Submitted:
06 March 2026
Posted:
09 March 2026
You are already at the latest version
Abstract
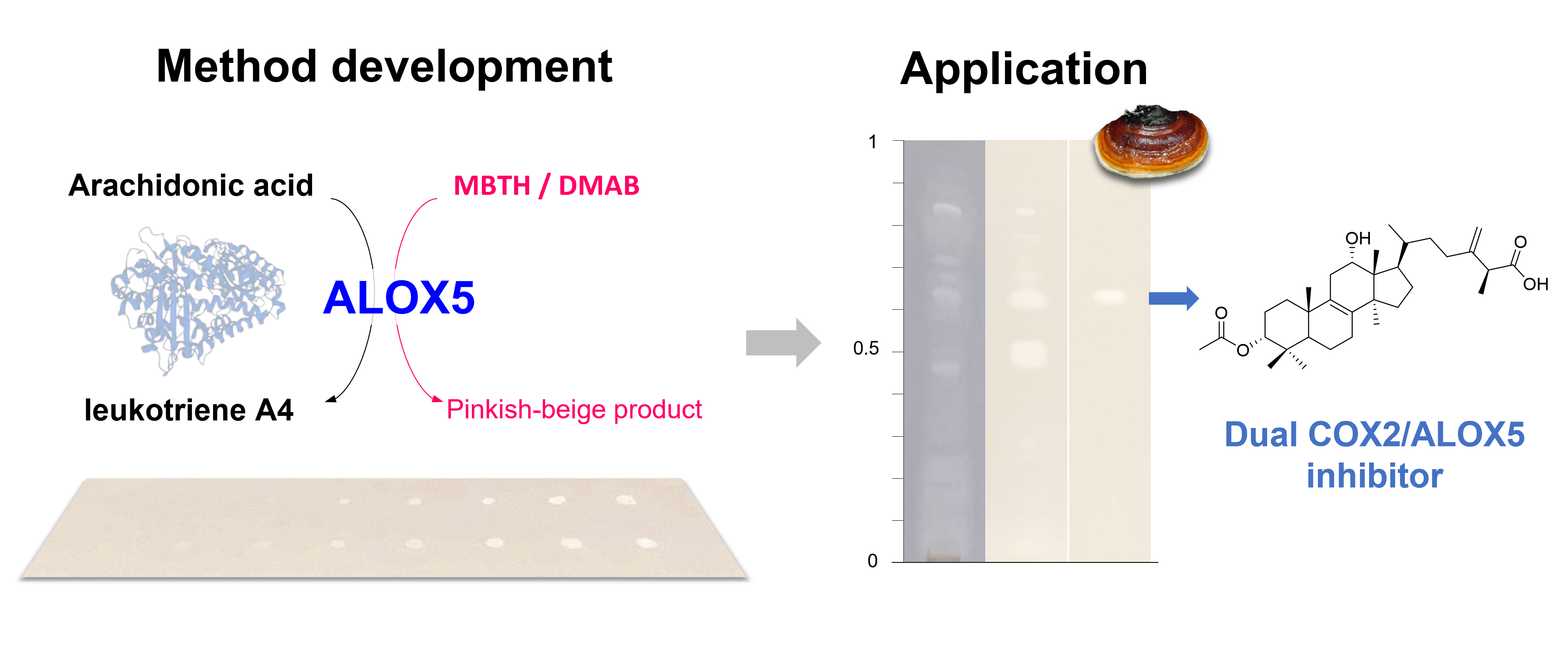
Arachidonate 5-lipoxygenase (ALOX5) is a key enzyme implicated in several inflammatory disorders, including asthma and allergic rhinitis. Despite its therapeutic importance, only one compound is currently approved as an ALOX5 inhibitor in the United States, highlighting the urgent need for new drug candidates. Progress in this area is often hindered by conventional bioassays, which can be labor-intensive, costly, and unsuitable for complex mixtures. To overcome these challenges, we developed a simple thin-layer chromatography (TLC) bioautographic assay for the rapid detection of ALOX5 inhibitors in natural extracts, a rich source of pharmacologically active compounds. The method exploits the oxidative coupling of 3-methyl-2-benzothiazolinone hydrazone (MBTH) with 3-(dimethylamino)benzoic acid (DMAB) during the ALOX5-catalyzed conversion of arachidonic acid, producing a colored indamine dye. Experimental parameters influencing chromogenic reaction were investigated and optimized to minimize reagent consumption while ensuring accuracy and sensitivity of the method. The assay was then applied to a panel of natural products and to crude mushroom extracts, enabling the rapid identification of several active compounds within complex extracts, including the dual COX2/ALOX5 inhibitor 3α-acetylpolyporenic acid A. Easy to implement, cost-efficient, and well suited for screening and bioguided fractionation, this TLC bioassay provides a powerful tool to accelerate the discovery of novel anti-inflammatory compounds.
Keywords:
thin-layer chromatography
; arachidonate 5-lipoxygenase
; bioautography
; enzyme inhibition
; anti-inflammatory drugs
; dual COX/LOX inhibitor
1. Introduction
Inflammation is a multifaceted process mediated by several enzymatic pathways, among which cyclooxygenases (COX) and lipoxygenases (LOX) play central roles. In particular, arachidonate 5-lipoxygenase (ALOX5) contributes critically to inflammatory, allergic, and other immune-related disorders through the conversion of arachidonic acid into leukotrienes [1]. While numerous COX inhibitors are clinically available, therapeutic options targeting the ALOX5 pathway remain limited and are primarily indicated for the prophylaxis and long-term management of asthma. These include zileuton, an orally active ALOX5 inhibitor, and leukotriene receptor antagonists such as montelukast, although the latter has been linked to potential neuropsychiatric adverse effects [2]. Zileuton itself may cause headaches, nausea, diarrhea, or sinusitis. Another ALOX5 inhibitor often used as positive control in bioassays, the plant-derived lignan nordihydroguaiaretic acid (NDGA), has not been approved as a drug due to toxicity concerns and insufficient clinical evidence. Recently, ALOX5 has been also identified as a promising target against cancer and age-related brain disorders [3,4]. The broad clinical relevance of ALOX5, coupled with the limited availability of safe and effective inhibitors, underscores the urgent need to identify and develop novel drugs with improved safety profiles.
Current assays for ALOX5 inhibition primarily rely on quantification of metabolic products using HPLC-MS or ELISA, or employ chromogenic or fluorogenic agents that yield a measurable change in color or fluorescence during enzymatic reactions [5,6,7,8]. However, these methods are frequently tedious, expensive, and poorly suited for the evaluation of complex natural extracts, as they fail to provide insights into the activity of individual constituents. Nevertheless, numerous natural products possess significant ALOX5-inhibitory potential [9,10]. In a previous work, we developed a rapid, cost-effective and sensitive TLC assay for detecting COX-2 inhibitors in complex extracts, based on a chromogenic reaction [11]. Building on this approach, we present here an enzyme-based TLC assay designed to detect ALOX5 inhibitors. Several chromogenic reagents have been reported for monitoring lipoxygenase activity in microplate assays, including: xylenol orange, used in the ferrous-oxidized xylenol orange (FOX) assay, which forms a pink-purple complex under acidic conditions [12]; a colorless precursor that yields methylene blue in the presence of hemoglobin [13]; and the combination of 3-methyl-2-benzothiazolinone hydrazone (MBTH) with 3-(dimethylamino)benzoic acid (DMAB), which produces a colored indamine dye [14,15]. Based on a preliminary testing, we adopted the MBTH-DMAB reagent to develop a TLC assay to detect rapidly ALOX5 inhibitors.
2. Materials and Methods
2.1. Chemicals and Reagents
Recombinant human 5-lipoxygenase (ALOX5), arachidonic acid (AA), 3-methyl-2-benzothiazolinone (MBTH), 3-(dimethylamino)benzoic acid (DMAB), Tris-HCl, calcium chloride (CaCl2), adenosine triphosphate (ATP) disodium salt, bovine hemoglobin, and hematin porcine, were all purchased from Sigma-Aldrich. Ethanol, acetonitrile, and methyl tert-butyl ether were obtained from Fisher Scientific, tetrahydrofuran from VWR, ethyl acetate from Carlo Erba, and cyclohexane from Sigma-Aldrich. Zileuton, nordihydroguaiaretic acid (NDGA), piperine, uvaol, caffeine, salicylic acid, and paracetamol were also purchased from Sigma-Aldrich. Betulin was from Carl Roth. The other standard natural compounds were purchased either from Sigma-Aldrich or Extrasynthese.
2.2. Mushroom Material
Mushroom samples were collected from natural habitats in and around Strasbourg (Bas-Rhin, France) between 2018 and 2024. The species Xylaria polymorpha, Fomes fomentarius, Fomitopsis pinicola, and Fomitopsis betulina were identified using a combination of macroscopic examination and DNA-based analyses. An additional specimen belonging to the genus Hebeloma was included in the study; however, its species could not be conclusively determined using macroscopic, microscopic, or molecular sequencing approaches. The fruiting bodies were freeze-dried using a Labconco Freezone 4.5 L freeze dryer, ground into a fine powder, and extracted with ethyl acetate via ultrasonication with a ratio of 25 mL of solvent per gram of dry matter. The resulting extracts were concentrated under reduced pressure to remove the solvent and stored at 4 °C. A voucher specimen of each species is deposited at the Laboratory CAMBAP, University of Strasbourg.
2.3. Purification and Identification of 3α-Acetylpolyporenic Acid A
A finely powdered sample of Fomitopsis betulina (37.5g) was successively extracted by sonication during 30 min with n-heptane and ethyl acetate at room temperature (ratio of 10 mL per gram). A portion (8.2 g) of the dry ethyl acetate extract was subjected to solid phase extraction (SPE) using a 60 mL Chromabond cartridge loaded with silica gel (15g) with mixtures of acetonitrile (ACN) and water of decreasing polarity: 100%, 80%, 60%, and 40% of ACN. The fraction obtained with 80% ACN was purified by preparative HPLC using a C18 Kinetex Axia preparative column (5 µm, 150 × 21.2 mm, Phenomenex) and a Gilson 322 pump delivering the mobile phases A (H₂O + 0.1% formic acid) and B (ACN + 0.1% formic acid) at a flow rate of 30 mL/min. A sample at a concentration of 20 mg/ml was loaded via a valve equipped with a 2.5 mL loop, and eluted with a gradient from 15% to 85% of solvent B in 12 min, then kept at 85% B to 21 min. The fractions were collected using a Gilson FC 204 fraction collector, and combined on the basis of their TLC and analytical HPLC profiles. The fraction eluted at approximately 13.1 min yielded a product identified as 3α-acetylpolyporenic acid A, by comparing its spectral data with those in the literature [16].
2.4. Preparation of Solutions
ALOX5 was prepared in 100 mM Tris-HCl buffer (pH 7.5) to a final concentration of 2 U/mL, supplemented with 5 µM CaCl₂. Stock solutions of arachidonic acid (AA), DMAB, and MBTH were prepared in ethanol. Immediately before use, the chromogenic mixture was prepared by combining these stock solutions in Tris-HCl buffer to yield final concentrations of 20 µM AA, 0.1 mM MBTH, and 0.1 mM DMAB. To evaluate their influence on the chromogenic reaction, hematin or hemoglobin was added to the reaction mixture at a final concentration of 100 µM, while ATP (1 µM) was added directly to the ALOX5 enzyme solution. Standard compounds were dissolved in ethanol and mushroom extracts were resuspended in ethyl acetate at appropriate concentrations for TLC analysis.
2.5. Thin-Layer Chromatography
Experiments were performed on pre-washed (MeOH) aluminum TLC silica gel plates 60 F254 Merck. Appropriate volumes of standard compound solutions were applied as dot spots on the TLC plate without any elution. Ethyl acetate crude extracts of mushrooms (50 µg) were deposited onto TLC plates as bands of 8 mm in length, with a Camag Linomat 5 equipped with a 100 μL Hamilton syringe under a stream of nitrogen. The plate was then developed over 8 cm with methyl tert-butyl ether/tetrahydrofurane/cyclohexane (5:1:4 v/v/v).
2.6. Arachidonate 5-Lipoxygenase Inhibition Assay
After thorough drying to remove any residual solvent, the TLC plates (10 × 20 cm) were uniformly sprayed with 2 mL of ALOX5 solution using a Camag Derivatizer. The plates were then placed in a Petri dish lined with moist filter paper and incubated at 37 °C in a Binder KB 23 incubator. Following a 5-minute incubation, the chromogenic reaction was initiated by spraying 2 mL of arachidonic acid/MBTH/DMAB solution with the Derivatizer. After an additional 5 minutes, images of the TLC plates were captured under visible light using a Camag Reprostar 3.
2.7. Data Processing
To obtain quantitative values during method development and assess the influence of experimental parameters on the contrast between inhibition zones and the colored background, plate images were analyzed using ImageJ® 1.53k. Photographs were first converted to 32-bit grayscale to standardize intensity measurements. Background homogeneity was evaluated, and the contrast Cw was calculated using Weber-Fechner’s law (1) defined as:
where Linhib and Lback refer to the luminous intensity (in candelas) measured within the inhibition zone and in the surrounding background, respectively.
2.8. Optimization of Reagents Concentrations and Statistical Analyses
To evaluate the effects of enzyme, substrate, and chromogenic reagent concentrations on the contrast obtained with ALOX5 inhibitors (zileuton, NDGA, and piperine), a two-level factorial design was implemented, resulting in eight experimental conditions, each performed in triplicate. The tested factors and levels were: ALOX5 concentration (0.5 and 2 U/mL), arachidonic acid concentration (0.02 and 1 mM), and both DMAB and MBTH concentrations (0.1 and 1 mM), with the MBTH:DMAB ratio fixed at 1:1. Contrast values were analyzed using descriptive statistics, with a significance threshold of α = 0.05, in MATLAB R2023b. Analysis of variance (ANOVA) was applied to assess the overall effect of experimental conditions, followed by Tukey’s HSD post hoc test to identify pairwise differences between conditions.
2.9. Sensitivity and Linearity
The linearity and limit of detection (LOD) were assessed by applying decreasing amounts of inhibitors (ranging from 0.01 to 100 µg) using a Linomat 5 (Camag). Bioassays were conducted in triplicate without chromatographic elution. Regression equations were derived for each inhibitor, and the visual LOD were determined by independent observers.
3. Results and Discussion
3.1. Development and Validation of the TLC-based Enzyme Assay
3.1.1. Specificity and Selectivity
In order to assess the suitability of the method for TLC bioautography, preliminary tests were performed in the absence of inhibitors to evaluate color development on silica gel plates and identify any potential interactions. The initial concentrations of enzyme and reagents were adapted from literature-reported conditions used in microplate assays, as follows: ALOX5 2 U/mL, AA 20 µM, MBTH 1 mM, and DMAB 10 mM. During the enzymatic reaction, MBTH was oxidized to an intermediate that condenses with DMAB to form an indamine dye (Figure 1).
Although this dye should exhibit an intense blue coloration in acidic aqueous solution, incubation on TLC plates resulted in a pinkish-beige coloration. This shift can be attributed to the markedly different environment on silica, where interactions with surface silanol groups and increased exposure to air could alter chromophore stability. While coloration was observed across all plates, the intensity was markedly higher on the plate treated with the complete reaction mixture. These findings confirm the feasibility of the bioautographic approach.
Selectivity was subsequently evaluated using zileuton, NDGA, piperine, caffeine, and salicylic acid. Inhibition of ALOX5 by piperine has been previously reported with an IC50 value of 54 μM [17], whereas only a very weak effect was documented for salicylic acid (IC50 value of 168 μM) [18]; the lack of reported data for caffeine may suggest an absence of inhibitory activity. As expected, zileuton, NDGA, and piperine yielded distinct clear spots against the colored background, whereas caffeine and salicylic acid showed no effect. Zileuton and piperine produced uniform clear spots, while NDGA generated a halo-shaped spot with a darker core (Figure 2).
3.1.2. Optimization of Experimental Parameters
- Heme cofactors
Our assay is based on the work of Anthon and Barrett used for plant linoleate lipoxygenase [15], which itself builds upon the peroxidase assay described by Ngo and Lenhoff [14]. Anthon and Barrett added a heme cofactor (hemoglobin) in their chromogenic reaction, acting as a peroxidase-type catalyst, thus accelerating the oxidation of MBTH by lipid peroxide. We decided to run experiment adding hemoglobin or hematin (100 µM) to the reaction mixture and without any heme factor. Experiments using hemoglobin and hematin were not conclusive. Nebulization of hemoglobin in the derivatizer resulted in excessive foaming which prevented uniform application onto the TLC plate. Although background staining was slightly increased in the presence of heme cofactors, the intrinsic coloration of both hemoglobin and hematin posed a significant limitation: it obscured the inhibition zones, thereby substantially diminishing their contrast and complicating result interpretation. Subsequent experiments were therefore carried out without any heme compounds.
- Incubation conditions
Regarding incubation, conditions were set at 37 °C in moist atmosphere, with a pH of 7.5 (100 mM Tris-HCl buffer) for an optimized enzyme activity. During optimization steps, coloration and contrast were evaluated after 5, 10, 20, 30, 40, and 60 minutes of incubation, once the reaction mixture is applied. Five minutes of incubation were found to be sufficient to clearly detect inhibition zone, with the highest contrast obtained between 5 and 10 minutes, and the coloration was relatively stable during 40 minutes (it has to be noted that the inhibition zone of NDGA turned darker than the background after 25 minutes).
- ATP and calcium
In vivo, the activity of ALOX5 is critically dependent on calcium ions and ATP: Ca²⁺ facilitates the enzyme translocation to the nuclear membrane while ATP stabilizes its active conformation [19]. Although these cofactors are not strictly required for assays using purified enzyme, some studies include ATP and Ca²⁺ in their in vitro assays. We investigated the influence of these two factors in our TLC bioassay and found that coloration – and thus contrast – was slightly enhanced in the presence of Ca²⁺ (5 µM) in the enzyme solution, whereas ATP had no observable effect.
- Reagent concentrations
Our optimization approach focused on three key parameters: enzyme concentration, substrate concentration, and chromogenic reagent concentration, as well as their respective ratios. According to Ngo and Lenhoff [14], a DMAB:MBTH ratio of 50:1 yields the most intense staining in a 96-well plate format, though this was established for fluorescence detection. In our bioassay, which is read under visible light, we determined that a DMAB:MBTH ratio of 1:1 produced more intense staining. Then, we employed a two-level factorial design, generating eight experimental conditions, each performed in triplicate. The tested factors and levels were as follows: ALOX5 concentration (0.5 and 2 U/mL), arachidonic acid concentration (0.02 and 1 mM), and DMAB and MBTH concentrations (0.1 and 1 mM), with the MBTH:DMAB ratio fixed at 1:1. Statistical analysis revealed that, while one-way ANOVA did not indicate a significant overall difference (p = 0.083), Tukey’s multiple comparisons test identified certain conditions as more favorable. These conditions represented the best compromise among contrast, detection limit, and linearity, while minimizing the diffusion halo observed for NDGA. Results were validated through contrast quantification using ImageJ software.
- Final Protocol
The optimized protocol involves pre-incubation of the developed TLC plate at 37 °C for 5 minutes in a moist atmosphere with 100 mM Tris-HCl buffer (pH 7.5) containing 2 U/mL of 5-lipoxygenase and 5 µM CaCl₂. The reaction is initiated by spraying a reaction mixture containing 20 µM arachidonic acid, 0.1 mM MBTH, and 0.1 mM DMAB, followed by an additional 5-minute incubation.
3.1.3. Limit of Detection and Linear Regression
The visual limit of detection (LOD) was determined for five compounds: zileuton, NDGA, piperine, uvaol, and betulin (Table 1). Zileuton exhibited a detectable inhibition zone at quantities as low as 1 µg, whereas the LOD of both nordihydroguaiaretic acid (NDGA) and piperine was determined at 0.01 µg. The limit of detection for the triterpene derivatives betulin and uvaol was 0.05 µg and 0.5 µg, respectively. These findings demonstrate that ALOX5 inhibitors can be visually detected below the microgram level according to their inhibitory potency.
Regression analyses were performed for each inhibitor, yielding good coefficients of determination (R² = 0.98 for zileuton, piperine and betulin). Linear relationships were observed for zileuton, betulin, and uvaol, whereas NDGA and piperine exhibited logarithmic regression profiles (Table 1). The nonlinear responses may be attributed to diffusion effects or, in the case of piperine, to interference from its yellow coloration during luminance measurements.
3.2. Application to Natural Products
3.2.1. Application to Pure Compounds
A total of 40 natural products were evaluated for their ability to inhibit ALOX5 using the developed bioassay. The compounds were selected to represent a broad range of metabolite classes, including 13 phenolic compounds, 19 terpene and sterol derivatives, 4 alkaloids, 2 vitamins (ascorbic acid and α-tocopherol), caffeine, and sinigrin. Zileuton and NDGA were used as positive controls while paracetamol, known to have no effect on ALOX5 activity, served as negative control. Each compound (10 µg) was spotted onto the TLC plate without elution. After incubation, 21 of the 40 compounds induced inhibition zones (Table 2), more or less clearly defined (Figure A1); these slight diffusion effects – not disturbing for a screening purpose – could be due to the physicochemical properties of compounds, affecting interactions with both the silica phase and the enzyme/reagents solutions.
Steroid and triterpenoid derivatives produced clearly demarcated inhibition spots, with triterpenoids exhibiting particularly sharp and well-defined outlines. The activities observed for uvaol, madecassic acid, and α-hederin, are in line with previous reports: inhibition of human ALOX5 by oleanane- and ursane-type triterpenoids has been described, predominantly for boswellic acid derivatives, and for uvaol, with an IC50 of 42 µM [20,21]. In our bioassay, lupeol, betulin, and ginsenoside Rd also produced detectable inhibition zones. Although in vitro inhibition of ALOX5 by lupane-type triterpenes has not been demonstrated, inhibition of other lipoxygenase isoforms suggests that the lupane scaffold can interact with lipoxygenase active sites, as exemplified by the activity of lupeol against soybean 15-LOX (IC50 35 µM) [22]. Furthermore, the inhibitory activity of ginsenoside Rd was recently predicted using machine learning models [23]. This computational approach also unveiled the inhibitory potential of gitoxin, a cardenolide closely-related to digitoxin, found to be active in our TLC bioassay [23]. Stigmasterol, β-sitosterol, and the steroidal glycoalkaloid tomatine also generated an inhibition zone on the plate, although no inhibition of ALOX5 activity by phytosterols has been reported to date. Alkaloids were also active, with piperine demonstrating the most pronounced inhibitory effect. While the activity of piperine and its derivative against ALOX5 is well known [17,24,25], information about the LOX inhibitory effect of quinine, senecionine, or lobeline is scanty.
Regarding mono- and sesquiterpenes, they yielded no to poorly defined inhibition zones, that might be attributable to their volatile nature and evaporation during incubations. Carvacrol gave a large diffuse inhibition zone, in accordance with literature [26]. No inhibition was observed for ascorbic acid and α-tocopherol; given that vitamins C and E are recognized as potent antioxidants, these findings further substantiate that antioxidant activity does not yield false-positive results in this assay, despite the oxidative nature of the enzymatic reaction.
Although most of our results are consistent with the literature, this was not the case for phenolic compounds, for which no inhibitory effect was observed. Because the yellow coloration of flavonoid derivatives could potentially interfere with the bioassay, luminance data were analyzed. These data showed a slight shift following the enzymatic reaction, indicating the formation of the colored reaction product and thereby confirming the absence of inhibition of ALOX5 activity. This finding contrasts with several reports describing ALOX5 inhibitory activity for polyphenols [27,28]. The discrepancy may be explained by the low amounts applied to the TLC plate, which were well below the reported IC₅₀ values. For example, 10 µg of kaempferol corresponds to approximately 35 nmol, whereas its IC₅₀ has been reported to be 20 µM [29].
3.2.2. Application to Mushroom Extracts
The TLC bioautographic assay was subsequently applied to complex crude natural extracts from mushrooms: 50 µg of ethyl acetate extracts from Hebeloma sp., Xylaria polymorpha, Fomes fomentarius, Fomitopsis pinicola, and Fomitopsis betulina were applied onto a silica TLC plate and eluted with an appropriate solvent system. After thorough drying to remove residual solvent, the bioassay was initiated. Following incubation, the plate developed a uniform pinkish-beige background, against which distinct inhibition zones were observed in all extracts except that of F. fomentarius (Figure 3). A single inhibition zone was detected for X. polymorpha (Rf 0.86), while the Hebeloma extract exhibited two zones (Rf 0.04 and 0.87). Extracts from Fomitopsis species displayed the highest diversity of ALOX5 inhibitors, with F. pinicola producing seven distinct inhibition zones most likely corresponding to at least seven different active compounds within the crude extract.
Interestingly, we previously demonstrated the presence of COX-2 inhibitors in F. pinicola, using a TLC-based enzymatic assay developed by our group [11]. Comparison of the COX-2 and ALOX5 bioautograms revealed that several compounds act as dual COX/LOX inhibitors (Figure 4), a feature considered promising for the development of safer drug candidates [30,31,32]. This rapid insight was enabled by the chromatographic separation inherent to bioautography, which allows direct comparison of multiple biological activities without prior purification, thereby substantially accelerating the identification of compounds with complementary properties. One of these compounds was purified and identified as 3α-acetylpolyporenic acid A, a lanostane-type triterpenoid previously isolated from F. betulina, known to inhibit COX-1 and 3α-hydroxysteroid dehydrogenase [16].
The other dual COX-2/ALOX5 inhibitors detected in Fomitopsis species have also been preliminarily characterized as triterpenoid derivatives, and more in-depth studies of their anti-inflammatory activity are currently underway. These results underline the anti-inflammatory potential of macromycetes metabolites [33].
4. Conclusions
The developed ALOX5 TLC bioautographic assay demonstrates high sensitivity, enabling the detection of biologically active compounds even at low concentrations in crude extracts. In contrast to commercially expensive kits, this method offers several key advantages: it provides early insight into both the number and physicochemical nature of active constituents prior to fractionation, and it minimizes the influence of synergistic or antagonistic interactions that may obscure individual activities and lead to false-negative outcomes [34]. The assay is simple to implement, cost-effective, and well suited for high-throughput screening as well as for bioguided fractionation monitoring. Furthermore, its compatibility with complementary bioautographic assays allows the parallel identification of compounds acting through multiple mechanisms, thereby facilitating and accelerating the discovery of novel anti-inflammatory agents, as illustrated in this study with the rapid identification in a crude natural extract of a potential dual COX/LOX inhibitor.
Author Contributions
Conceptualization, AU; methodology, AU; validation, AU and YP; formal analysis, YP, MM and AU; investigation, YP, MM and AU; data curation, AU, YS, LB, CC and VB; writing-review and editing, AU, YS, LB, CC, VB and YP; supervision, AU; project administration, AU, LB, CC and VB; funding acquisition, VB. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
This work was supported by PiLeJe. We thank Antoine Mamet, training student, for his contribution to this project, Elodie Lepeltier for preliminary research and assays, and Claude Blondeau (PiLeJe) for proofreading.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ALOX5 | Arachidonate 5-lipoxygenase |
| COX | Cyclooxygenase |
| TLC | Thin-layer chromatography |
| HPLC | High-performance liquid chromatography |
| ELISA | enzyme-linked immunosorbent assay |
| DMAB | 3-(dimethylamino)benzoic acid |
| MBTH | 3-methyl-2-benzothiazolinone hydrazone |
| NDGA | Nordihydroguaiaretic acid |
| FOX | Ferrous-oxidized xylenol orange assay |
| EtOAc | Ethyl acetate |
Appendix A
Figure A1.
ALOX5 TLC bioassay of standard natural products (spotted at 10 µg).

References
- Haeggström, J.Z.; Funk, C.D. Lipoxygenase and Leukotriene Pathways: Biochemistry, Biology, and Roles in Disease. Chem. Rev. 2011, 111, 5866–5898. [Google Scholar] [CrossRef]
- Paljarvi, T.; Forton, J.; Luciano, S.; Herttua, K.; Fazel, S. Analysis of Neuropsychiatric Diagnoses after Montelukast Initiation. JAMA Netw. Open 2022, 5, e2213643. [Google Scholar] [CrossRef]
- Vishnupriya, P.; Aparna, A.; Viswanadha, V.P. Lipoxygenase (LOX) Pathway: A Promising Target to Combat Cancer. Curr. Pharm. Design 2021, 27, 3349–3369. [Google Scholar] [CrossRef]
- Yan, M.; Zhang, S.; Li, C.; Liu, Y.; Zhao, J.; Wang, Y.; Yang, Y.; Zhang, L. 5-Lipoxygenase as an Emerging Target against Age-Related Brain Disorders. Ageing Res. Rev. 2021, 69, 101359. [Google Scholar] [CrossRef] [PubMed]
- Pufahl, R.A.; Kasten, T.P.; Hills, R.; Gierse, J.K.; Reitz, B.A.; Weinberg, R.A.; Masferrer, J.L. Development of a Fluorescence-Based Enzyme Assay of Human 5-Lipoxygenase. Anal. Biochem. 2007, 364, 204–212. [Google Scholar] [CrossRef]
- Nair, D.G.; Funk, C.D. A Cell-Based Assay for Screening Lipoxygenase Inhibitors. Prostaglandins Other Lipid Mediat. 2009, 90, 98–104. [Google Scholar] [CrossRef]
- Lu, W.; Zhao, X.; Xu, Z.; Dong, N.; Zou, S.; Shen, X.; Huang, J. Development of a New Colorimetric Assay for Lipoxygenase Activity. Anal. Biochem. 2013, 441, 162–168. [Google Scholar] [CrossRef]
- Huang, X.; Li, Y.; Sabier, M.; Si, J.; Wang, P.; Shen, Y.; Zhang, X.; Liu, J. Guidelines for the in Vitro Determination of Anti-Inflammatory Activity. eFood 2024, 5, e160. [Google Scholar] [CrossRef]
- Schneider, I.; Bucar, F. Lipoxygenase Inhibitors from Natural Plant Sources. Part 1: Medicinal Plants with Inhibitory Activity on Arachidonate 5-Lipoxygenase and 5-Lipoxygenase/Cyclooxygenase. Phytother Res 2005, 19, 81–102. [Google Scholar] [CrossRef]
- Werz, O. Inhibition of 5-Lipoxygenase Product Synthesis by Natural Compounds of Plant Origin. Planta Med 2007, 73, 1331–1357. [Google Scholar] [CrossRef] [PubMed]
- Urbain, A.; Trabelssi, N.; Bardot, V. Development of an Enzyme-Based Thin-Layer Chromatographic Assay for the Detection of Cyclooxygenase-2 Inhibitors. Separations 2022, 9, 238. [Google Scholar] [CrossRef]
- Cho, Y.S.; Kim, H.S.; Kim, C.H.; Cheon, H.G. Application of the Ferrous Oxidation-Xylenol Orange Assay for the Screening of 5-Lipoxygenase Inhibitors. Anal. Biochem. 2006, 351, 62–68. [Google Scholar] [CrossRef]
- Auerbach, B.; Kiely, J.; Cornicelli, J. A Spectrophotometric Microtiter-Based Assay for the Detection of Hydroperoxy Derivatives of Linoleic-Acid. Anal. Biochem. 1992, 201, 375–380. [Google Scholar] [CrossRef]
- Ngo, T.T.; Lenhoff, H.M. A Sensitive and Versatile Chromogenic Assay for Peroxidase and Peroxidase-Coupled Reactions. Anal. Biochem. 1980, 105, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Anthon, G.E.; Barrett, D.M. Colorimetric Method for the Determination of Lipoxygenase Activity. J Agric Food Chem 2001, 49, 32–37. [Google Scholar] [CrossRef]
- Wangun, H.V.K.; Berg, A.; Hertel, W.; Nkengfack, A.E.; Hertweck, C. Anti-Inflammatory and Anti-Hyaluronate Lyase Activities of Lanostanoids from Piptoporus betulinus. J Antibiot (Tokyo) 2004, 57, 755–758. [Google Scholar] [CrossRef]
- Prasad, N.S.; Raghavendra, R.; Lokesh, B.R.; Naidu, K.A. Spice Phenolics Inhibit Human PMNL 5-Lipoxygenase. Prostaglandins Leukot Essent Fatty Acids 2004, 70, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Lapenna, D.; Ciofani, G.; Pierdomenico, S.D.; Neri, M.; Cuccurullo, C.; Giamberardino, M.A.; Cuccurullo, F. Inhibitory Activity of Salicylic Acid on Lipoxygenase-Dependent Lipid Peroxidation. Biochim. Biophys. Acta 2009, 1790, 25–30. [Google Scholar] [CrossRef]
- Häfner, A.-K.; Kahnt, A.S.; Steinhilber, D. Beyond Leukotriene Formation—The Noncanonical Functions of 5-Lipoxygenase. Prostaglandins Other Lipid Mediat. 2019, 142, 24–32. [Google Scholar] [CrossRef]
- Vo, N.N.Q.; Nomura, Y.; Muranaka, T.; Fukushima, E.O. Structure–Activity Relationships of Pentacyclic Triterpenoids as Inhibitors of Cyclooxygenase and Lipoxygenase Enzymes. J. Nat. Prod. 2019, 82, 3311–3320. [Google Scholar] [CrossRef]
- Gilbert, N.C.; Gerstmeier, J.; Schexnaydre, E.E.; Börner, F.; Garscha, U.; Neau, D.B.; Werz, O.; Newcomer, M.E. Structural and Mechanistic Insights into 5-Lipoxygenase Inhibition by Natural Products. Nat. Chem. Biol. 2020, 16, 783–790. [Google Scholar] [CrossRef]
- Gutierrez-Lugo, M.-T.; Deschamps, J.D.; Holman, T.R.; Suarez, E.; Timmermann, B.N. Lipoxygenase Inhibition by Anadanthoflavone, a New Flavonoid from the Aerial Parts of Anadenanthera colubrina. Planta Med. 2004, 70, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Mikutis, S.; Lawrinowitz, S.; Kretzer, C.; Dunsmore, L.; Sketeris, L.; Rodrigues, T.; Werz, O.; Bernardes, G.J.L. Machine Learning Uncovers Natural Product Modulators of the 5-Lipoxygenase Pathway and Facilitates the Elucidation of Their Biological Mechanisms. ACS Chem Biol 2023, 19, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Tomy, M.J.; Sharanya, C.S.; Dileep, K.V.; Prasanth, S.; Sabu, A.; Sadasivan, C.; Haridas, M. Derivatives Form Better Lipoxygenase Inhibitors than Piperine: In Vitro and in Silico Study. Chem Biol Drug Des 2015, 85, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Stöhr, J.R.; Xiao, P.-G.; Bauer, R. Constituents of Chinese Piper Species and Their Inhibitory Activity on Prostaglandin and Leukotriene Biosynthesis in Vitro. J Ethnopharmacol 2001, 75, 133–139. [Google Scholar] [CrossRef]
- Fatima, K.; Luqman, S.; Meena, A. Carvacrol Arrests the Proliferation of Hypopharyngeal Carcinoma Cells by Suppressing Ornithine Decarboxylase and Hyaluronidase Activities. Front. Nutr. 2022, 9. [Google Scholar] [CrossRef]
- Chen, S. Natural Products Triggering Biological Targets- A Review of the Anti-Inflammatory Phytochemicals Targeting the Arachidonic Acid Pathway in Allergy Asthma and Rheumatoid Arthritis. Curr. Drug Targets 2010, 12, 288–301. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; González-Sarrías, A.; Laparra-Llopis, J.M.; Schneider, C.; Espín, J.C. Targeting Mammalian 5-Lipoxygenase by Dietary Phenolics as an Anti-Inflammatory Mechanism: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 7937. [Google Scholar] [CrossRef]
- Laughton, M.J.; Evans, P.J.; Moroney, M.A.; Hoult, J.R.S.; Halliwell, B. Inhibition of Mammalian 5-Lipoxygenase and Cyclo-Oxygenase by Flavonoids and Phenolic Dietary Additives: Relationship to Antioxidant Activity and to Iron Ion-Reducing Ability. Biochem. Pharmacol. 1991, 42, 1673–1681. [Google Scholar] [CrossRef]
- Mukhopadhyay, N.; Shukla, A.; Makhal, P.N.; Kaki, V.R. Natural Product-Driven Dual COX-LOX Inhibitors: Overview of Recent Studies on the Development of Novel Anti-Inflammatory Agents. Heliyon 2023, 9, e14569. [Google Scholar] [CrossRef]
- Agrawal, N. A Comprehensive Review on the Advancements of Dual COX-2/5-LOX Inhibitors as Anti-Inflammatory Drugs. Chem. Biol. Drug Des. 2025, 105, e70114. [Google Scholar] [CrossRef]
- Chatzipieris, F.P.; Petsas, E.; Lambrinidis, G.; Vassiliou, S.; Chasapis, C.T. Recent Advances in Dual COX/LOX Inhibitor Design (2020-2024): Establishing “The Rule of Four for Inflammation.”. Life-Basel 2026, 16, 163. [Google Scholar] [CrossRef]
- Taofiq, O.; Martins, A.; Barreiro, M.F.; Ferreira, I.C.F.R. Anti-Inflammatory Potential of Mushroom Extracts and Isolated Metabolites. Trends Food Sci. Technol. 2016, 50, 193–210. [Google Scholar] [CrossRef]
- Urbain, A.; Simões-Pires, C.A. Thin-Layer Chromatography for the Detection and Analysis of Bioactive Natural Products. In Encyclopedia of Analytical Chemistry; Meyers, R.A., Ed.; Wiley, 2020; pp. 1–29. ISBN 978-0-471-97670-7. [Google Scholar]
Figure 1.
Chromogenic reaction of MBTH with DMAB during ALOX5 conversion of arachidonic acid into leukotriene A4.
Figure 1.
Chromogenic reaction of MBTH with DMAB during ALOX5 conversion of arachidonic acid into leukotriene A4.

Figure 2.
Preliminary ALOX5 TLC assay with 10 µg of (a) zileuton, (b) NDGA, and (c) piperine, used as positive controls, and (d) caffeine and (e) salicylic acid, used as negative controls.
Figure 2.
Preliminary ALOX5 TLC assay with 10 µg of (a) zileuton, (b) NDGA, and (c) piperine, used as positive controls, and (d) caffeine and (e) salicylic acid, used as negative controls.

Figure 3.
TLC of mushroom extracts (50 µg); (a) after ALOX5 bioassay under visible light; (b): prior to bioassay at 254 nm; (1) Hebeloma sp., (2) Xylaria polymorpha, (3) Fomes fomentarius, (4) Fomitopsis pinicola, (5) Fomitopsis betulina.
Figure 3.
TLC of mushroom extracts (50 µg); (a) after ALOX5 bioassay under visible light; (b): prior to bioassay at 254 nm; (1) Hebeloma sp., (2) Xylaria polymorpha, (3) Fomes fomentarius, (4) Fomitopsis pinicola, (5) Fomitopsis betulina.

Figure 4.
(a) COX-2 bioautogram of F. pinicola EtOAc extract (50 µg) [11]; (b) ALOX5 bioautogram of F. pinicola EtOAc extract (50 µg); (c) ALOX5 bioautogram of 3α-acetylpolyporenic acid A (1 µg).
Figure 4.
(a) COX-2 bioautogram of F. pinicola EtOAc extract (50 µg) [11]; (b) ALOX5 bioautogram of F. pinicola EtOAc extract (50 µg); (c) ALOX5 bioautogram of 3α-acetylpolyporenic acid A (1 µg).


Table 1.
Limit of detection (LOD), regression equation, and correlation coefficient of five ALOX5 inhibitors.
Table 1.
Limit of detection (LOD), regression equation, and correlation coefficient of five ALOX5 inhibitors.
| Compound | LOD (µg) | LOD (nmol) | Regression equation | R2 |
|---|---|---|---|---|
| zileuton | 1.00 | 4.23 | y = 2.4175x + 232.75 | 0.98 |
| NDGA | 0.01 | 3.31 | y = - 15.64ln(x) + 140.72 | 0.97 |
| piperine | 0.01 | 1.65 | y = - 10.01ln(x) + 176.94 | 0.98 |
| uvaol | 0.50 | 1.13 | y = 0.3882x + 241.03 | 0.91 |
| betulin | 0.05 | 0.226 | y = 0.0194x + 241.53 | 0.98 |
Table 2.
Evaluation of ALOX5 inhibitory activity of selected natural products using the developed TLC bioassay.
Table 2.
Evaluation of ALOX5 inhibitory activity of selected natural products using the developed TLC bioassay.
| Compound | ALOX5 inhibition a |
Compound | ALOX5 inhibition a |
|
|---|---|---|---|---|
| NDGA | + | lobeline | ± | |
| betulin | + | caffeine | - | |
| digitoxin | + | anethol | - | |
| ginsenoside Rd | + | eucalyptol | - | |
| α-hederin | + | menthol | - | |
| lupeol | + | α-tocopherol | - | |
| madecassic acid | + | ascorbic acid | - | |
| piperine | + | epigallocatechin | - | |
| quinine | + | kampferol | - | |
| senecionin | + | naringin | - | |
| β-sitosterol | + | protocatechuic acid | - | |
| stigmasterol | + | quercetin | - | |
| tomatine | + | quercetol | - | |
| uvaol | + | resveratrol | - | |
| borneol | ± | rutin | - | |
| carvacrol | ± | salicylic acid | - | |
| caryophyllene | ± | scopoletin | - | |
| farnesene | ± | taxifolin | - | |
| humulene | ± | tyrosol | - | |
| santonin | ± | sinigrin | - |
a inhibition was indicated by a clear visible spot on the TLC plate after bioautography for a spotted quantity of 10 µg.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



