Submitted:
24 February 2026
Posted:
02 March 2026
You are already at the latest version
Abstract
Background: Determining loss of consciousness during slaughter in calves is fundamentally a neurobiological challenge requiring distinction between cortical awareness and subcortical reflex activity. Electroencephalography (EEG) and evoked potentials directly assess cortical responsiveness, while cerebral blood flow (CBF) constitutes the upstream physiological determinant of cortical viability. Aim: To synthesize current evidence on neurologic effects of slaughter modalities in calves, integrating electrophysiological and cerebral perfusion data within a mechanistic framework relevant to welfare monitoring and policy. Methods: A structured narrative review was conducted following SANRA principles. PubMed/MEDLINE was searched from inception to January 2026 using combinations of terms related to calves, cattle, EEG, evoked potentials, cerebral blood flow, stunning, and exsanguination. Inclusion was limited to peer-reviewed, PubMed-indexed studies reporting neurophysiologic endpoints in cattle or calves. EFSA scientific opinions were consulted for contextual interpretation. Results: Effective penetrating captive bolt and appropriately applied electrical stunning produce cortical lesions compatible with rapid unconsciousness. Exsanguination without prior stunning induces rapidly progressive cerebral hypoperfusion leading to cortical failure typically within seconds, with measurable inter-individual variability. Animal-based measures (ABMs) demonstrate imperfect concordance with cortical endpoints. Integration of EEG with perfusion metrics would provide a physiologically coherent method to distinguish concussive suppression, post-ictal states, and ischemic cortical failure. Conclusions: EEG and evoked potentials remain the strongest markers of cortical unconsciousness in calves. Complementary assessment of CBF can reduce interpretive uncertainty and enhance welfare monitoring precision.
Keywords:
calf
; electroencephalography
; cerebral blood flow
; unconsciousness
; slaughter
; animal welfare
; evoked potentials
; neuroethics
1. Introduction
The determination of unconsciousness in calves during slaughter must be grounded in cortical neurobiology rather than inferred solely from observable motor behavior. Conscious perception, particularly the conscious experience of pain, requires preserved thalamocortical connectivity and functional integration across distributed cortical networks, including primary sensory, associative, and limbic regions [1,2,3]. Experimental neurophysiology in mammals consistently demonstrates that loss of evoked cortical responses and collapse of organized electroencephalographic (EEG) activity are more reliable indicators of loss of consciousness than persistence or absence of brainstem reflexes. In cattle, Daly et al. [4] showed that visually and somatosensory evoked cortical responses were immediately abolished following effective captive bolt stunning, whereas behavioral signs alone did not provide equivalent resolution of cortical state. Similarly, studies in calves undergoing exsanguination demonstrated that electrocortical suppression occurred within seconds, yet some reflexive responses could transiently persist beyond EEG-defined unconsciousness [5,6]. These findings underscore that corneal reflexes, gasping, or limb movements may be mediated at subcortical or spinal levels and therefore cannot independently define conscious awareness.
Within the European Union, slaughter is regulated under Regulation (EC) No 1099/2009, with scientific interpretation guided by the European Food Safety Authority (EFSA). EFSA opinions acknowledge that operational monitoring relies on animal-based measures (ABMs) but emphasize their limitations and the importance of combined indicators to infer unconsciousness. Validation studies in veal calves confirm imperfect concordance between ABMs and EEG-defined cortical states [7].
Calves constitute a distinct neurophysiological subgroup compared with mature cattle, given differences in cranial anatomy, cerebral perfusion patterns, and developmental maturation. Skull thickness influences mechanical energy transfer during stunning, while age-related cerebrovascular characteristics may affect the dynamics of ischemic cortical failure. Integrating electrophysiologic endpoints (EEG and evoked potentials) with cerebral perfusion metrics would provide a mechanistically coherent framework for defining loss of consciousness and clarifying modality-specific pathways to cortical failure in calves.
The finality of this research is to establish a physiologically grounded, multimodal framework integrating EEG and cerebral perfusion metrics to define loss of consciousness in calves during slaughter. By reducing epistemic uncertainty and harmonizing cortical endpoints with operational monitoring, it aims to strengthen scientific rigor, regulatory precision, and welfare assurance within the European context.
2. Methods
This research was conducted as a structured narrative review, in accordance with the Scale for the Assessment of Narrative Review Articles (SANRA) [8]. The SANRA framework was selected to ensure methodological transparency, explicit justification of the review question, systematic literature identification, critical appraisal of included studies, and coherent synthesis of heterogeneous evidence. Given the complexity of neurophysiologic endpoints and the diversity of experimental designs in slaughter research, a narrative approach was deemed more appropriate than a quantitative meta-analysis. The primary objective was interpretative integration of electrophysiologic and cerebral perfusion data rather than statistical aggregation of effect sizes.
A structured literature search was performed in PubMed/MEDLINE from database inception through January 2026. Search strings combined controlled vocabulary terms and free-text keywords related to species (“calf,” “veal,” “cattle”), neurophysiologic measurement (“electroencephalography,” “EEG,” “electrocorticography,” “evoked potentials”), cerebral hemodynamics (“cerebral blood flow,” “perfusion,” “oxygenation”), slaughter modalities (“stunning,” “captive bolt,” “electrical stunning,” “exsanguination,” “neck cutting”), and state descriptors (“unconsciousness,” “insensibility”). Boolean operators were used to maximize sensitivity while preserving conceptual specificity. Reference lists of retrieved articles were manually screened to identify additional eligible studies, particularly earlier experimental reports frequently cited in regulatory opinions.
Studies were eligible if they met the following criteria: 1 – peer-reviewed and indexed in PubMed; 2 – conducted in cattle or calves, with calf-specific data preferentially extracted where available; 3 – included direct neurophysiologic measurements, specifically EEG, electrocorticography (ECoG), or evoked potentials, with or without concomitant cerebral perfusion assessment; and 4 – published in English. Studies were excluded if they lacked direct cortical or perfusion endpoints, relied solely on behavioral indicators without physiologic validation, or involved non-bovine species without clear translational relevance to calf neurophysiology.
Given substantial heterogeneity in experimental paradigms, EEG criteria, perfusion methodologies, and outcome definitions, findings were synthesized qualitatively, emphasizing mechanistic interpretation and cross-study coherence rather than pooled quantitative estimates. The results were synthesized in 4 different areas of interest: Neurobiological Foundations; Electrophysiologic Evidence Across Slaughter Modalities; Cerebral Blood Flow and Cortical Viability; Comparative Mechanistic Interpretation.
3. Neurobiological Foundations
Conscious awareness in mammals depends on preserved thalamocortical processing and large-scale cortical network integration [9]. Contemporary neurobiological models emphasize that wakeful experience requires functional connectivity between thalamic relay nuclei and distributed cortical regions, with dynamic synchronization across sensory and associative networks [10]. Disruption of this integration, whether through mechanical trauma, electrical seizure induction, or ischemia, leads to loss of conscious processing [11]. In large animal models relevant to slaughter pathophysiology, cortical function has been assessed using electroencephalography (EEG) and evoked potentials [12,13]. EEG reflects spontaneous population-level cortical activity, while stimulus-locked evoked responses provide more specific evidence of preserved thalamocortical transmission. In cattle, Daly et al. [4] demonstrated that visually and somatosensory evoked cortical responses were abolished rapidly following effective captive bolt stunning, supporting their utility as markers of cortical non-responsiveness.
Similarly, Gregory et al. [6] showed rapid loss of visually evoked cortical responsiveness during exsanguination in calves. Cerebral perfusion provides the metabolic substrate necessary to sustain cortical electrical activity [14]. Experimental calf data indicate that progressive cerebral hypoperfusion during exsanguination leads to electrocortical suppression within seconds [5]. When cerebral blood flow declines below critical thresholds, organized cortical activity collapses, reinforcing the mechanistic linkage between perfusion failure and loss of consciousness [15]. These concepts are summarized in Figure 1.
4. Electrophysiologic Evidence Across Slaughter Modalities
4.1. Penetrating Captive Bolt (PCB)
Experimental neurophysiologic investigations in calves provide direct cortical evidence regarding the efficacy of penetrating captive bolt (PCB) stunning. In a controlled study using implanted EEG electrodes in veal calves, Lambooy et al. [16] demonstrated that correct frontal or occipital bolt placement induced rapid EEG changes characterized by high-amplitude slow activity rapidly transitioning toward suppression, consistent with abrupt loss of cortical function. In contrast, nuchal (neck) placement resulted in delayed electrocortical suppression, with a mean interval of approximately 21 seconds before EEG criteria compatible with unconsciousness were reached; corneal reflexes persisted until cortical suppression occurred. These findings highlight the critical importance of correct shot placement in determining the temporal profile of cortical failure.
Complementing these observations, Daly et al. [4] assessed cortical function in cattle using electrocorticography (ECoG) combined with visually and somatosensory evoked potentials. They reported rapid and irreversible abolition of evoked cortical responses following effective PCB stunning, indicating interruption of thalamocortical transmission after the impact. Because evoked responses represent stimulus-locked cortical processing, their disappearance provides strong evidence of loss of conscious responsiveness. Together, these studies support the conclusion that correctly applied PCB produces rapid cortical failure, while technical inaccuracies may introduce measurable delay and variability in the onset of unconsciousness. This concept is strongly reinforced by more recent data published in an important scientific journal [17].
4.2. Electrical Stunning
Electrical stunning produces a characteristic neurophysiologic sequence consisting of generalized epileptiform activity followed by post-ictal cortical suppression [5,18]. The initial high-amplitude, synchronized seizure activity reflects widespread depolarization across cortical networks, whereas the subsequent suppression phase corresponds to functional disruption of thalamocortical integration. In adult cattle, Devine et al. [19] documented marked EEG amplitude suppression and altered frequency patterns rapidly following electrical stunning, demonstrating electrocortical changes incompatible with conscious processing.
Head-only electrical stunning induces a reversible state [20]; if exsanguination is delayed, cortical activity may recover as cerebral perfusion is maintained. In contrast, head-to-body electrical stunning combines seizure induction with ventricular fibrillation or cardiac arrest, leading to cessation of cerebral perfusion and eliminating the possibility of neurologic recovery [21]. These mechanistic distinctions are central to interpreting EEG endpoints and assessing the temporal reliability of unconsciousness across electrical stunning modalities.
4.3. Non-Penetrating Captive Bolt
Non-penetrating captive bolt (NPCB) stunning relies exclusively on concussive transmission of kinetic energy without cranial penetration, aiming to induce cerebral concussion sufficient to disrupt cortical integration. In calves, Gibson et al. [22] reported electrocortical suppression following NPCB application under controlled conditions. However, subsequent EEG assessments in bovines indicated greater variability and occasional incomplete cortical suppression compared with penetrating captive bolt methods, underscoring the sensitivity of NPCB efficacy to energy delivery and anatomical factors [17].
4.4. Slaughter Without Stunning
Experimental investigations of exsanguination without prior stunning provide direct neurophysiologic insight into the temporal dynamics of cortical failure in calves. Gregory et al. [6] assessed visually evoked cortical responses during slaughter and reported loss of stimulus-locked cortical activity at approximately 17 seconds following bilateral carotid severance, indicating cessation of functional thalamocortical transmission within this interval. Complementing these findings, a different group of research evaluated spontaneous electrocortical activity in calves and observed rapid EEG suppression within seconds in most animals; however, rare prolonged cases were documented, attributed to carotid problems and incomplete exsanguination, thereby illustrating inter-individual variability in cerebral perfusion dynamics [5]. Importantly, validation studies comparing animal-based measures (ABMs) with electrophysiologic endpoints demonstrate imperfect concordance [7]. This study showed that certain behavioral indicators persisted beyond EEG-defined unconsciousness, while others disappeared prior to cortical suppression, underscoring the limitations of ABMs as standalone markers of conscious state. All together, these data highlight the progressive ischemic mechanism of cortical failure during exsanguination and the potential usefulness of electrophysiologic validation in defining unconsciousness.
All the examined literature is synthesized in Table 1. It is the summary of what has been described before.
5. Cerebral Blood Flow and Cortical Viability
Stunning and non-stunning slaughter modalities converge on the endpoint of cortical non-responsiveness, yet they do so through fundamentally distinct neurophysiologic mechanisms. Penetrating captive bolt stunning produces immediate mechanical disruption of cortical networks, abolishing evoked cortical responses at the moment of impact, as demonstrated by Daly et al. [4] using electrocorticography and stimulus-locked potentials in cattle. Electrical stunning induces generalized epileptiform activity followed by post-ictal suppression of cortical activity, with EEG changes incompatible with conscious processing documented in cattle [19]. In contrast, exsanguination without prior stunning leads to rapid progressive cerebral hypoperfusion, with loss of visually evoked cortical responsiveness occurring approximately 17 seconds after carotid severance in calves [6].
The integration of EEG with cerebral blood flow (CBF) assessment , illustrated in Figure 2, enables mechanistic differentiation between concussive suppression (direct mechanical network disruption), post-ictal suppression (seizure-related transient cortical inactivity), and ischemic cortical failure (perfusion-dependent collapse) [26]. While EEG alone reflects cortical electrical status, concurrent perfusion monitoring provides upstream physiological context [27]. Non-invasive technologies such as near-infrared spectroscopy (NIRS), which estimates regional cerebral oxygenation, and Doppler ultrasonography for flow velocity measurement, offer experimentally feasible methods to correlate perfusion decline with electrophysiologic endpoints [28]. Multimodal integration thus enhances interpretive precision and reduces the risk of misclassifying transient suppression states as definitive loss of consciousness [29].
6. Comparative Mechanistic Interpretation
The principal slaughter modalities differ in their primary neurophysiologic mechanisms yet converge on the endpoint of cortical non-responsiveness. Penetrating captive bolt (PCB) stunning produces rapid mechanical disruption of cortical and subcortical structures. When correctly positioned, abolition of evoked cortical responses occurs in a few seconds, after the impact, supporting rapid loss of thalamocortical integration [4]. However, incorrect placement can delay cortical suppression, as demonstrated in calves, underscoring significant placement-dependent variability [16].
Electrical stunning induces generalized epileptiform activity followed by post-ictal suppression; EEG patterns incompatible with conscious processing have been documented in cattle [19]. Technical parameters, including current density, electrode positioning, and stun-to-stick interval, determine reliability. Non-penetrating captive bolt (NPCB) relies on concussive force; while EEG suppression has been observed, variability is greater than with PCB, reflecting a very high sensitivity to delivered energy and anatomical variables [22].
In contrast, neck cutting without prior stunning induces progressive cerebral hypoperfusion, with loss of visually evoked cortical responsiveness typically within 10–20 s, though inter-individual variability occurs [5,6]. Collectively, the literature supports high probability of rapid unconsciousness under optimized conditions but does not justify claims of absolute certainty across all individuals and operational contexts.
In Figure 3 there is a graphical representation of what is described by the published research. Several aspects are definitely not clear yet, and would deserve further deep investigation.
7. Discussion
7.1. Integrative Interpretation
The convergence of electrophysiologic and cerebral perfusion evidence consistently indicates that cortical non-responsiveness, particularly the abolition of stimulus-locked evoked potentials, constitutes the most defensible operational endpoint for defining unconsciousness in calves [30,31]. Studies using electrocorticography and evoked responses have demonstrated rapid and irreversible cortical disruption following correctly applied penetrating captive bolt stunning [4], while ischemia-driven loss of visually evoked cortical responsiveness during exsanguination occurs within seconds [5,6]. Electrical stunning induces a characteristic seizure-suppression sequence incompatible with conscious processing when technically adequate [19]. Mechanistically, stunning modalities disrupt cortical network integrity directly, whereas exsanguination produces progressive metabolic collapse secondary to cerebral hypoperfusion.
Despite mechanistic coherence, several limitations temper interpretive certainty. Experimental cohorts are often small, limiting robust estimation of tail-risk events. EEG endpoint criteria vary across studies, reducing comparability. Multicenter EU-scale data reflecting operational heterogeneity remain limited. Moreover, simultaneous EEG-cerebral blood flow (CBF) measurements in calves are sparse, constraining physiologic triangulation of cortical failure.
7.2. Future Research Agenda
Aligned with Horizon Europe priorities for sustainable food systems and digital innovation, future investigations should prioritize multicenter EU-scale EEG-CBF validation in calves, development of field-deployable multimodal EEG-oxygenation headsets, and application of machine-learning classifiers to detect cortical failure in real time. Quantification of 99th percentile time-to-cortical-failure distributions would enhance reliability modeling. Harmonization of cortical endpoint definitions across Member States would further strengthen regulatory precision and reduce residual epistemic uncertainty in welfare assessment.
Limitations: This structured narrative review is characterized by several methodological and evidentiary constraints that warrant careful consideration. First, the body of primary literature addressing electrophysiologic determination of unconsciousness in calves remains limited in both scale and breadth. Many of the seminal investigations were conducted in relatively small experimental cohorts under controlled laboratory conditions, which may not fully capture the operational variability encountered in commercial slaughter environments. As a consequence, the capacity to estimate low-frequency or tail-risk events, particularly delayed cortical failure, is inherently restricted.
Second, substantial heterogeneity exists in EEG acquisition and interpretation methodologies. Variations in electrode configuration, recording parameters, signal processing approaches, and criteria for defining cortical suppression or isoelectricity complicate direct cross-study comparison. Moreover, the inconsistent application of evoked potential paradigms limits the harmonization of cortical endpoints across investigations and precludes meaningful quantitative synthesis.
Third, the simultaneous assessment of cerebral blood flow or cerebral oxygenation in conjunction with EEG remains uncommon in bovine research. Thus, mechanistic conclusions linking perfusion dynamics to cortical electrical failure are supported largely by indirect inference rather than robust multimodal datasets.
Finally, as a narrative synthesis, even when structured according to SANRA principles, this review does not meet the methodological rigor of a systematic review or meta-analysis and may be susceptible to publication bias and selective reporting. These limitations, together with all the published data, underscore the necessity for coordinated, multicenter, multimodal investigations aimed at elucidating the precise relationship between cortical electrophysiology and cerebral perfusion in calves.
8. Conclusions
Electroencephalography and stimulus-locked evoked potentials represent the most robust cortical indicators of unconsciousness currently available in calves, as they directly assess thalamocortical responsiveness. Measurement of cerebral perfusion offers complementary physiological validation by defining the metabolic substrate necessary for sustained cortical activity. The integration of electrophysiologic and perfusion metrics therefore constitutes the most scientifically rigorous approach to enhancing welfare monitoring and minimizing residual uncertainty in the assessment of unconsciousness during slaughter.
Author Contributions
Conceptualization, G.V.; methodology, D.M., G.F., M.L.G.L., G.V.; formal analysis, M.L.G.L., N.C., P.V.P., V.YT.; writing: original draft preparation, A.C., M.L.G.L., G.V.; writing: review and editing, A.A.A.A., G.F., N.C., P.V.P., V.YT. All authors have read and agreed to the submitted version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author.
Acknowledgments
We extend our sincere gratitude to the Fondazione Paolo Procacci for their invaluable support and assistance throughout the publication process.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ABMs | Animal-based Measures |
| CBF | Cerebral Blood Flow |
| ECoG | Electrocorticography |
| EEG | Electroencephalography |
| EFSA | European Food Safety Authority |
| EP | Evoked Potentials |
| EU | European Union |
| FFT | Fast Fourier Transform |
| HALF | High-amplitude low-frequency |
| LVFA | Low voltage fast activity |
| NIRS | Near-infrared spectroscopy |
| NPCB | Non-penetrating captive bolt |
| PCB | Penetrating captive bolt |
| Ptot | Total power |
| RMS | Root Mean Square |
| SANRA | Scale for the Assessment of Narrative Review Articles |
| SEF50 | Spectral Edge Frequency 50 |
| SEF95 | Spectral Edge Frequency 95 |
| VER | Visual evoked responses |
References
- Sgourdou, P. The Consciousness of Pain: A Thalamocortical Perspective. NeuroSci 2022, 3, 311–320. [Google Scholar] [CrossRef]
- Ong, W.-Y.; Stohler, C.S.; Herr, D.R. Role of the Prefrontal Cortex in Pain Processing. Mol. Neurobiol. 2018, 56, 1137–1166. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Larrea, L.; Bastuji, H. Pain and consciousness. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2018, 87, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Daly, C.; Kallweit, E.; Ellendorf, F. Cortical function in cattle during slaughter: Conventional captive bolt stunning followed by exsanguination compared with shechita slaughter. Veter- Rec. 1988, 122, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Bager, F.; Braggins, T.; Devine, C.; Graafhuis, A.; Mellor, D.; Tavener, A.; Upsdell, M. Onset of insensibility at slaughter in calves: Effects of electroplectic seizure and exsanguination on spontaneous electrocortical activity and indices of cerebral metabolism. Res. Veter- Sci. 1992, 52, 162–173. [Google Scholar] [CrossRef]
- Gregory, N.; Wotton, S. Time to loss of brain responsiveness following exsanguination in calves. Res. Veter- Sci. 1984, 37, 141–143. [Google Scholar] [CrossRef]
- Verhoeven, M.T.W.; Gerritzen, M.A.; Hellebrekers, L.J.; Kemp, B. Validation of indicators used to assess unconsciousness in veal calves at slaughter. Animal 2016, 10, 1457–1465. [Google Scholar] [CrossRef]
- Baethge, C.; Goldbeck-Wood, S.; Mertens, S. SANRA—a scale for the quality assessment of narrative review articles. Res. Integr. Peer Rev. 2019, 26, 5. [Google Scholar] [CrossRef]
- Müller, E.J.; Munn, B.R.; Redinbaugh, M.J.; Lizier, J.; Breakspear, M.; Saalmann, Y.B.; Shine, J.M. The non-specific matrix thalamus facilitates the cortical information processing modes relevant for conscious awareness. Cell Rep. 2023, 42, 112844. [Google Scholar] [CrossRef]
- Whyte, C.J.; Redinbaugh, M.J.; Shine, J.M.; Saalmann, Y.B. Thalamic contributions to the state and contents of consciousness. Neuron 2024, 112, 1611–1625. [Google Scholar] [CrossRef]
- Cacciatore, M.; Magnani, F.G.; Barbadoro, F.; Ippoliti, C.; Stanziano, M.; Clementi, L.; Nigri, A.; Nanetti, L.; Marino, S.; La Porta, F.; et al. Thalamus and consciousness: A systematic review on thalamic nuclei associated with consciousness. Front. Neurol. 2025, 16, 1509668. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Abubakar, A.A.; Sazili, A.Q.; Kaka, U.; Goh, Y.-M. Application of Electroencephalography in Preslaughter Management: A Review. Animals 2022, 12, 2857. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, I.; Goh, Y.M.; Norbaiyah, B.; Sazili, A.Q.; Lotfi, M.; Soleimani, A.F.; Small, A.H. Changes in blood parameters and electroencephalogram of cattle as affected by different stunning and slaughter methods in cattle. Anim. Prod. Sci. 2013, 54, 187–193. [Google Scholar] [CrossRef]
- Aubert, A.; Costalata, R.; Agnèsaubertab. A Model of the Coupling between Brain Electrical Activity, Metabolism, and Hemodynamics: Application to the Interpretation of Functional Neuroimaging. NeuroImage 2002, 17, 1162–1181. [Google Scholar] [CrossRef]
- Van Lieshout, J.J.; Wieling, W.; Karemaker, J.M.; Secher, N.H. Syncope, cerebral perfusion, and oxygenation. J. Appl. Physiol. 2003, 94, 833–848. [Google Scholar] [CrossRef]
- Lambooy, E.; Spanjaard, W. Effect of the shooting position on the stunning of calves by captive bolt. Veter- Rec. 1981, 109, 359–361. [Google Scholar] [CrossRef]
- Gibson, T.J.; Oliveira, S.E.O.; Costa, F.A.D.; Gregory, N.G. Electroencephalographic assessment of pneumatically powered penetrating and non-penetrating captive-bolt stunning of bulls. Meat Sci. 2019, 151, 54–59. [Google Scholar] [CrossRef]
- Devine, C.; Tavener, A.; Graafhuis, A.; Gilbert, K. Electroencephalographic studies of calves associated with electrical stunning, throat cutting and carcass electro-immobilisation. New Zealand Veter- J. 1987, 35, 107–112. [Google Scholar] [CrossRef]
- Devine, C.; Tavener, A.; Gilbert, K.; Day, A. Electroencephalographic studies of adult cattle associated with electrical stunning, throat cutting and carcass electro-immobilization. New Zealand Veter- J. 1986, 34, 210–213. [Google Scholar] [CrossRef]
- Beausoleil, N.; Farouk, M.; Webster, J.; Johnson, C.; Dowling, S.; Sazili, A.; Cameron, C. Comparison of recovery of sheep, goats, and calves from reversible electrical head-only and head-to-body stunning for halal meat production. New Zealand Veter- J. 2024, 72, 288–299. [Google Scholar] [CrossRef]
- Musk, G.C.; Johnson, C.B. Commentary: A Comparison of the Methods of the Pre-Slaughter Stunning of Cattle in Australia—Mechanical, Electrical, and Diathermic Syncope. Animals 2024, 14, 3141. [Google Scholar] [CrossRef] [PubMed]
- Gibson, T.; Johnson, C.; Murrell, J.; Mitchinson, S.; Stafford, K.; Mellor, D. Electroencephalographic responses to concussive non-penetrative captive-bolt stunning in halothane-anaesthetised calves. New Zealand Veter- J. 2009, 57, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Imlan, J.C.; Kaka, U.; Goh, Y.-M.; Idrus, Z.; Awad, E.A.; Abubakar, A.A.; Ahmad, T.; Nizamuddin, H.N.Q.; Sazili, A.Q. Effects of Slaughter Knife Sharpness on Blood Biochemical and Electroencephalogram Changes in Cattle. Animals 2020, 10, 579. [Google Scholar] [CrossRef] [PubMed]
- Imlan, J.C.; Kaka, U.; Goh, Y.-M.; Idrus, Z.; Awad, E.A.; Abubakar, A.A.; Ahmad, T.; Nizamuddin, H.N.Q.; Sazili, A.Q. Effects of Slaughter Positions on Catecholamine, Blood Biochemical and Electroencephalogram Changes in Cattle Restrained Using a Modified Mark IV Box. Animals 2021, 11, 1979. [Google Scholar] [CrossRef]
- Small, A.; Jenson, I.; Phillips, A.; McLean, D.; Kalinowski, T.; Ralph, J. Cattle recover completely from unconsciousness induced by controlled application of 150–180 kJ of 915 MHz microwave energy to the forehead. Veter- Anim. Sci. 2025, 29, 100466. [Google Scholar] [CrossRef]
- Sherman, D.L.; Thakor, N.V. EEG Signal Processing: Theory and Applications. In Neural Engineering; He, B., Ed.; Springer: Cham, Switzerland, 2020; pp. 97–129. [Google Scholar] [CrossRef]
- Chung, C.K.E.; Poon, C.C.M.; Irwin, M.G. Peri-operative neurological monitoring with electroencephalography and cerebral oximetry: A narrative review. Anaesthesia 2022, 77, 113–122. [Google Scholar] [CrossRef]
- Nischal, S.A.; Patel, S.; Yuen, J.; Mortimer, A.; Patel, N.K. Cerebral blood flow and modern approaches for clinical assessment & monitoring: A view to the future. Br. J. Neurosurg. 2025, 1–15. [Google Scholar] [CrossRef]
- Gallucci, A.; Varoli, E.; Del Mauro, L.; Hassan, G.; Rovida, M.; Comanducci, A.; Casarotto, S.; Re, V.L.; Lauro, L.J.R. Multimodal approaches supporting the diagnosis, prognosis and investigation of neural correlates of disorders of consciousness: A systematic review. Eur. J. Neurosci. 2023, 59, 874–933. [Google Scholar] [CrossRef]
- Meyer, R.E. Physiologic Measures of Animal Stress during Transitional States of Consciousness. Animals 2015, 5, 702–716. [Google Scholar] [CrossRef]
- Hascalovici, J.R.; Schipper, H.M.; Regenstein, J.M.; Rosen, S.D.; Zivotofsky, A.; St-Jean, G.; Freilich, S.; Morrison, T.J.; Rabkin, S.W.; Haut, S. Rapid loss of consciousness in cattle following nonstun slaughter: Evidence from a systematic review. Am. J. Veter- Res. 2025, 86, 1–10. [Google Scholar] [CrossRef]
Figure 1.
– Neurobiological model of conscious awareness and cortical collapse. Conscious awareness is sustained by integrated, dynamically synchronized thalamocortical interactions between thalamic relay nuclei and distributed cortical regions, enabling functional connectivity across sensory and associative networks. Disruption by mechanical trauma (e.g., stunning), electrically induced seizures, or ischemia/perfusion failure produces a breakdown of integration with loss of thalamocortical processing. In the setting of cerebral hypoperfusion (e.g., exsanguination), crossing a critical metabolic threshold rapidly precipitates electrocortical suppression and loss of consciousness, reflecting collapse of organized cortical activity within seconds. Conscious state and network integrity can be assessed using EEG (spontaneous population-level cortical activity) and evoked potentials (EPs) (stimulus-locked thalamocortical transmission).
Figure 1.
– Neurobiological model of conscious awareness and cortical collapse. Conscious awareness is sustained by integrated, dynamically synchronized thalamocortical interactions between thalamic relay nuclei and distributed cortical regions, enabling functional connectivity across sensory and associative networks. Disruption by mechanical trauma (e.g., stunning), electrically induced seizures, or ischemia/perfusion failure produces a breakdown of integration with loss of thalamocortical processing. In the setting of cerebral hypoperfusion (e.g., exsanguination), crossing a critical metabolic threshold rapidly precipitates electrocortical suppression and loss of consciousness, reflecting collapse of organized cortical activity within seconds. Conscious state and network integrity can be assessed using EEG (spontaneous population-level cortical activity) and evoked potentials (EPs) (stimulus-locked thalamocortical transmission).
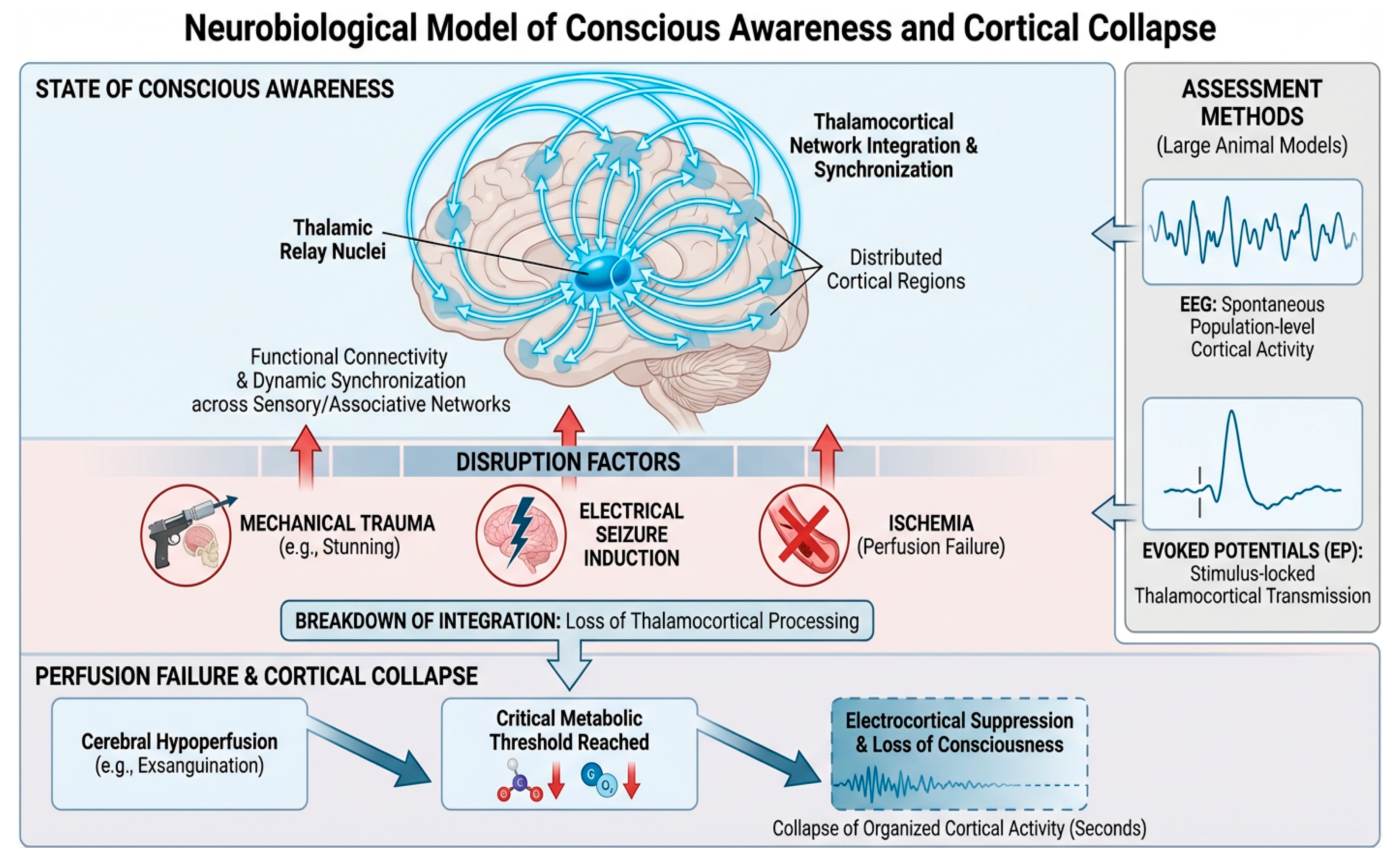
Figure 2.
– Mechanistic integration of electrophysiologic and cerebral perfusion pathways to cortical unconsciousness across slaughter modalities in calves. The schematic illustrates how PCB, electrical stunning, NPCB, and exsanguination without prior stunning converge on a shared cortical endpoint (cortical non-responsiveness) through distinct neurophysiologic pathways. Mechanical methods primarily induce concussive/mechanical cortical disruption, electrical stunning produces an ictal (seizure-like) phase followed by post-ictal cortical suppression, and exsanguination leads to progressive cerebral hypoperfusion with ischemic cortical failure. EEG and (EPs are shown as the principal direct cortical markers, whereas CBF/cerebral oxygenation provides physiologic context for mechanistic interpretation. Animal-based measures are included as supportive operational indicators but not standalone markers of consciousness.
Figure 2.
– Mechanistic integration of electrophysiologic and cerebral perfusion pathways to cortical unconsciousness across slaughter modalities in calves. The schematic illustrates how PCB, electrical stunning, NPCB, and exsanguination without prior stunning converge on a shared cortical endpoint (cortical non-responsiveness) through distinct neurophysiologic pathways. Mechanical methods primarily induce concussive/mechanical cortical disruption, electrical stunning produces an ictal (seizure-like) phase followed by post-ictal cortical suppression, and exsanguination leads to progressive cerebral hypoperfusion with ischemic cortical failure. EEG and (EPs are shown as the principal direct cortical markers, whereas CBF/cerebral oxygenation provides physiologic context for mechanistic interpretation. Animal-based measures are included as supportive operational indicators but not standalone markers of consciousness.
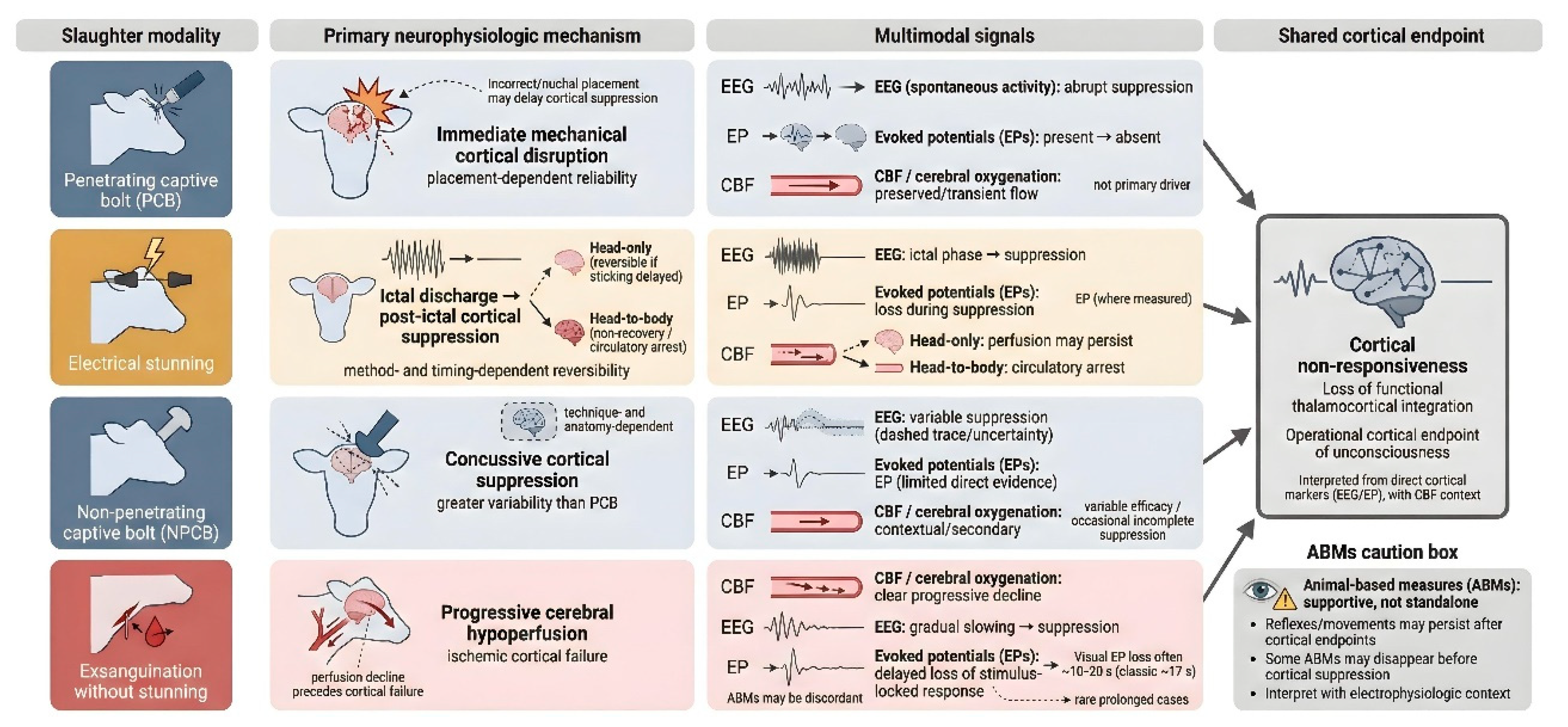
Figure 3.
– Comparative temporal trajectories and relative utility of monitoring markers for cortical unconsciousness assessment across slaughter modalities in calves. Panel A shows a conceptual, modality-specific timeline to cortical non-responsiveness (relative time scale). PCB typically produces immediate mechanical cortical disruption when correctly positioned, but malposition (e.g., nuchal placement) may delay electrocortical suppression. Electrical stunning follows an ictal (seizure-like) phase with subsequent post-ictal cortical suppression, with reversibility influenced by stunning method and stun-to-stick interval. NPCB can induce cortical suppression but shows greater variability and occasional incomplete suppression. Exsanguination without prior stunning follows a progressive hypoperfusion/ ischemic trajectory, in which cerebral perfusion decline precedes cortical failure; EEG suppression may occur within seconds, whereas loss of visual evoked cortical responses generally occurs later. Panel B summarizes the relative utility of monitoring markers: EEG and EPs are the principal direct cortical markers (with variable evidence density across modalities), CBF/cerebral oxygenation provides physiologic context (particularly for exsanguination), and ABMs are supportive but not standalone due to imperfect concordance with electrophysiologic endpoints. This schematic is a conceptual synthesis of heterogeneous studies, not a pooled quantitative estimate.
Figure 3.
– Comparative temporal trajectories and relative utility of monitoring markers for cortical unconsciousness assessment across slaughter modalities in calves. Panel A shows a conceptual, modality-specific timeline to cortical non-responsiveness (relative time scale). PCB typically produces immediate mechanical cortical disruption when correctly positioned, but malposition (e.g., nuchal placement) may delay electrocortical suppression. Electrical stunning follows an ictal (seizure-like) phase with subsequent post-ictal cortical suppression, with reversibility influenced by stunning method and stun-to-stick interval. NPCB can induce cortical suppression but shows greater variability and occasional incomplete suppression. Exsanguination without prior stunning follows a progressive hypoperfusion/ ischemic trajectory, in which cerebral perfusion decline precedes cortical failure; EEG suppression may occur within seconds, whereas loss of visual evoked cortical responses generally occurs later. Panel B summarizes the relative utility of monitoring markers: EEG and EPs are the principal direct cortical markers (with variable evidence density across modalities), CBF/cerebral oxygenation provides physiologic context (particularly for exsanguination), and ABMs are supportive but not standalone due to imperfect concordance with electrophysiologic endpoints. This schematic is a conceptual synthesis of heterogeneous studies, not a pooled quantitative estimate.
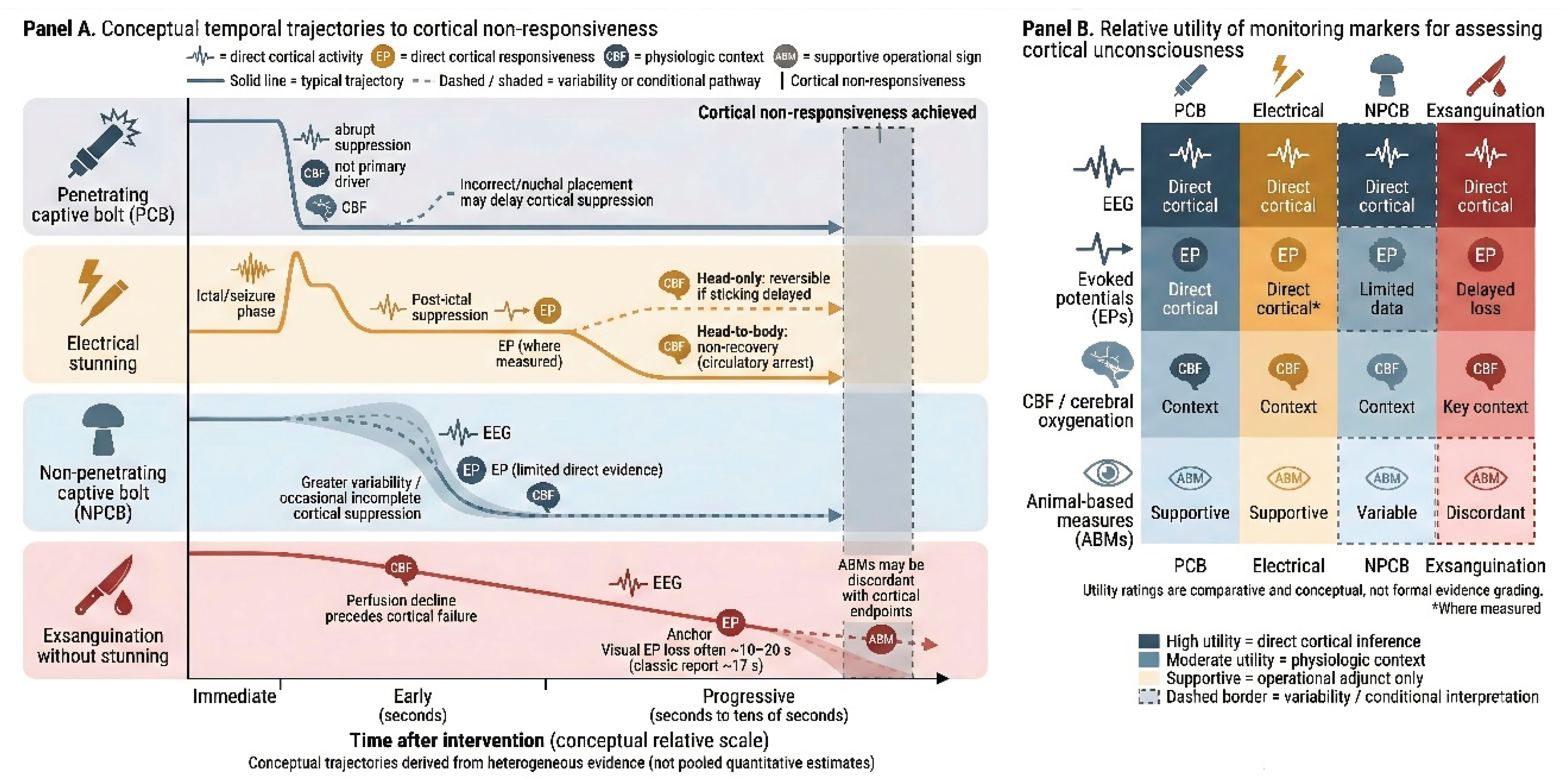

Table 1.
Key studies examined in the research.
| Domains | Study (Year) | Animal | Group | EEG/ ECoG | EEG Criterion for Unconsciousness | Key Findings |
|---|---|---|---|---|---|---|
| Penetrating Captive Bolt (PCB) | Lambooy (1981) [16] | Veal calves | Frontal vs occipital vs nape/neck placement | EEG (implanted electrodes) | Delta/theta tending to isoelectric line; consciousness judged from EEG | Frontal & occipital: 'almost certainly immediate' unconsciousness; nape: mean 21±6 s to unconsciousness; calves conscious until then |
| Daly et al (1987) [4] | Cattle | Bolt velocity comparison: 41 vs 47 vs 55 vs 58 m/s | ECoG (spontaneous cortical activity; epidural/dural electrodes) + VER (visual evoked responses) | - Not an EEG-only criterion; - Use loss of VER as primary indicator of insensibility; “severe reduction” of spontaneous ECoG defined as <10% of pre-shot baseline; VER considered “present” if >10% baseline |
- Higher bolt velocity (55–58 m/s) ↓ likelihood of VER presence in the first ~8 s and ↑ time to VER recovery; however VER could persist immediately after stun at all velocities. Spontaneous ECoG suppression (<10%) did not differ significantly by velocity. - 55 m/s as minimum recommended velocity |
|
| Electrical stunning | Devine et al (1986) [19] | Adult cattle | Immobilization (n=10) vs no immobilization (n=2) | Implanted EEG (frontal bone screw electrodes; transhemispheric bipolar); recorder disconnected during stun/ immobilization (no EEG during current) | Uses “window of sensibility” based on LVFA amplitude ~10–40 µV - insensibility inferred when EEG falls <10 µV (toward isoelectric) and LVFA absent/ outside window |
EEG amplitude <10 μV at end of immobilization; in non-immobilized animals reached <10 μV at 57 s and 63 s after stun initiation |
| Bager et al. (1992) [5] | Calves | Head-only electrical stun + slaughter vs no-stun slaughter | ECoG+ FFT analysis of power bands 8–30 Hz (sensibility-associated) and 2–8 Hz (insensibility/ anoxia-associated) | Loss of sensibility inferred from electrocortical activity analysis (FFT; HALF etc.) | Stunned calves: permanently insensible at time of stun. Non-stunned: 6 calves lost sensibility within 10 s; one calf with carotid clot had cortical activity >52 s but sensibility not regained (FFT) |
|
| Non-penetrating captive bolt (NPCB) | Gibson et al. (2009) [22] | Calves | Single group | EEG + FFT to compute total power (Ptot); visual classification: active / transitional / high-amplitude low-frequency / isoelectric | Visual inspection + changes in total power (Ptot) | Changes sufficient to produce insensibility within 0–14 s in conscious animals (inference); biphasic Ptot response in 5 animals |
| Verhoeven et al. (2016) [7] | Veal calves | (1) Inverted captive bolt + neck cut (n=25) (2) Upright no-stun (n=7) (3) Inverted no-stun (n=25); (4) Upright no-stun then captive bolt at ~34±8 s (n=25) |
Non-invasive EEG electrodes | Visual EEG staging: baseline → transitional → unconscious when low-freq/high-amp dominates → minimal brain activity/flat EEG defined as <10% baseline amplitude | After captive bolt, absence of threat/withdrawal/corneal/eyelid reflex indicated unconsciousness per EEG; after non-stun, threat/withdrawal lost before EEG-defined unconsciousness; eyelid/corneal were conservative, persisting until ~76±50 and 85±45 s after EEG loss of consciousness | |
| Gibson et al. (2019) [17] | Bulls | PCB (n=20) vs NPCB (n=11) | EEG; visual classification + spectral analysis | Visual EEG states: Normal, Transitional (suppressed; <½ baseline amp), HALF (high-amp low-freq), Isoelectric (amp <1/8 = 12.25% baseline, little/no low-freq). | PCB: 20/20 inconsistent with consciousness; NPCB: 82% complete unconsciousness; 2 bulls showed normal EEG activity periods indicating incomplete concussion | |
| Slaughter Without Stunning | Gregory & Wotton (1984) [6] | Calves | Exsanguination after cutting carotids/jugulars | ECoG (implanted electrodes) + flash visually evoked cortical responses (VER/EP) | Loss of visually evoked cortical response to flash used as indicator of loss of brain responsiveness | Time from sticking to loss of brain responsiveness: 17 ± 4 s (range 11.5–23 s). “Flat” ECoG occurred later on average (23 ± 11 s), supporting that evoked response loss is a more direct marker than waiting for isoelectric ECoG. |
| Imlan et al. (2020) [23] | Cattle | Sharp knife (n=10) vs commercial sharp knife (n=10) | EEG + FFT spectral analysis | Not an unconsciousness criterion study. - - F50 and Ptot (and band powers) as indices of pain/stress after neck cut, not as thresholds for loss of consciousness | Higher EEG median frequency and total power in commercial sharp knife group post-slaughter, interpreted as higher pain/stress | |
| Imlan et al. (2021) [24] | Cattle | Lateral recumbency (LP) n=10 vs Upright position (UP) n=10 | EEG + FFT spectral analysis | Not unconsciousness; EEG changes interpreted as pain/stress | Differences in catecholamines; EEG responses measured; details require full text | |
| Small et al. (2025) [25] | Cattle | EEG recorded in 6; one handled commercially without EEG delay | Non-invasive EEG; offline analysis: RMS power, FFT Ptot (0–30 Hz), SEF50, SEF95, and band power | No fixed numeric EEG threshold; “insensibility duration” operationally based on absence of eye reflexes and awareness, with EEG used to document suppression and frequency shifts (↓SEF50/95, ↓Ptot) during the unconscious period | - Behavioural loss of consciousness within 10 s; EEG-based insensibility duration ~63 s or more; - EEG showed suppression and shift to lower frequencies (↓SEF50/95; ↓Ptot) for ~60–90 s or longer, with gradual recovery - Full behavioural recovery observed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



