Submitted:
23 February 2026
Posted:
25 February 2026
You are already at the latest version
Abstract
Seeds are the central innovation of angiosperms, earliest developmental steps of which hinge on tightly coordinated fate decisions across sporophytic and gametophytic tissues. This review synthesizes recent advances in the regulation of the early phase of seed devel-opment, beginning with ovule primordium growth and the specification of a single mega-spore mother cell (MMC), through female gametophyte maturation, double fertilization, and the onset of embryo–endosperm–seed coat programs. Canonical regulators (e.g., SPL/NZZ-centered networks and auxin transport/threshold control) operate alongside re-pressive safeguards mediated by small-RNA pathways, transposon silencing, and Poly-comb/RBR1-linked cell-cycle gates that prevent premature proliferation and autonomous seed formation. After fertilization, parent-of-origin epigenetic asymmetries and dynamic chromatin states reshape transcriptional competence in endosperm and embryo, with auxin emerging as a key mobile coordinator of endosperm growth, integument differentia-tion, and seed coat initiation. Mechanical inputs, including endosperm turgor and mech-anosensitive responses in maternal tissues, are integrated with hormone signaling to tune seed growth and the timing of endosperm cellularization. Together with the accompany-ing figures, the review provides an illustrative depiction of the earliest regulatory logic underlying successful seed formation.
Keywords:
seed
; auxin
; imprinting
; mechanosensing
1. Introduction
Seeds are fundamental to the life cycles of flowering plants, serving as the primary means of reproduction. Beyond their biological significance, they are also the key food sources for human and other animal consumption. Normally, seed development begins with a double fertilization event wherein two sperm cells from pollen fuse with two female gametes in the ovule—one sperm fertilizes the egg cell to form the diploid zygote (future embryo), and the other fertilizes the central cell to form the triploid endosperm (Adhikari et al., 2020b). These two fertilization products, together with the surrounding maternal tissue that forms the seed coat, undergo a coordinated development that results in a mature seed (Baroux and Grossniklaus, 2019). The regulation of early-stage seed development involves multiple checkpoints and signals that integrate maternal and paternal inputs (Adhikari et al., 2020b). Ovule maturation produces a fertilization-ready structure in which growth is arrested awaiting fertilization (Ingram, 2010). In the present context, the term ‘developmental block’ refers to pre-fertilization and early post-fertilization barriers that prevent autonomous embryo/endosperm development, enforced by cell-cycle gating, epigenetic repression, and constrained signaling. Fertilization alleviates these barriers and initiates synchronized growth of embryo, endosperm, and seed coat.
Several studies have addressed seed development. The present review focuses on the earliest developmental window, from MMC speciation to ovule maturation, double fertilization, and progression of seed development through onset of cellularization and selected early embryo–endosperm interface modules. It summarizes current mechanistic models for sporophyte-gametophyte communication and coordinated growth of the embryo, endosperm, and seed coat. Evidence is drawn primarily from Arabidopsis, with selected comparisons to other species where they sharpen or challenge Arabidopsis-based models.
2. Evolutionary Triggers Behind Seed (Plant) Development
Angiosperm success on land is closely linked to innovations in reproductive development, including ovules, seeds, and associated maternal–offspring coordination. Comparative syntheses highlight that evolutionary inference for ovule/seed traits remains constrained by sampling and analytical limitations, and mechanistic generalizations should therefore be treated cautiously (Mathews and Kramer, 2012, Rudall, 2021, Benton et al., 2022, Boyko and Vasconcelos, 2025).
One influential hypothesis proposes that the ovule represents a modified shoot-derived developmental program, motivated by structural similarities between the nucellar apex and shoot apex and by nucellar expression of the shoot stem-cell regulator WUSCHEL (WUS) (Figure 1) (Mathews and Kramer, 2012). This view is compatible with ‘nested program’ models (synonymously referred to as ‘babushka doll’ and ‘hierarchical shifts’) in which inflorescence, flower, and ovule like complex branching structures development emerge through hierarchical redeployment of organogenesis modules. Integument regulators such as YABBY factors (including INNER NO OUTER (INO)) further illustrate the reuse of laminar organ-growth regulators in reproductive structures (Yamada et al., 2011, Arnault et al., 2018, Rudall, 2021).
Comparative anatomy also supports multiple evolutionary trajectories for the outer integument (OI) in bitegmic ovules. The OI has been proposed to represent a later innovation in angiosperms and, in some lineages, a convergent feature; ovules of Gnetales also possess an outer envelope that is suggested to originate from fusion of lateral bracts, consistent with initiation from two primordia (Rydin et al., 2010, Rudall, 2021). Given that auxin–cytokinin interplay can modulate integument initiation and bitegmic–unitegmic-like features in Arabidopsis, hormonal rewiring is a plausible route by which developmental programs were evolutionarily repurposed (Bencivenga et al., 2012).
At the cellular level, meiosis and double fertilization represent major evolutionary transitions that frame the pre- and post-fertilization ‘developmental blocks’ discussed below. The origin of meiosis has been argued to reflect selection for recombination in large eukaryotic genomes and avoidance of the long-term mutational accumulation (Muller’s ratchet) (Figure 2) (Muller, 1964, Takeuchi et al., 2014, Berger, 2024). It is an integral part of angiosperm reproductive success as it plays a fundamental role in genetic and epigenetic reset during the process. Angiosperm double fertilization typically produces zygote and endosperm. Two principal hypotheses for endosperm origin have been advanced primarily based on the inferential evidences: derivation from a supernumerary embryo versus derivation from a sexualized female gametophyte (FM). Conserved features such as micropylar–chalazal asymmetry of nuclear division are observed across basal angiosperms (Floyd and Friedman, 2000, Baroux et al., 2002, Becker et al., 2025). These evolutionary perspectives motivate the mechanistic focus of subsequent sections on how fertilization alleviates developmental blocks and coordinates early embryo, endosperm, and seed coat growth.
3. Ovule Development
At the onset of megasporogenesis, a single L2 cell differentiates into the megaspore mother cell (MMC), committing to meiosis and the sporophytic-to-gametophytic transition. The to-be-MMC cell speciates specifically from a single cell of the layer 2 (L2) in the developing ovule primordium indicating a precisely targeted molecular regulation during the process (Figure 3). Some of the key factors involved in the MMC speciation is shown in Figure 4, Figure 5 and Figure 6. One of the key factors behind the speciation process is the PRC1-mediated repression of an RdDM-associated 24-nt miRNA-partner effector protein encoding gene, ARGONAUTE 9 (AGO9). The process is facilitated by the PRC1 core component, E3 ligases, Really Interesting New Gene 1 (RING1) and RING2-mediated histone monoubiquitylation (H2Aub1) of the locus. Mutations in RING1A/B cause defects in MMC/FM specification and FM mitoses and coincide with ectopic expression of AGO family genes (including AGO9), resulting in aborted ovules (Lv et al., 2024). PRC1 and PRC2 complexes are known to play pivotal role during developmental transitions. PRC1 typically monoubiquitylates the histone (H2Aub1), which is suggested to be a PRC2 recruitment signal for subsequent histone methylation (H3K27me3). In such case, the PRC2 recruitment process likely requires H2Aub binding protein, which however, has not been identified yet. The PRC2 recruitment independent of ubiquitylation is also a common occurrence in plants (Liu et al., 2025b). As one recent study observed, H2Aub predominantly marks the repressed genes with no H3K27me3 marks. On transposable elements (TEs) however, both marks tend to colocalize. Nevertheless, the ubiquitylation in the former and methylation of the latter case play deterministic regulatory role (Figure 4) (Wu et al., 2025).
Typically, the PRC1-regulated AGO9 is expressed in the nucleus of MMC (Rodríguez-Leal et al., 2015) and cytoplasm of the apical somatic cells (Olmedo-Monfil et al., 2010). Furthermore, the defect in the AGO9-interacting partner miRNA822 leads to the persistence of the more than one post-meiotic cells. The ago9 mutant or (ovule-specific) miRNA822-target genes defective mutants exhibit similar phenotype (Tovar-Aguilar et al., 2024). The ring1a/b mutants exhibit reduced expression of a MADS-box member, SEEDSTICK (STK) (Chen et al., 2016), a positive regulator of AGO9 and RNA-DEPENDENT RNA POLYMERASE 6 (RDR6), thereby negatively affecting SPL/NZZ expression and MMC speciation (Mendes et al., 2020). RDR6 is known to convert single-stranded precursors into double-stranded RNA, the process dependent on the function of a RNA binding protein SUPPRESSOR OF GENE SILENCING 3 (SGS3) during trans-acting siRNA biogenesis (Figure 4) (Yoshikawa et al., 2005).
As in ago9 mutants, sgs3 and rdr6 mutants develop supernumerary MMCs. This supports a model in which tasiRNA-mediated silencing moves between companion somatic cells to restrict MMC fate in a non-cell-autonomous manner (Olmedo-Monfil et al., 2010, Rodríguez-Leal et al., 2015). AGO9 also promotes DOMAIN REARRANGED METHYLTRANSFERASE 1 (DRM1) and DRM2-mediated silencing (as part of RdDM) at the SPL/NZZ locus and may contribute to SPL/NZZ mRNA degradation in surrounding hypodermal somatic cells, thereby allowing MMC fate acquisition by a single central cell (Figure 4) (Matzke and Mosher, 2014, Mendes et al., 2020). Recent work further suggests that single MMC specification depends on balanced methylation and demethylation rather than methylation alone (Jiang et al., 2025).
The study suggested that DRM2 and DME act on largely distinct target sets. Approximately 30% of ovule primordia showed co-expression of DRM2 and DME in surrounding L2 cells, and dme mutants produced more than one MMC-like cell. In the normal MMC, DRM2 and DME expression declines, whereas neighboring L2/L3 cells retain DME expression with reduced DRM2 (Figure 4) (Jiang et al., 2025).
SPL/NZZ facilitates the recruitment of TOPLESS (TPL) and co-repressor at the CINCINNATA (CIN)-like TEOSINTE BRANCHED1/CYCLOIDEA/PCF (TCP) bound locus to repress the CIN-like TCP-regulated genes via the EAR motif (LXLXL) at its C-terminus. The process is essential for MMC speciation and development (Wei et al., 2015). SPL/NZZ additionally acts as a part of a MADS-box-associated complex to target the canonical CArG motifs in the ANTEGUMENTA (ANT) promoter and contributes to its repression. When active, ANT represses the auxin flux pump encoding member PIN1, thereby hindering the auxin-mediated AUX/IAA degradation. Degradation of AUX/IAA is essential for AUXIN RESPONSE FACTOR (ARF) release to act on downstream targets involved in MMC development (Figure 5) (Cavalleri et al., 2025). PIN1-driven auxin transport establishes a micropylar auxin maximum and a chalazal minimum in the ovule primordium; SPL/NZZ (with miR160-restricted ARF17) supports the PIN1 expression domain underlying the micropylar maximum, whereas BEL1 restricts PIN1 at the chalaza to maintain the auxin minimum (Bencivenga et al., 2012, Huang et al., 2022, Yu et al., 2025).
The hypodermal somatic cells surrounding the MMC are repressed from attaining the MMC fate by the SWI/Snf-related group 1 (SWR1), an ATP-dependent chromatin remodeling complex in plants. The SWR1 complex mediates the deposition of histone variant H2A.Z around the transcription start site (TSS) of WRKY28 locus via its component ACTIN RELATED PROTEIN 6 (ARP6) and promotes its expression. The process is non-cell-autonomously enhanced by the inner integument expressed KLUH (KLU) encoding cytochrome P450 protein. WRKY28, then actively represses the hypodermal somatic cells from attaining the MMC fate (Figure 6) (Zhao et al., 2018a, Aslam et al., 2019). In the inner integument, KLU contributes to cell proliferation regulates the final seed size (Adamski et al., 2009), which is however counteracted by jasmonate signaling-associated network (Figure 6) (Zhang et al., 2025). Regarding gametogenesis, the SWR1 complex further regulates the gametophyte development post-MMC speciation. Its component ARP6 hinders the expression of DISRUPTED MEIOTIC cDNA1 (DMC1) in the megasporocyte and surrounding non-sporangenous ovule cells prior to meiosis. After the initiation of meiosis process, however, ARP6 activates DMC1 exclusively in the megasporocyte and the process is facilitated by the SWR1 complex-mediated incorporation of the histone variant H2A.Z at the DMC1 locus (Qin et al., 2014).
4. Ovule Maturity and Growth Progression Halt
Normal ovule maturation culminates in a state of readiness for fertilization (Figure 1) (Adhikari et al., 2020b). This pre-fertilization pause is not a passive cessation of activity but rather an actively maintained state. Rhe phase of cell cycle during this quiescent period may vary among different plant species (Liu et al., 2020). This regulatory control guarantees that seed development (and the provisioning of nutrients) proceeds only when a viable embryo is formed, typically after a successful fertilization. Due to such a developmental roadblock, mature ovules typically have a limited lifespan and undergo active programmed cell death afterwards (at 5-6 days after anthesis in Arabidopsis). As the ovules age, plants initiate senescence of the unfertilized ovules through ethylene signaling and activation of cell death-associated genes such as the triad of NAC-family transcription factors, in their outer tissues during the process (Van Durme et al., 2023).
The post-maturity developmental progression halt of the gametes is largely regulated by a plant RETINOBLASTOMA RELATED (RBR) homolog, RBR1 and its downstream targets in central cell. The whole process can be separated into three distinct modules- a) cell-cycle gate control, b) imprinting setup, and c) FIS-PRC2 execution (Figure 7). RBRs are attributed for their positive role in maintaining cellular quiescence at diverse tissues during plant development (Gombos et al., 2023). In the mature ovule, RBR1 suppresses the E2F-DP complex targets, which include the G1-to-S phase transition-associated genes, by directly interacting with the complex and halts the central cell division (Ebel et al., 2004).
CYCD7;1 phosphorylates the E2F-DP-bound RBR1 thereby sliding it off of its bound loci (Weimer et al., 2018). The rbr1 mutant and the lines ectopically expressing CYCD7;1 exhibit autonomous endosperm proliferation as the E2F-DP bound loci are freed of RBR1 in both cases (Ebel et al., 2004, Sornay et al., 2015, Weimer et al., 2018). While the rbr1 mutant ovules are rarely fertilized, the ovules ectopically expressing CYCD7;1 exhibits successful fertilization producing seeds relatively larger than that produced by WT (Figure 7) (Ebel et al., 2004, Sornay et al., 2015). In addition to its role in cell cycle control, RBR1, along with the WD40 domain harboring PRC2 interacting protein, MSI1, suppresses MET1 expression (Johnston et al., 2008), which leads to the gradual decrease in methylation of maternally imprinted genes such as FIS2 and FWA during female gametogenesis after each cycle of DNA replication. The residual methylation in the loci is further removed by DME thereby activating the FIS2-led developmental block of central cell in a mature ovule (Figure 7) (Jullien et al., 2008). Analogous observation was made on a gene encoding an additional PRC2 subunit, MEA (Xiao et al., 2003).
The paternal MEA and RBR1 are repressed by PRC2-mediated H3K27me3 in pollens. The paternal RBR1 repression continues after fertilization as well, which is mediated by MEA-PRC2 (Johnston et al., 2008). The third module governing growth progression halt in mature ovule is FIS-PRC2, which deposits H3K27me3 marks to its target loci. As observed in Arabidopsis a HETEROCHROMATIN PROTEIN 1 family member TFL2/LHP1 broadly co-localizes with H3K27me3-marked genes (>85-90% overlap at the mapped sites), and H3K27me3 profiles remain largely similar in lhp1 seedlings, indicating that LHP1 likely acts downstream as a reader/effector of Polycomb chromatin (Turck et al., 2007). LHP1 interacts with MSI1 thereby contributing to spread in H3K27me3 deposition toward 3′-ends of the target gene in a context dependent manner and shape local/3D chromatin topology (Veluchamy et al., 2016). A recent Arabidopsis study showed that DEK proteins physically interact with LHP1 and loss of DEK causes genome-wide H3K27me3 increases, consistent with a role in H3K27me3 homeostasis (Nakamura et al., 2026). In mammalian systems, DEK binds nucleosomes (including via H2A/H2B-dependent contacts), compacts polynucleosomes, and enhances PRC2 methyltransferase activity in vitro (Alexiadis et al., 2000, Kujirai et al., 2025). The different observations made by the two studies on the H3K27me3 change likely reflect differences between reconstituted in vitro and in vivo regulatory contexts (Kujirai et al., 2025, Nakamura et al., 2026) (Figure 8).
Although DEK proteins have been linked to chromatin compaction and Polycomb-associated states in multiple systems, caution is warranted when extrapolating mechanistic conclusions across kingdoms or experimental contexts. Much of the mechanistic detail for DEK derives from in vitro or cultured-cell studies where chromatin composition, cell-cycle state, and accessory factors can differ markedly from those operating in planta during ovule maturation. In Arabidopsis, genetic and molecular associations between DEK, LHP1, and Polycomb-linked repression are consistent with a role for DEK in shaping repressive chromatin environments, but they do not, on their own, establish that DEK functions as a conserved structural subunit of LHP1/PRC machinery or that it executes an identical biochemical mechanism to that inferred in mammalian systems {Nakamura, 2026 #2367;Alexiadis, 2000 #2368}. Accordingly, the DEK–LHP1–PRC2 relationship is best framed as a working model in which DEK may modulate Polycomb occupancy, chromatin accessibility, or higher-order chromatin organization in a context-dependent manner, pending direct evidence from ovule-stage biochemical interaction tests and locus-resolved chromatin profiling.
Rather than maintaining central-cell quiescence via a single downstream target, FIS–PRC2 is best framed as preventing premature activation of an endosperm-like program before fertilization, with evidence that this repression can involve auxin-related outputs and type I MADS-box regulators including AGL62. Unlike the lines with defects in the FIS-PRC2 module, ovules with perturbed cell-cycle gating (rbr1 and ectopic CYCD7;1 expression) can show autonomous endosperm-like proliferation but do not initiate seed coat development, indicating that fertilization-linked signals are required for full sporophytic responses. A recent study highlighted that FIS-PRC2 also suppresses embryo-nourishing programs, including auxin biosynthesis genes, before fertilization (Heidemann et al., 2025). Consistent with this framework, ovule maturation is linked to repression of auxin biosynthesis in the central cell and to constrained sugar influx, and integument growth ceases unless fertilization-associated auxin export and/or pollen tube reception signaling is perceived.
In contrast to the central cell, the egg cell exhibits a stricter pre-fertilization developmental block, and the underlying mechanisms remain less well resolved. Available evidence supports at least three partially separable modules: (i) a cell-cycle gate, (ii) Polycomb-linked repression involving MEA–PRC2, and (iii) suppression of embryo-program execution (Figure 9).
While RBR1 is normally expressed in central cell before fertilization and acts as one of the key fertilization-dependent gate (at S-phase), it is typically not expressed at the egg cell until fertilization (Ingouff et al., 2007, Simonini et al., 2024). Instead, as more recent Arabidopsis studies suggest egg cell is typically arrested at the G2 stage of cell division (Simonini et al., 2024, Simonini, 2025). Typically, the anaphase-promoting complex/cyclosome (APC/C) is known to regulate the activation of the genes associated with mitosis - the subsequent stage in the cell cycle – via ubiquitylation of the repressive Cyclin B protein (CycB). An Arabidopsis study presented inferential evidence strongly suggesting presence of such mechanism in mature plant egg cell (Figure 9). The study highlighted that its APC/C member, APC4 is expressed during the mega-gametogenesis and embryogenesis processes and perturbation of APC4 leads to the accumulation of CycB protein, aberration in gametogenesis, and defect in embryogenesis (Wang et al., 2012).
RBR1 in embryo is attributed for its role in embryo patterning rather than fertilization gating. The MEA-PRC2-mediated controlled regulation of CYCD1.1 by depositing H3K27me3 marks at its locus contributes to the process by affecting the RBR1 phosphorylation rate in the patterning embryo (Figure 9) (Simonini et al., 2021). Additionally, defect in the PRC2 interacting protein, MSI1 brings autonomous embryo (n) and endosperm (2n) development as observed in Arabidopsis (Guitton and Berger, 2005). The mutant ovules, however abort at the pre-globular to globular stage (Köhler et al., 2003). It had also been proposed that MSI1 and FIE, subunits of MEA-PRC2 complex, likely contribute to the RBR1-mediated cell cycle gate (Figure 9) (Guitton and Berger, 2005). FIS2 and MEA exhibit detectable exclusive expression at the central cell and endosperm while MSI1 and FIE expression is extended to sporophytic tissues as well . The consequence of MEA defect on embryogenesis however is more severe when it is in egg cell/embryo (hyperproliferative with disorganized development) than when it is in the central cell/endosperm (Simonini et al., 2021).
The third module (embryo program) largely depends on a PLETHORA member BABY BOOM (BBM). In Arabidopsis, an RK domain-containing factor, RKD5, contributes to repression of BBM in the mature unfertilized egg cell; rkd5 mutants show a low frequency of parthenogenic seed development, implying that additional repressors constrain BBM activity (Figure 9) . Native BBM expression is silenced in the egg cell before fertilization and is detected in sperm cells, followed by biparental expression shortly after fertilization (Khanday et al., 2019). An aposporous millet accession expresses BBM-like genes before fertilization, and promoter-gene combinations from this system can trigger apomixis-like outcomes when introduced into sexual backgrounds (Conner et al., 2015). Ectopic BBM expression in the egg cell using heterologous constructs can induce parthenogenesis more efficiently, consistent with potential species-specific constraints on embryogenic competence (Chen et al., 2022).
In addition to the independent regulatory networks of the egg and central cells, studies indicate their interaction effect as well. The two component cells have functional plasmodesmatal connections between them facilitating the central cell to egg cell siRNA movement (Schröder et al., 2023). It suggests that the hypomethylation of central cell, mainly by DEMETER (and subsequent 24nt-siRNAs biogenesis and their movement to the egg cell is one of the key factors for the hypermethylation of egg cell (Gehring et al., 2009, Bauer and Fischer, 2011, Lafon-Placette and Köhler, 2014).
5. Double Fertilization and Alleviation of the Molecular Block Against Seed Development
Double fertilization coordinates parental-genome union and triggers rapid transcriptional/epigenetic reprogramming. After the release of the sperm cells in the ovule, the independent fertilization of the two female gametes, egg and central cells, occurs almost simultaneously with the event commencing slightly earlier on the former than on the latter. Nevertheless, endosperm proliferation normally precedes post-zygotic proliferation (Figure 10) (Lafon-Placette and Köhler, 2014, Doll et al., 2023).
Studies show that erasure of old epigenetic marks (parental) and incorporation of fresh ones (gametophytic) occurs in two successive stages of gametophyte development. Strong evidence on it comes from a Merchantia study, in which the authors observed that meiosis in the species coincides with the overall epigenetic marks erasure, while the subsequent sporulation step deposits fresh epigenetic marks by gamete maturation (Montgomery and Berger, 2023). Such instance also explains how it is possible for each of the FM component cells to exhibit unique epigenetic marks as observed in Arabidopsis (Migicovsky and Kovalchuk, 2012).

Table 1.
Developmental timeline from pollen tube reception to early heart stage (Arabidopsis).
| Stage / window | Endosperm | Embryo | Maternal integument / testa |
|---|---|---|---|
| Pollen tube reception / pre-syngamy (POEM) | Central cell/endosperm-like responses can be triggered by pollen tube content release without gamete fusion (Kasahara et al., 2016, Honma et al., 2020). | No zygote formed. | PTC can initiate chalazal seed-coat wall formation; auxin can phenocopy aspects of initiation (Figueiredo et al., 2016, Liu et al., 2019). |
| Double fertilization | Paternal miR159 helps relieve maternal barriers by targeting GAMYB members (MYB33/MYB65) (Zhao et al., 2018b). | Fertilization initiates embryogenesis and relieves pre-fertilization arrest programs (Adhikari et al., 2020b). | Maternal tissues become responsive to fertilization-derived signals that promote seed coat initiation (Figueiredo et al., 2016). |
| Early coenocytic endosperm (pre-globular) | AGL62 supports/maintains the coenocytic program and prevents premature cellularization; an imprinted PHE1 network supports proliferation and delays cellularization (Hehenberger et al., 2012, Batista et al., 2019). | Early patterning is established as zygotic programs become active (Nodine and Bartel, 2012). | Endosperm expansion generates mechanical load on integuments (turgor) (Beauzamy et al., 2016). |
| Late coenocytic endosperm / late globular | Endosperm turgor approaches a maximum near the end of the coenocytic stage and then declines before cellularization (Beauzamy et al., 2016). | Embryo reaches globular stage prior to heart-stage organogenesis (Figure 10). | Chalazal seed coat differentiation includes activation of BAN and PA deposition (Debeaujon et al., 2003). |
| Early heart stage (cellularization onset) | Cellularization initiates micropylar-to-chalazal; ABA/ABI5 represses the SHB1–MIN3–IKU2 proliferation module (Hehenberger et al., 2012, Cheng et al., 2014, Li et al., 2022). | Heart-stage morphogenesis proceeds alongside increasing zygotic genome activity (Zhao et al., 2019). | As cellularization proceeds, mechanosensitive testa responses are proposed to constrain growth via GA deactivation (ELA1/2) and wall stiffening (Creff et al., 2015, Creff et al., 2023). |
Stages are presented qualitatively; exact timing varies with accession and growth conditions.
5.1. Pre-Syngamy Changes
The ovules exhibit distinct feature change upon pollen tube reception even in absence of subsequent gamete fusion. As the observations show, the central cell response can simply be triggered by pollen tube content release inside the ovule without subsequent gamete fusion (Kasahara et al., 2016, Honma et al., 2020). In rice, such phenomenon facilitates unperturbed sugar influx, which however, cannot be converted into starch - as would normally occur in the fertilized ovule (seed) - thereby developing the sugary grain (Honma et al., 2020). In Arabidopsis, a typical dicot, the ovule exhibits a typical ‘pregnant’ characteristic – POEM-like ovule enlargement morphology – the feature termed as pollen tube-dependent ovule enlargement morphology (POEM) (Figure 10) (Kasahara et al., 2016). Additionally, some studies show that the release of pollen tube content (PTC) alone is sufficient to trigger the cell wall initiation at the chalazal end (Liu et al., 2019). However, earlier study showed that the post-fertilization auxin biosynthesis in the central cell is essential for the cell wall initiation process and demonstrated that exogenous auxin applied ovules phenocopy the trait (Figueiredo et al., 2016). Since the synergid cells are known to constitute auxin albeit at a relatively low level (Sun et al., 2021), whether the seed coat initiation observed by (Liu et al., 2019) is the outcome of the burst of the pollen tube receiving synergid or the PTC itself remains to be determined.
5.2. The Coenocyte
Paternal miR159 copy plays a key role in clearance of maternal barrier of endosperm proliferation. Among others, its targets include GAMYB members, MYB33 and MYB65, which are highly expressed in the pre-fertilized central cell in Arabidopsis (Zhao et al., 2018b). The RBR1-mediated cell-cycle arrest is released upon gamete fusion by putative RBR1 phosphorylation (and degradation) mediated by the sperm cell derived CYCD7;1 and CDKA;1 in the central cell/endosperm thereby easing the cell-cycle gate (Figure 7) (Sabelli et al., 2013, Simonini et al., 2024). Fertilization also brings changes to the MET1/DME-mediated methylation status. While MET1 is expressed at both pre- and post-fertilized endosperms, expression of DME is significantly reduced after fertilization and contributes to the suppression of paternal MEA allele (Choi et al., 2002, Xiao et al., 2003, Gehring et al., 2006). Arabidopsis DME reportedly harbors repressive and enhancing cis-elements at its post-TSS region likely contributing to its fertilization event-regulated expression behavior (Figure 11) (Park et al., 2017). The maternal MEA-PRC2, still active in the post-fertilized endosperm, contributes to the H3K27me3 marks deposition and silencing of the paternal MEA and RBR1 (Figure 7) (Gehring et al., 2006, Johnston et al., 2008).
With the diminished level of functional FIS-PRC2 in the post-fertilized endosperm, expression of AGL62 increases (Hehenberger et al., 2012). AGL62 supports the coenocytic endosperm program and influences seed growth in a dosage-sensitive manner, with genetic evidence placing it within an early endosperm transcriptional module that includes PHE1 and related type I MADS factors (Kradolfer et al., 2013). In strawberry, AGL62 co-acts with AGL80 in support of early endosperm proliferation programs (Guo et al., 2022).
A mechanistic model for parental asymmetry at PHE1 target loci proposes that PHE1 binds CArG-like motifs and activates AGL62 and other type I MADS-box genes, while imprinting outcomes are shaped by allele-specific chromatin and cis-element configurations. In this model, many paternally expressed PHE1 targets are associated with RC/Helitron transposable-element-derived sequences near the PHE1-binding region and lack gene-body H3K27me3 on the paternal allele, whereas maternally expressed PHE1 targets more frequently exhibit non-CG methylation at the PHE1-binding site on the maternal allele (Batista et al., 2019).
The coenocytic endosperm proliferation is mediated by facilitating the expression of the paternally imprinted crucial auxin biosynthetic genes YUC10 and TAR1 via repression of their repressors, the homeodomain-leucine zipper transcription factors - the ATHB members (Figure 11) (Figueiredo et al., 2015, Figueiredo et al., 2016, Guo et al., 2022). The persistent synergid is eliminated by merging with the developing endosperm by its second division which is pivotal for blocking polytubey (Maruyama et al., 2015). Additionally, F-actin plays a crucial role in the nuclear positioning within the endosperm at its coenocytic phase. The process is dependent on the microtubules, which prepares the foundation for the F-actin anchorage to the divided nuclei. Perturbation of either microtubule or F-actin polymerization negatively affects nuclear division and positioning, reduces coenocytic cavity size, and compromises seed development and final seed size (Ali et al., 2023).
Arabidopsis fis2 mutant ovules show ectopic expression of YUC10 and TAR1 together with AGL62 and trigger autonomous endosperm proliferation, but do not initiate seed coat development (Roszak and Köhler, 2011, Pankaj et al., 2024a). In addition to failed cellularization, the mutant exhibits increased hexose levels (Hehenberger et al., 2012). AGL62 defects perturb auxin efflux from the endosperm, consistent with a role for endosperm-derived auxin in coordinating seed coat initiation and growth (Figueiredo et al., 2016).
The endosperm-synthesized auxin is quickly exported to the integument potentially via the AGL62-regulated ABCB transporter, PGP10 in the fertilized ovule. The process may additionally be facilitated by the genes such as TWD1 by contributing to the membrane localization of PGP10 (Wu et al., 2010, Figueiredo et al., 2016, Tsering et al., 2025). The transported auxin is distributed among the inner and outer integument layers potentially via PIN3 and AUX1 respectively (Figure 11) (Larsson et al., 2017, Liu et al., 2023).
5.3. Seed Coat Development
This fertilization derived endosperm signal contributes to the suppression of sporophytic PRC2 (EMF/VRN-PRC2). As a mutation study showed defect in either or both of integument expressed PRC2 components VRN2, encoding a PRC2 Su(z)12 subunit, and SWN, a MEA homolog, leads to an increase in the autonomous seed development and seed coat formation (Roszak and Köhler, 2011). The process is mediated by the de-repression of the otherwise EMF/VRN-PRC2-repressed gibberellin (GA) biosynthesis and TRANSPARENT TESTA (TT) pathway-associated genes (Figueiredo and Köhler, 2018). It is notable to mention that the PRC2-repression-led activation of TT16 and GOA in the endothelium (inner layer of the seed coat) and nucellus redundantly promote nucellus degeneration via repression of HVA22d, a suppressor of GA-mediated PCD and autophagy, while the endosperm proliferates. TT16 is also essential for subsequent seed coat development and serves as a key factor linking the (central cell) fertilization signal to maternal response (Figure 11) (Xu et al., 2016).
The unfertilized ovules also constitute auxin but at its inactive (conjugated) form (Aloni et al., 2006). Auxin conjugation has been suggested to be facilitated by GH3.5 based on its synthesis in the unfertilized (at the micropylar end of central cell) and fertilized ovules (at the apical end of funiculus) (Larsson et al., 2017). Post-fertilized ovules exhibit synthesis of an auxin deconjugate, a basic helix-loop-helix leucine zipper protein, ILR1 at its micropylar region (Rampey et al., 2004) suggesting its potential role in activating/freeing the conjugated IAA (Vanneste et al., 2025). ARF2, on the other hand, negatively regulates seed size in response to auxin by conferring repressive effect in the integument cell proliferation and organ growth (Figure 11) (Schruff et al., 2006).
Endosperm-to-integument signaling is proposed to intersect with carbohydrate gating at the chalaza. Auxin has been suggested to enhance starch accumulation and maternal sucrose responsiveness by promoting SWEET10 and INV4 expression and by influencing callose-gate dynamics at the chalazal phloem end, which could increase sucrose flux and elevate T6P. The sucrose transport is additionally facilitated by the downregulation of callose biosynthesis-associated genes, CALS5/6/7 (Xu et al., 2025). Fertilization event is known to trigger the active degradation of the phloem end callose gate at the chalaza (Kasahara, 2025, Liu et al., 2025a, Nakajima and Kasahara, 2025). The carbohydrate changes in the developing seed may connect to TOR signaling and GA-linked outputs that accompany fertilization responses (seed coat initiation and seed development); however, the TOR-centered branch should be framed as emerging rather than fully established, pending targeted genetic tests and stage-resolved quantification (Figure 11).
Studies have shown that very early stage (pre-globular to globular) seeds exhibit high level of GA-biosynthesis and GA-responsive genes in its micropylar region including the embryo tissues such as suspensor and embryo neck (Solfanelli et al., 2005, Ziegler et al., 2019). When observed in a broader developmental time-frame (from globular stage to maturity), in Arabidopsis, a significantly higher expression of GA biosynthesis genes, GA3ox3 and GA3ox4, in addition to those associated with cytokinin and abscisic acid biosynthesis have been detected in the chalazal endosperm (CZE) region (Belmonte et al., 2013). The seed coat (proanthocyanidin (PA) deposition) exclusively initiates at the chalazal integument region adjoining the putative CZE, which is consistent even in the unfertilized ovule when GA or auxin are exogenously applied (Figueiredo et al., 2016). An Arabidopsis study showed that expression of a key gene involved in seed coat PA biosynthesis, BANYULS (BAN) (Figure 12), peaks at late globular to early heart stage (Debeaujon et al., 2003), while bioactive GA peaks at the bent cotyledon stage with a gradual increase in DELLA proteins in subsequent developmental stages peaking at the maturity (Hu et al., 2018).
The coenocytic phase and the cavity volume of the coenocytic endosperm is often regarded as positive attributes behind the seed size. The internal turgor pressure of the endosperm can reach up to 1.5 MPa near the end of the coenocytic stage in Arabidopsis before dropping down to about 0.6 MPa and subsequent cellularization (Beauzamy et al., 2016). The high turgor pressure confers positive effect on seed size while also contributing to the rigidity (wall stiffening) of the testa (Creff et al., 2023).
5.4. Seed Size Regulation (Endosperm-Dependent)
Endosperm turgor pressure is a major mechanical determinant of seed growth, reflecting both promotion of expansion during the coenocytic phase and restriction as maternal tissues stiffen (Beauzamy et al., 2016, Creff et al., 2023). In Arabidopsis, paternally imprinted genes promote auxin biosynthesis and endosperm proliferation, increasing turgor, whereas maternally imprinted PRC2 members contribute to a later reduction (Figueiredo et al., 2015, Beauzamy et al., 2016). Elevated turgor reorients and bundles microtubules in outer integument L2 cells (oi L2) and is associated with stiffening of the adaxial inner wall (wall 3), a response linked to the wall-localized mechanosensitive CYP module ELA1 and GA deactivation (Figure 12 and Figure 13). Seed growth slows once cellularization begins (early heart stage), and ELA1 expression peaks around torpedo stage, consistent with subsequent restriction of size expansion by wall-3 stiffening (Creff et al., 2015, Creff et al., 2023). Additionally, the wall 3 of iku2 mutant exhibits relatively higher instances of demethylesterified homogalacturonans and higher stiffness as compared to col-0 suggesting the higher level of Ca2+-mediated cell wall reinforcement by cross-linking demethylesterified homogalacturonans at the testa cell wall of the mutant ovule (Hamann, 2012, Creff et al., 2023).
FER is additionally known to mediate ROS production and TIR1/AFB1 oxidation thereby contributing to the auxin-signaling-dependent growth attributes (Duan et al., 2010, Lu et al., 2024). Arabidopsis seeds exhibit spike in FER accumulation in the abaxial layer (L1) of the outer integument (oi)/testa at globular stage (50 hours after pollination) and FER defect leads to an increase in seed size. Additionally, overexpression of GEF1, potential FER-interacting partner, leads to a significant decrease in the seed size suggesting its potential role in GEF-ROP/ARAC-facilitated mechanosensing (Figure 13) and seed size regulation. Furthermore, in both cases, visibly significant difference of the transgenic/mutant line to WT was observed at the globular stage, indicating that the seed size regulation up to the globular stage is largely controlled by FER-regulated oi L1 (Yu et al., 2014). Later study by Creff et al. (2015) showed that this oi L1 layer cells exhibit less mechano-responsiveness to the endosperm turgor as compared to the oi L2 layer cells at later developmental stages.
FER is known to contribute to the elongation of hypocotyl by integrating brassinosteroid (BR) signaling (Deslauriers and Larsen, 2010). Furthermore, BR-signaling facilitates GA-mediated testa cell elongation in the developing seeds. BZR1, a BR signaling enzyme affecting the BR biosynthetic pathway in a negative feedback loop, interacts with a DELLA member GAI, which renders both of the interacting partners inactive (Figure 12) (Gallego-Bartolomé et al., 2012) thereby facilitating the GA-responsive gene expression and cell elongation. Furthermore, BR acts as a non-cell autonomous seed coat-to-endosperm signal and contributes to the positive regulation of the endosperm proliferation associated genes such as SHB1, IKU2, and MIN3 (Jiang et al., 2013, Lima et al., 2024, Pankaj et al., 2024a, Pankaj et al., 2024b). The active synthesis and signaling of BR in the maternal tissue (integuments/testa) has been suggested to potentiate the tissue to sustain endosperm proliferation (Figure 12) (Lima et al., 2024).
The BR-effect in seed size regulation is likely due to its role in counteracting the PRC2-mediated repressive effect in the tissue. In the pre-fertilized ovules, seed coat development is actively repressed by the PRC2-mediated H3K27me3 marks deposition in the integuments. As observed in Arabidopsis, BR function couples with the demethylation activity of the seed coat localized JUMONJI-type (JMJ) histone demethylases (ELF6 and REF6) and BR effectors (BES1 and BZR1) recruit the JMJ proteins to the target loci for demethylation (Pankaj et al., 2024b). The study postulated that BRI1, the BR-signaling associated protein involved in JMJs recruitment can facilitate the histone demethylation in a BR-independent manner unlike which, the process mediated by a BRI1 ortholog, BRL3 is BR-dependent (Figure 12). Studies indicate that the BR-mediated seed size regulation is likely mediated by microtubule reorganization (Abe et al., 2010, Jiang et al., 2013, Delesalle et al., 2025)
Unlike auxin, GA, and BR, ABA is discussed here as a potentially growth-restrictive input that may limit endosperm cavity expansion. Non-seed-associated studies indicate that the GAMYB members contribute to ABA signaling by promoting the accumulation of ABI5 transcript, a key ABA-signaling associated gene (Guo et al., 2021, Wyrzykowska et al., 2022). Earlier study additionally showed that ABA itself promotes miR159 accumulation in ABI3-dependent manner leading to GAMYB silencing (Figure 12) (Reyes and Chua, 2007). As observed in a maize study, its zma-miR159-target GAMYB members ZmMYB74 and ZmMYB139, negatively affect seed size by enhancing the expression of key downstream genes ABI5, mir156, and CDK. Furthermore, knock-out of the GAMYB members or overexpression miR159 enhanced the grain size (Wang et al., 2023).
5.5. Endosperm Cellularization
Endosperm cellularization typically commences after 10th consecutive nuclear division in Arabidopsis endosperm (Ali et al., 2023). The PRC2-mediated drop in endosperm turgor pressure contributes to its proliferation halt followed by subsequent cellularization (Weinhofer et al., 2010) via their putative downstream clustered ARFs (cARFs) (Figure 12) (Moreno-Romero et al., 2016, Butel et al., 2024).
The endosperm cellularization process is positively correlated with the activation of the genes associated with the ABA biosynthesis and signaling (Cheng et al., 2014). However, its potential role in the endosperm turgor pressure reduction has remained unexplored. Nevertheless, a parallel can be made with the operation of stomata, which is typically regulated by the guard cell turgor pressure. ABA reportedly increases the cytosolic Ca2+ leading to activation of slow anion (S-type) channel and downregulation of K+ influx channel in the guard cells thereby reducing its turgor and stomatal closure (Murata et al., 2001).
Typically, cellularization initiates at the micropylar region and progresses towards the chalaza in a wave fashion (Figure 10) (Hehenberger et al., 2012, Li et al., 2022). The process also incorporates the cues from the chalazal region bridged by ABA signaling. In brief, TFL1 - a PEBP member exclusively synthesized at the chalazal endosperm - is normally transported into the peripheral endosperm mediated by the small soluble GTP-binding RAN proteins, where it contributes to ABA signaling by stabilizing ABI5 (Zhang et al., 2020). ABI5 suppresses the accumulation of a key gene involved in endosperm proliferation, SHB1, by binding to the ABRE element in its promoter in ABA-dependent manner (Cheng et al., 2014). In alternate case, SHB1 typically binds to the W-box within the promoters of a WRKY transcription factor MIN3 and LRR protein family member IKU2 facilitated by MIN3 itself thereby promoting endosperm proliferation and the seed cavity volume increase (Kang et al., 2013). Defect in ABA biosynthesis and signaling leads to de-repression of the SHB1-MIN3-IKU2 pathway leading to an increase in seed size (Figure 12) (Cheng et al., 2014).
The SHB1-MIN3-IKU2 pathway is also negatively regulated by FIS-PRC2 in AGL9/15 dependent manner (Figure 12) in such a way that AGL9/15 binds to the CArG motifs located in the IKU and MIN3 promoter and contribute to recruiting FIS-PRC2 by interacting with MEA for the H3K27me3 marks deposition. Defect in AGL9 and AGL15 leads to increased seed cavity and seed mass (Zhang et al., 2024). Studies show that AGL15 is activated by auxin and it perturbs auxin signaling while contributing to somatic embryogenesis (Zheng et al., 2016). Since defect in either of AGL9/AGL15 and TFL1 leads to increase in seed cavity and mass, whether both of them act on the same pathway during endosperm cellularization yet remains unclear.
5.6. Embryo Development
While the endosperm proliferation commences almost immediately after karyogamy, zygotic division initiates several hours after fertilization (~10 to 20+ hrs depending on species). Immediately after the egg cell-to-sperm cell fusion, the zygote contracts briefly before returning to its initial state as suggested by an in vitro observation made in maize (Antoine et al., 2000). Some isolated-gamete or in vitro fertilization assays employ non-physiological medium conditions to enable handling and fusion (Faure et al., 1994, Kranz and Lörz, 1994). In intact/semi-in vivo double fertilization, fertilization triggers egg-cell Ca²⁺ transients/spikes, supporting Ca²⁺ as an activation signal under near-physiological tissue context (Figure 14) (Antoine et al., 2000, Denninger et al., 2014, Hamamura et al., 2014). The influx contributes to the cell wall formation around the freshly formed zygote (Denninger et al., 2014). The fertilized egg cell secretes, the aspartic endopeptidases ECS1, and ECS2, upon its successful fertilization as observed in Arabidopsis. The peptides actively contribute to blocking polytubey primarily by destroying the pollen tube attraction-associated LURE1 peptides (Figure 14) (Yu et al., 2021).
As mentioned earlier, RBR1 is inactive in mature egg cell. Nevertheless, RBR1-regulated cell cycle arrest in the tissue has been put forth by some studies based on the presence of G1/S phase-associated replication factors TMPK1 and DPB2, putative E2F-regulated outputs of the RBR1/E2F/DP gate, after fertilization (Ronceret and Devic, 2008). However, initiation of the first zygotic division requires APC/C-dependent turnover of mitotic cyclin activity, indicating that proteolysis-driven cell-cycle licensing is essential during early zygote activation (Figure 9) (Wang et al., 2012, Guo et al., 2016). Hence, while studies indicate that APC/C likely regulates the fertilization-dependent gating and RBR1/E2F regulates the embryo patterning, additional evidence is required to address the ongoing disagreement about whether the Arabidopsis egg cell is arrested in a pre-replicative (G0/G1-like) state or in G2 (Simonini, 2025).
Early embryogenesis involves a maternal-to-zygotic transition (MZT), during which maternally supplied transcripts are progressively removed and de novo zygotic transcription increases. In Arabidopsis, transcriptome dynamics support a transition from an egg/early-zygote program to a globular-stage program, followed by stronger zygotic genome activation (ZGA) coincident with elongation and patterning (Figure 14) (Kao and Nodine, 2019, Zhao et al., 2019, Brantley and Di Talia, 2024). Although transcripts from both parental alleles are detected early, key specification events are largely governed by zygotic gene products (Nodine and Bartel, 2012). The zygotic transcription is initiated in both fertilized egg and central cells almost simultaneously, even though endosperm proliferation precedes embryonic divisions (Kao and Nodine, 2019).
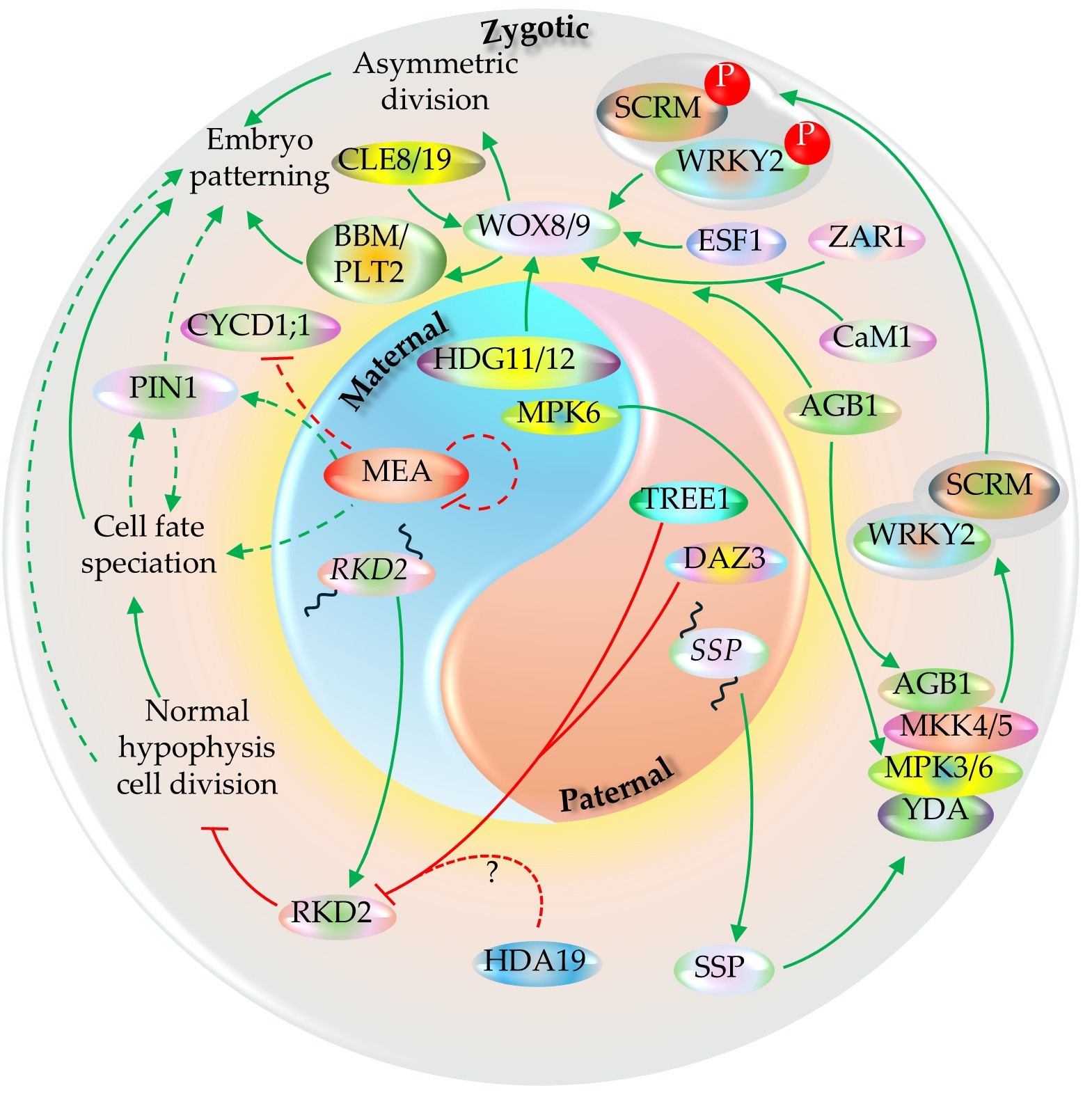
The paternal and maternal regulatory inputs establish zygote polarity and instruct the first asymmetric division - the earliest visible mark of embryo patterning. One of the paternally derived triggers is an RLCK-II family member, SHORT SUSPENSOR (SSP), mRNA of which is synthesized in the pollen but translation occurs in the zygote after fertilization. SSP subsequently potentiates a zygote MAPK cascade comprising the MAPKKK YODA (YDA), the MAPKKs MKK4/MKK5, and the MAPKs MPK3/MPK6, which together promote zygote elongation and asymmetric division {Bayer, 2009 #2421;Ueda, 2017 #2423;Lukowitz, 2004 #2444;Wang, 2007 #2435;Zhang, 2017 #2434}. Normal embryo development requires functional maternal copy of at least MPK6 during the process (Zhang et al., 2017). An LRR-RLK member, ZAR1, after the activation of its kinase activity via its direct interaction with CaM1 and AGB1, is also required for WOX8 expression and asymmetric division of the zygote (Yu et al., 2016). The heterotrimeric G-protein β subunit AGB1 also interacts with YDA–MKK4/5–MPK3/6 components and functions as the scaffold for the complex {Yuan, 2017 #2470}. Additionally, the WOX8 activation is also dependent on the endosperm-derived maternally imprinted factor, ESF1 (Figure 15) (Costa et al., 2014). WOX8 activates the PLETHORA member BBM and other genes associated with embryogenesis (Zhao et al., 2024). BBM and its close homolog, PLT2, are known to induce parthenogenesis independently upon their ectopic expression (Figure 15) (Chen et al., 2022).
The activated YDA-MAPK phosphorylates WRKY2, which is required for repolarization of the zygote. It directly activates WOX8 transcription after fertilization, thereby linking fertilization to the earliest embryo axis programs (Ueda et al., 2017). These paternal (SSP-to-YDA) and maternal (HDG11/12-to-WOX8) pathways are integrated into a framework in which independent parental cues converge to ensure robust zygote elongation and asymmetric division thereby initiating reliable apical-basal axis (Figure 15) (Ueda et al., 2011, Ueda et al., 2017). After the very first division, the PIN1-mediated auxin transport module facilitates to stabilizing the axis formation and embryo regionalization and patterning (Figure 15) (Möller and Weijers, 2009). For a normal hypophysis development in the embryo, RKD2 needs to be repressed, which is normally active in the egg cell. The paternally derived one zinc finger domain and two repressive EAR motifs harboring proteins, DAZ3 and TREE1 redundantly facilitate the process by binding to a AGCTAAAG motif at RKD2 promoter (Cheng et al., 2024). A separate study on TREE1-mediated shoot growth repression proposed the involvement of TREE1 EAR motif on HDA19 recruitment thereby leading to the deacetylation-mediated repression of the target gene (Wang et al., 2020). While such case is also plausible on RKD2 regulation in the zygote (Figure 15), it requires direct evidence to confirm as such.
Studies indicate that the WOX8/9-mediated patterning of early-stage embryo is facilitated by CLAVATA3/ESR-RELATED (CLE) proteins. CLE19, typically expressed in the abaxial epidermal layer of the embryo can confer embryo development defect non-cell autonomously when its antagonistic counterpart, CLE19G6T ectopically expressed in the endosperm, with no significant defect in the endosperm itself (Xu et al., 2015). Defect in CLE8, normally expressed in both embryo and endosperm, confers development defect in embryo as well as endosperm (Fiume and Fletcher, 2012). The expression domain of WUS is regulated by accompanying CLE signals in WUS niches, including regulation of the WOX8 expression domain by CLE8 (Figure 15) (Laux et al., 1996, Jun et al., 2010, Fiume et al., 2017). CLE is known to modulate WUS gradient at the post-transcriptional and post-translational level (Plong et al., 2021). Whether the CLE8/19-mediated embryo development regulation is conferred by modulating WOX8 gradient in similar way remains to be unraveled.
At the chromatin level, fertilization is accompanied by broad epigenetic reconfiguration of parental genomes, including resetting of histone variant landscapes and chromatin states that limit transgenerational carryover of gametic epigenetic information. In Arabidopsis, live cell imaging of histone H3 variants in fertilization products and subsequent studies support active remodeling/resetting in the zygote as part of early embryonic reprogramming. Furthermore, the zygote and endosperm exhibit distinctly different patterns of parental H3 marks. While the paternal H3 marks are gradually diluted in the endosperm over several rounds of nuclear divisions, zygote exhibits replication independent H3 mark removal (before its first division) as observed in Arabidopsis (Ingouff et al., 2007). This indicates distinctly different PRC2-mediated regulation in the two tissues.
PRC2 component, MEA regulates PIN1 to bring changes to the auxin distribution in the developing embryo and facilitates patterning via the regulation of cell cycle component, CYCD1;1 (Figure 15) (Simonini et al., 2021, Simonini et al., 2024). Furthermore, it is autoregulated in a negative feedback loop (independent of the PRC2 complex); hence the heterozygous mutant (MEA/mea) ovules exhibit higher level of MEA expression than WT (Baroux et al., 2006). Additionally, as transgenic study showed, translation of its transcript is dynamically regulated and pathogen infection leads to higher MEA protein synthesis (Rani et al., 2025). However, the complete picture of MEA-PRC2 mode of action during MZT and ZGA phases of embryo development is not fully clear yet.
Unlike the fertilization-dependent sexual ovules, the apomictic ovules exhibit parthenocarpy (fertilization-independent seed development). A comparative assessment on the apomictic and sexual accessions of buffelgrass showed that the ovules of the former exhibits higher expression of the genes encoding calcium receptor protein and its effector, calcium/calmodulin-dependent protein kinases (Ke et al., 2021). However, since apomixis is a convergent trait with its features appearing independently at different lineages (Goeckeritz et al., 2024), it remains yet unclear how deterministic is the role of calcium receptor proteins in apomictic trait induction and persistence across species.
5.7. Embryo-Endosperm Growth Coordination
It has now been well accepted that molecular handshake between endosperm and embryo is essential for their timely progression in a developing seed. However, an Arabidopsis study with the developmental stage-specific marker genes observed that key developmental changes - endosperm cellularization, embryo growth, and embryo maturation - are not strictly correlated but primarily dependent on the developmental stage (O'Neill et al., 2019). Nevertheless, normal embryo and endosperm development is essential for a typical seed development. While egg cell fertilization alone might be sufficient to trigger the division of the unfertilized central cell, its further development requires successful double fertilization as the seed development does not progress beyond post-globular stage (Nowack et al., 2006). Similarly, the proliferation, cellularization, and developmental programmed cell death (dPCD) of the endosperm can progress in absence of successful zygote formation as well. The development of dicot seeds after the initiation of the endosperm degeneration, however, relies on the embryo growth as observed in Arabidopsis (Xiong et al., 2021).
Embryo growth is partly facilitated by LEC1, encoding a CCAAT-binding motif atypical subunit of NF-Y(B) transcription factor, which is normally expressed at both zygote and endosperm (Figure 11) (Lee et al., 2003, Song et al., 2025). It plays a positive role during gametic and somatic embryogenesis (SE) as well as in post-embryonic development (Zhao et al., 2016, Brand et al., 2019). It is expressed in the central cell (endosperm) immediately after fertilization, before exhibiting expression in the embryo. Furthermore, its endosperm expression is more pivotal than its expression within the embryo itself during embryogenesis. As observed in Arabidopsis, significant proportion (87%) of the LEC1-target-genes are uniquely active in embryos as compared to the endosperm at any developmental stage (Song et al., 2025) while the LEC1-targets vary in the developing seeds depending on the endosperm growth phase- proliferation, cellularization, or degeneration (Pelletier et al., 2017, Song et al., 2025). The endosperm-derived LEC1 enters embryo suspensor symplastically via plasmodesmata (Figure 16) and is detectable by the two-celled zygotic stage (Song et al., 2021). It promotes the expression of key maturity-associated genes including itself, LEC2, FUS3, and ABI3, which are essential for the seed storage protein accumulation during the later developmental stage (Song et al., 2021). The across-cell-movement of the typical nuclear localizing LEC1 protein suggests that it either constitutes yet unidentified sorting signal within it - similar to that suggested for the KNOTTED1 protein (Lucas et al., 1995) - or it is accompanied with yet unidentified signaling factor facilitating the transport. While LEC1-defect does not bring any pronounced defect during very early developmental stage, the mutant embryo exhibits desiccation intolerance and development halt at its linear cotyledonary stage (Lee et al., 2003).
Additional endosperm-dependent embryo feature is its cuticle development, which typically commences at the globular stage in Arabidopsis (Szczuka and Szczuka, 2003, De Giorgi et al., 2021). Embryo at globular stage and onwards expresses GSO1/2 encoding putative leucine-rich repeat transmembrane-type receptor kinase (Tsuwamoto et al., 2008), and TWS1, encoding an ER-localized peptide (Doll et al., 2020b). The TWS1 needs to be activated before it is secreted to the apoplast and the process is facilitated by tyrosyl-protein sulfotransferase (TPST) by sulfating (Tyr-sulfating) TWS1 in the golgi complex. After being secreted, the sulfated TWS1 at the apoplast is further cleaved at the close downstream of its sulfation site by an endosperm-derived subtilase family protease, ALE1. Then the N-terminal TWS1 fragment harboring the sulfated site interacts with the extracellular region of the embryo plasma membrane-bound GSO1/2 (Tanaka et al., 2001, Doll et al., 2020b). The TWS1-to-GSO1/2 binding confirms the cuticle absence/damage and triggers the repair machinery (Figure 16). In later stage, the intact cuticle prevents TWS1 leakage to the apoplastic region thereby bringing the repair process to a halt (Doll et al., 2020b).
GSO1/2 also contributes to later developmental stages by promoting formation of the ‘sheath’, a thin layer above the cuticle that supports non-sticky embryo growth during development and cotyledon emergence during germination (Moussu et al., 2017). This process involves deposition of the ZHOUPI (ZOU)-regulated cysteine-rich protein KERBEROS (KRS) (Figure 16) (Doll et al., 2020a). KRS expression initiates at the heart stage, peaks at the torpedo stage, and declines toward maturity in Arabidopsis. KRS shares sequence similarity with the tomato stigma-exudate protein STIG1, which functions as a ligand for the pollen-tube LRR receptor kinase LePRK2 and modulates pollen-tube redox potential during elongation (Tang et al., 2004, Huang et al., 2014). By analogy, KRS has been proposed to act as a ligand for an embryo-localized LRR receptor kinase to promote invasive embryo growth and ROS accumulation (Figure 16). However, direct biochemical evidence that KRS binds or activates GSO1/2 or other LRR receptor kinase is not yet available, which can be tested by genetic epistasis (krs × gso1/2), peptide rescue assays with synthetic KRS, and binding/activation measurements (SPR/ITC; receptor phosphorylation or reporter outputs) using GSO1/2 ± SERK coreceptors.
Along with its interacting bHLH counterpart, INDUCER OF CBF EXPRESSION1 (ICE1), ZOU affects the embryo surrounding region (ESR) (Denay et al., 2014). The perturbation of ZOU function affects the cell wall properties thereby increasing tissue-stiffness. Defect in either of ZOU and ICE1 hinders the expression of the dPCD-associated NAC TFs (KIR1, ANAC087, ANAC046, ORE1) and their downstream nucleic acid degradation associated genes (PASPA3 and BFN1) (Figure 16) leading to a halt to the embryo growth and shriveled seed development (Yang et al., 2008, Denay et al., 2014, Fourquin et al., 2016, Doll et al., 2023). The ZOU defect in Arabidopsis delays the expression the dPCD-associated genes by 4-5 days as compared to its wild type counterpart, which typically exhibits their expression by 8 DAP (Doll et al., 2023). The cumulative defect in KIR1 and additional ESR-expressed genes (anac046-1 anac087-1 kir1-1 ore1-1) leads to the persistence of uncleared corpse cells and halts the embryonic growth (Doll et al., 2023). Analogous observation in maize showed that its KIR homologs KIL1/2 play role on PCD and corpse cell residues clearance at the endosperm region adjacent to the growing embryo, referred to as embryo scutellum. Defects in those genes affect the PCD and corpse cell clearance processes thereby strongly hindering the embryo growth. Furthermore, their expression is regulated by the paternally imprinted endosperm-specific DOSAGE-EFFECT DEFECTIVE1 (DED1), the closest maize homolog of Arabidopsis MYB64/119 (Dai et al., 2022, Doll et al., 2025).
The endosperm dPCD at the ESR, however, rely on the normal embryo development. The halt in post-globular stage embryonic growth led by the defect in DEK1, a gene encoding an integral membrane protein with a calpain protease domain, brings the normal endosperm development with no dPCD, even though the ZOU-dependent cell wall thinning and expression of pre-cell death markers remains normal in the dek1 mutant (Johnson et al., 2005, Fourquin et al., 2016). Normal DEK1 expression, however, is indispensable for its L1 layer development in Arabidopsis (Johnson et al., 2005). The gene is attributed for its role in the endosperm aleurone cell (layer) development (Lid et al., 2002). It strongly suggests that embryo L1 layer functions as the harbinger of the molecular factors more directly involved in embryo-endosperm communication at their interface. Such postulation also aligns with the globular stage growth arrest observed in the L1 layer development defective Arabidopsis dual mutant pdf2; atml1 (Ogawa et al., 2015). While the endosperm dPCD is essential for monocot seed development as well (Domínguez and Cejudo, 2014), a study showed that defect in two FERONIA homologs FERONIA-LIKE RECEPTOR 3 (FER3) and FER14 leads to a surge in ROS and upregulation of the PCD associated genes and caspases. Furthermore, the mutant endosperm exhibited disruption in starch biosynthesis, and decrease in seed storage protein accumulation (He et al., 2023) suggesting potential role of FERONIA-like CrRLK1l members to perceive the pressure exerted by the growing embryo against endosperm and translating it to the controlled execution of PCD of the endosperm cells (Figure 16). Future monocots and dicots studies are expected to shed more light on the case
6. Bottlenecks and Future Perspectives
Despite rapid progress, seed development remains a patchwork of tissue-specific insights that are still difficult to unify into a predictive, cross-species framework. Below are key bottlenecks prevailing in the field.
6.1. Correlative vs Causative Evidence Behind MMC Speciation
Many proposed MMC speciation regulators (auxin threshold, SPL/NZZ-associated networks, small RNAs, chromatin states, and mechanics) are supported by strong correlations. Accompanying direct causal chains of evidence in a narrow spatial and temporal windows would benefit the case. Potential such avenues include (i) True spatiotemporal perturbation (cell-type specific, inducible, reversible): e.g., transient alteration of auxin transport or chromatin regulators specifically in L2 neighbors versus the enlarging candidate MMC, then measuring reversibility and fate stability. (ii) Integration of mechanics with gene regulation: primordium geometry, cell wall mechanics, and turgor-driven growth likely contribute to which L2 cell becomes MMC-competent. Study linking the mechanical changes to transcriptional/epigenetic switching would shed more light on the matter. Additionally, the coenocytic phase of endosperm development exhibit strong positive link with the microtubule and F-actin polymerization (Ali et al., 2023). However, potential factors associated with the process in the endosperm is yet to be identified. Study on the cytoskeleton dynamics machineries with reference to the studies on other turgor-influenced tissues such as pollen tube (Adhikari et al., 2020a), and stomata is expected to shed more light on the topic.
iku2 mutant exhibits increased testa stiffness in the developing seed (Hamann, 2012, Creff et al., 2023). IKU2 is a member leucine rich repeat (LRR) kinase 2 protein. A study on animal model (mouse fibroblast cell) showed that its LRR kinase 2 member, Lrrk2, modulates the intracellular cytoskeletal dynamics. The protein exhibits affinity to F-actin and its defect increases the cell’s perimeter-to-area ratio (Meixner et al., 2011). Furthermore, cellular auxin level is known to contributes to its actin dynamics in plant (Figure 12) (Adhikari et al., 2020a). As observed in an Arabidopsis study, degradation of either microtubules or F-actin leads to a significant decrease in the coenocytic endosperm area and exhibits a visibly elongated size (Ali et al., 2023). Whether IKU2 defect confers cytoskeleton (F-actin in particular) dysregulation leading to the development of misshaped endosperm thereby affecting its turgor remains to be determined.
Regarding the ELA-controlled GA-regulation and testa stiffening, while GA biosynthesis is suppressed via the ELA1/2-mediated mechanosensing, their ER-localization (Zhang et al., 2011) strongly suggests for upstream mechanosensory factors at play. In addition, ER is known to ‘sense’ the external mechanical stress based on Ca2+ homeostasis changes in its lumen potentially activated by the stress-triggered Ca2+ influx-led imbalance in cytoplasmic Ca2+ level (Howell, 2013, Yoshimura et al., 2021). Mechanical strain that alters cell-wall/pectin status can be perceived by FERONIA (FER) and homologs at the cell surface, engaging downstream Ca²⁺-channel activity to shape cytosolic Ca²⁺ signaling. This FER-dependent Ca²⁺ signaling can operate alongside tension-gated mechanosensitive channels to form the overall Ca²⁺ “mechano-signature” (Figure 13) (Feng et al., 2018). Furthermore, Ca2+ is linked to GA biosynthesis and signaling (Du et al., 2023, Gai et al., 2024). It can rapidly elevate cytosolic Ca²⁺ in Arabidopsis via a pathway that is independent of DELLA degradation, indicating that GA signaling can interface directly with Ca²⁺ dynamics {Okada, 2017 #2467}. However, direct evidence linking FER-dependent Ca²⁺ signaling to ELA1/2-mediated GA deactivation in the seed coat is not yet available. In parallel, membrane tension–gated mechanosensitive Ca²⁺-permeable channels (e.g., MCA-family, and other plant MS channel families such as MSL/OSCA/PIEZO) could contribute independently or alongside FER to shape the Ca²⁺ ‘mechano-signature’ that precedes downstream hormonal responses {Yoshimura, 2021 #2406;Guichard, 2022 #2469}.
6.2. Sporophyte-Gametophyte Cross-Talk Conduits
Multiple lines of evidence support sporophyte-gametophyte and embryo-endosperm cross-talk. However, the identity of mobile signals, their transport distances, and their routes (symplastic, apoplastic, or uptake-mediated) remain incompletely defined. Priority directions include: (i) identifying mobile molecules and directionality (small RNAs, peptides, hormones, metabolites) using lineage-restricted production and receiver-specific reporters; (ii) quantifying timing-dependent connectivity changes around fertilization as a measurable developmental variable; and (iii) testing conservation across taxa (eudicots vs monocots; persistent vs transient connectivity patterns).
6.3. Translational Perspective
Despite substantial progress on the molecular and mechanical basis of seed development, comparatively fewer studies translate these advances into crop improvement pipelines. Prioritizing translational frameworks could clarify cross-species conservation of regulatory modules while accelerating delivery of validated targets to breeding programs. Institutionally supported multi-team collaborations that integrate genetics, phenomics, and seed biology are expected to facilitate such efforts.
7. Conclusion
The earliest phase of seed development is governed by coordinated hormonal, positional, and genetic/epigenetic regulators that enforce pre-fertilization developmental blocks and then enable rapid post-fertilization reprogramming. MMC restriction, ovule maturation, and fertilization-dependent activation of the coenocytic endosperm are coupled to maternal responses that initiate and pattern the seed coat, with auxin functioning as a central inter-tissue coordinator. Polycomb-associated chromatin states, small-RNA-linked silencing, and RBR1/APC-mediated cell-cycle gates provide robustness against ectopic proliferation, yet are selectively remodeled after fertilization to support parent-of-origin programs in endosperm and embryo. In parallel, mechanical forces generated by endosperm expansion and their perception by maternal tissues emerge as key determinants of seed growth and the timing of cellularization. Outstanding challenges include resolving causal signal flow across tissue interfaces (hormones, peptides, small RNAs, metabolites) and integrating mechanics with transcriptional and chromatin state changes at cell-type resolution.
Author Contributions
P.B.A. conceptualized and drafted the manuscript and associated figures; R.D.K. assisted during the editing process.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Abe, Y.; Mieda, K.; Ando, T.; Kono, I.; Yano, M.; Kitano, H.; Iwasaki, Y. The SMALL AND ROUND SEED1 (SRS1/DEP2) gene is involved in the regulation of seed size in rice. Genes & genetic systems 2010, 85, 327–339. [Google Scholar]
- Adamski, N.M.; Anastasiou, E.; Eriksson, S.; O'Neill, C.M.; Lenhard, M. Local maternal control of seed size by KLUH/ CYP78A5-dependent growth signaling. Proceedings of the National Academy of Sciences 2009, 106, 20115–20120. [Google Scholar] [CrossRef]
- Adhikari, P.B.; Liu, X.; Kasahara, R.D. Mechanics of pollen tube elongation: A perspective. Frontiers in Plant Science 2020a, 11, 589712. [Google Scholar] [CrossRef]
- Adhikari, P.B.; Liu, X.; Wu, X.; Zhu, S.; Kasahara, R.D. Fertilization in flowering plants: an odyssey of sperm cell delivery. Plant Molecular Biology 2020b, 103, 9–32. [Google Scholar] [CrossRef]
- Alexiadis, V.; Waldmann, T.; Andersen, J.; Mann, M.; Knippers, R.; Gruss, C. The protein encoded by the proto-oncogene DEK changes the topology of chromatin and reduces the efficiency of DNA replication in a chromatin-specific manner. Genes Dev 2000, 14, 1308–1312. [Google Scholar] [CrossRef]
- Ali, M.F.; Shin, J.M.; Fatema, U.; Kurihara, D.; Berger, F.; Yuan, L.; Kawashima, T. Cellular dynamics of coenocytic endosperm development in Arabidopsis thaliana. Nature Plants 2023, 9, 330–342. [Google Scholar] [CrossRef]
- Aloni, R.; Aloni, E.; Langhans, M.; Ullrich, C.I. Role of auxin in regulating Arabidopsis flower development. Planta 2006, 223, 315–328. [Google Scholar] [CrossRef]
- Antoine, A.F.; Faure, J.E.; Cordeiro, S.; Dumas, C.; Rougier, M.; Feijó, J.A. A calcium influx is triggered and propagates in the zygote as a wavefront during in vitro fertilization of flowering plants. Proceedings of the National Academy of Sciences 2000, 97, 10643–10648. [Google Scholar] [CrossRef]
- Arnault, G.; Vialette, A.C.M.; Andres-Robin, A.; Fogliani, B.; Gâteblé, G.; Scutt, C.P. Evidence for the extensive conservation of mechanisms of ovule integument development since the most recent common ancestor of living angiosperms. Frontiers in Plant Science 2018, 9–2018. [Google Scholar] [CrossRef]
- Aslam, M.; Fakher, B.; Jakada, B.H.; Cao, S.; Qin, Y. SWR1 chromatin remodeling complex: A key transcriptional regulator in plants. Cells 2019, 8. [Google Scholar] [CrossRef]
- Baroux, C.; Gagliardini, V.; Page, D.R.; Grossniklaus, U. Dynamic regulatory interactions of Polycomb group genes: MEDEA autoregulation is required for imprinted gene expression in Arabidopsis. Genes & Development 2006, 20, 1081–1086. [Google Scholar] [CrossRef]
- Baroux, C.; Grossniklaus, U. Seeds—An evolutionary innovation underlying reproductive success in flowering plants. In Current Topics in Developmental Biology; Grossniklaus, U., Ed.; Academic Press, 2019; pp. 605–642. [Google Scholar]
- Baroux, C.; Spillane, C.; Grossniklaus, U. Evolutionary origins of the endosperm in flowering plants. Genome Biology 2002, 3, reviews1026.1021. [Google Scholar] [CrossRef]
- Batista, R.A.; Moreno-Romero, J.; Qiu, Y.; van Boven, J.; Santos-González, J.; Figueiredo, D.D.; Köhler, C. The MADS-box transcription factor PHERES1 controls imprinting in the endosperm by binding to domesticated transposons. eLife 2019, 8, e50541. [Google Scholar] [CrossRef]
- Bauer, M.J.; Fischer, R.L. Genome demethylation and imprinting in the endosperm. Current Opinion in Plant Biology 2011, 14, 162–167. [Google Scholar] [CrossRef]
- Bayer, M.; Nawy, T.; Giglione, C.; Galli, M.; Meinnel, T.; Lukowitz, W. Paternal control of embryonic patterning in Arabidopsis thaliana. Science 2009, 323, 1485–1488. [Google Scholar] [CrossRef]
- Beauzamy, L.; Fourquin, C.; Dubrulle, N.; Boursiac, Y.; Boudaoud, A.; Ingram, G.C. Endosperm turgor pressure decreases during early Arabidopsis seed development. Development 2016, 143, 3295–3299. [Google Scholar] [CrossRef]
- Becker, A.; Chen, X.; Dresselhaus, T.; Gutsche, N.; Müller-Schüssele, S.J.; Sprunck, S.; Theißen, G.; de Vries, S.; Zachgo, S. Sexual reproduction in land plants: an evolutionary perspective. Plant Reproduction 2025, 38, 12. [Google Scholar] [CrossRef]
- Belmonte, M.F.; Kirkbride, R.C.; Stone, S.L.; Pelletier, J.M.; Bui, A.Q.; Yeung, E.C.; Hashimoto, M.; Fei, J.; Harada, C.M.; Munoz, M.D.; Le, B.H.; Drews, G.N.; Brady, S.M.; Goldberg, R.B.; Harada, J.J. Comprehensive developmental profiles of gene activity in regions and subregions of the Arabidopsis seed. Proceedings of the National Academy of Sciences 2013, 110, E435–E444. [Google Scholar] [CrossRef]
- Bencivenga, S.; Simonini, S.; Benková, E.; Colombo, L. The transcription factors BEL1 and SPL are required for cytokinin and auxin signaling during ovule development in Arabidopsis. The Plant Cell 2012, 24, 2886–2897. [Google Scholar] [CrossRef]
- Benton, M.J.; Wilf, P.; Sauquet, H. The angiosperm terrestrial revolution and the origins of modern biodiversity. New Phytologist 2022, 233, 2017–2035. [Google Scholar] [CrossRef]
- Berger, F. Meiosis as a mechanism for epigenetic reprogramming and cellular rejuvenation. Development 2024, 151. [Google Scholar] [CrossRef]
- Boyko, J.D.; Vasconcelos, T. Trait lability as a predictor of diversification dynamics in flowering plants. bioRxiv. 2025. 2024.2006.2003.597046.
- Brand, A.; Quimbaya, M.; Tohme, J.; Chavarriaga-Aguirre, P. Arabidopsis LEC1 and LEC2 orthologous genes are key regulators of somatic embryogenesis in cassava. Frontiers in Plant Science 2019, 10–2019. [Google Scholar] [CrossRef]
- Brantley, S.; Di Talia, S. The maternal-to-zygotic transition. Current Biology 2024, 34, R519–R523. [Google Scholar] [CrossRef]
- Butel, N.; Qiu, Y.; Xu, W.; Santos-González, J.; Köhler, C. Parental conflict driven regulation of endosperm cellularization by a family of Auxin Response Factors. Nature Plants 2024, 10, 1018–1026. [Google Scholar] [CrossRef]
- Cavalleri, A.; Astori, C.; Manrique, S.; Bruzzaniti, G.; Smaczniak, C.; Mizzotti, C.; Ruiu, A.; Spano, M.; Movilli, A.; Gregis, V.; Xu, X.; Kaufmann, K.; Colombo, L. (2025) SPOROCYTELESS/NOZZLE acts together with MADS-domain transcription factors to regulate an auxin-dependent network controlling the Megaspore Mother Cell development. bioRxiv 2025.2003, 2011.641985. [Google Scholar]
- Chen, B.; Maas, L.; Figueiredo, D.; Zhong, Y.; Reis, R.; Li, M.; Horstman, A.; Riksen, T.; Weemen, M.; Liu, H.; Siemons, C.; Chen, S.; Angenent, G.C.; Boutilier, K. BABY BOOM regulates early embryo and endosperm development. Proceedings of the National Academy of Sciences 2022, 119, e2201761119. [Google Scholar] [CrossRef]
- Chen, D.; Molitor, A.M.; Xu, L.; Shen, W.-H. Arabidopsis PRC1 core component AtRING1 regulates stem cell-determining carpel development mainly through repression of class I KNOX genes. BMC Biology 2016, 14, 112. [Google Scholar] [CrossRef]
- Cheng, T.; Liu, Z.; Li, H.; Huang, X.; Wang, W.; Shi, C.; Zhang, X.; Chen, H.; Yao, Z.; Zhao, P.; Peng, X.; Sun, M.-X. Sperm-origin paternal effects on root stem cell niche differentiation. Nature 2024, 634, 220–227. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Zhao, X.Y.; Shao, X.X.; Wang, F.; Zhou, C.; Liu, Y.G.; Zhang, Y.; Zhang, X.S. Abscisic acid regulates early seed development in Arabidopsis by ABI5-mediated transcription of SHORT HYPOCOTYL UNDER BLUE1. The Plant Cell 2014, 26, 1053–1068. [Google Scholar] [CrossRef]
- Choi, Y.; Gehring, M.; Johnson, L.; Hannon, M.; Harada, J.J.; Goldberg, R.B.; Jacobsen, S.E.; Fischer, R.L. DEMETER, a DNA glycosylase domain protein, is required for endosperm gene imprinting and seed viability in Arabidopsis. Cell 2002, 110, 33–42. [Google Scholar] [CrossRef]
- Conner, J.A.; Mookkan, M.; Huo, H.; Chae, K.; Ozias-Akins, P. A parthenogenesis gene of apomict origin elicits embryo formation from unfertilized eggs in a sexual plant. Proceedings of the National Academy of Sciences 2015, 112, 11205–11210. [Google Scholar] [CrossRef]
- Costa, L.M.; Marshall, E.; Tesfaye, M.; Silverstein, K.A.T.; Mori, M.; Umetsu, Y.; Otterbach, S.L.; Papareddy, R.; Dickinson, H.G.; Boutiller, K.; VandenBosch, K.A.; Ohki, S.; Gutierrez-Marcos, J.F. Central cell–derived peptides regulate early embryo patterning in flowering plants. Science 2014, 344, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Creff, A.; Ali, O.; Bied, C.; Bayle, V.; Ingram, G.; Landrein, B. Evidence that endosperm turgor pressure both promotes and restricts seed growth and size. Nature Communications 2023, 14, 67. [Google Scholar] [CrossRef]
- Creff, A.; Brocard, L.; Ingram, G. A mechanically sensitive cell layer regulates the physical properties of the Arabidopsis seed coat. Nature Communications 2015, 6, 6382. [Google Scholar] [CrossRef]
- Dai, D.; Mudunkothge, J.S.; Galli, M.; Char, S.N.; Davenport, R.; Zhou, X.; Gustin, J.L.; Spielbauer, G.; Zhang, J.; Barbazuk, W.B.; Yang, B.; Gallavotti, A.; Settles, A.M. Paternal imprinting of dosage-effect defective1 contributes to seed weight xenia in maize. Nature Communications 2022, 13, 5366. [Google Scholar] [CrossRef]
- De Giorgi, J.; Fuchs, C.; Iwasaki, M.; Kim, W.; Piskurewicz, U.; Gully, K.; Utz-Pugin, A.; Mene-Saffrane, L.; Waridel, P.; Nawrath, C. The Arabidopsis mature endosperm promotes seedling cuticle formation via release of sulfated peptides. Developmental cell 2021, 56, 3066–3081. e3065. [Google Scholar] [CrossRef]
- Debeaujon, I.; Nesi, N.; Perez, P.; Devic, M.; Grandjean, O.; Caboche, M.; Lepiniec, L. Proanthocyanidin-accumulating cells in Arabidopsis testa: regulation of differentiation and role in seed development. Plant Cell 2003, 15, 2514–2531. [Google Scholar] [CrossRef]
- Delesalle, C.; Montiel-Jorda, A.; Aiba, R.; Spielmann, J.; Neveu, J.; Fujita, S.; Vert, G. Arabidopsis microtubule-BRI1-associated proteins negatively regulate hypocotyl elongation by controlling brassinosteroid-dependent cortical microtubule reorientation. Plant Communications 2025, 101637. [Google Scholar] [CrossRef]
- Denay, G.; Creff, A.; Moussu, S.; Wagnon, P.; Thévenin, J.; Gérentes, M.-F.; Chambrier, P.; Dubreucq, B.; Ingram, G. Endosperm breakdown in Arabidopsis requires heterodimers of the basic helix-loop-helix proteins ZHOUPI and INDUCER OF CBP EXPRESSION 1. Development 2014, 141, 1222–1227. [Google Scholar] [CrossRef]
- Denninger, P.; Bleckmann, A.; Lausser, A.; Vogler, F.; Ott, T.; Ehrhardt, D.W.; Frommer, W.B.; Sprunck, S.; Dresselhaus, T.; Grossmann, G. Male-female communication triggers calcium signatures during fertilization in Arabidopsis. Nat Commun 2014, 5, 4645. [Google Scholar] [CrossRef]
- Deslauriers, S.D.; Larsen, P.B. FERONIA is a key modulator of brassinosteroid and ethylene responsiveness in Arabidopsis hypocotyls. Molecular Plant 2010, 3, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Doll, N.M.; Bovio, S.; Gaiti, A.; Marsollier, A.-C.; Chamot, S.; Moussu, S.; Widiez, T.; Ingram, G. The endosperm-derived embryo sheath is an anti-adhesive structure that facilitates cotyledon emergence during germination in Arabidopsis. Current Biology 2020a, 30, 909–915.e904. [Google Scholar] [CrossRef]
- Doll, N.M.; Fierlej, Y.; Eekhout, T.; Elias, L.; Bellot, C.; Sun, G.; Grones, C.; Aesaert, S.; Coussens, G.; De Rycke, R.; Šimášková, M.; Montes, E.; Plagnard, C.; Rogowsky, P.M.; Abebaw, Y.M.; Bendahmane, M.; De Rybel, B.; Pauwels, L.; Widiez, T.; Nowack, M.K. KIL transcription factors facilitate embryo growth in maize by promoting endosperm elimination via lytic cell death. The Plant Cell 2025, 37. [Google Scholar] [CrossRef]
- Doll, N.M.; Royek, S.; Fujita, S.; Okuda, S.; Chamot, S.; Stintzi, A.; Widiez, T.; Hothorn, M.; Schaller, A.; Geldner, N.; Ingram, G. A two-way molecular dialogue between embryo and endosperm is required for seed development. Science 2020b, 367, 431–435. [Google Scholar] [CrossRef]
- Doll, N.M.; Van Hautegem, T.; Schilling, N.; De Rycke, R.; De Winter, F.; Fendrych, M.; Nowack, M.K. Endosperm cell death promoted by NAC transcription factors facilitates embryo invasion in Arabidopsis. Current Biology 2023, 33, 3785–3795.e3786. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, F.; Cejudo, F.J. Programmed cell death (PCD): an essential process of cereal seed development and germination. Frontiers in Plant Science 2014, 5–2014. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Zhao, Y.; Xiao, C.; Ren, D.; Ding, Y.; Xu, J.; Jin, H.; Jiao, H. Mechanism analysis of calcium nitrate application to induce gibberellin biosynthesis and signal transduction promoting stem elongation of Dendrobium officinale. Industrial Crops and Products 2023, 195, 116495. [Google Scholar] [CrossRef]
- Duan, Q.; Kita, D.; Li, C.; Cheung, A.Y.; Wu, H.-M. FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair development. Proceedings of the National Academy of Sciences 2010, 107, 17821–17826. [Google Scholar] [CrossRef]
- Ebel, C.; Mariconti, L.; Gruissem, W. Plant retinoblastoma homologues control nuclear proliferation in the female gametophyte. Nature 2004, 429, 776–780. [Google Scholar] [CrossRef]
- Faure, J.E.; Digonnet, C.; Dumas, C. An in vitro system for adhesion and fusion of maize gametes. Science 1994, 263, 1598–1600. [Google Scholar] [CrossRef]
- Feng, W.; Kita, D.; Peaucelle, A.; Cartwright, H.N.; Doan, V.; Duan, Q.; Liu, M.-C.; Maman, J.; Steinhorst, L.; Schmitz-Thom, I.; Yvon, R.; Kudla, J.; Wu, H.-M.; Cheung, A.Y.; Dinneny, J.R. The FERONIA receptor kinase maintains cell-wall integrity during salt stress through Ca2+ signaling. Current Biology 2018, 28, 666–675.e665. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, D.D.; Batista, R.A.; Roszak, P.J.; Hennig, L.; Köhler, C. Auxin production in the endosperm drives seed coat development in Arabidopsis. eLife 2016, 5, e20542. [Google Scholar] [CrossRef]
- Figueiredo, D.D.; Batista, R.A.; Roszak, P.J.; Köhler, C. Auxin production couples endosperm development to fertilization. Nature Plants 2015, 1, 15184. [Google Scholar] [CrossRef]
- Figueiredo, D.D.; Köhler, C. Auxin: a molecular trigger of seed development. Genes & Development 2018, 32, 479–490. [Google Scholar] [CrossRef]
- Fiume, E.; Coen, O.; Xu, W.; Lepiniec, L.; Magnani, E. Developmental patterning of sub-epidermal cells in the outer integument of Arabidopsis seeds. PLOS ONE 2017, 12, e0188148. [Google Scholar] [CrossRef]
- Fiume, E.; Fletcher, J.C. Regulation of Arabidopsis embryo and endosperm development by the polypeptide signaling molecule CLE8. Plant Cell 2012, 24, 1000–1012. [Google Scholar] [CrossRef]
- Floyd, Sandra K.; Friedman, William E. Evolution of endosperm developmental patterns among basal flowering plants. International Journal of Plant Sciences 2000, 161, S57–S81. [Google Scholar] [CrossRef]
- Fourquin, C.; Beauzamy, L.; Chamot, S.; Creff, A.; Goodrich, J.; Boudaoud, A.; Ingram, G. Mechanical stress mediated by both endosperm softening and embryo growth underlies endosperm elimination in Arabidopsis seeds. Development 2016, 143, 3300–3305. [Google Scholar] [CrossRef]
- Gai, W.; Liu, C.; Yang, M.; Li, F.; Xin, H.; Gai, S. Calcium signaling facilitates chilling- and GA- induced dormancy release in tree peony. Frontiers in Plant Science 2024, 15–2024. [Google Scholar] [CrossRef]
- Gallego-Bartolomé, J.; Minguet, E.G.; Grau-Enguix, F.; Abbas, M.; Locascio, A.; Thomas, S.G.; Alabadí, D.; Blázquez, M.A. Molecular mechanism for the interaction between gibberellin and brassinosteroid signaling pathways in Arabidopsis. Proceedings of the National Academy of Sciences 2012, 109, 13446–13451. [Google Scholar] [CrossRef] [PubMed]
- Gehring, M.; Bubb, K.L.; Henikoff, S. Extensive demethylation of repetitive elements during seed development underlies gene imprinting. Science 2009, 324, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Gehring, M.; Huh, J.H.; Hsieh, T.-F.; Penterman, J.; Choi, Y.; Harada, J.J.; Goldberg, R.B.; Fischer, R.L. DEMETER DNA glycosylase establishes MEDEA polycomb gene self-imprinting by allele-specific demethylation. Cell 2006, 124, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Goeckeritz, C.Z.; Zheng, X.; Harkess, A.; Dresselhaus, T. Widespread application of apomixis in agriculture requires further study of natural apomicts. iScience 2024, 27, 110720. [Google Scholar] [CrossRef] [PubMed]
- Gombos, M.; Raynaud, C.; Nomoto, Y.; Molnár, E.; Brik-Chaouche, R.; Takatsuka, H.; Zaki, A.; Bernula, D.; Latrasse, D.; Mineta, K.; Nagy, F.; He, X.; Iwakawa, H.; Őszi, E.; An, J.; Suzuki, T.; Papdi, C.; Bergis, C.; Benhamed, M.; Bögre, L.; Ito, M.; Magyar, Z. The canonical E2Fs together with RETINOBLASTOMA-RELATED are required to establish quiescence during plant development. Commun Biol 2023, 6, 903. [Google Scholar] [CrossRef]
- Guitton, A.-E.; Berger, F. Loss of function of MULTICOPY SUPPRESSOR OF IRA 1 produces nonviable parthenogenetic embryos in Arabidopsis. Current Biology 2005, 15, 750–754. [Google Scholar] [CrossRef]
- Guo, C.; Jiang, Y.; Shi, M.; Wu, X.; Wu, G. ABI5 acts downstream of miR159 to delay vegetative phase change in Arabidopsis. New Phytologist 2021, 231, 339–350. [Google Scholar] [CrossRef]
- Guo, L.; Jiang, L.; Zhang, Y.; Lu, X.-L.; Xie, Q.; Weijers, D.; Liu, C.-M. The anaphase-promoting complex initiates zygote division in Arabidopsis through degradation of cyclin B1. The Plant Journal 2016, 86, 161–174. [Google Scholar] [CrossRef]
- Guo, L.; Luo, X.; Li, M.; Joldersma, D.; Plunkert, M.; Liu, Z. Mechanism of fertilization-induced auxin synthesis in the endosperm for seed and fruit development. Nature Communications 2022, 13, 3985. [Google Scholar] [CrossRef]
- Hamamura, Y.; Nishimaki, M.; Takeuchi, H.; Geitmann, A.; Kurihara, D.; Higashiyama, T. Live imaging of calcium spikes during double fertilization in Arabidopsis. Nature Communications 2014, 5, 4722. [Google Scholar] [CrossRef]
- Hamann, T. Plant cell wall integrity maintenance as an essential component of biotic stress response mechanisms. Frontiers in plant science 2012, 3, 77. [Google Scholar] [CrossRef]
- He, W.; Li, W.; Luo, X.; Tang, Y.; Wang, L.; Yu, F.; Lin, Q. Rice FERONIA-LIKE RECEPTOR 3 and 14 affect grain quality by regulating redox homeostasis during endosperm development. Journal of Experimental Botany 2023, 74, 3003–3018. [Google Scholar] [CrossRef]
- Hehenberger, E.; Kradolfer, D.; Köhler, C. Endosperm cellularization defines an important developmental transition for embryo development. Development 2012, 139, 2031–2039. [Google Scholar] [CrossRef]
- Heidemann, B.; Primetis, E.; Zahn, I.E.; Underwood, C.J. To infinity and beyond: recent progress, bottlenecks, and potential of clonal seeds by apomixis. Plant J 2025, 121, e70054. [Google Scholar] [CrossRef]
- Honma, Y.; Adhikari, P.B.; Kuwata, K.; Kagenishi, T.; Yokawa, K.; Notaguchi, M.; Kurotani, K.; Toda, E.; Bessho-Uehara, K.; Liu, X.; Zhu, S.; Wu, X.; Kasahara, R.D. High-quality sugar production by osgcs1 rice. Communications Biology 2020, 3, 617. [Google Scholar] [CrossRef]
- Howell, S.H. Endoplasmic reticulum stress responses in plants. Annual Review of Plant Biology 2013, 64, 477–499. [Google Scholar] [CrossRef]
- Hu, Y.; Zhou, L.; Huang, M.; He, X.; Yang, Y.; Liu, X.; Li, Y.; Hou, X. Gibberellins play an essential role in late embryogenesis of Arabidopsis. Nature Plants 2018, 4, 289–298. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, L.; Malik, S.; Gentile, B.R.; Xiong, V.; Arazi, T.; Owen, H.A.; Friml, J.; Zhao, D. Specification of female germline by microRNA orchestrated auxin signaling in Arabidopsis. Nature Communications 2022, 13, 6960. [Google Scholar] [CrossRef]
- Huang, W.J.; Liu, H.K.; McCormick, S.; Tang, W.H. Tomato pistil factor STIG1 promotes in vivo pollen tube growth by binding to phosphatidylinositol 3-phosphate and the extracellular domain of the pollen receptor kinase LePRK2. The Plant Cell 2014, 26, 2505–2523. [Google Scholar] [CrossRef]
- Ingouff, M.; Hamamura, Y.; Gourgues, M.; Higashiyama, T.; Berger, F. Distinct dynamics of HISTONE3 variants between the two fertilization products in plants. Curr Biol 2007, 17, 1032–1037. [Google Scholar] [CrossRef]
- Ingram, G.C. Family life at close quarters: communication and constraint in angiosperm seed development. Protoplasma 2010, 247, 195–214. [Google Scholar] [CrossRef]
- Jiang, T.; Li, L.; Hu, Q.; Kuang, X.; Zhang, L.; Zeng, W.; Miki, D.; Zheng, B. The DNA methylation–demethylation balance prevents development of multiple megaspore mother cells in Arabidopsis. The Plant Cell 2025, 37. [Google Scholar] [CrossRef]
- Jiang, W.-B.; Huang, H.-Y.; Hu, Y.-W.; Zhu, S.-W.; Wang, Z.-Y.; Lin, W.-H. Brassinosteroid regulates seed size and shape. Arabidopsis Plant Physiology 2013, 162, 1965–1977. [Google Scholar] [CrossRef]
- Johnson, K.L.; Degnan, K.A.; Ross Walker, J.; Ingram, G.C. AtDEK1 is essential for specification of embryonic epidermal cell fate. The Plant Journal 2005, 44, 114–127. [Google Scholar] [CrossRef]
- Johnston, A.J.; Matveeva, E.; Kirioukhova, O.; Grossniklaus, U.; Gruissem, W. A dynamic reciprocal RBR-PRC2 regulatory circuit controls Arabidopsis gametophyte development. Current Biology 2008, 18, 1680–1686. [Google Scholar] [CrossRef]
- Jullien, P.E.; Mosquna, A.; Ingouff, M.; Sakata, T.; Ohad, N.; Berger, F. Retinoblastoma and its binding partner MSI1 control imprinting in Arabidopsis. PLoS Biol 2008, 6, e194. [Google Scholar] [CrossRef]
- Jun, J.; Fiume, E.; Roeder, A.H.K.; Meng, L.; Sharma, V.K.; Osmont, K.S.; Baker, C.; Ha, C.M.; Meyerowitz, E.M.; Feldman, L.J.; Fletcher, J.C. Comprehensive analysis of CLE polypeptide signaling gene expression and overexpression activity in Arabidopsis. Plant Physiology 2010, 154, 1721–1736. [Google Scholar] [CrossRef]
- Kang, X.; Li, W.; Zhou, Y.; Ni, M. A WRKY Transcription Factor Recruits the SYG1-Like Protein SHB1 to Activate Gene Expression and Seed Cavity Enlargement. PLOS Genetics 2013, 9, e1003347. [Google Scholar] [CrossRef]
- Kao, P.; Nodine, M.D. Transcriptional activation of Arabidopsis zygotes is required for initial cell divisions. Scientific Reports 2019, 9, 17159. [Google Scholar] [CrossRef]
- Kasahara, R.D. Fertilization initiates seed nutrition via phloem end by a callose degradation enzyme. DNA and Cell Biology 2025, 44, 407–410. [Google Scholar] [CrossRef]
- Kasahara, R.D.; Notaguchi, M.; Nagahara, S.; Suzuki, T.; Susaki, D.; Honma, Y.; Maruyama, D.; Higashiyama, T. Pollen tube contents initiate ovule enlargement and enhance seed coat development without fertilization. Science Advances 2016, 2, e1600554. [Google Scholar] [CrossRef]
- Ke, Y.; Podio, M.; Conner, J.; Ozias-Akins, P. Single-cell transcriptome profiling of buffelgrass (Cenchrus ciliaris) eggs unveils apomictic parthenogenesis signatures. Sci Rep 2021, 11, 9880. [Google Scholar] [CrossRef]
- Khanday, I.; Skinner, D.; Yang, B.; Mercier, R.; Sundaresan, V. A male-expressed rice embryogenic trigger redirected for asexual propagation through seeds. Nature 2019, 565, 91–95. [Google Scholar] [CrossRef]
- Köhler, C.; Hennig, L.; Bouveret, R.; Gheyselinck, J.; Grossniklaus, U.; Gruissem, W. Arabidopsis MSI1 is a component of the MEA/FIE Polycomb group complex and required for seed development. The EMBO Journal 2003, 22, 4804–4814. [Google Scholar] [CrossRef]
- Kradolfer, D.; Hennig, L.; Kohler, C. Increased maternal genome dosage bypasses the requirement of the FIS Polycomb Repressive Complex 2 in Arabidopsis seed development. PLoS Genet 2013, 9, e1003163. [Google Scholar] [CrossRef]
- Kranz, E.; Lörz, H. In vitro fertilisation of maize by single egg and sperm cell protoplast fusion mediated by high calcium and high pH. Zygote 1994, 2, 125–128. [Google Scholar] [CrossRef]
- Kujirai, T.; Echigoya, K.; Kishi, Y.; Saeki, M.; Ito, T.; Kato, J.; Negishi, L.; Kimura, H.; Masumoto, H.; Takizawa, Y.; Gotoh, Y.; Kurumizaka, H. Structural insights into how DEK nucleosome binding facilitates H3K27 trimethylation in chromatin. Nature Structural & Molecular Biology 2025, 32, 1183–1192. [Google Scholar] [CrossRef]
- Lafon-Placette, C.; Köhler, C. Embryo and endosperm, partners in seed development. Current Opinion in Plant Biology 2014, 17, 64–69. [Google Scholar] [CrossRef]
- Larsson, E.; Vivian-Smith, A.; Offringa, R.; Sundberg, E. Auxin homeostasis in arabidopsis ovules is anther-dependent at maturation and changes dynamically upon fertilization. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef]
- Laux, T.; Mayer, K.F.X.; Berger, J.; Jürgens, G. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar] [CrossRef]
- Lee, H.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Arabidopsis LEAFY COTYLEDON1 represents a functionally specialized subunit of the CCAAT binding transcription factor. Proceedings of the National Academy of Sciences 2003, 100, 2152–2156. [Google Scholar] [CrossRef]
- Li, C.; Hu, F.; Chen, H.; Zhao, J. Transcriptome characteristics during cell wall formation of endosperm cellularization and embryo differentiation in Arabidopsis. Front Plant Sci 2022, 13, 998664. [Google Scholar] [CrossRef]
- Lid, S.E.; Gruis, D.; Jung, R.; Lorentzen, J.A.; Ananiev, E.; Chamberlin, M.; Niu, X.; Meeley, R.; Nichols, S.; Olsen, O.-A. The defective kernel 1 (dek1) gene required for aleurone cell development in the endosperm of maize grains encodes a membrane protein of the calpain gene superfamily. Proceedings of the National Academy of Sciences 2002, 99, 5460–5465. [Google Scholar] [CrossRef]
- Lima, R.B.; Pankaj, R.; Ehlert, S.T.; Finger, P.; Fröhlich, A.; Bayle, V.; Landrein, B.; Sampathkumar, A.; Figueiredo, D.D. Seed coat-derived brassinosteroid signaling regulates endosperm development. Nature Communications 2024, 15, 9352. [Google Scholar] [CrossRef]
- Liu, H.; Luo, Q.; Tan, C.; Song, J.; Zhang, T.; Men, S. Biosynthesis- and transport-mediated dynamic auxin distribution during seed development controls seed size in Arabidopsis. The Plant Journal 2023, 113, 1259–1277. [Google Scholar] [CrossRef]
- Liu, X.; Adhikari, P.; Kasahara, R. Pollen tube contents from failed fertilization contribute to seed coat initiation in Arabidopsis. F1000Research 2019, 8. [Google Scholar] [CrossRef]
- Liu, X.; Nakajima, K.P.; Adhikari, P.B.; Wu, X.; Zhu, S.; Okada, K.; Kagenishi, T.; Kurotani, K.-i.; Ishida, T.; Nakamura, M.; Sato, Y.; Kawakatsu, Y.; Xie, L.; Huang, C.; He, J.; Yokawa, K.; Sawa, S.; Higashiyama, T.; Bradford, K.J.; Notaguchi, M.; Kasahara, R.D. Fertilization-dependent phloem end gate regulates seed size. Current Biology 2025a, 35, 2049–2063.e2043. [Google Scholar] [CrossRef]
- Liu, X.Q.; Shi, J.J.; Fan, H.; Jiao, J.; Gao, L.; Tan, L.; Nagawa, S.; Wang, D.Y. Nuclear DNA replicates during zygote development in Arabidopsis and Torenia fournieri. Plant Physiology 2020, 185, 137–145. [Google Scholar] [CrossRef]
- Liu, Y.; Xiao, S.; Yang, M.; Guo, G.; Zhou, Y. The impact of Polycomb Group proteins on 3D chromatin structure and environmental stresses in plants. Plants 2025b, 14, 1038. [Google Scholar] [CrossRef]
- Lu, B.; Wang, S.; Feng, H.; Wang, J.; Zhang, K.; Li, Y.; Wu, P.; Zhang, M.; Xia, Y.; Peng, C.; Li, C. FERONIA-mediated TIR1/AFB2 oxidation stimulates auxin signaling in Arabidopsis. Molecular Plant 2024, 17, 772–787. [Google Scholar] [CrossRef]
- Lucas, W.J.; Bouché-Pillon, S.; Jackson, D.P.; Nguyen, L.; Baker, L.; Ding, B.; Hake, S. Selective trafficking of KNOTTED1 homeodomain protein and its mRNA through plasmodesmata. Science 1995, 270, 1980–1983. [Google Scholar] [CrossRef]
- Lukowitz, W.; Roeder, A.; Parmenter, D.; Somerville, C. A MAPKK kinase gene regulates extra-embryonic cell fate in Arabidopsis. Cell 2004, 116, 109–119. [Google Scholar] [CrossRef]
- Lv, Y.; Li, J.; Wang, Z.; Liu, Y.; Jiang, Y.; Li, Y.; Lv, Z.; Huang, X.; Peng, X.; Cao, Y.; Yang, H. Polycomb proteins RING1A/B promote H2A monoubiquitination to regulate female gametophyte development in Arabidopsis. Journal of Experimental Botany 2024, 75, 4822–4836. [Google Scholar] [CrossRef]
- Maruyama, D.; Völz, R.; Takeuchi, H.; Mori, T.; Igawa, T.; Kurihara, D.; Kawashima, T.; Ueda, M.; Ito, M.; Umeda, M.; Nishikawa, S.I.; Groß-Hardt, R.; Higashiyama, T. Rapid elimination of the persistent synergid through a cell fusion mechanism. Cell 2015, 161, 907–918. [Google Scholar] [CrossRef]
- Mathews, S.; Kramer, E.M. The evolution of reproductive structures in seed plants: a re-examination based on insights from developmental genetics. New Phytologist 2012, 194, 910–923. [Google Scholar] [CrossRef]
- Matzke, M.A.; Mosher, R.A. RNA-directed DNA methylation: an epigenetic pathway of increasing complexity. Nature Reviews Genetics 2014, 15, 394–408. [Google Scholar] [CrossRef]
- Meixner, A.; Boldt, K.; Van Troys, M.; Askenazi, M.; Gloeckner, C.J.; Bauer, M.; Marto, J.A.; Ampe, C.; Kinkl, N.; Ueffing, M. A QUICK screen for Lrrk2 interaction partners--leucine-rich repeat kinase 2 is involved in actin cytoskeleton dynamics. Mol Cell Proteomics 2011, 10, M110.001172. [Google Scholar] [CrossRef]
- Mendes, M.A.; Petrella, R.; Cucinotta, M.; Vignati, E.; Gatti, S.; Pinto, S.C.; Bird, D.C.; Gregis, V.; Dickinson, H.; Tucker, M.R.; Colombo, L. The RNA-dependent DNA methylation pathway is required to restrict SPOROCYTELESS/NOZZLE expression to specify a single female germ cell precursor in Arabidopsis. Development 2020, 147. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Kovalchuk, I. Epigenetic modifications during angiosperm gametogenesis. Front Plant Sci 2012, 3, 20. [Google Scholar] [CrossRef]
- Möller, B.; Weijers, D. Auxin control of embryo patterning. Cold Spring Harb Perspect Biol 2009, 1, a001545. [Google Scholar] [CrossRef]
- Montgomery, S.A.; Berger, F. Epigenetic reprogramming of imprinting at meiosis. bioRxiv 2023, 2017.541143. [Google Scholar] [CrossRef]
- Moreno-Romero, J.; Jiang, H.; Santos-González, J.; Köhler, C. Parental epigenetic asymmetry of PRC2-mediated histone modifications in the Arabidopsis endosperm. The EMBO Journal 2016, 35, 1298–1311. [Google Scholar] [CrossRef]
- Moussu, S.; Doll, N.M.; Chamot, S.; Brocard, L.; Creff, A.; Fourquin, C.; Widiez, T.; Nimchuk, Z.L.; Ingram, G. ZHOUPI and KERBEROS mediate embryo/endosperm separation by promoting the formation of an extracuticular sheath at the embryo surface. Plant Cell 2017, 29, 1642–1656. [Google Scholar] [CrossRef]
- Muller, H.J. The relation of recombination to mutational advance. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 1964, 1, 2–9. [Google Scholar] [CrossRef]
- Murata, Y.; Pei, Z.M.; Mori, I.C.; Schroeder, J. Abscisic acid activation of plasma membrane Ca2+ channels in guard cells requires cytosolic NAD(P)H and is differentially disrupted upstream and downstream of reactive oxygen species production in abi1-1 and abi2-1 protein phosphatase 2C mutants. Plant Cell 2001, 13, 2513–2523. [Google Scholar] [CrossRef]
- Nakajima, K.P.; Kasahara, R.D. How do seeds get bigger? A newly identified phloem-terminal tissue controls seed enlargement through callose degradation. Academia Molecular Biology and Genomics 2025, 2. [Google Scholar] [CrossRef]
- Nakamura, M.; Bente, H.; Derkacheva, M.; Gentry, M.; Landberg, K.; Thelander, M.; Sundberg, E.; Hennig, L.; Köhler, C. Chromatin-associated DEK proteins maintain H3K27me3 balance and coordinate developmental transitions in plants. New Phytol 2026, 249, 930–944. [Google Scholar] [CrossRef]
- Nodine, M.D.; Bartel, D.P. Maternal and paternal genomes contribute equally to the transcriptome of early plant embryos. Nature 2012, 482, 94–97. [Google Scholar] [CrossRef]
- Nowack, M.K.; Grini, P.E.; Jakoby, M.J.; Lafos, M.; Koncz, C.; Schnittger, A. A positive signal from the fertilization of the egg cell sets off endosperm proliferation in angiosperm embryogenesis. Nature Genetics 2006, 38, 63–67. [Google Scholar] [CrossRef]
- O'Neill, J.P.; Colon, K.T.; Jenik, P.D. The onset of embryo maturation in Arabidopsis is determined by its developmental stage and does not depend on endosperm cellularization. Plant J 2019, 99, 286–301. [Google Scholar] [CrossRef]
- Ogawa, E.; Yamada, Y.; Sezaki, N.; Kosaka, S.; Kondo, H.; Kamata, N.; Abe, M.; Komeda, Y.; Takahashi, T. ATML1 and PDF2 play a redundant and essential role in Arabidopsis embryo development. Plant and Cell Physiology 2015, 56, 1183–1192. [Google Scholar] [CrossRef]
- Olmedo-Monfil, V.; Durán-Figueroa, N.; Arteaga-Vázquez, M.; Demesa-Arévalo, E.; Autran, D.; Grimanelli, D.; Slotkin, R.K.; Martienssen, R.A.; Vielle-Calzada, J.P. Control of female gamete formation by a small RNA pathway in Arabidopsis. Nature 2010, 464, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Pankaj, R.; Lima, R.B.; Figueiredo, D.D. Hormonal regulation and crosstalk during early endosperm and seed coat development. Plant Reprod 2024a, 38, 5. [Google Scholar] [CrossRef]
- Pankaj, R.; Lima, R.B.; Luo, G.-Y.; Ehlert, S.; del Toro-de León, G.; Bente, H.; Finger, P.; Sato, H.; Figueiredo, D.D. BRI1-mediated removal of seed coat H3K27me3 marks is a brassinosteroid-independent process. bioRxiv 2024b, 2007.569203. [Google Scholar]
- Park, J.-S.; Frost, J.M.; Park, K.; Ohr, H.; Park, G.T.; Kim, S.; Eom, H.; Lee, I.; Brooks, J.S.; Fischer, R.L.; Choi, Y. Control of DEMETER DNA demethylase gene transcription in male and female gamete companion cells in Arabidopsis thaliana. Proceedings of the National Academy of Sciences 2017, 114, 2078–2083. [Google Scholar] [CrossRef]
- Pelletier, J.M.; Kwong, R.W.; Park, S.; Le, B.H.; Baden, R.; Cagliari, A.; Hashimoto, M.; Munoz, M.D.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEC1 sequentially regulates the transcription of genes involved in diverse developmental processes during seed development. Proceedings of the National Academy of Sciences 2017, 114, E6710–E6719. [Google Scholar] [CrossRef]
- Plong, A.; Rodriguez, K.; Alber, M.; Chen, W.; Reddy, G.V. CLAVATA3 mediated simultaneous control of transcriptional and post-translational processes provides robustness to the WUSCHEL gradient. Nature Communications 2021, 12, 6361. [Google Scholar] [CrossRef]
- Qin, Y.; Zhao, L.; Skaggs, M.I.; Andreuzza, S.; Tsukamoto, T.; Panoli, A.; Wallace, K.N.; Smith, S.; Siddiqi, I.; Yang, Z.; Yadegari, R.; Palanivelu, R. ACTIN-RELATED PROTEIN6 regulates female meiosis by modulating meiotic gene expression in Arabidopsis. The Plant Cell 2014, 26, 1612–1628. [Google Scholar] [CrossRef]
- Rampey, R.A.; LeClere, S.; Kowalczyk, M.; Ljung, K.; Sandberg, G.; Bartel, B. A family of auxin-conjugate hydrolases that contributes to free indole-3-acetic acid levels during Arabidopsis germination. Plant Physiol 2004, 135, 978–988. [Google Scholar] [CrossRef]
- Rani, S.; Roy, S.; Nandi, A.K. Pathogen induces MEDEA gene in a parent-of-origin nonspecific manner and causes embryo lethality in MEDEA overexpressing plants. Journal of Plant Biochemistry and Biotechnology 2025, 34, 933–940. [Google Scholar] [CrossRef]
- Reyes, J.L.; Chua, N.-H. ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination. The Plant Journal 2007, 49, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Leal, D.; León-Martínez, G.; Abad-Vivero, U.; Vielle-Calzada, J.-P. Natural variation in epigenetic pathways affects the specification of female gamete precursors in Arabidopsis. The Plant Cell 2015, 27, 1034–1045. [Google Scholar] [CrossRef]
- Ronceret, A.; Devic, M. Redrawing the borderline: Control of DNA replication at fertilization. Plant Signal Behav 2008, 3, 554–555. [Google Scholar] [CrossRef]
- Roszak, P.; Köhler, C. Polycomb group proteins are required to couple seed coat initiation to fertilization. Proceedings of the National Academy of Sciences 2011, 108, 20826–20831. [Google Scholar] [CrossRef] [PubMed]
- Rudall, P.J. Evolution and patterning of the ovule in seed plants. Biological Reviews 2021, 96, 943–960. [Google Scholar] [CrossRef] [PubMed]
- Rydin, C.; Khodabandeh, A.; Endress, P.K. The female reproductive unit of Ephedra (Gnetales): comparative morphology and evolutionary perspectives. Botanical Journal of the Linnean Society 2010, 163, 387–430. [Google Scholar] [CrossRef]
- Sabelli, P.A.; Liu, Y.; Dante, R.A.; Lizarraga, L.E.; Nguyen, H.N.; Brown, S.W.; Klingler, J.P.; Yu, J.; LaBrant, E.; Layton, T.M.; Feldman, M.; Larkins, B.A. Control of cell proliferation, endoreduplication, cell size, and cell death by the retinoblastoma-related pathway in maize endosperm. Proc Natl Acad Sci U S A 2013, 110, E1827-1836. [Google Scholar] [CrossRef]
- Schröder, J.A.; Bonnet, D.M.V.; Jullien, P.E. Non-cell-autonomous small RNA silencing in Arabidopsis female gametes. Current Biology 2023, 33, 183–188.e183. [Google Scholar] [CrossRef] [PubMed]
- Schruff, M.C.; Spielman, M.; Tiwari, S.; Adams, S.; Fenby, N.; Scott, R.J. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 2006, 133, 251–261. [Google Scholar] [CrossRef]
- Simonini, S. Regulation of cell cycle in plant gametes: when is the right time to divide? Development 2025, 152. [Google Scholar] [CrossRef]
- Simonini, S.; Bemer, M.; Bencivenga, S.; Gagliardini, V.; Pires, N.D.; Desvoyes, B.; Van der Graaff, E.; Gutierrez, C.; Grossniklaus, U. The Polycomb group protein MEDEA controls cell proliferation and embryonic patterning in Arabidopsis. Developmental Cell 2021, 56, 1945–1960. e1947. [Google Scholar] [CrossRef]
- Simonini, S.; Bencivenga, S.; Grossniklaus, U. A paternal signal induces endosperm proliferation upon fertilization in Arabidopsis. Science 2024, 383, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Solfanelli, C.; Ceron, F.; Paolicchi, F.; Giorgetti, L.; Geri, C.; Ceccarelli, N.; Kamiya, Y.; Picciarelli, P. Expression of two genes encoding gibberellin 2- and 3-oxidases in developing seeds of Phaseolus coccineus. Plant and Cell Physiology 2005, 46, 1116–1124. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Xie, X.; Chen, C.; Shu, J.; Thapa, R.K.; Nguyen, V.; Bian, S.; Kohalmi, S.E.; Marsolais, F.; Zou, J.; Cui, Y. LEAFY COTYLEDON1 expression in the endosperm enables embryo maturation in Arabidopsis. Nature Communications 2021, 12, 3963. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Xie, X.; Mavraganis, I.; Yu, B.; Shen, W.; Yang, H.; Xiang, D.; Wei, Y.; Cui, Y.; Zou, J. Tracking the genome-wide occupancy of Arabidopsis LEAFY COTYLEDON1 in endosperm development. Commun Biol 2025, 8, 771. [Google Scholar] [CrossRef] [PubMed]
- Sornay, E.; Forzani, C.; Forero-Vargas, M.; Dewitte, W.; Murray, J.A.H. Activation of CYCD7;1 in the central cell and early endosperm overcomes cell-cycle arrest in the Arabidopsis female gametophyte, and promotes early endosperm and embryo development. The Plant Journal 2015, 84, 41–55. [Google Scholar] [CrossRef]
- Szczuka, E.; Szczuka, A. Cuticle fluorescence during embryogenesis of Arabidopsis thaliana (L.) Heynh. Acta Biol. Cracov. Ser. Bot 2003, 45, 63–67. [Google Scholar]
- Takeuchi, N.; Kaneko, K.; Koonin, E.V. Horizontal gene transfer can rescue prokaryotes from muller’s ratchet: Benefit of DNA from dead cells and population subdivision. G3 GenesGenomesGenetics 2014, 4, 325–339. [Google Scholar] [CrossRef]
- Tanaka, H.; Onouchi, H.; Kondo, M.; Hara-Nishimura, I.; Nishimura, M.; Machida, C.; Machida, Y. A subtilisin-like serine protease is required for epidermal surface formation in Arabidopsis embryos and juvenile plants. Development 2001, 128, 4681–4689. [Google Scholar] [CrossRef]
- Tang, W.; Kelley, D.; Ezcurra, I.; Cotter, R.; McCormick, S. LeSTIG1, an extracellular binding partner for the pollen receptor kinases LePRK1 and LePRK2, promotes pollen tube growth in vitro. The Plant Journal 2004, 39, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Tovar-Aguilar, A.; Grimanelli, D.; Acosta-García, G.; Vielle-Calzada, J.-P.; Badillo-Corona, J.A.; Durán-Figueroa, N. The miRNA822 loaded by ARGONAUTE9 modulates the monosporic female gametogenesis in Arabidopsis thaliana. Plant Reproduction 2024, 37, 243–258. [Google Scholar] [CrossRef]
- Tsering, T.; Di Donato, M.; Samakovli, D.; Milioni, D.; Iacobini, F.R.; Panagiotopoulos, K.; Plitsi, P.-K.; Azarello, E.; Mancuso, S.; Pukyšová, V.; Zwiewka, M.; Nodzynski, T.; Stumpe, M.; Ludwig-Müller, J.; Bailly, A.; Hatzopoulos, P.; Geisler, M.M. HSP90 differentially stabilizes plant ABCB-type auxin transporters on the plasma membrane. Nature Communications 2025, 16, 8643. [Google Scholar] [CrossRef]
- Tsuwamoto, R.; Fukuoka, H.; Takahata, Y. GASSHO1 and GASSHO2 encoding a putative leucine-rich repeat transmembrane-type receptor kinase are essential for the normal development of the epidermal surface in Arabidopsis embryos. The Plant Journal 2008, 54, 30–42. [Google Scholar] [CrossRef]
- Turck, F.; Roudier, F.; Farrona, S.; Martin-Magniette, M.-L.; Guillaume, E.; Buisine, N.; Gagnot, S.; Martienssen, R.A.; Coupland, G.; Colot, V. Arabidopsis TFL2/LHP1 specifically associates with genes marked by trimethylation of histone H3 lysine 27. PLOS Genetics 2007, 3, e86. [Google Scholar] [CrossRef]
- Ueda, M.; Aichinger, E.; Gong, W.; Groot, E.; Verstraeten, I.; Dai Vu, L.; De Smet, I.; Higashiyama, T.; Umeda, M.; Laux, T. Transcriptional integration of paternal and maternal factors in the Arabidopsis zygote. Genes & development 2017, 31, 617–627. [Google Scholar]
- Ueda, M.; Zhang, Z.; Laux, T. Transcriptional activation of Arabidopsis axis patterning genes WOX8/9 links zygote polarity to embryo development. Developmental Cell 2011, 20, 264–270. [Google Scholar] [CrossRef]
- Van Durme, M.; Olvera-Carrillo, Y.; Pfeiffer, M.L.; Doll, N.M.; De Winter, F.; Lin, Z.; Nowack, M.K. Fertility loss in senescing Arabidopsis ovules is controlled by the maternal sporophyte via a NAC transcription factor triad. Proceedings of the National Academy of Sciences 2023, 120, e2219868120. [Google Scholar] [CrossRef]
- Vanneste, S.; Pei, Y.; Friml, J. Mechanisms of auxin action in plant growth and development. Nature Reviews Molecular Cell Biology 2025, 26, 648–666. [Google Scholar] [CrossRef] [PubMed]
- Veluchamy, A.; Jégu, T.; Ariel, F.; Latrasse, D.; Mariappan, K.G.; Kim, S.K.; Crespi, M.; Hirt, H.; Bergounioux, C.; Raynaud, C.; Benhamed, M. LHP1 regulates H3K27me3 spreading and shapes the three-dimensional conformation of the Arabidopsis genome. PLoS One 2016, 11, e0158936. [Google Scholar] [CrossRef]
- Wang, H.; Ngwenyama, N.; Liu, Y.; Walker, J.C.; Zhang, S. Stomatal development and patterning are regulated by environmentally responsive mitogen-activated protein kinases in Arabidopsis. Plant Cell 2007, 19, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ko, E.E.; Tran, J.; Qiao, H. TREE1-EIN3–mediated transcriptional repression inhibits shoot growth in response to ethylene. Proceedings of the National Academy of Sciences 2020, 117, 29178–29189. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wan, J.; Dang, K.; Meng, S.; Hu, D.; Lin, Y.; Qiu, X.; Guo, Z.; Fu, Z.; Ding, D.; Tang, J. zma-miR159 targets ZmMYB74 and ZmMYB138 transcription factors to regulate grain size and weight in maize. Plant Physiology 2023, 193, 2430–2441. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, Y.; Gu, H.; Kang, D.; Chen, Z.; Liu, J.; Qu, L.-J. The Arabidopsis APC4 subunit of the anaphase-promoting complex/cyclosome (APC/C) is critical for both female gametogenesis and embryogenesis. The Plant Journal 2012, 69, 227–240. [Google Scholar] [CrossRef]
- Wei, B.; Zhang, J.; Pang, C.; Yu, H.; Guo, D.; Jiang, H.; Ding, M.; Chen, Z.; Tao, Q.; Gu, H.; Qu, L.-J.; Qin, G. The molecular mechanism of SPOROCYTELESS/NOZZLE in controlling Arabidopsis ovule development. Cell Research 2015, 25, 121–134. [Google Scholar] [CrossRef]
- Weimer, A.K.; Matos, J.L.; Sharma, N.; Patell, F.; Murray, J.A.H.; Dewitte, W.; Bergmann, D.C. Lineage- and stage-specific expressed CYCD7;1 coordinates the single symmetric division that creates stomatal guard cells. Development (Cambridge, England) 2018, pp. dev160671. [Google Scholar] [CrossRef]
- Weinhofer, I.; Hehenberger, E.; Roszak, P.; Hennig, L.; Köhler, C. H3K27me3 profiling of the endosperm implies exclusion of polycomb group protein targeting by DNA methylation. PLOS Genetics 2010, 6, e1001152. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Otegui, M.S.; Spalding, E.P. The ER-Localized TWD1 immunophilin is necessary for localization of multidrug resistance-like proteins required for polar auxin transport in Arabidopsis roots. The Plant Cell 2010, 22, 3295–3304. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, X.; Chen, J.X.; Chaumier, T.; Tirichine, L. The RING1 subunit of Polycomb Repressive Complex 1 monoubiquitinates histone H2A and mediates repression independently of Polycomb Repressive Complex 2 in the model diatom Phaeodactylum tricornutum. New Phytologist 2025. [Google Scholar] [CrossRef] [PubMed]
- Wyrzykowska, A.; Bielewicz, D.; Plewka, P.; Sołtys-Kalina, D.; Wasilewicz-Flis, I.; Marczewski, W.; Jarmolowski, A.; Szweykowska-Kulinska, Z. The MYB33, MYB65, and MYB101 transcription factors affect Arabidopsis and potato responses to drought by regulating the ABA signaling pathway. Physiologia Plantarum 2022, 174, e13775. [Google Scholar] [CrossRef]
- Xiao, W.; Gehring, M.; Choi, Y.; Margossian, L.; Pu, H.; Harada, J.J.; Goldberg, R.B.; Pennell, R.I.; Fischer, R.L. Imprinting of the MEA polycomb gene is controlled by antagonism between MET1 methyltransferase and DME glycosylase. Developmental Cell 2003, 5, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Wang, W.; Sun, M.-X. Endosperm development is an autonomously programmed process independent of embryogenesis. The Plant Cell 2021, 33, 1151–1160. [Google Scholar] [CrossRef]
- Xu, T.-T.; Ren, S.-C.; Song, X.-F.; Liu, C.-M. CLE19 expressed in the embryo regulates both cotyledon establishment and endosperm development in Arabidopsis. Journal of Experimental Botany 2015, 66, 5217–5227. [Google Scholar] [CrossRef]
- Xu, W.; Fiume, E.; Coen, O.; Pechoux, C.; Lepiniec, L.; Magnani, E. Endosperm and nucellus develop antagonistically in Arabidopsis seeds. Plant Cell 2016, 28, 1343–1360. [Google Scholar] [CrossRef]
- Xu, W.; Iannaccone, M.; Gomez-Paez, D.-M.; Choinard, S.; Lu, J.; Le Hir, R.; Dinant, S.; Kalmbach, L.; Feil, R.; Lunn, J.E.; Meyer, C.; Magnani, E. Too much sugar makes plants ‘pregnant’: maternal sucrose signals fertilization in Arabidopsis seeds. bioRxiv 2025, 2025.2005.2016.654273. [Google Scholar] [CrossRef]
- Yamada, T.; Yokota, S.y.; Hirayama, Y.; Imaichi, R.; Kato, M.; Gasser, C.S. Ancestral expression patterns and evolutionary diversification of YABBY genes in angiosperms. The Plant Journal 2011, 67, 26–36. [Google Scholar] [CrossRef]
- Yang, S.; Johnston, N.; Talideh, E.; Mitchell, S.; Jeffree, C.; Goodrich, J.; Ingram, G. The endosperm-specific ZHOUPI gene of Arabidopsis thaliana regulates endosperm breakdown and embryonic epidermal development. Development 2008, 135, 3501–3509. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Peragine, A.; Park, M.Y.; Poethig, R.S. A pathway for the biogenesis of trans-acting siRNAs in Arabidopsis. Genes & Development 2005, 19, 2164–2175. [Google Scholar]
- Yoshimura, K.; Iida, K.; Iida, H. MCAs in Arabidopsis are Ca2+-permeable mechanosensitive channels inherently sensitive to membrane tension. Nature Communications 2021, 12, 6074. [Google Scholar] [CrossRef]
- Yu, F.; Li, J.; Huang, Y.; Liu, L.; Li, D.; Chen, L.; Luan, S. FERONIA receptor kinase controls seed size in Arabidopsis thaliana. Molecular Plant 2014, 7, 920–922. [Google Scholar] [CrossRef]
- Yu, T.-Y.; Wang, P.; Lv, Y.; Wang, B.; Zhao, M.-R.; Dong, X.-W. Auxin orchestrates germ cell specification in Arabidopsis. International Journal of Molecular Sciences 2025, 26, 3257. [Google Scholar] [CrossRef]
- Yu, T.Y.; Shi, D.Q.; Jia, P.F.; Tang, J.; Li, H.J.; Liu, J.; Yang, W.C. The Arabidopsis receptor kinase ZAR1 is required for zygote asymmetric division and its daughter cell fate. PLoS Genet 2016, 12, e1005933. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, X.; Zhao, P.; Peng, X.; Chen, H.; Bleckmann, A.; Bazhenova, A.; Shi, C.; Dresselhaus, T.; Sun, M.X. Fertilized egg cells secrete endopeptidases to avoid polytubey. Nature 2021, 592, 433–437. [Google Scholar] [CrossRef]
- Zhang, B.; Li, C.; Li, Y.; Yu, H. Mobile TERMINAL FLOWER1 determines seed size in Arabidopsis. Nature Plants 2020, 6, 1146–1157. [Google Scholar] [CrossRef]
- Zhang, J.; Yao, J.; He, K.; Yu, C.; Du, J.; Du, J.; Fu, Q.; Yao, R.; Howe, G.A.; Han, X.; Hu, Y. Jasmonate signaling coordinates with the SOD7–KLU pathway to regulate seed size in Arabidopsis thaliana. The Plant Cell 2025, 37. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, H.; Su, J.; Wang, H.; Zhu, Q.; Liu, Y.; Xu, J.; Lukowitz, W.; Zhang, S. Maternal control of embryogenesis by MPK6 and its upstream MKK4/MKK5 in Arabidopsis. The Plant Journal 2017, 92, 1005–1019. [Google Scholar] [CrossRef]
- Zhang, S.; Mohanty, D.; Muzaffar, A.; Ni, M. Two MADS-box proteins, AGL9 and AGL15, recruit the FIS-PRC2 complex to trigger the phase transition from endosperm proliferation to embryo development in Arabidopsis. Molecular Plant 2024, 17, 1110–1128. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, B.; Yan, D.; Dong, W.; Yang, W.; Li, Q.; Zeng, L.; Wang, J.; Wang, L.; Hicks, L.M.; He, Z. Two Arabidopsis cytochrome P450 monooxygenases, CYP714A1 and CYP714A2, function redundantly in plant development through gibberellin deactivation. The Plant Journal 2011, 67, 342–353. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, D.; Kong, F.; Lin, K.; Zhang, H.; Li, G. The Arabidopsis thaliana nuclear factor Y transcription factors. Front Plant Sci 2016, 7, 2045. [Google Scholar] [CrossRef]
- Zhao, L.; Cai, H.; Su, Z.; Wang, L.; Huang, X.; Zhang, M.; Chen, P.; Dai, X.; Zhao, H.; Palanivelu, R.; Chen, X.; Qin, Y. KLU suppresses megasporocyte cell fate through SWR1-mediated activation of WRKY28 expression in Arabidopsis. Proceedings of the National Academy of Sciences 2018a, 115, E526–E535. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Zhou, X.; Shen, K.; Liu, Z.; Cheng, T.; Liu, D.; Cheng, Y.; Peng, X.; Sun, M.-x. Two-step maternal-to-zygotic transition with two-phase parental genome contributions. Developmental Cell 2019, 49, 882–893.e885. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chen, A.; Gao, Z.; Hou, F.; Chen, Y.; Liu, Y. Overexpression of HmWOX8 promotes callus proliferation and shoot regeneration by regulating hormone signaling and shoot development-related genes. Ornamental Plant Research 2024, 4, e026. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, S.; Wu, W.; Li, L.; Jiang, T.; Zheng, B. Clearance of maternal barriers by paternal miR159 to initiate endosperm nuclear division in Arabidopsis. Nature Communications 2018b, 9, 5011. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Zheng, Y.; Ji, H.; Burnie, W.; Perry, S.E. Gene regulation by the AGL15 transcription factor reveals hormone interactions in somatic embryogenesis. Plant Physiology 2016, 172, 2374–2387. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.J.; Khan, D.; Kalichuk, J.L.; Becker, M.G.; Belmonte, M.F. Transcriptome landscape of the early Brassica napus seed. Journal of Integrative Plant Biology 2019, 61, 639–650. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structural similarities between shoot apical meristem (SAM) and ovule primordium (highlighted panels), which subsequently develop into shoot and ovule respectively. ii = inner integument; oi = outer integument.
Figure 1.
Structural similarities between shoot apical meristem (SAM) and ovule primordium (highlighted panels), which subsequently develop into shoot and ovule respectively. ii = inner integument; oi = outer integument.

Figure 2.
Illustrative depiction of the meiosis-with-recombination led persistence of the eukaryotic population over long period of time (green). Any population lacking functional recombination likely succumbed to the Muller’s rachet due to the accumulation of unmanageable deleterious mutation bringing it to its subsequent extinction (red).
Figure 2.
Illustrative depiction of the meiosis-with-recombination led persistence of the eukaryotic population over long period of time (green). Any population lacking functional recombination likely succumbed to the Muller’s rachet due to the accumulation of unmanageable deleterious mutation bringing it to its subsequent extinction (red).
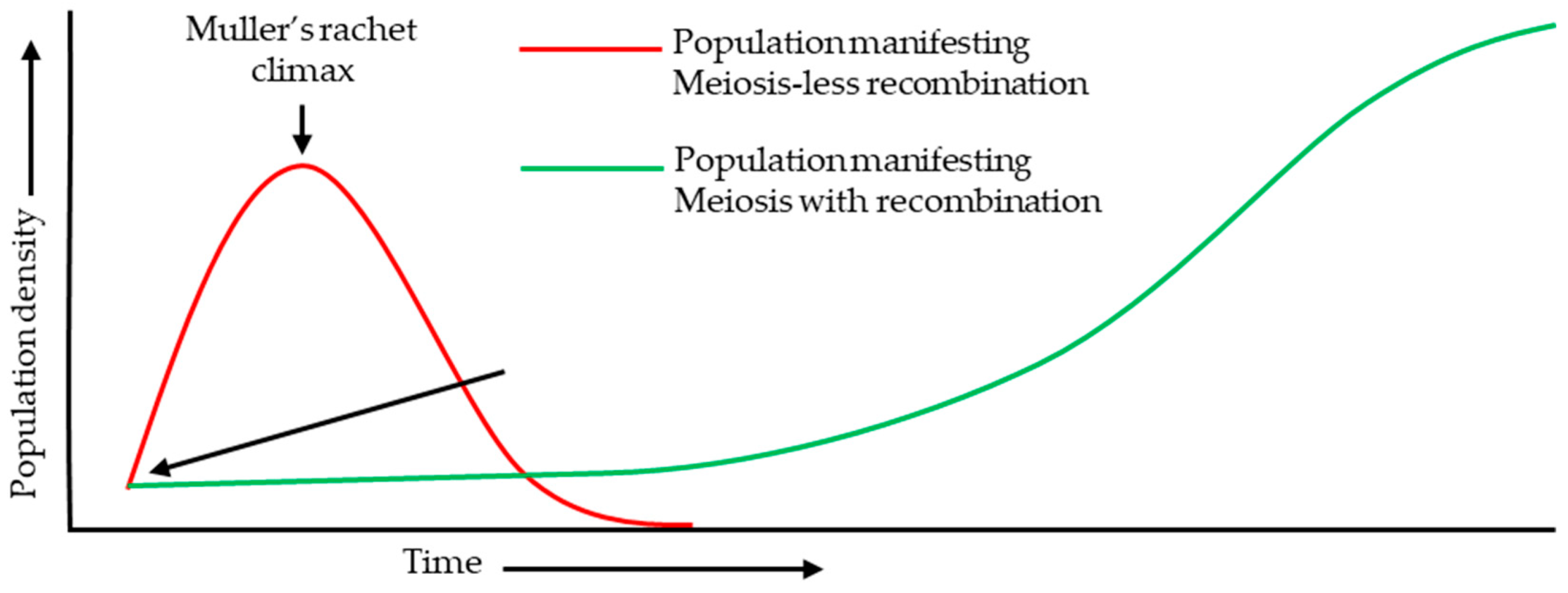
Figure 3.
Typical developmental progression of ovule primordia and MMC speciation. The two images of stage 2 represent for the ovule primordia with normal single MMC speciation (former) and aberrant primordia with more than one L2 cell exhibiting MMC fate.
Figure 3.
Typical developmental progression of ovule primordia and MMC speciation. The two images of stage 2 represent for the ovule primordia with normal single MMC speciation (former) and aberrant primordia with more than one L2 cell exhibiting MMC fate.
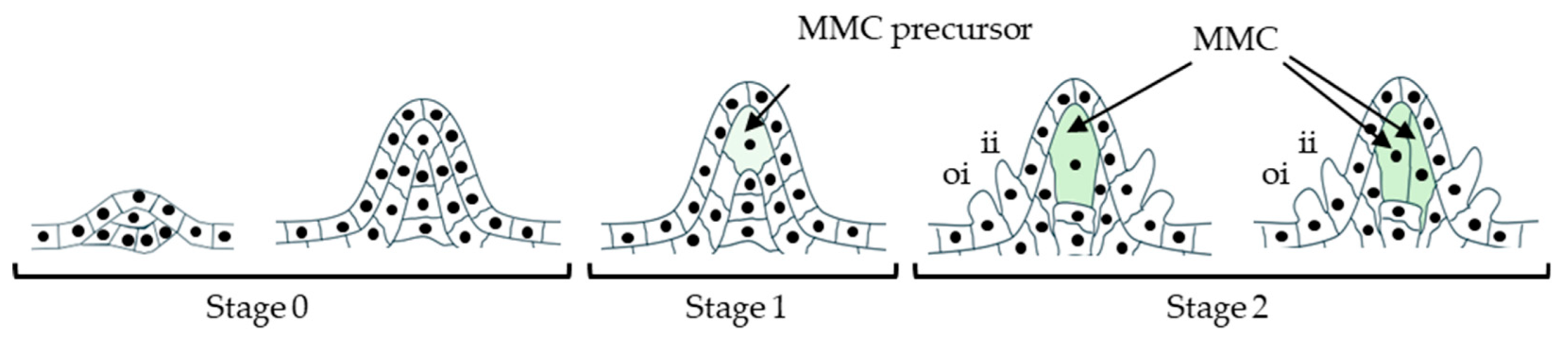
Figure 4.
PRC1/PRC2 and small-RNA–directed epigenetic pathways restrict MMC fate to a single L2 cell and support downstream FM development. (a) PRC1 represses AGO9 via H2Aub1 at target gene loci; in contrast, at transposable elements (TEs), PRC1 + PRC2 co-marking (H2Aub1 + H3K27me3) is enriched, highlighting locus-type–dependent Polycomb outputs (Lv et al., 2024; Wu et al., 2025; Liu et al., 2025b). (b) STK promotes AGO9 and the tasiRNA pathway component RDR6. tasiRNA biogenesis is schematized as precursor processing and stabilization (DCL1, SGS3), dsRNA synthesis (RDR6), and DCL4-dependent production of 21-nt tasiRNAs, which contribute to restricting SPL/NZZ via cytoplasmic RNA decay and/or reinforcing nuclear silencing in surrounding L2 companion cells (Yoshikawa et al., 2005; Matzke and Mosher, 2014; Chen et al., 2016; Mendes et al., 2020).(Bottom left) AGO9-associated RdDM (via DRM1/2) represses SPL/NZZ in MMC-adjacent cells, preventing supernumerary MMC formation; single-MMC outcome is proposed to depend on a balance of methylation (DRM2) and demethylation (DME) rather than methylation alone (Matzke and Mosher, 2014; Mendes et al., 2020; Jiang et al., 2025). (c) miR822–AGO9-linked mRNA degradation of ovule targets (e.g., AT2G13900/ AT5G02330/AT5G02350) supports normal post-meiotic FM development; disruption yields aberrant FG outcomes (Tovar-Aguilar et al., 2024). Green arrows indicate activation/positive effects; red bars indicate repression; dashed lines indicate indirect/putative links.
Figure 4.
PRC1/PRC2 and small-RNA–directed epigenetic pathways restrict MMC fate to a single L2 cell and support downstream FM development. (a) PRC1 represses AGO9 via H2Aub1 at target gene loci; in contrast, at transposable elements (TEs), PRC1 + PRC2 co-marking (H2Aub1 + H3K27me3) is enriched, highlighting locus-type–dependent Polycomb outputs (Lv et al., 2024; Wu et al., 2025; Liu et al., 2025b). (b) STK promotes AGO9 and the tasiRNA pathway component RDR6. tasiRNA biogenesis is schematized as precursor processing and stabilization (DCL1, SGS3), dsRNA synthesis (RDR6), and DCL4-dependent production of 21-nt tasiRNAs, which contribute to restricting SPL/NZZ via cytoplasmic RNA decay and/or reinforcing nuclear silencing in surrounding L2 companion cells (Yoshikawa et al., 2005; Matzke and Mosher, 2014; Chen et al., 2016; Mendes et al., 2020).(Bottom left) AGO9-associated RdDM (via DRM1/2) represses SPL/NZZ in MMC-adjacent cells, preventing supernumerary MMC formation; single-MMC outcome is proposed to depend on a balance of methylation (DRM2) and demethylation (DME) rather than methylation alone (Matzke and Mosher, 2014; Mendes et al., 2020; Jiang et al., 2025). (c) miR822–AGO9-linked mRNA degradation of ovule targets (e.g., AT2G13900/ AT5G02330/AT5G02350) supports normal post-meiotic FM development; disruption yields aberrant FG outcomes (Tovar-Aguilar et al., 2024). Green arrows indicate activation/positive effects; red bars indicate repression; dashed lines indicate indirect/putative links.
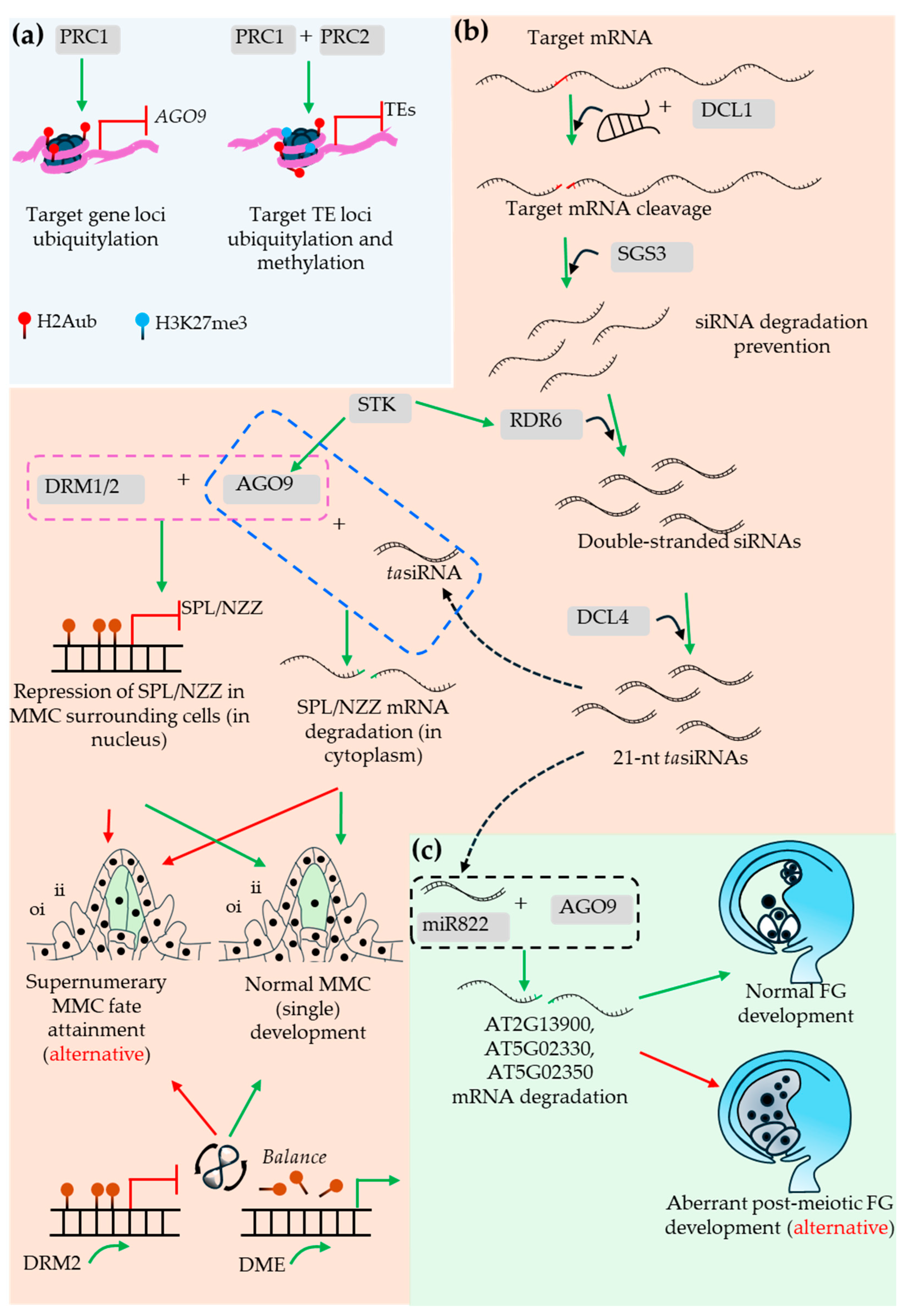
Figure 5.
Key SPL/NZZ-mediated positive regulators (primarily) players behind single-MMC speciation. SPL/NZZ recruits the co-repressor TPL/TPR (via its C-terminal EAR motif) to CIN-like TCP–bound loci and, together with HDA19, represses TCP downstream genes, thereby restricting MMC fate and supporting normal single-MMC development (Wei et al., 2015). SPL/NZZ also functions in a STK/SEP3-associated MADS-box complex that targets CArG motifs in the ANT promoter to repress its expression; ANT otherwise represses PIN1, so ANT repression permits PIN1-dependent auxin flux to reach an optimal auxin threshold that promotes AUX/IAA degradation, releases ARFs, and enables activation of MMC developmental programs (Cavalleri et al., 2025). Green arrows, activation; red bars, repression.
Figure 5.
Key SPL/NZZ-mediated positive regulators (primarily) players behind single-MMC speciation. SPL/NZZ recruits the co-repressor TPL/TPR (via its C-terminal EAR motif) to CIN-like TCP–bound loci and, together with HDA19, represses TCP downstream genes, thereby restricting MMC fate and supporting normal single-MMC development (Wei et al., 2015). SPL/NZZ also functions in a STK/SEP3-associated MADS-box complex that targets CArG motifs in the ANT promoter to repress its expression; ANT otherwise represses PIN1, so ANT repression permits PIN1-dependent auxin flux to reach an optimal auxin threshold that promotes AUX/IAA degradation, releases ARFs, and enables activation of MMC developmental programs (Cavalleri et al., 2025). Green arrows, activation; red bars, repression.
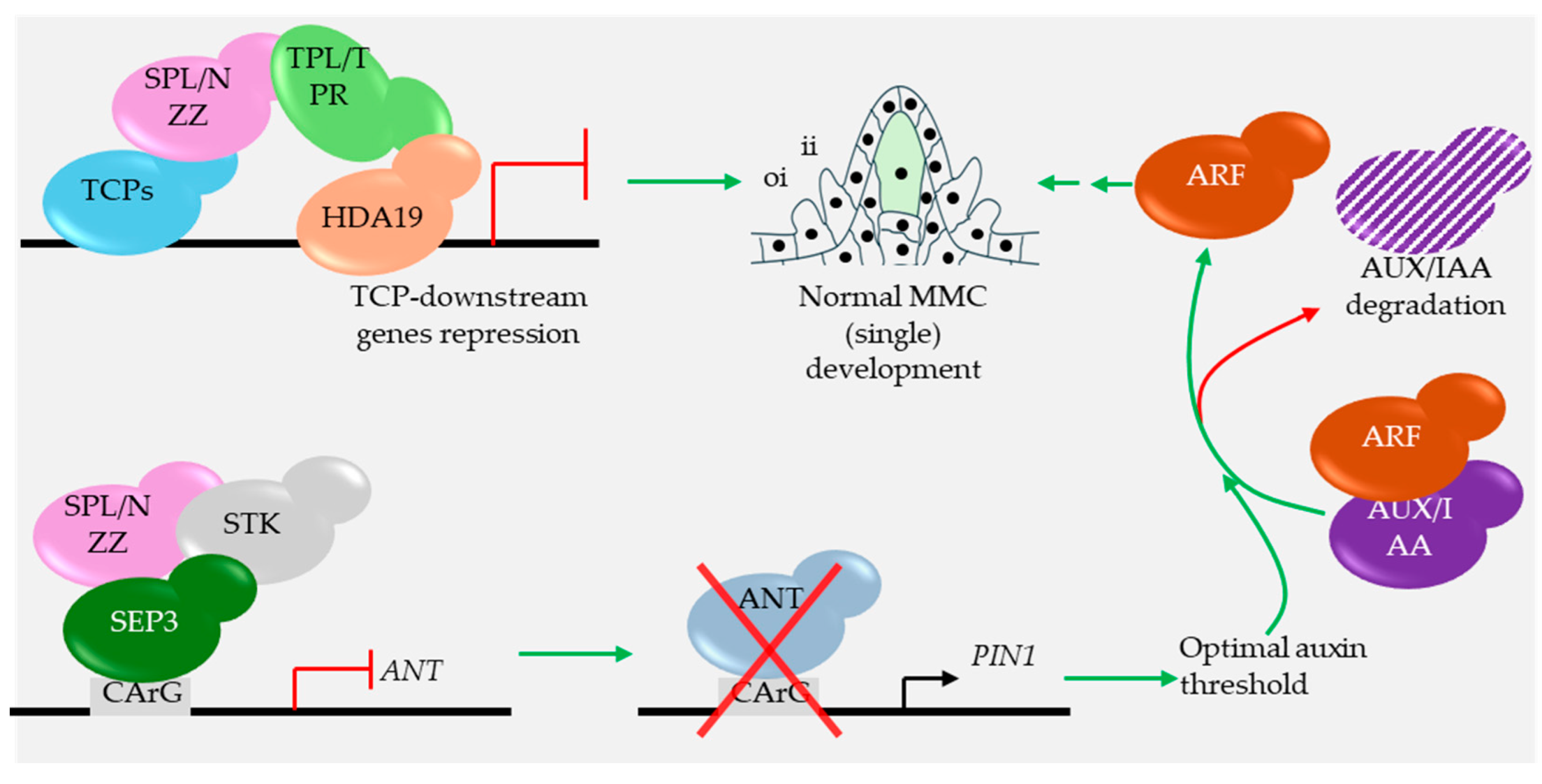
Figure 6.
Factors associated with the hypodermal somatic cells, inner integument, and outer integument development. Left (hypodermal somatic cells): the SWR1 chromatin-remodeling complex, via ARP6, promotes H2A.Z deposition around the WRKY28 transcription start site (TSS), enhancing its expression and thereby repressing ectopic acquisition of MMC fate. The process is non-cell-autonomously promoted by inner-integument KLU activity (Zhao et al., 2018a; Aslam et al., 2019). Middle (inner integument): KLU promotes cell proliferation and contributes to final seed size (Adamski et al., 2009), while jasmonate (JA) signaling through COI1/SCFCOI1 and the 26S proteasome (via JAZ–MYC control) counteracts this proliferative program (Zhang et al., 2025). Right (outer integument): a GIF1/GRF growth module is integrated with JA-responsive regulators (including MYC–TPL and PPD1/2–KIX8/9 acting at G-box elements), with proteasome-linked turnover shaping proliferation/elongation outputs (Zhang et al., 2025). Green arrows indicate activation; red bars indicate repression; dashed connectors indicate indirect and/or non-cell-autonomous effects.
Figure 6.
Factors associated with the hypodermal somatic cells, inner integument, and outer integument development. Left (hypodermal somatic cells): the SWR1 chromatin-remodeling complex, via ARP6, promotes H2A.Z deposition around the WRKY28 transcription start site (TSS), enhancing its expression and thereby repressing ectopic acquisition of MMC fate. The process is non-cell-autonomously promoted by inner-integument KLU activity (Zhao et al., 2018a; Aslam et al., 2019). Middle (inner integument): KLU promotes cell proliferation and contributes to final seed size (Adamski et al., 2009), while jasmonate (JA) signaling through COI1/SCFCOI1 and the 26S proteasome (via JAZ–MYC control) counteracts this proliferative program (Zhang et al., 2025). Right (outer integument): a GIF1/GRF growth module is integrated with JA-responsive regulators (including MYC–TPL and PPD1/2–KIX8/9 acting at G-box elements), with proteasome-linked turnover shaping proliferation/elongation outputs (Zhang et al., 2025). Green arrows indicate activation; red bars indicate repression; dashed connectors indicate indirect and/or non-cell-autonomous effects.
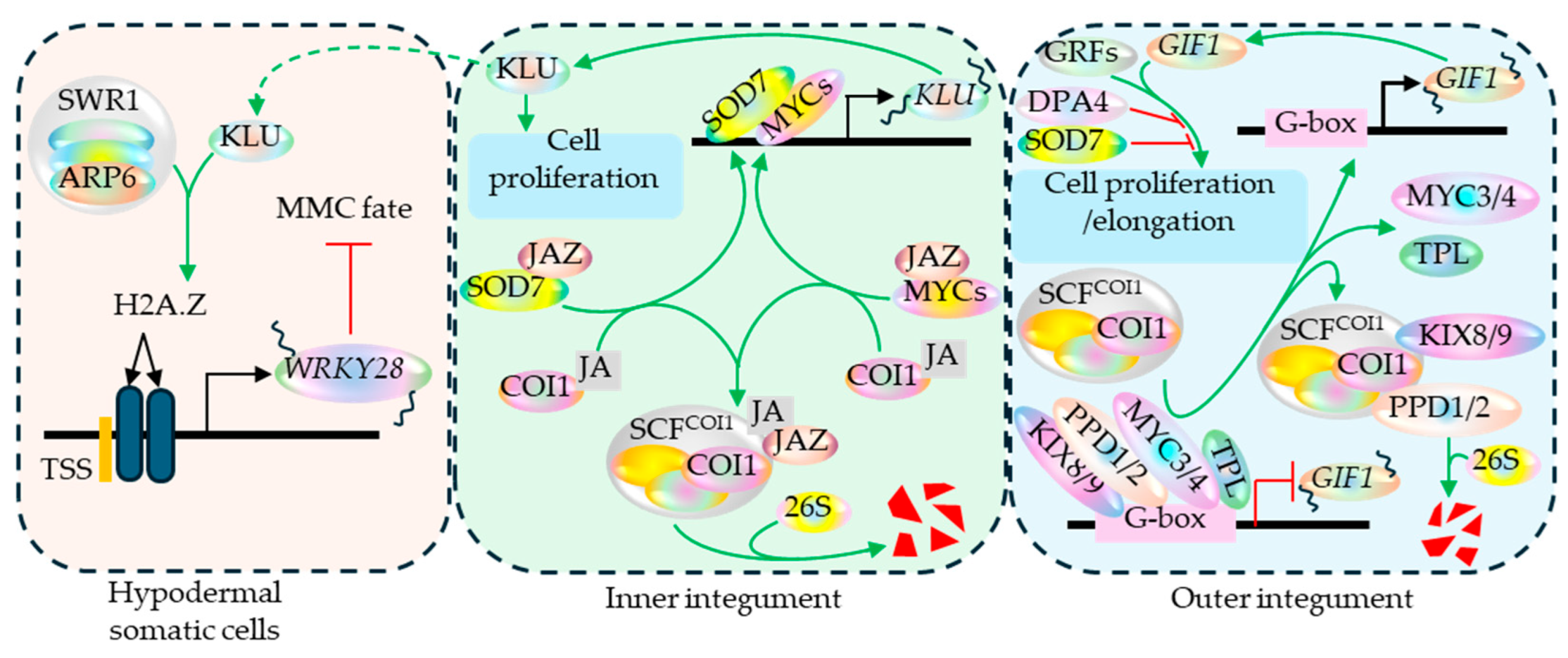
Figure 7.
Factors regulating the growth-progression halt and post-fertilization proliferation of central cell (secondary endosperm) in the mature ovule and underlying events in Arabidopsis. In the unfertilized FM, RBR1 restricts G1/S by repressing E2F/DP-driven S-phase genes, enforcing the mature-ovule growth halt; loss of RBR1 causes failure of arrest with ectopic central-cell proliferation and autonomous endosperm-like divisions (Ebel et al., 2004). Central-cell CYCD7;1–CDKA;1 promotes RBR1 phosphorylation/inactivation, thereby sliding it off and releasing E2F/DP to drive extra nuclear proliferation when ectopically expressed (Sornay et al., 2015). Normally, sperm cell-derived (paternal) CYCD7;1 and CDKA facilitates the RBR1 phosphorylation process after fertilization. RBR1–MSI1 represses MET1, enabling DME-linked demethylation and maternal activation of MET1-sensitive imprinted genes (MEA, FIS2, FWA) (Jullien et al., 2008; Johnston et al., 2008; Xiao et al., 2003). After fertilization, maternal MEA-PRC2 contributes to the H3K27me3-mediated repression of the paternal RBR and MEA alleles (Gehring et al., 2006; Johnston et al., 2008). FIS–PRC2 prevents fertilization-independent activation of an endosperm-like program before fertilization; after fertilization, AGL62 supports the coenocytic program and delays cellularization (Figueiredo et al., 2016).
Figure 7.
Factors regulating the growth-progression halt and post-fertilization proliferation of central cell (secondary endosperm) in the mature ovule and underlying events in Arabidopsis. In the unfertilized FM, RBR1 restricts G1/S by repressing E2F/DP-driven S-phase genes, enforcing the mature-ovule growth halt; loss of RBR1 causes failure of arrest with ectopic central-cell proliferation and autonomous endosperm-like divisions (Ebel et al., 2004). Central-cell CYCD7;1–CDKA;1 promotes RBR1 phosphorylation/inactivation, thereby sliding it off and releasing E2F/DP to drive extra nuclear proliferation when ectopically expressed (Sornay et al., 2015). Normally, sperm cell-derived (paternal) CYCD7;1 and CDKA facilitates the RBR1 phosphorylation process after fertilization. RBR1–MSI1 represses MET1, enabling DME-linked demethylation and maternal activation of MET1-sensitive imprinted genes (MEA, FIS2, FWA) (Jullien et al., 2008; Johnston et al., 2008; Xiao et al., 2003). After fertilization, maternal MEA-PRC2 contributes to the H3K27me3-mediated repression of the paternal RBR and MEA alleles (Gehring et al., 2006; Johnston et al., 2008). FIS–PRC2 prevents fertilization-independent activation of an endosperm-like program before fertilization; after fertilization, AGL62 supports the coenocytic program and delays cellularization (Figueiredo et al., 2016).
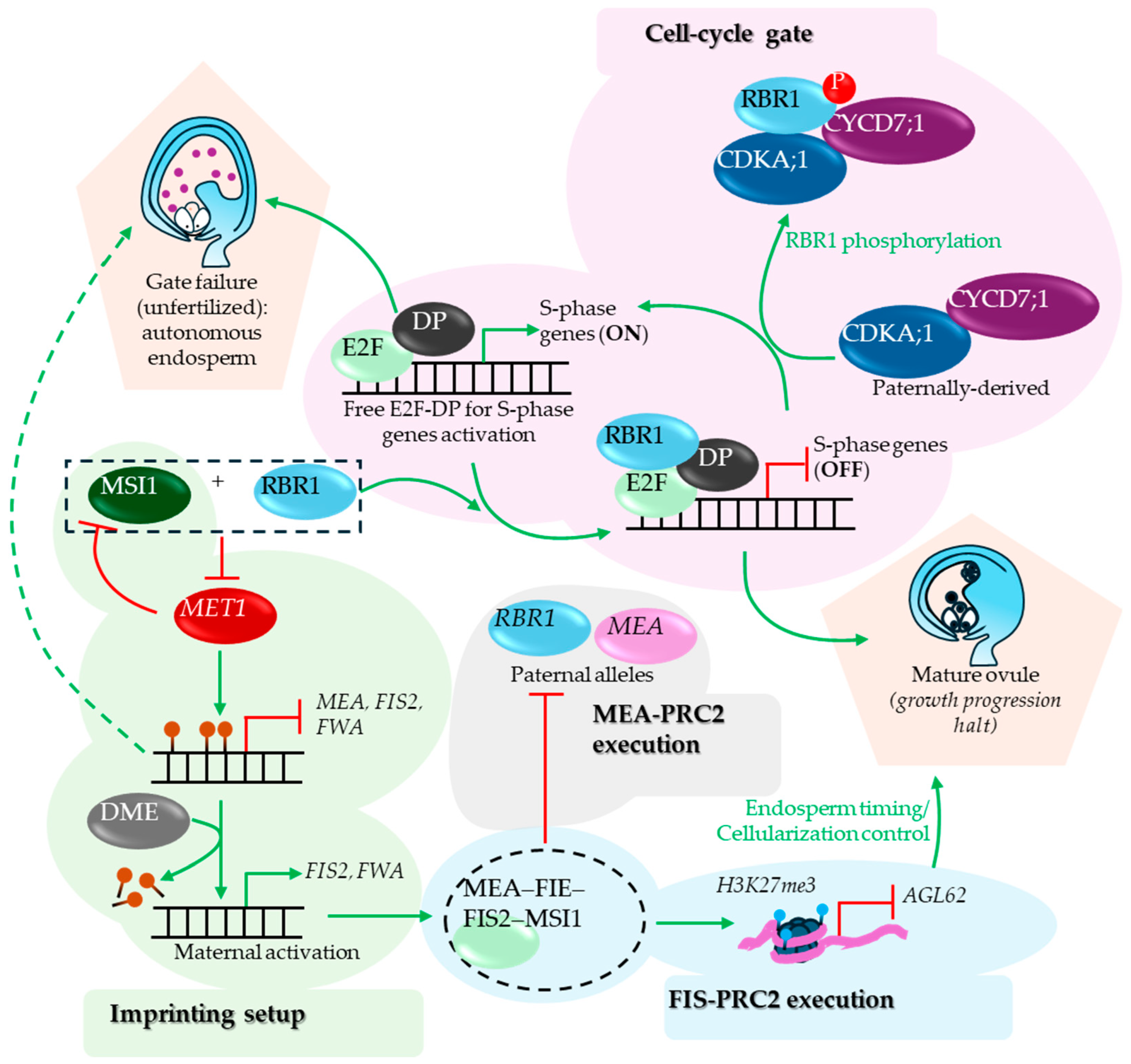
Figure 8.
Schematic representation of putative PRC2-mediated H3K27me3 marks deposition and associated mechanism. The histone methylation marks are recognized and bound by TFL2/LHP1 (Turck et al., 2007) and bridges PRC2 to the complex by interacting with MSI1 and facilitating the spread of histone methylation toward the gene’s 3′-ends in a context-dependent manner (Veluchamy et al., 2016). TFL2/LHP1 additionally interacts with DEK proteins (Nakamura et al., 2026), which, as suggested by the observations made in the animal reconstituted systems, interacts with H2A/H2B, induces kink in the nucleosome linker DNA and promotes chromatin compaction (Alexiadis et al., 2000, Kujirai et al., 2025).
Figure 8.
Schematic representation of putative PRC2-mediated H3K27me3 marks deposition and associated mechanism. The histone methylation marks are recognized and bound by TFL2/LHP1 (Turck et al., 2007) and bridges PRC2 to the complex by interacting with MSI1 and facilitating the spread of histone methylation toward the gene’s 3′-ends in a context-dependent manner (Veluchamy et al., 2016). TFL2/LHP1 additionally interacts with DEK proteins (Nakamura et al., 2026), which, as suggested by the observations made in the animal reconstituted systems, interacts with H2A/H2B, induces kink in the nucleosome linker DNA and promotes chromatin compaction (Alexiadis et al., 2000, Kujirai et al., 2025).
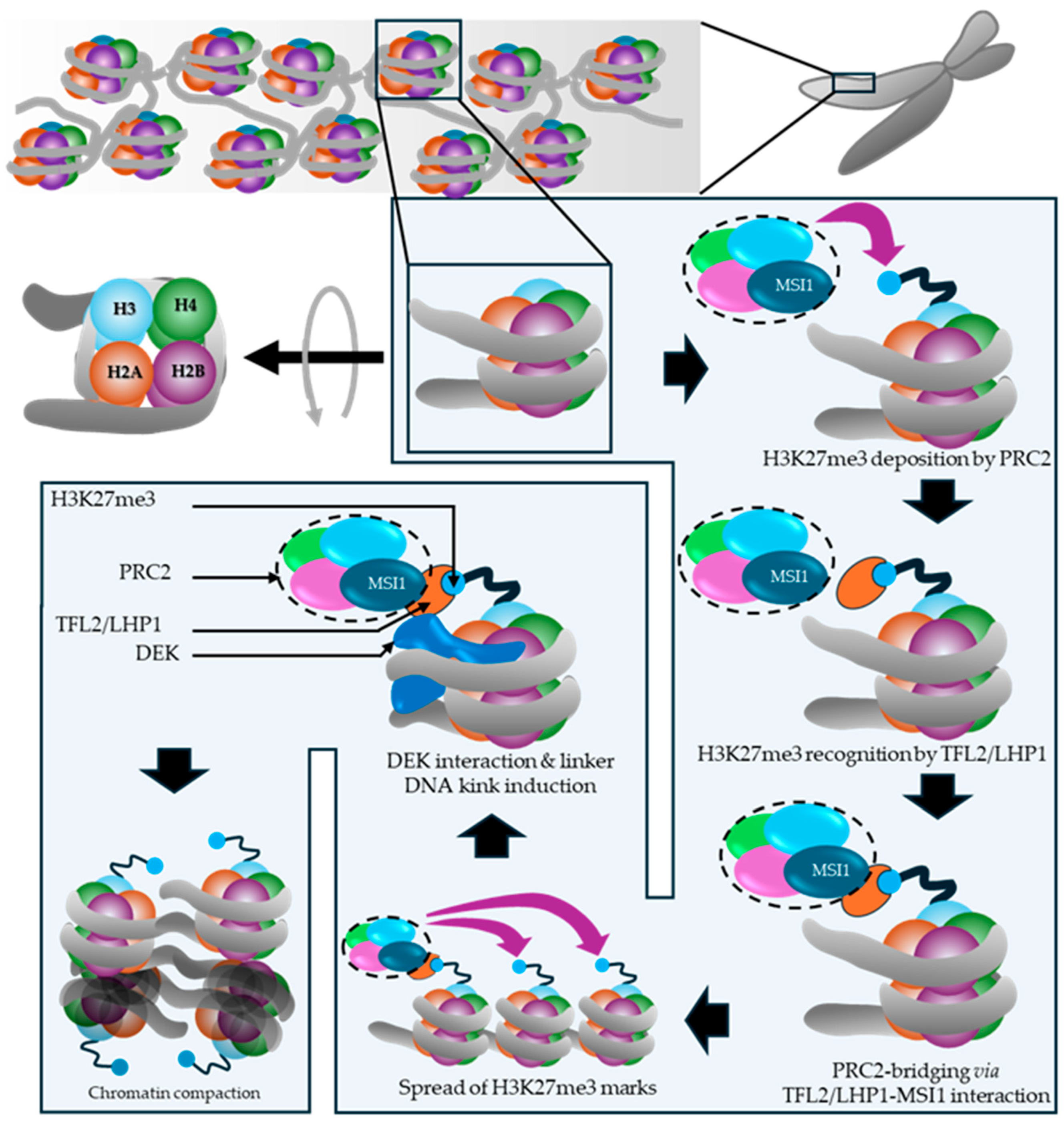
Figure 9.
Factors enforcing developmental halt in egg cell. In the mature FM, The Polycomb components MSI1 and FIE are proposed to interface with RBR1-mediated cell-cycle gate during embryo patterning (Guitton and Berger , 2005). Furthermore, the MEA-PRC2 complex deposits histone methylation marks at CYCD1;1 locus thereby potentially affecting the RBR1 phosphorylation and G1/S-phase transition (Simonini et al., 2021). The key factors associated with the post-maturity developmental halt in the egg cell are those associated with the G2/M-phase transition (Simonini et al., 2021). Likely factors include the Cyclin B member conferring repressive effect to the M-phase progression genes and maternal APC4 at its inactive state awaiting activation via yet unidentified paternal factor (Wang et al., 2012). Additionally, the embryogenesis trigger BABY BOOM (BBM) is kept OFF in egg cells by egg-cell repression mechanisms including its 3’-bound AtRKD5 (Liu et al., 2024). Green arrows indicate positive regulation; red bars indicate repressive effect; dashed connections indicate proposed/indirect links.
Figure 9.
Factors enforcing developmental halt in egg cell. In the mature FM, The Polycomb components MSI1 and FIE are proposed to interface with RBR1-mediated cell-cycle gate during embryo patterning (Guitton and Berger , 2005). Furthermore, the MEA-PRC2 complex deposits histone methylation marks at CYCD1;1 locus thereby potentially affecting the RBR1 phosphorylation and G1/S-phase transition (Simonini et al., 2021). The key factors associated with the post-maturity developmental halt in the egg cell are those associated with the G2/M-phase transition (Simonini et al., 2021). Likely factors include the Cyclin B member conferring repressive effect to the M-phase progression genes and maternal APC4 at its inactive state awaiting activation via yet unidentified paternal factor (Wang et al., 2012). Additionally, the embryogenesis trigger BABY BOOM (BBM) is kept OFF in egg cells by egg-cell repression mechanisms including its 3’-bound AtRKD5 (Liu et al., 2024). Green arrows indicate positive regulation; red bars indicate repressive effect; dashed connections indicate proposed/indirect links.
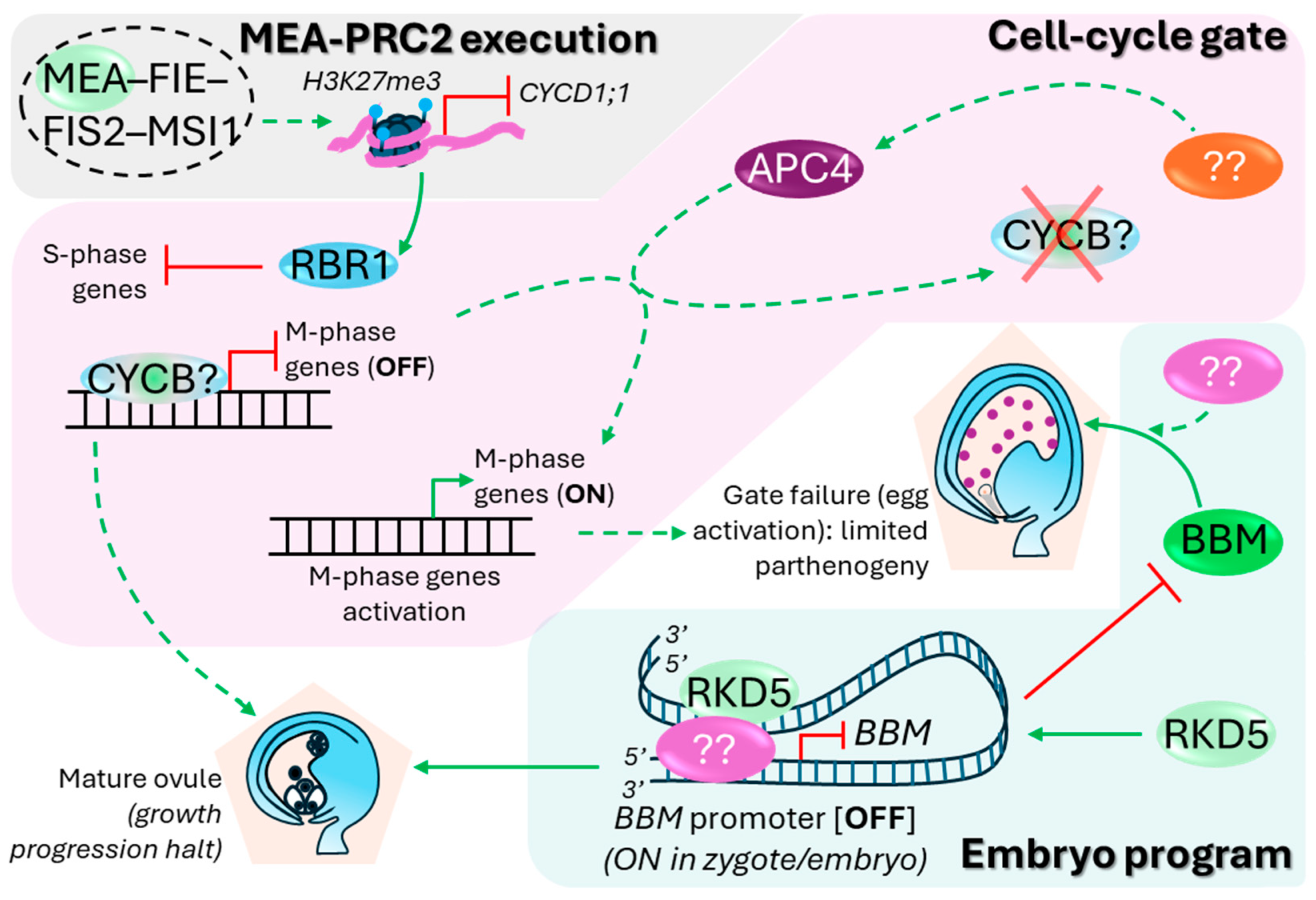
Figure 10.
Key developmental stages of early seed development. Mature ovule exhibits burst of the synergid cell receiving the pollen tube. The ovule initiates seed coat initiation (brown region at the chalaza) irrespective of the successful syngamy (Liu et al., 2019). The ovule with failed syngamy enlarges before its subsequent abortion (Kasahara et al., 2016). The successfully fertilized ovule on the other hand, initiates the coenocytic endosperm proliferation. It follows the persistent synergid removal and suspensor-supported embryo development until the embryonic heart stage, when the endosperm cellularization initiates in a wave fashion starting at the micropylar region.
Figure 10.
Key developmental stages of early seed development. Mature ovule exhibits burst of the synergid cell receiving the pollen tube. The ovule initiates seed coat initiation (brown region at the chalaza) irrespective of the successful syngamy (Liu et al., 2019). The ovule with failed syngamy enlarges before its subsequent abortion (Kasahara et al., 2016). The successfully fertilized ovule on the other hand, initiates the coenocytic endosperm proliferation. It follows the persistent synergid removal and suspensor-supported embryo development until the embryonic heart stage, when the endosperm cellularization initiates in a wave fashion starting at the micropylar region.
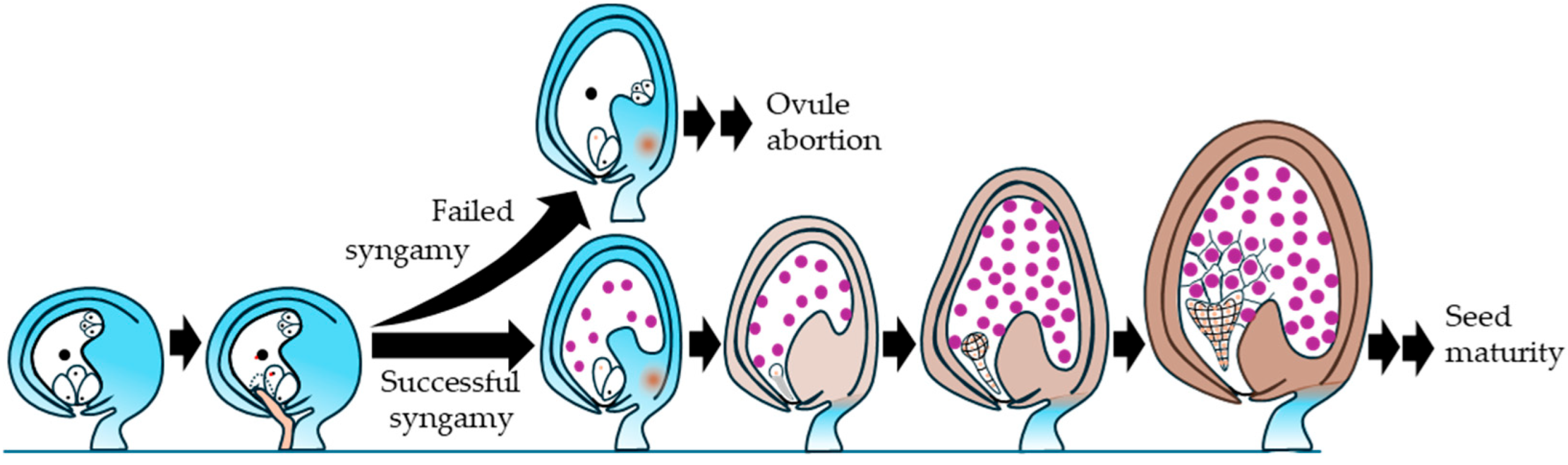
Figure 11.
Key factors associated with the post-fertilization seed development. Fertilization reduces DME expression via post-TSS regulatory elements (putative enhancer X and repressor Y are shown) (Park et al., 2017), altering MET1/DME-associated methylation status and contributing to paternal MEA silencing (Choi et al., 2002; Xiao et al., 2003). Reduced FIS-PRC2 constraint permits increased AGL62 (Hehenberger et al., 2012) (positively supported by LEC1 (Lee et al., 2003, Song et al., 2025)) which, together with PHE1/AGL80, promotes paternally imprinted programs including auxin biosynthetic genes YUC10 and TAR1 via repression of ATHB factors (Roszak and Köhler, 2011, Kradolfer et al., 2013, Batista et al., 2019, Guo et al., 2022, Pankaj et al., 2024). Auxin homeostasis includes GH3.5-mediated conjugation (prevalent in unfertilized ovule) and ILR1-linked deconjugation (in fertilized ovule) (Aloni et al., 2006; Rampey et al., 2004; Larsson et al., 2017; Vanneste et al., 2025). Endosperm auxin export is proposed via PGP10/TWD1 (Wu et al., 2010, Figueiredo et al., 2016, Tsering et al., 2025) and integument distribution via PIN3/AUX1 (Larsson et al., 2017, Liu et al., 2023), antagonizing sporophytic EMF/VRN-PRC2 to enable GA and TT pathway-associated genes thereby contributing to the seed coat initiation and seed development (Roszak and Köhler, 2011, Figueiredo and Köhler, 2018). The TT-pathway associated TT16 additionally represses the suppressor of GA-mediated PCD thereby facilitating the nucellus degeneration process (Xu et al., 2016). The integument-exported auxin enhances the sucrose transport and conversion-associated genes, SWEET10 and INV4 respectively, in addition to the downregulation and upregulation of the callose biosynthesis (CALS5/6/7) and callose degradation (BG_PPAP) associated genes respectively at the chalaza. Recent reports propose that endosperm-derived auxin enhances chalazal sucrose unloading and T6P accumulation, potentially linking carbohydrate status to TOR activation and downstream GA-associated responses during early seed development; these connections remain provisional and warrant genetic/temporal validation (Liu et al., 2025a, Xu et al., 2025). Green arrows, activation/transport; red bars, repression; dashed lines, putative/indirect links.
Figure 11.
Key factors associated with the post-fertilization seed development. Fertilization reduces DME expression via post-TSS regulatory elements (putative enhancer X and repressor Y are shown) (Park et al., 2017), altering MET1/DME-associated methylation status and contributing to paternal MEA silencing (Choi et al., 2002; Xiao et al., 2003). Reduced FIS-PRC2 constraint permits increased AGL62 (Hehenberger et al., 2012) (positively supported by LEC1 (Lee et al., 2003, Song et al., 2025)) which, together with PHE1/AGL80, promotes paternally imprinted programs including auxin biosynthetic genes YUC10 and TAR1 via repression of ATHB factors (Roszak and Köhler, 2011, Kradolfer et al., 2013, Batista et al., 2019, Guo et al., 2022, Pankaj et al., 2024). Auxin homeostasis includes GH3.5-mediated conjugation (prevalent in unfertilized ovule) and ILR1-linked deconjugation (in fertilized ovule) (Aloni et al., 2006; Rampey et al., 2004; Larsson et al., 2017; Vanneste et al., 2025). Endosperm auxin export is proposed via PGP10/TWD1 (Wu et al., 2010, Figueiredo et al., 2016, Tsering et al., 2025) and integument distribution via PIN3/AUX1 (Larsson et al., 2017, Liu et al., 2023), antagonizing sporophytic EMF/VRN-PRC2 to enable GA and TT pathway-associated genes thereby contributing to the seed coat initiation and seed development (Roszak and Köhler, 2011, Figueiredo and Köhler, 2018). The TT-pathway associated TT16 additionally represses the suppressor of GA-mediated PCD thereby facilitating the nucellus degeneration process (Xu et al., 2016). The integument-exported auxin enhances the sucrose transport and conversion-associated genes, SWEET10 and INV4 respectively, in addition to the downregulation and upregulation of the callose biosynthesis (CALS5/6/7) and callose degradation (BG_PPAP) associated genes respectively at the chalaza. Recent reports propose that endosperm-derived auxin enhances chalazal sucrose unloading and T6P accumulation, potentially linking carbohydrate status to TOR activation and downstream GA-associated responses during early seed development; these connections remain provisional and warrant genetic/temporal validation (Liu et al., 2025a, Xu et al., 2025). Green arrows, activation/transport; red bars, repression; dashed lines, putative/indirect links.
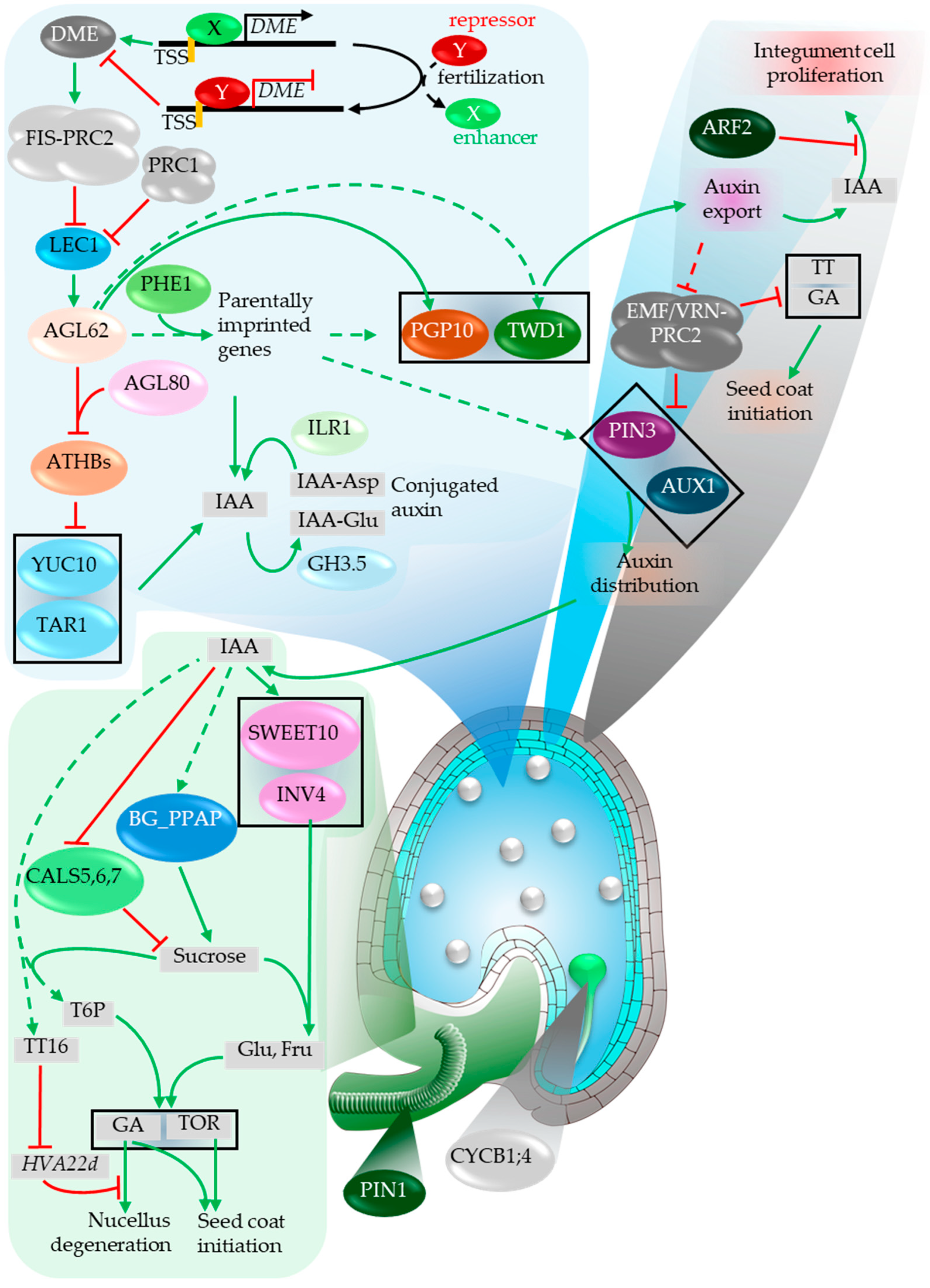
Figure 12.
Key factors associated with the endosperm cellularization and seed coat development. During the coenocytic phase, the endosperm generates a paternally biased expansion force (PEF) that promotes cavity growth, while maternal programs impose a maternal restrictive force (MRF) to constrain final seed size (inset). Rising endosperm turgor pressure is proposed to act as a mechanical cue; once a threshold is reached, maternal pathways reduce turgor and trigger cellularization (Weinhofer et al., 2010; Beauzamy et al., 2016; Butel et al., 2024). ABA signaling from the chalazal domain integrates into this transition: TFL1 transport (via RAN) stabilizes ABI5, which represses the pro-proliferative SHB1–MIN3–IKU2 module (Zhang et al., 2020; Cheng et al., 2014; Kang et al., 2013). In the testa, chalazal PA accumulation (via BAN) and GA/BR modules are shown as key outputs and regulators of cell division/elongation and coat differentiation (Debeaujon et al., 2003; Figueiredo et al., 2016; Hu et al., 2018). Mechanosensitive ELA1/2-mediated GA deactivation promotes wall stiffening and restricts late growth, while BR signaling (BRI1/BAK1 → BES1/BZR1) can support seed coat activation via chromatin remodeling (Creff et al., 2015; Creff et al., 2023; Jiang et al., 2013; Pankaj et al., 2024b). Green arrows, activation; red bars, repression; dashed lines, indirect/putative links.
Figure 12.
Key factors associated with the endosperm cellularization and seed coat development. During the coenocytic phase, the endosperm generates a paternally biased expansion force (PEF) that promotes cavity growth, while maternal programs impose a maternal restrictive force (MRF) to constrain final seed size (inset). Rising endosperm turgor pressure is proposed to act as a mechanical cue; once a threshold is reached, maternal pathways reduce turgor and trigger cellularization (Weinhofer et al., 2010; Beauzamy et al., 2016; Butel et al., 2024). ABA signaling from the chalazal domain integrates into this transition: TFL1 transport (via RAN) stabilizes ABI5, which represses the pro-proliferative SHB1–MIN3–IKU2 module (Zhang et al., 2020; Cheng et al., 2014; Kang et al., 2013). In the testa, chalazal PA accumulation (via BAN) and GA/BR modules are shown as key outputs and regulators of cell division/elongation and coat differentiation (Debeaujon et al., 2003; Figueiredo et al., 2016; Hu et al., 2018). Mechanosensitive ELA1/2-mediated GA deactivation promotes wall stiffening and restricts late growth, while BR signaling (BRI1/BAK1 → BES1/BZR1) can support seed coat activation via chromatin remodeling (Creff et al., 2015; Creff et al., 2023; Jiang et al., 2013; Pankaj et al., 2024b). Green arrows, activation; red bars, repression; dashed lines, indirect/putative links.
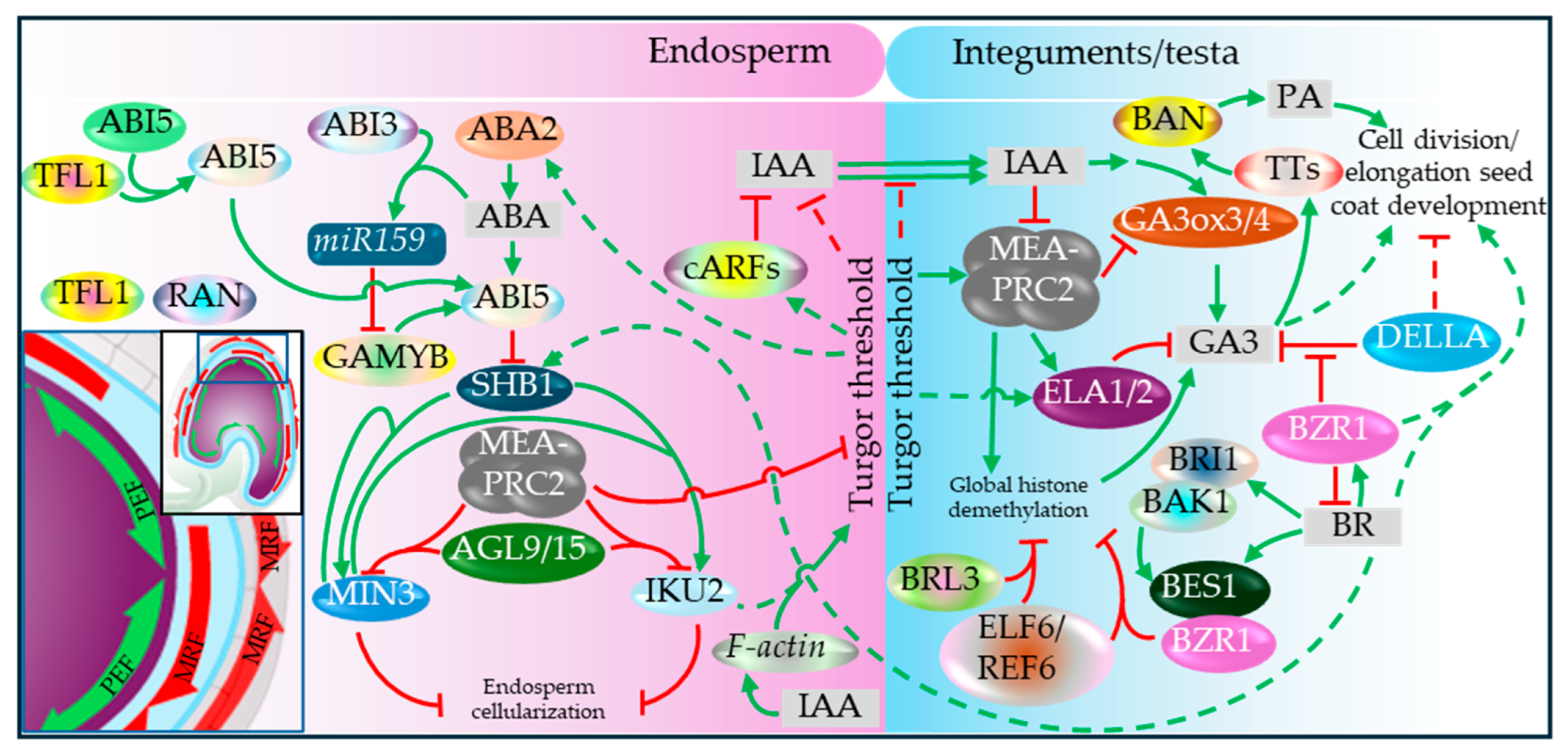
Figure 13.
Schematic representation of putative mechanosensing in the fertilized ovule. The very initial seed enlargement is largely contributed by the outer integument, in which FER contributes to the process (Yu et al., 2014). The expanding volume (cavity) of the coenocytic endosperm is perceived by the cell wall property change at the inner wall of the outer integument (OI) L1 layer. The FER-complex has been proposed to interact with the cell-wall freed RALF peptide and trigger actin dynamics, Ca2+ influx as well as RBOH-mediated ROS accumulation thereby contributing to the integument elongation and division. The actin dynamics is proposed to be regulated by IKU2 as well. The stiffening of the inner cell wall of the inner integument L2 layer and seed expansion limit is mediated by the ER-localized ELA1/2 via the deactivation of bioactive GA3 (Creff et al., 2015, Creff et al., 2023), which likely plays role during the final stage of the seed maturation.
Figure 13.
Schematic representation of putative mechanosensing in the fertilized ovule. The very initial seed enlargement is largely contributed by the outer integument, in which FER contributes to the process (Yu et al., 2014). The expanding volume (cavity) of the coenocytic endosperm is perceived by the cell wall property change at the inner wall of the outer integument (OI) L1 layer. The FER-complex has been proposed to interact with the cell-wall freed RALF peptide and trigger actin dynamics, Ca2+ influx as well as RBOH-mediated ROS accumulation thereby contributing to the integument elongation and division. The actin dynamics is proposed to be regulated by IKU2 as well. The stiffening of the inner cell wall of the inner integument L2 layer and seed expansion limit is mediated by the ER-localized ELA1/2 via the deactivation of bioactive GA3 (Creff et al., 2015, Creff et al., 2023), which likely plays role during the final stage of the seed maturation.
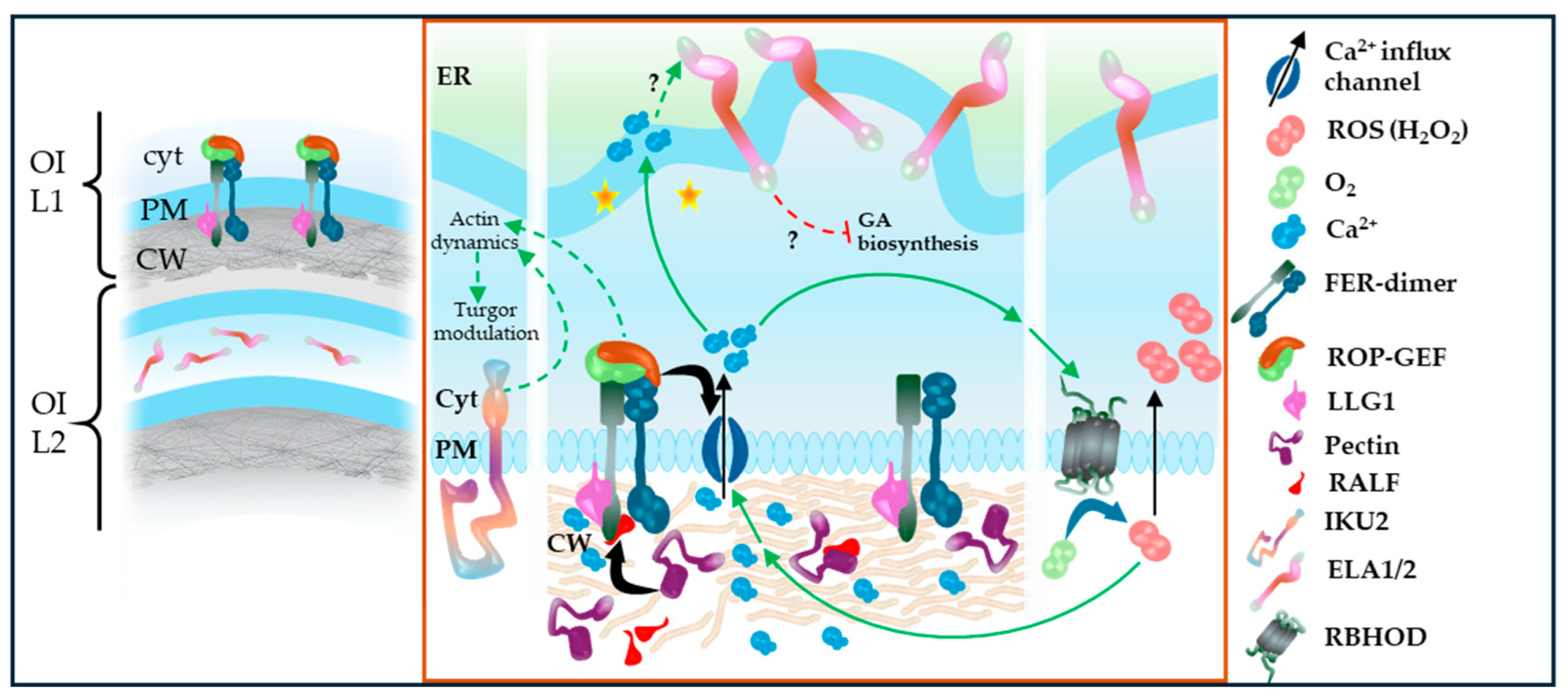
Figure 14.
Early stages of sexual embryo development in angiosperm. Immediately after the egg cell (EC) and sperm cell (SC) fusion, the fresh zygotic cell contracts briefly before returning to the initial state (Antoine et al., 2000). Fertilization triggers Ca2+ influx (Denninger et al., 2014, Hamamura et al., 2014) and ECS1/2 proteins secretion. ECS1/2 actively contributing to the degradation of LURE1 peptide thereby hindering potential polytubey cases (Yu et al., 2021). The zygote exhibits the expression of maternal genome up until it reaches to the elongated zygotic stage when it shows the expression of mixed genome of the zygote (Zhao et al., 2019).
Figure 14.
Early stages of sexual embryo development in angiosperm. Immediately after the egg cell (EC) and sperm cell (SC) fusion, the fresh zygotic cell contracts briefly before returning to the initial state (Antoine et al., 2000). Fertilization triggers Ca2+ influx (Denninger et al., 2014, Hamamura et al., 2014) and ECS1/2 proteins secretion. ECS1/2 actively contributing to the degradation of LURE1 peptide thereby hindering potential polytubey cases (Yu et al., 2021). The zygote exhibits the expression of maternal genome up until it reaches to the elongated zygotic stage when it shows the expression of mixed genome of the zygote (Zhao et al., 2019).
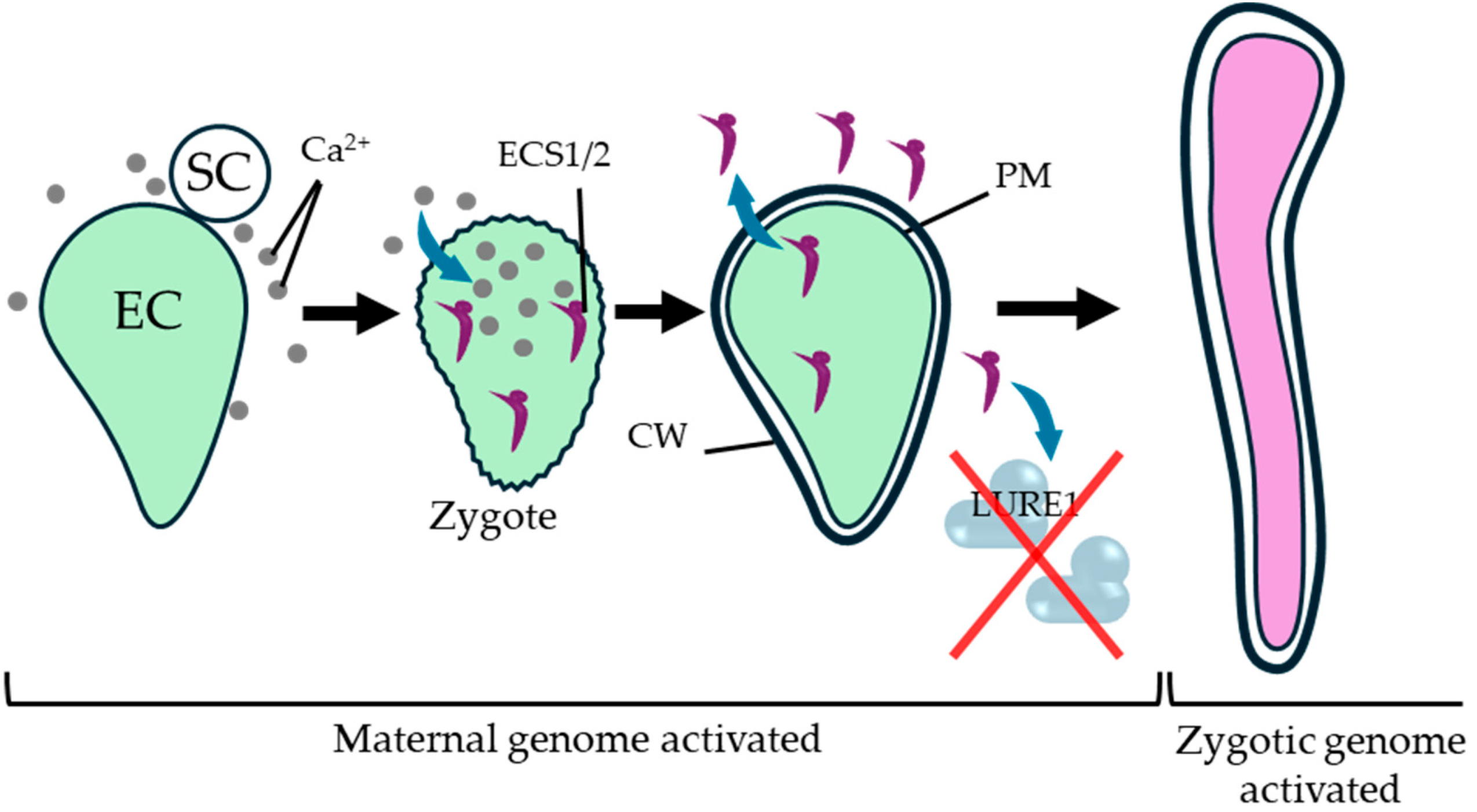
Figure 15.
Factors associated with the maternal and paternal genome interaction in a zygote. WOX8/9 plays a central role in the very initial asymmetric division and subsequent embryo patterning. Its expression niche is controlled by the secretory CLAVATA peptides CLE8/19. Normal WOX8 expression relies on the maternal (HDG11/12) and paternal (SSP-dependent YDA-MAPK) cascades. Sperm cell-derived SSP mRNA is translated in the zygote, which in turn activates YDA. AGB1 acts as a scaffold for the components of the MAPK complex by directly interacting with the associated components (Yuan et al., 2017), YDA forms complex with MPK3/6 and MKK4/5 thereby potentiating the complex for the phosphorylation of WRKY2 and SCRM, which directly contribute to WOX8 activation (Wang et al., 2007, Zhang et al., 2017). The activation is additionally facilitated by the Cam1 and AGB1-interaction-activated ZAR1 (Yu et al., 2016) as well as the endosperm-derived maternally imprinted member ESF1 (Costa et al., 2014). The egg cell constitutes normal RKD2 expression, which hinders normal hypophysis cell division in the embryo. The process is alleviated by the sperm cell-derived TREE1 and DAZ3 (Cheng et al., 2024), which are postulated to be facilitated via the HDA19-mediated deacetylation of RKD2 locus. MEA contributes to the patterning process by regulating CYCD1;1 and itself in a dose-dependent manner thereby affecting auxin distribution and cell fate speciation in the developing embryo (Baroux et al., 2006, Simonini et al., 2021, Simonini et al., 2024). Promoting and suppressive effects are linked with green and red arrows respectively. The dotted arrows represent postulated/indirect links.
Figure 15.
Factors associated with the maternal and paternal genome interaction in a zygote. WOX8/9 plays a central role in the very initial asymmetric division and subsequent embryo patterning. Its expression niche is controlled by the secretory CLAVATA peptides CLE8/19. Normal WOX8 expression relies on the maternal (HDG11/12) and paternal (SSP-dependent YDA-MAPK) cascades. Sperm cell-derived SSP mRNA is translated in the zygote, which in turn activates YDA. AGB1 acts as a scaffold for the components of the MAPK complex by directly interacting with the associated components (Yuan et al., 2017), YDA forms complex with MPK3/6 and MKK4/5 thereby potentiating the complex for the phosphorylation of WRKY2 and SCRM, which directly contribute to WOX8 activation (Wang et al., 2007, Zhang et al., 2017). The activation is additionally facilitated by the Cam1 and AGB1-interaction-activated ZAR1 (Yu et al., 2016) as well as the endosperm-derived maternally imprinted member ESF1 (Costa et al., 2014). The egg cell constitutes normal RKD2 expression, which hinders normal hypophysis cell division in the embryo. The process is alleviated by the sperm cell-derived TREE1 and DAZ3 (Cheng et al., 2024), which are postulated to be facilitated via the HDA19-mediated deacetylation of RKD2 locus. MEA contributes to the patterning process by regulating CYCD1;1 and itself in a dose-dependent manner thereby affecting auxin distribution and cell fate speciation in the developing embryo (Baroux et al., 2006, Simonini et al., 2021, Simonini et al., 2024). Promoting and suppressive effects are linked with green and red arrows respectively. The dotted arrows represent postulated/indirect links.
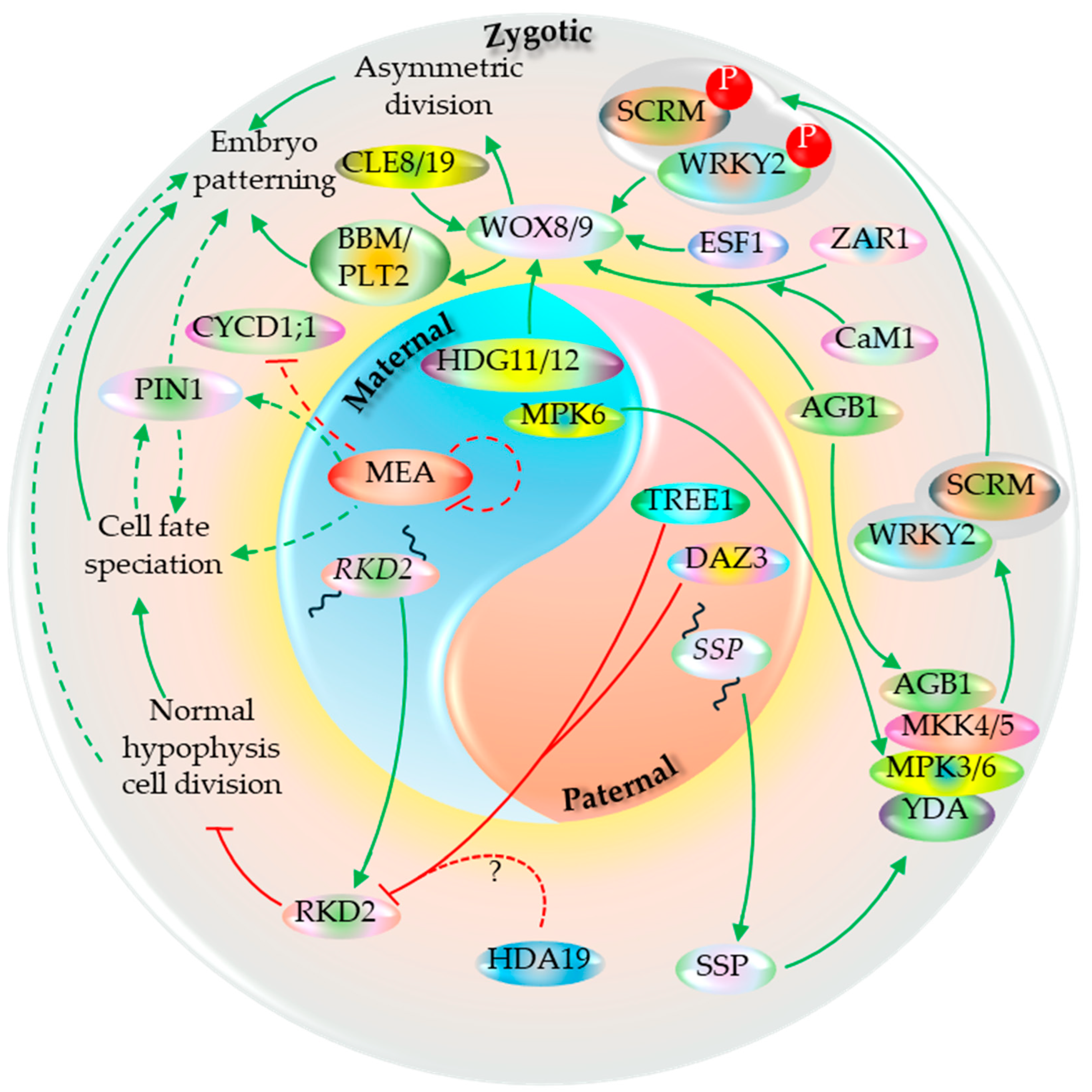
Figure 16.
Factors associated with the embryo-endosperm interaction. The endosperm-derived LEC1 moves to the embryo by its binucleate stage and contributes to the expression of key genes including itself (LEC2, ABI3, FUS3 regulating the embryo patterning and maturation processes during its later development stages (Song et al., 2021). At the globular stage, embryo cuticle development commences, which is mediated by the embryo membrane-localized GSO1/2. The signal depends on the endosperm-derived ALE1-mediated cleavage of the embryo-derived sulfated peptide TWS1 (at the apoplast), which act as ligand for GSO1/2 (Tanaka et al., 2001, Doll et al., 2020b). In the later developmental stage, after endosperm cellularization, the invasive embryonic growth in the endosperm and developmental PCD (dPCD) of the latter are actively supported by the endosperm-expressed ZOU. ZOU, along with its ortholog ICE1, regulates the PCD-associated genes encoding NAC domain harboring proteins (KIR1, ANAC087, ANAC046, ORE1) involved in controlling the genes such as PASPA3 and BFN1 directly involved in the nucleic acid degradation (Yang et al., 2008, Denay et al., 2014, Fourquin et al., 2016, Doll et al., 2023). The process is further contributed by the embryo membrane bound DEK1 via mechanism yet unclear. ZOU also facilitates the deposition of KRS to the developing embryo sheath (Moussu et al., 2017, Doll et al., 2020a). Putative KER-interacting LRR receptor kinase (GSO1/2 or other) contributing to ROS accumulation and dPCD facilitation has been shown. Potential role of FER-complex in regulating controlled ROS homeostasis and downstream dPCD has been postulated. Factors with known mode of action has been shown in shapes with accompanying legend while others are presented in unique spheres. Promoting and suppressive effects are linked with green and red arrows respectively. The dotted arrows represent postulated/indirect links.
Figure 16.
Factors associated with the embryo-endosperm interaction. The endosperm-derived LEC1 moves to the embryo by its binucleate stage and contributes to the expression of key genes including itself (LEC2, ABI3, FUS3 regulating the embryo patterning and maturation processes during its later development stages (Song et al., 2021). At the globular stage, embryo cuticle development commences, which is mediated by the embryo membrane-localized GSO1/2. The signal depends on the endosperm-derived ALE1-mediated cleavage of the embryo-derived sulfated peptide TWS1 (at the apoplast), which act as ligand for GSO1/2 (Tanaka et al., 2001, Doll et al., 2020b). In the later developmental stage, after endosperm cellularization, the invasive embryonic growth in the endosperm and developmental PCD (dPCD) of the latter are actively supported by the endosperm-expressed ZOU. ZOU, along with its ortholog ICE1, regulates the PCD-associated genes encoding NAC domain harboring proteins (KIR1, ANAC087, ANAC046, ORE1) involved in controlling the genes such as PASPA3 and BFN1 directly involved in the nucleic acid degradation (Yang et al., 2008, Denay et al., 2014, Fourquin et al., 2016, Doll et al., 2023). The process is further contributed by the embryo membrane bound DEK1 via mechanism yet unclear. ZOU also facilitates the deposition of KRS to the developing embryo sheath (Moussu et al., 2017, Doll et al., 2020a). Putative KER-interacting LRR receptor kinase (GSO1/2 or other) contributing to ROS accumulation and dPCD facilitation has been shown. Potential role of FER-complex in regulating controlled ROS homeostasis and downstream dPCD has been postulated. Factors with known mode of action has been shown in shapes with accompanying legend while others are presented in unique spheres. Promoting and suppressive effects are linked with green and red arrows respectively. The dotted arrows represent postulated/indirect links.
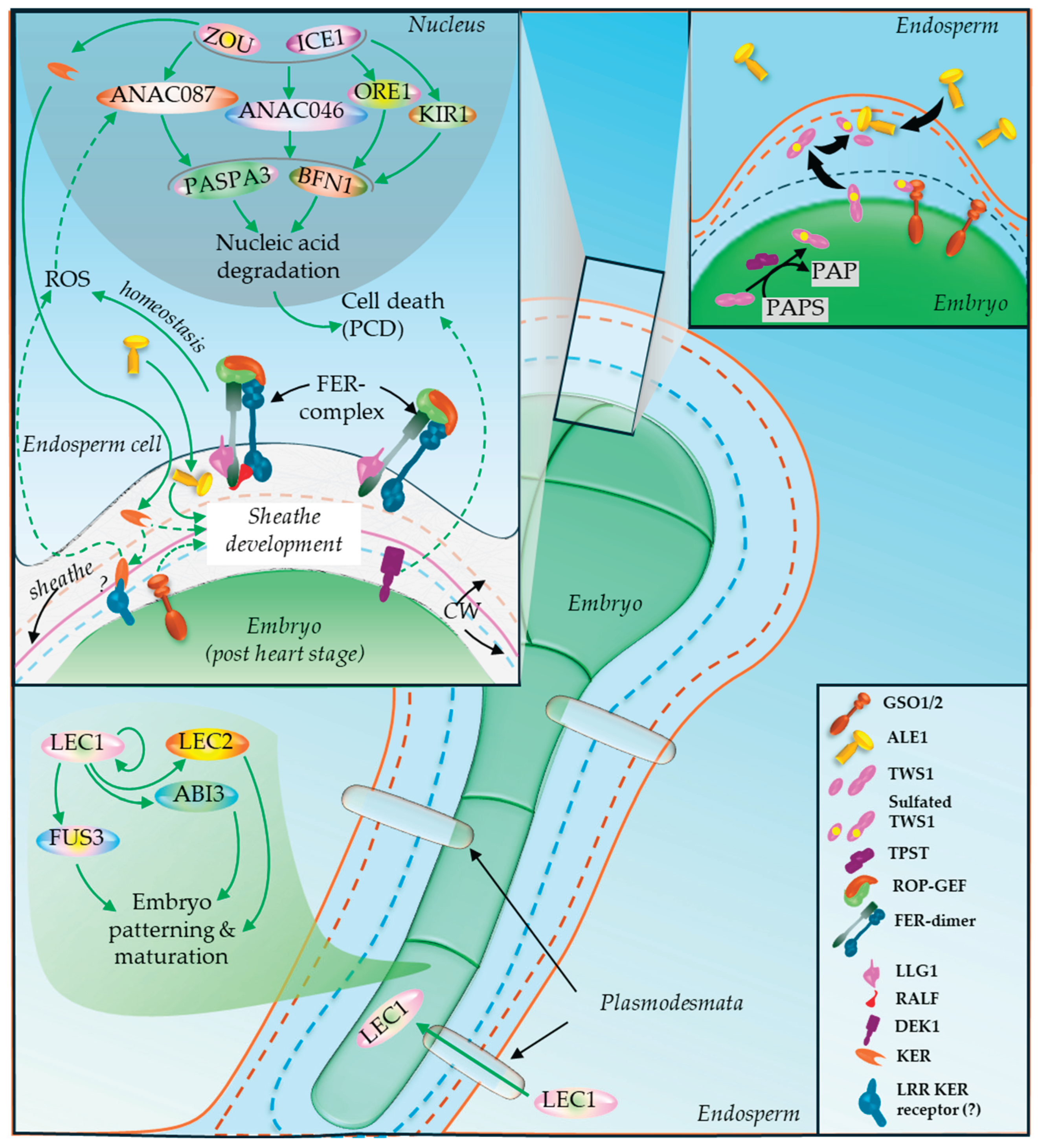
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



