Submitted:
18 March 2026
Posted:
19 March 2026
You are already at the latest version
Abstract
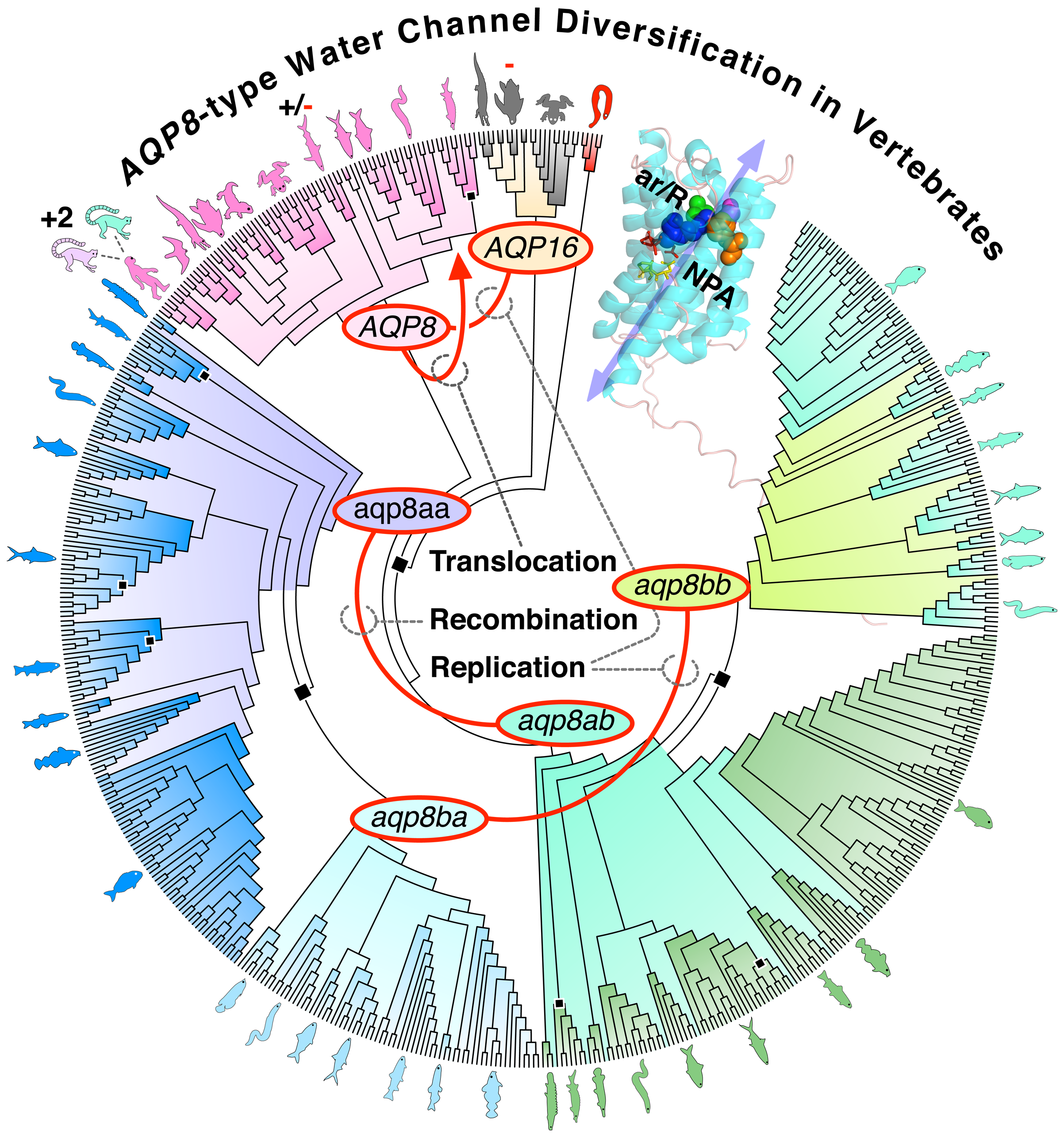
AQP8-type water channels are expressed superficially in the plasma membrane or intracellularly in the inner mitochondrial membrane where they respectively function in osmohomeostasis or as peroxiporins to alleviate oxidative stress. To date only single-copy AQP8 or AQP16 genes are known in tetrapods and two binary gene clusters composed of aqp8aa-aqp8ab and aqp8ba-aqp8bb in teleost fishes. Here using phylogenomic and synteny analyses we revise this view and show that bony fish aqp8aa, -ab, -ba and -bb genes are non-canonical co-orthologs that independently arose at chromosomal breakpoints. Conversely, canonical orthologs of tetrapod AQP8 are now detected in all vertebrate classes except hagfishes. In cartilaginous fishes, intact aqp8 orthologs and linked pseudogenes exist in squalomorph sharks, but only fractionated aqp8-like pseudogenes in galeomorph sharks. Some isolated aqp8-like exons are detected batoid ray genomes, while no aqp8-type coding sequences are currently found in holocephalan genomes. In ray-finned fishes, the canonical ortholog of tetrapod AQP8 underwent gene translocation in their common ancestor ~400 million years ago, but was subsequently inactivated or lost in many descendant lineages. In close temporal proximity to this gene translocation event, the actinopterygian aqp8aa-aqp8ab binary gene cluster was generated in the original syntenic locus potentially as a result of meiotic recombination. Our data support a model of total chromosmal replication for the generation of tetropod AQP16 genes and the teleost aqp8ba-aqp8bb gene cluster. We further uncover additional duplicates in Strepsirrhini primates that provide an eminent example of the stochastic nature of neofunctionalization. The present data thus suggest that chromosomal translocation, recombination and replication events contributed to the diversification of vertebrate AQP8-type genes.
Keywords:
1. Introduction
2. Results
2.1. Canonical AQP8 Orthologs Exist in All Vertebrate Lineages

2.2. Pseudogenes Confirm the Loss of AQP8-Type Orthologs in Piscine Genomes


2.3. Differential Retention of the aqp8 Binary Gene Clusters in Osteoglossomorpha

2.4. Synteny Reveals Gene Translocation of Canonical aqp8 in Actinopterygii
2.5. AQP8 Is Duplicated in the Strepsirrhini Primates
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability
Conflicts of Interest
References
- Agemark, M.; Kowal, J.; Kukulski, W.; Nordén, K.; Gustavsson, N.; Johanson, U.; Engel, A.; Kjellbom, P. Reconstitution of water channel function and 2D-crystallization of human aquaporin 8. Biochim. Biophys. Acta 2012, 1818, 839-850. [CrossRef]
- Kirscht, A.; Sonntag, Y.; Kjellbom, P.; Johanson, U. A structural preview of aquaporin 8 via homology modeling of seven vertebrate isoforms. BMC Struct. Biol. 2018, 18, 1-5. [CrossRef]
- Ishibashi, K.; Kuwahara, M.; Kageyama, Y.; Tohsaka, A.; Marumo, F.; Sasaki, S. Cloning and functional expression of a second new aquaporin abundantly expressed in testis. Biochem. Biophys. Res. Commun. 1997, 237, 714-718. [CrossRef]
- Jahn, T.P.; Møller, A.L.; Zeuthen, T.; Holm, L.M.; Klærke, D.A.; Mohsin, B.; Kühlbrandt, W.; Schjoerring, J.K. Aquaporin homologues in plants and mammals transport ammonia. FEBS lett. 2004, 574, 31-36. [CrossRef]
- Holm, L.M.; Jahn, T.P.; Møller, A.L.; Schjoerring, J.K.; Ferri, D.; Klaerke, D.A.; Zeuthen, T. NH3 and NH4+ permeability in aquaporin-expressing Xenopus oocytes. Pflügers Archiv 2005, 450, 415-428. [CrossRef]
- Liu, K.; Nagase, H.; Huang, C.G.; Calamita, G.; Agre, P. Purification and functional characterization of aquaporin-8. Biol. Cell 2006, 98, 153-161. [CrossRef]
- Saparov, S.M.; Liu, K.; Agre, P.; Pohl, P. Fast and selective ammonia transport by aquaporin-8. J. Biol. Chem. 2007, 282, 5296-5301. [CrossRef]
- Tingaud-Sequeira, A.; Calusinska, M.; Finn, R.N.; Chauvigné, F.; Lozano, J.; Cerdà, J. The zebrafish genome encodes the largest vertebrate repertoire of functional aquaporins with dual paralogy and substrate specificities similar to mammals. BMC Evol. Biol. 2010, 10, 38. [CrossRef]
- Engelund, M.B.; Chauvigné, F.; Christensen, B.M.; Finn, R.N.; Cerdà, J.; Madsen, S.S. Differential expression and novel permeability properties of three aquaporin 8 paralogs from seawater-challenged Atlantic salmon smolts. J. Exp. Biol. 2013, 216, 3873-3885. [CrossRef]
- Bienert, G.P.; Chaumont, F. Aquaporin-facilitated transmembrane diffusion of hydrogen peroxide. Biochim. Biophys. Acta 2014, 1840, 1596-1604. [CrossRef]
- Chauvigné, F.; Boj, M.; Finn, R.N.; Cerdà, J. Mitochondrial aquaporin-8-mediated hydrogen peroxide transport is essential for teleost spermatozoon motility. Sci. Rep. 2015, 5, 7789. [CrossRef]
- Bertolotti, M.; Farinelli, G.; Galli, M.; Aiuti, A.; Sitia, R. AQP8 transports NOX2-generated H2O2 across the plasma membrane to promote signaling in B cells. J. Leucocyte Biol. 2016, 100, 1071-1079. [CrossRef]
- Bestetti, S.; Medraño-Fernandez, I.; Galli, M.; Ghitti, M.; Bienert, G.P.; Musco, G.; Orsi, A.; Rubartelli, A.; Sitia, R. A persulfidation-based mechanism controls aquaporin-8 conductance. Sci. Adv. 2018, 4, eaar5770. [CrossRef]
- Chauvigné, F.; Ducat, C.; Ferré, A.; Hansen, T.; Carracal, M.; Albián, J.; Finn, R.N.; Cerdà, J. A multiplier peroxiporin signal transduction pathway powers piscine spermatozoa. Proc. Natl. Acad. Sci. USA 2021, 118, (10) e2019346118. [CrossRef]
- Krüger, C.; Waldeck-Weiermair, M.; Kaynert, J.; Pokrant, T.; Komaragiri, Y.; Otto, O.; Michel, T.; Elsner, M. AQP8 is a crucial H2O2 transporter in insulin-producing RINm5F cells. Redox Biol. 2021, 43, 101962. [CrossRef]
- Calamita, G.; Ferri, D.; Bazzini, C.; Mazzone, A.; Botta, G.; Liquori, G.E.; Paulmichl, M.; Portincasa, P.; Meyer, G.; Svelto, M. Expression and subcellular localization of the AQP8 and AQP1 water channels in the mouse gall bladder epithelium. Biol. Cell 2005, 97, 415-423. [CrossRef]
- Wellner, R.B.; Redman, R.S.; Swaim, W.D.; Baum, B.J. Further evidence for AQP8 expression in the myoepithelium of rat submandibular and parotid glands. Pflügers Archiv 2005, 451, 642-645. [CrossRef]
- Molinas, S.M.; Trumper, L.; Marinelli, R.A. Mitochondrial aquaporin-8 in renal proximal tubule cells: evidence for a role in the response to metabolic acidosis. Am. J Physiol. - Renal Physiol. 2012, 303, F458-F466. [CrossRef]
- Dong, C.; Chen, L.; Feng, J.; Xu, J.; Mahboob, S.; Al-Ghanim, K.; Li, X.; Xu, P. Genome wide identification, phylogeny, and expression of aquaporin genes in common carp (Cyprinus carpio). PLoS One 2016, 11, (12):e0166160. [CrossRef]
- Medrano-Fernandez, I.; Bestetti, S.; Bertolotti, M.; Bienert, G.P.; Bottino, C.; Laforenza, U.; Rubartelli, A.; Sitia, R. Stress regulates aquaporin-8 permeability to impact cell growth and survival. Antiox. Redox Signaling 2016, 24, 1031-1044. [CrossRef]
- Finn, R.N.; Cerdà, J., (2018) ‘Aquaporin’, In: Choi, S. (Ed), Encyclopedia of Signaling Molecules, 2nd Edition, Springer, New York, pp 1-18.
- Cutler, C.P.; Mainer, S.; Ojo, T. The aquaporin 8 (AQP8) membrane channel gene is present in the elasmobranch dogfish (Squalus acanthias) genome and is expressed in brain but not in gill, kidney or intestine. Comp. Biochem. Physiol. 2022, B260, 110730. [CrossRef]
- Wang, S.; Qin, Y.; Sheng, J.; Duan, X.; Shen, L.; Liu, D. Aquaporin 8ab is required in zebrafish embryonic intestine development. Acta Biochim. Biophys. Sinica 2022, 54, 952. [CrossRef]
- Huo, X.; et al.Wu, F. Hepatocyte aquaporin 8-mediated water transport facilitates bile dilution and prevents gallstone formation in mice. J. Hepatol. 2025, 82, 464-479. [CrossRef]
- Calamita, G.; Ferri, D.; Gena, P.; Liquori, G.E.; Cavalier, A.; Thomas, D.; Svelto, M. The inner mitochondrial membrane has aquaporin-8 water channels and is highly permeable to water. J. Biol. Chem. 2005, 280, 17149-17153. [CrossRef]
- Huang, B.; Jin, L.; Zhang, L.; Cui, X.; Zhang, Z.; Lu, Y.; Yu, L.; Ma, T.; Zhang, H. Aquaporin-8 transports hydrogen peroxide to regulate granulosa cell autophagy. Front. Cell Dev. Biol. 2022, 10, 897666. [CrossRef]
- Cerdà, J.; Chauvigné, F.; Finn, R.N., (2023) ‘Evolution and function of peroxiporins in piscine spermatozoa’, In: Bienert, F.S. (Ed), Peroxiporins: Redox Signal Mediators in and Between Cells, pp 158-171.
- Abascal, F.; Irisarri, I.; Zardoya, R. Diversity and evolution of membrane intrinsic proteins. Biochim. Biophys. Acta 2014, 1840, 1468-1481. [CrossRef]
- Finn, R.N.; Cerdà, J. Evolution and functional diversity of aquaporins. Biol. Bull. 2015, 229, 6-23.
- Finn, R.N.; Chauvigné, F.; Stavang, J.A.; Belles, X.; Cerdà, J. Insect glycerol transporters evolved by functional co-option and gene replacement. Nature Communications 2015, 6, 7814. [CrossRef]
- Stavang, J.A.; Chauvigné, F.; Kongshaug, H.; Cerdà, J.; Nilsen, F.; Finn, R.N. Phylogenomic and functional analyses of salmon lice aquaporins uncover the molecular diversity of the superfamily in Arthropoda. BMC Genomics 2015, 16, 618. [CrossRef]
- Catalán-García, M.; Chauvigné, F.; Ferré, A.; Stavang, J.A.; Nilsen, F.; Cerdà, J.; Finn, R.N. Lineage-level divergence of copepod glycerol transporters and the emergence of isoform-specific trafficking regulation. Communications Biol. 2021, 4, 643. [CrossRef]
- Wang, W.; Zhang, X.S.; Wang, Z.N.; Zhang, D.X. Evolution and phylogenetic diversity of the aquaporin gene family in arachnids. Int. J. Biol. Macromol. 2023, 240, 124480. [CrossRef]
- Finn, R.N.; Chauvigné, F.; Hlidberg, J.B.; Cutler, C.P.; Cerdà, J. The lineage-specific evolution of aquaporin gene clusters facilitated tetrapod terrestrial adaptation. PloS One 2014, 9, (11):e113686. [CrossRef]
- Crow, K.D.; Smith, C.D.; Cheng, J.F.; Wagner, G.P.; Amemiya, C.T. An independent genome duplication inferred from Hox paralogs in the American paddlefish - a representative basal ray-finned fish and important comparative reference. Genome Biol. Evol. 2012, 4, 937-953. [CrossRef]
- Du, K.; Stöck, M.; Kneitz, S.; Klopp, C.; Woltering, J.M.; Adolfi, M.C.; Feron, R.; Prokopov, D.; Makunin, A.; Kichigin, I. The sterlet sturgeon genome sequence and the mechanisms of segmental rediploidization. Nat. Ecol. Evol. 2020, 4, 841-852. [CrossRef]
- Finn, R.N.; Cerdà, J. Genetic adaptations for the oceanic success of fish eggs. Trends Genet. 2024, 40, 6.
- Fraser, J.A.; Huang, J.C.; Pukkila-Worley, R.; Alspaugh, J.A.; Mitchell, T.G.; Heitman, J. Chromosomal translocation and segmental duplication in Cryptococcus neoformans. Eukaryotic Cell 2005, 4, 401-406. [CrossRef]
- Kuzmin, E.; Taylor, J.S.; Boone, C. Retention of duplicated genes in evolution. Trends Genet. 2021, 38, 59-72. [CrossRef]
- Albalat, R.; Cañestro, C. Evolution by gene loss. Nature Reviews Genetics. Nature Rev. Genetics 2016, 2016, 379-391. [CrossRef]
- Mitelman, F.; Johansson, B.; Mertens, F. The impact of translocations and gene fusions on cancer causation. Nature Revi. Cancer 2007, 7, 233-245. [CrossRef]
- Harewood, L.; Schütz, F.; Boyle, S.; Perry, P.; Delorenzi, M.; Bickmore, W.A.; Reymond, A. The effect of translocation-induced nuclear reorganization on gene expression. Genome Res. 2010, 20, 554-564. [CrossRef]
- Wadsworth, M.E.; Page, M.L.; Heberle, B.A.; Miller, J.B.; Steely, C.J.; Ebbert, M.T. Dark and camouflaged genomic regions remain challenging in CHM13. Sci. Rep. 2026, 16, 1557. [CrossRef]
- Litman, T.; Søgaard, R.; Zeuthen, T., (2009) ‘Ammonia and urea permeability of mammalian aquaporins’, In: Beitz, E. (Ed), Aquaporins, Springer, Berlin, pp 327-358. [CrossRef]
- Li, C.; Wang, W., (2014) ‘Urea transport mediated by aquaporin water channel proteins’, In: Yang, B.; Sands, J.M. (Eds), Urea Transporters, Springer, London, pp 227-265.
- Evans, D.H.; Piermarini, P.M.; Choe, K.P., (2004) ‘Homeostasis: osmoregulation, pH regulation, and nitrogen excretion’, In: Carrier, J.C.; Musick, J.A.; Heithaus, M.R. (Eds), Biology of Sharks and their Relatives, CRC Press, Taylor Francis Group, pp 247-268.
- Sands, J.M.; Blount, M.A., (2014) ‘Genes and proteins of urea transporters’, In: Yang, B.Y.; Sands, J.M. (Eds), Urea Transporters, Springer, Dordrecht, pp 45-63.
- Yilmaz, O.; Chauvigné, F.; Ferré, A.; Nilsen, F.; Fjelldal, P.G.; Cerdà, J.; Finn, R.N. Unravelling the complex duplication history of deuterostome glycerol transporters. Cells 2020, 9, (7):1663. [CrossRef]
- Assis, R.; Bachtrog, D. Neofunctionalization of young duplicate genes in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, 17409-17414. [CrossRef]
- Ferré, A.; Chauvigné, F.; Vlasova, A.; Norberg, B.; Bargelloni, L.; Guigó, R.; Finn, R.N.; Cerdà, J. Functional Evolution of Clustered Aquaporin Genes Reveals Insights into the Oceanic Success of Teleost Eggs. Mol. Biol. Evol. 2023, 40, (4):msad071. [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673-4680. [CrossRef]
- Suyama, M.; Torrents, D.; Bork, P. PAL2NAL: robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Res. 2006, 34, (suppl_2):W609. [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572-1574. [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Systematic Biol. 2018, 67, 901-904. [CrossRef]
- Nguyen, N.T.; Vincens, P.; Dufayard, J.F.; Roest Crollius, H.; Louis, A. Genomicus in 2022: comparative tools for thousands of genomes and reconstructed ancestors. Nucleic Acids Res. 2022, 50, D1025-D1031. [CrossRef]
- Steiper, M.E.; Young, N.M., (2009) ‘Primates (Primates)’, In: Hedges, S.B.; Kumar, S. (Eds), The Timetree of Life, Oxford University Press, pp 482-486.
- Naylor, G.J.; Caira, J.N.; Jensen, K.; Rosana, K.A.; Straube, N.; Lakner, C., (2012) ‘Elasmobranch phylogeny: a mitochondrial estimate based on 595 species’, In: Carrier, J.C.; Musick, J.A.; Heithaus, M.R. (Eds), Biology of sharks and their relatives, CRC Press, Taylor & Francis Group, pp 31-56.
- Hughes, L.C. et al.Shi, Q. Comprehensive phylogeny of ray-finned fishes (Actinopterygii) based on transcriptomic and genomic data. Proc. Natl. Acad. Sci. USA 2018, 115, 6249-6254. [CrossRef]
- Kumar, S.; Suleski, M.; Craig, J.M.; Kasprowicz, A.E.; Sanderford, M.; Li, M.; Stecher, G.; Hedges, S.B. TimeTree 5: an expanded resource for species divergence times. Mol. Biol. Evol. 2022, 39, (8):msac174. [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



