Submitted:
13 February 2026
Posted:
27 February 2026
Read the latest preprint version here
Abstract
This article addresses a current point of contention in the field of single molecule/single particle tracking, as well as relevant literature, and supplements it with some published cell-based experiments to illustrate our conclusions and known theorems. We attempt to explain the controversy surrounding the differing biophysical and cell biological results of studies on the individual molecule and those “at the single-molecule level” as well as at the level of many molecules in such a way that even readers who are unfamiliar with the subject can understand it without having to read all the mathematical, physical, and biophysical references.
Keywords:
translational diffusion
; spectroscopy
; imaging
; super-resolution microscopy
; single molecule
; single-molecule tracking
; single-particle tracking
; individual molecule
; selfsame molecule
; thermodynamic temporal single-molecule limits in diffraction-limited and diffraction-unlimited optical systems
; hardware
1. Introduction
From a physical point of view, “particle” is a general term that covers atoms, ions, and molecules, while a molecule is a specific type of particle made up of two or more covalently bonded atoms. Chemically speaking, a molecule is a type of particle. Here, we use the term commonly used in biochemistry, namely that molecules are particles. SMT or SPT representing transformative tools in the field of life sciences and offering unparalleled insights into the dynamic behaviors of biomolecules within live cells. By capturing the movement, interaction, and localization of single molecules, SMT has revolutionized our understanding of cellular mechanisms at the nanoscale. This article provides an in-depth exploration into the principles, methodologies, applications, challenges, and future directions of SMT for live cell research.
Understanding cellular processes at the molecular level is key to unravelling the complexities of life. Unlike traditional ensemble-based methods that average behaviors across populations, SMT may uniquely captures the individuality and heterogeneity of biomolecular dynamics. The implications of these observations extend beyond fundamental biology into medical research, drug discovery, and synthetic biology, where molecular-level insights dictate advancements.
As the field evolves, SMT continues to integrate interdisciplinary approaches from mathematics, physics, chemistry, biochemistry, computer science, engineering and cell biology making it an indispensable tool for life scientists. This detailed overview seeks to unravel the theoretical underpinnings, technical advancements, and transformative applications of SMT, while critically examining existing challenges and providing a glimpse into future innovations.
The paper also serves to provide a basic understanding of highly sophisticated measuring instruments and methods for evaluating measurement data or evaluation algorithms independent of the manufacturers and software developers or providers, which is important for cell biologists and molecular biologists who are not primarily trained in physics and optics (see section 3. Some Challenges in Single-Molecule Tracking).
Since the topic is too broad for a single article, we hope that this paper will convey the excitement surrounding the very latest achievements in the field of single-molecule tracking for live cells.
2. Fundamentals of Single-Molecule Tracking
2.1. What is SMT?
The SMT involves the visualization and analysis of single molecules within living systems. SMT reveals rare events at the molecular level. It leverages advanced optical tools and computational methods to follow the trajectory of single molecules over time. At its core, SMT provides quantitative metrics, such as diffusion coefficients, binding rates, and molecular dwell times, which are crucial for understanding dynamic processes. This method solves longstanding questions regarding molecular interactions, localization, and their role in cellular function. For instance, SMT allows to deconstruct the stochastic movement of receptors on cellular membranes or analyze protein transport within the cytoskeleton framework.
2.2. Historical Context
The concept of tracking individual molecules emerged in the late 20th century, driven by advancements in fluorescence microscopy and biophysics. Milestones include the introduction of fluorescent tagging and high-resolution detectors that enabled researchers to overcome the diffraction limit of light.
Foundational work by Albert Einstein on Brownian motion laid the theoretical groundwork for understanding molecular dynamics [1]. However, it was not until the advent of fluorescence-based techniques, such as fluorophore tagging and Förster resonance energy transfer (FRET), that scientists could directly observe single molecules. Early pioneers, such as W. E. Moerner and E. Betzig, revolutionized the field by demonstrating single-molecule fluorescence in biological systems, earning them the Nobel Prize in Chemistry [2,3].
In 1989, W. E. Moerner became the first scientist to measure the light absorption of a single molecule at liquid helium temperatures (1.6 Kelvin corresponding to -257.15 degree Celsius) [4]. In this case, it was a pentacene molecule in a p-terphenyl host crystal and the first optical detection and spectroscopy of single molecules in the condensed phase.
In 1997, W. E. Moerner spread a variant of GFPs all over a gel. The distance between the individual proteins after immobilization was greater than Abbe’s diffraction limit of 0.2 µm [5]. The protein fluoresced at an excitation wavelength of 488 nm. After some time, the fluorescence decreased. Light with a wavelength of 405 nm reactivated the protein, whereupon it fluoresced again at an excitation wavelength of 488 nm. Because the protein molecules were only sparsely distributed, the glow of individual molecules could be observed with a conventional microscope. The fluorescence of individual molecules was controlled, thus solving a problem that Eric Betzig had formulated in 1995 [6]. In 2006, Eric Betzing excited proteins to fluorescence by means of a laser pulse. Because the pulse was very weak, only a fraction of the proteins began to fluorescence. Due to their small number, almost all protein molecules were located at a distance greater than the Abbe diffraction limit of 0.2 μm. This allowed the position of each glowing protein to be determined very precisely under the microscope. After a certain time, the fluorescence faded, and a new subset of proteins was excited by a laser pulse. This procedure was then repeated. The images taken were superimposed and a super-resolution image of the lysosome membrane was obtained far better than Abbe’s diffraction limit [3]. In the biological and medical sciences, single molecules are one of the cutting-edge technologies in methodologies and far-reaching applications. Single molecule tracking and single molecule analysis, respectively, revolutionized how researchers view problems such as molecular motor diseases, protein folding diseases and enzymatic kinetics just to name a few.
Stefan Hell, who also received the Nobel Prize in Chemistry, theoretically proposed a method called stimulated emission depletion (STED) in 1994 [7]. One light pulse excited all fluorescent molecules, while another light pulse quenched the fluorescence of all molecules except those in a nanometer-sized volume in the center and the fluorescence of this volume was recorded. By scanning the sample and continuously measuring the light intensity, a comprehensive image was obtained. The smaller the volume that fluoresces at a given time, the higher the spatial resolution of the resulting image. Therefore, optical microscopes were no longer inherently limited by a spatial resolution limit. In 2000, Stefan Hell proved that his ideas worked in practice [8].
The search for ways to utilize the optical signals of individual molecules began in the three research groups through the application of single-molecule fluorescence. However, the beginning of the investigation of single molecules is primarily attributed to Professor W. Moerner. Based on the discoveries of these three research groups, we can now study biochemistry “at the single-molecule level” without immobilization on biological or artificial surfaces (e.g., cover slips) and without hydrodynamic focussing [9]. Biochemistry and chemistry traditionally deal with large quantities of molecules and their effects. Most research groups neglect the thermodynamic jitter of molecules/particles in their so-called single-molecule or single-particle studies, or in their studies “at the single-molecule level” in dilute liquids or living cells without immobilization or hydrodynamic flow. The time resolution of translational diffusion for a single molecule/particle (an individual molecule/particle) in dilute liquids and living cells, including membranes, is also referred to as “meaningful time Tm” in order to separate the single molecule/particle in time from another single molecule/particle of the same kind without immobilizing it on solid phases or membranes and without hydrodynamic flow [10]. The temporal resolution of freely difussing individual molecules (single molecules) in dilute solutions (liquids), membranes of live cells depends on the concentration of molecules of the same kind in the bulk phase [10,11]; it is the reference measure for all time-resolved calculations/algorithms based upon parameter sets of (specific) measurement techniques, for example, photon counting statistics, Nyquist limit, ‘single-molecule FRET’, etc., in order to ultimately justify whether or not a single molecule/particle (individual molecule/particle, self-same molecule/particle) was measured during the observation/detection time [11,12]. To be clear, this work in the field of single-molecule biophysics deals with the observation and analysis of the individual molecule, meaning one and the same molecule, in solutions and membranes as well as in living cells without immobilization or hydrodynamic focusing. For this reason, the concept of the “re-entries” into the detection/observation volume and the concept of the “meaningful time Tm” were introduced to analyze specifically the behavior of the individual molecule over longer periods of time when it repeatedly re-enters the detection/observation volume [13]. In other words: Only one and the same molecule diffuses into and out of the detection/observation volume during the meaningful time Tm, which must be the measurement time. These original articles emphasize the study of individual molecules in their natural state, without the need for surface immobilization or the use of external forces like hydrodynamic focusing. The articles developed theoretical frameworks for detecting individual molecules, including the use of the thermodynamic signature of a single molecule or particle (thermodynamic jitter) in dilute liquids and living cells [14]. Thus, they laid the foundation for single-molecule biophysics and biochemistry based on the stochastic nature of diffusion (thermodynamic jitter). These articles suggested that the light (photons) we perceive from individual molecules is not just a simple quantum emission but also reflects the random thermal motion (thermodynamic jitter) of these molecules in their environment, making photon statistics a secondary “epiphenomenon” of the fundamental molecular motion. Essentially, the irregular motion of the individual molecule changes its position relative to the detector or light field, thereby influencing when and how photons are emitted or detected, making the photon signal an indicator of the underlying thermodynamic fluctuations [11]. However, nearly all single-molecule studies in dilute solutions or living cells without immobilization on surfaces or hydrodynamic focusing are not based exclusively on facts about the individual molecules or puzzle out each single molecule (individual molecule) within the surrounding phase (bulk phase). Combining single-molecule methods with ensemble averages yields interesting results. Bridging approaches utilizing ensemble averages are currently being implemented.
2.3. Key Techniques
Two opposing views can be found in both recent and older articles. One focuses on fluctuations in molecular motion and concludes that, for example, no significant thermodynamic jitter occurs at -233 °C (see above). The other view, which corresponds to the prevailing opinion, ignores thermodynamic jitter such as Brownian motion when it comes to the temporal resolution of measurement or investigation methods, or their calculation [15]. Therefore, single-molecule studies do not always refer exclusively to details of the individual molecules or interpret each molecule in the context of the bulk phase [13], which is particularly true for single-molecule/single-particle tracking experiments. The bridge between these two perspectives is built by combining approaches ‘at the single molecule level’ with ensemble averages, which, as shown here, leads to stimulating and challenging outcomes and could differ from those obtained at the level of many molecules [16].
SMT/SPT are quantitative and non-invasive tools for monitoring the movement and interactions of labeled protein molecules within living cells. SMT/SPT have been extended to study the temporal dynamics. SMLM improves the spatial resolution capability of optical imaging to examine length scales below the diffraction limit of light. It comprises techniques of super-resolution imaging that enable visualizing the distribution of proteins with high precision. Currently, the lateral resolution is typically in the range of ∼30-20 nm, compared to the resolution limit of ∼250 nm in conventional microscopy. Newer SMLM-based methods such as MinFlux achieve a 3D spatial resolution of 1–3 nm [17,18,19].
SMT and SMLM generate point clouds of spatial data points. These point clouds consist of highly precise coordinates representing the positions of single molecules within a biological sample. This point cloud data format is a fundamental feature of these techniques, enabling an unprecedented visualization of molecular organization and dynamics that is not possible with conventional microscopy.
In the latest literature, some authors consider SMT to be a subset/subfield or example of SMLM techniques [20].
2.4. Core Principles
A conceptual framework for thinking about crowded environment like biological cells and their compartments is that anomalous subdiffusive motions of macromolecules can occur due to geometric and/or energetic disorder [21,22]. The motions of proteins, RNAs or DNA can be hindered either by molecular crowding modeled by spatial, geometric restraints of fractals or by chemical binding represented by continuous-time random walks with heavy tails [23]. This has been coined a coexistence of an ergodic anomalous diffusion and a nonergodic process of dynamics of macromolecules in living cells and their complex environment. Deciphering the molecular nature of the coexistence of ergodic anomalous process and nonergodic behavior constitutes an important quest of the field. In principle, widely used experimental methods such as fluorescence fluctuation spectroscopy and imaging like fluorescence correlation spectroscopy reveal only apparent values of the diffusion coefficients [24].
- (i)
- Fluorescent Labeling
Various methods for fluorophore labeling have been developed, such as fluorescent proteins, fluorophore-labeled ligands or nanobodies, and self-labeling protein tags such as HaloTag or SNAP-Tag (see [25]). The development of advanced fluorophores has dramatically enhanced SMT capabilities [26,25]. Organic dyes, quantum dots, and genetically encoded fluorescent proteins offer brightness and photostability, critical for tracking in dynamic environments.
Quantum dots have revolutionized SMT due to their superior photostability and brightness [27]. The nanoscale semiconductor particles enable prolonged imaging sessions, allowing researchers to monitor molecular dynamics spanning several minutes or hours.
Moreover, genetically encoded fluorescent proteins, such as mCherry and eGFP, provide specificity, enabling the tagging of intracellular targets without altering their native function.
Site-specific, bio-orthogonal dye-labeling of proteins in vivo is used for SMT due to their superior photophysical properties compared to fluorescent proteins [28].
- (ii)
- High-Resolution Microscopy
Techniques like TIRF microscopy and confocal microscopy are utilized for precision imaging (see [29,30]). These methods reduce background noise and improve spatial resolution. Super-resolution techniques, such as STORM and PALM, have pushed the boundaries of spatial resolution, enabling the visualization of subcellular structures [20].
Wide-field microscopy illuminates the whole sample simultaneously, capturing a broad 2D view. It is ideal for thin samples such as cell cultures and uses either transmitted light (brightfield, phase contrast) or fluorescence light (fluorophores absorb and emit light). It is simple, fast, and excellent for live-cell microscopy and SMT [31].
Recent innovations in light-sheet microscopy have further enhanced SMT by providing three-dimensional spatial resolution while minimizing phototoxicity [32]. These methods allow researchers to track molecules in thicker samples, such as organoids or tissue slices, expanding SMT applications beyond single cells.
- (iii)
- Two-Color Single-Molecule Tracking
Two-color SMT is a fluorescence microscopy-based method for detecting and quantifying specific protein-protein interactions at the single-molecule level, which offers high sensitivity to heterogeneities and rare events [33,34].
- (iv)
- Tracking Algorithms
Computational methods are employed to extract trajectories from noisy data and stochastic movement. Advanced algorithms account for drift correction, localization precision, and trajectory mapping [35].
- (v)
- Machine Learning Integration
Artificial intelligence and machine learning algorithms are increasingly used to analyze SMT datasets [36]. These tools improve the accuracy of high-precision coordinates (point cloud, trajectory mapping) of proteins or other labelled targets in a sample and uncover patterns that were previously undetectable in SMLM [37]. Machine learning plays a pivotal role in distinguishing signal from noise in SMT experiments [38]. Neural networks trained on experimental datasets can identify molecular trajectories, classify movement patterns, and predict cellular interactions. These computational advancements streamline the analysis process and open avenues for mining large-scale SMT data [39].
3. Some Challenges in Single-Molecule Tracking
Currently, most research groups use devices that enable direct tracking for SMT. This approach is described in the literature as superior to other methods and offers a flexible tool, as generally shown in Figure 1. From a biophysical perspective, quantitative single-molecule applications with a time resolution of currently up to 40 ms are possible, as shown in Figure 2, which is generally independent of the device manufacturer.
beads was tracked at 535 nm. The scanning mirror circled the bead and Fourier transformed to indicate a direction of the bead’s next move and followed. The z-direction was tracked up and down of this circled xy-direction pattern and determined which direction the bead’s next z-direction moved. The units in the x-pos, y-pos and z-pos were in micrometer. The full tracking time was 500 seconds. This figure is from the ISS laboratory and illustrates our known conclusions or theorems.
In the literature, trajectories of single molecules are initially analyzed in a manner like that described in [40], but the particle motion is considered in three dimensions [41] rather than two. The equations consider normal (Brownian) diffusion in three dimensions [42]. For the Brownian motion of the spheres (see Figure 2), a D value of (apparent diffusion coefficient) was estimated, which was certainly overestimated; this example is intended to illustrate a problem in the literature with quantitative measurements, namely when the Brownian molecular motion is superimposed by other physical motions (here superdiffusion). Turbulent flow occurred when the particle solution was injected into the sample-well or when attempting to mix it using pipettes by infusion/withdrawal. The turbulent flow was even visible to eyes in the binocular image.
Since the start and end points of 16 consecutive tracks were close together (see Figure 2), they were usually combined into a single track, as shown in Figure 3. The individual tracks represented independent measurements. The Figure 3 clearly shows the entire path from start to finish as a single path. In the subsequent analysis step, different lengths of random walks for the total single track were generated and average over n = 1000, where n is the number of different lengths in the calculation (see in the literature [43]). We processed the measurement data in the original units (μm). To check the assumption of a Brownian particle the mean quadratic deviation of a track is calculated in the literature [43], which means the average
where is the origin and is the end point of a track [43]. The datasets were subsequently assembled and sorted. This yielded 261 data points, which are depicted in Figure 4.
The graph of Figure 4 demonstrates the various observations, as explained in the legend. In our example illustrating conclusions on the data evaluation of Brownian motion in the current literature (see also Section 4.1. Protein dynamics), we divided the data into groups according to certain features [44]. To be clear, the experimentally obtained data were used and statistical averaging with variation in the “step length” was performed as described in [43], i.e., in a very traditional way. No simulation of the path was performed; instead reference is made exclusively to the measurements provided. The statistical averaging and calculation of the “standard deviation” yielded the diffusion coefficient as the slope in a mathematical least squares method. When converting per unit of time, taking superdiffusion (β = 1.06 in Figure 4) into account, a value of was estimated. It is in the order of magnitude of the D value calculated without correction for turbulent flow but is a better apparent estimate than the distorted value of (a Bayesian optimized parameter selection). For diameter beads (radius ), the calculation using the Stokes-Einstein equation with low Reynolds number yields a diffusion coefficient of approximately in deionized water at room temperature (approximately 300 Kelvin or 27 °C, see also [45]).
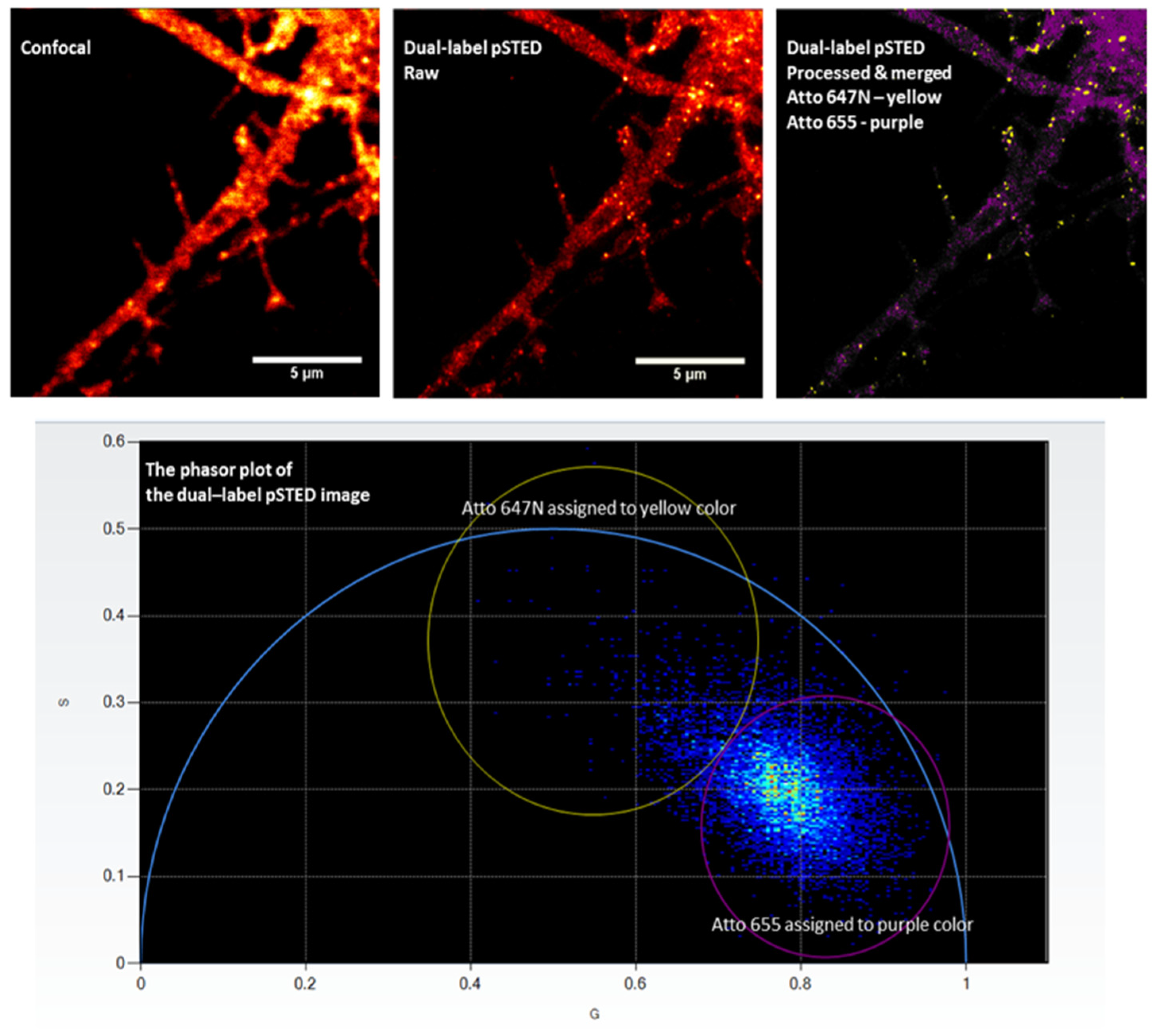
An additional increase on the spatial resolution is provided by STED as illustrated for this review in Figure 5 and the lifetime information acquired through the digital frequency domain lifetime imaging (FastFLIM) and the phasor plot (see [46]). The photons emitted by the fluorophores are separated from the photons of the fluorophores with shorter decay time partially or fully activated by the STED excitation laser. A spatial resolution of 30 nm was measured on standard samples. The combination of the pSTED and the lifetime analysis using phasor diagrams enables dual labeling excitation (see Figure 6). This means that it is possible to distinguish between two fluorophores with similar emission patterns but different decay times [47].
4. Applications of Single-Molecule Tracking Techniques in Live Cells
4.1. Protein Dynamics
SMT has been pivotal in elucidating protein-protein interactions, conformational changes, and their role in signaling pathways [48]. For example, studies on kinase activation have provided insights into signal transduction mechanisms at nanometer resolution [49]. Another example is the study of the dynamics of epidermal growth factor receptor (EGFR) dynamics, where SMT revealed heterogeneity in receptor activation and their clustering patterns on cell membranes [50]. Such insights are critical for understanding cancer biology and potential therapeutic targets. The detection of moving particles using the likelihood ratio and subsequent subpixel localization in living cells by SMT revealed how biomolecules move in crowded intracellular environments; most tracking software programs assume Brownian motion, an approximation that fails in the case of anomalous diffusion [51].
4.2. Membrane Transport
Tracking single molecules has shed light on vesicular trafficking, endocytosis, and exocytosis (see [52]). SMT has revealed the stochastic nature of vesicle docking and fusion events. Additionally, SMT in synaptic vesicle dynamics has unravelled the mechanisms behind neurotransmitter release and recycling, providing a deeper understanding of neuronal communication (see [53]).
4.3. Drug Discovery
By observing the binding and dissociation of drug molecules to their targets in real-time, SMT contributes to the development of novel therapeutic agents. For example, SMT has been employed to study the binding of smaller molecules to nuclear ribonucleoproteins enhancing the pharmacological profiling of candidate drugs by robing an endogenous regulatory network in live cells [54].
4.4. RNA Dynamics
SMT has uncovered the movement and localization of mRNA within the cellular cytoplasm, providing a deeper understanding of translation regulation. Recent studies using SMT have explored how RNA granules move within neuronal cells, shedding light on their role in synaptic plasticity and learning [55].
5. Present Demands in Single-Molecule Tracking
5.1. Photobleaching and Phototoxicity
The fluorescence labeling required for SMT can lead to photobleaching of dyes and cellular damage, limiting observation time [56]. Minimizing laser intensity or developing novel fluorophores with reduced photobleaching properties represents an active area of research. Fluorescence-regenerative markers represent a promising solution to this problem, but they impose stricter requirements on signal contrast and labeling kinetics. The systematic development of fluorogenicity and reversible binding has created a template for the development of an innovative, bright, and regenerative array marker that has been used in STED nanoscopy and SMT on living cells [57].
5.2. Complexity of Cellular Systems
The dense and heterogeneous environment of live cells poses challenges for molecule identification and trajectory reconstruction [24,58]. Strategies such as multi-color imaging [59] and deconvolution algorithms [60] have emerged to address these complexities, providing a clearer representation of molecular interactions in crowded cellular environments (see [61,62]). Computational modeling was used for kinetic Monte Carlo simulations after image processing and data anlaysis in the SMT application of interferometric scattering microscopy [63].
5.3. Data Analysis Bottlenecks
Fluorescence from single molecules can be distinguished from the small background associated with a femtoliter of solvent [64]. At a solution concentration of approximately 1 nM and less at room temperature, the probability that an analyte molecule is in the probe volume is less than one [65,66]. However, we have pointed out that fluorescence correlation spectroscopy (FCS) or two-color (dual color) fluorescence cross-correlation spectroscopy (FCCS), with today’s temporal resolution and sensitivity, is only suitable for measuring individual molecules/particles, i.e., for measuring the same molecule/particle, with measurement times of only a few milliseconds [10,67]. Although the fluorescence of individual molecules can be detected, in the case of fluorescence fluctuations spectrometry, the autocorrelation and two-color (dual color) cross-correlation functions are recorded and analyzed [68]. FCS/two-color (dual collor) FCCS provide averaged data, and the conclusions apply only to systems with multiple molecules (many molecules). Properties of a single molecules (an individual molecule) are not obtained [1]. We have called for increased sensitivity in the time domain of measurements [14].
Processing large amounts of SMT data requires sophisticated algorithms and computational power, which can be resource intensive. Cloud-based solutions and parallel computing frameworks are being integrated into SMT workflows to handle high-throughput data analysis efficiently [69].
5.4. Plasmonic Platforms
DNA-based plasmonic nanostructures have established themselves as powerful tools for the optical signal detection of single molecules due to their unique self-organization properties and their excellent optical performance (see [70]). Plasmonic effects during electrochemical reactions were recently studied by single-molecule electrochemiluminescence microscopy [71]. A time-resolved deep-UV confocal microscope for autofluorescence correlation spectroscopy (UV-FCS) was developed [72]. Aluminium zero-mode waveguides, UV horn antennas, and resonant rhodium dimer nanoslit antennas were used to enhance protein autofluorescence by integrating these nanophotonic techniques with antioxidants and background reduction. A multiplex biosensor that enables continuous monitoring using plasmon-enhanced single-molecule fluorescence was investigated at the single-molecule level [73].
6. Future Directions
6.1. Multi-Color SMT
New strategies allow for simultaneous tracking of multiple types of molecules, enabling the study of complex interactions in space and time [74]. This advancement will enable researchers to observe molecular networks and signaling cascades in real-time, providing holistic insights into cellular functionality.
6.2. Non-Labeling Methods
Emerging label-free imaging techniques, such as scattering-based methods, may reduce phototoxicity and offer new avenues for SMT (see [75]). Techniques such as Raman scattering and holographic imaging present promising paths forward, eliminating the need for external labels while preserving molecular integrity. Interferometric imaging on a microfiber surface to provide near-field illumination broadens the applications of label-free methods [76].
6.3. Integration with Systems Biology
7. Conclusions
SMT has emerged as a cornerstone technology in life sciences, offering unprecedented insights into live-cell dynamics. By overcoming technical challenges and integrating cutting-edge innovations, SMT continues to expand our scientific horizons. The implications of this technology extend from fundamental biology to clinical applications, promising breakthroughs in the quest to decode the complexities of life. Here, we show a way through the maze that surrounds individual molecules/particles interpretations in living cells. We attempt to flesh out the controversy so that the uninitiated reader can understand it without having to go and read all the references.
Supplementary Materials
The following supporting information can be downloaded at: https://www.youtube.com/watch?v=MkdPAGxfI14, Video S1: Fixed HeLa cell, Mitochondria labeled with Atto 647N.
Author Contributions
Conceptualization, Z.F.-P.; methodology, Z.F.-P, G.B., B.B., S.-C.L.; writing—original draft preparation, Z.F.-P.; writing—review and editing, Z.F.-P., G.B., S.-C.L., B.B; supervision, Z.F.-P., G.B., S.-C.L., B.B. All authors have read and agreed to the submitted version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data are presented in the review article. The Figures are from the ISS laboratory and illustrate our known conclusions or theorems.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
The following abbreviations are used in this article:
| SMT | Single-Molecule Tracking |
| SPT | Single-Particle Tracking |
| SMLM | Single-Molecule Localization Microscopy |
| STORM | Stochastic Optical Reconstruction Microscopy |
| PALM | Photo-Activated Localization Microscopy |
| MinFlux | Minimal photon Flux |
| FRET | Förster Resonance Energy Transfer |
| TIRF | Total Internal Reflection Fluorescence microscopy |
| FLIM | Fluorescence Lifetime Microscopy |
| FCS | Fluorescence Correlation Spectroscopy |
| FCCS | Fluorescence Cross-Correlation Spectroscopy |
| SPSM-FCS | Single-Phase Single-Molecule Fluorescence Correlation Spectroscopy (autocorrelation and two-color (dual color) cross-correlation) |
| UV | Ultraviolet |
| IR | Infrared |
| NIR | Near-Infrared |
| KHz | Kilohertz, a unit of frequency, representing 1,000 cycles or oscillations per second |
| MSD | Mean Squared Displacement |
| D | Diffusion coefficient |
| 3D | Three dimensional |
| 2D | Two dimensional |
| EGFR | Epidermal Growth Factor Receptor |
| mCherry | Member of the mFruits family of monomeric red fluorescent proteins |
| eGFP | Enhanced Green Fluorescent Protein |
| DNA | Deoxyribonucleic Acid |
| RNA | Ribonucleic Acid |
| mRNA | Messenger Ribonucleic Acid |
References
- Földes-Papp, Z.; Baumann, G.; Kinjo, M.; Tamura, M. Single-Phase Single-Molecule Fluorescence Correlation Spectroscopy (SPSM-FCS). Distinguished article entry. In Encyclopedia of Medical Genomics & Proteomics; Fuchs, J., Podda, M. Editors, Eds.; Taylor & Francis, New York, USA, 2005, pp. 1-7.
- Moerner, W. E. Nobel Lecture: Single-molecule spectroscopy, imaging, and photocontrol. Reviews of Modern Physics 2015, 87, 1183-1212.
- Betzig, E.; Patterson, G. H.; Sougrat, R.; Lindwasser, O. W.; Olenych, S.; Bonifacino, J. S.; Davidson, M. W.; Lippincott-Schwartz, J.; Hess, H. F. Imaging intracellular fluorescent proteins at nanometer resolution. Science 2006, 313, 1642-1645.
- Moerner, W.E.; Kador, L. Optical detection and spectroscopy of single molecules in a solid. Physical Reviews Letters 1989, 62, 2535-2538.
- Dickson, R. M.; Cubitt, A. B.; Tsien, R. Y.; Moerner, W. E. On/Off blinking and switching behavior of single molecules of green fluorescent protein. Nature 1997, 388, 355–358.
- Betzig, E. Proposed Method for molecular optical imaging. Optics Letters 1995, 20, 237-239.
- Hell, S. W.; Wichmann, J. Breaking the diffraction resolution limit by stimulated emission: stimulated-emission-depletion fluorescence microscopy. Optic Letters 1994, 19, 780-782.
- Klar, T. A.; Jakobs, S.; Dyba, M.; Egner, A.; Hell, S. Fluorescence microscopy with diffraction resolution barrier broken by stimulated emission. Proceedings of the National Academy of Sciences 2000, 97, 8206-8210.
- Földes-Papp, Z. Individual macromolecule motion in a crowded living cell. Current Pharmaceutical Biotechnology 2015, 16, 1-2.
- Földes-Papp, Z. Fluorescence Fluctuation Spectroscopic Approaches to the Study of a Single Molecule Diffusing in Solution and a Live Cell without Systemic Drift or Convection: A Theoretical Study. Current Pharmaceutical Biotechnology 2007, 8, 261-273.
- Földes-Papp, Z. Single-molecule time resolution in dilute liquids and live cells at the molecular scale: Constraints on the measurement time. American Journal of Translational Medicine 2021, 5, 154-165.
- Földes-Papp, Z. The thermodynamic signature of a single molecule or a single particle in dilute liquids and live cells: Single-Molecule Biophysics & Biochemistry based on the stochastic nature of diffusion. American Journal of Translational Medicine 2023, 7, 74-77.
- Földes-Papp, Z. Measurements of Single Molecules in Solution and Live Cells Over Longer Observation Times Than Those Currently Possible: The Meaningful Time. Current Pharmaceutical Biotechnology 2013, 14, 441-444.
- Baumann, G.; Földes-Papp, Z. Single-Molecule Tracking in Live Cell without Immobilization or without Hydrodynamic Flow by Simulations: Thermodynamic Jitter. Biophysica 2024, 3, 442-452.
- Baumann, G.; Földes-Papp, Z. Study on Single-Molecule Biophysics and Biochemistry in dilute liquids and live cells without immobilization or significant hydrodynamic flow: The thermodynamic Single-Molecule DEMON. Current Pharmaceutical Biotechnology 2022, 23, 1750-1757.
- Tang, W.; Fuentenebro Navas, D.; Vermeer, B.; Schmid S. Disentangling a Complex Biomolecular World with Singel-Molecule Resolution. Chimia 2025, 79, 770-779.
- F. Balzarotti, F.; Eilers, Y.; Gwosch, K. C.; Gynna, A. H.; Westphal, V.; Stefani, F. D.; Elf, J.; Hell, S. W. Nanometer resolution imaging and tracking of fluorescent molecules with minimal photon fluxes. Science 2017, 355, 606–612.
- Gwosch, G. K.; Pape, J. K.; Balzarotti, F.; Hoess, P.; Ellenberg, J.; Ries, J.; Hell, S. W. Minflux nanoscopy delivers 3d multicolor nanometer resolution in cells. Nature Methods 2020, 17, 217–224.
- Schultze, S.; Grubmüller, H. Bayesian MINIFLUX localization microscopy. Cornell University arXiv:2510.25654 [physics.comp-ph] 2025, (1-5).
- Driouchi, A.; Bretan, M.; Davis, B. J.; Heckert, A.; Seeger, M.; Silva, M. B.; Forrest, W.S.R.; Hsiung, J.; Tan, J.; Yang, H.; McSwiggen, D.T.; Song, L.; Sule, A.; Abaie, B.; Chen, H.; Chhun, B.; Conroy, B.; Elliott, L. A.; Gonzalez, E.; Ilkov, F.; Isaacs, J.; Labaria, G.; Lagana, M.; Larsen, D. D.; Margolin, B.; Nguyen, M. K.; Park, E.; Rine, J.; Tang, Y.; Vana, M.; Wilkey, A.; Zhang, Z.; Basham, S.; Ho, J. .J; Johnson, S.; Klammer, A. A.; Lin, K.; Darzacq, X.; Betzig, E.; Berman, R.T.; Anderson, D.J. Oblique line scan illumination enables expansive, accurate and sensitive single-protein measurements in solution and in living cells. Nature Methods 2025, 22, 559-568.
- Meroz, Y.; Sokolov, I.M.; Klafter, J. Subdiffusion of mixed origins: When ergodicity and nonergodicity coexist. Physical Review E 2010, 81, 0101001.
- Földes-Papp, Z.; Baumann, G.; Li, L.-C. Visualization of subdiffusive sites in a live single cell. Journal of Biological Methods 2021, 8, e142 (1-8).
- Földes-Papp, Z.; Baumann, G. Fluorescence molecule counting for single-molecule studies in crowded environment of living cells without and with broken ergodicity. Current Pharmaceutical Biotechnology 2011, 12, 824-833.
- Baumann, G.; Place, R. F.; Földes-Papp, Z. Meaningful interpretation of subdiffusive measurements in living cells (crowded environment) by fluorescence fluctuation microscopy. Current Pharmaceutical Biotechnology 2010, 11, 527-543.
- Catapano, C.; Dietz, M. S.; Kompa, J.; Jang, S.; Freund, P.; Johnsson, K.; Heilemann, M. Long-Term Single-Molecule Tracking in Living Cells using Weak-Affinity Protein Labeling. Angewandte Chemie International Edition 2025, 64, e202413117 (1-8).
- König, A. I.; Sorkin, R.; Alon, A.; Nachmias, D.; Dhara, K.; Brand, G.; Yifrach, O.; Arbely, E.; Roichman, Y.; Elia, N. Live cell single molecule tracking and localization microscopy of bioorthogonally labeled plasma membrane proteins. Nanoscale 2020, 12, 3236-3248.
- Sakuragi, S.; Kato, N.; Uchida; T.; Katagiri, T.; Enomoto, M.; Kato, R.; Yoshimura, H.; Oyama, C.; Katayama, I.; Chikuma, A.; Teramura, Y.; Bannai, H. Oligo DNA-based quantum dot (QD) single-particle tracking for multicolor single-molecule imaging. bioRxv 2025, preprint . [CrossRef]
- Ilievski, F.; Wikström, L.; Borg, A.; Volkov, I. L.; Brandis, G.; Johansson, M. Optimization of the genetic code expansion technology for intracellular labelling and single-molecule tracking of proteins in genomically re-coded E. coli. RSC Chemical Biology 2026, Advance Article online DOI: 10.1039/d5cb00221d.
- Bjerneld, E. J.; Földes-Papp; Z.; Käll, M.; Rigler, R. Single-molecule surface-enhanced Raman and fluorescence correlation spectroscopy of horseradish peroxidase. Journal of Physical Chemistry B 2002, 106, 1213-1218.
- Földes-Papp, Z.; Liao, S.-C.; Barbieri, B.; Gryczynski, K.; Luchowski, R.; Gryczynski, Z.; Gryczynski, I.; Borejdo, J.; You, T. Single actomyosin motor interactions in skeletal muscle. Biochimica et Biophysica Acta 2011, 1813, 856-866.
- Steves, M. A.; Xu, K. Direct microsecond wide-field single-molecule tracking and super-resolution mappimg via CCD vertical shift. Nature Communications 2025, 16, 10503 (1-19).
- Cabillic, M.; Forriere, H.; Bettarel, L.; Butler, C.; Neuhaus, A.; Idrissi, I.; Sambrano-Lopez, M. E.; Rossbroich, J.; Müller, L.-R.; Ries, J.; Grenci, G.; Viasnoff, V.; Levet, F.; Sibarita, J.-B.; Galland, R. In-depth single molecule localization microscopy using adaptive optics and single objective light-sheet microscopy. Nature Communications 2025, 16, 8362 (1-15).
- Ruprecht, V.; Brameshuber, M.; Schütz, G. J. Two-color single molecule tracking combined with photobleaching for the detection of rare molecular interactions in fluid biomembranes. Soft Matter 2010, 6, 568-581.
- Hänselmann. S.; Herten, D. P. Two-Color Single-Molecule Tracking in Live Cells. Methods Molecular Biology 2017, 1663, 127-138.
- Saavedra, L. A.; Barrantes, F. J. Temporal convolutional networks work as general feature extractions for single-particle diffusion analysis. Journal of Physics: Photonics 2025, 7, 025017 (1-15).
- Umney, O.; Slaney, H.; Williams, C. J. M.; Quirke P.; Peckham, M.; Curd, A. P. ClusterNet: Classifying Single-Molecule localization Microscopy Datasets with Graph-Based Deep Learning of Supracluster Structure. Small Science 2025, e202500255 (1-11).
- Pineda, J.; Masó-Orriols, S.; Masoliver, M.; Bertran, J.; Golsör, M.; Volpe, G.; Manzo C. Enhanced spatial clustering of single-molecule localizations with graph neural networks. Nature Communications 2025, 16, 9693 (1-17).
- Saguy, A.; Xiao, D.; Narayanasamy, K. K., Nakatani, Y.; Saliba, N.; Gagliano, G.; Gustavsson, A.-K.; Heilemann, M., Shechtman, Y. One-click reconstruction in single-molecule localization microscopy via experimental parameter-aware deep learning. npj Imaging 2025, 3, 61(1-10).
- Khater, I. M.; Nabi, I. R.; Hamarneh, G. HDCluster: High-Degree Graph Clustering for Robust Analysis of Single Molecule Localization Microscopy. bioRxiv [Preprint] 2025 (Oct 24), 2025.10.23.684134.
- Saxton, M. J.; Jacobson, K. Single-particle tracking: applications to membrane dynamics. Annual Review of Biophysics and Biomolecular Structure 1997, 26, 373-399.
- Jeon, J.-H.; Leijnse, N; Odderschede, L. B.; Metzler, R. Anomalous diffusion and power-law relaxation of the time averaged mean squared displacement in worm-like micellar solution. New Journal of Physics 2013, 15, 045011 (1-16).
- Ghosal, A.; Wang, Y.-H.; Nguyen, N.; Troyer, L.; Kim, S. Practical considerations for accurate estimation of diffusion parameters from single-particle tracking in living cells. The Journal of Physical Chemistry 2025, 163, 134202 (1-11).
- Baumann, G.; Gryczynski, I.; Földes-Papp, Z. Anomalous behavior in length distributions of 3D random Brownian walks (three dimensional random Brownian walks) and measured photon count rates within observation volumes of single-molecule trajectories in fluorescence fluctuation microscopy. Optics Express 2010, 18, 17883-17896.
- Lanza, A.; Martinez-Martin, I.; Tapia-Rojo, R.; Bo, S. Measuring irreversibility in stochastic systems by categorizing single-molecules displacements. Cornell University arXiv:2511.09183v1 [cond-mat.stat-mech] 2025, 1-12.
- Khrapak, S.; Khrapak, A. Stokes–Einstein Relation in Different Models of Water. Molecules 2024, 29, 5587. [CrossRef]
- Földes-Papp, Z.; Liao, S.-C. J.; You, T.; Barbieri, B. Reducing Background Contributions in Fluorescence Fluctuation Time-Traces for Single-Molecule Measurements in Solution. Current Pharmaceutical Biotechnology 2009, 10, 532-542.
- Sun, Y.; Tortarolo, G.; Teng, K.-W.; Ishitsuka, Y.; Coskun, U. C.; Liao, S.-C. J.; Diaspro, A.; Vicidomini, G.; Selvin, P.R.; Barbieri, B. A novel pulsed STED microscopy method using FastFLIM and the phasor plots. In Proceedings SPIE 10069, Proceedings volume: Multiphoton Microscopy in the Biomedical Sciences XVII, San Francisco, California, United States of America, February 21, 2017; 100691C. [CrossRef]
- McSwiggen, D. T.; Liu, H.; Tan, R.; Puig, S. A.; Akella, L. B.; Berman, R.; Bretan, M.; Chen, H.; Darzacq, X.; Ford, K.; Godbey, R.; Gonzalez, E.; Hanuka, A.; Heckert, A.; Ho, J. J.; Johnson, S. L.; Kelso, R.; Klammer, A.; Krishnamurthy, R.; Li, J.; Lin, K.; Margolin, B.; McNamara, P.; Meyer, L.; Pierce, S. E.; Sule, A.; Stashko, C.; Tang, Y., Anderson, D. J.; Beck, H. P. A high-throughput platform for single-molecule tracking identifies drug interaction and cellular mechanisms. eLife 2023, 12, RP93183 (1-26).
- Liu, J.; Zhang, X.; Zhao, B.; Ling, H.; Li, Y.; Sun, K.; Chen, S.; Zhang, Y.; Zhai, T.; Zhang, Y.; Li, F.; Qian Liu, Q. In Situ Monitoring of Membrane Protein Dynamics Using High-Throughput Red-Light-Activated Single-Molecule Tracking. ACS Nano 2025, 19, 13466–13478.
- Watanabe, D., Hiroshima, M., Yasui, M.; Ueda, M. Single molecule tracking based drug screening. Nature Communication 2024, 15, 8975 (1-14).
- Park, J.; Sokolovska, N.; Cabriel, C.; Kobayashi, A.; Corsin, E.; Garcia-Fernandez, F.; Izeddin, I.; Miné-Hattab, J. Novel estimation of memory in molecular dynamics with extended and comprehensive single-molecule tracking software: FreeTrace. bioRxiv 2026, preprint (1-23), . [CrossRef]
- Zalejski, J.; Sun, J.; Sharma, A. Unravelling the Mystery inside Cells by Using Single-Molecule Fluorescence Imaging. Journal of Imaging 2023, 9, 192 (1-25).
- Park, C.; Jung, S.; Park, H. Single vesicle tracking for studying synaptic vesicle dynamics in small central synapses. Current Opinion in Neurobiology 2022,76, 102596.
- Graham, T. G. W.; Dugast-Darzacq, C.; Dailey, G. M.; Weng, B.; Anantakrishnan, S.; Darzacq, X.; Tjian, R. Single-molecule live imaging of subunit interactions and exchange within cellular regulatory complexes. Molecular Cell 2025, 85, 2854-2868.
- Bühler, B.; Sunbul, M. Single-Molecule RNA Imaging in Live Cells with an Avidity-Based Fluorescent Light-Up Aptamer biRhoBAST. Methods in Molecular Biology 2024; 2822, 87-100.
- Hou, S.; Exell, J.; Welsher, K. Real-time 3D single molecule tracking. Nature Communication 2020, 11, 3607.
- Si, D.; Dong, Z.; Xing, G.; Yao, L.; Lu, J.; Ma, M.; Yuan,Y.; Zhao, H.; Dong, B.; Li, C.; Yuan, L.; Chen, Z.; Wang, L. A bright and regenerative array tag with fluorogenic ligands for long-term live-cell nanoscopy and single-molecule tracking. Nature Communications 2026, 17, 576 (1-14).
- Place, R. F; Noonan, E. J.; Földes-Papp, Z.; Li, L.-C. Defining Features and Exploring Chemical Modifications to Manipulate RNAa Activity. Current Pharmaceutical Biotechnology 2010, 11, 518-526.
- Butler, C.; Saraceno G.E.; Kechkar, A.; Bénac, N.; Studer, V.; Dupuis, J.P.; Groc, L.; Galland, R.; Sibarita J.-B. Multi-Dimensional Spectral Single Molecule Localization Microscopy. Frontiers in Bioinformatics 2022, 2, 813494 (1-14).
- Rabiee, N.; Lan, X. Advancing Multicolor Super-Resolution Volume Imaging: Illuminating Complex Cellular Dynamics. Journal of the American Chemical Society Au 2025, 5, 2388–2419.
- Gebhardt, C. M. Single-molecule Tracking and Kinetic Analysis in Living Cells and Multicellular Organisms. Journal of Molecular Biology 2026, 438, 169308.
- Kim, K.; Jang, J.; Cho, J.; Ahn, Y.; Jeong, S.; Shin, J.; Yea, K.; Lee, W. J.; Seo, D. Reconciling a Kinetic Model for Dimerization of the EGFR Using Single-Molecule Tracking in Living Cells. The Journal of Physical Chemistry B 2025,129, 9567-957.
- Mondal, S.; Bonventre, E.; Hancock; W.O.; Rice, L.M. Single-molecule microtubule dynamics measurements reveal an intermediate state and clarify the role of nucleotide. Nature Communications 2026, 17, 559 (1-12).
- Földes-Papp, Z. What it means to measure a single molecule in a solution by fluorescence fluctuation spectroscopy. Experimental and Molecular Pathology 2006, 80, 209-218.
- Földes-Papp, Z; Rigler, R. Quantitative Two-Color Fluorescence Cross-Correlation Spectroscopy in the Analysis of Polymerase Chain Reaction. Biological Chemistry 2001, 382, 473-478.
- Földes-Papp, Z; Demel, U.; Tilz, G. P. Ultrasensitive detection and identification of fluorescent molecules by FCS: Impact for immunobiology. Proceedings of the National Academy of Sciences of the United States of America 2001, 98, 11509-11514.
- Földes-Papp, Z. ’True’ single-molecule molecule observations by fluorescence correlation spectroscopy and two-color fluorescence cross-correlation spectroscopy. Experimental and Molecular Pathology 2007, 82, 147-155.
- Földes-Papp, Z. How the molecule number is correctly quantified in two-color fluorescence cross-correlation spectroscopy: corrections for cross-talk and quenching in experiments. Current Pharmaceutical Biotechnology 2005, 6, 437-444.
- Muñoz-Gil, G.; Bachimanchi, H.; Pineda, J.; Midtvedt, B.; Fernández-Fernández, G.; Requena, B.; Ahsini, Y.; Asghar, S.; Bae, J.; Barrantes, F. J.; Bender, S. W. B.; Cabriel, C.; Conejero, J. A.; Escoto, M.; Feng, X.; Haidari, R.; Hatzakis, N. S.; Huang, Z.; Izeddin, I.; Jeong, H.; Jiang, Y.; Kæstel-Hansen, J.; Miné-Hattab, J.; Ni, R.; Park, J.; Qu, X.; Saavedra, L. A.; Sha, H.; Sokolovska, N.; Zhang, Y.; Volpe, G.; Lewenstein, M.; Metzler, R.; Krapf, D.; Volpe, G.; Manzo, C. Quantitative evaluation of methods to analyze motion changes in single-particle experiments. Nature Communication 2025, 16, 6749 (1-17).
- Niu, R.; Shao, J.; Wu, M.; Liu, C.; Chao, J. Single-Molecule Detection of Optical Signals Using DNA-Based Plasmonic Nanostructures. Biosensors 2025, 15, 398 (1-18). [CrossRef]
- Huang, X.; Shi, Q.; Lu, Y.; Li, B.; Ning, Y.; Feng, L.; Sojic, N.; Jiang, D.; Liu, B. Single-Molecule Electrochemiluminescence Imaging of Plasmonic Hot Spot Reactivity. Angewandte Chemie International Edition 2025, 64: e202508266.
- Wenger, J. UV plasmonics to enhance protein autofluorescence detection at the single molecule level. In Proceedings SPIE PC1358, Proceedings volume: UV and Higer Energy Photonics: From Materials to Applications 2025, San Diego, California, United States of America, September 19, 2025; PC135830A. [CrossRef]
- Nooteboom, S.W.; Lamberti, V.; Schellekens, T.; Zijlstra, P. Multiplexed and continuous single-molecule biosensing by spectral demixing of plasmon-enhanced fluorescence. Biosensors and Bioelectronics 2026, 298, 118384 (1-11).
- Ito, Y.; Sakata-Sogawa, K.; Tokunaga, M. Multi-color single-molecule tracking and subtrajectory analysis for quantification of spatiotemporal dynamics and kinetics upon T cell activation. Scientific Reports 2017, 7, 6994 (1-14).
- Fujita, H.; Zhong, C.; Arai, S.; Suzuki, M. Bright Dots and Smart Optical Microscopy to Probe Intracellular Events in Single Cells. Frontiers in Bioengineering and Biotechnology 2019, 6, 204 (1-7).
- Li, G.; Gong, C. High-Contrast Interferometric Imaging of Single-Molecule Dynamics on Optical Fibers. Cornell University arXiv:2510.10061v1 [physics.optics] 2025, 1-19.
- Dahal, L.; Walther, N.; Tjian, R.; Darzacq, X.; Graham, T. G. W. Single-molecule tracking (SMT): a window into live-cell transcription biochemistry. Biochemical Society Transactions 2023, 51, 557–569.
- Kamarulzaman, L.; Kim, S.; Hidaka, T.; Tsuchida, M.; Taniguchi, Y. Comprehensive molecular profiling of single-cell proteome via gel electrophoresis and 3D single-molecule imaging. bioRxiv 2026, preprint 1-30, . [CrossRef]
Figure 1.
The particle tracking used the Scanning Mirrors to circulate the laser beam around the particle, with a value for the z-position up and down to find the new locations to track (see legend of Figure 2). UV: Ultraviolet, VIS: Visible. NIR: Near-Infrared, M1: Mirror 1, D1: Dichroic 1, D2: Dichroic 2, H: Halfwave Plate, P: Polarizer, SC: Scanning Lens, M2: Mirror 2, MDW: Multi-Dichroic Wheel, FL: Focal Lens, VP: Variable Pinhole, S: Shutter, PMT: Photomultiplier-Tube. This figure is from the ISS laboratory and illustrates our known conclusions or theorems.
Figure 1.
The particle tracking used the Scanning Mirrors to circulate the laser beam around the particle, with a value for the z-position up and down to find the new locations to track (see legend of Figure 2). UV: Ultraviolet, VIS: Visible. NIR: Near-Infrared, M1: Mirror 1, D1: Dichroic 1, D2: Dichroic 2, H: Halfwave Plate, P: Polarizer, SC: Scanning Lens, M2: Mirror 2, MDW: Multi-Dichroic Wheel, FL: Focal Lens, VP: Variable Pinhole, S: Shutter, PMT: Photomultiplier-Tube. This figure is from the ISS laboratory and illustrates our known conclusions or theorems.
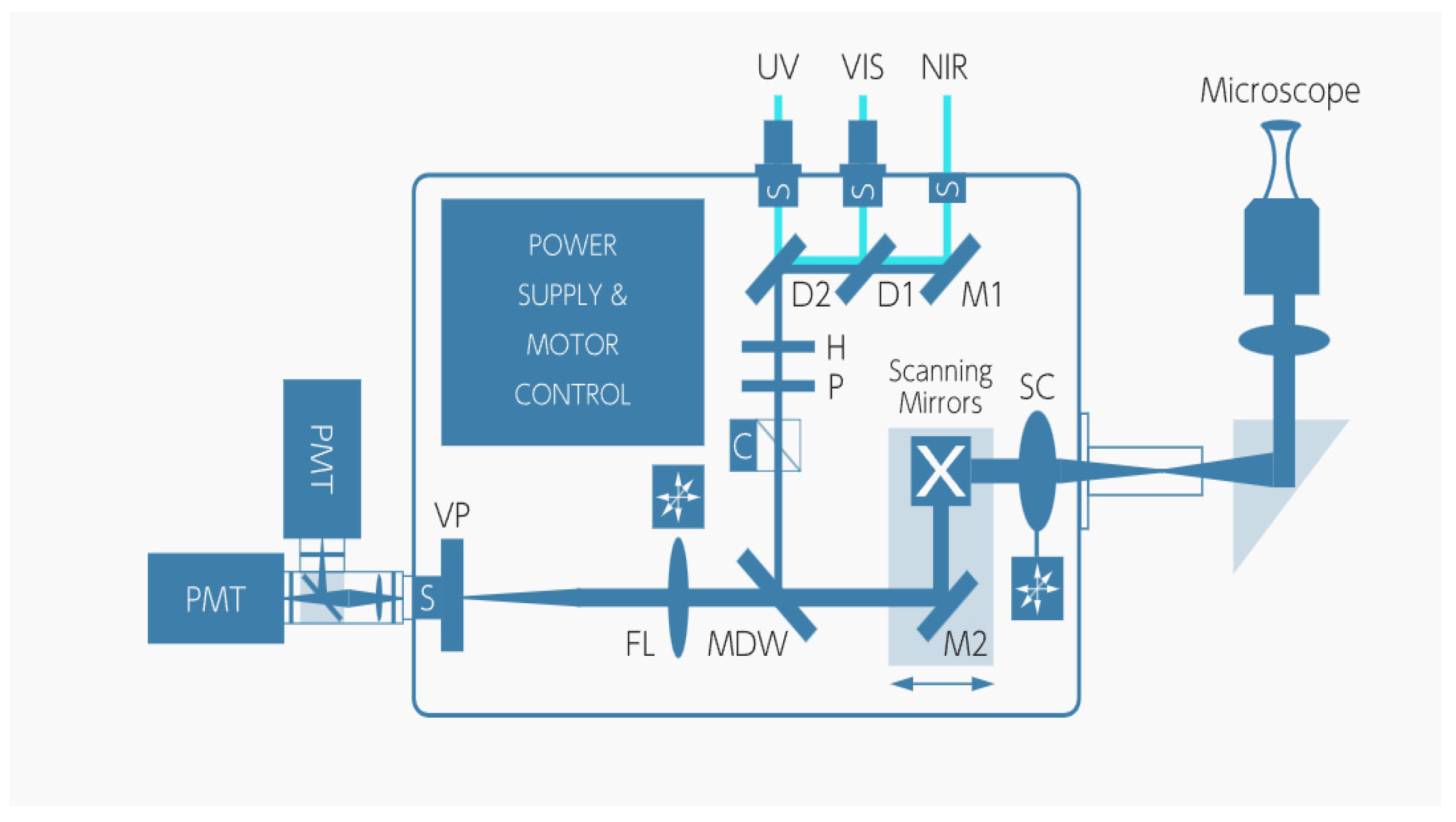
Figure 2.
The concentration of the 0.5 μm beads was 0.1 nM in distilled water at room temperature. The beads were purchased from Duke Scientific Corp., cat. no. G500, diameter 0.499 µm, 15 ml polymer microspheres, green fluorescing 1% solids. IOTech3001 PCI board was used to perform the 3D particle tracking. The 488 nm laser was applied for excitation and green fluorescence of single beads was tracked at 535 nm. The scanning mirror circled the bead and Fourier transformed to indicate a direction of the bead’s next move and followed. The z-direction was tracked up and down of this circled xy-direction pattern and determined which direction the bead’s next z-direction moved. The units in the x-pos, y-pos and z-pos were in micrometer. The full tracking time was 500 seconds. This figure is from the ISS laboratory and illustrates our known conclusions or theorems.
Figure 2.
The concentration of the 0.5 μm beads was 0.1 nM in distilled water at room temperature. The beads were purchased from Duke Scientific Corp., cat. no. G500, diameter 0.499 µm, 15 ml polymer microspheres, green fluorescing 1% solids. IOTech3001 PCI board was used to perform the 3D particle tracking. The 488 nm laser was applied for excitation and green fluorescence of single beads was tracked at 535 nm. The scanning mirror circled the bead and Fourier transformed to indicate a direction of the bead’s next move and followed. The z-direction was tracked up and down of this circled xy-direction pattern and determined which direction the bead’s next z-direction moved. The units in the x-pos, y-pos and z-pos were in micrometer. The full tracking time was 500 seconds. This figure is from the ISS laboratory and illustrates our known conclusions or theorems.
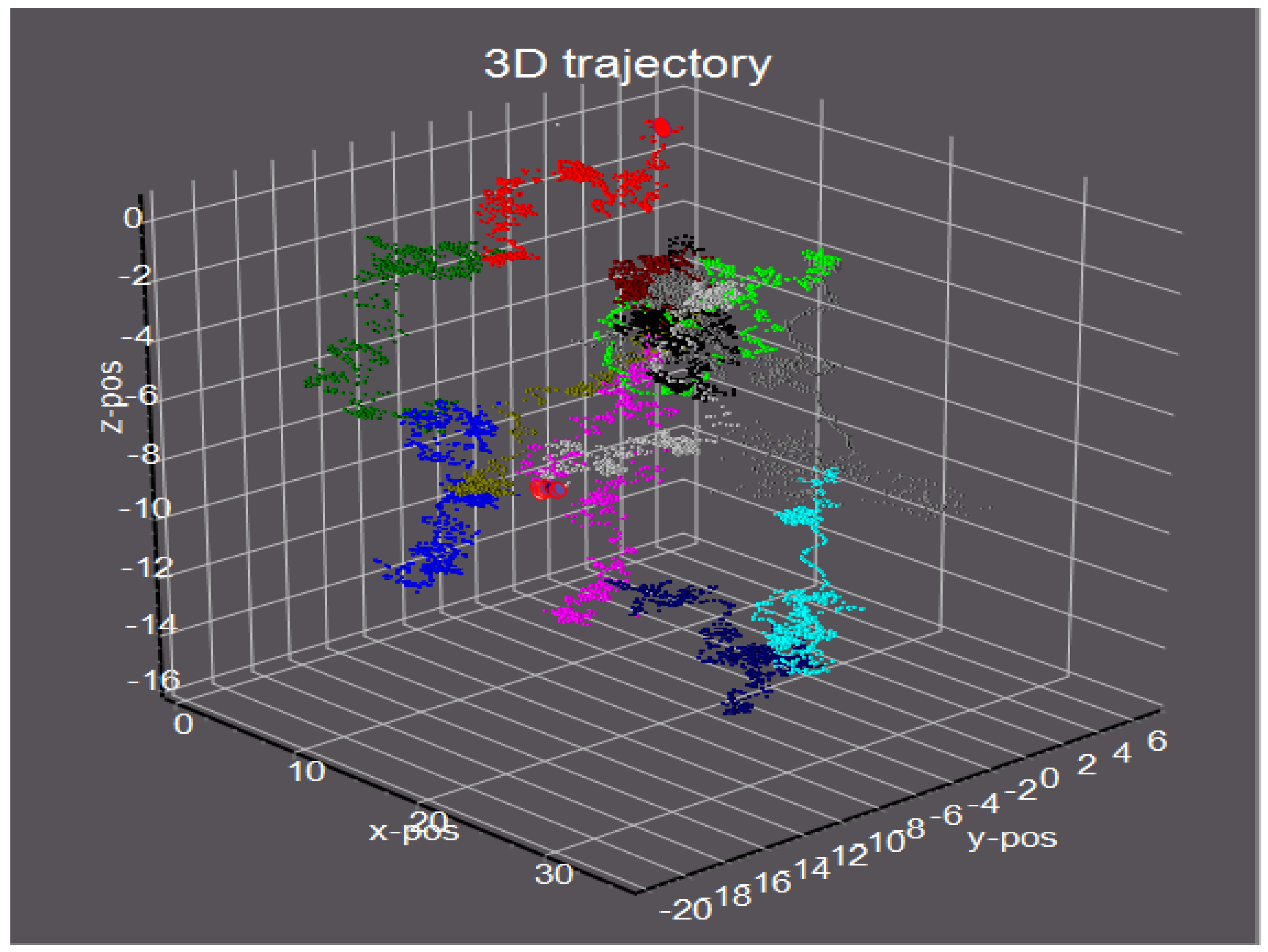
Figure 3.
The graph shows the single total track. A blue dot marks the track’s starting and ending points. This figure is from the ISS laboratory and illustrates our known conclusions or theorems.
Figure 3.
The graph shows the single total track. A blue dot marks the track’s starting and ending points. This figure is from the ISS laboratory and illustrates our known conclusions or theorems.
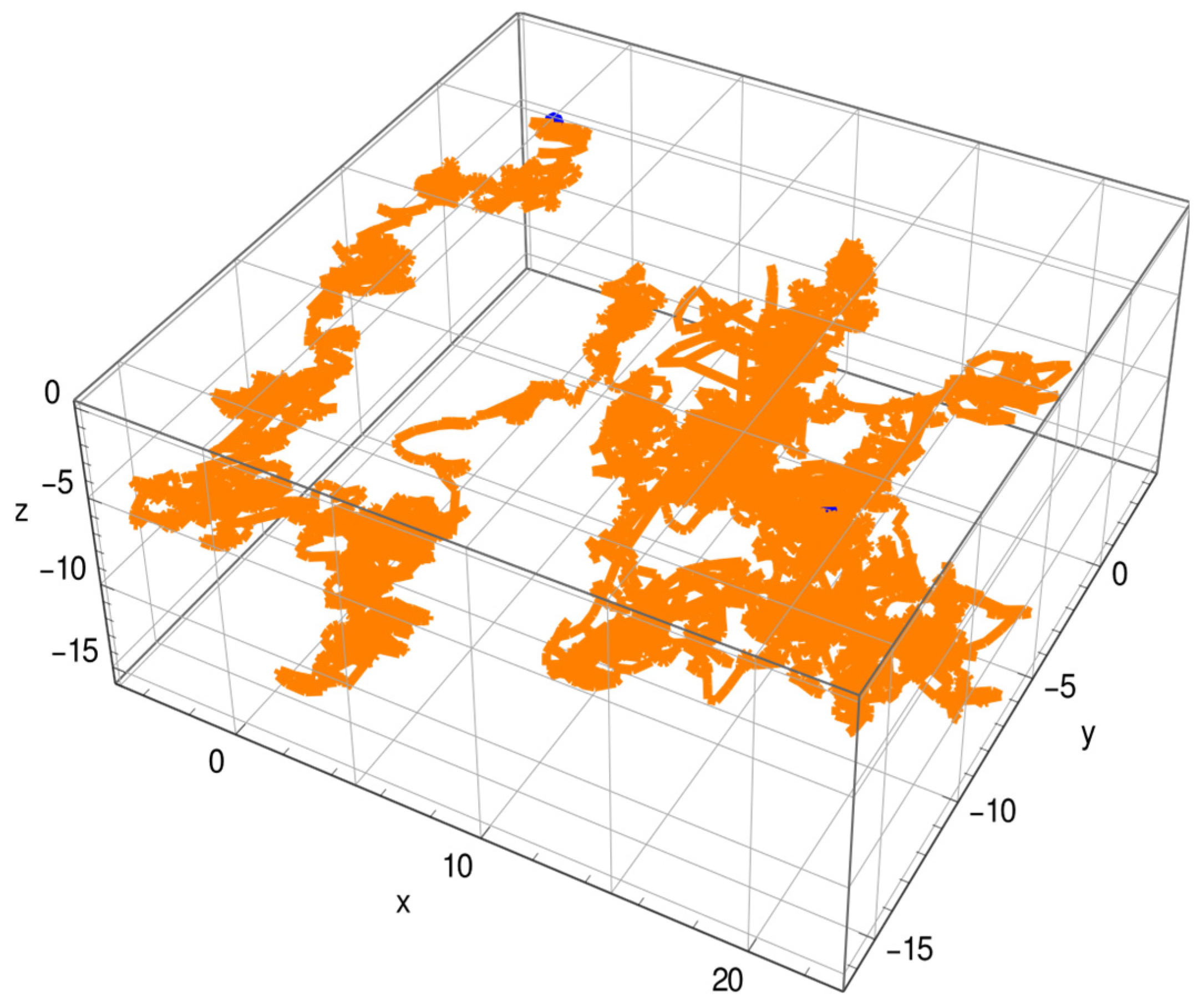
Figure 4.
The dataset depicts two areas with linear increases. The first interval stretches from n ∈ (0, 400), whereas the second interval runs from n ∈ (500, 900). The slopes for these intervals are α = 0.071 and α = 0.085, respectively. Fitting the whole interval n ∈ (0, 900) with produces α = 0.048, β = 1.06 with a time unit of 40 ms, indicating that there was another physical process in addition to diffusion. Since the first interval controled Brownian motion, it is assumed that stands for the diffusion coefficient with a time unit of 40 ms. This figure is from the ISS laboratory and illustrates our known conclusions or theorems.
Figure 4.
The dataset depicts two areas with linear increases. The first interval stretches from n ∈ (0, 400), whereas the second interval runs from n ∈ (500, 900). The slopes for these intervals are α = 0.071 and α = 0.085, respectively. Fitting the whole interval n ∈ (0, 900) with produces α = 0.048, β = 1.06 with a time unit of 40 ms, indicating that there was another physical process in addition to diffusion. Since the first interval controled Brownian motion, it is assumed that stands for the diffusion coefficient with a time unit of 40 ms. This figure is from the ISS laboratory and illustrates our known conclusions or theorems.
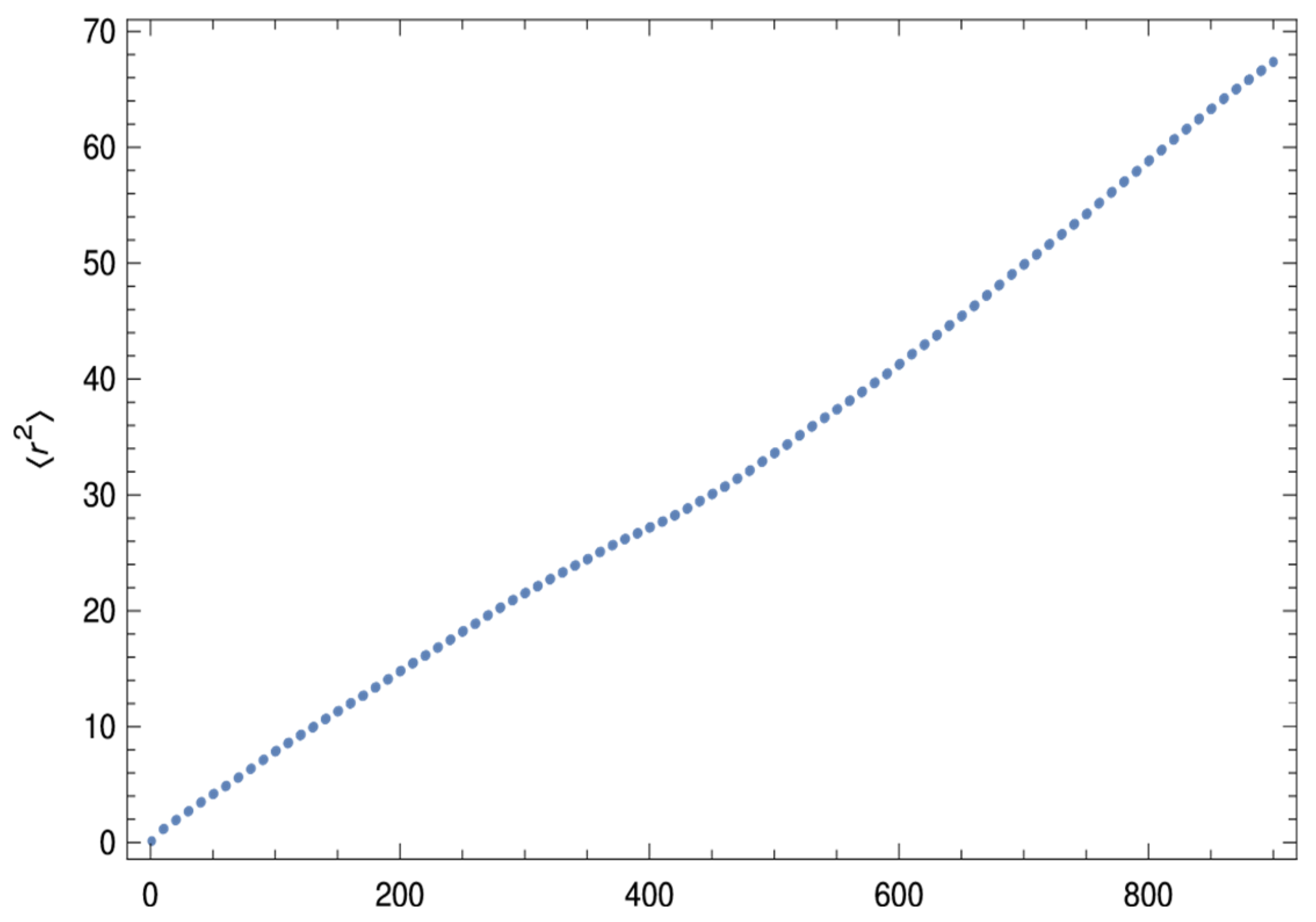
Figure 5.
Fixed HeLa cell, mitochondria labeled with Atto 647N and imaged by STED. The figure is shown with the permission of ISS com. STED z-stack, 34 x 34 x 1.2 µm3,1024 x 1024 x 10 voxels. This figure is from the ISS laboratory and illustrates our known conclusions or theorems. See also our website of ISS Inc. and https://www.youtube.com/watch?v=MkdPAGxfI14 .
Figure 5.
Fixed HeLa cell, mitochondria labeled with Atto 647N and imaged by STED. The figure is shown with the permission of ISS com. STED z-stack, 34 x 34 x 1.2 µm3,1024 x 1024 x 10 voxels. This figure is from the ISS laboratory and illustrates our known conclusions or theorems. See also our website of ISS Inc. and https://www.youtube.com/watch?v=MkdPAGxfI14 .
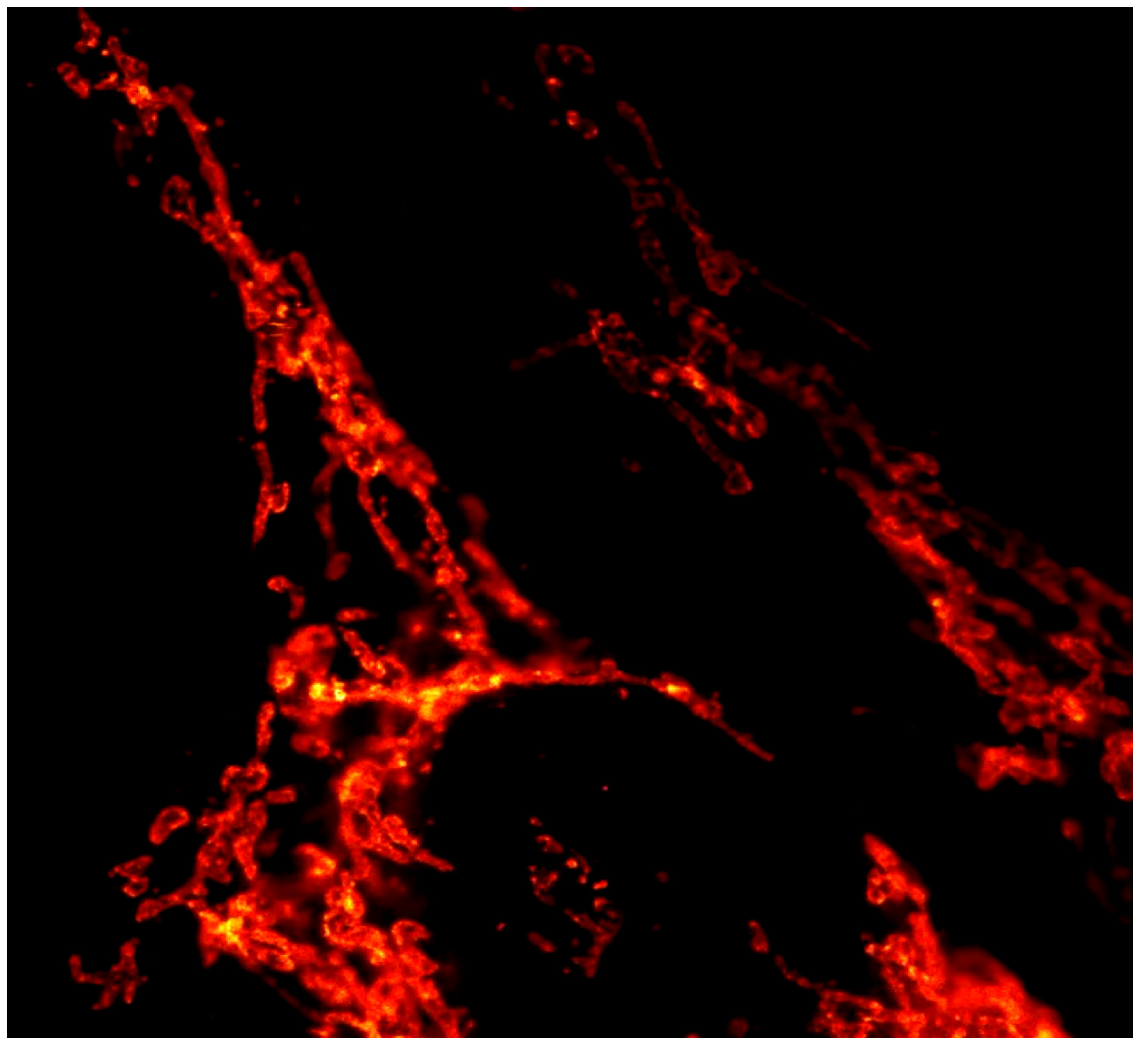
Figure 6.
Live Hela cell microtubule labeled with Atto 647N and Atto 655. Both makers (labels) were excited with a 640 nm laser, separated by pSTED and FastFLIM and then displayed with two different ‘false’ colors. Atto 647N is depicted in yellow and Atto 655 is depicted in violet to generate the processed and fused pSTED image of the two markers. The figure is shown with the permission of ISS com (see also our website of ISS com) and illustrates our known conclusions or theorems.
Figure 6.
Live Hela cell microtubule labeled with Atto 647N and Atto 655. Both makers (labels) were excited with a 640 nm laser, separated by pSTED and FastFLIM and then displayed with two different ‘false’ colors. Atto 647N is depicted in yellow and Atto 655 is depicted in violet to generate the processed and fused pSTED image of the two markers. The figure is shown with the permission of ISS com (see also our website of ISS com) and illustrates our known conclusions or theorems.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



