Submitted:
19 February 2026
Posted:
27 February 2026
You are already at the latest version
Abstract
An influential intuition about the biology of aging is that organisms are born with a fixed amount of “life stuff” that they burn through in the process of living. According to this mindset, the key to living longer is simply to burn through this life stuff as slowly as possible. We instead reinterpret this “life stuff” as Survival Capital: resources allocated to maintenance and growth that can compound over time to facilitate future reproduction. Contrasting with classical evolutionary theories that treat life-history expenditures as linear substitutes, we develop an explicit framework in which investments in survival and reproduction act as synergistic complements. We first show how early-life investments in survival traits generate complementary incentives for later-life investments, thereby increasing the value of survival into older ages. We then add multiplicative damage to survival, capturing biologically plausible risk accumulation without altering the qualitative logic of the first model, thereby reinforcing the effects of complementarity among survival investments. Next, we demonstrate that such complementarities naturally imply bi-stability of life-history strategies when trade-offs between survival and reproduction across discrete life stages are considered. Finally, we show that when investments can be made to affect multiple components of health and interact synergistically, they can produce runaway increases in health investment, driving extreme healthspans. This moves beyond the classical “fast–slow” continuum by making the value of later life endogenous. Linking these allocation trade-offs to existing evidence concerning real world counterparts of Survival Capital generates powerful empirical predictions for both evolutionary biology and synergistic geroscience.
Keywords:
senescence
; healthspan
; aging
; geroscience
; life-history theory
; synergistic epistasis
; fitness capital
; resource allocation
1. Introduction
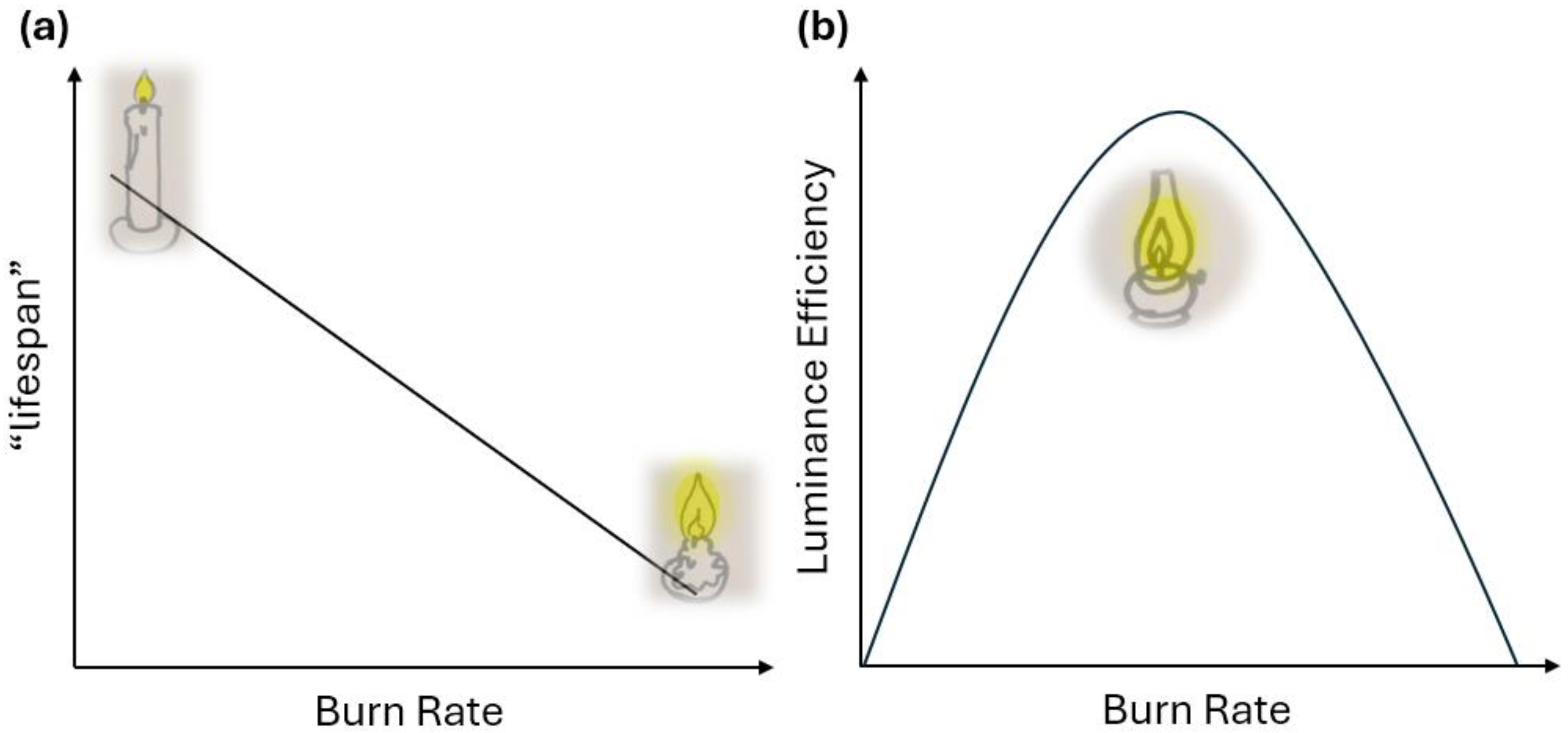
Metaphors matter because they shape not only how existing facts are perceived, but also which facts become discoverable. Many ideas about aging share an assumption that can be captured by the metaphor of candlelight. Once lit, a candle draws down on a finite fuel source, the wick. If life is like light, this metaphor implies a trade-off between lasting longer by burning more slowly (emitting less light) and burning brighter but extinguishing sooner. This intuition recurs in various biological concepts, ranging from the rate of telomere shortening (seen as limiting a cell’s “replicative budget”) to the notion that animals have a finite number of lifetime heartbeats that can either be spent quickly or conserved over time, forming a continuum from the fast-paced life of short-lived mice to the slower pace of long-lived elephants. This "candle mindset" implies that longevity is a zero-sum game of conservation. However, a living organism is less like a candle and more like an automobile with a combustion engine. Its "fuel" is not a fixed reservoir but a dynamic flow, where the supply of fuel and continual maintenance is regulated by the value the work it produces. In this view, longevity is not about minimizing activity (dimming the flame), but about maximizing the efficiency with which resources are translated into sustained functional output — a concept we define here as “healthspan”.
While the car engine highlights the necessity of flow and maintenance, the older metaphor of the oil lamp allows us to see why simply “minimizing the burn” can be as lethal as overexertion. On this analogy, the flame’s brightness again represents an organism’s functional output, such as activities, growth and repair, reproduction, and so forth. This brightness depends on the fuel, the working wick, and airflow, with the wick drawing fuel from the reservoir by capillary action. A small knob allows the precise manual adjustment of the wick’s height (raising the wick increases the burn rate and thus the flame’s brightness). Unlike the candle metaphor, simply minimizing the rate of burn is clearly suboptimal: if the wick is too thin, or not fed enough fuel, only a weak, sputtering flame can burn. Energy barely converts into light, soot accumulates reducing combustion efficiency, and the flame is prone to extinguishing despite abundant fuel below. Conversely, brighter is not always better: setting the wick too high or burning too intensely causes smoke, wasted heat, and reduced efficiency. There is a “sweet spot,” an optimal combustion rate where fuel converts efficiently to light, heat loss is minimized, and the flame burns steadily over time.
Figure 1.
Contrasting Metaphors for Life (a) If life is candlelight, slowing the rate of burn may be seen as extending the total time the candle is lit (“lifespan”), whereas burning too brightly (i.e., to burn the candle at both ends”) shortens it. (b) Alternatively, an oil lamp operating within a sweet spot of fuel-to-light conversion is more likely to be used for longer than one with either too weak a flame (vulnerable to sputtering out), or too strong may even pose fire hazards as if a Molotov cocktail.
Figure 1.
Contrasting Metaphors for Life (a) If life is candlelight, slowing the rate of burn may be seen as extending the total time the candle is lit (“lifespan”), whereas burning too brightly (i.e., to burn the candle at both ends”) shortens it. (b) Alternatively, an oil lamp operating within a sweet spot of fuel-to-light conversion is more likely to be used for longer than one with either too weak a flame (vulnerable to sputtering out), or too strong may even pose fire hazards as if a Molotov cocktail.
Classical evolutionary theories, like those following Williams (1957) and Hamilton (1966; 1996), essentially treat life as an oil lamp with a fixed reservoir. Optimal life, like optimal flame, is not dim, but well-regulated and efficient. In nature, metabolic output, nutrient supply, growth and repair processes all interact to determine both fitness outcomes and longevity. A heart or muscle operating too slowly, sluggish cellular repair processes, chronic catabolic states, etc., may be more like a lamp with a sputtering flame, symptoms of not being long for this world rather than signatures of long-term survival. On these views, the decision that Natural Selection shapes is primarily a matter of when reproductive effort is to be focused. This means that resources spent on maintaining health in later life usually come at the expense of breeding earlier in life and overall reproductive output. In such models, survival investments are therefore usually seen acting like as “substitutes”. Hence, there tends to be a gradated deterioration in health that follows (sometimes steeply) from the point where reproductive efforts are best concentrated (a process we recognize as “aging”) (for reviews, see Flatt & Partridge, 2018; McAuley, 2025).
We seek to extend these ideas by importing insights from Becker’s (1965; 1975; 2002) and Grossman’s (1972) economic framework of Health Capital. In this light, health is not a depleting fuel, but an investable and accumulative asset. Because survival is a prerequisite for all future payoffs, investments in health are fundamentally complementary. Improving one aspect of survival (e.g., immune function) increases the expected "dividend" of all other survival investments (e.g., cancer resistance), because the organism is now more likely to be alive to reap those benefits. This complementarity operates symmetrically. An adaptation that increases the efficiency of one repair system amplifies the marginal returns to all other systems. Just as a gaslight is maintained only as long as its illumination justifies the gas consumed, an organism’s selective gradient is not a fixed downward slope, but an endogenous outcome of its own "survival capital" investment. When this feedback is strong, the traditional trade-off between reproduction and longevity can disappear. Thus, we would not find as paradoxical findings that reproductive output and lifespan often correlate positively. This is a finding that runs contrary to traditional approaches, a phenomenon sometimes attributed to deleterious mutational load (Flatt & Partridge, 2018; Van Noordwijk & de Jong, 1986). However, in our terms, we may characterise this as a “wealth effect”, expected to arise in organisms who can afford higher fertility and longevity because living longer enables them to acquire more resources.
The present thesis suggests that the force of selection acting to shape the healthspan of species is not an exogenous law of nature that simply decays with time, but a dynamic variable amplified through intertemporal feedback. By modelling health as a stock of synergistic capital rather than a depleting candle, we can see that organisms may not simply slide into senescence; they can be pushed, for adaptive reasons, toward different life history strategies. In some cases, a bistable threshold emerges, contrasting a 'life fast, die young' regime (where low survival efficacy makes maintenance a wasted effort), or a “Survival Capital” regime (where synergistic returns drive a runaway increase in health investment). In the sections that follow, we formalize this logic. We first show how complementarity between early and late survival creates an 'uplift' effect on the marginal benefit of maintenance. We then introduce the multiplicative damage to illustrate how repair systems reinforce one another, creating a “synergy window” through which investment in health can flip the direction of life-history evolution. Ultimately, we show that when the efficiency of survival investment passes a threshold, costly investments in health no longer act as a tether limiting reproduction, but as a lever that extends lifetime reproductive potential by expanding the healthspan, providing a new theoretical foundation for the diversity of aging in nature.
2. The Model
2.1. The Basic Becker-Hamilton Model
To formalize how survival investments reinforce one another across time, we begin by integrating Becker’s resource allocation logic with Hamilton’s age-structured selection framework. This section derives an “uplift effect”: the mechanism where surviving to one stage endogenously increases the marginal value of survival in the next. By treating survival as a series of interlinked probabilities rather than a fixed decay, we recast the selective gradient not an exogenous given, but a product of the organism’s own maintenance strategy.
Consider an organism with two ages at which it can reproduce (age 1 and age 2). Let be the probability an individual survives from birth to the end of age 1, while represents the probability that the individual who has reached age 1 will also survive to the end of age 2. Thus, the probability of surviving from birth through to age the end of age 2 is .
Let and represent the expected number of offspring produced during age 1 and age 2, respectively (we think of these as age-specific fertility payoffs). Expected lifetime reproduction (a simple measure of fitness being expected offspring), is then:
This means that the expected reproduction of an individual at age 1 is (because it must survive to reproduce), while the expected reproduction at age 2 being the probability of also surviving through age 2 (which is multiplied by fecundity ). Suppose that animals can make investments that increase dissociable aspects of survival, and that these investments cost. We can model the cost as a simple quadratic function (producing a standard, convex cost with increasing marginal cost) as follows:
With , representing costs parameters affecting each type of survival investment, such that larger means survival is costlier to increase. The net payoff (what selection will “favor” if costs translate into trade-offs) is:
If increasing raises the marginal benefit of increasing (and/or vice versa), then investments in survival are complementary. To determine the optimal investments, we take partial derivatives and set them to zero. First, taking the derivative with respect to :
Then, taking the derivative with respect to gives:
These two first order conditions determine the interior optimum (). To solve this from (2):
Substitute into (1):
So, if we presume that the denominator is positive, optimal investment in surviving stage 1 is:
Then, optimal investment in surviving stage 2 is:
To ensure an interior solution, we require and . The marginal fitness contributions are thus . That is, the marginal fitness contribution of increasing probability of surviving to time 2 is proportional to the probability of surviving to time 1. This means that the marginal fitness benefit of raising increases linearly with . Put another way, this is the Becker-style complementarity: improving one aspect of survival is not just a defensive measure to delay death; it is a fitness-growing investment that expands the potential for future reproductive output1. Because the cross partial of fitness is positive
This means that the effect of increasing survival at time 2 increases the marginal benefit to fitness of surviving to time 1. From the first-order conditions we thus see explicit interdependence: the optimal depends on and on (via ). This shows a clear feedback loop between investment in one aspect of survival and investment in others.
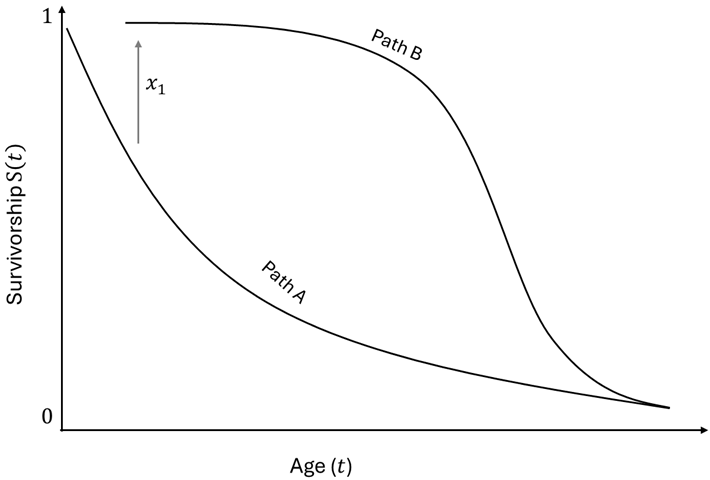
If (fitness payoff at age 2) increases, then the denominator shrinks, so increases. That is, greater value of reproduction at later life raises selection to invest in early-life survival because early survival unlocks the later payoffs. Conversely, a mutation that raises (e.g., better juvenile disease resistance) increases the marginal benefit of increasing (e.g., later life-maintenance), so selection should favor higher . This is the complementarity feedback loop. Thus, the term that appears in the first-order condition represents a sort of marginal “shadow value” or an “uplift” effect. As shown in Figure 2, the vertical gap between the baseline marginal benefit and the uplifted curve represents the value of future reproduction, which if increased shifts the optimal survival through stage 1 rightward. Note that the term can thus be thought of as a measure capturing the “shadow value” of future fertility feedback, in that it represents the feedback term in the denominator that amplifies early survival investment when later fertility is high.
The feasibility condition mentioned () captures biological constraints: if payoffs at later ages are extremely high relative to costs, the simple convex-like cost model predicts very large investments. In nature, this would likely be tempered by other constraints, such as resource limitations, pleiotropic costs, ecological hazards, etc., that are not considered in this minimal model.
If we think of Hamilton as showing how selection pressure shifts mechanically with age, given a fixed survival curve, then the Becker-inspired extension shows how those curves themselves are interlinked, because improving one part of the survival schedule endogenously changes the selective landscape for others. This shows that when survival probabilities at different life stages are complementary, improvements in early survival raise the marginal value of later survival (and vice versa). This creates feedback loops in the evolution of survival traits: populations that extend survival in one phase experience increased selection for survival in others.
Notice also that, although we have illustrated complementarity between investments made at different life stages, an analogous complementarity can arise among distinct survival mechanisms within a single stage. For example, concurrent investments made in defences against predation and avoidance of dietary toxins can also be seen as complements in the same sense, since each increases the marginal value of the other when overall survival depends multiplicatively on both. In the stage based models considered here, complementarity arises because the marginal value of one survival investment increases with the level of another when survival between stages enters fitness in a mutually reinforcing way. Formally, the cross partial derivative is whenever survival probabilities compound across investments. The same structure applies within a single stage if survival during that period is produced by various, jointly necessary investments. E.g., if survival at the period is:
So that, for instance, if investment in both skills enabling the organism to evade predators and learning to avoid consumption of poisonous food both contribute to the probability of reaching the next stage. Then:
And because lifetime fitness multiplies each stage of survival across stages, the positive complementarity simply “nests” (within-stage complemetnarity feeds into between stage complementarity). Although the remainder of this paper further sketches a model based on stage-specific investments for illustrative purposes, it should be remembered that the same logic that applies to consecutive investments should equally apply to concurrent investments in health.
2.2. Multiplicative Damage and Survival Capital
We can reinforce this notion of complementarity among survival investments by introducing some biological realism to the basic model. In nature, many challenges to fitness are not immediately fully fatal, but are incrementally insidious: near misses, degree of survival parasite load, injuries, and so on, may not be immediately fatal, but add up to have weightier impacts on fitness later in life. Just as it may be harder to guard against death from a lightning bolt than heat exposure, these may also be the sorts of gradated challenges where investments in health would be most amenable to selection. By introducing multiplicative damage, we demonstrate how investments in somatic maintenance do not merely prevent death in the present, but actively slow the rate of cumulative decay, thereby amplifying the returns of every subsequent investment.
To explore this sort of biological reality, we keep the two-age structure introduced above. Let and denote investments into specific survival “technologies” (these correspond to variables that determine survival probabilities). Define stage-specific trait-based survival contributions and with:
Damage accumulates additively across stages. Let be the damage increment accrued during stage , such that investing more reduces damage accrual. So,
Define cumulative damage after stage by:
The probability of surviving from birth through to the end of stage is now denoted as 2. With additive damage and multiplicative effects on survival, the natural per-stage expressions are:
And
Writing survivorship this way preserves the intuitive multiplicative structure while making the effect of cumulative damage explicit. Age-specific fecundities are and as before. Expected lifetime reproduction is now expressed by:
As before, we impose convex costs on investments writing these costs as quadratic in :
With . The objective net payoff is:
We next calculate the first order conditions with respect to and set to zero. The derivative of with respect to . Letting , then using the product and then the chain rule:
Where and . Intuitively, the first bracketed term is the direct survival gain from increasing the trait, and the second term () is the indirect benefit from reducing damage accrual. To take the derivative of with respect to , let and treat and as constants when differentiate with respect to :
This shows that increasing has a marginal effect on survival to age 2 that is scaled by survival to age 1 (the factor ). This is the explicit Becker-style complementarity with the payoff to later-life investment being multiplied by the probability of surviving to that age.
The cross derivative shows how affects . This is taken by differentiating with respect to , using the product and chain rule.
Equivalently,
Notice that this is another form of complementarity: the marginal changes in early investments propagate forward to affect later survival multiplicatively. Finally, we take the derivatives of with respect to investments and set equal to zero to find the optimal investment levels. With respect to :
Substituting the expanded expressions for the -derivatives gives a condition that combines the direct and indirect returns form increasing investment to improve immediate survival and reduce damage sustained that affects later survival and equates these to marginal cost . In biological terms, the first term represents the immediate survival benefit of the investment, while the second term captures the payoff from living a healthier life: the reduction in cumulative damage that makes later stages of life more worthwhile for selection to maintain. In other words, whereas in section 2.1, investment only affected stage 1 survival, here, investment persists to affect fitness in both stages. Deriving with respect to :
I.e. the marginal benefit of stage 2 investment (scaled by the probability of reaching stage 2) equals marginal cost. For clarity, the abstract expansions can be replaced with the explicit bracketed forms from above. The factors multiplies the return to investment in stage 2. Thus, the marginal value of later-life survival depends on the probability of reaching that stage of life (the same Becker-style complementarity emphasized above). Damage terms introduce channels: Investment reduces damage accrual (since ), which both increases immediate survival and raises later-life survivorship multiplicatively. Hence has both a direct effect on and an indirect effect on through . Compared with the minimal of section 2.1, this treatment makes explicit the mechanism of what we may refer to here as “health” or “survival capital”: investments reduce cumulative damage (or slow down the rate of wear and tear), thereby raising the realized fitness payoff to complementary investments later.
In this extension, survivorship is decomposed into both a “trait-mediated” component and a “cumulative-damage” component: investments both raise instantaneous survival and reduce damage accrual. Because later-stage payoffs are multiplied by the probability of reaching those stages, the model illustrates a sort of Becker-style complementarity and makes explicit the role of “survival capital” in amplifying the returns to investment across traits affecting health. Thus, introducing damage accumulation bolsters our earlier conclusion: investments at one life stage increase the marginal payoff of investments at other stages, naturally giving rise to a notion of survival capital that can be leveraged across time.
2.3. Tradeoffs Between Survival Capital and Reproduction
It is important to note that survival capital is not a “free lunch” but must be purchased at the expense of immediate reproduction. With the synergy of survival capital established, we now address the central life-history problem: the trade-off between current reproduction and the longer-term compounding of fitness through health. This section identifies the first-order conditions that govern how an organism splits its finite 'effort.' To explore how the complementarity logic interacts with explicit trade-offs between spending on reproduction and investing in survival, consider again an organism with two stages at which it can invest in survival and reproduce (ages 1 and 2). As in 2.1, let and again denote the probabilities of surviving through age 1 and 2, respectively, and and the age-specific fecundities. Lifetime expected reproduction is as before:
Rather than treating and as independent parameters, however, suppose that both depend on how much of its finite resources it allocates between invstment in survival and on reproducing at each stage. Let total avialable effort at each age be normalized to 1, and let denote the proportion of resources invested in survival, and be the proportion devoted to reproduction. We assume a simple functional relationship linking these to survival and fecundity as follows:
Where represent the efficiencies of investment into survival and reproduction, respectively. These can be thought of as the biological conversion rates of effort into survival probability or fecundity. Substituting into the expression for , lifetime reproductive fitness becomes:
This comprises two distinct intertemporal effects. One is early investment which affects both current reproduction (via ), and future reproduction (via survival to period 2). The other is related to later investment , which affects only the second term, but the marginal benefit of depends directly on the level of . In other words, there are two types of “spillover effects” happening across time. First, what the animal invests early in life affects both how much it can reproduce right away and how likely it is to survive long enough to reproduce when older. Early effort has both an immediate payoff and a delayed one. Secondly, the value of investing later in life depends on what the animal did earlier, so if it invested more in surviving the first stage, then later investments become more worthwhile.
To find the optimal allocations, we take the partial derivatives of with respect to and and setting to zero. For :
And for :
From the second condition, . substituting into the first condition:
This captures the complementarity feedback. The optimal amount of early-life investment increases with the efficiency of later survival and as the fertility potential at older age. When reproduction later in life is valuable or achievable (large ), selection favors greater early survival investment because surviving to that age enables substantial returns in fitness. If, however, is small, then approaches 1/2 or less, and selection may favor diverting more effort toward reproduction at age 1. Obviously, in extreme cases (where is roughly zero), selection maximizes immediate reproduction with minimal survival investment.
This extension points to a kind of threshold characteristic. When later life fertility and survival are both sufficiently rewarding, survival investments become mutually supportive across stages. But when fertility later in life is negligible, survival and reproduction become substitutes rather than complements, producing classic life-history tradeoffs (i.e., making it a matter of choice between early versus late effort). This model therefore unifies two regimes of life-history under the same framework. The Becker-Hamilton complementarity operates when the marginal value of later reproduction is high enough to induce reinforcing survival investments, while traditional G. C. Williams-style tradeoffs emerge when later payoffs are discounted or unreliable. In this way, our logic does not replace life-history tradeoffs, but points to a wrinkle where intertemporal feedback (positive or negative) depends on the relative magnitude of late-life reproducing potential.
This analysis helps to clarify how complementary and substitutive life-history strategies emerge from the same underlying allocation problem. When later survival and fecundity are sufficiently worthwhile, early-life investment in survival capital is favoured because it unlocks access to future payoffs — producing an iterative reinforcement of longevity investments across life stages. Biologically, this corresponds to “slow” life-history strategies in stable environments where delayed reproduction yields higher returns (e.g., primates, large mammals, and many long-lived birds). Figure 3 depicts the requirement for this complementary regime to be sustainable. The figure contrasts the efficacy of investment in health or maintenance under three hypothetical scenarios. Unlike a “substitutes” model (which shows diminishing, concave returns), or a simple linear model, we predict a synergistic window (within which is a steep, locally convex region). This represents the notion that marginal returns are maximised only after a threshold level of investment is made, enabling a system to reach a superior biological potential (higher asymptote). This non-linear efficacy is a necessary condition for our inter-survival investments to amplify and stabilize a “slow” life history strategy.
Conversely, when the potential for future reproduction is low — such as might be the case if later survival is uncertain or late fertility yields minimal benefits — selection shifts toward front-loading reproductive effort. Organisms then invest less in maintenance and more in immediate fecundity, consonant with “fast” strategies such as semelparous insects or annual plants. In this regime, early survival and later survival investments become substitutes rather than complements.
Thus, this Becker–Hamilton framework naturally generates a continuum of life-history strategies without invoking separate evolutionary mechanisms for aging or reproductive timing. Complementarity in survival investments and classic reproductive trade-offs appear as quantitative outcomes of the same principle: the intertemporal feedback between the value of survival and the value of reproduction. Intrinsic differences in factors determining which pathway a species evolves are treated here as given. We next consider how environmental perturbances might push a species one way or another through impacting the quantity of resources organisms have available for making these trade-offs between survival and reproduction.
2.4. Multiple Stage Tradeoff Model and Response to Resource Effects
In this part, we explore how these evolved strategies respond to environmental shifts. To do this, we begin by setting up the 2-stage allocation problem and derive the first order conditions for early survival and investment to give a simple case showing threshold/bidirectional effects of exogenous shifts in resources. This enables us to see how exogenous resource shocks — such as a sudden abundance of food — impact optimal survival investment. This analysis demonstrates that the relationship between resources and lifespan may not be universal but instead determined by whether an organism’s strategy favours immediate reproductive reinvestment or long-term somatic reinforcement.
Again, we have two stages, . At each stage the organism has resources which it divides between survival investment and reproduction :
Stage 1 resources are exogenous baseline , stage 2 resources increase with survival of stage 1:
Where is the stage 1 effective survival probability (it can include multiplicative damage terms). Survival to stage 2 is , and stage 2 survival given investment is . Lifetime expected reproduction is:
The decision variables are , , and (implicitly, , ). Constraints are coupled because depends on . The Lagrangian is:
Taking derivatives (as done in the multi-stage case) gets the multipliers:
The first order condition important for early survival investment (after collecting terms and substituting multipliers) is:
Where and . This has the same intuitive interpretation mentioned earlier: organisms ought to invest in early survival until the marginal increase in surviving to stage 2 (left factor) multiplied by the total downstream value of that survival (direct continuation of reproduction plus indirect resource feedback ) equals the marginal cost (one unit of stage 1 reproduction foregone).
If we denote the left-hand side of the first order condition minus 1 by :
Implicit differentiation gives
Appendix A provides a full derivation of from the first-order condition, showing formally how the effect of baseline resources on early survival depends on downstream allocation and survival feedback. We then compute the numerator and then the denominator. First, the numerator:
Note here that may respond to , as . This must be solved from the stage 2 first order condition or approximated under some policy rule. Next, the denominator:
So,
This expression is of little use until we make assumptions about both the function and how responds to and . The sign of depends on the sign of in the numerator and the sign of the denominator (which will tend to be concave for most purposes). Thus, the direction of change can be either positive or negative depending on how stage 2 allocation adjusts when changes. This suggests a threshold.
To produce a relevant example, assume there is a linear effect of investment on survival with constant . Thus and . Assume also that organisms allocate during stage 2 according to a simple rule: during stage 2, a fixed fraction () of stage 2 resources are invested in survival: . So . This rule is a tractable heuristic that stands in for how organisms may split later resources between maintenance and reproduction. Let’s also assume that costs and any nonlinearities are “folded” into the choice of and for this illustrative example. Under these assumptions and . Since , the first order condition now becomes:
Or:
Collect terms in :
And therefore:
Solve for :
Now we take derivative with respect to to find the effect of exogenous resource shocks during early life:
The key point is that how stage-2 resources are allocated (captured by ) determines the sign. This means that, if then and . That is: in this simple linear example, increasing baseline resources reduces optimal early survival investment . This makes intuitive sense: extra baseline resources mainly increase later reproduction (since ), which reduces the marginal value of spending scarce early resources on survival. Organisms thus shift effort toward immediate reproduction (or allow stage-2 resources to be spent in later stages), thereby reducing . If, however, is exactly 1 (all stage-2 resources go to survival, i.e. ), then and in this linear special case.
Increasing or (more feedback or more efficient conversion of early investment into survival) reduces the magnitude of the negative derivative, that is, when feedback is stronger or early investment more effective, the downward effect of increasing on is smaller.
Thus, even in this simple analytically tractable case the sign is not universal. The effect of a resources shock depends on how later resources are spent. In particular, if later resources are channelled chiefly into reproduction (small ), an increase in baseline resources encourages more reproduction and thus less early survival investment; if later resources are directed mainly into survival (large ), baseline increases do not depress early survival investment and may even support it (in richer parametrizations of and non-linear cases one can get ).
Figure 4.
Paradoxical Impact of Enrichment. The optimal response of early survival investment to an increase in baseline resources depends on the downstream allocation policy. Organisms with a “reproductive bias” (low θ) primarily allocated resources to fecundity, and so increased baseline resources reduce the marginal value of survival, leading to a negative reaction norm (lower investment in longevity). Meanwhile, organisms characterized by higher θ according to the linear model, primarily reinvest future resources into maintenance, and so the negative effect of an increase in resources is neutralized (flat slope). Also depicted is an imagined non-linear regime where survival efficacy is synergistic, making the reaction positive (dashed line) predicting that resource abundance actively drives the evolution or development of an extended healthspan.
Figure 4.
Paradoxical Impact of Enrichment. The optimal response of early survival investment to an increase in baseline resources depends on the downstream allocation policy. Organisms with a “reproductive bias” (low θ) primarily allocated resources to fecundity, and so increased baseline resources reduce the marginal value of survival, leading to a negative reaction norm (lower investment in longevity). Meanwhile, organisms characterized by higher θ according to the linear model, primarily reinvest future resources into maintenance, and so the negative effect of an increase in resources is neutralized (flat slope). Also depicted is an imagined non-linear regime where survival efficacy is synergistic, making the reaction positive (dashed line) predicting that resource abundance actively drives the evolution or development of an extended healthspan.
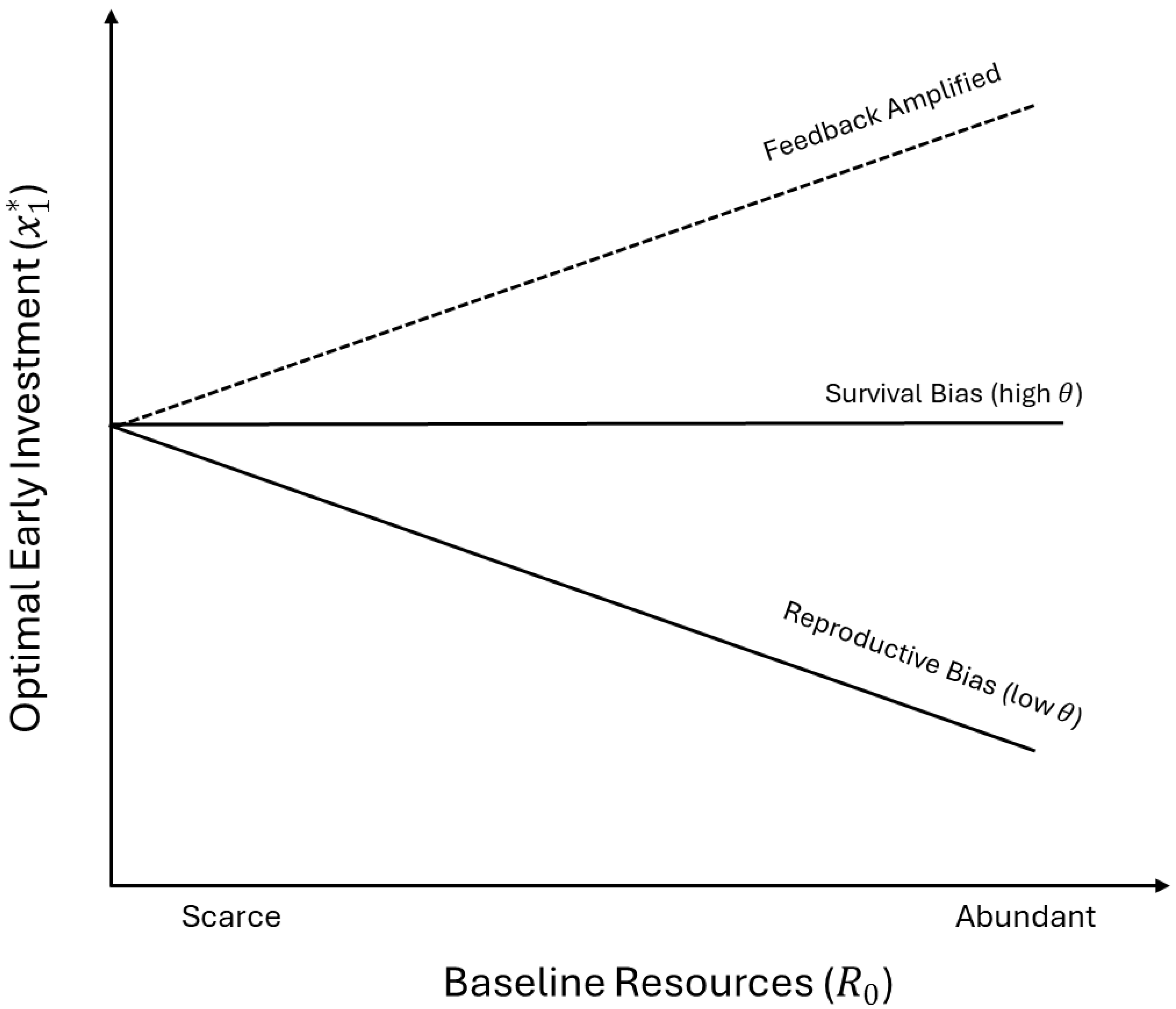
What this demonstrates is that threshold / bidirectional behaviour is likely and simple. The multi-stage model yields situations where the same beneficial environmental change (raising baseline resources or lowering extrinsic mortality) can either lower early investment in survival (shifting organisms to earlier reproduction), or raise early investment (shifting toward longevity), depending on how later resources are used and how effective survival investments are. It also shows that which regime arises depends on parameters that are, in principle, all biologically interpretable, namely (1) how much of later resources are invested into survival versus reproduction (plastic allocation rules); (2) strength of resource feedback from survival; (3) efficiency with which early investment raises survival (survival capital productivity); (4) the curvature of (nonlinearity) and the stage-2 optimization rule for (whether is chosen optimally rather than fixed fraction) — both can flip the sign in richer models. In other words, if gains in baseline resources primarily enable immediate reproduction (organisms “use” extra resources to breed more), then selection favours shifting current resources away from costly early maintenance toward reproduction. Conversely, if extra resources are funnelled into maintenance (either because organisms or ecology favour investment into survival, or because survival investments are highly productive), additional resources increase the returns to early survival, and organisms respond by investing more in longevity. An interesting question that we next turn to is whether this effect can increasingly extend lifespan when both the quantity of resources an animal can obtain and the number of stages they can survive are determined by the individual’s decisions.
2.5. Endogenizing Stages and Resources
While the previous section analysed how fixed biological policies respond to external resource shocks, it treated the number of life stages as a static constraint. In this final part of the model, we lift that restriction by endogenizing the life course itself. By allowing survival at one stage to generate the very resources required to reach the next, we uncover a critical threshold where life history shifts from a story of inevitable decay to one of self-amplifying longevity.
For stages ., the organism determines survival investment and reproduction given the budget available at each the stage :
The probability of surviving through stage (i.e. surviving from to ) is
with . For simplicity, we treat the survival “technology” as stationary across stages; generalisation to stage-dependent is straightforward. Surviving a stage gives a resource payoff that contributes to the next stage’s budget. A simple rule to capture this is:
Where is a baseline per-stage endowment (e.g. parental gift, minimal foraging), and converts survival at a stage into extra resources available next stage (experience, territory, reputation, accumulated stores, social position, etc.) Notice that this is the same feedback idea used earlier but written stage-by-stage.
Next, we treat a stage as evolutionarily meaningful (i.e. the individual has a real reproductive opportunity) if its budget exceeds a minimal viability threshold . Thus, the realized number of stages is
If the individual is effectively unable to mount any meaningful reproductive or survival investment that would constitute an additional and stage that is meaningful. Contributing to lifetime fitness, reproductive output per stage is . The expected lifetime reproduction, then, is:
Where, as before, each stage’s reproduction is weighted by cumulative survival to that stage. To identify conditions where we might get runaway evolution of extended lifespan, we want to derive a simple condition that tells us when a small improvement in survival investment (or in efficiency) can generate self-amplifying resource gains that produce more stages.
Consider a representative stage and linearise the effect of a small increase in investment around a baseline strategy. The logic of this is that a small increment in effective resource-equivalent value at stage produces some increment at stage and, if , then the effect amplifies and can grow geometrically, unlocking many additional stages. To find this, let denote the marginal survival gain per unit of extra investment at the baseline, and represents the marginal downstream value (or “shadow price”) of a small unit increase in resources at the next stage, measured in units of expected lifetime reproduction. The term equals the derivative of lifetime fitness with respect to a unit increase in (holding other decisions fixed). Leaving the same as above, we start with a small one-unit increase in effective resource-like value at stage (for example, because is slightly higher or because “technology” improved). That leads to a change in survival at stage of approximately . This extra survival then also increases the expected resources available next stage by . The downstream value of that resource increment is logically equal to times the resource increment. Therefore, the net downstream value increase is approximately
If the multiplicative factor3 is
and is greater than , then and the perturbation grows from stage to stage, with small improvements becoming amplified. Here, the notation “” is used to indicate a definition, i.e. that the quantity on the left-hand side is defined to be equal to the expression on the right-hand side. The effect of such a multiplication of an early investment conceptually expressed in Figure 5. This also shows that, if instead , then perturbations decay. Thus, the instability / runaway criterion is that there is potentially runaway extension in the number of stages if . This is directly analogous to the ecological/bio-mathematical notion of a reproduction number producing growth (e.g., Lewis, Shaui, & Driessche, 2019). Appendix B provides the linearization and matrix derivation underlying this stage-extension criterion, showing that perturbations amplify across stages when the product of resource feedback, survival efficacy, and investment responsiveness exceeds unity. Here, the product combines resource-feedback strength (), the effectiveness of survival investment (), and the marginal fitness worth of resources later (). This makes sense because, if survival investment is very effective (large ), a small extra investment can produce a large increase in survival probability. That may, in turn, produce a relatively large resource gain () for the next stage. If extra resources are very valuable (large ) such as if they enable much extra reproduction or further feedback, the loop has strong leverage. Moreover, if resource feedback is too small (surviving gains very little additional resource), no amplification occurs.
2.6. Endogenizing Stages and Resources
While the previous section analysed how fixed biological policies respond to external resource shocks, it treated the number of life stages as a static constraint. In this final part of the model, we lift that restriction by endogenizing the life course itself. By allowing survival at one stage to generate the very resources required to reach the next, we uncover a critical threshold where life history shifts from a story of inevitable decay to one of self-amplifying longevity.
For stages ., the organism determines survival investment and reproduction given the budget available at each the stage :
The probability of surviving through stage (i.e. surviving from to ) is
with . For simplicity, we treat the survival “technology” as stationary across stages; generalisation to stage-dependent is straightforward. Surviving a stage gives a resource payoff that contributes to the next stage’s budget. A simple rule to capture this is:
Where is a baseline per-stage endowment (e.g. parental gift, minimal foraging), and converts survival at a stage into extra resources available next stage (experience, territory, reputation, accumulated stores, social position, etc.) Notice that this is the same feedback idea used earlier but written stage-by-stage.
Next, we treat a stage as evolutionarily meaningful (i.e. the individual has a real reproductive opportunity) if its budget exceeds a minimal viability threshold . Thus, the realized number of stages is
If the individual is effectively unable to mount any meaningful reproductive or survival investment that would constitute an additional and stage that is meaningful. Contributing to lifetime fitness, reproductive output per stage is . The expected lifetime reproduction, then, is:
Where, as before, each stage’s reproduction is weighted by survival to that stage. To identify conditions where we might get runaway evolution of extended lifespan, we want a simple condition that tells us when a small improvement in survival investment (or in efficiency) can generate self-amplifying resource gains that produce more stages.
We can take a representative stage and linearise the effect of a small increase in investment around a baseline strategy. The logic of this is that a small increment in effective resource-equivalent value at stage produces some increment at stage and, if , then the effect amplifies and can grow geometrically, unlocking many additional stages. To find this, let denote the marginal survival gain per unit of extra investment at the baseline, and represents the marginal downstream value (or “shadow price”) of a small unit increase in resources at the next stage, measured in units of expected lifetime reproduction. The term equals the derivative of lifetime fitness with respect to a unit increase in (holding other decisions fixed). Leaving the same as above, we start with a small one-unit increase in effective resource-like value at stage (for example, because is slightly higher or because “technology” improved). That leads to a change in survival at stage of approximately . This extra survival then also increases the expected resources available next stage by . The downstream value of that resource increment is logically equal to times the resource increment. Therefore, the net downstream value increase is approximately
If the multiplicative factor4 is
and is greater than , then and the perturbation grows from stage to stage, with small improvements becoming amplified. Here, the notation “” is used to indicate a definition, i.e. that the quantity on the left-hand side is defined to be equal to the expression on the right-hand side. The effect of such a multiplication of an early investment conceptually expressed in Figure 5. This also shows that, if instead , then perturbations decay. Thus, the instability / runaway criterion is that there is potentially runaway extension in the number of stages if . This is directly analogous to the ecological/bio-mathematical notion of a reproduction number producing growth (e.g., Lewis, Shaui, & Driessche, 2019). Appendix B provides the linearization and matrix derivation underlying this stage-extension criterion, showing that perturbations amplify across stages when the product of resource feedback, survival efficacy, and investment responsiveness exceeds unity. Here, the product combines resource-feedback strength (), the effectiveness of survival investment (), and the marginal fitness worth of resources later (). This makes sense because, if survival investment is very effective (large ), a small extra investment can produce a large increase in survival probability. That may, in turn, produce a relatively large resource gain () for the next stage. If extra resources are very valuable (large ) such as if they enable much extra reproduction or further feedback, the loop has strong leverage. Moreover, if resource feedback is too small (surviving gains very little additional resource), no amplification occurs.
3. Discussion
We have explored a very different way to looking at aging from both candle and oil lamp approaches, with health seen as a form of multifaceted “survival capital”. On this view, expenditures on health like growth and repair are not merely drains on a fixed amount of life. Rather, they can alter the selective environment and increase the value of further investments. Just as maintenance of the engine in a car makes it more worthwhile to fix the brakes, replace old tyres, change the oil, learning to evade predators more effectively can, for instance, make it more worthwhile to also invest in long-term disease resistance, and vice versa.
The idea at the heart of this is that investments in health are often complementary: improving one aspect of survival raises the marginal value of improving others. Our model’s purpose is to make that intuition rigorous — to show how the survival curve itself can be reshaped endogenously by mutually reinforcing investments, rather than being viewed as a fixed slope set once and for all by declining selection pressure. In other words, complementarity means the marginal fitness return to investing in trait A increases with the level of trait B — formally the cross-partial of fitness with respect to the two investments is positive. That small statement carries a lot of weight. When late-life payoffs are small, the classic “fast” strategy can emerge: investments in early survival and early reproduction act more like substitutes, and selection favours front-loaded reproduction (recovering the familiar Williams/Hamilton intuition). But when later payoffs are substantial, the system flips: early survival and later maintenance are complements — investing early unlocks value later, and so selection favours a “slow” life history that concentrates on longevity and sustained performance. The model therefore unifies these regimes as two ends of a single allocation problem, rather than as competing, unrelated explanations.
For brevity, we hereinafter refer to this perspective as the Synergistic Survival Capital (SSC) model. We adopt this name because the core mechanism is that survival investments function like capital: gains in one domain raise the marginal returns to investments in all complementary domains. These synergies reshape the selective landscape, creating conditions under which survival, health, and future resource prospects become mutually reinforcing rather than zero-sum. The SSC model thus provides a unified, mechanistic way to understand how organisms can evolve an extended healthspan, threshold-like transitions in life-history strategies, and coordinated maintenance across multiple biological systems.
The most novel prediction is a simple, tester-friendly threshold: survival extension can self-amplify when a single multiplier exceeds unity. Think of a small investment in maintenance as producing three linked effects: (1) it raises the probability of surviving to the next stage, (2) surviving to the next stage tends to raise the resources available then (territory, status, stores), and (3) those additional resources make further survival investments more profitable. Multiply those three sensitivities together (the resource feedback from survival, the efficacy of survival investment, and the marginal fitness value of the unlocked resources) and we obtain a single number we have denoted as . If , a small improvement at one stage produces a bigger improvement at the next, and the process can grow geometrically: a little longevity begets more resource, which begets more longevity, and so on. If, on the contrary, , perturbations damp out and lifespan is bounded. This “runaway” criterion is both mathematically crisp and biologically interpretable: it identifies why some lineages or ecological contexts (strong resource feedbacks, high repair efficacy, steep returns to late reproduction) are predisposed to evolve extreme healthspan and lifespan, while others are not. Our framework therefore translates into testable predictions across multiple domains, including the ecological (which environments or social systems produce the resource feedbacks that favour ?), physiological (which mechanisms raise investment efficacy and thus tip the multiplier?), and life-history (where do threshold and bistability effects predict surprising switches between fast and slow strategies?).
The SSC also has implications for applied biomedicine. It warns against naive extrapolation from simple “slow the candle” interventions: increasing resource intake or reducing a single source of mortality may sometimes shorten rather than lengthen lifespan if it shifts allocation toward early reproduction unless the surrounding parameters push over the threshold. More formally, feedback increases the slope of the reproductive value function at later ages, complementarity increases the cross-partial of fitness with respect to maintenance investments, and the threshold appears only when the product of these sensitivities exceeds unity, allowing small perturbations in survival investment to compound across ages. The remainder of the discussion unpacks these implications, links them to empirical literatures in ecology and the emerging field of “geroscience”, and proposes experiments and comparative tests that will allow researchers to decide which regime (substitute, complement, or runaway) best describes the organisms they study.
3.1. Other Theories
To put the theory in the proper context it is first necessary to describe influential classic theories of aging. From this it will be clearer where they contrast in empirical predictions and that many can be seen as subsets within the SSC model’s wider parameter space.
3.1.1. Disposable Soma and Antagonistic Pleiotropy
The Disposable Soma theory (Kirkwood, 1977; Kirkwood & Holliday, 1979) argues that organisms face a strict budget constraint: every unit of resource spent repairing the body is a unit not spent on reproduction. Optimal life history is therefore characterised by a “just-enough” level of somatic maintenance to ensure reproductive success but not indefinite survival. This is emblematic of the classic candle metaphor: a finite reservoir of fuel that must be allocated between burning now (reproduction) and preserving wax (maintenance). A core prediction of Disposable Soma is that increasing investment in one domain makes investment in others less valuable — a trade-off widely documented across taxa where increased reproductive effort is linked to accelerated physiological decline (Reznick, 1985; Stearns, 1989). Maintenance traits are substitutes: more of one reduces the marginal benefit of another. Thus, organisms with heavy reproductive effort should be shorter-lived, while organisms under reduced reproductive load should become longer-lived because the saved resources can be diverted into repair.
The SSC model diverges precisely on this point. Here, investments in survival mutually enhance each other: improving one form of maintenance increases the marginal return to additional improvements elsewhere. Rather than a zero-sum allocation, the model predicts positive cross-effects: better early maintenance raises the payoff to later maintenance, and vice versa. This leads to nonlinear thresholds rather than smooth trade-offs: small increases in maintenance can cross a tipping point where self-reinforcing improvements become optimal5.
Empirically, the contrast is sharper. Disposable Soma predicts negative covariance between reproduction and maintenance6(Reznick, 1985; Stearns, 1989), and substitution among maintenance traits. The complementarity at the heart of SSC predicts positive covariance among maintenance traits and potentially a positive association between early survival investment and later reproduction if survival increases resource acquisition. While a meta-analysis of 117 mammal species confirms that preventing reproduction can extend lifespan by 10–20% by "releasing" resources (Garratt et al., 2025), the SSC model explains why this effect is highly variable and species-dependent. Removing reproduction should always extend lifespan under Disposable Soma; under complementarity, the effect appears only when the released resources push the organism across a synergy threshold or feed back into complementary investments. In short, the direction of the effect on lifespan of manipulating reproductive output on lifespan should be conditional on the resource allocation of the species organism in question.
3.1.2. Mutation Accumulation and Constraints
Antagonistic Pleiotropy (AP) posits that alleles beneficial early in life are favoured even if they impose costs late in life (Williams, 1957; 1966). Because early-life fitness is weighted heavily by selection, genes with early benefits and late penalties can spread (McAuley, 2025). The empirical hallmark of AP is age-flipped genetic effects: alleles that enhance early survival, maturation, or fecundity but accelerate physiological decline later. For example, in humans, this is evidenced by genetic variants linked to enhanced early-life immunity or fertility that simultaneously increase the risk of cancers, autoimmune disorders, and neurodegeneration in later life (Byars & Voskarides, 2020). AP treats each locus largely independently: what matters is the sign of its effects before and after reproduction. SSC alters this picture because complementarity among investments in health changes the value of late-life survival itself. If early survival investment increases the marginal returns to late survival (via synergies or resource feedback), the net selection gradient on a pleiotropic allele can be reversed. That is, an allele that would be favoured under traditional AP may become disfavoured when complementarity boosts the weight of later survival.
AP and SSC somewhat differ in their predicted population signatures. AP predicts common alleles with opposite effects at different ages, widespread trade-offs between early and late performance, and “seesaw” patterns of gene expression. SSC predicts positive covariation in survival and maintenance traits across age: individuals (or lineages) that invest effectively early tend to invest effectively late. Rather than seesaws, we expect ramps—mutually reinforcing effects across ages. Empirical evidence from human exceptional longevity supports this 'ramping' signature. Centenarian humans often exhibit superior maintenance characteristics typical of younger individuals, particularly regarding genomic stability (López-Otín et al., 2013). Rather than showing the age-flipped trade-offs predicted by AP, they possess higher constitutive levels of key DNA repair enzymes7, such as PARP1 and Ku70, compared to the general older population (Bürkle et al., 2005). This suggests that enhanced, sustained investment in genomic integrity acts as a protective, complementary trait: by maintaining the genomic 'template', the marginal benefit of other somatic repair remains high even at advanced ages. Put another way, whereas AP predicts antagonistic pleiotropic effects of genes, SSC predicts synergistic pleiotropic effects, potentially reversing the selection gradient expected by AP.
3.1.3. Life History Strategies and Senescence
Mutation Accumulation (MA) also emphasises that because selection weakens with age, deleterious late-acting mutations drift to moderate frequencies (Medawar, 1952). The signature prediction here is that there should be increasing additive genetic variance in health with age, reflecting the neutral accumulation of late-acting defects. MA expects little structured covariation among later-life traits as variation is largely stochastic.
SSC provides a contrasting mechanism. Because early survival investment can raise the value of later survival, complementarity can strengthen selection at older ages. This means that the “neutral zone” predicted by MA can shrink: late-acting deleterious alleles may no longer drift if later survival is sufficiently valuable due to synergies. Conversely, if complementarity is weak or absent, MA-like patterns would be expected to prevail. Thus, the two mechanisms can coexist, but complementarity modulates the age-structure of selection rather than accepting its decline as fixed. Empirically, MA predicts late-life traits should show high variance and weak selection signatures, while complementarity predicts coordinated selection on multi-stage maintenance systems, reduced late-life variance, and structured rather than random deterioration. Under stress (e.g., mutation accumulation lines), MA predicts earlier collapse; complementarity predicts lines with stronger early survival investment should evolve expanded later maintenance. In sum, to the degree that SSC is more important in nature relative to MA in explaining patterns of aging, the neutral zone theoretically predicted by MA should be empirically small.
3.1.4. Exercise and Other Elixirs
Perhaps the strongest empirical challenge to classical evolutionary theories of aging is the robust set of behavioural and social factors that increase human healthspan, despite apparently imposing substantial energetic or opportunity costs. Exercise is the clearest example: demanding physical activity should, under Disposable Soma or strict trade-off logics, divert resources away from reproduction or repair. Yet in humans and many other long-lived species, physical activity improves maintenance and slows aging. This is naturally explained by SSC, where activity functions as a cue of survival capacity and opportunity (Liberman et al., 2021), upregulating coordinated maintenance pathways. The candle-like mindset has no problem seeing the telomeres, for instance, as acting as a finite and inexorably declining limiting factor on health, that repeated cycles of cell replication “burn through”. But it would be blind-sided by the possibility that regular physical activity powerfully induces the upregulation of telomerase, and so actively replenish the telomere “counter” (this is in fact the case (Werner et al., 2019)). Indeed, candle-like thinking may predict the opposite: that exercise (or any other factor that demands increased investment in growth and repair) should burn through the fixed amount of life stuff more rapidly.
Exercise is not the only case where human healthspan correlates positively with traits that should be costly under traditional models. Social behaviours such as marriage, pair bonding, parental investment, and broader social integration are among the strongest known predictors of longevity and delayed morbidity in humans. Under a strict trade-off framework, such behaviours should divert resources toward mating effort, childcare, or social competition, leaving less for somatic repair. Reproducing early and often, in particular, should reduce maintenance if all investments draw from a single budget. There is some evidence for such negative trade-offs in short-lived, r-selected species, consistent with classical expectations. In humans, however, the pattern is often reversed: married individuals tend to live longer, have lower mortality risk, and exhibit slower cognitive decline than unmarried or childless peers (Manzoli et al., 2007; Lillard & Waite, 1995). Larger social networks predict better immune function, slower aging biomarkers, and reduced dementia risk (Holt-Lunstad et al., 2010; Steptoe et al., 2013). These associations may be dismissed as mere artefacts of healthier individuals self-selecting into marriage or social activity. But they may instead reflect a more general and deeper pattern: behaviours that indicate embeddedness in a cooperative social environment, access to future resources, or ongoing reproductive opportunity are regularly associated with enhanced maintenance rather than diminished maintenance.
SSC provides a positive and mechanistic account of these phenomena. In the model, investments in survival increase the marginal value of future survival by unlocking resources, social support, mating opportunities, and cooperative gains later in life. Signals that the environment is worth surviving for — such as stable pair bonds, active parenting, strong alliances, or deep social integration — should upregulate maintenance. Rather than drawing from a fixed pool, such behaviours increase the expected returns to living longer. Social bonds, like exercise, become indicators of favourable future conditions and thus trigger or justify complementary investments in survival. This is a distinctive prediction absent in classical theories: Social, cognitive, and behavioural traits that signal greater access to resources or more future reproductive opportunities should elevate maintenance investment and improve healthspan—even if they carry short-term energetic costs. This interpretation accommodates an array of empirical outcomes that are puzzling under trade-off theory alone: the longevity benefits of marriage, social connectivity, cooperative breeding, grandparenting (Hawkes et al., 1998; Kaplan et al., 2000), culturally scaffolded resource acquisition, and even culturally transmitted skills that increase productivity at older ages (e.g., see the “active life” hypothesis of Liberman et al., 2021). They are all behaviours that, in a complementarity framework, raise the future value of survival itself and therefore strengthen selection for extended maintenance.
3.2. Evidence and Implications
3.2.1. Resource Feedback, Survival Capital, and the Value of Living Longer
Recall that a central implication of the present thesis is that successfully surviving in any stage in life may provide more than the simple avoidance of death. In the model, each stage’s survival investment increases the chance of reaching the next stage, and that surviving stage can increase the resource budget for stage (via the feedback rule where converts survival into additional future resources). This means that early-life survival investment raises not only the chance of reaching later stages but also the value of being alive in those later stages. The marginal benefit of later survival therefore depends positively on prior survival, which is the essence of health as “survival capital.”
In the general multi-stage model (Section 2.5), the downstream reproductive value of an increment in next-stage resources is captured by the marginal fitness value , all else being equal. Likewise, the effectiveness of any investment in survival at time is captured by (the marginal improvement in stage- survival per unit of investment). A small increase in this effectiveness therefore produces a downstream benefit that is multiplied by . This determines whether the effect of an incremental survival improvement decays or amplifies across stages. Where it amplifies (), survival investments generate self-reinforcing increases in future resources and reproductive value. That is, if a bit of resources can have a large impact on fitness (high ) and if surviving a bit longer can enable a lot of extra resources to be gathered (high ), and if investment in health is effective at increasing survival (high ), we predict extremes in longevity to evolve.
How well do these theoretical assumptions compare with reality? Can we find cases where early-life advantages compound over time, producing persistent individual differences in survival, reproductive performance, and life-history trajectories? Empirically, such compounding effects appear to be the rule rather than the exception across long-lived, cognitively complex, and socially structured species. In many primates, for instance, early-life conditions (maternal care, social integration, ecological learning, etc.) have been suggested to produce long-lasting and multiplicative effects on adult performance (French & Carp, 2016. An interesting example is the spotted hyena (Crocuta crocuta), where offspring of high-ranking females grow faster, begin to breed earlier, and rear more total offspring over their lifetime than those from low-ranking mothers (Holekamp et al., 2012). These “silver spoon” effects agree with SSC predictions: individuals who begin life with higher survival capital enjoy elevated for later resources and often higher effective survival efficacy because well-regulated physiology and competent behavioural repertoires enhance the payoff to subsequent investment. As the SSC model predicts, early advantages shift individuals onto trajectories where each subsequent stage becomes more valuable, increasing the returns to sustained survival.
Elephants (Loxodonta africana) illustrate a system with an unusually large feedback parameter . Each year of survival yields disproportionately valuable ecological knowledge, such as the ability of matriarchs to distinguish infrequent predator vocalizations or locate distant water sources during rare droughts (McComb et al., 2011). This means the marginal value of late-life survival rises sharply with age, and orphaning—which severs this intergenerational capital transfer—stunts growth and reduces the survival efficacy of the remaining kin (Parker & Wittemyer, 2022). Because these knowledge gains are frequently realized only in extreme environmental states (e.g., multi-year droughts), their payoff function is highly convex. This means the marginal value of late-life survival rises sharply with age, and that the compounding benefits of accumulated knowledge imply sustained or rising survival efficacy across adulthood. In our terms, elephants lie in the parameter region where so that selection favours unusually long adult survival because many of the highest value returns on survival capital are realized only very late in life.
Similar patterns would be predicted in other species wherever knowledge and social structure play an important role that spikes during periods of intense survival challenge. For example, meerkats (Suricata suricatta) are animals that live in groups and cooperatively breed. Social interactions powerfully influence their ability to deal with challenging environmental conditions, with more interactions (i.e., larger group size) reducing likelihood of mortality during crises (Clutton-Brock et al., 1999; Silk, 2007). Given meerkats are less than 1 kg, sexually mature in their first year of life, and reproduce repeatedly over many years, this places them quite firmly in the “slow” end of the traditional life-history spectrum, consistent with our predictions that they should be relatively long-lived. For comparison, they tend to live to about 5-15 years in the wild, while similarly sized insectivorous mammals like rodents rarely live for more than 3 years in the wild.
Several avian lineages, notably parrots, corvids and some long-lived seabirds, also combine large brains plus long lifespans and extended developmental periods, during which juveniles remain dependent on parents and have prolonged opportunities for learning. Comparative work in parrots shows that species with larger brains relative to body size also have greater life expectancy, and that many parrot species have juvenile periods of up to about six years, which are thought to facilitate extensive social learning and skill acquisition (Smeele et al., 2022). In corvids, phylogenetic analyses likewise link larger relative brain size, extended development and prolonged parental care to the evolution of advanced cognitive abilities, with extended parenting proposed as a key facilitator of cognitive skillsets that support flexible responses to environmental challenges (Griesser et al., 2017). More broadly, across birds, larger relative brain size and higher rates of behavioural innovation are associated with greater exposure to environmental variability and with success in coping with novel or fluctuating conditions, suggesting that cognition and extended learning can be favoured in variable or challenging environments.
Some cetaceans illustrate the same principle even more strikingly, where the ultimate expression of survival capital may be the evolution of a prolonged post-reproductive lifespan (“grandmother” effects) (Foster et al., 2012). For example, post-menopausal killer whales (Orcinus orca) provide crucial leadership and foraging knowledge during periods of resource scarcity. The loss of such individuals produces dramatic spikes in mortality risk for their kin: a ~6.7-fold increase when a post-reproductive mother dies, and ~4.5-fold when a non-reproductive grandmother dies (Nattrass et al., 2019). These effects intensify in low-salmon years, indicating that the marginal fitness value of their knowledge is sharply crisis-dependent. In SSC terms, the fitness value of knowledge manifested in grandmothers is an instance of a sharply increasing for older ages, since the benefit of an additional year of survival increases precisely when ecological conditions deteriorate. Because each additional year of survival increases both the stock of knowledge and the contexts in which that knowledge pays off, the system exhibits a strongly convex resource feedback function. In SSC terms, this corresponds to an elevated and a large , causing . Under these conditions, a prolonged post-reproductive lifespan becomes an adaptive allocation that maximizes the high marginal value of survival at older ages. In other words, because these benefits cannot be fully realized without extended survival, natural selection favors unusually long lifespans as the mechanism enabling individuals to accumulate and deploy this capital8.
Although our minimal formal model does not explicitly represent ecological, cognitive, or social “capital” as a state variable, these forms of accumulated advantage map cleanly onto the model’s parameters. Any process that raises the fitness value of surviving into later stages (knowledge acquisition, social integration, territory control, cooperative alliances, or intergenerational skill transfer) increases , the marginal reproductive value of later life. Likewise, processes in which survival generates access to additional resources or opportunities inflate , the resource–survival feedback elasticity. Together, these forces determine the multiplier . The SSC model therefore predicts that taxa experiencing (i) substantial accumulation of survival-relevant capital across age, (ii) strong resource feedback from continued survival, or (iii) high marginal reproductive value of later stages, should exhibit the strongest complementarity among maintenance traits and be most likely to exceed the threshold . Elephants, killer whales, many primates, and certain long-lived birds are canonical cases: their social learning, ecological knowledge, and interdependent group structures generate precisely the capital-dependent increases in and that favour extended maintenance and extreme healthspans. Where resource acquisition, social status, or ecological knowledge accumulate synergistically, longevity ceases to be a mere byproduct of low extrinsic mortality and instead becomes an adaptive strategy for harvesting the increasingly valuable returns on early-life investments. In this context, Survival Capital functions as a specialized form of “Fitness Capital” — an accumulated stock of biological assets that can be augmented through both individual resource acquisition and, in social species like the orca, through altruistic investment from kin (Roy, 2026). This is the hallmark prediction of the SSC framework and a key domain in which it offers explanatory power beyond classical aging theories.
3.2.2. Physiological Synergy and Survival Efficacy
An important assumption of the SSC framework is that investments in survival traits exhibit strong complementarity: the marginal return of one maintenance system depends positively on the state of others. This concerns the internal survival efficacy parameter , which captures how effectively investment translates into improved survival probabilities. High implies that health-related traits are deeply interconnected where improvements in one domain amplify the effect on mortality risk of improvements in others. This “maintenance gap” (the distance between the actual and potential somatic health) suggests that selection for longevity is fundamentally a selection for the integration of interacting physiological traits (Wensink et al.,2012). Evidence for this underscores our more general view that each unit of investment in survival — whether metabolic, behavioural, social, or physiological — can generate a disproportionately large reduction in mortality risk. When combined with the feedback parameter and the marginal value of future resources , high values of can elevate the SSC multiplier above unity, generating positive feedback across life stages.
Natural systems provide abundant evidence of such synergistic physiological architectures, where health-related traits form deeply interconnected modules. In these cases, improvements in one domain (e.g., immune response) amplify performance in others (e.g., tissue repair, thermal tolerance), causing the payoff to survival investment to be likely multiplicative rather than additive. Many vertebrates exhibit highly integrated systems where somatic maintenance mechanisms cooperate to maintain low mortality even under ecological stress, such as DNA repair, immunocompetence, stress resistance, and metabolic buffering (López-Otín et al., 2013). When these systems reinforce one another, investment in one mechanism effectively enhances several. This biological interdependence matches the theoretical structure of a high survival efficacy : the derivative becomes large because the investment has cross-domain effects.
Across molecular, cellular, and organismal levels, key maintenance pathways, such as mTOR, AMPK (AMP-activated protein kinase), Sirtuins, mitochondrial quality control, immune surveillance, form a conserved and functionally integrated network. These systems do not operate in isolation. Rather, each sets the preconditions for the others to function effectively. For example, AMPK governs mitochondrial quality control, establishing the ATP supply and redox balance required for virtually all energy-intensive maintenance processes (Koklesova et al., 2022), effectively serving as a "universal resource booster" (Herzig & Shaw, 2018). Furthermore, AMPK suppresses oxidative stress by upregulating antioxidant gene expression, a highly complementary effect that preserves the functional capacity of damage-sensitive components like DNA and lipids (Wu & Zou, 2020). This bioenergetic rebalancing has broad, clinically validated benefits; for instance, Metformin (a partial AMPK activator) exhibits pleiotropic protective effects against cardiovascular disease, cancer, and neurodegeneration, confirming that targeting bioenergetic homeostasis is a highly effective, complementary investment in survival (Froldi, 2024; Prattichizzo et al., 2018).
Sirtuins, particularly SIRT1 and SIRT6, link cellular energy status to genomic stability (Chang & Guarente, 2014): SIRT1-mediated deacetylation is required to activate SIRT6’s DNA repair functions, meaning that genomic maintenance is gated by metabolic state. They serve as a mechanistically quantifiable example of molecular complementarity. For instance, SIRT1 and SIRT6 interact synergistically to recognize DNA breaks and potentiate the DNA damage response; SIRT1-mediated deacetylation is often a prerequisite to activate SIRT6’s repair functions (Meng et al., 2020). Because Sirtuins and other repair enzymes like PARPs compete for a shared pool of NAD+, investment in NAD+ bioavailability yields amplified, complementary returns across both metabolic regulation and DNA repair kinetics (Barcena et al., 2024; Yu et al., 2024). In the SSC model, this represents a high : a single investment (in NAD+ or SIRT activation) produces system-wide gains in both cardiovascular stability and genomic integrity (Qiu et al., 2023).
Experimental studies also repeatedly demonstrate supra-additive effects when complementary maintenance pathways are co-targeted. In model organisms, drug combinations that simultaneously engage distinct longevity pathways extend lifespan far beyond the sum of the individual components. In C. elegans, drug combinations that simultaneously engage distinct longevity pathways (such as IGF and SREBP signalling) extend lifespan far beyond the sum of individual components, recruiting new regulatory modules for lipid metabolism and stress response that are not activated by single interventions (Admasu et al., 2018).
A parallel line of evidence emerges from studies of immunometabolism. T-cell function is strongly constrained by mitochondrial health. In metabolically repressed environments, such as the microenvironment of tumours, exhausted T cells respond weakly to immune checkpoint blockade because their mitochondrial systems are operating below the threshold required to mount an effective proliferative response. Augmenting metabolic capacity with 4-1BB agonism to engage PGC-1α-mediated mitochondrial biogenesis can be seen in our terms as functionally a foundational investment. When combined with PD-1 blockade, the result is robust, quantitatively synergistic antitumor immunity (Menk et al., 2018): the metabolic intervention amplifies the marginal efficacy of the immune intervention. A brief “metabolic preconditioning” window is sufficient to unlock this synergy, showing that even short-lived boosts to foundational maintenance systems can disproportionately raise the returns on downstream repair or defence mechanisms.
Distinguishing between pleiotropy and synergy helps clarify why these findings matter for evolutionary theory. “Pleiotropy” refers to the fact that key nutrient-sensing pathways (e.g., mTOR, AMPK, Sirtuins) each regulate many biological processes. “Synergy” refers to the emergent phenomenon that coordinated investment across these systems produces non-linear, supra-additive returns. The former explains why master regulators are evolutionarily conserved, but the latter explains why longevity gains require simultaneous optimization of multiple systems. Comparative evidence across primates shows that physiological dysregulation can accelerate in a coordinated fashion, indicating that vulnerabilities are themselves systemically coupled (Cohen et al., 2022). This conserved interdependence is exactly the biological structure required for a high survival efficacy parameter to be a realistic evolutionary assumption.
Such physiological synergy is well documented in birds (Holmes et al., 2001) and bats (Wilkinson & Adams, 2019) with those showing longer-than-expected lifespans relative to body size typically also exhibiting unusually low baseline mortality hazards and high repair capacities. Strikingly, rather than showing the reproductive decline predicted by classical theory, several long-lived avian species actually exhibit increased reproductive output or success as they age — a pattern that suggests heightened investment in survival may pay off by unlocking periods where the accumulation of 'capital' (such as foraging expertise or social status) can translate into greater overall reproductive output (Healy et al., 2014; Holmes et al., 2001). Each increment of investment produces improvements across multiple maintenance systems, producing precisely the kind of convex survival–investment function assumed in SSC. In other words, if there is low physiological modularity then this implies low (investment works only locally). But where there is high physiological synergy, is likely high, (investment produces system-wide gains). The empirical distribution of lifespans across taxa may be well explained by which side of this distinction they fall on.
In sum, it seems that energy production, damage repair, immune defence, and metabolic regulation form a hierarchical, mutually reinforcing health maintenance complex. Mitochondrial quality control sets the energetic ceiling for genomic repair; proteostasis capacity shapes immune efficacy; metabolic state gates DNA repair machinery; and immune-mediated clearance protects the integrity of these systems over time. Interventions that fail to account for this architecture exhibit sharply diminishing returns, whereas those that exploit it produce disproportionate gains in healthspan and survival. Taken together, the emerging picture of molecular and systemic findings provides physiological validation for the SSC’s core assumption of complementarity, and undergirds the view that longevity evolves not because a single pathway suppresses aging, but because survival capital is built by the synergistic functioning of multiple maintenance systems whose benefits compound over time.
3.2.3. Ecological Shifts and Threshold Dynamics
As we have seen, a major prediction of the SSC is that sharp qualitative shifts in life-history strategy may evolve when populations cross the critical threshold where the survival-capital multiplier exceeds unity, . This is because crossing this threshold changes the evolutionary logic of how resources should be allocated, as survival investments begin to amplify the value of future resources. This makes a “slow”, longevity-oriented strategy optimal. In contrast, when , the marginal returns of future resources remain low, favouring a “fast” strategy centred on early reproduction. Crucially, this threshold is not fixed. It shifts as the environment alters the three key parameters of resource feedback, survival efficacy, and the marginal value of future resources.
Although more work is needed to make investigation these predictions empirically, there is already some empirical evidence to show that populations can move bidirectionally across this threshold in real time. One line of support for the SSC framework comes from environments in which external mortality risks change rapidly. These contexts can reveal the behavioural and physiological signatures of the SSC threshold (the point at which crosses 1 causing organisms to switch between fast and slow life-history allocations). Crucially, multiple experimental and natural systems show that life histories adjust bidirectionally and often paradoxically, consistent with an internal allocation mechanism perhaps governed by , not simply by the external environment.
Island colonisation and invasion biology provide cases where predation or parasitism is suddenly reduced. These may be seen as exogenously increasing survival efficacy because baseline mortality is lower, making each unit of investment in other aspects of survival yield a larger marginal reduction in hazard. The reprieve from predators or parasites can also increase resource feedback because individuals may plausibly accumulate skills, energy reserves, or structural advantages more easily in a benign environment. The upshot is that, when both parameters rise, this increase in the multiplier should shift the organism from a fast to a slow allocation strategy. This shift sometimes results in "island syndrome" characterized by delayed reproduction, larger adult size, and enhanced somatic investment seen in island birds and rodents (Covas, 2012).
However, some well-documented cases show the opposite response, i.e., safer environments leading to faster life histories (Zera & Harshman, 2001). These cases in some crucial ways help validate SSC because they demonstrate that external conditions do not automatically dictate the life-history trajectory. Instead, in SSC’s terms, the organism’s internal resource-allocation policy governs where resources flow depending on the values of , , and . In general, the paradoxical outcome of faster aging or reduced investment in maintenance despite lower extrinsic mortality should occur if the resource reallocation maximizes early fitness, achieving fitness saturation rapidly. If the organism effectively maximizes its net reproductive rate by exhausting its resources early, then the marginal value of future resources at older ages approaches zero quickly due to intrinsic resource depletion rather than external risk. In other words, when freed from their old enemies, members of an invasive population may experience (1) lower defence costs (e.g., reduced calcium needed for exoskeleton thickness thus, freeing up energy for use in reproduction or somatic investment), (2) lower metabolic load (increasing baseline resources), and crucially (3) reduced marginal value of future survival (lower ) if reproductive rate is maximised by front-loading all reproduction early. This means that, even though the environment is safer, the drop in means the future is worth relatively less, making , and so the optimal policy shifts toward earlier reproduction even at the expense of longevity. The result is intrinsically truncated lifespan through accelerated maturation and early-life allocation. Thus, ecological release does not guarantee slower life histories but depends on how safety changes the marginal value of future investments.
The crustacean Daphnia (water fleas) provides a compelling experimental system to explore the rapid, plastic, and conditional nature of life-history responses to changes in extrinsic mortality risk, often mediated by predator-released kairomones (Riessen, 1999). The observed shifts in Daphnia are often complex and bidirectional, contingent not merely on the overall rate of mortality, but on the specific age or size class targeted by the predator. Equally important are cases where greater extrinsic mortality selects for longer lifespans in Daphnia, contrary to classical predictions but consistent with SSC when the internal parameters are considered. The key mechanism is a sharp increase in the marginal value of future reproduction . This occurs when: the cost of failed reproduction is extremely high, growth reduces predation risk, and energy availability is not limiting. In such cases, individuals benefit from delaying reproduction and allocating heavily to survival until they reach a “safe” size or condition that makes mating success more likely (Agrawal et al., 1999). Daphnia under Chaoborus predation behave exactly this way. Because predation on juveniles is intense, only individuals that grow rapidly beyond the vulnerable size and stage survive. Selection therefore favours delayed maturation, strong early-life survival investment, and so an extended lifespan relative to the baseline (Riessen, 1999). The SSC explanation is that predation raises survival efficacy (survival investment becomes more valuable), resource feedback (growth itself increases future survival dramatically), and especially the late-life marginal value , because maturity confers a disproportionate reproductive payoff. Together, these push above 1, generating the slow-life-history phenotype9.
Guppies provide a uniquely well-studied case of reversible movement across the threshold, because natural populations occur both above and below waterfalls that separate low-predation and high-predation environments (Fitzpatrick et al., 2014). Selection experiments and observational studies show that guppies rapidly evolve delayed senescence (Byrant & Reznick, 2004) and increased maintenance in low-predation sites, earlier reproduction, smaller offspring, and faster senescence in high-predation sites (Reznick et al., 1990; Reznick, Butler, & Rodd, 2001), and strong phenotypic plasticity in both directions. In SSC terms lower predation raises and because individuals can survive to take fuller advantage of future reproduction. Conversely, higher predation lowers because the future becomes less reliably valuable (even if survival investment remains effective). The fact that guppies flip between slow and fast strategies depending on the ecological setting demonstrates that the life-history strategy is controlled by internal allocation laws, and that these respond exactly in ways that might have been predicted by changes in the components of .
In sum, across invasive species, island fauna, experimental evolution in guppy and Daphnia systems, facts so far known appear to show life-history strategies change when the SSC multiplier crosses 1. Both directions are observed: safer environments can speed up or slow down life histories depending on how they affect , , and particularly . Finally, these responses cannot be simply explained by exogenous factors such as extrinsic mortality alone but can be easily handled if we suppose there are internal allocation rules that track the marginal return of investment. We predict that further ecological and experimental evidence will continue to support the SSC threshold prediction and validates the model’s emphasis on the joint roles of resource feedback, survival efficacy, and the marginal value of future fitness.
3.3. Limitations and Future Directions
While our framework provides a rigorous basis for intertemporal complementarity, it relies on several simplifying assumptions for mathematical transparency. We utilize quadratic cost functions and stationary parameters (such as a fixed efficiency) to isolate the mechanisms of the threshold. In biological reality, maintenance efficacy may itself decline with age as damage accrues in the very pathways responsible for repair (Vijg & Gampisi, 2008). This may be an important aspect of the feedback loop not modelled here. Furthermore, our focus is on species that exhibit high metabolic activity and social or ecological feedback loops. As such, the model does not currently describe extreme lifespans achieved through "passive" metabolic suppression or absolute "cold-storage" strategies, such as those observed in the Greenland shark (Somniosus microcephalus) (Nielsen et al., 2016), deep-sea bivalves, and many types of plant. Future theoretical work should explore whether these low-energy regimes represent a distinct evolutionary equilibrium or if the multiplier can be re-parameterized for environments where resource feedback is low but extrinsic hazard is essentially negligible.
A key simplification of the present model is the treatment of resources as a single, fungible currency (). In nature, organisms operate in a multi-dimensional nutritional landscape, where different “currencies” may contribute to the multiplier in distinct ways. Following the Geometric Framework (Simpson & Raubenheimer, 2012), we can distinguish between structural capital (e.g., protein and amino acids needed for somatic repair and enzymatic maintenance) and metabolic fuel (e.g., carbohydrates and lipids required to power these processes). If the effectiveness of survival investment () is specifically limited by protein intake, while the costs of survival are paid in energy, the runaway condition () depends on the balance of nutrients rather than total caloric abundance. In other words, while our approach assumes that the potential for self-amplifying survival capital depends on total resources, this should not obscure the fact that 'effective' resources depend on the specific balance and timing of different nutrient currencies. This implies that the 'railroad switch' to a long-lived strategy may be triggered not simply by resource volume, but by shifts in the availability or acquisition efficiency of repair-limiting nutrients. Mechanistically, the complementarity between maintenance investments appears inherently dynamic: while catabolic states (low mTORC1) are required for autophagy and the clearance of damaged organelles, the successful completion of repair and restorative synthesis requires intermittent periods of mTOR-driven anabolism (Mannick & Lamming, 2023). Efficient somatic maintenance thus relies on a complementary oscillation between these states. If repair processes are 'gated' by the synthesis of new metabolic enzymes and repair proteins, then systemic synergy is constrained by the efficiency of resource delivery and the 'metabolic rewiring' that occurs during genomic assault (Ma & Zhou, 2025).
Future extensions could explicitly model such nutrition-related trade-offs, exploring how nutrient-specific appetites such as for protein might have evolved to “prime” the survival capital feedback loop. Experiments that feed animals continuously without opportunities for metabolic oscillation, environmental challenges, or physical activity, for example, may fail to capture this dynamic. Moreover, our model highlights the fact that “r-selected” species will naturally prioritize rapid growth and early reproduction, confounding lifespan effects. Future empirical and theoretical work could therefore benefit from (1) distinguishing protein and non-protein resources, (2) implementing protocols that facilitate the translation of these resources into health (e.g., intermittent or cyclical feeding protocols that allow more optimal or natural anabolic–catabolic dynamics), and (3) using species a range of slower life history strategies, to better probe how nutrient-specific allocation rules influence the development and extension of survival capital.
The core value of this model lies in its ability to generate testable predictions regarding the shift between fast (substitutive) and slow (complementary) life histories. We emphasize that the emergence of long life should correlate not just with low predation and less exposure to disease, but with the presence of compounding resource payoffs and integrated maintenance architectures. Systematic reviews or Meta-analyses using phylogenetic comparative methods could map these ecological and social feedback scores across taxa to confirm if acts as a universal predictor of healthspan extension. Furthermore, our threshold logic predicts that life-history shifts can be non-linear and bidirectional; experimental evolution studies in the lab (e.g., using Daphnia or Drosophila) could manipulate the resource feedback of survival to observe if tipping the multiplier triggers a step-change in physiological investment.
The framework has significant implications for human geroscience, particularly in the shift from single-target therapies to multi-target synergies. If human somatic maintenance is governed by a complementary network of mTOR, AMPK, and Sirtuins, the model suggests that single-pathway interventions may exhibit sharply diminishing returns. Applied research should therefore prioritize quantifying the synergistic "cross-talk" between these pathways. If co-targeting multiple master regulators can raise integrated repair efficacy above the dampened state often associated with advanced age, it may trigger a partial restoration of the self-reinforcing maintenance state characteristic of longer-lived species.
4. Summary and Conclusions
The SSC framework departs from the classic intuition that senescence is an inevitable consequence of declining selection with age. However, it does not contradict the underlying logic of classic theory but rather offers to extend and help clarify it. For example, Hamilton’s force-of-selection schedule shows only that if the reproductive value of later life is fixed and typically low, then late-acting deleterious effects will be weakly selected and survival investment should decline with age. What it does not specify is why the value of later life must be low in the first place. The SSC model fills precisely this gap by making the value of later life endogenous: investment in health as a sort of survival capital can increase the budget of ecological, social, cognitive, or physiological resources an organism has at other stages, which in turn can increase the marginal returns to additional survival. Early investment can therefore raise the future reproductive value curve, potentially steepening it rather than letting it decline. In this regime, the logic shifts in a subtle but important way: the same sort of structure that makes early mortality so costly also magnifies the long-horizon benefits of early survival investment when complementarities are strong. Under complementarity, small improvements in early survival can compound through resource feedbacks to elevate the fitness payoffs of later ages and sustain strong selection on extended healthspan. Thus, the “oil lamp with finite fuel” metaphor are not violated; rather, the model shows that organisms can, under certain ecological and social conditions, invest to enlarge the fuel reservoir and improve its burn efficiency. SSC therefore helps to identify the conditions under which senescence is not inevitable but contingent, with candle-like patterns of aging emerging only when capital feedback is insufficient to push the survival–resource multiplier above unity.
Looked at this way, the extraordinary lifespans and post-reproductive periods observed in orcas and elephants are not biological accidents; they are the natural expression of survival capital amplification. In these species, the accumulation of social and ecological knowledge creates a sharply increasing marginal value of future reproduction, pushing the evolutionary multiplier into a self-reinforcing, 'runaway' state. This suggestion arrives at a time when traditional life-history paradigms, such as the fast-slow continuum, are being criticised by inabilities to account for the diversity and non-linearity of aging patterns across taxa (Mc Auley 2025; Stott et al., 2024). We interpret the biological reality of intertwined homeostatic networks hints that Natural Selection indeed favours coordinated investment in these systems, precisely the conditions under which survival efficacy becomes high and synergistic, generating the non-linear returns on survival that underpin extended healthspan. For applied geroscience and human longevity research, this suggests a strategic shift. If human healthspan is ultimately governed by the same intertemporal logic, then single-target interventions that act only on the 'candle' (slowing basic metabolism or targeting one isolated disease) may be intrinsically limited or even counterproductive. Instead, significant gains in somatic maintenance likely require interventions that artificially replicate high survival efficacy through molecular synergies (e.g., the simultaneous modulation of mTOR, AMPK, and Sirtuins) and enhance the feedback returns of health investment. By treating healthspan as a form of survival capital rather than a fixed stock of resilience, research can move toward a model of longevity or “Darwinian medicine” that exploits the same intertemporal complementarities that define nature's long-lived lineages.
In conclusion, the evolution of healthspan is not merely an endurance contest against entropic decay, nor is it a simple trade-off against a fixed and given amount of resource. By formalizing survival as a form of capital that builds on itself through intertemporal feedback, the SSC model offers a bridge between classical life-history trade-offs and the biological reality of synergistic somatic maintenance. It recasts longevity and the healthspan as an adaptive, endogenously shaped strategy — one that lineages actively "choose" when the returns in Darwinian fitness are worthwhile. Reframing our research questions around this logic may allow for a deeper understanding of why nature produces such vast diversity in life strategies and aid in the formulation of interventions that work with, rather than against, the grain of our physiological architecture.
Conflicts of Interest and Funding Declaration
I have no funding or conflict of interest to declare. No new data was generated in the creation of this manuscript.
Appendix A: Implicit Differentiation of Early Survival Investment
The purpose of this appendix is to show formally how the partial derivative of optimal early survival investment with respect to baseline resources can be derived from the first-order condition (FOC) using implicit differentiation. This derivation underpins the threshold/bidirectional response of early survival to exogenous resource shocks discussed in Section 2.4.
Recall from the main text that for the two-stage model, the organism chooses early survival investment to satisfy the FOC:
where is stage-2 reproduction, is the stage-2 resource budget, and captures the feedback from survival into future resources.
To find how the optimal early investment changes with baseline resources, we differentiate implicitly:
The numerator is
Since , we have:
Thus, the numerator becomes
Intuitively, this captures how an increase in baseline resources affects the downstream reproductive payoff through stage-2 investment decisions.
Meanwhile, the denominator is
Here, accounts for how early investment affects stage-2 resources and allocation. The denominator captures the curvature of the FOC, determining whether perturbations in are amplified or diminished.
Combining numerator and denominator, the derivative is
This is the general expression. The sign of depends on: (1) the numerator (whether extra baseline resources increase stage-2 reproduction net of any induced change in stage-2 survival investment ()), and (2) the denominator: concavity of the survival FOC and sensitivity of stage-2 allocation to early investment. Physiologically, this means that if an organism is 'wired' to funnel windfall resources into immediate breeding (low ), an increase in, say, environmental quality lowers the selective pressure to maintain the body, as the relative payoff of current offspring outweighs the 'survival capital' dividend. To illustrate, assume survival is linear, e.g.,
So, that ,
Assume also that stage-2 allocation is That is, the allocation of investment into survival is a fixed fraction of resources. Then,
Substituting into the FOC:
Which simplifies to
Differentiating with respect to gives
This means that, if (i.e., some stage-2 resources go to reproduction), then the derivative is negative. On this basis, providing the organism with additional resources at baseline will reduce early survival investment, as the relative benefit of survival is reduced. By contrast, if (i.e., all stage-2 resources go to survival), then the derivative becomes zero: any extra resources automatically get reinvested into survival at stage 2. Early survival already fully secures future survival, so the marginal value of additional early investment does not change. This confirms the threshold / bidirectional effect described in Section 2.4.
The implicit differentiation shows formally that the effect of baseline resources on early survival depends critically on how later resources are allocated and the productivity of early investment. The general expression allows for more complex survival functions and stage-2 policies , providing a rigorous foundation for the intuitive results in Figure 4.
Appendix B: Linearization and Matrix Derivation of the Stage-Extension Criterion
The purpose here is to show that a small perturbation in the organism’s “resource-like” state at stage will amplify across stages when a certain product of sensitivities exceeds unity. This feature provides the potential for stage-extension or “runaway” increase in realized stages/budgets that was mentioned in the main text. We derive the scalar condition first, then give a compact matrix form that generalizes to multiple state variables.
Recall from the main text that we had the number of stages indexed by , etc. At stage the organism has a resource-like state . This can be stage-specific budget representing stored capital, territory quality, skills, etc. Baseline steady state is denoted by . At stage , the organism chooses an investment (e.g., effort on survival) as a function of current state:
We assume a smooth, differentiable policy . Probability of survival through stage is a function of investment in survival capital or health:
where . Successful survivorship produces an increment to the next stage’s resource state according to a feedback rule:
where is baseline endowment organisms are born with, and parameter converts survival into next-stage resources (same notation as in main text).
We next ask what can be expected to arise from small perturbations
That is, do they decay or grow over the organism’s lifetime? Growth indicates the potential to unlock additional viable life stages and/or to amplify budgets. To answer this, we linearize about the steady state :
We define
and
Then,
The linearized forward map for small perturbations are therefore
And, to first order,
Iterating this expression over stages gives
Perturbations grow away from the steady state whenever
This gives the local amplification or “runaway” condition: If perturbations decay. If , then the linearization is neutral.
The biologically interpretation of these three components is straightforward. As in the text, represents how strongly surviving stage raises next-stage resource endowment (what we have called “resource feedback”). The term represents how effective additional investment into survival is (i.e., the marginal survival gain per unit investment). Here, . This conveys how extra resources translate into higher investment in survival (plastic or strategic responsiveness). If all three are sufficiently large so that their product exceeds 1, a small improvement can be amplified across stages: living a bit longer yields more resources, which elicit higher investment, which further increases survival, etc.
In the main text, the scalar instability condition is expressed instead as , where denotes the marginal fitness value of resources at the next stage. These formulations emphasize complementary aspects of the same feedback loop. The present appendix derives the mechanical instability condition governing state dynamics, whereas the main text presents the value-based condition in terms of lifetime fitness. Under optimal and smooth investment behavior, the policy responsiveness is itself an increasing function of , since higher fitness returns to resources induce stronger investment responses. Consequently, the criteria and are closely related and mutually reinforcing ways of characterizing the same underlying amplification mechanism.
If the organism’s state is vector valued, , representing multiple resource channels or forms of capital, and investments are likewise vector valued, , the state-transition rule generalizes to
where is an matrix mapping survival-related effects into resource increments, and returns an -vector of survival probabilities or survival contributions.
Linearizing around the steady state gives
where the Jacobian matrix is
The Jacobian captures the local sensitivity of next-stage resources to small changes in current resources; its dominant eigenvalue determines whether survival capital is dampened or amplified across stages. Here denotes the matrix of partial derivatives evaluated at , and is the Jacobian of the investment policy. The linearized dynamics exhibit exponential growth of perturbations if and only if the spectral radius of exceeds unity:
In intuitive terms, this means that if the 'best' combination of resource feedback and investment responsiveness yields a return of more than 100%, the organism's health-state will compound rather than decay. If , perturbations decay and the steady state is locally stable. The scalar case derived above is recovered as the special case in which reduces to the scalar , whose spectral radius is simply its absolute value. As a simple illustration, consider the scalar case with , , and . Then , so small perturbations grow and stage extension is dynamically feasible. By contrast, if , then , and perturbations decay. Modest changes in parameter values can therefore move the system across the threshold.
Finally, we emphasize that the condition (or ) is a local linear stability result. It identifies when cascading stage extension is dynamically possible, not when unbounded growth will occur in practice. Nonlinearities, physiological limits, ecological constraints, and saturation effects will ultimately bound resource accumulation and lifespan. The utility of the criterion lies in identifying when small innovations or environmental changes can trigger nonlinear shifts in realized stages and budgets, a question that is ultimately empirical.
Research Highlights
- Aging as endogenous survival capital, not fixed resource depletion
- Maintenance and reproduction evolve as intertemporal complements
- The > 1 threshold triggers a switch from fast to slow life histories
- Feedback synergies drive the runaway evolution of extreme healthspans
- Provides a theoretical basis for multi-target geroscience
| 1 | While we formalize the evolution of healthspan through the lens of marginal resource allocation, this optimization approach remains logically rooted in population genetics. In this framework, the marginal fitness gains represent the selection coefficient acting on an allele that shifts allocation toward maintenance. We follow the principle that natural selection produces phenotypic designs that maximize fitness subject to trade-offs and constraints (Parker & Maynard Smith, 1990). Under this view, an 'optimum' where marginal benefit equals marginal cost corresponds to a genetic equilibrium where no mutant allele for a different allocation strategy can invade the population (Maynard Smith, 1982). See Grafen (2002, 2006) for the formal correspondence between optimization programs and gene-frequency dynamics via the Price Equation. |
| 2 | Note that L1 is exactly the probability of surviving to age 1 which was p1 in the previous section, and L2 is p1p2. |
| 3 | The use of a script-M () here is intended to signal to the reader that this is a special, model-specific multiplier that encapsulates the product of three interacting forces. It separates the conceptual object “runaway survival capital multiplier” from ordinary variables or constants, reducing ambiguity. |
| 4 | The use of a script-M () here is intended to signal to the reader that this is a special, model-specific multiplier that encapsulates the product of three interacting forces. It separates the conceptual object “runaway survival capital multiplier” from ordinary variables or constants, reducing ambiguity. |
| 5 | This is a concept echoing the "unmaintainability" thresholds explored in recent systems-biology views of aging (Wensink & Cohen, 2022; McAuley, 2025). In those frameworks, senescence is viewed not merely as a consequence of resource scarcity, but as a systemic transition into a state where repair is no longer physically or informationally viable—a constraint that, in our terms, would be represented by a collapse in the survival efficacy parameter as systemic complexity or damage accumulation exceeds a critical limit. orse": an organism may evolve an inherently fragile or "unmaintainable" architecture precisely because its ecological niche disincentivizes long-term survival capital. In this sense, fragility is a designed strategy rather than an unavoidable constraint. A classic example is the Pacific salmon (Oncorhynchus spp.), which undergoes a programmed, systemic collapse—characterized by massive cortisol surges and immune failure—immediately following spawning (Dickhoff, 1989). The salmon does not simply "wear out"; it crosses a programmed unmaintainability threshold because the marginal fitness value of post-reproductive survival has been selected to zero. In short, our view is as consistent with the notion that selection “disincentivises” maintenance beyond a certain age and that complexity is then allows to become fragile or brittle than the reverse direction of causation. |
| 6 | To take a dramatic real-world example, some species of octopus are semelparous, meaning that adults die after a single bout of reproduction. It seems that the act of spawning causes female Octopus hummelincki to undergo a hormonal shift that leads to a rapid deterioration in physical health and then death (Wodinsky, 1977). This involves a sudden drop in neuropeptides and a surge in steroid signalling that causes metabolic collapse (Wang & Ragsdale, 2018). Experimentally preventing this endocrinological change by removing the optic gland can artificially extend their lifespan by several months, effectively switching the organism from a "suicide" program to something of a maintenance regime. |
| 7 | The capacity to maintain genomic stability is a central determinant of aging. Deficient DNA repair systems are tightly linked to the pathology of premature aging syndromes in both human and mouse models, as unrepaired DNA damage triggers cellular senescence and systemic dysfunction (Hoeijmakers, 2009). |
| 8 | This analytical result provides a formal mechanism for the simulation-based findings of Pang (2020), who demonstrated that late-life investment strategies in descendants incur the adaptive evolution of prolonged post-reproductive lifespans. |
| 9 | This logic of "gated" survival may provide a novel explanation for terrestrial gigantism, such as that seen in Mesozoic dinosaurs. While environmental productivity is often cited, the SSC framework suggests a "Size-Refuge" feedback loop: if predators (e.g., theropods) are gape-limited and lack the cooperative hunting strategies of modern mammalian carnivores (e.g., wolves, lions, humans), the survival efficacy of increasing body size becomes extremely high. Once an organism crosses a size threshold that effectively nullifies extrinsic predation, the marginal value of future resources spikes, justifying further massive investment in somatic capital (size and armour) over early reproduction. This creates a runaway selection for gigantism: a strategy that, in our terms, persists until the "multiplier" is disrupted by ecological collapse or the evolution of more efficient, cooperative predatory strategies that lower the survival efficacy of size. (For similar views in the explanation of large sizes among Dinosauria see Codron et al., 2012; Hone & Benton, 2005; Sander et al., 2011). |
References
- Admasu, T. D.; et al. Drug synergy slows aging and improves healthspan through IGF and SREBP lipid signaling. Developmental Cell (2018), 47(1), 67–79. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A. A.; Laforsch, C.; Tollrian, R. Transgenerational induction of defences in animals and plants. Nature (1999), 401(6748), 60–63. [Google Scholar] [CrossRef]
- Barcena, M. L.; et al. Role of AMPK and sirtuins in aging heart: basic and translational aspects. Aging and Disease (2024), 16(6), 3335. [Google Scholar] [PubMed]
- Becker, G. S. A Theory of the Allocation of Time. The Economic Journal (1965), 75(299), 493–517. [Google Scholar] [CrossRef]
- Becker, G. S. (1975); Human capital: A theoretical and empirical analysis; National Bureau of Economic Research. [Google Scholar]
- Becker, G. S. Human capital. The Concise Encyclopedia of Economics (2002), 2, 1–12. [Google Scholar]
- Bryant, M. J.; Reznick, D. Comparative studies of senescence in natural populations of guppies. The American Naturalist (2004), 163(1), 55–68. [Google Scholar] [CrossRef]
- Bürkle, A.; Beneke, S.; Brabeck, C.; Leake, A.; Meyer, R.; Muiras, M. L.; Pfeiffer, R. Poly (ADP-ribose) polymerase-1, DNA repair and mammalian longevity. Experimental gerontology (2002), 37(10-11), 1203–1205. [Google Scholar] [CrossRef]
- Byars, S. G.; Voskarides, K. Antagonistic pleiotropy in human disease. Journal of Molecular Evolution (2020), 88(1), 12–25. [Google Scholar] [CrossRef]
- Clutton-Brock, T. H.; O'riain, M. J.; Brotherton, P. N.; Gaynor, D.; Kansky, R.; Griffin, A. S.; Manser, M. Selfish sentinels in cooperative mammals. science (1999), 284(5420), 1640–1644. [Google Scholar] [CrossRef]
- Codron, D.; Carbone, C.; Müller, D. W.; Clauss, M. Ontogenetic niche shifts in dinosaurs influenced size, diversity and extinction in terrestrial vertebrates. Biology Letters (2012), 8(4), 620–623. [Google Scholar] [CrossRef]
- Cohen, A. A.; Ferrucci, L.; Fülöp, T.; Gravel, D.; Hao, N.; Kriete, A.; Varadhan, R. A complex systems approach to aging biology. Nature aging (2022), 2(7), 580–591. [Google Scholar] [CrossRef]
- Covas, R. Evolution of reproductive life histories in island birds worldwide. Proceedings of the Royal Society B: Biological Sciences (2012), 279(1733), 1531–1537. [Google Scholar] [CrossRef]
- Dickhoff, W. W. (1989); Salmonids and annual fishes: Death after sex. In ‘‘Development, Maturation, and Senescence of Neuroendocrine Systems: A Comparative Approach’’; Schreibman, MP, Scanes, CG, Eds.; [Google Scholar]
- Fitzpatrick, S. W.; Torres-Dowdall, J.; Reznick, D. N.; Ghalambor, C. K.; Chris Funk, W. Parallelism isn’t perfect: could disease and flooding drive a life-history anomaly in Trinidadian guppies? The American Naturalist (2014), 183(2), 290–300. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T.; Partridge, L. Horizons in the evolution of aging. BMC Biology (2018), 16(1), 93. [Google Scholar] [CrossRef]
- Foster, E. A.; Franks, D. W.; Mazzi, S.; Darden, S. K.; Balcomb, K. C.; Ford, J. K.; Croft, D. P. Adaptive prolonged postreproductive life span in killer whales. Science (2012), 337(6100), 1313–1313. [Google Scholar] [CrossRef]
- French, J. A.; Carp, S. B. Early-life social adversity and developmental processes in nonhuman primates. Current opinion in behavioral sciences (2016), 7, 40–46. [Google Scholar] [CrossRef]
- Froldi, G. View on metformin: antidiabetic and pleiotropic effects. Pharmaceuticals (2024), 17(4), 478. [Google Scholar] [CrossRef]
- Garratt, M.; Lagisz, M.; Staerk, J.; Neyt, C.; Stout, M. B.; Isola, J. V.; Nakagawa, S. Sterilization and contraception increase lifespan across vertebrates. Nature (2025), 1–9. [Google Scholar] [CrossRef]
- Grafen, A. A first formal link between the Price equation and an optimization program. Journal of theoretical biology (2002), 217(1), 75–91. [Google Scholar] [CrossRef] [PubMed]
- Grafen, A. Optimization of inclusive fitness. Journal of theoretical Biology (2006), 238(3), 541–563. [Google Scholar] [CrossRef] [PubMed]
- Griesser, M.; Drobniak, S. M.; Nakagawa, S.; Botero, C. A. Family living sets the stage for cooperative breeding and ecological resilience in birds. PLoS biology (2017), 15(6), e2000483. [Google Scholar] [CrossRef] [PubMed]
- Grossman, M. On the concept of health capital and the demand for health. Journal of Political economy (1972), 80(2), 223–255. [Google Scholar] [CrossRef]
- Hamilton, W. D. The moulding of senescence by natural selection. Journal of theoretical biology (1966), 12(1), 12–45. [Google Scholar] [CrossRef]
- Hamilton, W. D. (1996); Narrow Roads of Gene Land: The Collected Papers of W.D. Hamilton Volume 1: Evolution of Social Behaviour; Oxford University Press: Oxford; Volume 1. [Google Scholar]
- Healy, K.; Guillerme, T.; Finlay, S.; Kane, A.; Kelly, S.; McClean, D.; Cooper, N. Ecology and mode-of-life explain lifespan variation in birds and mammals. Proceedings of the Royal Society B: Biological Sciences (2014), 281, 1784. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R. J. AMPK: guardian of metabolism and mitochondrial homeostasis. Nature Reviews Molecular Cell Biology (2018), 19(2), 121–135. [Google Scholar] [CrossRef]
- Hoeijmakers, J. H. DNA damage, aging, and cancer. New England journal of medicine (2009), 361(15), 1475–1485. [Google Scholar] [CrossRef]
- Holekamp, K. E.; Smith, J. E.; Strelioff, C. C.; Van Horn, R. C.; Watts, H. E. Society, demography and genetic structure in the spotted hyena. Molecular Ecology (2012), 21(3), 613–632. [Google Scholar] [CrossRef]
- Holmes, D. J.; Flückiger, R.; Austad, S. N. Comparative biology of aging in birds: an update. Experimental gerontology (2001), 36(4-6), 869–883. [Google Scholar] [CrossRef]
- Hone, D. W.; Benton, M. J. The evolution of large size: how does Cope's Rule work? Trends in ecology & evolution (2005), 20(1), 4–6. [Google Scholar]
- Holt-Lunstad, J.; Smith, T. B.; Layton, J. B. Social relationships and mortality risk: a meta-analytic review. PLoS Medicine (2010), 7(7), e1000316. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, H.; Hill, K.; Lancaster, J.; Hurtado, A. M. A theory of human life history evolution: Diet, intelligence, and longevity. Evolutionary Anthropology (2000), 9(4), 156–185. [Google Scholar] [CrossRef]
- Kirkwood, T. B. Evolution of longevity. Nature (1977), 270, 301–304. [Google Scholar] [CrossRef]
- Kirkwood, T. B.; Holliday, R. The evolution of ageing and longevity. Proceedings of the Royal Society of London. Series B. Biological Sciences (1979), 205(1161), 531–546. [Google Scholar] [CrossRef]
- Lewis, M. A.; Shuai, Z.; van den Driessche, P. A general theory for target reproduction numbers with applications to ecology and epidemiology. Journal of mathematical biology (2019), 78(7), 2317–2339. [Google Scholar] [CrossRef]
- Lieberman, D. E.; Kistner, T. M.; Richard, D.; Lee, I. M.; Baggish, A. L. The active grandparent hypothesis: Physical activity and the evolution of extended human healthspans and lifespans. Proceedings of the National Academy of Sciences (2021), 118(50), e2107621118. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M. A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell (2013), 153(6), 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Zhou, S. Metabolic rewiring in the face of genomic assault: integrating DNA damage response and cellular metabolism. Biomolecules (2025), 15(2), 168. [Google Scholar] [CrossRef]
- Mannick, J. B.; Lamming, D. W. Targeting the biology of aging with mTOR inhibitors. Nature aging (2023), 3(6), 642–660. [Google Scholar] [CrossRef]
- Manzoli, L.; et al. Marital status and mortality in the elderly: a systematic review and meta-analysis. Social Science & Medicine (2007), 64(1), 77–94. [Google Scholar] [CrossRef]
- Maynard Smith, J. (1982); Evolution and the Theory of Games.; Cambridge University Press. [Google Scholar]
- Mc Auley, M. T. The evolution of ageing: classic theories and emerging ideas. Biogerontology (2025), 26(1), 6. [Google Scholar] [CrossRef] [PubMed]
- McComb, K.; Shannon, G.; Durant, S. M.; Sayialel, K.; Slotow, R.; Poole, J.; Moss, C. Leadership in elephants: the adaptive value of age. Proceedings of the Royal Society B: Biological Sciences (2011), 278(1722), 3270–3276. [Google Scholar] [CrossRef]
- Medawar, P. B. An Unsolved Problem of Biology: An inaugural lecture delivered at University College, London, 6 December 1951; H. K. Lewis & Co: London, (1952). [Google Scholar]
- Meng, F.; et al. Synergy between SIRT1 and SIRT6 helps recognize DNA breaks. eLife (2020), 9, e55828. [Google Scholar] [CrossRef]
- Menk, A. V.; et al. 4-1BB costimulation induces T cell mitochondrial function and biogenesis enabling cancer immunotherapeutic responses. Journal of Experimental Medicine (2018), 215(4), 1091–1100. [Google Scholar] [CrossRef]
- Murphy, D.; Mumby, H. S.; Henley, M. D. Age differences in the temporal stability of a male African elephant (Loxodonta africana) social network. Behavioral Ecology (2020), 31(1), 21–31. [Google Scholar] [CrossRef]
- Nattrass, S.; Croft, D. P.; Ellis, S.; Cant, M. A.; Weiss, M. N.; Wright, B. M.; Franks, D. W. Postreproductive killer whale grandmothers improve the survival of their grandoffspring. Proceedings of the National Academy of Sciences (2019), 116(52), 26669–26673. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Hedeholm, R. B.; Heinemeier, J.; Bushnell, P. G.; Christiansen, J. S.; Olsen, J.; Steffensen, J. F. Eye lens radiocarbon reveals centuries of longevity in the Greenland shark (Somniosus microcephalus). Science (2016), 353(6300), 702–704. [Google Scholar] [CrossRef]
- Pang, T. Y. On age-specific selection and extensive lifespan beyond menopause. Royal Society Open Science (2020), 7(5). [Google Scholar] [CrossRef]
- Parker, G. A.; Maynard Smith, J. Optimality theory in evolutionary biology. Nature (1990), 348(6296), 27–33. [Google Scholar] [CrossRef]
- Parker, J. M.; Wittemyer, G. Orphaning stunts growth in wild African elephants. Conservation Physiology (2022), 10(1), coac053. [Google Scholar] [CrossRef]
- Reznick, D. Costs of reproduction: an evaluation of the empirical evidence. Oikos (1985), 257–267. [Google Scholar] [CrossRef]
- Reznick, D. A.; Bryga, H.; Endler, J. A. Experimentally induced life-history evolution in a natural population. Nature (1990), 346(6282), 357–359. [Google Scholar] [CrossRef]
- Reznick, D.; Butler, M. J., IV; Rodd, H. Life-history evolution in guppies. VII. The comparative ecology of high-and low-predation environments. The American Naturalist (2001), 157(2), 126–140. [Google Scholar] [CrossRef]
- Roy, D. (2026); Investment Cascades and Fitness Capital; Preprints. [Google Scholar] [CrossRef]
- Riessen, H. P. Predator-induced life history shifts in Daphnia: a synthesis of studies using meta-analysis. Canadian Journal of Fisheries and Aquatic Sciences (1999), 56(12), 2487–2494. [Google Scholar] [CrossRef]
- Sander, P. M.; Christian, A.; Clauss, M.; Fechner, R.; Gee, C. T.; Griebeler, E. M.; Witzel, U. Biology of the sauropod dinosaurs: the evolution of gigantism. Biological Reviews (2011), 86(1), 117–155. [Google Scholar] [CrossRef]
- Simpson, S. J.; Raubenheimer, D. (2012); The nature of nutrition: a unifying framework from animal adaptation to human obesity; Princeton university press. [Google Scholar]
- Silk, J. B. The adaptive value of sociality in mammalian groups. Philosophical Transactions of the Royal Society B: Biological Sciences (2007), 362(1480), 539–559. [Google Scholar] [CrossRef]
- Smeele, S. Q.; Conde, D. A.; Baudisch, A.; Bruslund, S.; Iwaniuk, A.; Staerk, J.; Aplin, L. Coevolution of relative brain size and life expectancy in parrots. Proceedings of the Royal Society B (2022), 289(1971), 20212397. [Google Scholar] [CrossRef]
- Stearns, S. C. Trade-offs in life-history evolution. Functional ecology (1989), 3(3), 259–268. [Google Scholar] [CrossRef]
- Stott, I.; Salguero-Gómez, R.; Jones, O. R.; Ezard, T. H.; Gamelon, M.; Lachish, S.; Hodgson, D. J. Life histories are not just fast or slow. Trends in Ecology & Evolution (2024), 39(9), 830–840. [Google Scholar] [CrossRef] [PubMed]
- Vijg, J.; Campisi, J. Puzzles, promises and a cure for ageing. Nature (2008), 454(7208), 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. Y.; Ragsdale, C. W. Multiple optic gland signaling pathways implicated in octopus maternal behaviors and death. Journal of Experimental Biology (2018), 221(19), jeb185751. [Google Scholar] [CrossRef] [PubMed]
- Wensink, M. J.; Cohen, A. A. The Danaid theory of aging. Frontiers in cell and developmental biology (2022), 9, 671208. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; et al. Differential effects of endurance, interval, and resistance training on telomerase activity and telomere length among 266 healthy adults. European Heart Journal (2019), 40(1), 34–46. [Google Scholar] [CrossRef] [PubMed]
- Wensink, M. J.; et al. The maintenance gap: a new theoretical perspective on the evolution of aging. Biogerontology (2012), 13(2), 197–201. [Google Scholar] [CrossRef]
- Wilkinson, G. S.; Adams, D. M. Recurrent evolution of extreme longevity in bats. Biology letters (2019), 15(4), 20180860. [Google Scholar] [CrossRef]
- Williams, G. C. Pleiotropy, Natural Selection, and the Evolution of Senescence. Evolution. (1957), 11(4), 398–411. [Google Scholar] [CrossRef]
- Williams, G. C. (1966); Adaptation and Natural Selection; Princeton University Press: Princeton, NJ. [Google Scholar]
- Wodinsky, J. Hormonal inhibition of feeding and death in octopus: control by optic gland secretion. Science (1977), 198(4320), 948–951. [Google Scholar] [CrossRef]
- Zera, A. J.; Harshman, L. G. The physiology of life history trade-offs in animals. Annual review of Ecology and Systematics (2001), 32(1), 95–126. [Google Scholar] [CrossRef]
Figure 2.
The Becker–Hamilton Optimum Survival. The optimal early-life survival probability, is determined by the intersection of the marginal cost of somatic maintenance (MC = ) and the integrated marginal benefit (MB ). Three hypothetical MB curves are drawn: (1) MB1 is the benefit of surviving if only the first stage matters, (2) MB2 (low ) represents an animal living dangerously in stage 2, and (3) MB3 (high ) represents an animal very likely to survive through until the end of stage 2. The marginal benefit of early survival is shifted upward by the "shadow value" of expected future reproduction (), illustrating how intertemporal complementarity endogenously increases selection for early-stage maintenance, even if fecundity is held constant. The shaded rectangle represents the portion of the organism’s total lifetime reproduction unlocked by surviving the first stage and being around for stage 2. The smaller patterned rectangle depicting this for an individual that is less likely to survive until the end than another, represented by the larger, solid grey rectangle that it is subsumed by, with the difference representing reproductive opportunities lost by perhaps hazardous living. .
Figure 2.
The Becker–Hamilton Optimum Survival. The optimal early-life survival probability, is determined by the intersection of the marginal cost of somatic maintenance (MC = ) and the integrated marginal benefit (MB ). Three hypothetical MB curves are drawn: (1) MB1 is the benefit of surviving if only the first stage matters, (2) MB2 (low ) represents an animal living dangerously in stage 2, and (3) MB3 (high ) represents an animal very likely to survive through until the end of stage 2. The marginal benefit of early survival is shifted upward by the "shadow value" of expected future reproduction (), illustrating how intertemporal complementarity endogenously increases selection for early-stage maintenance, even if fecundity is held constant. The shaded rectangle represents the portion of the organism’s total lifetime reproduction unlocked by surviving the first stage and being around for stage 2. The smaller patterned rectangle depicting this for an individual that is less likely to survive until the end than another, represented by the larger, solid grey rectangle that it is subsumed by, with the difference representing reproductive opportunities lost by perhaps hazardous living. .
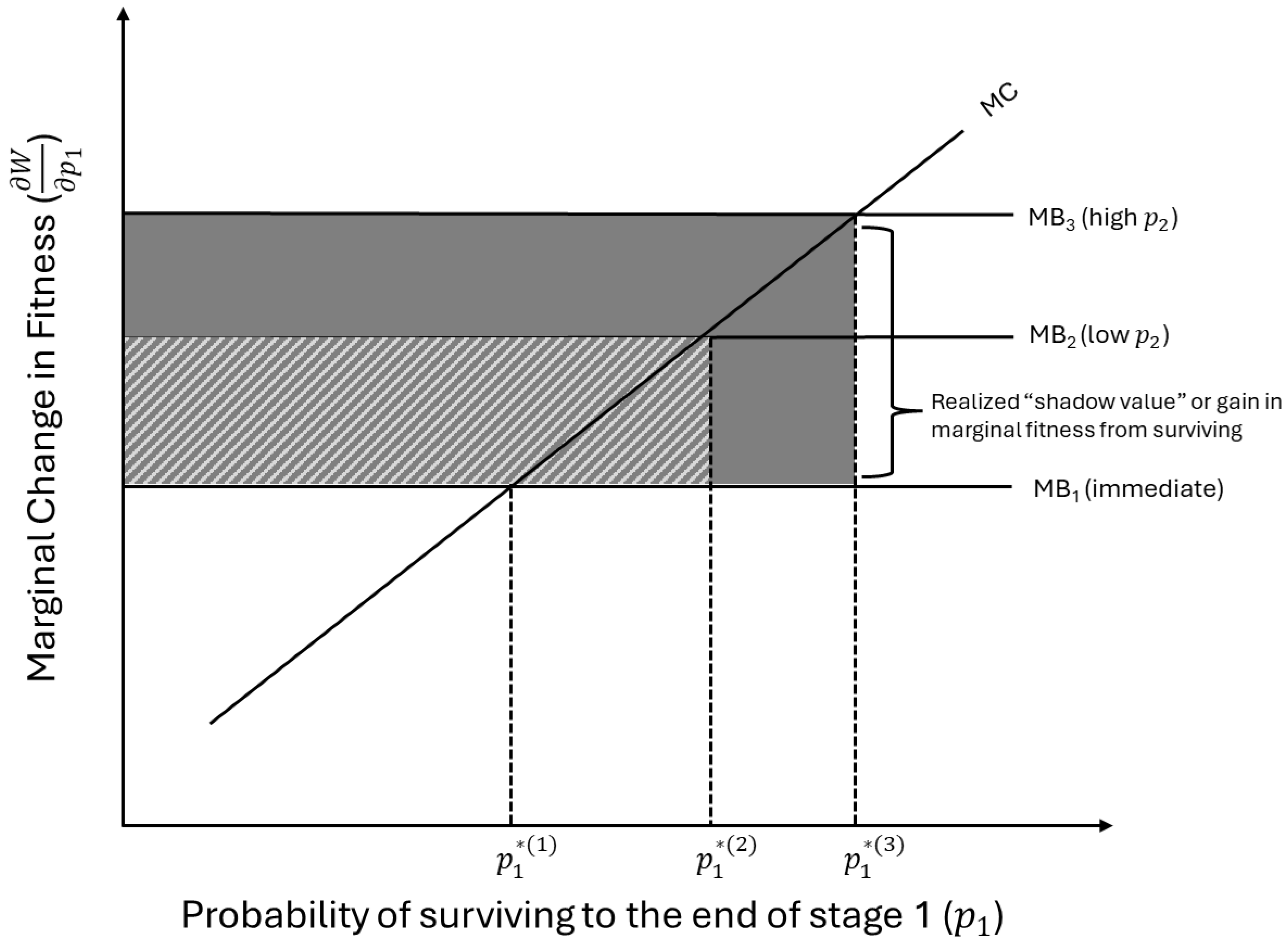
Figure 3.
Comparative Survival Efficacy. Contrasting three different “laws of biological conversion” by which investment in maintenance affects survival. Under the complementarity regime, returns on investments in any one aspect of survival may initially be slow but there is a steep “synergy window” within which incremental investments yield very large improvements in the probability of survival. On this view, once an organism invests enough to stabilize basic comatic integrity, the efficiency of all other repair and growth in health mechanisms may spike. This can be compared to either a simple linear returns on investment regime, where investments in health are simply added up to determine corresponding increases in survival, and a “substitutive” regime where investments in health quickly run into persistent and diminishing returns. This diagram suggests that, under the complementarity regime, organisms must invest a substantial and critical amount in survival to move beyond a low inflection point and unlock the regions where investments in health produce are mutually reinforcing. .
Figure 3.
Comparative Survival Efficacy. Contrasting three different “laws of biological conversion” by which investment in maintenance affects survival. Under the complementarity regime, returns on investments in any one aspect of survival may initially be slow but there is a steep “synergy window” within which incremental investments yield very large improvements in the probability of survival. On this view, once an organism invests enough to stabilize basic comatic integrity, the efficiency of all other repair and growth in health mechanisms may spike. This can be compared to either a simple linear returns on investment regime, where investments in health are simply added up to determine corresponding increases in survival, and a “substitutive” regime where investments in health quickly run into persistent and diminishing returns. This diagram suggests that, under the complementarity regime, organisms must invest a substantial and critical amount in survival to move beyond a low inflection point and unlock the regions where investments in health produce are mutually reinforcing. .
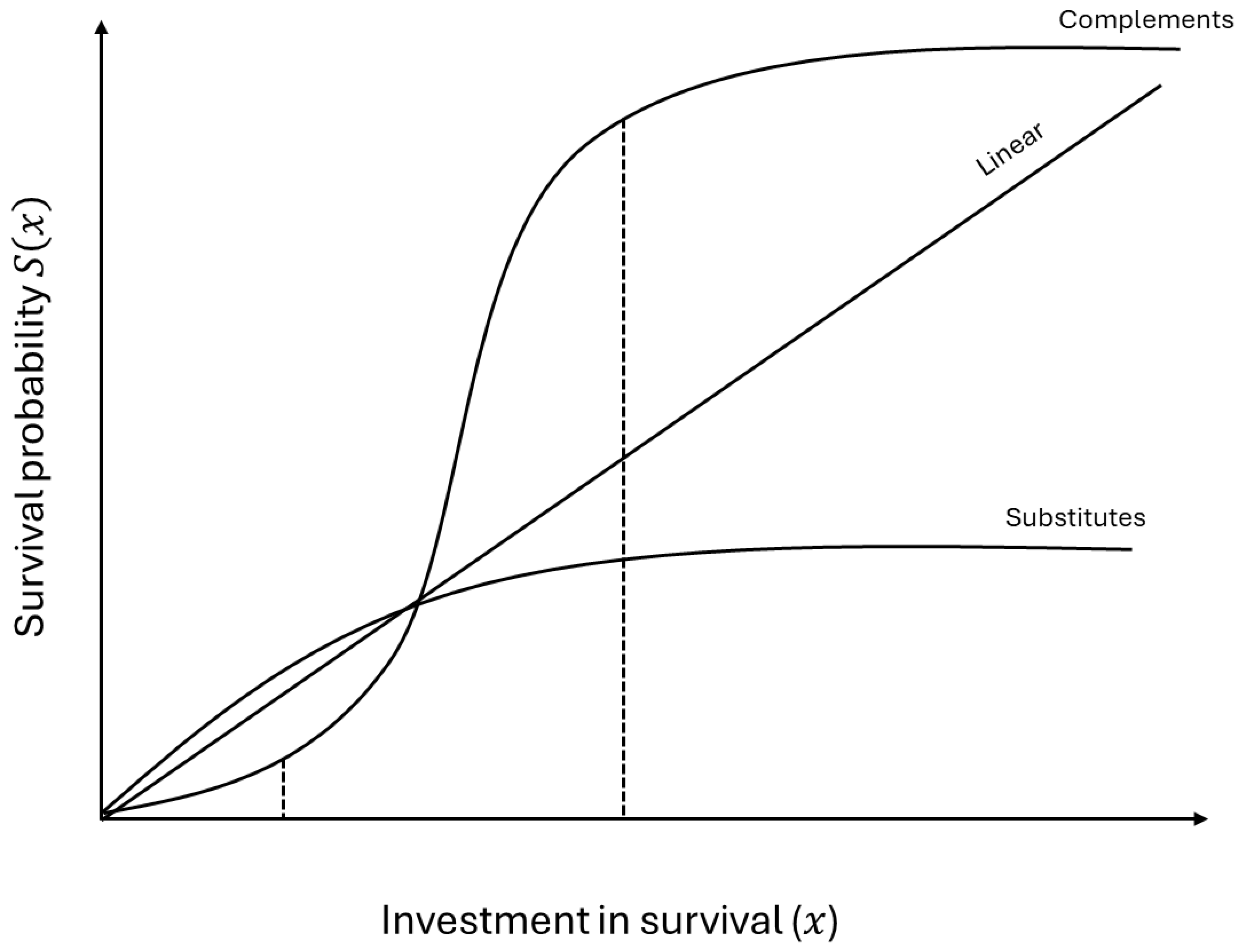
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



