Submitted:
18 February 2026
Posted:
25 February 2026
You are already at the latest version
Abstract
Drug repurposing offers a time- and cost-efficient strategy for accelerating the development of anticancer therapies by leveraging the established safety profiles of existing pharmaceuticals. In this study, we examined the anticancer potential of three tetracycline analogues—chemically modified tetracycline-3 (COL-3), doxycycline (DOX), and minocycline (MIN)- in leukemia by assessing their cytotoxic effects and modulation of the JAK2/STAT3 signaling pathway. Cytotoxicity was evaluated in K562, KG-1, and Jurkat cell lines using luminescence-based viability assays, whereas the mechanisms of cell death were analyzed by Annexin-V/7-AAD staining and Western blotting. COL-3 displayed the highest cytotoxic potency across all cell lines, with Jurkat cells showing the greatest overall sensitivity. Flow cytometry revealed that tetracycline analogues primarily induced apoptosis, although the molecular mechanisms differed between cell lines. In K562 and KG-1a cells, apoptosis occurred largely through JAK2/STAT3-independent mechanisms, involving differential regulation of BCL-2 family proteins: COL-3 reduced BCL-2 expression, whereas DOX and MIN increased BAX expression. In contrast, Jurkat cell apoptosis correlated with suppression of phosphorylated JAK2 and STAT3 and downregulation of BCL-2, implicating a JAK2/STAT3-dependent mechanism. Taken together, these findings demonstrate that tetracycline analogues exert cell line-specific anticancer activities through distinct molecular pathways. Among them, COL-3 emerges as the most potent analogue and acts through both JAK/STAT-dependent and -independent mechanisms. This work supports further investigation of COL-3 as a candidate for drug repurposing strategies in hematological malignancies.
Keywords:
JAK2/STAT3
; tetracycline-3 (COL-3)
; doxycycline (DOX) and minocycline (MIN)
; leukemia
1. Introduction
Drug repurposing, also known as drug repositioning, has emerged as an attractive strategy for accelerating the development of cancer therapeutics. By identifying new indications for approved drugs with known safety, pharmacokinetic and toxicological profiles, this approach reduces the time and cost associated with traditional drug discovery pipelines [1,2,3]. Increasing evidence supports the feasibility of repurposing antimicrobials as antineoplastic agents, particularly in settings where resistance, toxicity and limited durable responses remain significant clinical challenges [4,5,6].
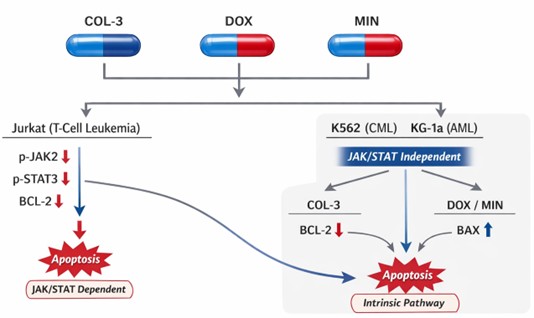
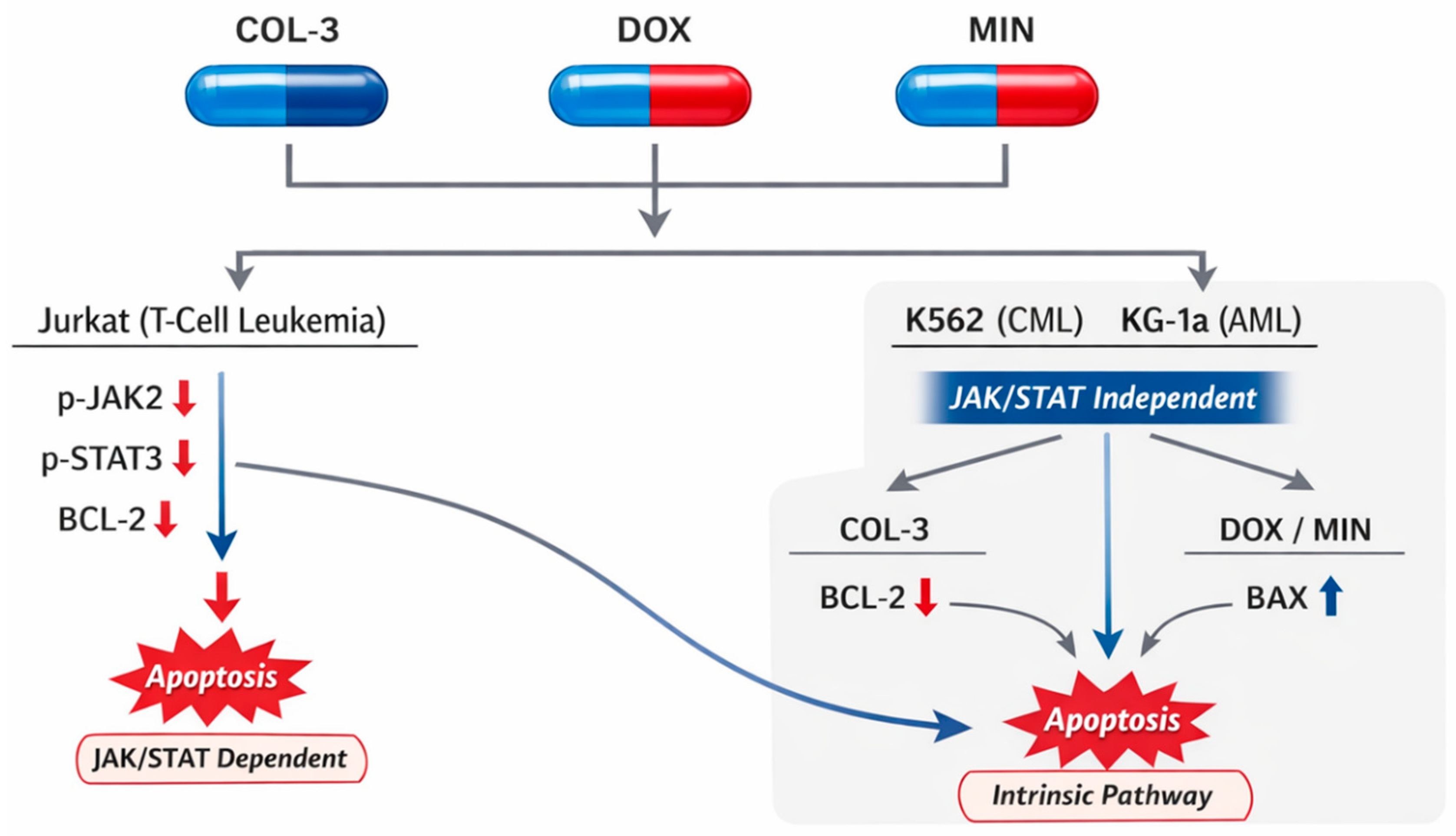
Figure 1.
Graphical Abstract: Mechanistically distinct apoptotic responses across leukemia subtypes support repurposing of tetracycline analogues in hematologic malignancies.
Figure 1.
Graphical Abstract: Mechanistically distinct apoptotic responses across leukemia subtypes support repurposing of tetracycline analogues in hematologic malignancies.
Leukemia encompasses a heterogeneous group of hematological malignancies characterized by uncontrolled proliferation of hematopoietic cells. Although considered relatively uncommon, leukemia represents a substantial clinical burden. In the United States, approximately 61,090 new cases were reported in 2021, accounting for ~3% of all newly diagnosed cancers [7,8]. In children, leukemia constitutes nearly one-third of all cancers, with acute lymphoblastic leukemia (ALL) representing 75–80% of pediatric cases [9,10,11]. Among adults, chronic myeloid leukemia (CML) comprises ~15% of leukemia diagnoses and predominantly affects older individuals, with a median age at diagnosis of 65 years [12,13,14]. Despite advances in targeted therapy and hematopoietic stem cell transplantation, resistance, relapse and treatment-related toxicities remain major obstacles, underscoring the need for novel therapeutic strategies [15,16,17].
Recent developments in molecular oncology have deepened the understanding of signaling networks driving leukemic cell survival and proliferation. Among these, the Janus kinase/signal transducer and activator of transcription (JAK/STAT) pathway plays a pivotal role in cytokine-mediated regulation of hematopoietic cell growth, differentiation and apoptosis. Aberrant JAK/STAT activation, whether driven by mutations, cytokine microenvironments or upstream kinase dysregulation, has been widely documented across leukemia subtypes and linked to therapeutic resistance and disease progression [18,19,20]. Inhibition of JAK kinases or blockade of STAT3 activation therefore represents a rational therapeutic approach, and several small-molecule inhibitors targeting this axis are currently under clinical evaluation.
Parallel to these efforts, the tetracycline class of antibiotics—including doxycycline (DOX), minocycline (MIN) and chemically modified tetracycline-3 (COL-3)—has attracted attention due to reported anti-metastatic, antiproliferative and pro-apoptotic activities in solid and hematologic malignancies [18,21,22,23,24]. Unlike conventional chemotherapeutics, tetracycline analogues exert a broad spectrum of cellular effects including mitochondrial dysfunction, inhibition of matrix metalloproteinases, interference with mitochondrial protein translation and induction of oxidative stress. Importantly, their interactions with apoptosis-regulating proteins such as members of the BCL-2 family suggest mechanistic links to intrinsic cell death pathways, making them attractive repurposing candidates.
However, the extent to which tetracycline analogues modulate oncogenic signaling pathways such as JAK2/STAT3 in leukemia remains insufficiently characterized. Given the pathway’s centrality in leukemogenesis and treatment resistance, elucidating these interactions may provide insight into repurposing opportunities and reveal mechanistic dependencies that vary across leukemia subtypes.
In this study, we investigated the cytotoxic effects of COL-3, DOX and MIN in three leukemia cell lines—K562, KG-1a and Jurkat—and examined their impact on cell death mechanisms and key signaling proteins. Specifically, we assessed modulation of the JAK2/STAT3 axis and the expression of BCL-2 and BAX, two major regulators of intrinsic apoptosis. Our findings reveal cell line-specific mechanistic responses and highlight COL-3 as the most potent analogue, acting through both JAK/STAT-dependent and independent pathways. These results support further evaluation of tetracycline derivatives as repurposed agents in hematologic malignancies.
2. Materials and Methods
2.1. Materials
Doxycycline hyclate (DOX) and minocycline hydrochloride (MIN) were purchased from Sigma Aldrich (Stockholm, Sweden) as crystalline powders and dissolved in distilled water. Chemically modified tetracycline-3 (COL-3) was provided by CollaGenex Pharmaceuticals Inc. (Newtown, PA, USA) and Galderma R&D (Sophia-Antipolis, France) and dissolved in dimethyl sulfoxide (DMSO). RPMI 1640 medium supplemented with GlutaMAX™ and HEPES, fetal bovine serum (FBS), penicillin–streptomycin, phosphate-buffered saline (PBS) and other cell culture reagents were obtained from Gibco (Thermo Fisher Scientific). Additional reagents included MycoAlert™ Mycoplasma Detection Kit (Lonza), CellTiter-Glo® Luminescent Cell Viability Assay (Promega), PE Annexin V Apoptosis Detection Kit I (BD Biosciences), and standard Western blotting materials. Antibodies against JAK2, phospho-JAK2, STAT3, phospho-STAT3, BCL-2, BAX and β-actin and corresponding HRP-conjugated secondary antibodies were sourced according to manufacturers’ specifications. Catalog numbers for all reagents are provided in the supplementary materials.
2.2. Cell Culture
K562, KG-1a and Jurkat cell lines were maintained in RPMI 1640 medium supplemented with 10% FBS and 1% penicillin–streptomycin at 37 °C in a humidified incubator with 5% CO2. KG-1a cells were cultured with 20% FBS. Cells were used at passage numbers below 10 to ensure consistency and were routinely screened for mycoplasma contamination using MycoAlert™, with only mycoplasma-free cultures included in experiments.
2.3. Cell Starvation and Activation of Jurkat Cells
Jurkat cells were serum-starved for 4 h in RPMI 1640 to facilitate subsequent activation of the JAK2/STAT3 pathway. Activation was induced using pervanadate (PERV), a protein tyrosine phosphatase inhibitor that enhances tyrosine phosphorylation and thereby promotes JAK2 and STAT3 activation. After starvation, cells were treated with an activation mixture containing PERV and FBS for 15 min at room temperature, washed with PBS and resuspended in complete medium prior to treatment with tetracycline analogues.
2.4. Drug Treatment Conditions
Cells were divided into eight experimental groups per cell line, consisting of untreated controls, vehicle controls (0.1% DMSO), and two concentrations each of COL-3, DOX and MIN. Concentration ranges were selected based on pilot experiments to approximate IC50 values. Jurkat cells were additionally analyzed in activated and non-activated states. Detailed concentration schemes for each cell line are provided in Supplementary Table 1.
2.5. Cell Viability Assay
Cytotoxicity was assessed using the CellTiter-Glo® Luminescent Cell Viability Assay according to the manufacturer’s instructions. Briefly, 1.5×104 cells were seeded per well in opaque 96-well plates and treated with drug concentrations for 24 h. CellTiter-Glo® reagent was added at a 1:1 ratio, mixed for 2 min and incubated for 10 min at room temperature to stabilize luminescence. Luminescence was recorded using a SpectraMax i3X plate reader and analyzed with SoftMax Pro 7. IC50 values were determined from nonlinear regression fits.
2.6. Assessment of Cell Death Mechanisms
Apoptosis and necrosis were evaluated by Annexin V/7-AAD staining. After treatment for 4 h and 24 h, cells were washed twice with cold PBS, resuspended in binding buffer and stained with PE Annexin V and 7-AAD for 15 min at room temperature in the dark. Samples were analyzed within 1 h using a MACSQuant Analyzer 10. Populations were gated as viable (Annexin V−/7-AAD−), early apoptotic (Annexin V+/7-AAD−), late apoptotic (Annexin V+/7-AAD+) or necrotic (Annexin V−/7-AAD+). Compensation controls were included for all fluorochromes.
2.7. Protein Extraction and Western Blotting
Cells (1×107) were lysed in RIPA buffer supplemented with protease and phosphatase inhibitors and incubated on ice for 30 min. Protein concentrations were determined using a BCA assay. Equal amounts of protein were mixed with sample buffer containing β-mercaptoethanol, denatured at 95 °C for 5 min and separated on 4–15% SDS-PAGE gels. Proteins were transferred onto PVDF membranes overnight at 4 °C, blocked with 5% BSA in TBS-T for 2 h and probed with primary antibodies against JAK2, phospho-JAK2, STAT3, phospho-STAT3, BCL-2, BAX and β-actin. Membranes were incubated with HRP-conjugated secondary antibodies, washed and visualized using a LI-COR scanner. Band intensities were quantified with Odyssey or ImageJ software.
2.8. Statistical Analysis
Data are presented as mean ± standard deviation (SD) unless otherwise indicated. Comparisons among multiple groups were performed using one-way ANOVA followed by post hoc tests. Differences were considered statistically significant at p < 0.05. IC50 values were calculated using nonlinear curve fitting. All analyses were performed using GraphPad Prism 10.
3. Results
3.1. Cytotoxic Effects of Tetracycline Analogues in Leukemia Cell Lines
The cytotoxicities of COL-3, DOX and MIN were evaluated in K562, KG-1a and Jurkat cells using CellTiter-Glo™. All compounds reduced cell viability in a dose-dependent manner, with potency varying among both analogues and cell lines.
In K562 cells, COL-3 exhibited the lowest IC50 value (29.57 µg/mL; 95% CI: 21.42–41.23; R2 = 0.9430), followed by MIN (178.5 µg/mL; 95% CI: 134.6–235.2; R2 = 0.9395) and DOX (238.6 µg/mL; 95% CI: 136.4–592.6; R2 = 0.9188) (Figure 2A–C; Supplementary Table S1). In KG-1a cells, COL-3 again showed the greatest potency (IC50 = 7.486 µg/mL; 95% CI: 5.912–9.619; R2 = 0.9677), whereas DOX and MIN exhibited higher IC50 values of 99.54 µg/mL (95% CI: 70.73–152.7; R2 = 0.9553) and 167.8 µg/mL (95% CI: 121.9–265.8; R2 = 0.9583), respectively (Figure 2D–F; Supplementary Table S2). Jurkat cells displayed the greatest overall sensitivity, with IC50 values of 3.625 µg/mL (COL-3; 95% CI: 3.2–4.1; R2 = 0.9898), 8.049 µg/mL (DOX; 95% CI: 7.32–8.86; R2 = 0.9792) and 9.485 µg/mL (MIN; 95% CI: 8.71–10.33; R2 = 0.9884) (Figure 2G–I; Supplementary Table S3). Overall, COL-3 demonstrated the highest cytotoxic potency across all three cell lines, followed by DOX and MIN, whereas Jurkat cells exhibited the greatest sensitivity and K562 cells the greatest resistance.
3.2. Tetracycline Analogues Induce Apoptosis with Cell Line-Specific Features
To determine the mode of cell death, Annexin-V/7-AAD staining was performed after 4 h and 24 h of exposure to each analogue.
In K562 Cells, COL-3 (12.5 and 25 µg/mL) increased Annexin-V+ populations at both time points (10.6–16.4% at 4 h; 29.6–32.1% at 24 h vs. 23% control), accompanied by modest increases in 7-AAD+ cells. DOX (100 and 200 µg/mL) showed minimal changes at 4 h but increased Annexin-V+ cells at 24 h (25.6% vs. 23.1%), with late apoptotic (double-positive) populations elevated at 200 µg/mL. MIN (100 and 200 µg/mL) produced significant increases in late apoptotic cells at 24 h (e.g., 31.8% at 200 µg/mL), accompanied by reduced viability. These data indicate apoptosis as the predominant death modality in K562 cells, with magnitude correlating with concentration and exposure time (Figure 3; Supplementary Figure S1, S2, Supplementary Table S4).
Next in KG-1a Cells, COL-3 (3 and 6 µg/mL) significantly increased early apoptosis at both 4 h (4.2–5.6% vs. 0.04% control) and 24 h (4.6–15.6% vs. 0.02% control), with moderate increases in late apoptosis at 24 h. DOX (50 and 100 µg/mL) induced similar patterns, with early apoptotic populations rising from 1.2–1.8% at 4 h to 14.5–17.2% at 24 h. MIN (100 and 200 µg/mL) elicited substantial apoptosis at 24 h (18–23.1%) with associated increases in late apoptotic populations (7.2–23.5%). Together, these results demonstrate that all three analogues induce apoptosis in KG-1a cells in a time-dependent manner (Figure4; Supplementary Figure S3, S4, Supplementary Table S5).
Figure 3.
Effect of COL-3, DOX, and MIN on Apoptosis in K562 cells assessed by Flow Cytometry. Flow cytometric analysis of K562 cells treated with COL-3, DOX, and MIN, showing the percentage of Annexin V–positive cells (early apoptosis), 7-AAD–positive cells (late apoptosis/necrosis), and double-stained cells (late apoptosis/necrosis). Data represent the mean ± standard error (SE) from three independent experiments. Bar graphs illustrate the percentage changes in cell populations. a: Statistically significant difference compared to 4-hour control cells (p < 0.05). b: Statistically significant difference compared to 24-hour control cells (p < 0.05).
Figure 3.
Effect of COL-3, DOX, and MIN on Apoptosis in K562 cells assessed by Flow Cytometry. Flow cytometric analysis of K562 cells treated with COL-3, DOX, and MIN, showing the percentage of Annexin V–positive cells (early apoptosis), 7-AAD–positive cells (late apoptosis/necrosis), and double-stained cells (late apoptosis/necrosis). Data represent the mean ± standard error (SE) from three independent experiments. Bar graphs illustrate the percentage changes in cell populations. a: Statistically significant difference compared to 4-hour control cells (p < 0.05). b: Statistically significant difference compared to 24-hour control cells (p < 0.05).
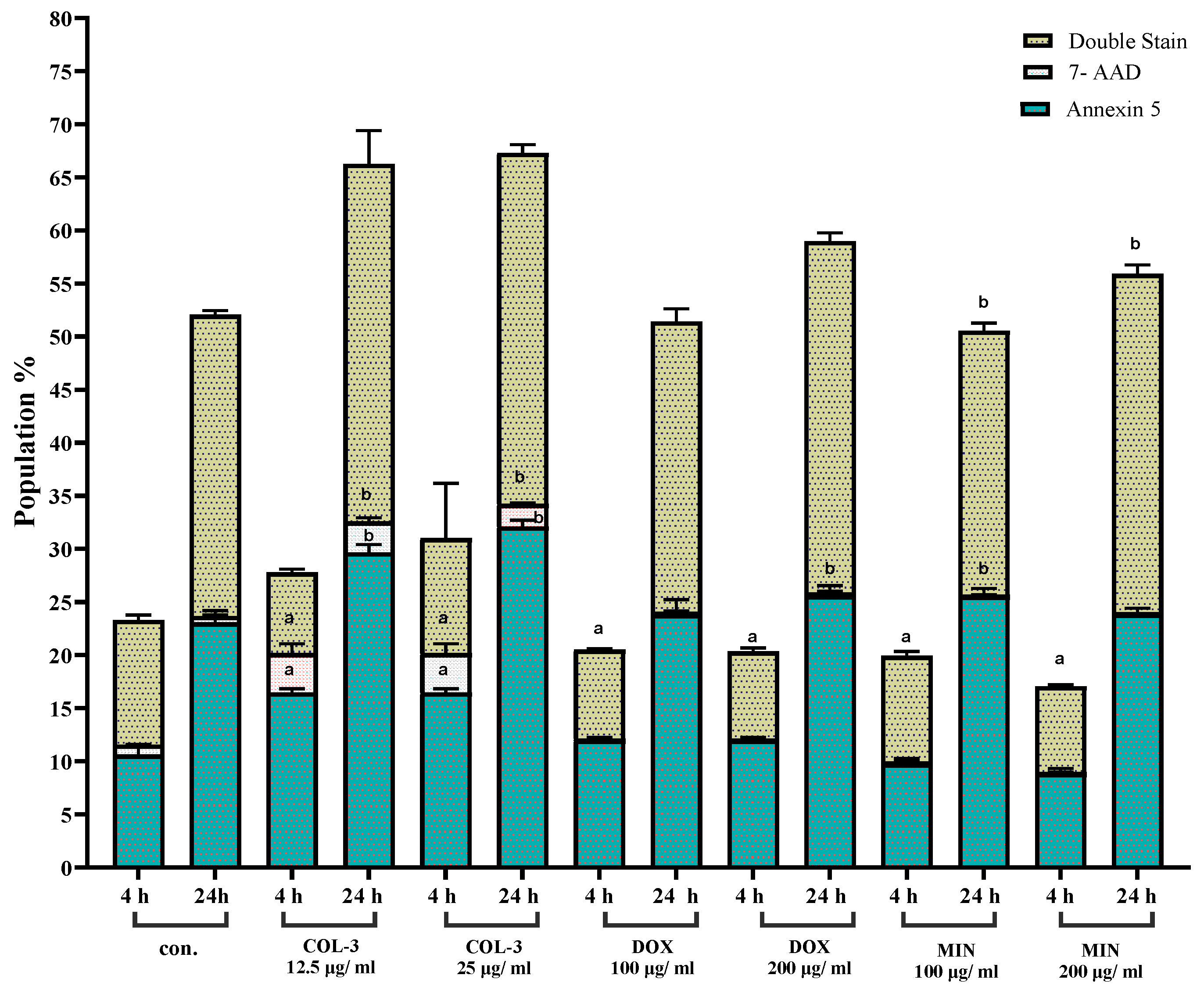
Figure 4.
Effect of COL-3, DOX, and MIN on Apoptosis in KG-1a cells assessed by Flow Cytometry. Flow cytometric analysis of K562 cells treated with COL-3, DOX, and MIN, showing the percentage of Annexin V–positive cells (early apoptosis), 7-AAD–positive cells (late apoptosis/necrosis), and double-stained cells (late apoptosis/necrosis). Data represent the mean ± standard error (SE) from three independent experiments. Bar graphs illustrate the percentage changes in cell populations. a: Statistically significant difference compared to 4-hour control cells (p < 0.05). b: Statistically significant difference compared to 24-hour control cells (p < 0.05).
Figure 4.
Effect of COL-3, DOX, and MIN on Apoptosis in KG-1a cells assessed by Flow Cytometry. Flow cytometric analysis of K562 cells treated with COL-3, DOX, and MIN, showing the percentage of Annexin V–positive cells (early apoptosis), 7-AAD–positive cells (late apoptosis/necrosis), and double-stained cells (late apoptosis/necrosis). Data represent the mean ± standard error (SE) from three independent experiments. Bar graphs illustrate the percentage changes in cell populations. a: Statistically significant difference compared to 4-hour control cells (p < 0.05). b: Statistically significant difference compared to 24-hour control cells (p < 0.05).
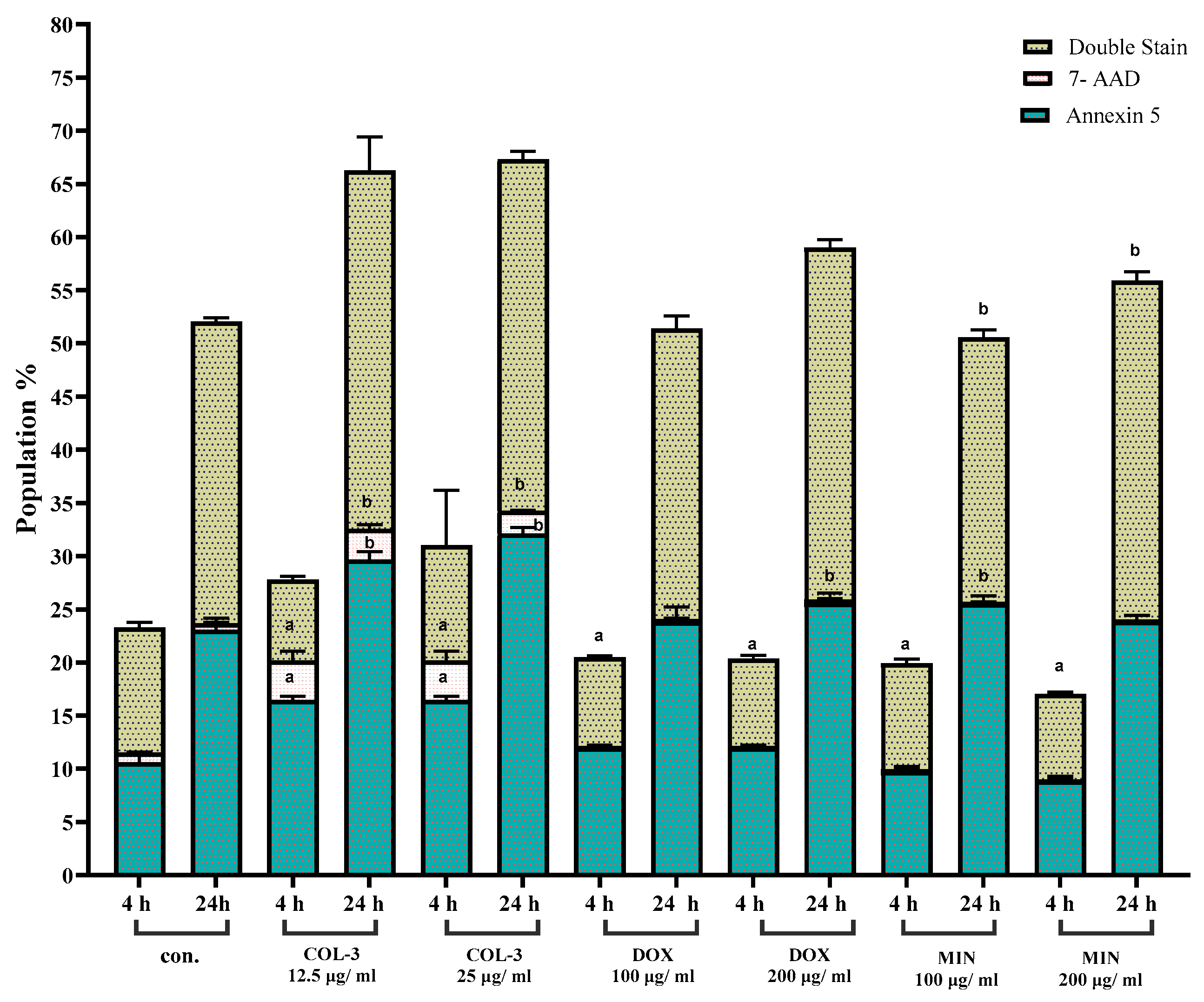
In Jurkat cells, COL-3 induced marked increases in late apoptotic populations at both 4 h and 24 h (e.g., 16–29% vs. 2.5–11% control), while DOX increased double-positive populations to 24–28.5% at 24 h. MIN induced the strongest apoptotic response at early time points, with Annexin-V+ populations reaching 6.3–6.5% at 4 h and 29% at 24 h. These findings indicate that Jurkat cells exhibit rapid apoptotic induction in response to tetracycline analogues (Figure 5; Supplementary Figure S5, S6, Supplementary Table S6).
4. Effects on the JAK2/STAT3 Pathway and BCL-2 Family Proteins
Western blotting was performed to determine whether apoptosis correlated with modulation of JAK2/STAT3 pathway components and BCL-2 family proteins.
In K562 Cells, all three analogues reduced total JAK2 and phospho-JAK2 levels, with COL-3 and MIN exhibiting statistically significant decreases. Total STAT3 expression also declined across conditions, whereas phospho-STAT3 increased, reaching significance in COL-3 and MIN groups. COL-3 and MIN reduced BAX expression, while DOX and MIN increased BCL-2 levels, suggesting apoptosis occurred independently of JAK2/STAT3 suppression and was associated with BCL-2 upregulation (Figure 5; Supplementary Figure S4).The other treatment groups showed small increases in BCL-2 expressions that were not statistically significant (Figure 6F, Supplementary Figure S7).
Next in KG-1a Cells, COL-3 and DOX (50 µg/mL) modestly reduced total JAK2 expression, whereas DOX (100 µg/mL) and MIN increased it. Phospho-JAK2 was reduced by COL-3 and MIN. STAT3 levels declined in all groups except MIN (100 µg/mL), while phospho-STAT3 increased significantly in DOX- and MIN-treated cells. COL-3 selectively reduced BCL-2, whereas DOX and MIN increased BAX expression, indicating apoptosis was associated with modulation of BCL-2 family proteins rather than JAK2/STAT3 inhibition (Figure 7; Supplementary Figure. S8).
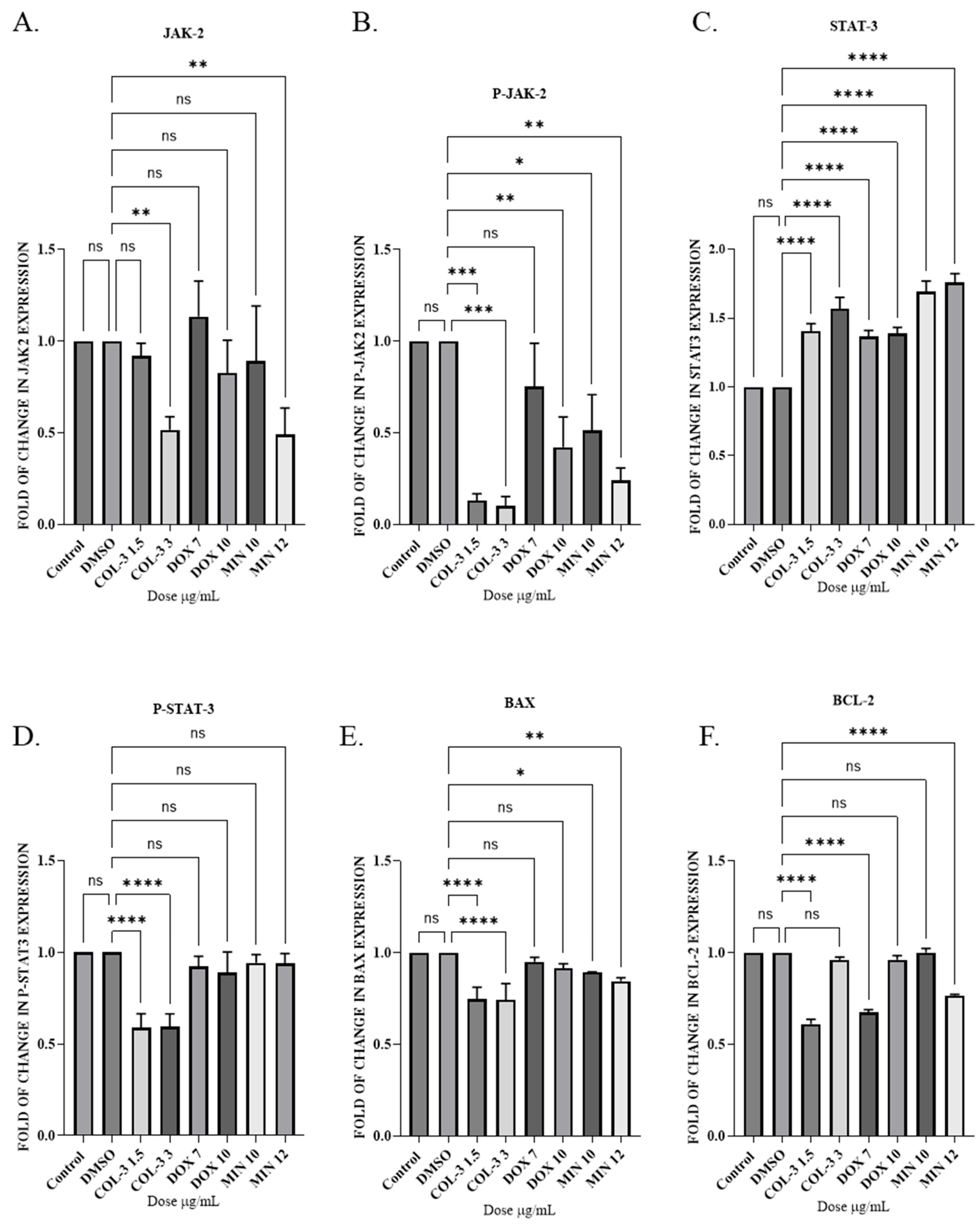
Subsequently in Jurkat cells, all analogues reduced total and phospho-JAK2 levels, with phospho-JAK2 suppression reaching statistical significance in all groups except DOX (7 µg/mL). Phospho-STAT3 levels were reduced by all treatments, with COL-3 showing significant effects. Total STAT3 levels increased across groups. BCL-2 expression was significantly reduced by COL-3, DOX and MIN (12 µg/mL), whereas BAX expression decreased following COL-3 and MIN treatment. These data indicate that apoptosis in Jurkat cells correlates with inhibition of the JAK2/STAT3 pathway and downregulation of BCL-2 (Figure 8; Supplementary Figure. S9).
5. Discussion
This study demonstrates that the tetracycline analogues COL-3, DOX and MIN exhibit differential cytotoxic and mechanistic effects in leukemia cell lines and modulate apoptotic pathways through both JAK2/STAT3-dependent and -independent mechanisms. Among the three compounds, COL-3 consistently showed the greatest potency, while Jurkat cells displayed the highest sensitivity. These findings extend emerging evidence that tetracycline derivatives possess anticancer activities distinct from their traditional antimicrobial roles and support their consideration as candidates for drug repurposing strategies in hematologic malignancies.
Previous studies have reported variable antitumor responses to tetracyclines in both solid and hematologic cancers, including effects on mitochondrial function, transcriptional regulation and apoptosis induction [21,23,24,25,26]. The IC50 values observed in the present work are broadly consistent with trends described by Fares et al. (2015), although absolute values differed, likely reflecting assay platforms and detection sensitivity[27]. The use of CellTiter-Glo in this study, which quantifies ATP and is considered more sensitive for assessing metabolic activity, may account for some of these differences. Collectively, these data reinforce that tetracycline analogues exhibit anti-leukemic effects in vitro and that their potency is influenced by both drug structure and leukemia subtype.
Mechanistically, the tetracycline analogues produced distinct effects on the JAK2/STAT3 axis and BCL-2 family proteins. In K562 and KG-1a cells, apoptosis did not correlate with suppression of phosphorylated JAK2 or phosphorylated STAT3, indicating that cell death occurred largely through alternative pathways. Instead, differential modulation of BCL-2 family proteins was observed: COL-3 reduced BCL-2 expression in KG-1a cells, whereas DOX and MIN increased BAX levels. These findings align with previous reports that mitochondrial and oxidative stress-related mechanisms contribute to tetracycline-induced cytotoxicity and that BCL-2 family proteins may mediate these effects [24,26,28]. The distinct profiles observed among cell lines underscore that leukemia subtypes may possess divergent apoptotic wiring, with implications for therapeutic targeting.
In contrast, Jurkat cells exhibited a clear relationship between apoptosis and suppression of phosphorylated JAK2 and STAT3, accompanied by reduced BCL-2 expression. Given that constitutive STAT3 activation has been associated with survival signaling in T-cell malignancies, the observed inhibition of this pathway suggests that COL-3, DOX and MIN may exert JAK/STAT-dependent effects in lymphoid contexts. These results differ from observations in non-activated Jurkat cells [29], suggesting that pathway priming state and cellular activation status may influence drug response. Together, these findings illustrate that tetracycline analogues can engage distinct apoptotic routes depending on leukemia subtype, pathway dependence and survival circuitry.
From a translational perspective, the heterogeneous responses observed across leukemia models highlight both opportunities and challenges for repurposing tetracycline analogues. The superior potency of COL-3, coupled with its dual pathway engagement, suggests that it may hold particular promise for further evaluation. Although DOX and MIN were less potent and required higher concentrations to elicit effects, their established clinical use, favorable safety profiles and widespread availability make them appealing candidates for combination or adjunctive approaches. Notably, prior work has demonstrated synergistic interactions between tetracyclines and cytotoxic agents in solid tumor models [21], indicating that combinatorial strategies in leukemia warrant investigation.
This study has several limitations. First, all experiments were conducted in established cell lines, which may not fully recapitulate primary leukemia biology. Second, no in vivo validation was performed, and pharmacokinetic considerations such as achievable plasma concentrations may influence translational feasibility. Third, the study did not interrogate downstream transcriptional consequences of STAT3 inhibition or mitochondrial function, which could provide additional mechanistic insight. Addressing these limitations through in vivo models, primary patient samples and pathway-level profiling will be essential to define therapeutic relevance more comprehensively.
Future work should prioritize evaluating COL-3 in preclinical leukemia models to determine dosing, tolerability and functional pathway engagement in vivo. Investigation into combinatorial regimens, particularly with JAK/STAT inhibitors or BCL-2 family modulators such as venetoclax, may yield synergistic effects given the mechanistic intersections identified. Furthermore, profiling leukemia subtypes with defined JAK/STAT activation states may enable stratification strategies for precision repurposing.
In conclusion, this study provides evidence that tetracycline analogues exert anti-leukemic effects through cell line-specific mechanisms and identifies COL-3 as the most potent candidate for further investigation. While additional validation is required, these findings support the continued exploration of tetracycline derivatives as repurposed agents in hematologic malignancies and highlight the value of targeting survival signaling pathways such as JAK/STAT3 in leukemia.
6. Conclusions
This study demonstrates that tetracycline analogues exert anti-leukemic effects through distinct molecular mechanisms that vary across leukemia cell lines. COL-3 showed the highest cytotoxic potency and engaged both JAK/STAT-dependent and -independent apoptotic pathways, whereas DOX and MIN induced apoptosis primarily through modulation of BCL-2 family proteins. Jurkat cells exhibited apoptosis associated with suppression of phosphorylated JAK2 and STAT3 and downregulation of BCL-2, suggesting pathway dependence in lymphoid cells. In contrast, apoptosis in K562 and KG-1a cells occurred independently of JAK/STAT signaling and involved differential regulation of BCL-2 or BAX. These findings highlight mechanistic heterogeneity in leukemia cell responses to tetracycline analogues and identify COL-3 as the most promising candidate for further exploration. While additional validation in primary models and in vivo systems is required, this work supports continued evaluation of tetracycline derivatives as repurposed candidates in hematological malignancies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: title; Table S1.
Author Contributions
Zienab Mohammad: Conceptualization, data curation, formal analysis, investigation, methodology, visualization, writing—original draft. Doste R. Mamand: Data curation, conceptualization, formal analysis, investigation, methodology, visualization, writing—original draft, writing—review & editing. Oscar P.B. Wiklander: Conceptualization, resources, funding acquisition, visualization, writing—review & editing. Moustapha Hassan: Conceptualization, resources, funding acquisition, investigation, methodology, supervision, visualization, writing—review & editing. Hoda W. El-Gawly, Naglaa A. El-Sherbeeny, Hala M.F. Mohammad, and Mohamed K. Elkherbetawy: Conceptualization, resources, investigation, methodology, supervision, visualization, writing—review & editing.
Funding
Zienab Mohammad Hassan was supported by a scholarship (JS15/19) from the Ministry of Higher Education of the Arab Republic of Egypt. Oscar P.B. Wiklander acknowledges funding from the Swedish Research Council (VR, 2022-02449), the Swedish Cancer Society (Cancerfonden, 23 2935 Pj), Radiumhemmet (Project #241392), the Center for Innovative Medicine (CIMED) junior investigator grants (FoUI-976434), and Karolinska Institutet (2-116/2023).
Institutional Review Board Statement
not applicable.
Informed Consent Statement
not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors acknowledge the Ministry of Higher Education of the Arab Republic of Egypt and financial support from the Swedish Research Council, the Swedish Cancer Society, Radiumhemmet, the Center for Innovative Medicine (CIMED) Junior Investigator Grants, and Karolinska Institutet.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| COL-3 | tetracycline-3 |
| DOX | doxycycline |
| MIN | minocycline |
References
- Chukhlovin, A.B. Drug Repurposing in Leukemia Treatment and Hematopoietic Stem Cell Transplantation. CTT 2019, 8, 12–19. [CrossRef]
- Langedijk, J.; Mantel-Teeuwisse, A.K.; Slijkerman, D.S.; Schutjens, M.-H.D.B. Drug Repositioning and Repurposing: Terminology and Definitions in Literature. Drug Discovery Today 2015, 20, 1027–1034. [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug Repurposing: Progress, Challenges and Recommendations. Nat Rev Drug Discov 2019, 18, 41–58. [CrossRef]
- Damery, E.; Solimando, D.A.; Waddell, J.A. Arsenic Trioxide and Tretinoin (AsO/ATRA) for Acute Promyelocytic Leukemia (APL). Hosp Pharm 2016, 51, 628–632. [CrossRef]
- Eisenstein, E.L.; Hill, K.D.; Wood, N.; Kirchner, J.L.; Anstrom, K.J.; Granger, C.B.; Rao, S.V.; Baldwin, H.S.; Jacobs, J.P.; Jacobs, M.L.; et al. Evaluating Registry-Based Trial Economics: Results from the STRESS Clinical Trial. Contemporary Clinical Trials Communications 2024, 38, 101257. [CrossRef]
- Nehme, R.; Hallal, R.; El Dor, M.; Kobeissy, F.; Gouilleux, F.; Mazurier, F.; Zibara, K. Repurposing of Acriflavine to Target Chronic Myeloid Leukemia Treatment. CMC 2021, 28, 2218–2233. [CrossRef]
- Pelcovits, A.; Niroula, R. Acute Myeloid Leukemia: A Review. R I Med J (2013) 2020, 103, 38–40.
- SEER data base; National Cancer Institute Acute Myeloid Leukemia—Cancer Stat Facts Available online: https://seer.cancer.gov/statfacts/html/amyl.html (accessed on 5 July 2024).
- Möricke, A.; Reiter, A.; Zimmermann, M.; Gadner, H.; Stanulla, M.; Dördelmann, M.; Löning, L.; Beier, R.; Ludwig, W.-D.; Ratei, R.; et al. Risk-Adjusted Therapy of Acute Lymphoblastic Leukemia Can Decrease Treatment Burden and Improve Survival: Treatment Results of 2169 Unselected Pediatric and Adolescent Patients Enrolled in the Trial ALL-BFM 95. Blood 2008, 111, 4477–4489. [CrossRef]
- Puumala, S.E.; Ross, J.A.; Aplenc, R.; Spector, L.G. Epidemiology of Childhood Acute Myeloid Leukemia. Pediatric Blood & Cancer 2013, 60, 728–733. [CrossRef]
- Seibel, N.L. Treatment of Acute Lymphoblastic Leukemia in Children and Adolescents: Peaks and Pitfalls. Hematology 2008, 2008, 374–380. [CrossRef]
- Cortes, J.E.; Gambacorti-Passerini, C.; Deininger, M.W.; Mauro, M.J.; Chuah, C.; Kim, D.-W.; Dyagil, I.; Glushko, N.; Milojkovic, D.; Le Coutre, P.; et al. Bosutinib Versus Imatinib for Newly Diagnosed Chronic Myeloid Leukemia: Results From the Randomized BFORE Trial. JCO 2018, 36, 231–237. [CrossRef]
- Copland, M.; Slade, D.; Byrne, J.; Brock, K.; De Lavallade, H.; Craddock, C.; Clark, R.; Smith, M.; Bishop, R.; Milojkovic, D.; et al. FLAG-IDA and Ponatinib in Patients with Blast Phase Chronic Myeloid Leukaemia: Results from the Phase I/II UK Trials Acceleration Programme Matchpoint Trial. Blood 2019, 134, 497–497. [CrossRef]
- Jabbour, E.J.; Faderl, S.; Kantarjian, H.M. Adult Acute Lymphoblastic Leukemia. Mayo Clinic Proceedings 2005, 80, 1517–1527. [CrossRef]
- Martinelli, G.; Boissel, N.; Chevallier, P.; Ottmann, O.; Gökbuget, N.; Topp, M.S.; Fielding, A.K.; Rambaldi, A.; Ritchie, E.K.; Papayannidis, C.; et al. Complete Hematologic and Molecular Response in Adult Patients With Relapsed/Refractory Philadelphia Chromosome–Positive B-Precursor Acute Lymphoblastic Leukemia Following Treatment With Blinatumomab: Results From a Phase II, Single-Arm, Multicenter Study. JCO 2017, 35, 1795–1802. [CrossRef]
- Molina, J.C.; Carraway, H.E. Treatment of Relapsed Acute Lymphocytic Leukemia in Adult Patients. Curr. Treat. Options in Oncol. 2024, 25, 993–1010. [CrossRef]
- Van Weelderen, R.E.; Klein, K.; Harrison, C.J.; Jiang, Y.; Abrahamsson, J.; Arad-Cohen, N.; Bart-Delabesse, E.; Buldini, B.; De Moerloose, B.; Dworzak, M.N.; et al. Measurable Residual Disease and Fusion Partner Independently Predict Survival and Relapse Risk in Childhood KMT2A -Rearranged Acute Myeloid Leukemia: A Study by the International Berlin-Frankfurt-Münster Study Group. JCO 2023, 41, 2963–2974. [CrossRef]
- Liang, D.; Wang, Q.; Zhang, W.; Tang, H.; Song, C.; Yan, Z.; Liang, Y.; Wang, H. JAK/STAT in Leukemia: A Clinical Update. Mol Cancer 2024, 23, 25. [CrossRef]
- Lin, M.; Nebral, K.; Gertzen, C.G.W.; Ganmore, I.; Haas, O.A.; Bhatia, S.; Fischer, U.; Kuhlen, M.; Gohlke, H.; Izraeli, S.; et al. JAK2 p.G571S in B-Cell Precursor Acute Lymphoblastic Leukemia: A Synergizing Germline Susceptibility. Leukemia 2019, 33, 2331–2335. [CrossRef]
- Liu, K.; Wu, Z.; Chu, J.; Yang, L.; Wang, N. Promoter Methylation and Expression of SOCS3 Affect the Clinical Outcome of Pediatric Acute Lymphoblastic Leukemia by JAK/STAT Pathway. Biomedicine & Pharmacotherapy 2019, 115, 108913. [CrossRef]
- Chhipa, R.; Singh, S.; Surve, S.; Vijayakumar, M.; Bhat, M. Doxycycline Potentiates Antitumor Effect of Cyclophosphamide in Mice. Toxicology and Applied Pharmacology 2005, 202, 268–277. [CrossRef]
- Sagar, J.; Sales, K.; Dijk, S.; Taanman, J.; Seifalian, A.; Winslet, M. Does Doxycycline Work in Synergy with Cisplatin and Oxaliplatin in Colorectal Cancer? World J Surg Onc 2009, 7, 2. [CrossRef]
- Dijk, S.N.; Protasoni, M.; Elpidorou, M.; Kroon, A.M.; Taanman, J.-W. Mitochondria as Target to Inhibit Proliferation and Induce Apoptosis of Cancer Cells: The Effects of Doxycycline and Gemcitabine. Sci Rep 2020, 10, 4363. [CrossRef]
- Zhu, C.; Yan, X.; Yu, A.; Wang, Y. Doxycycline Synergizes with Doxorubicin to Inhibit the Proliferation of Castration-Resistant Prostate Cancer Cells. ABBS 2017, 49, 999–1007. [CrossRef]
- Li, J.; Qin, Y.; Zhao, C.; Zhang, Z.; Zhou, Z. Tetracycline Antibiotics: Potential Anticancer Drugs. European Journal of Pharmacology 2023, 956, 175949. [CrossRef]
- Song, H.; Fares, M.; Maguire, K.R.; Sidén, Å.; Potácová, Z. Cytotoxic Effects of Tetracycline Analogues (Doxycycline, Minocycline and COL-3) in Acute Myeloid Leukemia HL-60 Cells. PLoS ONE 2014, 9, e114457. [CrossRef]
- Fares, M.; Abedi-Valugerdi, M.; Hassan, M.; Potácová, Z. DNA Damage, Lysosomal Degradation and Bcl-xL Deamidation in Doxycycline- and Minocycline-Induced Cell Death in the K562 Leukemic Cell Line. Biochemical and Biophysical Research Communications 2015, 463, 268–274. [CrossRef]
- Onoda, T.; Ono, T.; Dhar, D.K.; Yamanoi, A.; Nagasue, N. Tetracycline Analogues (Doxycycline and COL-3) Induce Caspase-dependent and -independent Apoptosis in Human Colon Cancer Cells. Intl Journal of Cancer 2006, 118, 1309–1315. [CrossRef]
- Ruiz-Moreno, C.; Velez-Pardo, C.; Jimenez-Del-Rio, M. Minocycline Induces Apoptosis in Acute Lymphoblastic Leukemia Jurkat Cells. Toxicology in Vitro 2018, 50, 336–346. [CrossRef]
Figure 2.
Dose-Dependent Effects and IC50 Determination of COL-3, DOX, and MIN in K562, KG-1a, and Jurkat cell lines. (A–C) Dose-response curves of COL-3, DOX, and MIN in K562 cells. (D–F) Dose-response curves of COL-3, DOX, and MIN in KG-1a cells. (G–I) Dose-response curves of COL-3, DOX, and MIN in Jurkat cells. Data are presented as mean ± standard error (SE) from three independent experiments. IC50 values were calculated using the profile likelihood method and are reported with 95% confidence intervals (CI).
Figure 2.
Dose-Dependent Effects and IC50 Determination of COL-3, DOX, and MIN in K562, KG-1a, and Jurkat cell lines. (A–C) Dose-response curves of COL-3, DOX, and MIN in K562 cells. (D–F) Dose-response curves of COL-3, DOX, and MIN in KG-1a cells. (G–I) Dose-response curves of COL-3, DOX, and MIN in Jurkat cells. Data are presented as mean ± standard error (SE) from three independent experiments. IC50 values were calculated using the profile likelihood method and are reported with 95% confidence intervals (CI).
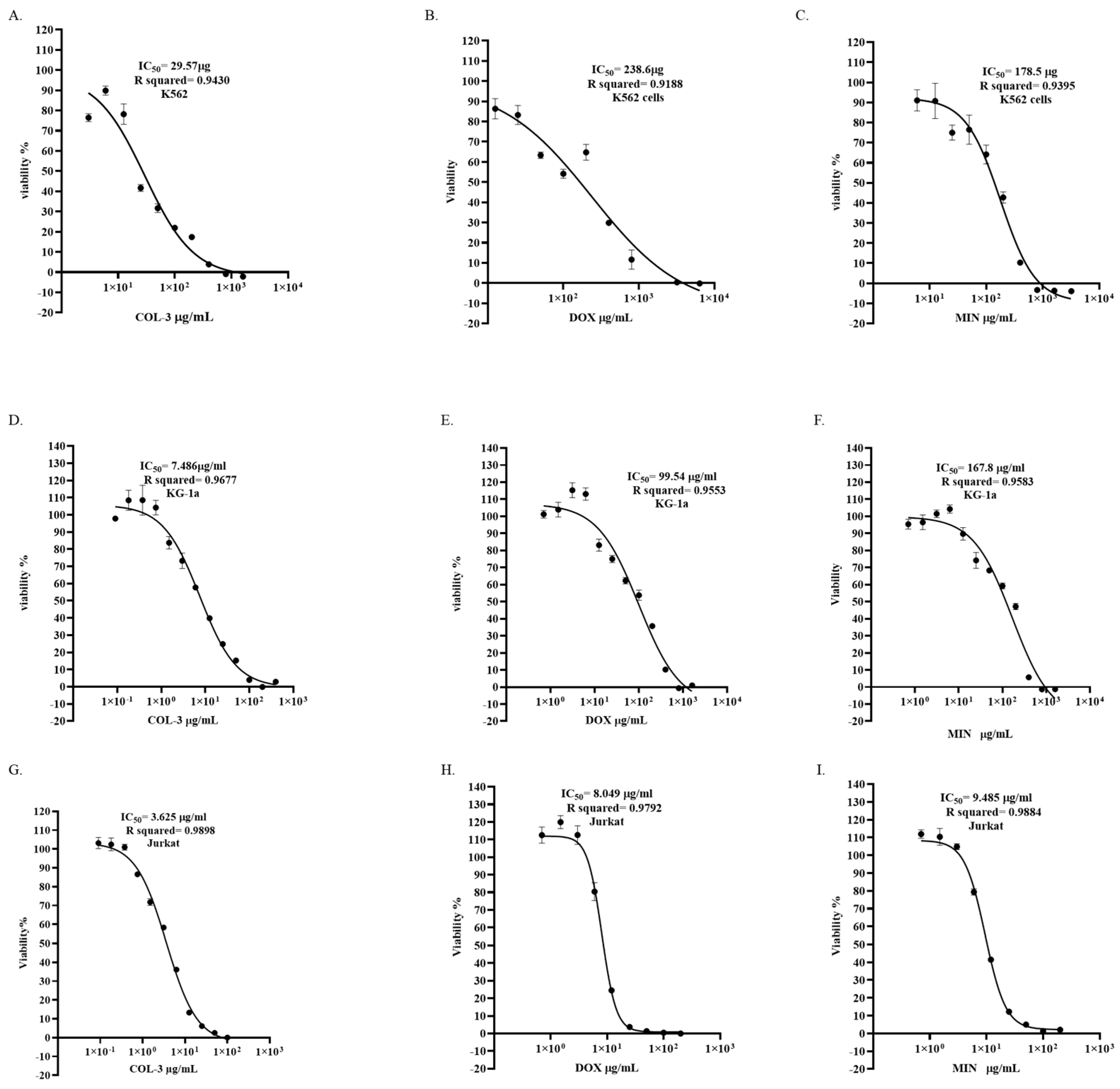
Figure 5.
Effect of COL-3, DOX, and MIN on Apoptosis in Jurkat cells assessed by Flow Cytometry. Flow cytometric analysis of Jurkat cells treated with COL-3, DOX, and MIN, showing the percentage of Annexin V–positive cells (early apoptosis), 7-AAD–positive cells (late apoptosis/necrosis), and double-stained cells (late apoptosis/necrosis). Data represent the mean ± standard error (SE) from three independent experiments. Bar graphs illustrate the percentage changes in cell populations. a: Statistically significant difference compared to 4-hour control cells (p < 0.05). b: Statistically significant difference compared to 24-hour control cells (p < 0.05).
Figure 5.
Effect of COL-3, DOX, and MIN on Apoptosis in Jurkat cells assessed by Flow Cytometry. Flow cytometric analysis of Jurkat cells treated with COL-3, DOX, and MIN, showing the percentage of Annexin V–positive cells (early apoptosis), 7-AAD–positive cells (late apoptosis/necrosis), and double-stained cells (late apoptosis/necrosis). Data represent the mean ± standard error (SE) from three independent experiments. Bar graphs illustrate the percentage changes in cell populations. a: Statistically significant difference compared to 4-hour control cells (p < 0.05). b: Statistically significant difference compared to 24-hour control cells (p < 0.05).
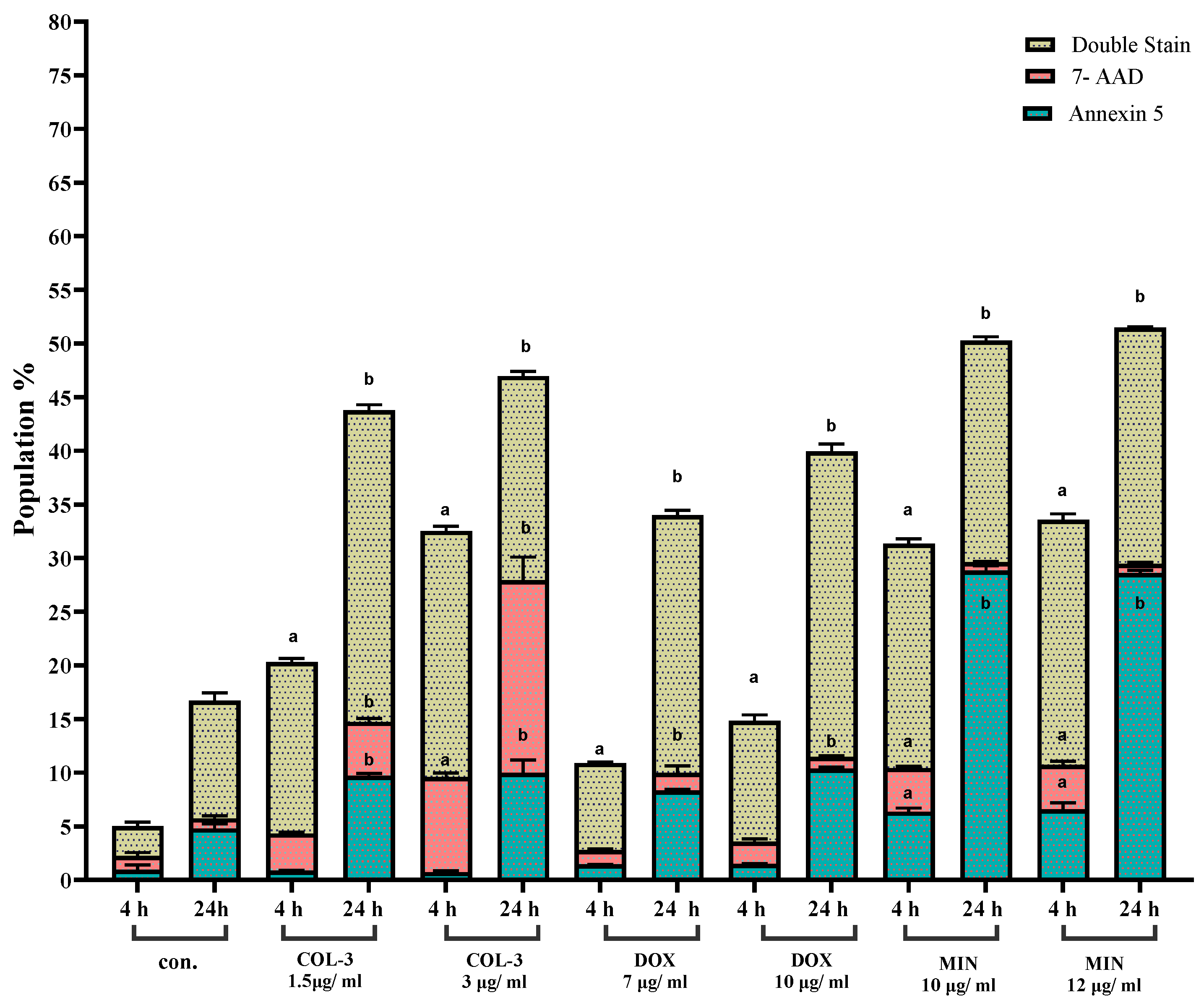
Figure 6.
The Effect of Tetracycline Analogues on the JAK2/STAT3 Pathway in K562 cells using WB. (A) Expression levels of JAK2 protein following treatment with COL-3, DOX, and MIN. (B) Expression levels of phosphorylated JAK2 (P-JAK2) after treatment with COL-3, DOX, and MIN. (C) Expression levels of STAT3 protein in response to COL-3, DOX, and MIN. (D) Expression levels of phosphorylated STAT3 (P-STAT3) following treatment with COL-3, DOX, and MIN. (E) Expression levels of pro-apoptotic protein BAX after treatment with COL-3, DOX, and MIN. (F) Expression levels of anti-apoptotic protein BCL-2 in response to COL-3, DOX, and MIN. Bar graphs represent fold changes in protein expression from at least three independent experiments (mean ± SD). Statistical significance was determined using one-way ANOVA followed by Dunnett’s multiple comparison test (*p < 0.05, ****p < 0.0001).
Figure 6.
The Effect of Tetracycline Analogues on the JAK2/STAT3 Pathway in K562 cells using WB. (A) Expression levels of JAK2 protein following treatment with COL-3, DOX, and MIN. (B) Expression levels of phosphorylated JAK2 (P-JAK2) after treatment with COL-3, DOX, and MIN. (C) Expression levels of STAT3 protein in response to COL-3, DOX, and MIN. (D) Expression levels of phosphorylated STAT3 (P-STAT3) following treatment with COL-3, DOX, and MIN. (E) Expression levels of pro-apoptotic protein BAX after treatment with COL-3, DOX, and MIN. (F) Expression levels of anti-apoptotic protein BCL-2 in response to COL-3, DOX, and MIN. Bar graphs represent fold changes in protein expression from at least three independent experiments (mean ± SD). Statistical significance was determined using one-way ANOVA followed by Dunnett’s multiple comparison test (*p < 0.05, ****p < 0.0001).
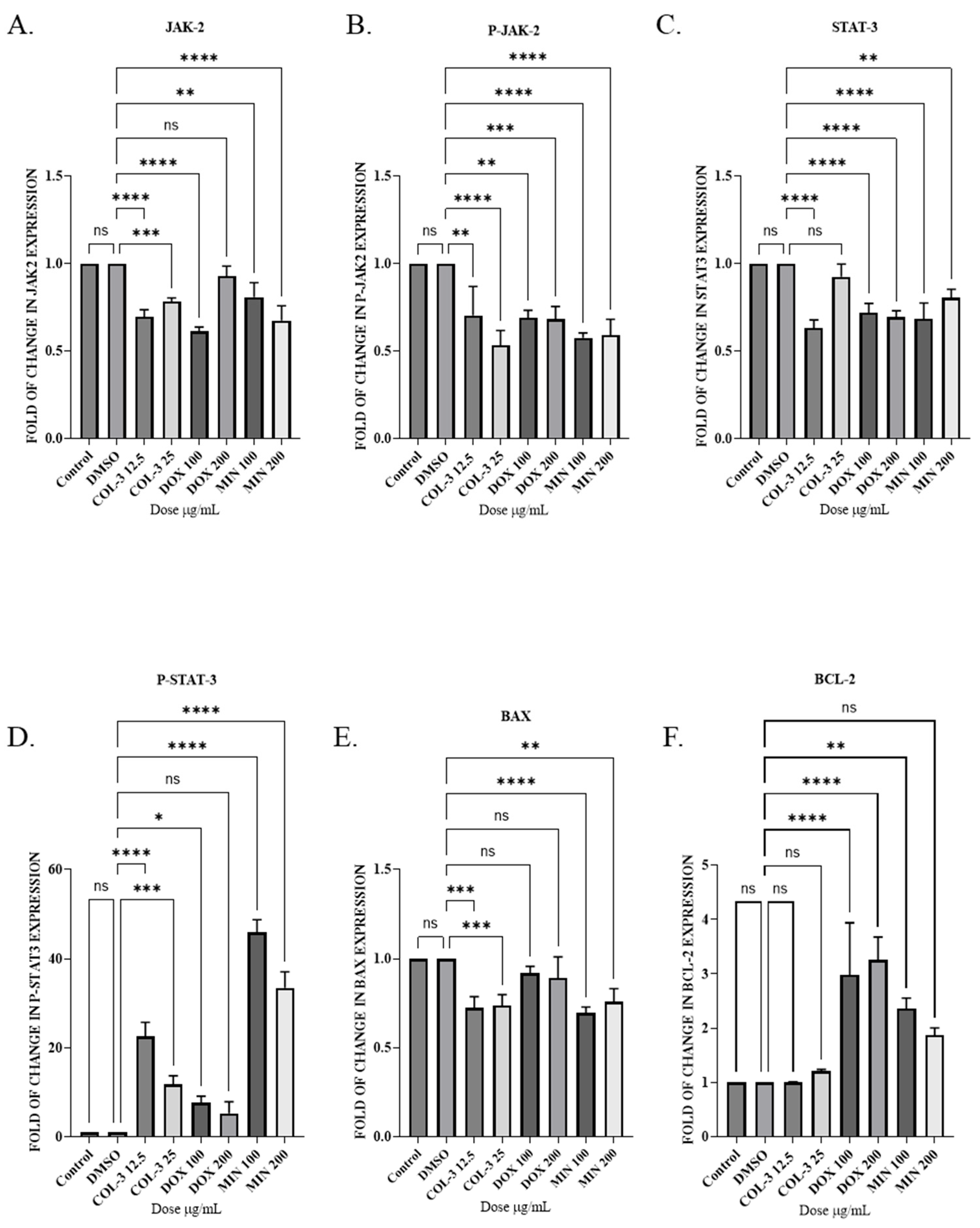
Figure 7.
The Effect of Tetracycline Analogues on the JAK2/STAT3 Pathway in KG -1a cells using WB. (A) Expression levels of JAK2 protein following treatment with COL-3, DOX, and MIN. (B) Expression levels of phosphorylated JAK2 (P-JAK2) after treatment with COL-3, DOX, and MIN. (C) Expression levels of STAT3 protein in response to COL-3, DOX, and MIN. (D) Expression levels of phosphorylated STAT3 (P-STAT3) following treatment with COL-3, DOX, and MIN. (E) Expression levels of pro-apoptotic protein BAX after treatment with COL-3, DOX, and MIN. (F) Expression levels of anti-apoptotic protein BCL-2 in response to COL-3, DOX, and MIN. Bar graphs represent fold changes in protein expression from at least three independent experiments (mean ± SD). Statistical significance was determined using one-way ANOVA followed by Dunnett’s multiple comparison test (*p < 0.05, ****p < 0.0001).
Figure 7.
The Effect of Tetracycline Analogues on the JAK2/STAT3 Pathway in KG -1a cells using WB. (A) Expression levels of JAK2 protein following treatment with COL-3, DOX, and MIN. (B) Expression levels of phosphorylated JAK2 (P-JAK2) after treatment with COL-3, DOX, and MIN. (C) Expression levels of STAT3 protein in response to COL-3, DOX, and MIN. (D) Expression levels of phosphorylated STAT3 (P-STAT3) following treatment with COL-3, DOX, and MIN. (E) Expression levels of pro-apoptotic protein BAX after treatment with COL-3, DOX, and MIN. (F) Expression levels of anti-apoptotic protein BCL-2 in response to COL-3, DOX, and MIN. Bar graphs represent fold changes in protein expression from at least three independent experiments (mean ± SD). Statistical significance was determined using one-way ANOVA followed by Dunnett’s multiple comparison test (*p < 0.05, ****p < 0.0001).
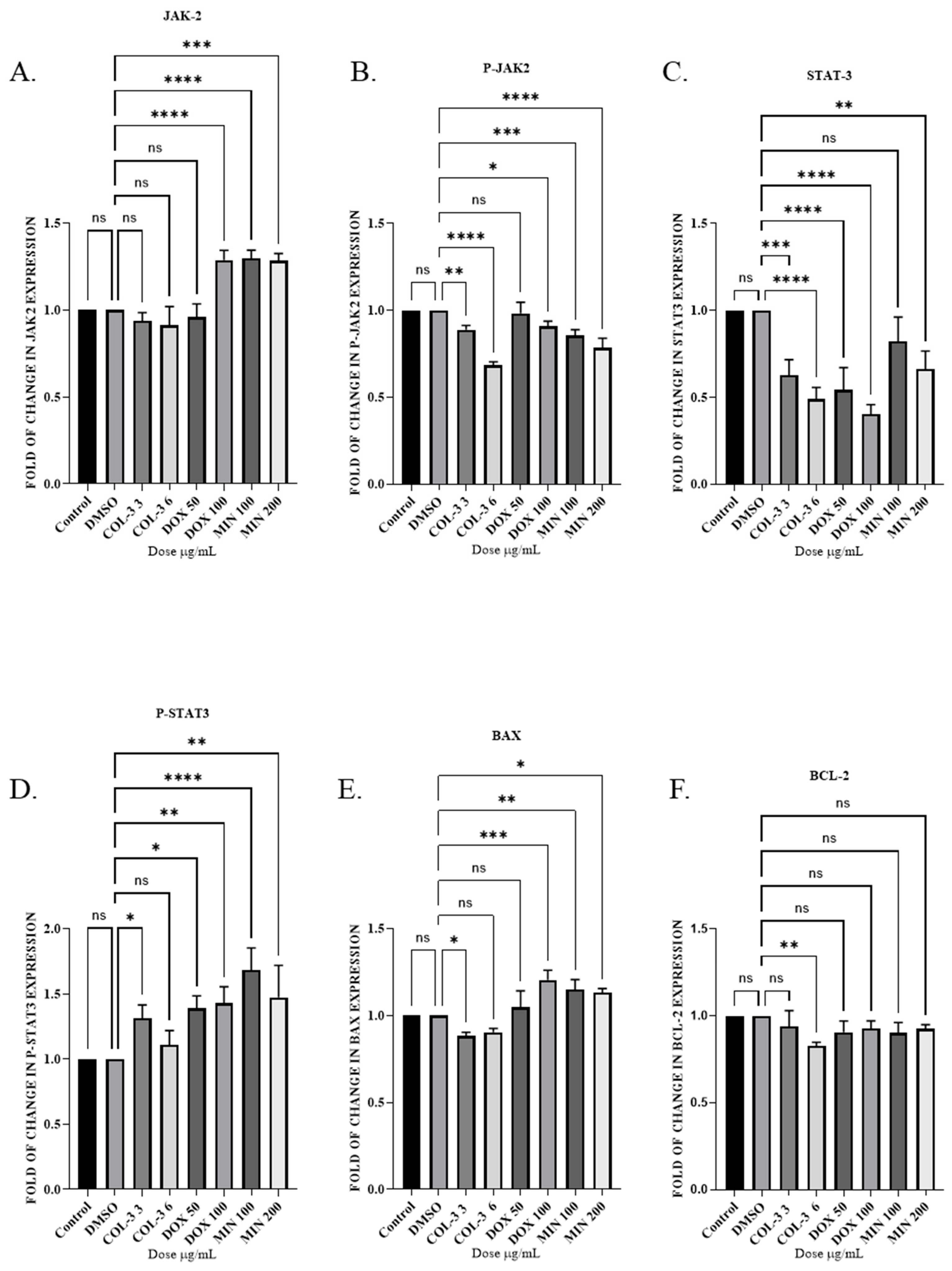
Figure 8.
The Effect of Tetracycline Analogues on the JAK2/STAT3 Pathway in Jurkat cells using WB. (A) Expression levels of JAK2 protein following treatment with COL-3, DOX, and MIN. (B) Expression levels of phosphorylated JAK2 (P-JAK2) after treatment with COL-3, DOX, and MIN. (C) Expression levels of STAT3 protein in response to COL-3, DOX, and MIN. (D) Expression levels of phosphorylated STAT3 (P-STAT3) following treatment with COL-3, DOX, and MIN. (E) Expression levels of pro-apoptotic protein BAX after treatment with COL-3, DOX, and MIN. (F) Expression levels of anti-apoptotic protein BCL-2 in response to COL-3, DOX, and MIN. Bar graphs represent fold changes in protein expression from at least three independent experiments (mean ± SD). Statistical significance was determined using one-way ANOVA followed by Dunnett’s multiple comparison test (*p < 0.05, ****p < 0.0001).
Figure 8.
The Effect of Tetracycline Analogues on the JAK2/STAT3 Pathway in Jurkat cells using WB. (A) Expression levels of JAK2 protein following treatment with COL-3, DOX, and MIN. (B) Expression levels of phosphorylated JAK2 (P-JAK2) after treatment with COL-3, DOX, and MIN. (C) Expression levels of STAT3 protein in response to COL-3, DOX, and MIN. (D) Expression levels of phosphorylated STAT3 (P-STAT3) following treatment with COL-3, DOX, and MIN. (E) Expression levels of pro-apoptotic protein BAX after treatment with COL-3, DOX, and MIN. (F) Expression levels of anti-apoptotic protein BCL-2 in response to COL-3, DOX, and MIN. Bar graphs represent fold changes in protein expression from at least three independent experiments (mean ± SD). Statistical significance was determined using one-way ANOVA followed by Dunnett’s multiple comparison test (*p < 0.05, ****p < 0.0001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



