Submitted:
22 February 2026
Posted:
25 February 2026
You are already at the latest version
Abstract
Light limitation constrains ecosystem function in large, shallow eutrophic lakes, yet the ecosystem-scale role of sediment resuspension is rarely isolated. We quantified wave- and bioturbation-driven resuspension effects on light attenuation in Utah Lake using repli-cated limnocorrals (~12 m diameter; 1.2–2.4 m depth) deployed over three growing seasons (2022–2024). Secchi depth (n = 363) and PAR profiles (7 depths; n = 126) were measured inside corrals and at paired open-lake controls. Mean Secchi depth was nearly twofold greater inside corrals (39.8 vs. 22.0 cm; p < 0.001), with lower light attenuation coefficients (k = 0.17 vs. 0.22). Ln(PAR) depth regressions consistently showed weaker attenuation under reduced disturbance, producing compensation depths 1.25× deeper inside corrals. Although near-surface PAR was sometimes higher in the open lake, PAR below 0.5 m was consistently greater inside corrals. Reduced attenuation was linked primarily to lower suspended solids and decreased resuspension of fine sediments and CaCO₃ precipitates; phytoplankton biomass further influenced late-summer attenuation. Sediment resuspension thus dominates underwater light climate, constrains benthic production, and reinforces phytoplankton-dominated pathways. Stabilizing sediments through invasive carp removal, macrophyte and mollusk recovery, and reduced dis-turbance should enhance light availability, benthic–pelagic coupling, and ecosystem function in turbid shallow lakes.

Keywords:
light attenuation
; sediment resuspension
; calcium carbonate precipitation
; shallow lakes
; photosynthetically active radiation
; benthic–pelagic coupling
; lake restoration
1. Introduction
Light penetration is fundamental to aquatic ecosystem functioning, regulating primary production by phytoplankton, benthic algae, and submerged macrophytes, and influencing predator–prey interactions and oxygen dynamics [1,2,3]. Reduced light availability suppresses photosynthesis, exacerbates hypoxia, and promotes cyanobacterial dominance [3,4].
Light attenuation, the exponential decline of irradiance with depth, is controlled by suspended particles, phytoplankton biomass [5], dissolved organic matter, and in some systems inorganic precipitates such as calcium carbonate (CaCO₃) [6,7,8,9,10,11,12]. Shallow eutrophic lakes typically exhibit high attenuation, but systems with abundant, easily resuspended CaCO₃ precipitates or other fine sediment materials may experience particularly strong light limitation [10,13].
Utah Lake exemplifies these conditions: chronic turbidity from wind driven wave action and invasive fish bioturbation limits light penetration across much of the basin, inhibiting benthic algae and macrophytes and reinforcing phytoplankton dominance [14,15,16,17]. Recent satellite analyses indicate that algal blooms in Utah Lake are primarily light-limited, with 94% of intense blooms (chlorophyll-a > 87 µg L⁻¹) occurring only when turbidity is < 120 nephelometric turbidity units (NTU)[18].
Wave action and benthic fish bioturbation are critical but under-quantified drivers of sediment resuspension. Mobilization of unconsolidated sediments increases turbidity and alters the underwater light climate. Understanding how physical disturbance regulates light attenuation is essential for guiding restoration and management.
We present results from limnocorral experiments designed to isolate the effects of wave disturbance and bioturbation on light attenuation in Utah Lake. We quantify and discuss implications for ecosystem function and restoration by comparing Secchi depth and PAR profiles inside corrals with open-lake controls, changes in light availability and key physical and chemical covariates.
2. Materials and Methods
The limnocorrals we used in this study were constructed of heavy-grade skirt material fitted with twelve foam-filled floats arranged in a cylindrical dodecagon (~12 m diameter; Figure 1; Supporting Information Figures S1–S2). We deployed the corrals in the northwest corner of Utah Lake (Figure 2) in spring (May–June) and removed in October during 2022–2024. We deployed ten corrals in 2022, twelve in 2023, and nine in 2024 at depths ranging from 1.2 to 2.4 m depending on lake level.
The limnocorrals partially isolated the water column from sediment resuspension by reducing wave energy and benthic bioturbation relative to open-lake conditions. This design allowed us to isolate and evaluate ecological effects of wave-driven and bioturbation-induced resuspension in Utah Lake and to assess the consequences of reducing these processes. By approximating conditions expected under restoration actions such as carp removal and sediment stabilization, the corrals provided a mechanistic basis for evaluating how reductions in resuspension can improve light availability and ecosystem function.
We measured Secchi depth between 18 May 2022 and 3 October 2023 inside (n = 218) and outside (n = 145) the corrals (Table 1). In 2024, we replaced Secchi measurements by direct PAR measurements using a Li-COR LI-1500 datalogger equipped with dual LI-192 underwater quantum sensors. We recorded PAR measurements at depths of 2, 24, 67, 87, 107, 127, and 147 cm inside corrals and at paired open-lake control sites (Table S1).
We converted Secchi depth measurements to light attenuation coefficients (k) using the nonlinear model of Padial and Thomaz [19]:
k = 2.00 ∙ SD-0.76
where k is the attenuation coefficient and SD is Secchi depth (cm).
We performed linear regression of ln-transformed PAR (µmol photons m⁻² s⁻¹) versus depth (cm) separately for corral and open-lake sites which follows the Beer–Lambert model [3,20]. We used a theoretical photosynthetic minimum of 0.01 µmol photons m⁻² s⁻¹ to estimate compensation depths [21,22,23].
The chemical and biological covariates we considered included phytoplankton biovolume, chlorophyll a, phycocyanin, conductivity, oxidation–reduction potential, total suspended solids, volatile suspended solids, total dissolved solids, and chemical oxygen demand. We used generalized linear regression models and mixed-effects regression models to statistically analyze the data. We performed these analyses in Stata/IC 16.1 [24].
No generative artificial intelligence (GenAI) has been used in this paper.
3. Results
Over the study period, Secchi depth was greater inside than outside corrals (mean 39.8 cm vs. 22.0 cm; t = 5.98, p < 0.001; Figure 3), ranging from 12–165 cm inside and 8–66 cm outside. Secchi-derived light attenuation coefficients (k) likewise differed between treatments (mean k = 0.17 inside vs. 0.22 outside; t = −7.07, p < 0.001; Figure 3), spanning 0.04–0.30 inside and 0.08–0.41 outside.
Seasonally, Secchi-derived attenuation inside and outside the corrals differed most strongly in early summer, with higher attenuation outside corrals in May–June (Figure 4). From July through October, attenuation increased at both corral and open-lake sites coincident with higher plankton densities, but attenuation remained lower inside corrals in mid- to late summer and was significantly lower in August (Figure 4).
In 2024, PAR profiles showed stronger attenuation outside corrals than inside (Figure 5). Predicted minimum-light threshold depths were 101 cm (95% CI: 99–104) inside corrals and 81 cm (95% CI: 77–85) outside, corresponding to a 1.25× increase inside corrals (95% CI for ratio: 1.22–1.29).
Across the 2024 sampling dates, near-surface PAR was often higher in the open lake, whereas PAR at deeper depths was consistently higher inside corrals (Supporting Information Figure S3; Table S2). We attribute this to near surface phytoplankton biomass inside the corrals, with less suspended solids in the water column compared to outside the corrals. Regression slopes of ln(PAR) versus depth varied among dates and generally indicated weaker attenuation inside corrals, except during a late-August bloom when phytoplankton biomass was elevated inside the corrals (Figure S1). Accordingly, the depth at which PAR reached the theoretical minimum occurred deeper inside corrals on most dates, with exceptions only when phytoplankton biomass was substantially higher inside corrals.
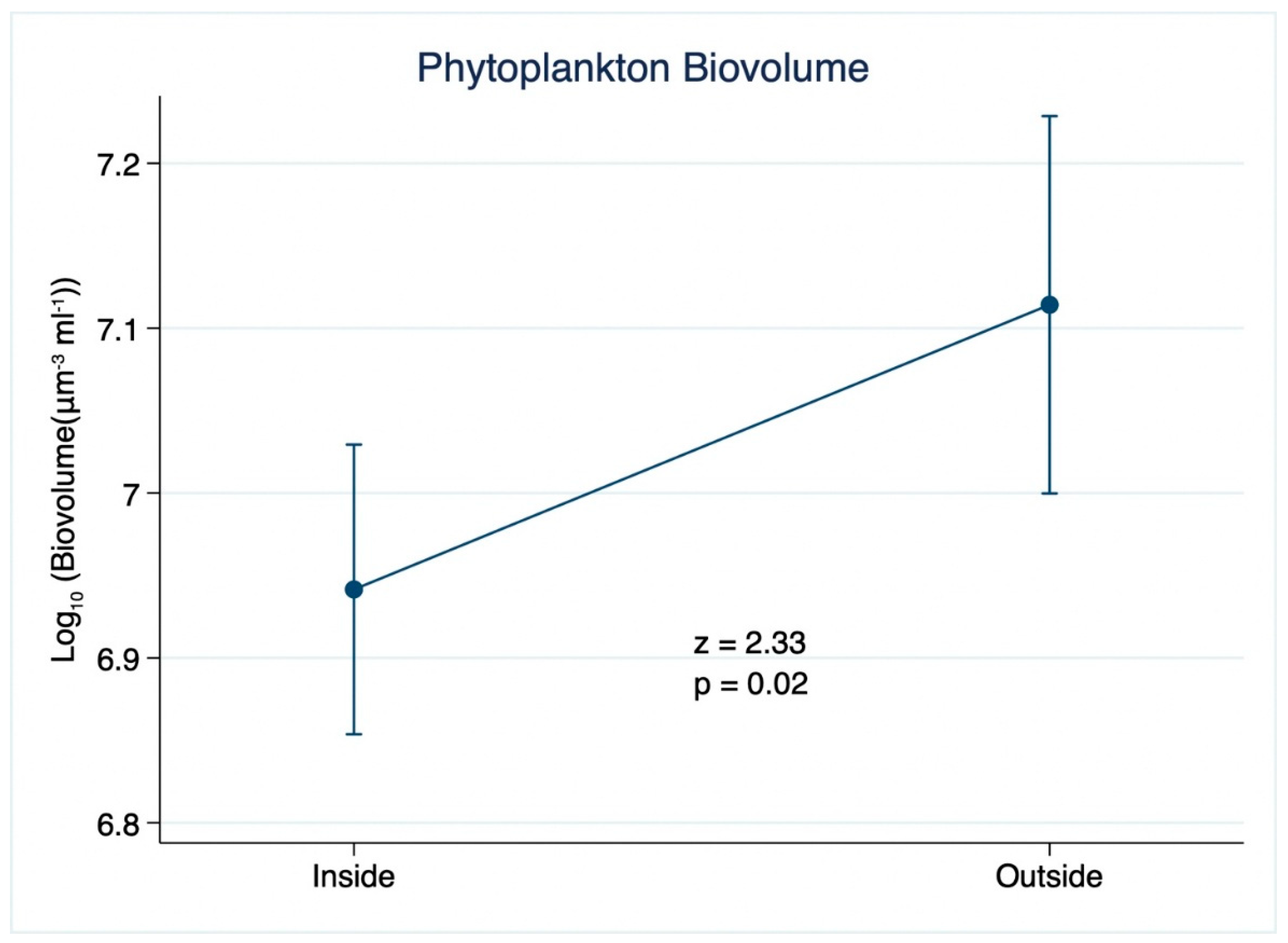
Phytoplankton biovolume (log₁₀ µm³ mL⁻¹) was significantly lower inside corrals (GLM: z = 2.23, p = 0.02; Figure 6; Table S3). Mixed-effects models indicated that relationships between PAR and covariates were depth-dependent (Table S4): Chlorophyll a and phycocyanin were associated with reduced PAR at the two shallowest depths but not deeper, consistent with pigment distributions concentrated near the surface. Total suspended solids reduced PAR across all depths, whereas volatile suspended solids, oxidation–reduction potential, and conductivity were most strongly associated with mid-depth PAR.
4. Discussion
Resuspension of fine sediments together with elevated phytoplankton biomass, emerged as the dominant controls on light attenuation in Utah Lake and represent a critical barrier to ecosystem recovery. By reducing wave energy and abiotic and biotic physical disturbance, the mesocosms promoted particulate settling, reduced resuspension, reduced optical backscatter, and increased subsurface light availability. These conditions are prerequisites for littoral zone benthic primary production and for re-establishing benthic–pelagic coupling in shallow lake ecosystems such as Utah Lake. However, under certain conditions these can also increase suspended phytoplankton biomass.
Phytoplankton biomass contributed to light attenuation through mechanisms that differed fundamentally from those of inorganic suspended solids. Whereas resuspended fine sediments primarily increase light attenuation via scattering and backscatter, phytoplankton blooms reduce light penetration mainly through absorption and self-shading. Prior work has demonstrated that high chlorophyll a concentrations can strongly attenuate PAR, with large-celled taxa such as the dinoflagellate Ceratium hirundinella capable of attenuating nearly 90% of incident radiation [25]. This species frequently dominates phytoplankton assemblages in Utah Lake and, together with other phytoplankton taxa particularly cyanobacteria, likely plays a substantial role in regulating underwater light conditions during blooms [26]. In addition, microbiologically mediated CaCO₃ precipitation may further elevate turbidity and light scattering, representing an understudied but potentially important process requiring targeted investigation [25]. While these processes contribute significantly to light limitation in Utah Lake, suspended sediments are the dominant process controlling light limitation outside of active blooms.
Interannual differences in lake level, with 2022–2023 characterized by low water that generally had higher suspended solids concentrations and 2024 by high water conditions which generally had lower concentrations both from dilution and increased depth limiting wave resuspension, likely influenced resuspension dynamics, particle residence time, and optical properties of the water column, complicating year-to-year comparisons. Although the corrals reduced turbation, we neither intended or achieved complete isolation from wave energy and bioturbation. Rather, our experimental design sought to approximate conditions expected under realistic restoration scenarios, including reduced bioturbation following carp removal and increased sediment stabilization by native macrophytes, benthic algae, and bivalve mollusks [26].
Chronic turbidity has cascading consequences for food-web structure and ecosystem function. Reduced light availability limits benthic primary production, favors phytoplankton-dominated energy pathways, and impairs visual foraging by higher trophic levels. Persistent light limitation in Utah Lake likely contributes to the low trophic transfer efficiencies documented in recent ecosystem models comparing food webs inside and outside limnocorrals [26]. These dynamics reinforce a degraded, phytoplankton-dominated state in Utah Lake characterized by inefficient energy transfer and accumulation of unconsumed primary production as detrital material.
In large, shallow lake ecosystems, turbidity is a well-established driver of light limitation and regime persistence [27,28,29,30]. Resuspension not only increases light attenuation but also enhances the release of sediment-bound nutrients into the water column, further stimulating phytoplankton growth ([31,32,33]. However, high concentrations of suspended solids, including CaCO₃ precipitates, can simultaneously constrain phytoplankton productivity by limiting light availability and altering nutrient uptake dynamics [34]. These opposing effects underscore the importance of sediment stabilization for restoring balanced primary production pathways.
The broader ecological implications of sustained turbidity are substantial. Resuspension impairs visual predators, reduces foraging efficiency of piscivorous fishes, and indirectly favors planktivorous fishes by providing visual refuge, thereby intensifying grazing pressure on zooplankton [35]. This process weakens top-down control of phytoplankton and further entrenches turbid conditions. Given that sediment resuspension in Utah Lake can persist for days following storm events, the cumulative effects on light climate and food-web interactions are likely profound.
Historical evidence suggests that Utah Lake was substantially less turbid prior to Euro-American settlement, with deeper light penetration and a more extensive littoral zone supported by abundant macrophytes, mollusks, and benthic algae ([36,37,38].. The transition to a phytoplankton-dominated system, combined with invasive bioturbators and altered hydrologic regimes, has fundamentally reshaped sediment dynamics and ecosystem functioning.
Given the pronounced spatial and temporal variability in PAR, long-term, high-resolution light monitoring is essential for evaluating restoration trajectories and quantifying functional recovery. Future work should explicitly couple optical dynamics with food-web and biogeochemical models to assess feedbacks among sediment stabilization, light availability, primary production pathways, and trophic transfer. Such integrative approaches will be critical for designing effective, science-based restoration strategies capable of shifting Utah Lake from a turbid, phytoplankton-dominated state toward a more resilient, benthic-supported ecosystem.
5. Conclusions
Our results emphasize that improvements in light availability will require basin-scale, integrated restoration strategies rather than localized interventions. Stabilizing sediments and retaining fine particles within benthic layers would promote benthic algal and macrophyte growth, enhance habitat complexity, and strengthen trophic transfer. Accordingly, restoration efforts should prioritize reestablishing sediment-stabilizing native vegetation and bivalves, control of bioturbating fishes, and water-level management that minimizes resuspension.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
David C. Richards: Conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, supervision, visualization, writing of original draft preparation and review and editing. Richard Mickelsen: Conceptualization, funding acquisition, resources, methodology, review and editing. Gustavious P. Williams: Conceptualization, funding acquisition, methodology, supervision, review and editing. Brett Marshall: Conceptualization, funding acquisition, methodology, supervision, review and editing. Sam Rushforth: Conceptualization, funding acquisition, methodology, data curation, review and editing. Sarah Rushforth: Conceptualization, funding acquisition, methodology, data curation, review and editing.
Funding
Funding from this study was provided by the Utah Department of Forestry, Fires, and State Lands.
Data Availability Statement
Pertinent data can be downloaded at: “Data for Sediment Resuspension Controls Light Attenuation and Ecosystem Function in a Large, Shallow, Eutrophic Lake”. Author: David C. Richards. Excel file name: Light attenuation Limnocorral Manuscript Data.xlsx DOI: 10.5281/zenodo.18263381
Acknowledgments
A hearty acknowledgement to all the team members of the Brigham Young University, Timpanogos Special Service District, River Continuum Concepts, and Wasatch Front Water Quality that designed and installed limnocorrals and collected data. None of this research would be possible without these dedicated citizens working to better understand and help restore Utah Lake. The authors have reviewed and edited the output and take full responsibility for the content of this publication.”
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Karlsson, J.; Byström, P.; Ask, J.; Ask, P.; Persson, L.; Jansson, M. Light limitation of nutrient-poor lake ecosystems. Nature 2009, 460, 506–509. [CrossRef]
- Kemp, W.M.; Boynton, W.R.; Adolf, J.E.; Boesch, D.F.; Boicourt, W.C.; Brush, G.; Cornwell, J.C.; et al. Eutrophication of Chesapeake Bay: historical trends and ecological interactions. Mar. Ecol. Prog. Ser. 2005, 303, 1–29. [CrossRef]
- Weiskerger, C.J.; Rowe, M.D.; Stow, C.A.; Stuart, D.; Johengen, T. Application of the Beer–Lambert model to attenuation of photosynthetically active radiation in a shallow, eutrophic lake. Water Resour. Res. 2018, 54, 8952–8962. [CrossRef]
- Medrano, E.A.; Uittenbogaard, R.E.; Pires, L.M.D.; van de Wiel, B.J.H.; Clercx, H.J.H. Coupling hydrodynamics and buoyancy regulation in Microcystis aeruginosa. Ecol. Model. 2013, 248, 41–56. [CrossRef]
- Whittington, J.; Sherman, B.; Green, D.; Oliver, R.L. Growth of Ceratium hirundinella in a subtropical reservoir. J. Plankton Res. 2000, 22, 1025–1045. [CrossRef]
- Babin, M.; Stramski, D.; Ferrari, G.M.; Claustre, H.; Bricaud, A.; Obolensky, G.; Hoepffner, N. Variations in light absorption coefficients in coastal waters. J. Geophys. Res. Oceans 2003, 108, 3211. [CrossRef]
- Bukata, R.P.; Jerome, J.H.; Kondratyev, K.Y.; Pozdnyakov, D.V. Optical Properties and Remote Sensing of Inland and Coastal Waters; CRC Press: Boca Raton, FL, USA, 1995.
- Effler, S.W.; Perkins, M.G.; O’Donnell, S.M.; Strait, C.M. Light attenuation in lakes. Inland Waters 2010, 1, 1–13.
- Kirk, J.T.O. Light and Photosynthesis in Aquatic Ecosystems, 3rd ed.; Cambridge Univ. Press: Cambridge, UK, 2011. [CrossRef]
- Morris, D.P.; Zagarese, H.; Williamson, C.E.; et al. Attenuation of solar UV radiation in lakes. Limnol. Oceanogr.1995, 40, 1381–1391. [CrossRef]
- Plummer, L.N.; Busenberg, E. Solubilities of calcite and aragonite. Geochim. Cosmochim. Acta 1982, 46, 1011–1040. [CrossRef]
- Wetzel, R.G. Limnology: Lake and River Ecosystems, 3rd ed.; Academic Press: San Diego, CA, USA, 2001.
- Robbins, L.L.; Blackwelder, P.L. Biochemical evidence for origin of whitings. Geology 1992, 20, 464–468.
- Richards, D.C.; Marshall, B.; Rushforth, S.; Wellard, B. Utah Lake mesocosm study. Zenodo 2025. [CrossRef]
- Taggart, J.B.; Ryan, R.L.; Williams, G.P.; et al. Historical phosphorus mass in Utah Lake. Water 2024, 16, 933. [CrossRef]
- Valek, R.A.; Tanner, K.B.; Taggart, J.B.; et al. ICP-OES detectable elements in Utah Lake. Water 2024, 16, 2170. [CrossRef]
- King, L.; Brahney, J.; Daly, S.; et al. Primary production modeling. Freshw. Sci. 2023, 42, 44–57. [CrossRef]
- Tanner, K.B.; Cardall, A.C.; Taggart, J.B.; Williams, G.P. Earth Observation investigation of algal blooms. Remote Sens. 2026, 18, 394. [CrossRef]
- Padial, A.A.; Thomaz, S.M. Prediction of light attenuation coefficient. Limnology 2008, 9, 143–151. [CrossRef]
- Ingle, J.D.; Crouch, S.R. Spectrochemical Analysis; Prentice Hall: Englewood Cliffs, NJ, USA, 1988.
- Hancke, K.; et al. Low light requirement for algae growth. J. Geophys. Res. Oceans 2018, 123, 985–1000. [CrossRef]
- Randelhoff, A.; et al. Arctic phytoplankton growth. Sci. Adv. 2020, 6, eabc2678. [CrossRef]
- Raven, J.A.; Kübler, J.E.; Beardall, J. Put out the light. J. Mar. Biol. Assoc. UK 2000, 80, 1–25. [CrossRef]
- StataCorp. Stata/IC 16.1 for Mac; StataCorp LLC: College Station, TX, USA, 2023.
- Mortensen, B.M.; Haber, M.J.; DeJong, J.T.; Caslake, L.F.; Nelson, D.C. Environmental factors on CaCO₃ precipitation. J. Appl. Microbiol. 2011, 111, 338–349. [CrossRef]
- Richards, D.C.; Marshall, B.; Rushforth, S.; Mickelsen, R.; Williams, G.; Miller, T. Effects of reduced turbation on food web function. Zenodo 2025. [CrossRef]
- Scheffer, M. Ecology of Shallow Lakes; Kluwer Academic: Dordrecht, Netherlands, 1998.
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [CrossRef]
- Edwards, K.F.; Thomas, M.K.; Klausmeier, C.A.; Litchman, E. Phytoplankton growth synthesis. Limnol. Oceanogr.2016, 61, 1232–1244. [CrossRef]
- Quinlan, R.; Filazzola, A.; Mahdiyan, O.; et al. Total phosphorus–chlorophyll relationships. Limnol. Oceanogr.2021, 66, 392–404. [CrossRef]
- Tammeorg, O.; Niemistö, J.; Möls, T.; et al. Wind-induced sediment resuspension. Aquat. Sci. 2013, 75, 559–570. [CrossRef]
- Tang, C.; Li, Y.; He, C.; Acharya, K. Sediment resuspension behavior. Sci. Total Environ. 2020, 708, 135131. [CrossRef]
- Zhang, S.; Yi, Q.; Buyang, S.; Cui, H.; Zhang, S. Bioavailable phosphorus enrichment. Sci. Total Environ. 2020, 710, 135672. [CrossRef]
- Schallenberg, M.; Burns, C.W. Sediment resuspension effects. Freshw. Biol. 2004, 49, 143–159. [CrossRef]
- Trochine, C.; Risholt, C.; Schou, M.; et al. Fish larvae diet in turbid lakes. Freshw. Biol. 2022, 67, 1601–1615. [CrossRef]
- Janetski, J.C. Utah Lake prehistory. Utah Hist. Q. 1990, 58, 5–31.
- Williams, G.P.; Nelson, S.; Rushforth, S.; et al. Human-driven trophic changes in Utah Lake. Water Air Soil Pollut.2023, 234, 218. [CrossRef]
- Richards, D.C.; Rushforth, S.; Marshall, B.; Williams, G.; Wellard, B. Utah Lake mesocosm study ecological section. Zenodo 2023. [CrossRef]
Figure 1.
General schematic of 10- meter diameter, 1.2 to 2.4- meter depth limnocorrals. Dodecagon connected floats attached to hollow black tubing provided buoyancy and stability for limnocorrals. Skirts were attached to floats. Sleeves were sown into bottom of skirts to house heavy chain. Sandbags were placed around the bottom of skirts. Sand and sediment were also pushed up against outside of skirts. Design was slightly modified each year to reduce damage from wind and waves.
Figure 1.
General schematic of 10- meter diameter, 1.2 to 2.4- meter depth limnocorrals. Dodecagon connected floats attached to hollow black tubing provided buoyancy and stability for limnocorrals. Skirts were attached to floats. Sleeves were sown into bottom of skirts to house heavy chain. Sandbags were placed around the bottom of skirts. Sand and sediment were also pushed up against outside of skirts. Design was slightly modified each year to reduce damage from wind and waves.
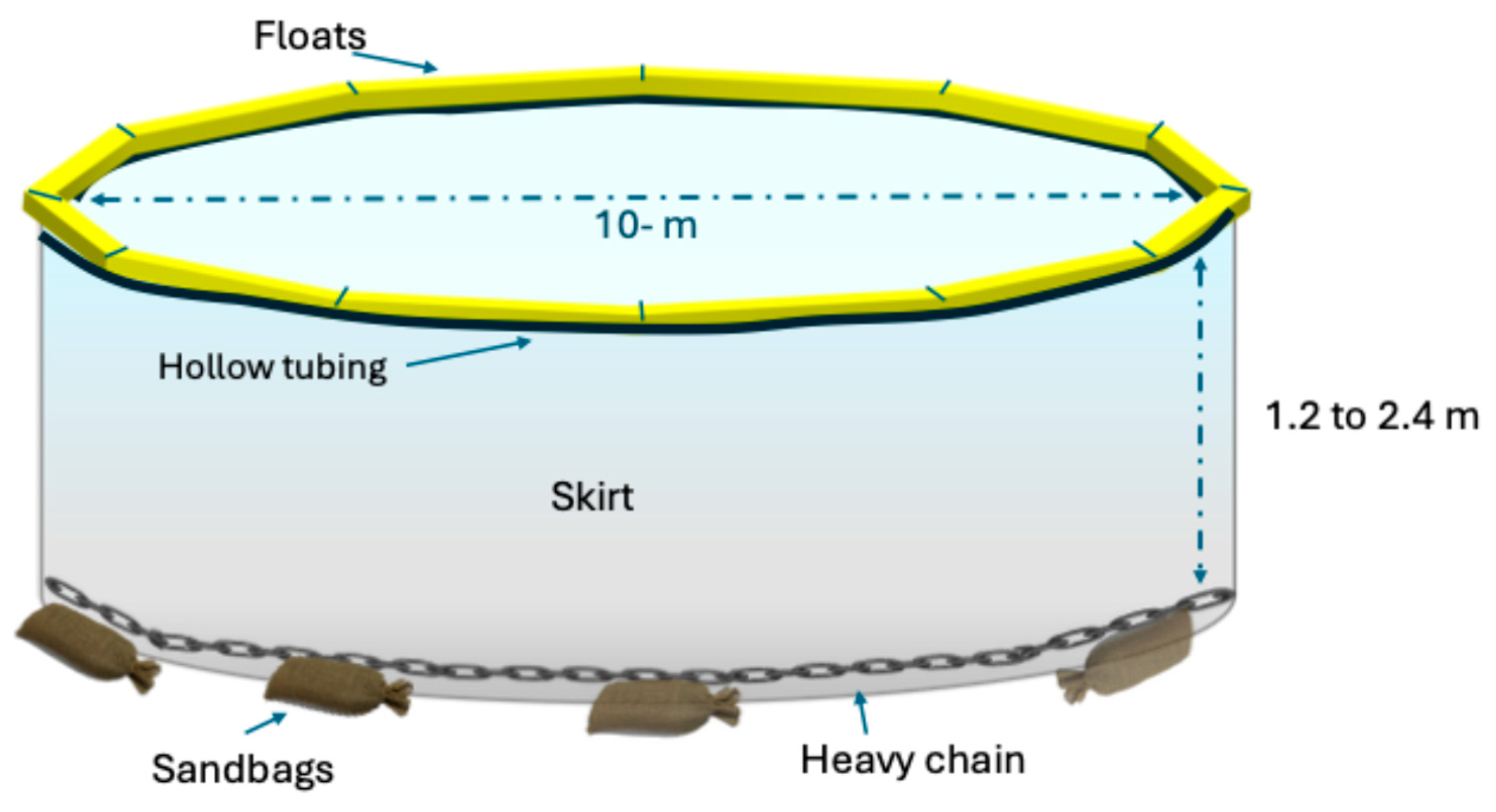
Figure 2.
Map of study area in Utah Lake, UT, USA. Limnocorrals were deployed in the upper right (NE) portion of Utah Lake marked with X. Approximate latitude: 40.330878°; longitude: -111.767755°.
Figure 2.
Map of study area in Utah Lake, UT, USA. Limnocorrals were deployed in the upper right (NE) portion of Utah Lake marked with X. Approximate latitude: 40.330878°; longitude: -111.767755°.
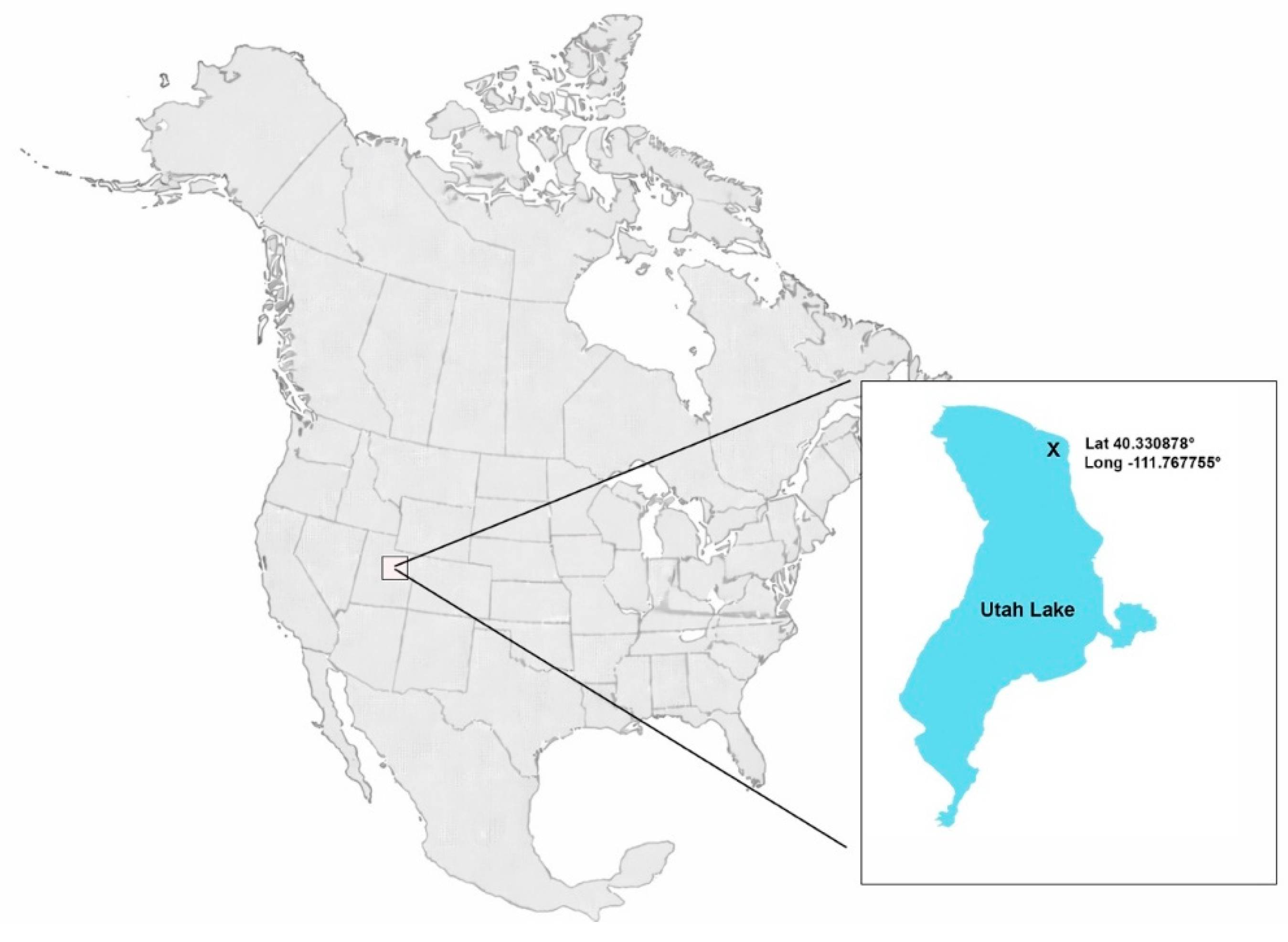
Figure 3.
Secchi depths and light attenuation coefficients inside and outside of limnocorrals, 2022/2023. Mean and 95% CIs with T-test results. N = 281 inside; 145 outside.
Figure 3.
Secchi depths and light attenuation coefficients inside and outside of limnocorrals, 2022/2023. Mean and 95% CIs with T-test results. N = 281 inside; 145 outside.
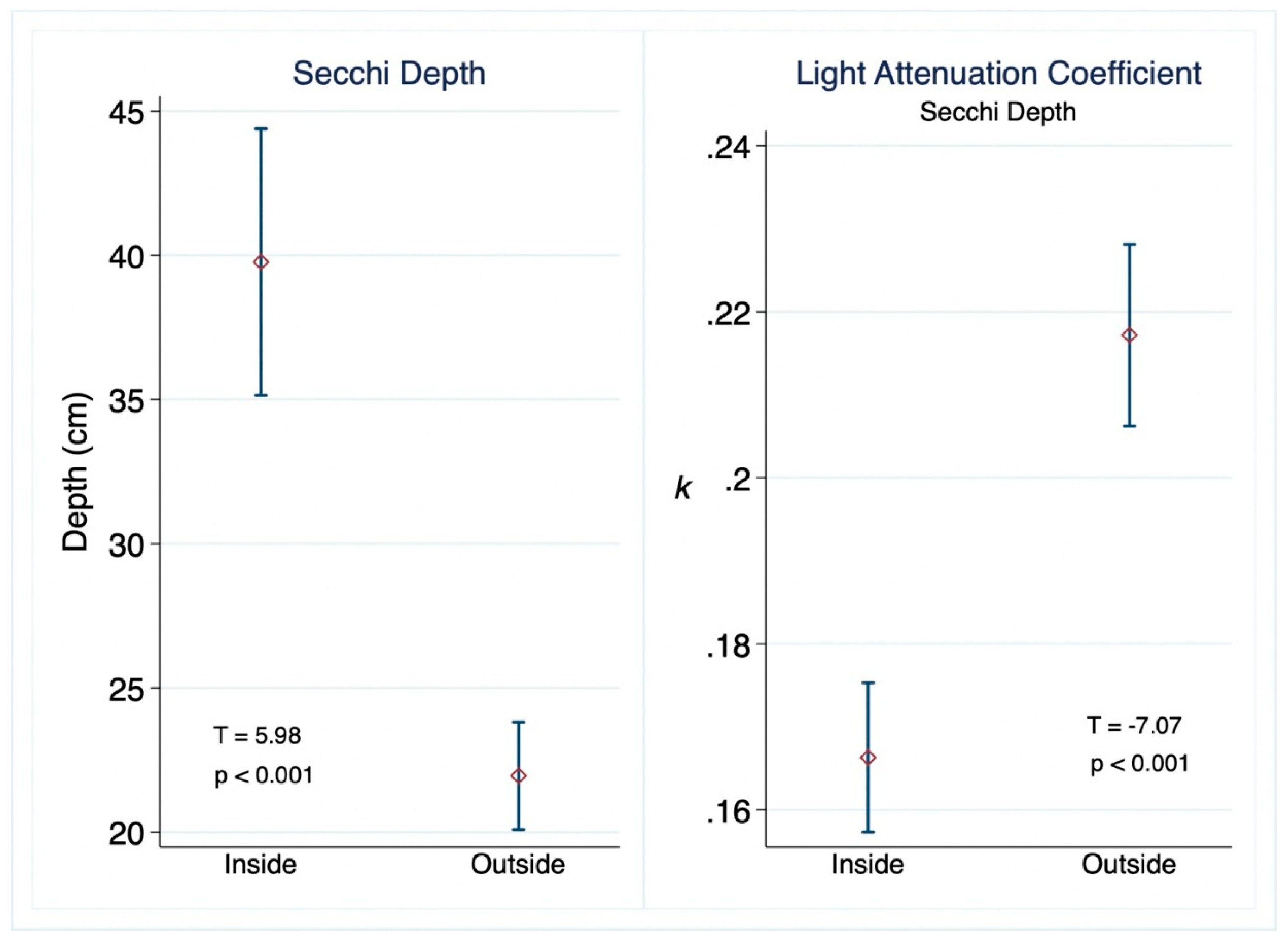
Figure 4.
Monthly differences in Secchi depth based light attenuation coefficient, k inside and outside corrals in 2022/2023. Numbers = sample size; * = inside vs outside significant difference at p = 0.05.
Figure 4.
Monthly differences in Secchi depth based light attenuation coefficient, k inside and outside corrals in 2022/2023. Numbers = sample size; * = inside vs outside significant difference at p = 0.05.
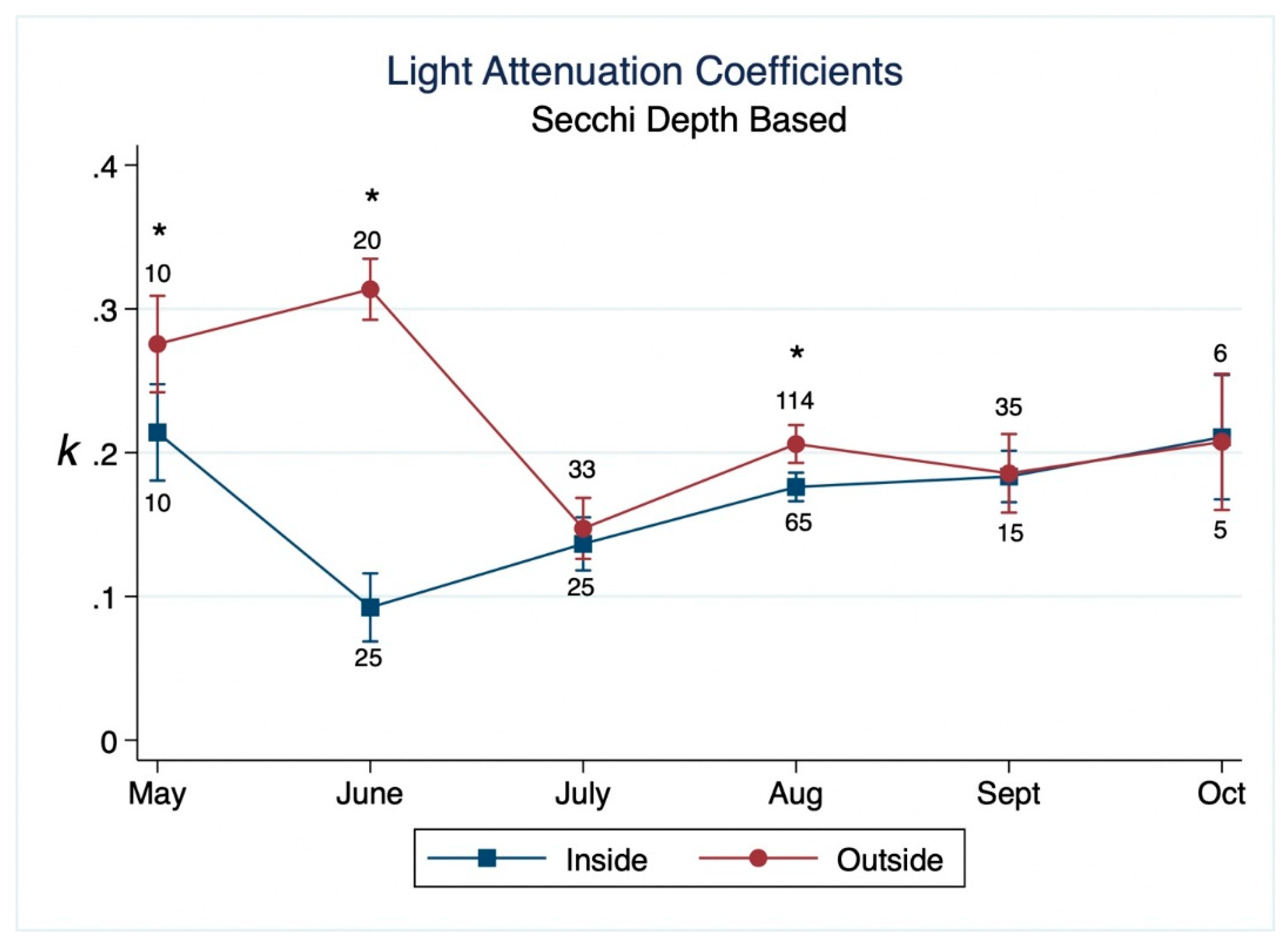
Figure 5.
Linear regression fit line with 95% CI lines of Ln(PAR (μmol photons m-2 s-1)) as a function of depth inside and outside of corrals in 2024. Dashed black x-axis reference line is the theoretical photosynthesis light limit Ln(0.01 μmol photons m-2 s-1) Numbers are predicted mean and 95% CIs at light limit from linear regression results.
Figure 5.
Linear regression fit line with 95% CI lines of Ln(PAR (μmol photons m-2 s-1)) as a function of depth inside and outside of corrals in 2024. Dashed black x-axis reference line is the theoretical photosynthesis light limit Ln(0.01 μmol photons m-2 s-1) Numbers are predicted mean and 95% CIs at light limit from linear regression results.
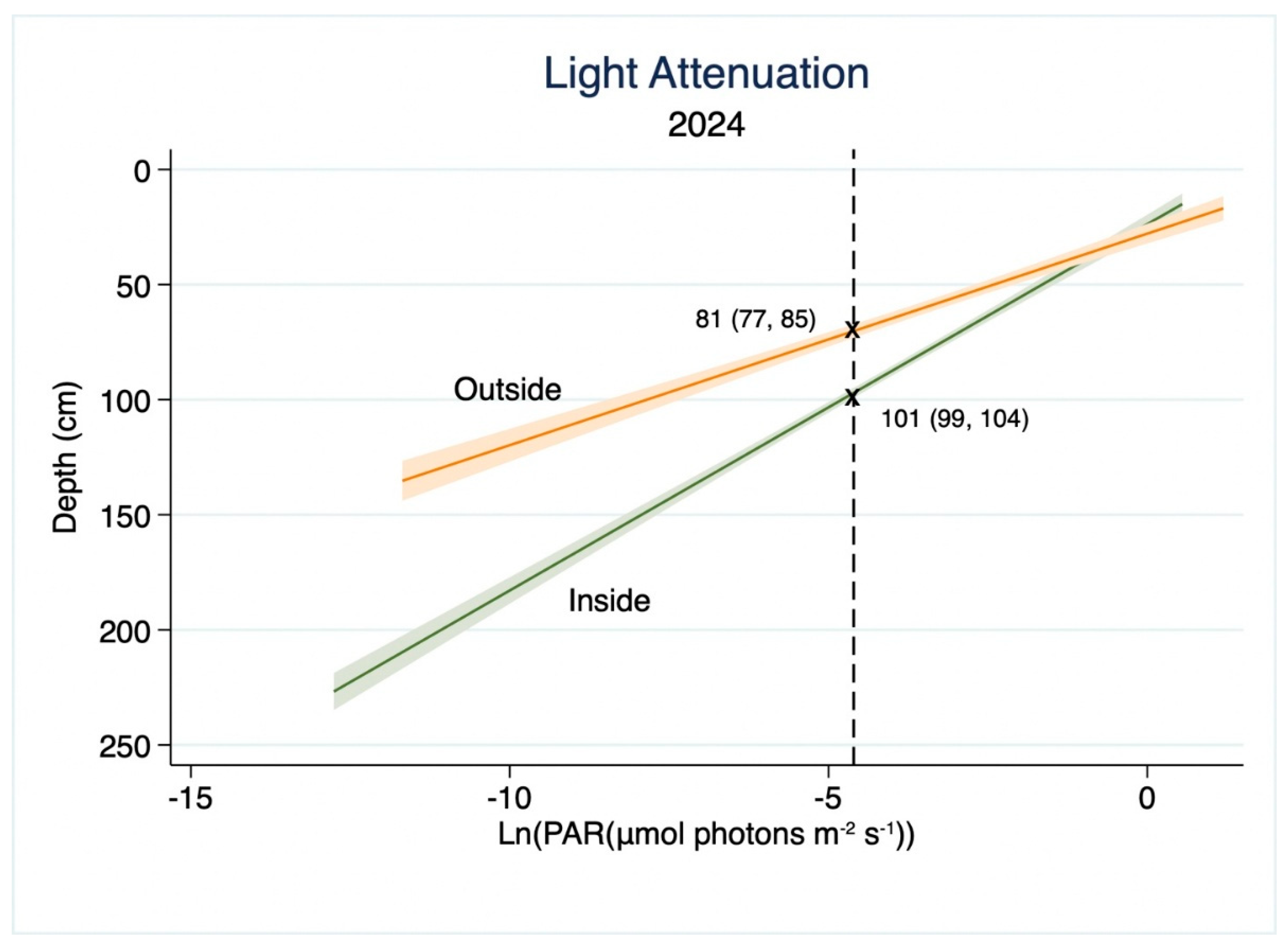
Figure 6.
Phytoplankton biovolume (μm-3 ml-1) (log10 transformed) inside vs outside (all years combined). Predicted mean and 95% CIs based on GLM regression using year and month as predictors.
Figure 6.
Phytoplankton biovolume (μm-3 ml-1) (log10 transformed) inside vs outside (all years combined). Predicted mean and 95% CIs based on GLM regression using year and month as predictors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



