Submitted:
25 February 2026
Posted:
26 February 2026
You are already at the latest version
Abstract
Sharp wave-ripples (SWRs) are transient hippocampal population events that coordinate neuronal ensemble activity and play a central role in memory consolidation and affective processing. Although SWRs exhibit marked functional specialization along the dorsoventral axis of the hippocampus, and many of the cellular mechanisms underlying SWRs are sex sensitive, systematic comparisons of SWR properties between females and males are lacking. Here, we examined sex- and region-dependent differences in SWRs and associated multiunit activity (MUA) in acute hippocampal slices from adult female and male rats. Spontaneous SWRs were recorded from the CA1 stratum pyramidale of dorsal and ventral hippocampus, and SWR amplitude, frequency, ripple oscillation properties, and SWR-locked neuronal firing were quantified. In the dorsal hippocampus, females exhibited a significantly higher SWR occurrence rate than males, whereas SWR amplitude was similar between sexes. In contrast, SWR-locked MUA was significantly higher in males, despite their lower SWR frequency. In the ventral hippocampus, females and males showed comparable SWR amplitude and frequency; however, males again displayed enhanced SWR-locked MUA, accompanied by reduced ripple power. Baseline MUA did not differ between sexes in either hippocampal region, indicating that sex-related effects were specific to the SWR state. These findings demonstrate that SWRs are regulated in a sex- and region-dependent manner, with dissociable effects on event recurrence, oscillatory structure, and neuronal recruitment. We propose that sex-sensitive mechanisms, including GABAergic interneuron function, steroid hormone signaling, and intrinsic and synaptic excitability, differentially tune SWR initiation and pyramidal cell recruitment along the dorsoventral axis. Our results reveal previously unrecognized dimensions of hippocampal network organization providing a mechanistic framework for understanding how sex-dependent circuit properties may shape hippocampal contributions to cognition and affective regulation and highlight the importance of incorporating sex as a fundamental biological variable in studies of hippocampal network dynamics.
Keywords:
sharp wave
; ripple
; multiunit activity
; network dynamics
; dorsoventral hippocampus
; sex
; gender
; septotemporal
; rat
1. Introduction
Sharp wave–ripples (SWRs) are transient, population events generated by the coordinated activity of hippocampal neuronal ensembles, and they consist of a large-amplitude sharp wave originating primarily in CA3 recurrent circuitry and a superimposed ripple oscillation (∼120–200 Hz) in CA1 [1]. Their excitatory output influences widespread cortical and subcortical targets and is thought to support key operations of episodic memory, systems consolidation, planning, and decision making, by supporting the reactivation of experience-dependent neuronal assemblies during sleep and awake immobility [1,2,3]. Disrupting SWRs impairs learning and adaptive behavior, underscoring their central role in hippocampal information processing. Mechanistically, generation of SWRs depends on the interaction of strong CA3 recurrent excitation, fast perisomatic inhibition by parvalbumin (PV)-containing interneurons, and additional synchronizing factors such as gap junctions [4,5]. Despite this rich literature, the overwhelming majority of SWR studies have used male rodents, and there is, to our knowledge, a complete absence of systematic comparisons of SWR properties between females and males in dorsal versus ventral hippocampus, either in vivo or in vitro.
The hippocampus is functionally and anatomically organized along its longitudinal (dorsoventral) axis. The dorsal (septal/posterior in human) segment of the hippocampus preferentially supports spatial and cognitive aspects of memory, whereas the ventral (temporal, anterior in human) hippocampus is more engaged in affective processing, stress regulation, and anxiety through its strong connectivity with the amygdala, hypothalamus, and medial prefrontal cortex [6,7,8,9,10]. Importantly, several studies indicate that ventral hippocampal manipulations affect anxiety-like behavior in a sex-dependent manner: ventral hippocampal inhibition or lesions modulate anxiety-like responses in male, but not necessarily female, mice, and androgen-dependent mechanisms shape excitability in ventral hippocampus-nucleus accumbens circuits [11,12].
Importantly, this functional gradient is reflected at the level of network physiology. Both in vivo [13,14,15,16] and in vitro [17,18,19,20,21] studies support dorsoventral specialization of SWRs. Dorsal SWRs are tightly linked to spatial memory replay and interactions with neocortical networks, whereas ventral SWRs show preferential coupling with limbic targets such as the amygdala and nucleus accumbens, consistent with a role in emotional and motivational processing [13,22]. Notably, recent work has demonstrated that ventral hippocampal SWRs can selectively reactivate stress-associated neuronal ensembles after aversive experience and causally influence stress susceptibility, highlighting ventral SWRs as key physiological substrates for affective regulation [22]. Together, these data establish dorsoventral differences in SWR properties function as a fundamental organizing principle of hippocampal network activity, indicating that SWR-generating circuits operate under distinct regimes along the hippocampal axis. Given that SWRs constitute the main offline population pattern through which hippocampal networks communicate with these downstream targets [1], it is highly plausible that SWR dynamics in dorsal and ventral hippocampus differ between sexes, yet this has not been tested. At the circuit level, SWR generation critically depends on precise inhibitory control, particularly from parvalbumin-positive (PV) and somatostatin-positive (SST) interneurons that synchronize pyramidal cell firing during ripple oscillations [5]. Variations in interneuron composition, intrinsic excitability, and receptor expression along the dorsoventral axis are therefore expected to profoundly shape SWR dynamics. Crucially, many of the cellular and synaptic mechanisms that govern SWRs are sex-sensitive.
Gonadal steroids, particularly estrogens, modulate hippocampal excitability, and inhibitory transmission. Estrogen receptors are expressed in hippocampal pyramidal cells and interneurons, and estradiol has been shown to regulate PV interneuron function, perineuronal net integrity, and high-frequency network oscillations such as gamma rhythms [23,24]. In mice, the adolescent trajectory of PV and GAD67 expression in hippocampal CA1/CA3 is steeper in females than males, with PV levels correlating positively with circulating estradiol; ovariectomy blunts this developmental increase, whereas estradiol replacement restores it [25,26]. Recent work demonstrates that gephyrin phosphorylation is crucial for the sexually dimorphic development and function of PV interneurons in hippocampus, with disruption of gephyrin signaling abolishing normal sex differences in PV cell density and leading to sex-specific cognitive impairments [27]. At the same time, estrogens exert rapid and long-term effects on hippocampal GABAergic inhibition and structural plasticity. Reviews and experimental work show that estradiol modulates inhibitory synapse density, GABAA receptor function, and the balance between excitation and inhibition in a sex- and region-dependent manner [28,29,30]. Structural plasticity studies further demonstrate estrous-cycle–linked fluctuations in dendritic spines and neurogenesis in hippocampus, with clear sex differences in the magnitude and direction of estrogen effects [31,32]. Because SWR initiation and ripple timing depend critically on interneuron-mediated inhibition and the fine-tuning of E/I balance, these sexually dimorphic GABAergic and estrogen-sensitive processes are a strong mechanistic reason to expect sex-related differences in SWRs along the dorsoventral axis.
Despite this strong mechanistic rationale, direct comparative evidence on SWRs in males and females is remarkably sparse. The vast majority of SWR studies have been conducted in male animals, and sex is often unreported or unexamined as a biological variable. To date, no study has systematically compared SWR properties between males and females, and none have done so with explicit dorsoventral resolution. This absence of data is particularly striking in light of the robust dorsoventral differences in SWR dynamics and the well-established sex differences in hippocampal circuitry and neuromodulatory systems. The lack of sex-comparative SWR data is especially consequential given pronounced sex differences in behavior and psychopathology linked to ventral hippocampal function. Anxiety disorders, post-traumatic stress disorder, and major depression are more prevalent in women than in men, with differences in symptom profiles, stress sensitivity, and disease trajectory [33,34,35]. Experimental studies further demonstrate sex-dependent differences in fear learning, fear generalization, and stress-induced hippocampal plasticity—processes that critically involve ventral hippocampal-amygdala circuits [36,37]. Recent rodent studies have also examined sex differences in limbic oscillatory activity in anxiety paradigms: for example, sex-dependent patterns of amygdalo–hippocampal oscillations and neuronal recruitment are seen in elevated plus maze and following infralimbic deep brain stimulation, suggesting that limbic network synchronization differs between males and females during anxiogenic conditions [38]. Human MEG/EEG studies report gender differences in hippocampal–parietal theta and gamma modulation during navigation and memory tasks, consistent with the idea that large scale hippocampal network dynamics are sexually differentiated [39,40]. Because ventral SWRs have been causally implicated in stress-related ensemble reactivation and affective regulation, sex differences in SWR dynamics represent a reasonable, yet unexplored, neurophysiological substrate for these behavioral biases.
In summary, converging lines of evidence demonstrate that SWRs differ fundamentally between dorsal and ventral hippocampus, the cellular and synaptic mechanisms underlying SWRs are strongly modulated by sex-dependent factors, and the behavioral domains linked to ventral hippocampal function exhibit pronounced sex differences in both physiology and pathology. Yet, comparative analyses of SWRs across sex and hippocampal axis are essentially absent. Addressing this gap is necessary to achieve a comprehensive understanding of hippocampal network function and may provide critical insights into the circuit-level mechanisms underlying sex-biased vulnerability to stress-related disorders. A systematic, sex-informed characterization of SWRs in dorsal and ventral hippocampus would therefore fill a critical gap in our understanding of hippocampal network physiology. It would also provide a much-needed mechanistic framework connecting sex differences in inhibitory circuitry, neuromodulation, synaptic plasticity, and affective behavior to the population events that regulate hippocampal output. This, in turn, could help explain sex-biased vulnerability and resilience to psychiatric disorders and inform the development of sex-specific biomarkers and interventions grounded in hippocampal network dynamics.
2. Materials and Methods
2.1. Animals and Hippocampal Slice Preparation
Transverse hippocampal slices (550 μm thick) were obtained from 3–4-month-old female and male Wistar rats housed in the Laboratory of Experimental Animals, Department of Medicine, University of Patras. Animals were maintained under stable temperature (20–22 °C) and a 12:12 h light-dark cycle with ad libitum access to food and water. All procedures complied with EU Directive 2010/63/EU and were approved by institutional and regional authorities (reg. no. 187531/626, 26/06/2018). Following deep anaesthesia with diethyl ether and decapitation, brains were quickly removed and placed in ice-cold oxygenated artificial cerebrospinal fluid (ACSF). Slices were prepared from the dorsal and ventral hippocampus (0.5–3.5 mm from each end) and transferred to an interface chamber perfused with ACSF (containing, in mM: 124 NaCl, 4 KCl, 2 CaCl2, 2 MgSO4, 26 NaHCO3, 1.25 NaH2PO4 and 10 glucose, equilibrated with 95% O2 and 5% CO2 gas mixture at pH=7.4) at 30 ± 0.5 °C. Slices were continuously humidified with a mixed gas consisting of 95% O2 and 5% CO2. After at least 90 minutes of recovery, electrophysiological recordings were initiated.
2.2. Electrophysiology and Data Acquisition
Spontaneous field potentials were recorded from the stratum pyramidale of the CA1 hippocampal field, using carbon fiber electrodes 7 μm-thick (Kation Scientific, Minneapolis, USA). Signal was acquired and amplified X500 and then filtered at 0.5 Hz–2 kHz using Neurolog systems (Digitimer Ltd, UK), consisting of AC preamplifier (NL 104A with NL 100AK headstage), AC/DC amplifier (NL 106) and band pass filter (NL 125/6). Analog signal was digitized at 10 kHz using a CED 1401-plus interface and the Spike software (Cambridge Electronic Design, Cambridge, UK), then, stored on a computer disk for off-line analysis using the same software. Activity consisted of sharp waves ridden by multiunit activity (MUA) (Figure 1). Events of SWRs were quantified by 1) their amplitude measured as the voltage difference between the positive peak and the baseline. 2) the duration of single SWR events measured as the time interval between the two points of the positive phase that intersect the baseline. 3) the inter-event interval (IEI) measured as the time between successive individual SWRs. Measures of SWRs were performed after original records were down sampled (at 1 kHz) and low-pass filtered at 35 Hz. Then, individual events were detected after setting a threshold at a level where all putative events were identified as verified by visual inspection as previously described [17]. Multiunit activity (MUA) was revealed in band-pass filtered records (at 400 Hz-1.5 kHz) and was detected by setting a threshold level at a value that all putative events (i.e., negative spikes) were identified as verified by visual inspection, as previously described [17]. MUA occurred between events of SWRs SWRs is called MUA-Base, and during SWRs SWRs called MUA-SWR. We quantified both MUA-Base and MUA-SWR by its frequency of occurrence (Hz). MUA-Base was measured by the frequency of MUA at steady state between consecutive events of SWRs. We measured MUA-SWR by the maximum frequency of MUA in peri-event histograms between SWRs and MUA, where we used the positive peaks of low-pass filtered SWRs SWRs as reference events (Figure 1D).
2.3. Statistical Analysis
Comparisons between two groups were performed using the independent t-test. The assumption of equal variances and normal distribution of data was evaluated using Levene’s test and the Shapiro–Wilk test, respectively. Each hippocampal slice was considered an independent experimental unit. Data are expressed as mean ± S.E.M. Diamond plots depict the median and interquartile range (25th to 75th percentiles; diamond-shaped box), mean (thick line), 5th and 95th percentiles (whiskers), and individual data points (superimposed). The number of slices is reported throughout the text. Statistical analyses were performed based on the number of slices. All analyses were conducted using IBM SPSS Statistics version 27.
3. Results
3.1. Higher Frequency of SWRs in Female than Male Dorsal Hippocampus
We recorded spontaneously occurring SWRs and MUA from the CA1 stratum pyramidale of the dorsal and ventral female and male hippocampus. From these recordings, we quantified three types of activity: sharp waves, ripple oscillation, and multiunit activity occurring either during events of SWRs (SWR-MUA) or between events of SWRs (baseline MUA, Figure 1). Considering that SWRs display different organizational properties between the dorsal and the ventral hippocampus [17,18,20,21], we compared them between the two sexes separately for each segment of the hippocampus. In the dorsal hippocampus, we found that SWRs display similar amplitude in females and males (Figure 2A,B,C). However, SWRs occurred with significantly higher frequency in female than male dorsal hippocampus (Figure 1A,D,E), suggesting a sex-related difference in the mechanisms that control the recurrency of SRW activity.
3.2. Higher SWR-Locked Multiunit Firing in Male than Female Dorsal Hippocampus
To determine whether sex-related differences extend beyond SWR occurrence to neuronal recruitment during SWRs, we compared MUA recorded during SWRs (MUA-SWR) and during inter-event periods (baseline MUA; MUA-Base) in the dorsal hippocampus of females and males. Peri-event time histograms revealed robust increases in MUA time-locked to the sharp wave peak in both sexes (Figure 3A). Quantitative analysis showed that baseline MUA frequency did not differ significantly between females and males (Figure 3C, left panel). In contrast, MUA-SWR frequency was significantly higher in males than in females (Figure 3C, right panel; Mann–Whitney test, p < 0.05), indicating a sex-related difference in the degree of neuronal firing recruited during SWRs in the dorsal hippocampus. Analysis of ripple oscillation parameters showed no significant sex differences in ripple peak frequency or ripple power in the dorsal hippocampus (Figure 3D). Thus, in dorsal hippocampus, males exhibit enhanced SWR-locked neuronal firing without accompanying changes in ripple frequency or power.
3.3. Similar Amplitude and Frequency of SWRs in Female and Male Ventral Hippocampus
We next examined whether sex-related differences in SWR properties were also present in the ventral hippocampus. Spontaneously occurring SWRs were recorded from ventral hippocampal slices of female and male rats (Figure 4A). Analysis of sharp wave amplitude revealed no significant differences between females and males, as shown by both time-course plots and cumulative distributions (Figure 4B,C). Similarly, comparison of inter-event interval (IEI) demonstrated that SWR occurrence rate did not differ between sexes in the ventral hippocampus (Figure 4D,E). Thus, unlike the dorsal hippocampus, the ventral hippocampus exhibited no sex-related differences in either SWR amplitude or frequency, indicating comparable generation and recurrence of SWRs in females and males in this hippocampal segment.
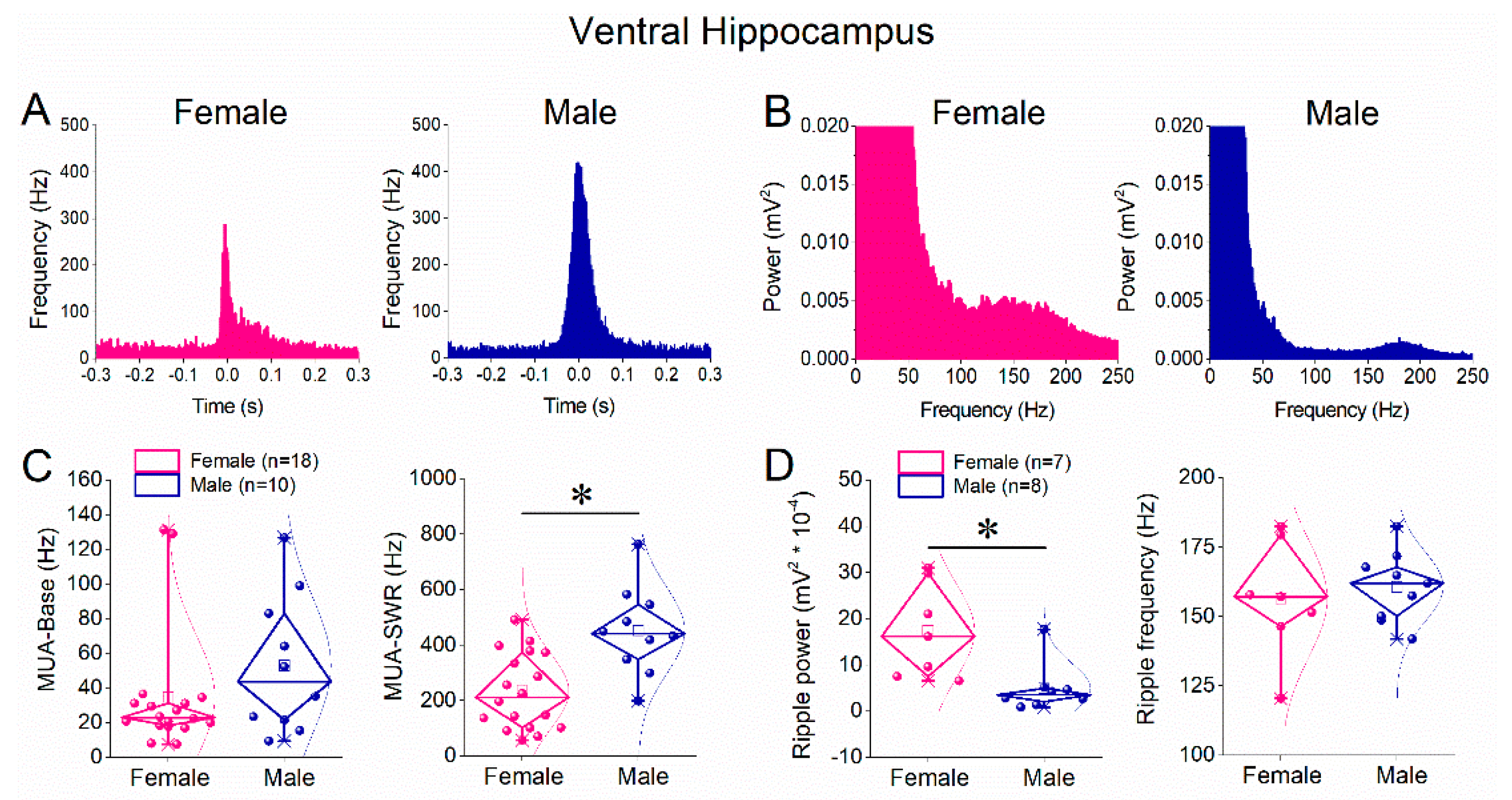
3.4. Higher SWR-Locked Multiunit Firing but Reduced Ripple Power in Male than Female Ventral Hippocampus
Although SWR amplitude and frequency were similar between sexes in the ventral hippocampus, we next assessed whether neuronal recruitment and ripple properties differed. Peri-event histograms showed prominent MUA increases time-locked to SWRs in both females and males (Figure 5A). Quantitative analysis revealed that baseline MUA frequency did not differ significantly between sexes (Figure 5C, left panel). In contrast, MUA-SWR frequency was significantly higher in males than in females (Figure 5C, right panel; Mann–Whitney test, p < 0.05), indicating enhanced SWR-associated neuronal firing in the male ventral hippocampus. Analysis of ripple oscillations revealed a significant reduction in ripple power in males compared with females (Figure 5D, left panel), whereas ripple peak frequency did not differ between sexes (Figure 5D, right panel). Thus, in the ventral hippocampus, males exhibit increased SWR-locked neuronal firing despite reduced ripple power.
4. Discussion
In the present study, we provide the first systematic comparison of SWRs and associated MUA between females and males along the dorsoventral axis of the hippocampus, and reveals pronounced, sex dependent differences that are themselves region specific along the dorsoventral axis of the adult rat hippocampus. Specifically, the principal findings are the following. In dorsal CA1, females showed a significantly higher frequency of SWRs than males, despite comparable sharp wave amplitude, whereas in ventral CA1 SWR amplitude and occurrence rate did not differ between sexes. In contrast, SWR locked MUA was higher in males than females in both dorsal and ventral hippocampus, while baseline MUA between SWRs showed smaller or no sex differences. Moreover, in ventral CA1, males combined stronger SWR locked MUA with reduced ripple power relative to females. Together, these results reveal a novel pattern: female dorsal hippocampus supports more frequent SWRs generated from apparently smaller neuronal populations, whereas male hippocampus, especially the ventral segment, generates fewer or similarly frequent SWRs that recruit more neurons with somewhat degraded ripple structure. These findings add a sex dimension to the well established dorsoventral specialization of SWR dynamics.
4.1. Mechanisms Underlying Sex-Dependent SWR Differences
The higher SWR frequency observed in the dorsal hippocampus of females suggests sex-related differences in the mechanisms underlying SWR initiation and recurrence. SWR generation depends critically on recurrent excitation in CA3 and the excitability of CA3–CA1 circuitry [1]. Sex differences in glutamatergic transmission and intrinsic neuronal excitability are well documented in hippocampus and are strongly modulated by gonadal steroids, particularly estrogens [28,41]. These effects could lower the threshold for SWR initiation or shorten the recovery period between successive events, leading to a higher event rate in females. The absence of sex differences in SWR amplitude indicates that once an SWR is generated, the magnitude of population depolarization is comparable between sexes, suggesting that sex-dependent mechanisms primarily influence event recurrence rather than event strength.
A central finding of this study is that males exhibit higher SWR-locked neuronal firing than females in both dorsal and ventral hippocampus, despite equal or lower SWR rates and, in ventral hippocampus, reduced ripple power. SWR-locked firing reflects the temporal precision and strength of neuronal recruitment during the ripple cycle and depends critically on fast perisomatic inhibition mediated by parvalbumin-positive (PV) interneurons [4,5]. Sex differences in PV interneuron development, density, and function have been reported in hippocampus and are shaped by gonadal steroids during adolescence [25,26]. More recently, gephyrin phosphorylation has been identified as a key organizer of sexually dimorphic PV interneuron development and function in CA1, with disruption of gephyrin signaling abolishing normal sex differences in PV cell density and producing sex- and task-specific cognitive deficits [31,32].
Within this framework, higher MUA-SWR in males may reflect tighter excitation–inhibition coupling during SWRs, promoting stronger spike synchronization even when ripple power is not increased. The lack of robust sex differences in baseline MUA supports the conclusion that these effects are specific to the SWR state and do not reflect global differences in excitability. In the ventral hippocampus, males exhibited higher SWR-locked firing together with reduced ripple power. Ventral hippocampal SWRs differ fundamentally from dorsal SWRs in their organization, intrinsic conductances, and neuromodulatory sensitivity [17,18,20,21]. Reduced ripple power in males may reflect altered synchronization of inhibitory networks or differences in dendritic current contributions, while preserved or enhanced MUA-SWR suggests efficient recruitment of spiking within a narrower temporal window. This dissociation highlights that ripple power and neuronal recruitment can be independently regulated and emphasizes the importance of analyzing multiple SWR features in parallel.
4.2. Functional Implications
SWRs constitute the primary population events through which hippocampal networks communicate with downstream cortical and subcortical targets [1]. Sex- and region-dependent differences in SWR rate, ripple structure, and SWR-locked firing are therefore expected to influence how information is transmitted to neocortical and limbic circuits. In the dorsal hippocampus, higher SWR frequency in females may support more frequent replay of spatial or contextual information, whereas stronger SWR-locked firing in males may enhance the impact or precision of individual replay events. These network-level differences may relate to reported sex biases in spatial strategies, pattern separation, and contextual memory, which have been linked to sex-specific plasticity and neurogenesis along the dorsoventral axis [42].
In the ventral hippocampus, SWRs have been causally linked to stress-related ensemble reactivation and affective regulation [13,22]. Sex differences in SWR-associated firing and ripple organization may therefore contribute to sex-biased vulnerability to anxiety and mood disorders. Epidemiological studies consistently report higher prevalence of anxiety and depression in women [33,34], and experimental work demonstrates sex-dependent hippocampal–amygdala network dynamics during stress and anxiety [8,38]. Although the present study was conducted in vitro, the observed sex-specific SWR signatures provide a plausible circuit-level substrate for these behavioral differences.
4.3. Limitations and Future Directions
This study was conducted in hippocampal slices, allowing precise dissection of intrinsic circuit mechanisms but excluding long-range inputs and acute hormonal fluctuations. Future in vivo approaches are required to determine whether the sex-dependent SWR differences observed here generalize to freely behaving animals across sleep-wake states. Also, future work should explicitly examine hormonal modulation of SWRs across the cycle to distinguish organizational from activational effects of sex steroids [28,29,30]. Mechanistically, future studies should combine SWR recordings with targeted manipulation of PV and SST interneurons, intrinsic conductances, and NMDA receptor signaling, which exhibits sex-dependent features in hippocampus [28,29,30,43]. Extending sex- and region-resolved SWR analyses to disease models characterized by sex-biased prevalence, such as anxiety disorders, depression, schizophrenia, fragile X syndrome, and Alzheimer’s disease, may also reveal whether pathological processes selectively disrupt SWR dynamics in one sex or hippocampal segment [20,44].
5. Conclusions
In conclusion, our findings demonstrate that SWRs are regulated in a sex- and region-dependent manner along the hippocampal dorsoventral axis. Females exhibit higher SWR recurrence in dorsal hippocampus, whereas males show enhanced SWR-locked neuronal firing in both dorsal and ventral hippocampus, accompanied by reduced ripple power in ventral circuits. These results reveal previously unrecognized dimensions of hippocampal network organization and underscore the necessity of incorporating sex as a biological variable in studies of hippocampal population dynamics. By linking sex-dependent circuit mechanisms to SWR physiology, this work provides a foundation for understanding sex-biased differences in cognition and affective behavior.
Author Contributions
A.M., G.T., and M.T., performed the experiments and data analysis. C.P. supervised the project and performed data analysis. A.M., and C.P. prepared and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The research project was supported by the Empeirikeion Foundation. A.M. is a recipient of a postgraduate fellowship from the Hellenic Foundation for Research and Innovation (HFRI). G.T. was financially supported by the “Polembros Shipping Limited”, as a recipient of Ph.D. fellowships.
Institutional Review Board Statement
The animal study protocol was approved by the Research Ethics Committee of the University of Patras and the Directorate of Veterinary Services of the Achaia Prefecture of Western Greece Region (reg. number: 5661/37, 18 January 2021). The treatment of animals and all experimental procedures used in this study were conducted in accordance with the European Communities Council Directive Guidelines for the care and use of Laboratory animals (2010/63/EU—European Commission).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data associated with this study are available from the corresponding author upon reasonable request.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments). Where GenAI has been used for purposes such as generating text, data, or graphics, or for study design, data collection, analysis, or interpretation of data, please add “During the preparation of this manuscript/study, the author(s) used [tool name, version information] for the purposes of [description of use]. The authors have reviewed and edited the output and take full responsibility for the content of this publication.”.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| SWR | sharp wave-ripple |
| MUA | multiunit activity |
| MUA-SWR | SWR-associated multiunit activity |
| MUA-Base | Baseline multiunit activity (outside SWRs) |
| IEI | inter-event interval |
| LFP | local field potential |
| PV | parvalbumin (interneurons) |
| SST | somatostatin (interneurons) |
References
- Buzsaki, G. Hippocampal sharp wave-ripple: A cognitive biomarker for episodic memory and planning. Hippocampus 2015, 25, 1073–1188. [Google Scholar] [CrossRef]
- Joo, H.R.; Frank, L.M. The hippocampal sharp wave-ripple in memory retrieval for immediate use and consolidation. Nature reviews. Neuroscience 2018, 19, 744–757. [Google Scholar] [CrossRef]
- Jadhav, S.P.; Kemere, C.; German, P.W.; Frank, L.M. Awake hippocampal sharp-wave ripples support spatial memory. Science 2012, 336, 1454–1458. [Google Scholar] [CrossRef]
- Csicsvari, J.; Hirase, H.; Czurko, A.; Mamiya, A.; Buzsaki, G. Oscillatory coupling of hippocampal pyramidal cells and interneurons in the behaving Rat. The Journal of neuroscience: the official journal of the Society for Neuroscience 1999, 19, 274–287. [Google Scholar] [CrossRef]
- Schlingloff, D.; Kali, S.; Freund, T.F.; Hajos, N.; Gulyas, A.I. Mechanisms of sharp wave initiation and ripple generation. The Journal of neuroscience: the official journal of the Society for Neuroscience 2014, 34, 11385–11398. [Google Scholar] [CrossRef]
- Fanselow, M.S.; Dong, H.W. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef]
- Strange, B.A.; Witter, M.P.; Lein, E.S.; Moser, E.I. Functional organization of the hippocampal longitudinal axis. Nat Rev Neurosci 2014, 15, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Felix-Ortiz, A.C.; Tye, K.M. Amygdala inputs to the ventral hippocampus bidirectionally modulate social behavior. The Journal of neuroscience: the official journal of the Society for Neuroscience 2014, 34, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Coreano, N.; Bolkan, S.S.; Pierce, G.M.; Blackman, D.R.; Hardin, W.D.; Garcia-Garcia, A.L.; Spellman, T.J.; Gordon, J.A. Direct Ventral Hippocampal-Prefrontal Input Is Required for Anxiety-Related Neural Activity and Behavior. Neuron 2016, 89, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.M.; Sprengel, R.; Sanderson, D.J.; McHugh, S.B.; Rawlins, J.N.; Monyer, H.; Seeburg, P.H. Hippocampal synaptic plasticity, spatial memory and anxiety. Nat Rev Neurosci 2014, 15, 181–192. [Google Scholar] [CrossRef]
- Maggio, N.; Segal, M. Corticosteroid regulation of synaptic plasticity in the hippocampus. TheScientificWorldJournal 2010, 10, 462–469. [Google Scholar] [CrossRef]
- Williams, E.S.; Manning, C.E.; Eagle, A.L.; Swift-Gallant, A.; Duque-Wilckens, N.; Chinnusamy, S.; Moeser, A.; Jordan, C.; Leinninger, G.; Robison, A.J. Androgen-Dependent Excitability of Mouse Ventral Hippocampal Afferents to Nucleus Accumbens Underlies Sex-Specific Susceptibility to Stress. Biological psychiatry 2020, 87, 492–501. [Google Scholar] [CrossRef]
- Sosa, M.; Joo, H.R.; Frank, L.M. Dorsal and Ventral Hippocampal Sharp-Wave Ripples Activate Distinct Nucleus Accumbens Networks. Neuron 2020, 105, 725–741.e728. [Google Scholar] [CrossRef] [PubMed]
- Nitzan, N.; Swanson, R.; Schmitz, D.; Buzsáki, G. Brain-wide interactions during hippocampal sharp wave ripples. Proceedings of the National Academy of Sciences of the United States of America 2022, 119, e2200931119. [Google Scholar] [CrossRef]
- De Filippo, R.; Schmitz, D. Differential ripple propagation along the hippocampal longitudinal axis. eLife 2023, 12. [Google Scholar] [CrossRef]
- Patel, J.; Schomburg, E.W.; Berenyi, A.; Fujisawa, S.; Buzsaki, G. Local generation and propagation of ripples along the septotemporal axis of the hippocampus. The Journal of neuroscience: the official journal of the Society for Neuroscience 2013, 33, 17029–17041. [Google Scholar] [CrossRef] [PubMed]
- Kouvaros, S.; Papatheodoropoulos, C. Prominent differences in sharp waves, ripples and complex spike bursts between the dorsal and the ventral rat hippocampus. Neuroscience 2017, 352, 131–143. [Google Scholar] [CrossRef]
- Trompoukis, G.; Rigas, P.; Leontiadis, L.J.; Papatheodoropoulos, C. I(h), GIRK, and KCNQ/Kv7 channels differently modulate sharp wave - ripples in the dorsal and ventral hippocampus. Molecular and cellular neurosciences 2020, 107, 103531. [Google Scholar] [CrossRef] [PubMed]
- Trompoukis, G.; Leontiadis, L.J.; Rigas, P.; Papatheodoropoulos, C. Scaling of Network Excitability and Inhibition may Contribute to the Septotemporal Differentiation of Sharp Waves-Ripples in Rat Hippocampus In Vitro. Neuroscience 2021, 458, 11–30. [Google Scholar] [CrossRef]
- Leontiadis, L.J.; Trompoukis, G.; Tsotsokou, G.; Miliou, A.; Felemegkas, P.; Papatheodoropoulos, C. Rescue of sharp wave-ripples and prevention of network hyperexcitability in the ventral but not the dorsal hippocampus of a rat model of fragile X syndrome. Frontiers in cellular neuroscience 2023, 17, 1296235. [Google Scholar] [CrossRef]
- Kandilakis, C.L.; Papatheodoropoulos, C. Serotonin Modulation of Dorsoventral Hippocampus in Physiology and Schizophrenia. Int. J. Mol. Sci. 2025, 26, 7253. [Google Scholar] [CrossRef] [PubMed]
- Kuga, N.; Nakayama, R.; Morikawa, S.; Yagishita, H.; Konno, D.; Shiozaki, H.; Honjoya, N.; Ikegaya, Y.; Sasaki, T. Hippocampal sharp wave ripples underlie stress susceptibility in male mice. Nature communications 2023, 14, 2105. [Google Scholar] [CrossRef]
- Blurton-Jones, M.; Tuszynski, M.H. Estradiol-induced modulation of estrogen receptor-beta and GABA within the adult neocortex: a potential transsynaptic mechanism for estrogen modulation of BDNF. The Journal of comparative neurology 2006, 499, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Taxier, L.R.; Gross, K.S.; Frick, K.M. Oestradiol as a neuromodulator of learning and memory. Nat Rev Neurosci 2020, 21, 535–550. [Google Scholar] [CrossRef]
- Willing, J.; Juraska, J.M. The timing of neuronal loss across adolescence in the medial prefrontal cortex of male and female rats. Neuroscience 2015, 301, 268–275. [Google Scholar] [CrossRef]
- Wu, Y.C.; Du, X.; van den Buuse, M.; Hill, R.A. Sex differences in the adolescent developmental trajectory of parvalbumin interneurons in the hippocampus: a role for estradiol. Psychoneuroendocrinology 2014, 45, 167–178. [Google Scholar] [CrossRef]
- Campbell, B.F.N.; Cruz-Ochoa, N.; Otomo, K.; Lukacsovich, D.; Espinosa, P.; Abegg, A.; Luo, W.; Bellone, C.; Földy, C.; Tyagarajan, S.K. Gephyrin phosphorylation facilitates sexually dimorphic development and function of parvalbumin interneurons in the mouse hippocampus. Molecular psychiatry 2024, 29, 2510–2526. [Google Scholar] [CrossRef]
- Woolley, C.S. Acute effects of estrogen on neuronal physiology. Annual review of pharmacology and toxicology 2007, 47, 657–680. [Google Scholar] [CrossRef] [PubMed]
- Li, F.R.; Wang, S.; Suarez, C.F.; Lévesque, M.; Avoli, M. Interneurons, GABA(A) signaling and their presumptive role in catamenial epilepsy. Neuroscience and biobehavioral reviews 2025, 176, 106291. [Google Scholar] [CrossRef] [PubMed]
- Choleris, E.; Galea, L.A.M.; Sohrabji, F.; Frick, K.M. Sex differences in the brain: Implications for behavioral and biomedical research. Neuroscience and biobehavioral reviews 2018, 85, 126–145. [Google Scholar] [CrossRef]
- Galea, L.A.; Wainwright, S.R.; Roes, M.M.; Duarte-Guterman, P.; Chow, C.; Hamson, D.K. Sex, hormones and neurogenesis in the hippocampus: hormonal modulation of neurogenesis and potential functional implications. Journal of neuroendocrinology 2013, 25, 1039–1061. [Google Scholar] [CrossRef]
- Loi, M.; Koricka, S.; Lucassen, P.J.; Joëls, M. Age- and sex-dependent effects of early life stress on hippocampal neurogenesis. Frontiers in endocrinology 2014, 5, 13. [Google Scholar] [CrossRef]
- Kendler, K.S.; Kessler, R.C.; Walters, E.E.; MacLean, C.; Neale, M.C.; Heath, A.C.; Eaves, L.J. Stressful life events, genetic liability, and onset of an episode of major depression in women. The American journal of psychiatry 1995, 152, 833–842. [Google Scholar] [CrossRef]
- Bangasser, D.A.; Cuarenta, A. Sex differences in anxiety and depression: circuits and mechanisms. Nat Rev Neurosci 2021, 22, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Altemus, M.; Sarvaiya, N.; Neill Epperson, C. Sex differences in anxiety and depression clinical perspectives. Frontiers in neuroendocrinology 2014, 35, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Clark, U.S.; Miller, E.R.; Hegde, R.R. Experiences of Discrimination Are Associated With Greater Resting Amygdala Activity and Functional Connectivity. Biological psychiatry. Cognitive neuroscience and neuroimaging 2018, 3, 367–378. [Google Scholar] [CrossRef]
- Trott, J.M.; Krasne, F.B.; Fanselow, M.S. Sex differences in contextual fear learning and generalization: a behavioral and computational analysis of hippocampal functioning. Learning & memory (Cold Spring Harbor, N.Y 2022, 29, 283–296. [Google Scholar] [CrossRef]
- Vila-Merkle, H.; González-Martínez, A.; Campos-Jiménez, R.; Martínez-Ricós, J.; Teruel-Martí, V.; Lloret, A.; Blasco-Serra, A.; Cervera-Ferri, A. Sex differences in amygdalohippocampal oscillations and neuronal activation in a rodent anxiety model and in response to infralimbic deep brain stimulation. Frontiers in behavioral neuroscience 2023, 17, 1122163. [Google Scholar] [CrossRef] [PubMed]
- Gron, G.; Wunderlich, A.P.; Spitzer, M.; Tomczak, R.; Riepe, M.W. Brain activation during human navigation: gender-different neural networks as substrate of performance. Nature neuroscience 2000, 3, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Cimadevilla, J.M.; Piccardi, L. Spatial skills. Handbook of clinical neurology 2020, 175, 65–79. [Google Scholar] [CrossRef]
- Scharfman, H.E.; MacLusky, N.J. Sex differences in the neurobiology of epilepsy: a preclinical perspective. Neurobiology of disease 2014, 72 Pt B, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Yagi, S.; Galea, L.A.M. Sex differences in hippocampal cognition and neurogenesis. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology 2019, 44, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, T.; Dos Santos Fabris, D.; de Oliveira, C.L.; Guimarães, F.S.; Gomes, F.V. Prefrontal and Hippocampal Parvalbumin Interneurons in Animal Models for Schizophrenia: A Systematic Review and Meta-analysis. Schizophrenia bulletin 2024, 50, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Papatheodoropoulos, C.J.B. Compensatory Regulation of Excitation/Inhibition Balance in the Ventral Hippocampus: Insights from Fragile X Syndrome. Biology 2025, 14, 363. [Google Scholar] [CrossRef]
Figure 1.
Representative sharp wave–ripples (SWRs) recorded from the stratum pyramidale of the CA1 hippocampal region. A. Raw local field potential (LFP) recording showing multiple SWR events (upper trace). The middle trace shows the same signal band-pass filtered in the ripple range (90–200 Hz), revealing the ripple oscillations. The lower trace shows the signal band-pass filtered (0.4 – 1.5 kHz), revealing multiunit activity (MUA). B. Example of a single SWR event. The raw signal was low-pass filtered (<30 Hz) to isolate the sharp wave component (upper trace), band-pass filtered (100–200 Hz) to reveal the ripple oscillation (middle trace), and high-pass filtered (0.4 – 1.5 kHz) to extract the associated MUA (bottom trace). C. Power spectrum (Fourier transform) of the LFP signal showing the characteristic ripple-band peak around ~150 Hz. An example of a ripple oscillation is shown in the inset. D. Peri-event time histogram of MUA aligned to the sharp wave peak. Horizontal lines indicate the time windows used to quantify MUA during SWRs (MUA-SWR) and baseline MUA occurring outside SWRs (MUA-Base).
Figure 1.
Representative sharp wave–ripples (SWRs) recorded from the stratum pyramidale of the CA1 hippocampal region. A. Raw local field potential (LFP) recording showing multiple SWR events (upper trace). The middle trace shows the same signal band-pass filtered in the ripple range (90–200 Hz), revealing the ripple oscillations. The lower trace shows the signal band-pass filtered (0.4 – 1.5 kHz), revealing multiunit activity (MUA). B. Example of a single SWR event. The raw signal was low-pass filtered (<30 Hz) to isolate the sharp wave component (upper trace), band-pass filtered (100–200 Hz) to reveal the ripple oscillation (middle trace), and high-pass filtered (0.4 – 1.5 kHz) to extract the associated MUA (bottom trace). C. Power spectrum (Fourier transform) of the LFP signal showing the characteristic ripple-band peak around ~150 Hz. An example of a ripple oscillation is shown in the inset. D. Peri-event time histogram of MUA aligned to the sharp wave peak. Horizontal lines indicate the time windows used to quantify MUA during SWRs (MUA-SWR) and baseline MUA occurring outside SWRs (MUA-Base).
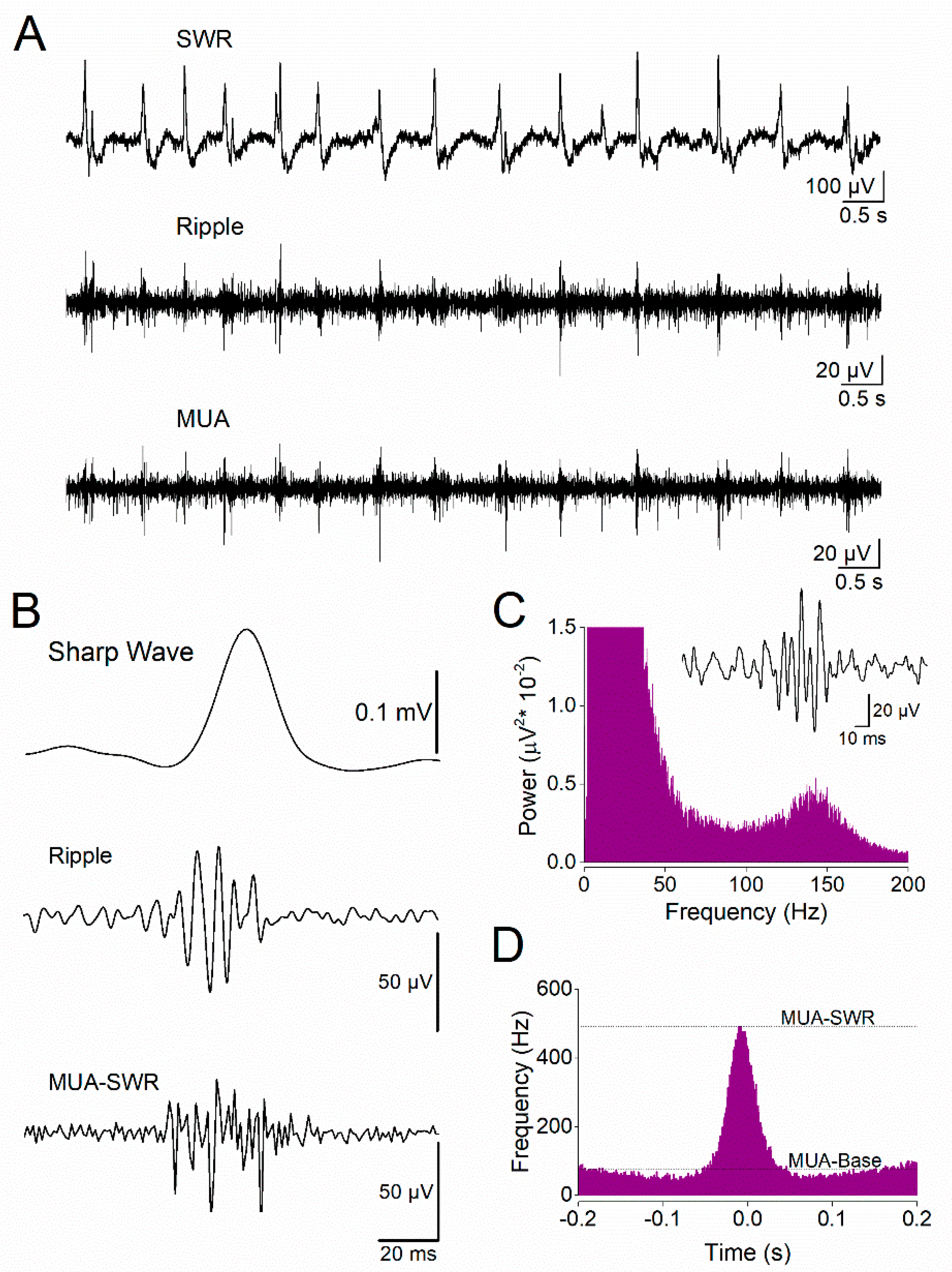
Figure 2.
Comparison of sharp wave–ripples (SWRs) between female and male dorsal hippocampus. A. Representative local field potential recordings from dorsal hippocampal slices obtained from a female (left) and a male (right) rat. B. Five-minute time course showing the amplitude of SWRs recorded from the dorsal hippocampus of female (pink) and male (blue) rats, demonstrating comparable SWR amplitudes between sexes. C. Diamond plots (left) and distribution histograms (right) of SWR amplitudes recorded from the dorsal hippocampus of female and male rats. D. Five-minute time course of inter-event interval (IEI), showing that SWRs occur at a higher rate (shorter IEI) in the dorsal hippocampus of female rats compared with males. E. Diamond plots (left) and distribution histograms (right) of IEI values from the dorsal hippocampus of female and male rats. The asterisk indicates a statistically significant sex-related difference in IEI (Mann–Whitney test, p < 0.05). Data are shown as mean ± SEM in time-course plots; individual data points are overlaid in diamond plots.
Figure 2.
Comparison of sharp wave–ripples (SWRs) between female and male dorsal hippocampus. A. Representative local field potential recordings from dorsal hippocampal slices obtained from a female (left) and a male (right) rat. B. Five-minute time course showing the amplitude of SWRs recorded from the dorsal hippocampus of female (pink) and male (blue) rats, demonstrating comparable SWR amplitudes between sexes. C. Diamond plots (left) and distribution histograms (right) of SWR amplitudes recorded from the dorsal hippocampus of female and male rats. D. Five-minute time course of inter-event interval (IEI), showing that SWRs occur at a higher rate (shorter IEI) in the dorsal hippocampus of female rats compared with males. E. Diamond plots (left) and distribution histograms (right) of IEI values from the dorsal hippocampus of female and male rats. The asterisk indicates a statistically significant sex-related difference in IEI (Mann–Whitney test, p < 0.05). Data are shown as mean ± SEM in time-course plots; individual data points are overlaid in diamond plots.
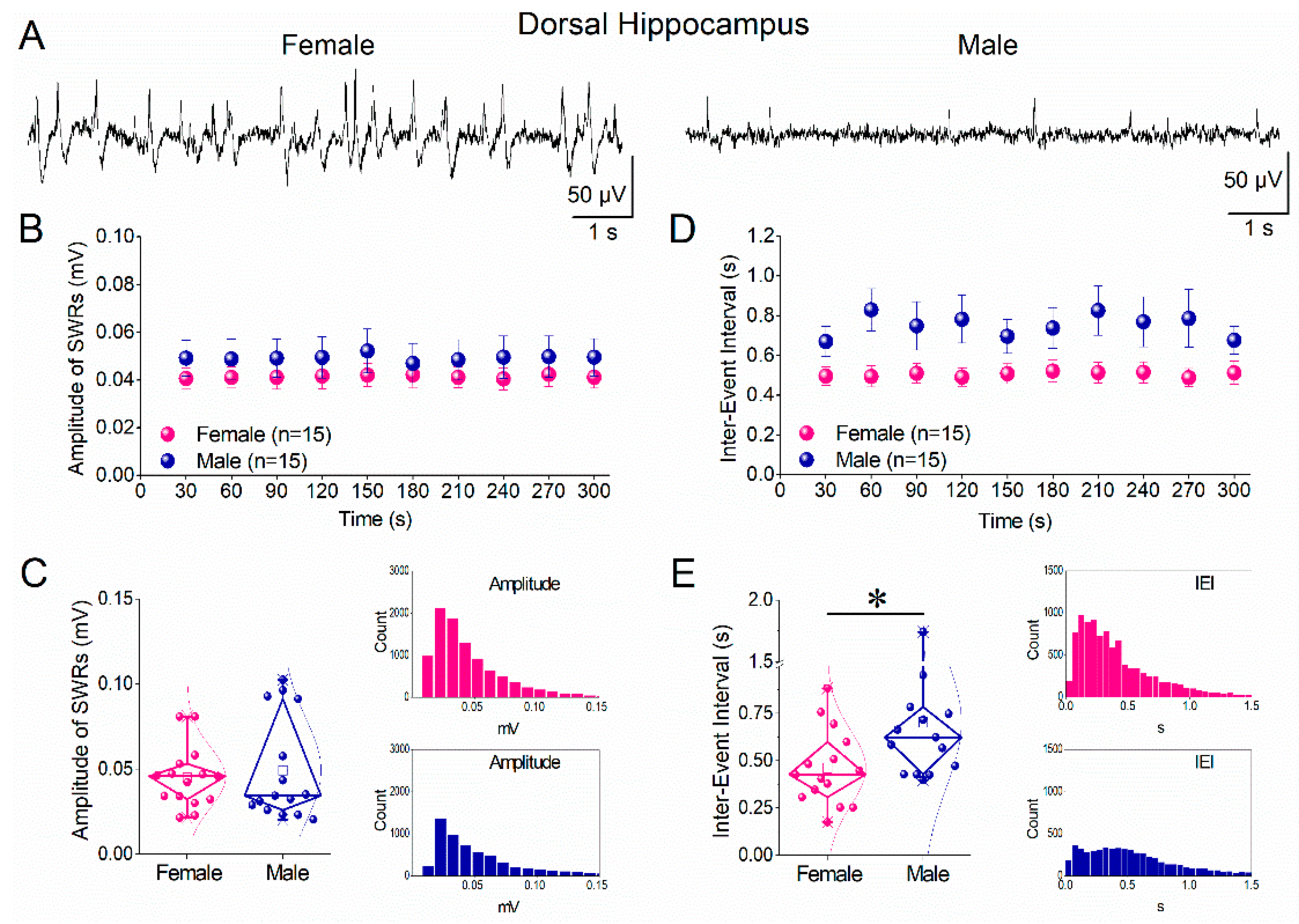
Figure 3.
Comparison of ripple oscillations and multiunit activity (MUA) between female and male dorsal hippocampus. A. Peri-event time histograms of MUA aligned to the sharp wave peak from dorsal hippocampal slices of a female (left) and a male (right) rat. B. Power spectra of local field potentials recorded from the dorsal hippocampus of female (left) and male (right) rats, illustrating ripple-band activity. C. Diamond plots showing the frequency of baseline MUA (MUA-Base; left) and SWR-associated MUA (MUA-SWR; right) recorded from the dorsal hippocampus of female (pink) and male (blue) rats. D. Diamond plots summarizing ripple power (left) and ripple peak frequency (right) in the dorsal hippocampus of female and male rats. Asterisks indicate a statistically significant sex-related difference in MUA-SWR frequency (Mann–Whitney test, p < 0.05). Individual data points are overlaid in diamond plots.
Figure 3.
Comparison of ripple oscillations and multiunit activity (MUA) between female and male dorsal hippocampus. A. Peri-event time histograms of MUA aligned to the sharp wave peak from dorsal hippocampal slices of a female (left) and a male (right) rat. B. Power spectra of local field potentials recorded from the dorsal hippocampus of female (left) and male (right) rats, illustrating ripple-band activity. C. Diamond plots showing the frequency of baseline MUA (MUA-Base; left) and SWR-associated MUA (MUA-SWR; right) recorded from the dorsal hippocampus of female (pink) and male (blue) rats. D. Diamond plots summarizing ripple power (left) and ripple peak frequency (right) in the dorsal hippocampus of female and male rats. Asterisks indicate a statistically significant sex-related difference in MUA-SWR frequency (Mann–Whitney test, p < 0.05). Individual data points are overlaid in diamond plots.
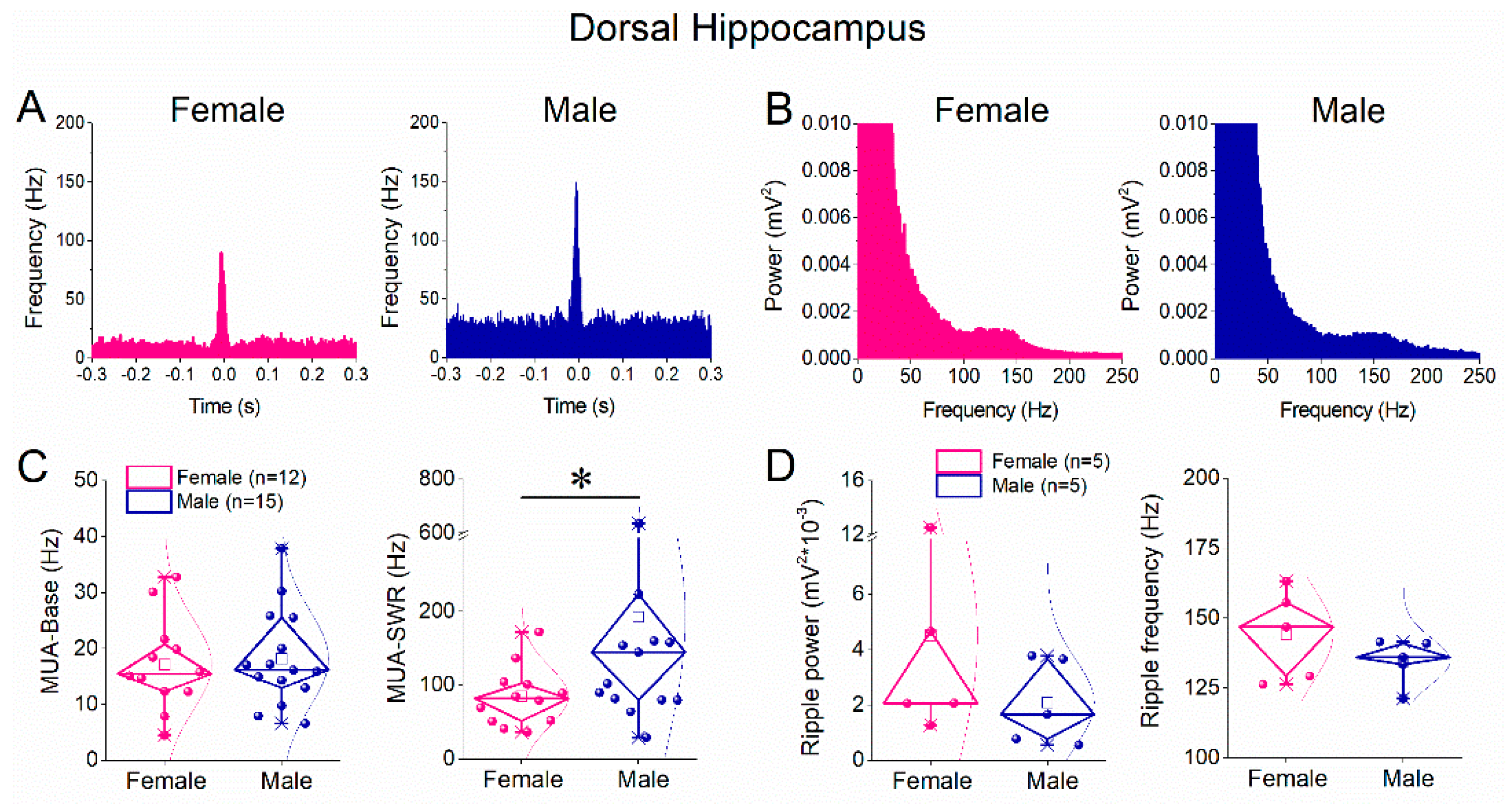
Figure 4.
Comparison of SWRs between female and male ventral hippocampus. A. Representative continuous local field potential recordings from ventral hippocampal slices obtained from a female (left) and a male (right) rat. B. Five-minute time course of SWR amplitude recorded from the ventral hippocampus of female (pink) and male (blue) rats, showing comparable amplitudes between sexes. C. Diamond plots (left) and distribution histograms (right) of SWR amplitudes from ventral hippocampal slices of female and male rats. D. Five-minute time course of inter-event interval (IEI), indicating similar SWR occurrence rates in the ventral hippocampus of female and male rats. E. Diamond plots (left) and distribution histograms (right) of IEI values from ventral hippocampal slices of female and male rats. Data indicate no significant sex-related differences in SWR amplitude or IEI in the ventral hippocampus. Data are shown as mean ± SEM in time-course plots; individual data points are overlaid in diamond plots.
Figure 4.
Comparison of SWRs between female and male ventral hippocampus. A. Representative continuous local field potential recordings from ventral hippocampal slices obtained from a female (left) and a male (right) rat. B. Five-minute time course of SWR amplitude recorded from the ventral hippocampus of female (pink) and male (blue) rats, showing comparable amplitudes between sexes. C. Diamond plots (left) and distribution histograms (right) of SWR amplitudes from ventral hippocampal slices of female and male rats. D. Five-minute time course of inter-event interval (IEI), indicating similar SWR occurrence rates in the ventral hippocampus of female and male rats. E. Diamond plots (left) and distribution histograms (right) of IEI values from ventral hippocampal slices of female and male rats. Data indicate no significant sex-related differences in SWR amplitude or IEI in the ventral hippocampus. Data are shown as mean ± SEM in time-course plots; individual data points are overlaid in diamond plots.
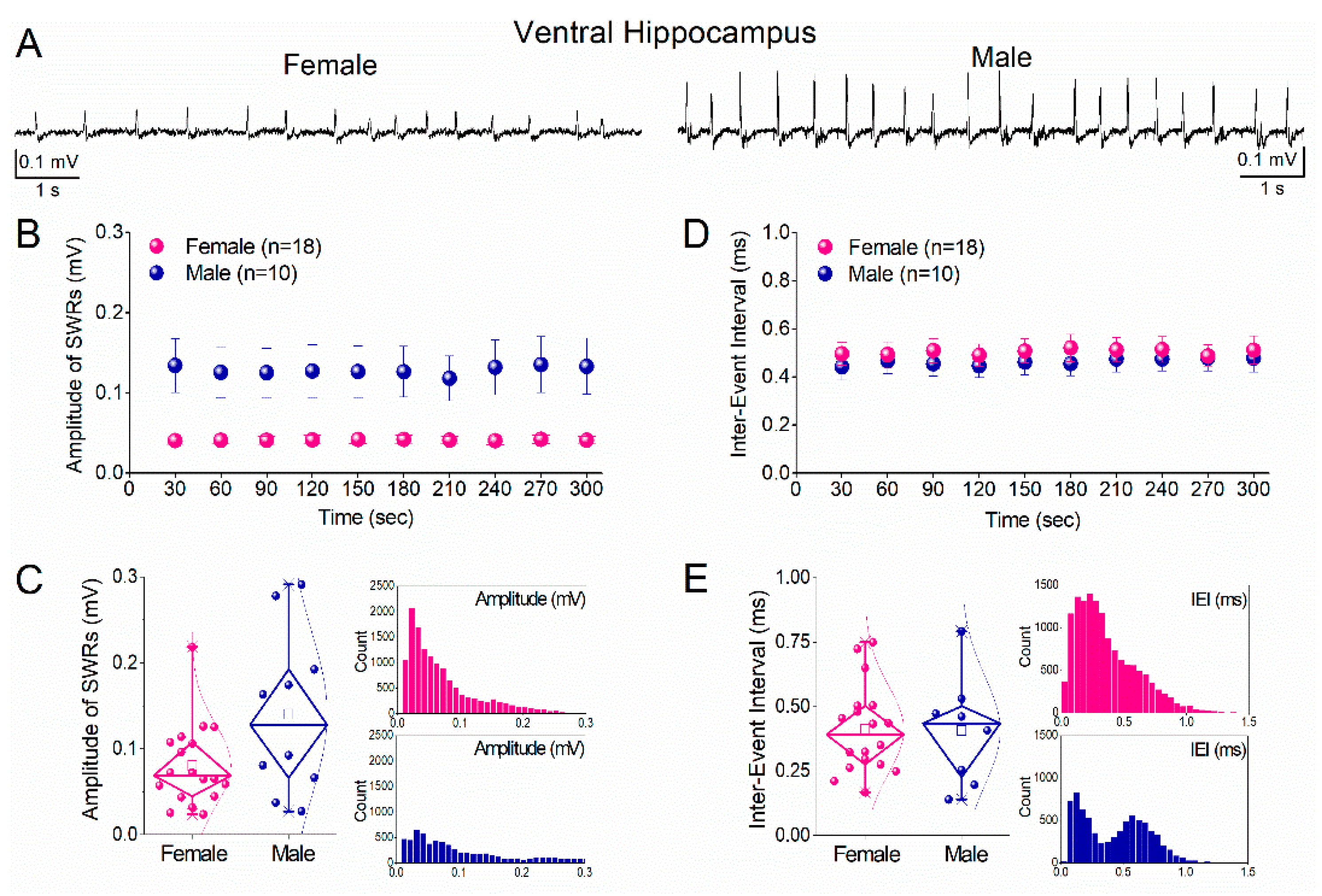
Figure 5.
Comparison of ripple oscillations and multiunit activity (MUA) between female and male ventral hippocampus. A. Peri-event time histograms of MUA aligned to the sharp wave peak from ventral hippocampal slices of a female (left) and a male (right) rat. B. Power spectra of local field potentials recorded from the ventral hippocampus of female (left) and male (right) rats, illustrating ripple-band activity. C. Diamond plots showing the frequency of baseline MUA (MUA-Base; left) and SWR-associated MUA (MUA-SWR; right) recorded from the ventral hippocampus of female (pink) and male (blue) rats. Asterisk indicates a statistically significant sex-related difference in MUA-SWR frequency. D. Diamond plots summarizing ripple power (left) and ripple peak frequency (right) in the ventral hippocampus of female and male rats. Individual data points are overlaid. Statistical comparisons were performed using the Mann–Whitney test (p < 0.05).
Figure 5.
Comparison of ripple oscillations and multiunit activity (MUA) between female and male ventral hippocampus. A. Peri-event time histograms of MUA aligned to the sharp wave peak from ventral hippocampal slices of a female (left) and a male (right) rat. B. Power spectra of local field potentials recorded from the ventral hippocampus of female (left) and male (right) rats, illustrating ripple-band activity. C. Diamond plots showing the frequency of baseline MUA (MUA-Base; left) and SWR-associated MUA (MUA-SWR; right) recorded from the ventral hippocampus of female (pink) and male (blue) rats. Asterisk indicates a statistically significant sex-related difference in MUA-SWR frequency. D. Diamond plots summarizing ripple power (left) and ripple peak frequency (right) in the ventral hippocampus of female and male rats. Individual data points are overlaid. Statistical comparisons were performed using the Mann–Whitney test (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



