,63,64]. Researchers have proposed numerous hypotheses to explain the functions of distress calls in birds: (1) to startle the predator, attract secondary predators to drive away the original predator, thereby facilitating escape [22,65,66]; (2) to warn conspecifics of the predator's presence [60,67]; (3) to provide other individuals with information about the predator, thus reducing their predation risk [66,68]; (4) to solicit help from kin or reciprocal altruists [62,63,69]; (5) to attract other individuals to mob the predator, which in turn facilitates the caller's escape [67,70]. Conover (1994) noted that these functions are not mutually exclusive, and distress signals may serve one or more functions [66].
Research on bat distress calls began in 1976, when Fenton et al. used field playback experiments to confirm that the distress calls of the Myotis Lucifugus could attract conspecific individuals to approach the sound source [71]. The results of this experiment are similar. Playback experiments indicate that when deconditioned acoustic files differing from the conditioning and habituation files were presented, R. ferrumequinum exhibited distinct behavioral responses—regardless of whether identification was based on sex, age, or individual level. It is demonstrated that R. ferrumequinum can distinguish NB-DFM calls from individuals of the same species but different sexes, ages, and individuals before and after sound wave conversion, thereby inferring its ability to recognize distress calls from conspecific individuals. The acoustic structure of bat distress calls shares similarities with other animal groups, as these calls contain noise components and are characterized by low-frequency, multi-harmonic patterns [72,73], enabling the encoding of information such as danger signals [74]. Consequently, bats' recognition of conspecific distress vocalizations within the same habitat facilitates the attraction of intraspecific individuals [20,28,75], thereby increasing escape opportunities. Eckenweber et al. (2016) found that the Saccopteryx bilineata can distinguish distress calls from conspecifics and heterospecifics within the same habitat, exhibiting significantly stronger responses to calls from conspecifics than from heterospecifics [25]. Additionally, bats not only eavesdrop on distress calls from conspecifics within the same territory but also respond strongly to distress signals from conspecifics within the same territory and even from heterospecific bats in different territories. Previous studies have confirmed that while bats can recognize distress calls from other species, there is no significant difference compared to intraspecific recognition. This is speculated to be related to predation risk assessment or interspecific cooperative mobbing, leading them to approach the direction of the sound source in both cases [51,76]. Additionally, researchers discovered that the European Pipistrellus pygmaeus can recognize distress calls from two co-occurring bat species, while exhibiting stronger responses to distress calls from four non-native bat species [20]. Research by Huang Xiaobin et al. indicates that the Rhinolophus sinicus, Myotis badius, and the Myotis laniger exhibit stronger responses to distress calls emitted by the Rhinolophus pusillus, the Miniopterus fuliginosus, and the Myotis altarium.
5. Conclusions
In summary, this study successfully applied machine learning to classify distress calls (NB-DFM) in R. ferrumequinum, demonstrating that these calls encode quantifiable signatures related to the caller's sex, age, and individual identity. The high classification accuracy achieved by the model, particularly for age and individual identification, provides strong evidence that the acoustic variation in these calls is perceptually relevant. Consequently, our findings support the hypothesis that R. ferrumequinum can utilize these acoustic cues to discriminate among conspecifics under stressful conditions. This work establishes a methodological framework for decoding complex animal vocalizations and advances our understanding of intraspecific communication in bats during distress.
Author Contributions
J.H. and J.L. designed and executed the experimental protocol. B.W. and M.G. assisted with field sampling and laboratory work. J.H. performed statistical analyses and drafted the methods and results sections of the manuscript, while J.H. and J.L. completed the manuscript. X.T. provided comments and revisions to the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Department of Science and Technology of Jilin Province, grant no. YDZJ202501ZYTS505.
Institutional Review Board Statement
According to the regulations of Wildlife Conservation of the People’s Republic of China (Chairman Decree (2016) No. 47), permits are required only for species included in the list of state-protected and region-protected wildlife species. None of the bats used in this study are endangered or region-protected species, so no specific permission is required. All animal experimental procedures were approved by the Forestry Bureau of Jilin Province, China (approval number: [2006] 178). All efforts were made to minimize the suffering of the animals. We confirm that all methods were performed in accordance with the relevant guidelines and regulations mentioned above.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest, financial or otherwise.
References
- Bradbury, J.W.; Vehrencamp, S.L. Principles of animal communication. Choice Rev. Online 1998, 49, 49–5059. [Google Scholar] [CrossRef]
- Gillam, E.; Fenton, M.B. Roles of acoustic social communication in the lives of bats. In Bat bioacoustics; Fenton, M.B., Grinnell, A.D., Popper, A.N., Fay, R.R., Eds.; Springer: New York, NY, USA, 2016; pp. 117–139. [Google Scholar]
- Hoi, H.; Darolová, A.; Krištofík, J. Slow song syllable rates provoke stronger male territorial responses in Eurasian reed warblers (Acrocephalus scirpaceus). J. Ornithol. 2023, 164, 193–202. [Google Scholar] [CrossRef]
- Golini, M.; Bell, M. The effects of anthrophony on song traits in European robins ( Erithacus rubecula ). Ecol. Evol. 2026, 16, e73018. [Google Scholar] [CrossRef] [PubMed]
- Elie, J.E.; Theunissen, F.E. Zebra finches identify individuals using vocal signatures unique to each call type. Nat. Commun. 2018, 9, 4026. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, R.A.; Amichaud, O.; Özcan, D.; Briefer, E.F. Biphonation in animal vocalizations: insights into communicative functions and production mechanisms. Philos. Trans. R. Soc. B Biol. Sci. 2025, 380, 20240011. [Google Scholar] [CrossRef]
- Shapiro, A.D. Recognition of individuals within the social group: signature vocalizations. In Handbook of behavioral neuroscience; Brudzynski, S.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; Vol. 19, pp. 495–503. [Google Scholar]
- Zimmermann, E.; Lerch, C. The complex acoustic design of an advertisement call in male mouse lemurs (Microcebus murinus, Prosimii, Primates) and sources of its variation. Ethology 1993, 93, 211–224. [Google Scholar] [CrossRef]
- Mitani, J.C. Male gibbon (Hylobates agilis) singing behavior: natural history, song variations and function. Ethology 1988, 79, 177–194. [Google Scholar] [CrossRef]
- Cowlishaw, G.U.Y. Song function in gibbons. Behaviour 1992, 121, 131–153. [Google Scholar] [CrossRef]
- Kanwal, J.S. Audiovocal communication in bats. Encycl. Neurosci. Acad. Press 2009, 681–690. [Google Scholar] [CrossRef]
- Salles, A.; Loscalzo, E.; Montoya, J.; Mendoza, R.; Boergens, K.M.; Moss, C.F. Auditory processing of communication calls in interacting bats. iScience 2024, 27, 109872. [Google Scholar] [CrossRef]
- Vargas-Mena, J.C.; Rocha, P.A.; Appel, G.; Melo Benathar, T.C.; Prous, X.; Tavares, V.D.C.; Trevelin, L. The echolocation calls and novel audible clicks of the thumbless bat, Furipterus horrens (Cuvier, 1828), in Eastern Amazonia, Brazil. Acta Chiropterologica 2026, 27. [Google Scholar] [CrossRef]
- Ma, J.; Kobayasi, K.; Zhang, S.; Metzner, W. Vocal communication in adult greater horseshoe bats, Rhinolophus ferrumequinum. J. Comp. Physiol. A 2006, 192, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Knörnschild, M.; Von Helversen, O. Nonmutual vocal mother–pup recognition in the greater sac-winged bat. Anim. Behav. 2008, 76, 1001–1009. [Google Scholar] [CrossRef]
- Davidson, S.M.; Wilkinson, G.S. Function of male song in the greater white-lined bat, Saccopteryx bilineata. Anim. Behav. 2004, 67, 883–891. [Google Scholar] [CrossRef]
- Wilkinson, G.S.; Boughman, J.W. Social calls coordinate foraging in greater spear-nosed bats. Anim. Behav. 1998, 55, 337–350. [Google Scholar] [CrossRef]
- Kloepper, L.N.; Tuninetti, A.; Bentley, I.; Harding, C.D.; Brighton, C.H.; Izadi, M.R.; Stevenson, R.L.; Taylor, G.K. Stereotyped active sensing in fast-diving echolocating bats. iScience 2025, 28, 114099. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, J.; Metzner, W. Different auditory feedback control for echolocation and communication in horseshoe bats. PLoS ONE 2013, 8, e62710. [Google Scholar] [CrossRef]
- González Noschese, C.S.; Olmedo, M.L.; Díaz, M.M. First characterization of the echolocation calls of Myotis dinellii (Chiroptera: Vespertilionidae). Mammal Res. 2024, 69, 53–58. [Google Scholar] [CrossRef]
- Huang, X.; Kanwal, J.S.; Jiang, T.; Long, Z.; Luo, B.; Yue, X.; Gu, Y.; Feng, J. Situational and age-dependent decision making during life threatening distress in Myotis macrodactylus. PLoS ONE 2015, 10, e0132817, 0doi. [Google Scholar] [CrossRef]
- Ruiz-Monachesi, M.R.; Labra, A. Complex distress calls sound frightening: The case of the weeping lizard. Anim. Behav. 2020, 165, 71–77. [Google Scholar] [CrossRef]
- Mariappan, S.; Bogdanowicz, W.; Raghuram, H.; Marimuthu, G.; Rajan, K.E. Structure of distress call: Implication for specificity and activation of dopaminergic system. J. Comp. Physiol. A 2016, 202, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Huang, X.; Wu, H.; Feng, J. Size and quality information in acoustic signals of Rhinolophus ferrumequinum in distress situations. Physiol. Behav. 2017, 173, 252–257. [Google Scholar] [CrossRef]
- Eckenweber, M.; Knörnschild, M. Responsiveness to conspecific distress calls is influenced by day-roost proximity in bats ( Saccopteryx bilineata ). R. Soc. Open Sci. 2016, 3, 160151. [Google Scholar] [CrossRef] [PubMed]
- Knörnschild, M.; Tschapka, M. Predator mobbing behaviour in the greater spear-nosed bat, Phyllostomus hastatus. Chiropt. Neotropical 2012, 18, 1132–1135. [Google Scholar]
- Lučan, R.K.; Šálek, M. Observation of successful mobbing of an insectivorous bat, Taphozous nudiventris (Emballonuridae), on an avian predator, Tyto alba (Tytonidae). Mammalia 2013, 77, 235–236. [Google Scholar] [CrossRef]
- Russ, J.M.; Racey, P.A.; Jones, G. Intraspecific responses to distress calls of the pipistrelle bat, Pipistrellus pipistrellus. Anim. Behav. 1998, 55, 705–713. [Google Scholar] [CrossRef]
- Manduva, V.C. Unlocking growth potential at the intersection of AI, robotics, and synthetic biology. Int. J. Mod. Comput. 2023, 6, 53–63. [Google Scholar]
- Jordan, M.I.; Mitchell, T.M. Machine learning: trends, perspectives, and prospects. Science 2015, 349, 255–260. [Google Scholar] [CrossRef]
- Huang, C.J.; Yang, Y.J.; Yang, D.X.; Chen, Y.J. Frog Classification using machine learning techniques. Expert Syst. Appl. 2009, 36, 3737–3743. [Google Scholar] [CrossRef]
- Acevedo, M.A.; Corrada-Bravo, C.J.; Corrada-Bravo, H.; Villanueva-Rivera, L.J.; Aide, T.M. Automated classification of bird and amphibian calls using machine learning: a comparison of methods. Ecol. Inform. 2009, 4, 206–214. [Google Scholar] [CrossRef]
- Shamir, L.; Yerby, C.; Simpson, R.; Von Benda-Beckmann, A.M.; Tyack, P.; Samarra, F.; Miller, P.; Wallin, J. Classification of large acoustic datasets using machine learning and crowdsourcing: application to whale calls. J. Acoust. Soc. Am. 2014, 135, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Redgwell, R.D.; Szewczak, J.M.; Jones, G.; Parsons, S. Classification of echolocation calls from 14 species of bat by support vector machines and ensembles of neural networks. Algorithms 2009, 2, 907–924. [Google Scholar] [CrossRef]
- Karaaslan, M.; Turkoglu, B.; Kaya, E.; Asuroglu, T. Voice analysis in dogs with deep learning: development of a fully automatic voice analysis system for bioacoustics studies. Sensors 2024, 24, 7978. [Google Scholar] [CrossRef] [PubMed]
- Duco, R.A.J.; Fontanilla, A.M.; M. Duya, M.R. Acoustic differentiation of horseshoe bats in Luzon Island, Philippines: developing a call identification key and uncovering potential cryptic species. Acta Chiropterologica 2026, 27. [Google Scholar] [CrossRef]
- Jin, L.; Yang, S.; Kimball, R.T.; Xie, L.; Yue, X.; Luo, B.; Sun, K.; Feng, J. Do pups recognize maternal calls in Pomona leaf-nosed bats, Hipposideros pomona? Anim. Behav. 2015, 100, 200–207. [Google Scholar] [CrossRef]
- Knörnschild, M.; Feifel, M.; Kalko, E.K.V. Mother–offspring recognition in the bat Carollia perspicillata. Anim. Behav. 2013, 86, 941–948. [Google Scholar] [CrossRef]
- Gelfand, D.L.; McCracken, G.F. Individual variation in the isolation calls of Mexican free-tailed bat pups (Tadarida brasiliensis mexicana). Anim. Behav. 1986, 34, 1078–1086. [Google Scholar] [CrossRef]
- Balcombe, J.P.; McCracken, G.F. Vocal recognition in Mexican free-tailed bats: Do pups recognize mothers? Anim. Behav. 1992, 43, 79–87. [Google Scholar] [CrossRef]
- Gillam, E.H.; Chaverri, G. Strong individual signatures and weaker group signatures in contact calls of Spix’s disc-winged bat, Thyroptera tricolor. Anim. Behav. 2012, 83, 269–276. [Google Scholar] [CrossRef]
- Fagerlund, S. Bird species recognition using support vector machines. EURASIP J. Adv. Signal Process. 2007, 2007, 038637. [Google Scholar] [CrossRef]
- Chen, W.P.; Chen, S.S.; Lin, C.C.; Chen, Y.Z.; Lin, W.C. Automatic recognition of frog calls using a multi-stage average spectrum. Comput. Math. Appl. 2012, 64, 1270–1281. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V. Scikit-Learn: machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar] [CrossRef]
- Arnold, B.D.; Wilkinson, G.S. Individual specific contact calls of pallid bats (Antrozous pallidus) attract conspecifics at roosting sites. Behav. Ecol. Sociobiol. 2011, 65, 1581–1593. [Google Scholar] [CrossRef]
- Carter, G.G.; Skowronski, M.D.; Faure, P.A.; Fenton, B. Antiphonal calling allows individual discrimination in white-winged vampire bats. Anim. Behav. 2008, 76, 1343–1355. [Google Scholar] [CrossRef]
- Ancillotto, L.; Russo, D. Selective aggressiveness in European free-tailed bats (Tadarida teniotis): influence of familiarity, age and sex. Naturwissenschaften 2014, 101, 221–228. [Google Scholar] [CrossRef]
- Kanwal, J.S.; Matsumura, S.; Ohlemiller, K.; Suga, N. Analysis of acoustic elements and syntax in communication sounds emitted by mustached bats. J. Acoust. Soc. Am. 1994, 96, 1229–1254. [Google Scholar] [CrossRef]
- Priyadarshani, N.; Marsland, S.; Castro, I. Automated birdsong recognition in complex acoustic environments: a review. J. Avian Biol. 2018, 49, jav–01447. [Google Scholar] [CrossRef]
- Adavanne, S.; Drossos, K.; Cakir, E.; Virtanen, T. Stacked convolutional and recurrent neural networks for bird audio detection. In Proceedings of the 2017 25th European Signal Processing Conference (EUSIPCO), August 2017; IEEE: Kos, Greece; pp. 1729–1733. [Google Scholar]
- Xiao, Y.H.; Wang, L.; Hoyt, J.R.; Jiang, T.L.; Lin, A.Q.; Feng, J. Stereotypy and variability of social calls among clustering female big-footed myotis (Myotis macrodactylus). Zool. Res. 2018, 39, 114. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, T.; Liu, M.; Li, A.; Xiao, Y.; Metzner, W.; Liu, Y. Comparing context-dependent call sequences employing machine learning methods: an indication of syntactic structure of greater horseshoe bats. J. Exp. Biol. 2019, 222, jeb.214072. [Google Scholar] [CrossRef]
- Bohn, K.M.; Schmidt-French, B.; Ma, S.T.; Pollak, G.D. Syllable acoustics, temporal patterns, and call composition vary with behavioral context in Mexican free-tailed bats. J. Acoust. Soc. Am. 2008, 124, 1838–1848. [Google Scholar] [CrossRef]
- Jin, L.; Wang, J.; Zhang, Z.; Sun, K.; Kanwal, J.S.; Feng, J. Postnatal development of morphological and vocal features in Asian particolored bat, Vespertilio sinensis. Mamm. Biol. 2012, 77, 339–344. [Google Scholar] [CrossRef]
- Wang, Z.; Hong, T.; Piette, M.A. Building thermal load prediction through shallow machine learning and deep learning. Appl. Energy 2020, 263, 114683. [Google Scholar] [CrossRef]
- Neyshabur, B.; Bhojanapalli, S.; McAllester, D.; Srebro, N. Exploring generalization in deep learning. Adv. Neural Inf. Process. Syst. 2017, 30, 5947–5956. [Google Scholar] [CrossRef]
- Wang, D.; Chen, J. Supervised speech separation based on deep learning: an overview. IEEEACM Trans. Audio Speech Lang. Process. 2018, 26, 1702–1726. [Google Scholar] [CrossRef]
- Kawaguchi, K.; Kaelbling, L.P.; Bengio, Y. Generalization in deep learning. Mach. Learn. Res. 2017, 18, 1–46. [Google Scholar] [CrossRef]
- Seyfarth, R.M.; Cheney, D.L. Signalers and receivers in animal communication. Annu. Rev. Psychol. 2003, 54, 145–173. [Google Scholar] [CrossRef]
- Koenig, W.D.; Stanback, M.T.; Hooge, P.N.; Mumme, R.L. Distress calls in the acorn woodpecker. The Condor 1991, 93, 637–643. [Google Scholar] [CrossRef]
- Avery, M.I.; Racey, P.A.; Fenton, M.B. Short distance location of hibernaculum by little brown bats (Myotis lucifugus). J. Zool. 1984, 204, 588–590. [Google Scholar] [CrossRef]
- Fong, L.J.M.; Navea, F.; Labra, A. Does Liolaemus lemniscatus eavesdrop on the distress calls of the sympatric weeping lizard? J. Ethol. 2021, 39, 11–17. [Google Scholar] [CrossRef]
- Stefanski, R.A.; Falls, J.B. A study of distress calls of song, swamp, and white-throated sparrows (Aves: Fringillidae). I. Intraspecific responses and functions. Can. J. Zool. 1972, 50, 1501–1512. [Google Scholar] [CrossRef]
- Brémond, J.C.; Aubin, T. Responses to distress calls by black-headed gulls, Larus ridibundus: the role of non-degraded features. Anim. Behav. 1990, 39, 503–511. [Google Scholar] [CrossRef]
- Driver, P.M.; Humphries, D.A. The significance of the high-intensity alarm call in captured passerines. Ibis 1969, 111, 243–244. [Google Scholar] [CrossRef]
- Conover, M.R. Stimuli eliciting distress calls in adult passerines and response of predators and birds to their broadcast. Behaviour 1994, 131, 19–37. [Google Scholar] [CrossRef]
- Fang, W.H.; Hsu, Y.H.; Lin, W.L.; Yen, S.C. The function of avian mobbing: an experimental test of ‘attract the mightier’ hypothesis. Anim. Behav. 2020, 170, 229–233. [Google Scholar] [CrossRef]
- Conover, M.R.; Perito, J.J. Response of starlings to distress calls and predator models holding conspecific prey. Z. Für Tierpsychol. 1981, 57, 163–172. [Google Scholar] [CrossRef]
- Rohwer, S.; Fretwell, S.D.; Tuckfield, R.C. Distress screams as a measure of kinship in birds. Am. Midl. Nat. 1976, 96, 418–430. [Google Scholar] [CrossRef]
- Neudorf, D.L.; Sealy, S.G. Distress calls of birds in a Neotropical cloud forest. Biotropica 2002, 34, 118. [Google Scholar] [CrossRef]
- Fenton, M.B.; Belwood, J.J.; Fullard, J.H.; Kunz, T.H. Responses of Myotis lucifugus (Chiroptera: Vespertilionidae) to calls of conspecifics and to other sounds. Can. J. Zool. 1976, 54, 1443–1448. [Google Scholar] [CrossRef]
- August, P.V. Acoustical properties of the distress calls of Artibeus jamaicensis and Phyllostomus hastatus (Chiroptera: Phyllostomidae). Southwest. Nat. 1985, 30, 371. [Google Scholar] [CrossRef]
- Carter, G.G.; Logsdon, R.; Arnold, B.D.; Menchaca, A.; Medellin, R.A. Adult vampire bats produce contact calls when isolated: acoustic variation by species, population, colony, and individual. PLoS ONE 2012, 7, e38791. [Google Scholar] [CrossRef]
- Hechavarría, J.C.; Beetz, M.J.; Macias, S.; Kössl, M. Distress vocalization sequences broadcasted by bats carry redundant information. J. Comp. Physiol. A 2016, 202, 503–515. [Google Scholar] [CrossRef]
- Ryan, J.M.; Clark, D.B.; Lackey, J.A. Response of Artibeus lituratus (Chiroptera: Phyllostomidae) to distress calls of conspecifics. J. Mammal. 1985, 66, 179–181. [Google Scholar] [CrossRef]
- Carter, G.; Schoeppler, D.; Manthey, M.; Knörnschild, M.; Denzinger, A. Distress calls of a fast-flying bat (Molossus molossus) provoke inspection flights but not cooperative mobbing. PLoS One 2015, 10, e0136146. [Google Scholar] [CrossRef]
Figure 1.
Syllable types and proportion of syllables produced by R. ferrumequinum in distress contexts. Syllables that constitute more than 1% of all vocalizations were plotted. NB-DFM, noise burst to downward frequency modulation; NB-SFM, noise burst to sinusoidal frequency modulation; BNBI, long broadband noise burst; SFM, sinusoidal frequency modulation; SND, sinusoidal frequency modulation to broadband noise burst to downward frequency modulation; NSND, broadband noise burst to sinusoidal frequency modulation to broadband noise burst to downward frequency modulation; SNS, sinusoidal frequency modulation to broadband noise burst to sinusoidal frequency modulation; UFM, upward frequency modulation; NB-UFM, noise burst to upward frequency modulation.
Figure 1.
Syllable types and proportion of syllables produced by R. ferrumequinum in distress contexts. Syllables that constitute more than 1% of all vocalizations were plotted. NB-DFM, noise burst to downward frequency modulation; NB-SFM, noise burst to sinusoidal frequency modulation; BNBI, long broadband noise burst; SFM, sinusoidal frequency modulation; SND, sinusoidal frequency modulation to broadband noise burst to downward frequency modulation; NSND, broadband noise burst to sinusoidal frequency modulation to broadband noise burst to downward frequency modulation; SNS, sinusoidal frequency modulation to broadband noise burst to sinusoidal frequency modulation; UFM, upward frequency modulation; NB-UFM, noise burst to upward frequency modulation.

Figure 2.
(a) Power spectrum, (b) oscillogram and (c) spectrogram of a NB-DFM syllables produced by greater horseshoe bats.
Figure 2.
(a) Power spectrum, (b) oscillogram and (c) spectrogram of a NB-DFM syllables produced by greater horseshoe bats.

Figure 3.
Diagram of sex characteristics of distress calls in R. ferrumequinum. (a) Receiver operator characteristic (ROC) graph of gender prediction by Support Vector Machine model; (b) Linear classification discriminant graph of gender prediction based on Support Vector Machine model. The number 1 represents female calls, the number 2 represents male calls; (c) Feature importance analysis and related acoustic parameters. The ranking of different features in the dataset by their relative importance. The features' importance was normalized, so that they sum to 1.0.
Figure 3.
Diagram of sex characteristics of distress calls in R. ferrumequinum. (a) Receiver operator characteristic (ROC) graph of gender prediction by Support Vector Machine model; (b) Linear classification discriminant graph of gender prediction based on Support Vector Machine model. The number 1 represents female calls, the number 2 represents male calls; (c) Feature importance analysis and related acoustic parameters. The ranking of different features in the dataset by their relative importance. The features' importance was normalized, so that they sum to 1.0.

Figure 4.
Diagram of age characteristics of distress calls in R. ferrumequinum. (a) Receiver operator characteristic (ROC) graph of age prediction by Support Vector Machine model; (b) Linear classification discriminant graph of age prediction based on Support Vector Machine model. The number 1 represents adult calls, the number 2 represents subadult calls; (c) Feature importance analysis and related acoustic parameters. The ranking of different features in the dataset by their relative importance. The features' importance was normalized, so that they sum to 1.0.
Figure 4.
Diagram of age characteristics of distress calls in R. ferrumequinum. (a) Receiver operator characteristic (ROC) graph of age prediction by Support Vector Machine model; (b) Linear classification discriminant graph of age prediction based on Support Vector Machine model. The number 1 represents adult calls, the number 2 represents subadult calls; (c) Feature importance analysis and related acoustic parameters. The ranking of different features in the dataset by their relative importance. The features' importance was normalized, so that they sum to 1.0.

Figure 5.
Diagram of individual characteristics of distress calls in R. ferrumequinum. (a) Receiver operator characteristic (ROC) graph of individual prediction by Support Vector Machine model; (b) Linear classification discriminant graph of individual prediction based on Support Vector Machine model. The numbers from 1 to 10 represent different individuals, respectively; (c) Feature importance analysis and related acoustic parameters. The ranking of different features in the dataset by their relative importance. The features' importance was normalized, so that they sum to 1.0.
Figure 5.
Diagram of individual characteristics of distress calls in R. ferrumequinum. (a) Receiver operator characteristic (ROC) graph of individual prediction by Support Vector Machine model; (b) Linear classification discriminant graph of individual prediction based on Support Vector Machine model. The numbers from 1 to 10 represent different individuals, respectively; (c) Feature importance analysis and related acoustic parameters. The ranking of different features in the dataset by their relative importance. The features' importance was normalized, so that they sum to 1.0.

Figure 6.
Results of the NB-DFM call type sex identification habituation-dishabituation playback experiment in R. ferrumequinum. F represents female playback files, M represents male playback files, FF represents female habituation sound waves to female de-habituation sound waves; FM represents from female habituation sound wave to male de-habituation sound wave; MF represents the change from male habituation sound wave to female de-habituation sound wave. MM represents the transition from male habituation to male de-habituation. The symbol * represents p < 0.05, and ** represents p < 0.01.
Figure 6.
Results of the NB-DFM call type sex identification habituation-dishabituation playback experiment in R. ferrumequinum. F represents female playback files, M represents male playback files, FF represents female habituation sound waves to female de-habituation sound waves; FM represents from female habituation sound wave to male de-habituation sound wave; MF represents the change from male habituation sound wave to female de-habituation sound wave. MM represents the transition from male habituation to male de-habituation. The symbol * represents p < 0.05, and ** represents p < 0.01.

Figure 7.
Results of the NB-DFM call type age identification habituation-dishabituation playback experiment in Rhinolophus ferrumequinum. "S" represents subadult playback files, and "A" represents adult playback files; "SS" represents subadult habituated sound waves to subadult de-habituated sound waves; "SA" represents subadult habituated sound waves to adult de-habituation sound wave; "AS" represents adult habituated sound waves to subadult de-habituation sound wave; "AA" represents adult habituated sound waves to adult de-habituation sound wave. The symbol * represents p < 0.05, and ** represents p < 0.01.
Figure 7.
Results of the NB-DFM call type age identification habituation-dishabituation playback experiment in Rhinolophus ferrumequinum. "S" represents subadult playback files, and "A" represents adult playback files; "SS" represents subadult habituated sound waves to subadult de-habituated sound waves; "SA" represents subadult habituated sound waves to adult de-habituation sound wave; "AS" represents adult habituated sound waves to subadult de-habituation sound wave; "AA" represents adult habituated sound waves to adult de-habituation sound wave. The symbol * represents p < 0.05, and ** represents p < 0.01.

Figure 8.
Results of the NB-DFM call type individual identification habituation-dishabituation playback experiment in R. ferrumequinum. A and B respectively represent two different individuals. AA' represents the transformation from the habituated sound file of individual A to another sound file of individual A. AB represents the change from the habituated sound file of individual A to the dehabituated sound file of individual B. The symbol ** represents p < 0.01.
Figure 8.
Results of the NB-DFM call type individual identification habituation-dishabituation playback experiment in R. ferrumequinum. A and B respectively represent two different individuals. AA' represents the transformation from the habituated sound file of individual A to another sound file of individual A. AB represents the change from the habituated sound file of individual A to the dehabituated sound file of individual B. The symbol ** represents p < 0.01.


Table 1.
Description of the acoustic parameters of NB-DFM syllables of different sexes in R. ferrumequinum.
Table 1.
Description of the acoustic parameters of NB-DFM syllables of different sexes in R. ferrumequinum.
| Parameters | Mean ± SD | Mean ± SD | Minimum | Maximum | ||
|---|---|---|---|---|---|---|
| Female | Male | Female | Male | |||
| Duration | 0.11 ± 0.02 | 0.05 ± 0.05 | 0.05 | 0.28 | 0.18 | 0.14 |
| RMS | -17.16 ± 2.42 | 2.10 ± -23.91 | -23.91 | -13.90 | -12.09 | -17.76 |
| Peak frequency in start position | 34.35 ± 18.75 | 19.39 ± 11.20 | 11.20 | 77.80 | 77.60 | 32.48 |
| Minimum frequency in start position | 14.15 ± 4.13 | 4.99 ± 7.50 | 7.50 | 44.40 | 24.10 | 14.37 |
| Maximum frequency in start position | 82.24 ± 13.80 | 15.86 ± 18.70 | 18.70 | 122.50 | 115.70 | 81.07 |
| Bandwidth in start position | 68.05 ± 14.12 | 16.12 ± 6.30 | 6.30 | 110.10 | 103.20 | 66.65 |
| Peak frequency in end position | 37.89 ± 17.41 | 15.85 ± 12.20 | 12.20 | 83.20 | 82.20 | 36.77 |
| Minimum frequency in end position | 20.23 ± 3.42 | 3.19 ± 11.40 | 11.40 | 41.90 | 37.80 | 19.95 |
| Maximum frequency in end position | 81.52 ± 14.05 | 12.95 ± 16.80 | 16.80 | 117.90 | 118.40 | 81.86 |
| Bandwidth in end position | 61.24 ± 13.79 | 13.54 ± 4.60 | 4.60 | 96.90 | 96.40 | 61.85 |
| Peak frequency in center position | 36.71 ± 16.15 | 16.56 ± 20.70 | 20.70 | 74.70 | 75.90 | 38.74 |
| Minimum frequency in center position | 20.11 ± 5.57 | 4.16 ± 6.10 | 6.10 | 25.10 | 49.00 | 18.74 |
| Maximum frequency in center position | 82.96 ± 12.12 | 14.66 ± 52.00 | 52.00 | 120.60 | 120.10 | 84.01 |
| Bandwidth in center position | 62.79 ± 14.23 | 16.34 ± 29.50 | 29.50 | 111.30 | 102.20 | 65.43 |
| Peak frequency in maximum amplitude position | 42.18 ± 17.30 | 15.17 ± 19.00 | 19.00 | 75.90 | 72.00 | 39.76 |
| Minimum frequency in maximum amplitude position | 23.31 ± 6.30 | 3.70 ± 11.40 | 11.40 | 45.80 | 55.90 | 22.08 |
| Maximum frequency in maximum amplitude position | 78.35 ± 13.59 | 12.34 ± 47.80 | 47.80 | 113.50 | 115.40 | 73.07 |
| Bandwidth in maximum amplitude position | 54.99 ± 15.27 | 12.60 ± 5.10 | 5.10 | 91.70 | 95.90 | 51.47 |
Note: The unit of duration and interval is second (s). rms has no unit and other acoustic acoustic parameters' unit is kHz. The same below.
Table 2.
Description of the acoustic parameters of adult and subadult NB-DFM syllables in R. ferrumequinum.
Table 2.
Description of the acoustic parameters of adult and subadult NB-DFM syllables in R. ferrumequinum.
| Parameters | Adult Mean ± SD | Subadult Mean ± SD | Minimum | Maximum | ||
|---|---|---|---|---|---|---|
| Adult | Subadult | Adult | Subadult | |||
| Duration | 0.11 ± 0.02 | 0.05 ± 0.05 | 0.05 | 0.28 | 0.18 | 0.14 |
| RMS | -17.16 ± 2.42 | 2.10 ± -23.91 | -23.91 | -13.90 | -12.09 | -17.76 |
| Peak frequency in start position | 34.35 ± 18.75 | 19.39 ± 11.20 | 11.20 | 77.80 | 77.60 | 32.48 |
| Minimum frequency in start position | 14.15 ± 4.13 | 4.99 ± 7.50 | 7.50 | 44.40 | 24.10 | 14.37 |
| Maximum frequency in start position | 82.24 ± 13.80 | 15.86 ± 18.70 | 18.70 | 122.50 | 115.70 | 81.07 |
| Bandwidth in start position | 68.05 ± 14.12 | 16.12 ± 6.30 | 6.30 | 110.10 | 103.20 | 66.65 |
| Peak frequency in end position | 37.89 ± 17.41 | 15.85 ± 12.20 | 12.20 | 83.20 | 82.20 | 36.77 |
| Minimum frequency in end position | 20.23 ± 3.42 | 3.19 ± 11.40 | 11.40 | 41.90 | 37.80 | 19.95 |
| Maximum frequency in end position | 81.52 ± 14.05 | 12.95 ± 16.80 | 16.80 | 117.90 | 118.40 | 81.86 |
| Bandwidth in end position | 61.24 ± 13.79 | 13.54 ± 4.60 | 4.60 | 96.90 | 96.40 | 61.85 |
| Peak frequency in center position | 36.71 ± 16.15 | 16.56 ± 20.70 | 20.70 | 74.70 | 75.90 | 38.74 |
| Minimum frequency in center position | 20.11 ± 5.57 | 4.16 ± 6.10 | 6.10 | 25.10 | 49.00 | 18.74 |
| Maximum frequency in center position | 82.96 ± 12.12 | 14.66 ± 52.00 | 52.00 | 120.60 | 120.10 | 84.01 |
| bandwidth in center position | 62.79 ± 14.23 | 16.34 ± 29.50 | 29.50 | 111.30 | 102.20 | 65.43 |
| Peak frequency in maximum amplitude position | 42.18 ± 17.30 | 15.17 ± 19.00 | 19.00 | 75.90 | 72.00 | 39.76 |
| Minimum frequency in maximum amplitude position | 23.31 ± 6.30 | 3.70 ± 11.40 | 11.40 | 45.80 | 55.90 | 22.08 |
| Maximum frequency in maximum amplitude position | 78.35 ± 13.59 | 12.34 ± 47.80 | 47.80 | 113.50 | 115.40 | 73.07 |
| Bandwidth in maximum amplitude position | 54.99 ± 15.27 | 12.60 ± 5.10 | 5.10 | 91.70 | 95.90 | 51.47 |
Table 3.
Summary of mean ± standard deviation (SD) and range for acoustic parameters of NB-DFM syllable for individual recognition in R. ferrumequinum.
Table 3.
Summary of mean ± standard deviation (SD) and range for acoustic parameters of NB-DFM syllable for individual recognition in R. ferrumequinum.
| Parameters | Minimum | Maximum | Mean | SD |
|---|---|---|---|---|
| Duration | 0.05 | 0.28 | 0.13 | 0.04 |
| RMS | -23.94 | -12.09 | -17.46 | 2.28 |
| Peak frequency in start position | 10.20 | 77.80 | 33.41 | 19.05 |
| Minimum frequency in start position | 4.80 | 44.40 | 14.26 | 4.57 |
| Maximum frequency in start position | 16.60 | 122.50 | 81.66 | 14.85 |
| Bandwidth in start position | 6.30 | 110.10 | 67.35 | 15.14 |
| Peak frequency in end position | 12.20 | 83.20 | 37.33 | 16.62 |
| Minimum frequency in end position | 11.20 | 41.90 | 20.09 | 3.30 |
| Maximum frequency in end position | 16.80 | 118.40 | 81.69 | 13.48 |
| Bandwidth in end position | 4.60 | 96.90 | 61.54 | 13.64 |
| Peak frequency in center position | 19.20 | 75.90 | 37.73 | 16.35 |
| Minimum frequency in center position | 6.10 | 49.00 | 19.42 | 4.95 |
| Maximum frequency in center position | 46.60 | 120.60 | 83.48 | 13.43 |
| Bandwidth in center position | 26.80 | 111.30 | 64.11 | 15.34 |
| Peak frequency in maximum amplitude position | 15.10 | 75.90 | 40.97 | 16.28 |
| Minimum frequency in maximum amplitude position | 9.50 | 55.90 | 22.69 | 5.19 |
| Maximum frequency in maximum amplitude position | 45.60 | 115.40 | 75.71 | 13.22 |
| Bandwidth in maximum amplitude position | 5.10 | 95.90 | 53.23 | 14.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



