Submitted:
13 February 2026
Posted:
27 February 2026
You are already at the latest version
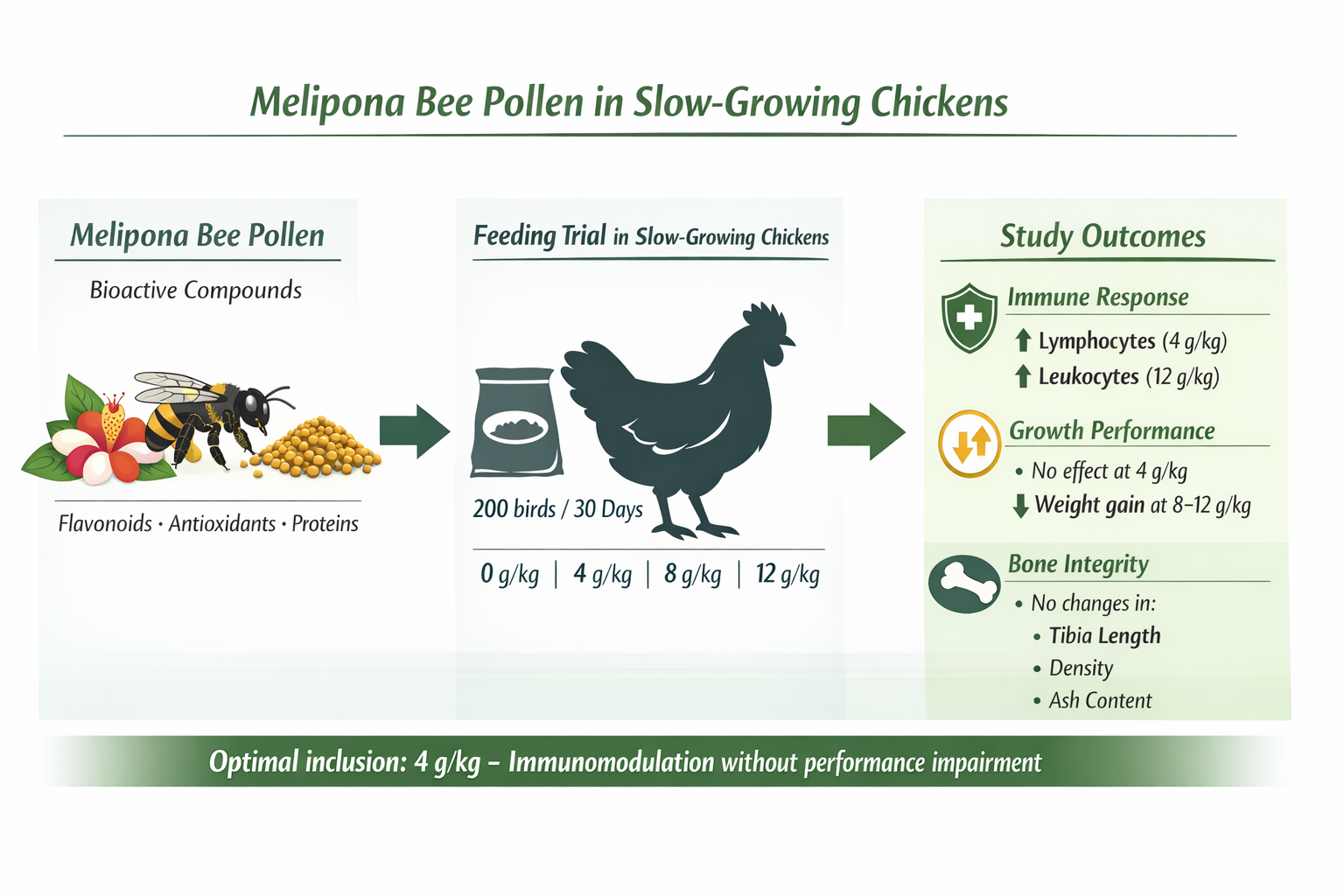
Abstract
This study evaluated the effects of stingless bee (Melipona) pollen supplementation on productive performance, carcass traits, hematological parameters, lymphoid organs, and bone histomorphometry in slow-growing chickens. A total of 200 one-day-old mixed-sex chickens were distributed in a completely randomized design with four dietary treatments: a control diet and diets supplemented with 4, 8, and 12 g of bee pollen per kg of feed, with five replicates of ten birds each, from 1 to 30 days of age. Productive performance was assessed, and carcass yield, edible viscera, lymphoid organs, blood parameters, and tibia morphometry were evaluated at 30 days. Data were analyzed using ANOVA followed by Tukey’s test (p < 0.05). Feed intake, carcass yield, noble cuts, and edible organs were not affected by pollen inclusion. However, weight gain and feed conversion ratio were negatively affected at inclusion levels above 4 g/kg. Hematological analysis revealed significant changes in total leukocytes, lymphocytes, eosinophils, and monocytes, suggesting immunomodulatory effects of pollen supplementation. No significant differences were observed in bone histomorphometric traits. In conclusion, dietary inclusion of 4 g/kg of Melipona pollen can be used as a functional feed additive in slow-growing chickens, providing immunological modulation without impairing productive performance or bone integrity.
Keywords:
stingless bee pollen
; functional feed additive
; hematology
; poultry nutrition
; slow-growing chickens
1. Introduction
Poultry of zootechnical interest raised in tropical environments are subject to adverse conditions, such as heat stress and exposure to pathogenic microorganisms, which cause low productive performance and increased disease prevalence. To mitigate these challenges, synthetic vitamins and antibiotics are traditionally used in order to minimize the effects of stress and control the growth of the pathogenic microbiota. Recent trials using natural products aimed at improving poultry performance and health have driven the investigation of bee-derived products—particularly pollen—as alternative and safe enhancers to replace antibiotic growth promoters in poultry diets [1]. Bee pollen consists of pollen grains mixed with nectar and secretions from the bees’ hypopharyngeal glands [2] and is considered a food supplement in animal nutrition due to its abundant nutrients [3], such as flavonoids [4], antioxidants [5], digestive enzymes [6] and antimicrobial compounds [7]; it is also rich in protein, essential amino acids, oils, vitamins, minerals, and carbohydrates [2,6]. However, most research has focused on pollen from Apis mellifera, resulting in limited information on the specific effects of pollen from Melipona bees, which are known to produce pollen with distinctive nutritional and chemical profiles shaped by their interaction with local and endemic flora [8]. Based on this background, the inclusion of Melipona bee pollen in the diet of slow-growing chickens may enhance weight gain, improve feed conversion ratio, stimulate the development of immune organs such as the bursa of Fabricius and thymus—thereby strengthening resistance to pathogens—and improve bone strength. Therefore, it was hypothesized that low dietary inclusion levels of stingless bee pollen could modulate immune-related parameters without negatively affecting productive performance in slow-growing chickens.
2. Materials and Methods
The trial was conducted on the campus of the National Autonomous University of Alto Amazonas, located in the city of Yurimaguas, Loreto region, Peru (5°53’51”S and 76°07’43”W, altitude of 184 meters above sea level), between May and September 2025. The animal research procedures were carried out in accordance with the guidelines of the Scientific Integrity Committee for the use of animals of the National Autonomous University of Alto Amazonas, Report No. 001-2024-UNAAA/CIC.
2.1. Facilities, Poultry and Treatments
The experiment was conducted in an 84-m² poultry shed constructed with concrete walls and a zinc-coated steel roof, enclosed with wire mesh, which provided adequate ventilation. The birds were housed in two metabolic battery units, each containing ten cages of 1 m² arranged in two columns of five levels. The cages were made of electro-galvanized mesh and equipped with feeder and drinker hoppers, which were cleaned and refilled twice daily to ensure free access to feed and water throughout the trial. Temperature was provided using incandescent bulbs during the first week to meet the thermal requirements of the birds, maintaining an average temperature of 29.6 ± 2.5 °C. Thereafter, birds were kept under ambient conditions, with an average temperature of 27.9 ± 2.1 °C and a mean relative humidity of 79 ±7%.
A total of two hundred one-day-old slow-growing chickens, mixed sex, were assigned to four treatments in a completely randomized design, with five replicates and ten birds per experimental unit. The experimental diets were supplemented with 4, 8, and 12 g of Melipona bee pollen per kg of feed, without the inclusion of synthetic antibiotics as growth promoters, along with a control diet containing an antibiotic. The pollen was collected with consent from regional meliponiculturists, then stored in a Thermo Scientific® Forma 900 series vertical ultra-freezer for 24 hours at –50 °C, freeze-dried in a Labconco® FreeZone model freeze-dryer at –85 °C for 24 hours under a pressure of 0.03 mbar, and finally ground in a porcelain mortar until a uniform powder was obtained. The experimental diets were offered in mash form, and their formulation was based on the nutritional requirements proposed by Pinheiro et al. (2014) [9] for free-range birds raised in semi-confined systems (Table 1). Proximate analyses were performed on both diets and the pollen (Table 2).
2.2. Productive Performance
Body weight and feed intake were recorded at the beginning and at 30 days to determine weight gain, feed intake, and feed conversion ratio. Mortality was recorded to adjust feed intake according to Sakomura & Rostagno (2016) [10].
2.3. Yield of Carcasses, Noble Cuts, Edible Viscera and Lymphoid Organs
At 30 days of age, two birds per experimental unit with representative body weight (±5%) were selected, fasted for 12 hours, weighed, and slaughtered by cervical dislocation, followed by bleeding, scalding, defeathering, and evisceration. The relative weights (%) of the carcasses (without feet, neck, and head) and the noble cuts (breast, thigh, drumstick, and wing) were evaluated. The relative weight of the defeathered and eviscerated carcass was calculated in relation to the fasting body weight.
Edible viscera (gizzard, heart, and liver) and lymphoid organs (thymus, spleen, and bursa) were collected during evisceration and individually weighed using a precision balance; All the adhered fat, its contents and the coiling membrane were removed from the gizzard. The relative weights of the cuts, edible viscera and lymphoid organs in relation to the eviscerated carcass were obtained.
2.4. Blood Analysis
From the birds selected for carcass evaluation, 1.5 mL of blood was collected beforehand via puncture of the brachial vein using 3-mL syringes with 21 × 1” needles, and the samples were placed in EDTA-containing tubes to preserve cellular integrity. Hematocrit was determined using the microhematocrit method, centrifuging heparinized glass capillary tubes at 12,000 rpm for 90 seconds. The proportion of erythrocytes relative to the total blood volume was estimated using percentage values printed according to the relative size of the blood fractions [11,12]. Hemoglobin concentration was indirectly estimated by multiplying the hematocrit value expressed as a percentage by 0.33 [13].
Total leukocyte count was performed using a Neubauer Improved Bright-Line hemocytometer (Kyntel®). A diluent was prepared consisting of 3 mL of saline solution (0.9% NaCl), 6 drops of methylene blue, and 6 drops of basic fuchsin, used as a supravital stain to highlight leukocyte nuclei and facilitate their identification in the hemocytometer [11,12].
From this solution, 1995 µL were extracted and mixed with 5 µL of whole blood using an Eppendorf micropipette, yielding a final dilution of 1:400. The suspension was carefully homogenized and loaded into the Neubauer chamber, avoiding bubble formation. Leukocytes were counted in the four large corner squares, corresponding to a total volume of 0.4 µL (each square: 1 mm² × 0.1 mm = 0.1 µL). The total leukocyte number (N) was converted to concentration using the formula:
where N is the number of leukocytes counted in the four large squares, 400 is the dilution factor (1:400), and 0.4 µL is the total volume counted in the chamber.
Leukocytes/µL = N × 400 / 0.4 = N × 1000
Manual counting using a Neubauer chamber is considered a standard method in avian hematology. For the differential leukocyte count, a drop of fresh blood was placed at one end of the microscope slide, and a smear was prepared using a second slide held at a 30–45° angle to produce a thin and uniform film. Each smear was stained with approximately eight drops of Wright’s stain, supplemented with drops of distilled water to enhance fixation and differential staining of the cellular components. The blood smears were then air-dried and examined using an optical microscope equipped with an integrated camera.
In each blood smear, a total of 100 leukocytes per sample were counted and classified into five categories—heterophils, lymphocytes, monocytes, eosinophils, and basophils—according to the morphological criteria described in avian hematology [11,12,14]. The results were expressed as the relative percentage (%) of each leukocyte type, allowing the identification of alterations consistent with inflammatory, infectious, or immunological processes.
2.5. Bone Histomorphometry
The methodology of Aguilar et al. (2018) [15] was followed. The right tibia was cleaned of muscles and adherent tissues, labeled, and stored frozen until morphometric analysis in the laboratory. Prior to measurement, tibiae were briefly boiled in water to facilitate the removal of any remaining residues and then dried. Bone length and midshaft diameter (measured as the medio-lateral diameter at the midpoint of the diaphysis) were recorded using a digital vernier caliper with an accuracy of 0.1 mm, and bones were subsequently weighed on a precision balance (after drying in an oven at 105 °C for 24 h to obtain dry weight). For ash analysis, dried bones were ground, and a known aliquot was incinerated in a muffle furnace at 600 °C for 3 h to determine the mineral content percentage.
2.6. Statistical Procedures
For the statistical analysis of performance, each cage was considered an experimental unit, using the average of the animal data per cage. For the statistical analysis of relative carcass weight, primal cuts, edible and lymphoid organs, hematological variables, and bone histomorphometry, the experimental unit was the mean of the measurements obtained from the two birds slaughtered per cage. Data were subjected to normality testing (Shapiro–Wilk) using the UNIVARIATE procedure, and homogeneity of variances was assessed using Bartlett’s test. All variables were analyzed using the General Linear Model (GLM) procedure of SAS® (Statistical Analysis System v. 9.4, 2016). For each variable, an analysis of variance was performed according to the following general model:
where Yᵢⱼ is the measured dependent variable; µ is the overall mean; αᵢ is the effect of treatments; and εᵢⱼ is the random error.
Yij = μ + αi + εij
Mean comparisons were performed using Tukey’s test at a 5% significance level.
3. Results
3.1. Growth Performance
The effects of the addition of bee pollen on the growth performance of slow-growing chickens are shown in Table 3. No significant differences (P>0.05) were detected in feed intake among treatments, with values ranging from 1283.0 g (T2) to 1344.4 g (T1); however, body weight gain was significantly higher (P<0.05) in the control group compared with the groups receiving 8 (674.8 g) and 12 (665.2 g) g of pollen per kilogram of feed. Likewise, feed conversion ratio increased as the dietary pollen level increased, with the control treatment (1.779 g/g) being more efficient and statistically superior to the 12 g dose (1.975 g/g).
The means followed by different letters in the row differ from each other according to Tukey’s test with a probability of 5%.
The effects of the addition of bee pollen on carcass yield and its main components of slow-growing chickens are shown in Table 4. The results of the percentage analysis of carcass yield, prime cuts, and edible viscera showed no significant differences among treatments (P>0.05). The lymphoid organs—thymus, bursa of Fabricius, and spleen—exhibited the same pattern.
3.2. Blood Parameters
Blood parameters of slow-growing chickens fed with stingless bee pollen are shown in Table 5. No significant differences (P>0.05) among treatments for hemoglobin and hematocrit; however, total leukocyte count increased significantly (P<0.05) in birds receiving 12 g (71,100 cells/mm³) of pollen compared with the other treatments. For lymphocytes, the 4-g dose (57.40%) exhibited a higher percentage, similar to the control (52.50%) and the 12-g treatment, but different (P<0.05) from the 8-g (42.30%) pollen inclusion. Eosinophils showed a higher response in the 8-g (11.30%) group, differing significantly from the 12-g (4.40%) treatment. Monocytes also differed significantly, with the 12-g (8.60%) dose presenting the highest percentage and differing from the control (2.20%) treatment. Heterophils and basophils were not affected by increasing levels of stingless bee pollen.
3.3. Bone Histomorphometry
The bone histomorphometry of slow-growing chickens fed with stingless bee pollen is shown in Table 5. No significant effects on tibia length, diameter, weight, volume, density, or ash content, indicating that pollen supplementation did not noticeably affect the macroscopic bone morphometry.

Table 6.
Bone histomorphometry of the tibia of slow-growing chickens fed bee pollen1.
| Variables | TC | T1 | T2 | T3 | SEM | P-value |
|---|---|---|---|---|---|---|
| Length (mm) | 77.54 | 76.15 | 77.33 | 76.06 | 0.988 | 0.617 |
| Diameter (mm) | 6.15 | 6.07 | 6.08 | 5.78 | 0.151 | 0.356 |
| Weight (g) | 2.36 | 2.40 | 2.34 | 2.22 | 0.129 | 0.767 |
| Volume (cm3) | 2.40 | 2.50 | 2.50 | 2.40 | 0.149 | 0.928 |
| Density (g/cm3) | 1.00 | 0.97 | 0.94 | 0.93 | 0.047 | 0.689 |
| Ash (%) | 45.68 | 45.86 | 42.14 | 46.35 | 1.648 | 0.287 |
1 Analyses carried out at the Biotechnology Laboratory of the National Autonomous University of Alto Amazonas – Yurimaguas, Peru.
4. Discussion
The results of the present study demonstrate that the inclusion of stingless bees (Melipona spp.) pollen in diets for slow-growing chickens induces differential responses in productive performance, particularly in body weight gain and feed conversion ratio. These findings are consistent with previous reports highlighting the potential of bee pollen as a functional feed additive in poultry production [4,16,17], mainly due to its antioxidant and immunomodulatory properties, which may positively contribute to overall animal health [18].
Feed intakes indicate that the inclusion of Melipona bee pollen, irrespective of inclusion level, did not negatively affect diet palatability nor alter feeding behavior. The consistency in feed intake is particularly relevant considering that bee pollen exhibits a nutritional profile markedly different from that of conventional feed ingredients, with a crude protein content of 45.55% and gross energy of 5259 kcal/kg (Table 2), values substantially higher than those of the primary dietary components [2]. The stability of feed consumption may be attributed to the pollen exine, the outer cell wall characterized by its resistance and complex polymeric structure. Studies on bee pollen composition indicate that this structure contains bioactive compounds capable of modulating physiological responses without affecting intake patterns [19]. Khalifa et al. (2021) [4] reported that bee pollen contains phenolic compounds, flavonoids, and other bioactive metabolites with therapeutic potential, suggesting that its dietary effects occur primarily at the metabolic level rather than through mechanisms related to organoleptic acceptability.
Higher levels of bee pollen inclusion were associated with lower weight gain, which contrasts with some previous findings, but is consistent with more recent studies analyzing nutrient bioavailability in the presence of pollen exine. The pollen exine is mainly composed of sporopollenin, a highly stable polymer that may limit enzymatic access to nutrients contained within the pollen grain. Although bee pollen is characterized by a high crude protein content (45.55%; Table 2), this protein is not entirely bioavailable due to the physical barrier imposed by the exine. This observation agrees with the findings of Hashmi et al. (2012) [20], who reported that the effective utilization of bee pollen in broiler diets requires careful consideration of its structural characteristics. The progressive decline in body weight gain observed with increasing pollen inclusion levels (from 741.2 g in the control treatment to 665.2 g in T3) may be attributed to a reduction in the effective availability of digestible nutrients. Although the chemical composition of the experimental diets remained relatively homogeneous (Table 2), the gradual replacement of conventional ingredients with Melipona pollen altered the fraction of nutrients effectively available for metabolic utilization. Li et al. (2018) [16] emphasized that, despite the high nutritional density of bee pollen, its practical value in animal feeding depends largely on the integrity of the exine and the digestive capacity of the host to degrade this structure. Previous studies have shown that pollen digestibility varies considerably according to pollen origin and processing methods. In the present study, pollen was incorporated in its natural, unprocessed form, without exine-disruption treatments, which may have limited its nutritional utilization.
Feed conversion ratio further supports this interpretation. A tendency toward reduced feed efficiency was observed as pollen inclusion levels increased. This reduction in efficiency suggests that birds require a greater amount of feed to achieve one unit of body weight gain when consuming diets containing higher concentrations of Melipona bee pollen. Overall, these results indicate that bee pollen acts primarily as a functional feed additive with bioactive properties rather than as a direct nutritional replacement ingredient. Similar conclusions were reported by Nemauluma et al. (2022, 2023) [21,22], who observed that bee pollen inclusion in broiler diets does not necessarily result in linear improvements in weight gain, particularly at high inclusion levels or when pollen is not processed to facilitate exine disruption.
Results of carcass yield and its main components (breast, thigh, drumstick, and wing) are particularly relevant, as it indicates that although total weight gain was lower in 8 and 12 g of pollen, carcass tissue composition remained proportional. This finding is consistent with the results reported by Nemauluma et al. (2023) [22], who likewise observed no alterations in body composition under similar experimental conditions. The absence of differences in the proportion of prime cuts suggests that the adverse effect of bee pollen on weight gain was not associated with changes in nutrient partitioning among tissues, but rather with a reduction in the total amount of tissue accreted. Additionally, the stability in the weights of immune-related organs (thymus, bursa of Fabricius, and spleen) suggests that, although bee pollen may exert immunomodulatory effects, it does not induce detectable morphological changes in these tissues under the conditions evaluated [23].
The presence of bioactive compounds such as flavonoids, phenolic compounds, and other antioxidants in the pollen structure offers potential immunological and physiological benefits, as reflected in hematological parameters. However, these benefits do not necessarily translate into increased body weight, since the metabolic energy required to process the pollen matrix with the intact exine competes with the energy available for somatic growth. Hemoglobin concentration and hematocrit values did not differ significantly the absence of changes in these parameters indicates that pollen inclusion did not affect oxygen-carrying capacity or induce anemia [24]. Hematological results support the hypothesis that Melipona bee pollen exerts its primary effects at the immunomodulatory level. The increased lymphocyte and monocyte count with higher pollen inclusion levels suggest stimulation of the immune system [18]. Eosinophils showed a greater response with 8 g/kg of feed, which could indicate increased immune activity or a mild inflammatory response triggered by the bioactive compounds present in the pollen [4,17]. This response pattern suggests activation of the immune system [25]. Pollen grains contain a complex matrix of immunologically active compounds. Khalifa et al. (2021) [4] and El Ghouizi et al. (2023) [2] extensively documented the immunomodulatory properties of bee pollen, attributed to its content of polysaccharides, proteins, flavonoids, and other secondary metabolites. Although this immune activation is beneficial for host defense, it requires substantial energy partitioning, which is reflected in the reduced somatic growth rate observed in treatments with higher pollen inclusion levels. Haščík et al. (2019) [19] reported similar findings in broilers supplemented with bee products, indicating that although body weight gain was affected, carcass quality and cut composition remained within acceptable ranges. This outcome has important practical implications for production systems in which carcass quality is a priority.
These results are consistent with previous studies indicating that supplementation, even at moderate concentrations, does not exert significant effects on variables such as bone weight or length in poultry with similar growth characteristics, including broilers and quail. However, other investigations have reported improvements in physicochemical properties of bone tissue associated with pollen supplementation, although under different experimental conditions and dosage levels, suggesting a potential dose-dependent response or modulation influenced by species and age [26,27]. On the other hand, some authors have reported unfavorable effects on bone mechanical strength when pollen supplementation exceeds certain levels, indicating that its inclusion does not always result in improved bone quality and may, in some cases, delay maturation or reduce tissue mineralization [27]. However, in the present study, the absence of variations in bone density and bone weight suggests that such adverse effects did not occur. The absence of significant differences among treatments may be attributed to the possibility that the inclusion levels tested were insufficient to elicit detectable histomorphometric changes, or to the adaptive capacity of slow-growing chickens, whose reduced growth rate may be associated with a more stable bone metabolism in response to nutritional interventions of this nature.
5. Conclusions
It can be concluded that the inclusion of 4 g/kg of Melipona bee pollen in diets for slow-growing chickens can be used as a natural enhancer without impairing productive performance. Moderate inclusion of bee pollen may provide immunological benefits without adversely affecting growth or general health. Furthermore, the levels of pollen evaluated did not produce alterations in the basic bone histomorphometry of the tibia of slow-growing chickens.
Further studies are recommended to investigate a broader range of inclusion levels and combinations of stingless bee products, with the aim of identifying optimal dosages capable of enhancing their effects in poultry production systems. Future research should incorporate a comprehensive assessment of productive, physiological, and biomechanical parameters to verify the potential structural and functional benefits of bee pollen on bone tissue and overall bird performance.
Author Contributions
J. V. Aguilar Vásquez: Conceptualization, Funding acquisition, Methodology, Project administration, Supervision, Writing – review & editing. W. Celis: Conceptualization, Supervision, Validation, Writing – original draft. M. A. Mathios-Flores: Data curation, Formal analysis, Visualization. J. Cáceres-Coral: Methodology, Resources, Software. N. C. Vilchez: Validation, Visualization, Writing – original draft. D. L. del Castillo: Investigation, Methodology. P. Mendoza-Tamani: Data curation, Formal analysis, Software. C. P. Perea: Investigation, Methodology. J. R. Lancha: Data curation, Formal analysis.
Funding
The authors express their gratitude to the Programa Nacional de Investigación Científica y Estudios Avanzados (PROCIENCIA – Perú) for financial support (CONTRACT Nº PE501086376-2024-PROCIENCIA), to the Universidad Nacional Autónoma de Alto Amazonas for logistical assistance, and to the Universidad Nacional Agraria La Molina for providing complementary laboratory analyses.
Institutional Review Board Statement
The animal research procedures were carried out in accordance with the guidelines of the Scientific Integrity Committee for the use of animals of the National Autonomous University of Alto Amazonas, Report No. 001-2024-UNAAA/CIC (24 December 2025, Yurimaguas, Peru).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data appear in the paper. However, additional data that support the findings of this study are available from the corresponding author, José Virgilio Aguilar Vásquez (jaguilar@unaaa.edu.pe).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Abd El-Aziz, A; Abo Ghanima, M; Mota-Rojas, D; Sherasiya, A; Ciani, F; El-Sabrout, K. Bee Products for Poultry and Rabbits: Current Challenges and Perspectives. Animals [Internet] 2023, 13(22), 3517. Available online: https://www.mdpi.com/2076-2615/13/22/3517. [CrossRef]
- El Ghouizi, A; Bakour, M; Laaroussi, H; Ousaaid, D; El Menyiy, N; Hano, C; et al. Bee Pollen as Functional Food: Insights into Its Composition and Therapeutic Properties. Antioxidants [Internet] 2023, 12(3), 557. Available online: https://www.mdpi.com/2076-3921/12/3/557. [CrossRef]
- Tsalsabilla, AT; Susilo, A; Amertaningtyas, D. Dietary Applications of Bee Pollen: Nutritional Composition, Health Benefits, and Applications. Asian J Food Res Nutr [Internet] 2025, 4(3), 758–66. Available online: https://journalajfrn.com/index.php/AJFRN/article/view/286. [CrossRef]
- Khalifa, SAM; Elashal, MH; Yosri, N; Du, M; Musharraf, SG; Nahar, L; et al. Bee Pollen: Current Status and Therapeutic Potential. Nutrients [Internet] 2021, 13(6), 1876. Available online: https://www.mdpi.com/2072-6643/13/6/1876. [CrossRef]
- Cavallero, A; Vidotto, F; Sbrana, C; Peres Fabbri, L; Petroni, G; Gabriele, M. Antioxidant-Rich Polyfloral Bee Pollen Exerts Antimicrobial Activity and Anti-Inflammatory Effect in A549 Lung Epithelial Cells by Modulating the NF-κB Pathway. Foods [Internet] 2025, 14(5), 802. Available online: https://www.mdpi.com/2304-8158/14/5/802. [CrossRef]
- Anjum, SI; Ullah, A; Gohar, F; Raza, G; Khan, MI; Hameed, M; et al. Bee pollen as a food and feed supplement and a therapeutic remedy: recent trends in nanotechnology. Front Nutr 2024, 11. [Google Scholar] [CrossRef]
- Urcan, AC; Criste, AD; Dezmirean, DS; Bobiș, O; Bonta, V; Burtescu, RF; et al. Enhancing Antioxidant and Antimicrobial Activities in Bee-Collected Pollen through Solid-State Fermentation: A Comparative Analysis of Bioactive Compounds. Antioxidants [Internet] 2024, 13(3), 292. Available online: https://www.mdpi.com/2076-3921/13/3/292. [CrossRef]
- Algethami, JS; El-Wahed, AAA; Elashal, MH; Ahmed, HR; Elshafiey, EH; Omar, EM; et al. Bee Pollen: Clinical Trials and Patent Applications. Nutrients [Internet] 2022, 14(14), 2858. Available online: https://www.mdpi.com/2072-6643/14/14/2858. [CrossRef]
- Pinheiro, SRF; Dourado, LRB; Silva, EP. Nutrição de aves caipiras criadas em sistema semiconfinado. In Nutrição de Não Ruminantes; Sakomura, NK, Silva, JH., Costa, FG., Fernandes, JB., Hauschild, L, Eds.; FUNEP: Jaboticabal, 2014; p. 678. [Google Scholar]
- Sakomura, NK; Rostagno, HS. Métodos de Pesquisa em Nutrição de Monogástricos, 2da ed.; Funep, 2016; p. 262 p. [Google Scholar]
- Yaeger, R. Atlas of Avian Hematology. Am J Trop Med Hyg [Internet] 1963, 12(1), 118. Available online: https://www.ajtmh.org/view/journals/tpmd/12/1/article-p118_a.xml. [CrossRef]
- Campbell, TW. Exotic Animal Hematology and Cytology; Wiley, 2015. [Google Scholar]
- Onasanya, GO; Oke, FO; Sanni, TM; Muhammad, AI. Parameters Influencing Haematological, Serum and Bio-Chemical References in Livestock Animals under Different Management Systems. Open J Vet Med [Internet] 2015, 05(08), 181–9. Available online: http://www.scirp.org/journal/doi.aspx?DOI=10.4236/ojvm.2015.58025. [CrossRef]
- Sánchez-Torres, L; Arredondo-Castro, M; Orozco-Benítez, G; Gutiérrez-Arenas, D; Carrillo-Beltrán, J; Lepe-Aguilar, R; et al. Variables hematológicas en aves deportivas, ganso común, pato doméstico, pato azteca, guajolote y pollo de engorda. In Abanico Agrofor; Jan 2021; p. 3. [Google Scholar]
- Aguilar, V. J; Zea, M. O; Vílchez, P. C. Rendimiento Productivo e Integridad Ósea de Pollos de Carne en Respuesta a Suplementación Dietaria con Cuatro Fuentes de Fitasa Comercial. Rev Investig Vet del Perú [Internet] 2018, 29(1), 169–79. Available online: https://revistas.gnbit.net/index.php/veterinaria/article/view/14078. [CrossRef]
- Li, QQ; Wang, K; Marcucci, MC; Sawaya, ACHF; Hu, L; Xue, XF; et al. Nutrient-rich bee pollen: A treasure trove of active natural metabolites. J Funct Foods [Internet] 2018, 49, 472–84. Available online: https://linkinghub.elsevier.com/retrieve/pii/S1756464618304845. [CrossRef]
- Batista Rodrigues, R; Avelar Pucci, LE; Uczay, J; Molinari, M; Lazzari, R; Uczay, M. Pólen apícola como aditivo em dietas para frangos de corte. Nativa [Internet] 2018, 6(5), 551–6. Available online: https://periodicoscientificos.ufmt.br/ojs/index.php/nativa/article/view/5865. [CrossRef]
- Al-Kahtani, SN; Alaqil, AA; Abbas, AO. Modulation of Antioxidant Defense, Immune Response, and Growth Performance by Inclusion of Propolis and Bee Pollen into Broiler Diets. Animals [Internet] 2022, 12(13), 1658. Available online: https://www.mdpi.com/2076-2615/12/13/1658. [CrossRef]
- Haščík, P; Pavelková, A; Arpášová, H; Čuboň, J; Kačániová, M; Kunová, S. The effect of bee products and probiotics on meat performance of broiler chickens. J Microbiol Biotechnol food Sci [Internet] 2019, 9(1), 88–92. Available online: https://office2.jmbfs.org/index.php/JMBFS/article/view/8894. [CrossRef]
- Hashmi, MS; Haseik, P; Elimam, I; Garlik, J; Marek, B; Miroslava, K. Effects of Bee Pollen on the Technical and Allocative Efficiency of Meat Production of Ross 308 Broiler. Int J Poult Sci [Internet] 2012, 11(11), 689–95. Available online: https://www.scialert.net/abstract/?doi=ijps.2012.689.695. [CrossRef]
- Nemauluma, MFD; Ng’ambi, JW; Kolobe, SD; Malematja, E; Manyelo, TG; Chitura, T. Bee pollen an alternative to growth promoters for poultry production-A review. Appl Ecol Environ Res [Internet] 2022, 20(5), 3817–32. Available online: https://aloki.hu/pdf/2005_38173832.pdf. [CrossRef]
- Nemauluma, MFD; Manyelo, TG; Ng’ambi, JW; Kolobe, SD; Malematja, E. Effects of bee pollen inclusion on performance and carcass characteristics of broiler chickens. Poult Sci [Internet] 2023, 102(6), 102628. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0032579123001529. [CrossRef]
- Prakatur, I; Miskulin, M; Pavic, M; Marjanovic, K; Blazicevic, V; Miskulin, I; et al. Intestinal Morphology in Broiler Chickens Supplemented with Propolis and Bee Pollen. Animals [Internet] 2019, 9(6), 301. Available online: https://www.mdpi.com/2076-2615/9/6/301. [CrossRef]
- Sevim, B. Effects of supplemental bee pollen on performance, meat quality, serum constituents and immunity system in growing quails. S Afr J Anim Sci [Internet] 2022, 51(6), 745–51. Available online: https://www.ajol.info/index.php/sajas/article/view/224910. [CrossRef]
- Genova, JL; Rodrigues, RB; Martins, JS; Uczay, M; Henriques, JKS. Própolis e pólen apícola na nutrição de animais não ruminantes. Arch Zootec [Internet] 2020, 69(265), 124–31. Available online: http://www.uco.es/ucopress/az/index.php/az/article/view/5048. [CrossRef]
- Korani, S; Khalesi, N; Korani, M; Jamialahmadi, T; Sahebkar, A. Applications of honeybee-derived products in bone tissue engineering. Bone Reports [Internet] 2024, 20, 101740. Available online: https://linkinghub.elsevier.com/retrieve/pii/S235218722400007X. [CrossRef] [PubMed]
- Tomaszewska E, Knaga S, Dobrowolski P, Lamorski K, Jabłoński M, Tomczyk-Warunek A, et al. The effect of bee pollen on bone biomechanical strength and trabecular bone histomorphometry in tibia of young Japanese quail (Coturnix japonica). Loor JJ, editor. PLoS One [Internet]. 2020 Mar 18;15(3):e0230240. Available from: https://dx.plos.org/10.1371/journal.pone.0230240.
Table 1.
Composition of experimental diets including bee pollen.
| Ingredient % | T control | T 1 | T 2 | T 3 |
|---|---|---|---|---|
| Fishmeal | 6.00 | 6.00 | 6.00 | 6.00 |
| Soy meal | 25.10 | 25.10 | 25.10 | 25.10 |
| Corn | 66.01 | 65.66 | 65.36 | 65.94 |
| Bee pollen | 0.00 | 0.40 | 0.80 | 1.20 |
| Calcium carbonate | 1.10 | 1.10 | 1.10 | 1.10 |
| Monocalcium phosphate | 0.80 | 0.80 | 0.80 | 0.80 |
| Choline chloride | 0.20 | 0.20 | 0.20 | 0.20 |
| Common salt | 0.20 | 0.20 | 0.20 | 0.20 |
| vit-min1 supplement | 0.10 | 0.10 | 0.10 | 0.10 |
| DL – methionine | 0.30 | 0.30 | 0.30 | 0.30 |
| L – lysine | 0.05 | 0.05 | 0.05 | 0.05 |
| Zinc bacitracina | 0.05 | 0.00 | 0.00 | 0.00 |
| Coccidiostat | 0.04 | 0.04 | 0.04 | 0.04 |
| Fungicab | 0.05 | 0.05 | 0.05 | 0.05 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 |
1 Vitamin and mineral supplement: Vitamin A (3,600,000-14,300,000 IU), vitamin D3 (1,090,000-3,300,000 IU), vitamin E (7270-110,000 IU), vitamin K3 (0.51-6.10 g), vitamin B1 (0.91-3.30 g), vitamin B2 (2.28-48.40 g), nicotinic acid (13.60-242 g), calcium pantothenate (4.55-60.50 g), vitamin B12 (0.009-0.033 g), vitamin B6 (1.64-6.60 g), folic acid (0.009-2.20 g), biotin (0.018-0.28 g), manganese oxide (49.14-110 g), zinc oxide (27.30-110 g), ferrous sulfate (1.36-110 g), copper sulfate (1.36-110 g), potassium iodide (0.38-4.40 g), cobalt carbonate (0.09-0.22 g), sodium selenite (0.09-0.33 g), ethoxyquin (20-130 g).
Table 2.
Chemical composition of experimental diets and pollen.
| Food | Humidity1 | Dry matter1 | Mineral matter1 | Ethereal extract1 | Crude fiber2 | Crude protein2 | Nitrogen-free extract3 | Gross energy2 kcal/kg |
|---|---|---|---|---|---|---|---|---|
| Control treatment | 11.84 | 88.16 | 6.64 | 2.89 | 2.18 | 20.51 | 55.95 | 3844 |
| Treatment 1 | 11.90 | 88.10 | 6.52 | 1.96 | 2.31 | 21.04 | 56.27 | 3897 |
| Treatment 2 | 11.93 | 88.07 | 6.33 | 2.02 | 2.49 | 21.84 | 55.40 | 3879 |
| Treatment 3 | 11.92 | 88.08 | 6.08 | 1.98 | 2.30 | 21.17 | 56.55 | 3876 |
| Pollen | 14.34 | 85.66 | 5.45 | 2.80 | 0.45 | 45.55 | 31.40 | 5259 |
1 Analyses carried out at the Biotechnology Laboratory of the National Autonomous University of Alto Amazonas – Yurimaguas, Peru. 2 Analyses carried out in the Laboratory of Nutritional Evaluation of Foods of the National Agrarian University La Molina – Lima, Peru. 3 Value determined by difference.
Table 3.
Productive performance of slow-growing chickens fed with bee pollen.
| Productive performance | TC | T1 | T2 | T3 | SEM | P-value |
|---|---|---|---|---|---|---|
| Initial weight (g) | 38.8 | 38.6 | 38.8 | 38.8 | 0.212 | 0.8795 |
| Final weight (g) | 780.2a | 772.2a | 713.4b | 704.0b | 11.707 | 0.0003 |
| Weight gain (g) | 741.2a | 733.4a | 674.8b | 665.2b | 11.707 | 0.0003 |
| Feed intake (g) | 1319.2 | 1344.4 | 1283.0 | 1300.0 | 24.042 | 0.3469 |
| Feed conversion (g/g) | 1.779b | 1,835ab | 1,904ab | 1.957a | 0.043 | 0.0494 |
Table 4.
Carcasses yield and relative weight noble cuts, edible viscera and lymphoid organs of slow-growing chickens fed with bee pollen.
Table 4.
Carcasses yield and relative weight noble cuts, edible viscera and lymphoid organs of slow-growing chickens fed with bee pollen.
| Variables | TC | T1 | T2 | T3 | SEM | P-value |
|---|---|---|---|---|---|---|
| Carcass (%) | 62.76 | 60.91 | 60.89 | 61.33 | 0.917 | 0.4494 |
| Breast (%) | 25.03 | 22.77 | 23.791 | 23.91 | 0.961 | 0.4527 |
| Thigh (%) | 15.92 | 16.71 | 16.94 | 15.99 | 0.504 | 0.4065 |
| Drumstick (%) | 17.53 | 17.22 | 17.47 | 17.79 | 0.487 | 0.8797 |
| Wing (%) | 14.99 | 15.11 | 15.34 | 15.05 | 0.300 | 0.8586 |
| Heart (%) | 0.87 | 0.89 | 0.96 | 0.91 | 0.023 | 0.0743 |
| Gizzard (%) | 3.57 | 3.93 | 3.93 | 3.94 | 0.183 | 0.4111 |
| Liver (%) | 3.39 | 3.39 | 3.19 | 3.20 | 0.104 | 0.3621 |
| Timo (%) | 0.61 | 0.64 | 0.59 | 0.78 | 0.086 | 0.4338 |
| Bursa (%) | 0.57 | 0.55 | 0.54 | 0.51 | 0.048 | 0.8767 |
| Spleen (%) | 0.23 | 0.22 | 0.24 | 0.23 | 0.018 | 0.9070 |
The means followed by different letters in the row differ from each other according to Tukey’s test with a probability of 5%.
Table 5.
Hematological values of slow-growing chickens fed bee pollen1.
| Variables | TC | T1 | T2 | T3 | SEM | P-value |
|---|---|---|---|---|---|---|
| Hematocrit (g/dL) | 27.00 | 27.80 | 26.90 | 28.40 | 0.639 | 0.3315 |
| Hemoglobin (g/dL) | 8.91 | 9.18 | 8.88 | 9.37 | 0.211 | 0.3364 |
| Leukocytes (cells/mm3) | 44600b | 43000b | 37000b | 71100a | 2736.101 | <0.0001 |
| Heterophiles (%) | 37.40 | 28.10 | 38.50 | 40.20 | 4.045 | 0.1882 |
| Lymphocytes (%) | 52.50ab | 57.40a | 42.30b | 46.20ab | 3.359 | 0.0274 |
| Eosinophils (%) | 7.30ab | 9.70a | 11.30a | 4.40b | 1.228 | 0.0061 |
| Basophils (%) | 0.60 | 0.70 | 0.90 | 0.60 | 0.260 | 0.8275 |
| Monocytes (%) | 2.20b | 4.10ab | 7.00ab | 8.60a | 1.246 | 0.0099 |
1 Analysis carried out in the Clinical Laboratory of the Medical Center of the National Autonomous University of Alto Amazonas. The means followed by different letters in the row differ from each other according to Tukey’s test with a probability of 5%.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



