Submitted:
12 February 2026
Posted:
13 February 2026
You are already at the latest version
Abstract
Background/Objectives: This article is a narrative review that examines the development of attachment from intrauterine life to the first thousand days of a child's life, integrating psychoanalytic, neuroscientific, genetic, and cross-cultural perspectives. Biological, relational, neurological, and cultural factors interact and determine individual differences in socio-emotional functioning. This paper aims to propose a reinterpretation of early attachment, describing it as both a clinical and relational phenomenon and an adaptive process inscribed in human evolutionary history, according to the described Four-Domain Integrative Framework.. Methods: The review examined three main areas of evidence: early attachment characteristics, cross-cultural caregiving variations, and genetic and epigenetic mechanisms underlying environmental sensitivity. Results: The first identified seven characteristics of early attachment (proximity seeking, emotional attunement, intrauterine experiences, maternal holding, security patterns, brain plasticity, and maternal stress) which represent developmental mechanisms that generate individual differences in trust, self-regulation, resilience, and psychopathological vulnerability. Second, cross-cultural variations in six distinct caregiving contexts were examined, demonstrating that secure attachment emerges through culturally specific pathways, differentially influencing motor development, sleep patterns, hypothalamic-pituitary-adrenal axis maturation axis maturation, and social skills. Finally, the differential susceptibility model was provided through the analysis of five genetic and epigenetic systems (oxytocin receptor gene, serotonin transporter gene, dopamine receptor gene, glucocorticoid receptor methylation, and fetal programming) that modulate environmental sensitivity. Conclusions: Biological, relational, neurological, and cultural factors interact and determine individual differences in socio-emotional functioning.
Keywords:
attachment
; early
; relationships
; psychoanalysis
; developmental neuroscience
; epigenetics
; brain plasticity
; emotion regulation
1. Introduction
Early emotional bonding is one of the key factors in human development. The relational experiences of the first thousand days of life—from gestation to early childhood—form the foundation on which the mind, the self, and the ability to relate to others are built. Psychoanalysts have always emphasized the importance of the primary relationship, from Bowlby’s attachment theories to Winnicott’s reflections on the role of the mother-environment. In recent decades, however, affective neuroscience has provided a solid basis for psychoanalytic theories by showing how interpersonal experiences in early childhood directly influence brain development, emotional regulation, and psychological resilience [1]. We will analyze some of the contributions from these disciplines, bringing together theoretical models and empirical data to understand how early emotional bonding a biological, psychological, and relational matrix is underlying human development. In this sense, intrauterine life has become a subject of fascination and reflection for those who study the birth and development of the human mind.
Freud [2] had already hypothesized that “There is much more continuity between intrauterine life and early childhood than the impressive break of the act of birth would have us believe” (p.286). Following this profound reflection, we can think about the continuum that characterizes life before and after birth and the impact, in evolutionary terms, that this prenatal space has on the future of the individual.Perinatal psychology invites us to reconsider the origin of individual developmental trajectories. Personal narratives do not begin at birth, but are already rooted in the prenatal stages of development. Earl relational and somatic experiences constitute a pre-history that shapes the emotional and psychological future of the individual.
Contrary to the traditional view that considered the fetus a passive organism, the most recent research shows that as early as the sixteenth week of gestation, the fetus exhibits exploratory behaviors, sensory responses, and implicit learning abilities. Intrauterine experiences—such as the tone of the mother’s voice, heart rate, and hormonal changes—are stimuli that the fetus perceives and integrates, contributing to the formation of bodily and emotional memory. Alessandra Piontelli’s pioneering studies [3] have documented a significant continuity between fetal behavioral characteristics and postnatal temperament. Her work has highlighted how individual differences predictive of subsequent development already emerge in the prenatal period. The research of psychoanalysts such as Negri and Piontelli [4], applied to the study of fetal behavior using ultrasound investigation techniques, has shown that the fetus is immersed in a relational environment that stimulates and shapes it, anticipating the construction of the bodily and affective self. It is an implicit dialogue that the mother, together with her environment, builds with the fetus in utero. Even before birth, the child is involved in an intersubjective relationship with the mother, mediated by neurophysiological and affective signals.
The impact of the mother, as the one who contains, and of the environment in which the woman is placed, as a space that in turn contains the mother-child dyad, is fundamental for the construction of the fetus’s mind, which is actively involved in this work of interaction and construction of a very early intersubjectivity. Neuroscientific studies [5] show, for example, that maternal stress, depression, and traumatic xperiences during pregnancy can alter the brain connectivity of the fetus, particularly in the amygdala and the hypothalamic-pituitary-adrenal axis, predisposing the child to difficulties in emotional regulation and stress response.
Prenatal sensory and motor experiences are stored in implicit memory, a predominantly emotional system that matures in the last trimester of pregnancy and involves the amygdala, basal ganglia, cerebellum, and cortical areas of the right hemisphere [6,7,8]. Neural connectivity develops progressively from the twenty fourth week, with a predominance of the right hemisphere until the first two years of life [9,10,11]. From the thirtieth week, the main connections that constitute the neurobiological substrate for basic affective functions are established. Fetal brain maturation follows a caudal-rostral progression (from the brainstem to the cerebral cortex). By 28 weeks, a biological framework is created that supports the emergence of autonomous primary mental activity [12]. Alterations in this early connectivity are implicated in developmental disorders, from autism spectrum disorders to major depression. These neurological maturation processes continue after birth, requiring appropriate relational experiences for the normal development of the limbic nuclei. The right amygdala and anterior cingulate cortex are strongly shaped by early caregiving experiences. These structures play a primary role in the development and maintenance of emotional and social attachment. From an evolutionary perspective, early attachment represents an adaptive strategy aimed at survival: the search for proximity to the caregiver, the ability for emotional attunement, and the emergence of individual differences already in the prenatal phase allow the child to adapt functionally to the environment. These characteristics not only promote immediate survival, but also constitute fundamental elements for the construction of personality, affective-relational regulation, and motivation in adulthood. To ensure clarity and consistency in the presentation, it is necessary to clarify the use of certain key terms which, although referring to related phenomena, are not perfectly interchangeable (Table 1).
1.1. The Four-Domain Integrative Framework: A Dynamic Model of Early Attachment
The development of early attachment is an extremely complex phenomenon that cannot be fully understood through single-perspective approaches. Different perspectives have analyzed this phenomenon in parallel, generating a fragmented understanding that constitutes a theoretical and clinical-applicative limitation and requires integration. While psychoanalytic theory has emphasized the relational dynamics that characterize early attachment, neuroscience has focused primarily on the brain mechanisms involved in the development of this phenomenon. Genetics has examined the influence of hereditary predispositions on attachment, and cross-cultural research has revealed contextual variations.
In light of these considerations, we propose a Four-Domain Integrative Framework that explains the dynamic, bidirectional interactions between: (1) Biology/Genetics, (2) Neuroscience, (3) Relational Experiences, and (4) Cultural Contexts.
This framework differs from traditional biopsychosocial models for several reasons. First, it emphasizes temporal cascades rather than static interactions. Rather than treating biological, psychological, and social factors as additive and relatively stable elements, our integrative framework emphasizes the dynamic interactions between these factors throughout development. Genetic predispositions do not add to environmental influences but create windows of differential susceptibility during which neural systems are most plastic and responsive to relational inputs. For example, variants of the short 5-HTTLPR allele have been associated with greater amygdaloid reactivity and greater sensitivity to environmental inputs [13,14], making this period a window of vulnerability in the case of adverse caregiving, but also of opportunity in supportive conditions [15].
Second, our integrative framework recognizes bidirectional causality at all levels. While traditional biopsychosocial models assume a unidirectional or additive relationship between different domains, our integrative framework emphasizes the properties that emerge from their interactions. Cultural practices (e.g., co-sleeping) do not simply reflect attachment patterns, but modulate physiological regulation, including potentially the HPA axis and cortisol rhythms [16], which in turn influence the child’s ability to regulate affect, thereby reinforcing culturally adaptive behavioral patterns. Similarly, maternal attunement is closely linked to oxytocin activity: recent studies show that oxytocin reactivity in infants and mothers during interaction is associated with maternal sensitivity and the subsequent development of secure attachment [17,18], demonstrating how relational experiences recursively modify the biological substrate.
Thirdly, it integrates an evolutionary-adaptive perspective that reformulates vulnerability as context dependent plasticity, rather than limiting itself to a dichotomous categorization of evolutionary outcomes in terms of health or pathology. What appears as “insecure attachment” in Western individualistic frameworks may represent adaptive strategies in collectivist or high-risk ecologies [19,20]. The differential susceptibility model [21] illustrates this principle: “orchid children” (carriers of sensitivity alleles) develop optimally in favorable environments but show greater vulnerability in adverse contexts,
while “dandelion children” remain stable in all conditions. This genetic diversity represents an evolutionary hedging strategy that maintains the adaptability of the population.
1.1.1. Points of mechanistic integration
The framework operates through four main mechanisms:
A. Epigenetic programming as biological memory of relational experience
The quality of maternal care modulates methylation patterns of the glucocorticoid receptor (NR3C1) during the prenatal and early postnatal periods [22,23], influencing the reactivity of the HPA axis. These epigenetic changes constitute a biological “memory” of the relational context, which interacts with genetic predispositions and cultural context without rigidly determining outcomes [14,15].
Concrete examples of this mechanism emerge from recent studies. Research conducted during the COVID-19 pandemic showed that mothers and newborns exposed to lockdown during the first trimester of pregnancy had significantly lower levels of methylation of NR3C1 and SLC6A4.COVID-19 pandemic showed that mothers and newborns exposed to lockdown during the first trimester of pregnancy had significantly lower levels of NR3C1 and SLC6A4 methylation than those exposed in the second or third trimester, with effects persisting even after controlling for confounding variables [24]. This evidence highlights how specific time windows during gestation represent critical periods of epigenetic susceptibility to maternal stress. Furthermore, longitudinal studies have documented that increased methylation of NR3C1 in infants, associated with prenatal maternal depression, predicts increased cortisol responses to stress at 3 months of age [25]. These epigenetic patterns are not deterministic but plastic: research on animal models shows that high-quality maternal care can reduce Nr3c1 methylation even in the presence of previous adverse experiences [22,23] suggesting windows of opportunity for early interventions that can remodel epigenetic programming.
B. Neural plasticity as a substrate of relational internalization
During the first years of life, the limbic and prefrontal circuits, together with the right hemisphere, show high plasticity and sensitivity to relational inputs [8,9,10]. The quality of caregiving and maternal attunement modulate these neural networks, promoting the regulation of emotions and the internalization of secure relational patterns [17,18]. Cultural differences in caregiving, such as the degree of physical contact and interactive synchrony, indirectly influence the development of structures such as the amygdala and insula, highlighting how relational experience and cultural context shape early neural plasticity [26].
Neural plasticity as a substrate for relational internalization is confirmed by specific evidence. Neuroimaging studies have revealed that mothers with greater responsive sensitivity show more coordinated activation patterns in the limbic and prefrontal regions during interaction with their children [27]. The role of oxytocin is particularly relevant: longitudinal research has shown that maternal sensitivity predicts secure infant attachment only when accompanied by an increase in maternal oxytocin during parent-child interaction, highlighting the bio-relational nature of this process [28]. Furthermore, low levels of basal maternal oxytocin at 3 months significantly predict emotional reactivity and withdrawal behaviors in the child at 3.5 years, even when controlling for maternal symptomatology and adult attachment style [29]. These data demonstrate how early relational experience, mediated by neurochemical systems such as oxytocin, literally embeds itself in emerging brain structures, creating neural templates that persist over time and influence subsequent socio-emotional functioning.
C. Cultural practices as scaffolds for gene expression and neural development
Cross-cultural variations in caregiving represent evolutionary adaptations that shape development according to environmental specificities. For example, co-sleeping in collectivist cultures maintains mother-child physiological synchrony and may influence cortisol rhythms, indirectly contributing to adaptive emotional regulation strategies [16]. These practices interact with gene variants such as OXTR, influencing attachment outcomes [30,31]. The interaction between cultural practices and genetic expression is supported by specific cross-cultural evidence. A comparative study has shown that co-sleeping, prevalent in collectivist cultures, not only maintains mother-child physiological synchrony but also modulates circadian cortisol rhythms, indirectly influencing the development of adaptive emotional regulation strategies [16]. These cultural practices are not biologically neutral: they interact with genetic variants such as OXTR rs53576, where the G allele is associated with greater social sensitivity and secure attachment, but its phenotypic effects vary significantly depending on the cultural context of caregiving [32].
Furthermore, recent research on babywearing (continuously carrying the baby) has shown that prolonged physical contact increases the release of oxytocin in both parents and children, while simultaneously reducing cortisol levels [18,26]. This example illustrates how specific cultural practices act as “scaffolds” that simultaneously shape gene expression, neural development, and behavioral patterns, creating culturally specific but biologically rooted developmental trajectories.
D. Developmental cascades: from the molecular to the behavioral level
The framework describes how genetic, epigenetic, neural, relational, and cultural factors influence each other throughout development:
1. Genetic: for example, DRD4 variants confer greater environmental sensitivity [13].
3. Neural: the plasticity of limbic and prefrontal circuits makes the brain receptive to relational experience [17,18].
4. Relational: the quality of maternal attunement or the extended network modulates emotional regulation and attachment patterns.
5. Cultural: Cultural practices can amplify or attenuate the effects of other domains, promoting adaptations consistent with the environment [16,19,33].
6. Behavioral outcome: By early childhood, differences in emotional regulation, social cognition, and exploratory behavior emerge, reflecting the integration of all four domains.
A paradigmatic example of this integrated cascade emerges from prospective longitudinal studies. Children carrying the short 5-HTTLPR variant (genetic level), exposed to prenatal maternal stress that alters NR3C1 methylation (epigenetic level), show increased amygdala reactivity to emotional stimuli in early life (neural level). This neural hyperreactivity predicts difficulties in emotional regulation observed during mother-child interactions (relational level), difficulties that may be amplified or attenuated by cultural caregiving practices such as co-sleeping or the availability of alloparental networks (cultural level). The behavioral outcome at 3-5 years of age, which includes differences in emotional regulation, social cognition, and exploratory behavior, thus reflects the dynamic integration of all these domains, rather than the additive contribution of each [23,24,34].
2. The First Thousand Days of Life: Neuroplasticity and Environment
The neural plasticity of the fetus and then of the child clearly shows how much researchers and clinicians focus on the environment as a determining factor in the development of the self, [35] Psychoanalytic literature provides us with solid theories on the importance of building the mother-child bond as the basis for this development.
At birth, the newborn is an active and competent subject, with an innate predisposition to relationships. The first interactions with the mother—through gaze, physical contact, and voice—constitute the first affective language, which allows the child to build a sense of continuity and internal coherence. This language is, as already described, implicit (embodied): interaction is contact, gaze, word/sound, movement, play.
Interaction conveys affection to the child in continuity with the prenatal stages of development. Winnicott [36] described the concept of “holding” as the maternal function of physical and emotional containment, which allows the child to feel safe and develop trust in the environment. Bion [37], with his theory of the α function, emphasized the importance of the maternal ability to transform raw experiences (β elements) into thinkability, promoting the birth of thought and mind. These qualities of bonding are the basis of healthy child development.
Early interactions are not simple behavioral exchanges, but deeply neurobiological processes that shape brain structure and mentalization capacity [38].
According to Bowlby’s attachment theory [39], children are biologically predisposed to seek the closeness of a caregiver in order to feel protected. Neuroscience confirms that secure attachment is associated with greater integration between the areas of the brain involved in emotional regulation, memory, and stress response. Maternal sensitivity—the ability to perceive, interpret, and respond appropriately to the signals of the newborn—is crucial for the maturation of these circuits. Edward Tronick [40], with his famous ‘still-face’ experiment, demonstrated how a lack of emotional reciprocity, even for short periods, can cause distress in children, highlighting the importance of intersubjective regulation for neurobehavioral development.
One might wonder why the first thousand days of life are the focus of so much attention. The first thousand days of life represent a critical window for brain development. During this period, the brain is characterized by extraordinary synaptic plasticity, which allows for rapid growth and reorganization of neural connections. At no other time do synapse formation and myelination grow so rapidly, after which they undergo a process of remodeling.
From the last trimester of pregnancy, there is an asymmetry in the growth of the hemispheres: the right hemisphere is more developed than the left until the latter catches up in a growth spurt from the second year of life. This means that brain development in this window favors the formation of attachment bonds and affective regulation, both of which are dominated by the right brain.
Early relational experiences, particularly those with the mother figure, modulate the activation of fundamental neurochemical circuits, such as those involving oxytocin, dopamine, and serotonin. These neurotransmitters not only regulate mood and attachment, but also influence the maturation of higher cortical structures. This last element helps us understand how even higher cognitive processes, learning, and thinking are the result of more archaic and unconscious emotional learning.
Early affective dysregulation, caused by trauma, neglect, or relational deficiencies, can compromise neuronal growth, alter the response to stress, and predispose individuals to mental disorders in adulthood, such as depression, anxiety, dissociative disorders, and relational difficulties. Many syndromes and diagnostic labels currently used in the field of development to classify forms of pathological adaptation take on a deeper meaning thanks to a more complex interpretation such as that of the intersubjective paradigm.
Antonio Imbasciati has broadened the perspective of child psychoanalysis, shifting the focus from the mind of the individual child to the parent-child relationship as a clinical and theoretical unit. Nonverbal affective communication between mother and newborn constitutes a psychic and neural structure that profoundly affects brain maturation. The early relationship is a structuring psychic situation, capable of producing both psychological and neural effects. Imbasciati [41,42,43] redefined the concept of brain maturation, arguing that it depends on primary relational learning and that the parent-child relationship acts as a vehicle for mental transformation through automatic and unconscious affective messages.
3. Aim
Building on the Four-Domain Integrative Framework presented in Section 1.2, this paper aims to propose a reinterpretation of early attachment, describing it as both a clinical and relational phenomenon and an adaptive process inscribed in human evolutionary history. As empirically demonstrated, biological, environmental, and relational factors influence and shape individual differences, starting from intrauterine life. Our integrative approach systematically explicates how biological predispositions, neralechanisms, relational experiences, and cultural practices interact dynamically to shape attachment trajectories across the first thousand days of life.
To achieve this aim, we examine three interconnected areas of evidence:
(1) Characteristics of early attachment (Table 1): We identify seven core features of early bonding—proximity seeking, emotional attunement, intrauterine experiences, maternal holding, security patterns, brain plasticity, and maternal stress. Table 1 [8,27,39,44,45,46] describes how these characteristics perform evolutionary functions of protection and adaptation, generating individual differences in emotional, cognitive, and relational development. genetic/epigenetic mechanisms (polymorphisms in OXTR, 5-HTTLPR, DRD4, and glucocorticoid receptor methylation) that modulate environmental sensitivity [22,23,30,31,34,48]; neurodevelopmental processes (hemispheric specialization, HPA-axis maturation, synaptic plasticity in the first 1000 days) that instantiate attachment at the neural level [8,16,22,23,27,46]; relational experiences (maternal attunement, holding, affect co-regulation) that activate or suppress genetic and neural potentials [36,39,40,45]; cultural-evolutionary contexts (caregiving ecologies such as co-sleeping, alloparenting, autonomy promotion) that shape which attachment strategies are adaptive [16,19,20,33].
(2) Cross-cultural variations in caregiving (Table 2): We examine six distinct cultural contexts to demonstrate that secure attachment emerges through culturally specific pathways, differentially influencing motor development, sleep patterns, HPA-axis maturation, and social skills [16,19,26,33,47].
This analysis challenges ethnocentric interpretations and reveals how cultural practices function as scaffolds for gene expression and neural development.
(3) Genetic and epigenetic foundations (Table 3): We analyze five genetic/epigenetic systems that modulate environmental sensitivity according to the differential susceptibility model [21,22,23,34,48,49], demonstrating how biological variability represents an evolutionary hedging strategy that maintains population adaptability.
For each area, we apply the Four-Domain Framework to elucidate mechanistic interactions rather than merely cataloging parallel findings. We show how biological predispositions create windows of differential susceptibility during which neural systems aremost plastic and responsive to relational inputs, which are themselves embedded in cultural scaffolds that shape adaptive outcomes. This integrative synthesis carries significant implications for clinical practice, early intervention protocols, and transcultural approaches to perinatal mental health.
4. Cross-Cultural and Genetic Research
Cross-cultural research highlights that, while universal patterns of attachment exist, caregiving practices adapt to specific ecological and social contexts, determining different developmental trajectories. The Table 2 [16,19,26,33,47] summarizes some of the main alternatives observed in attachment development across cultures. These findings suggests that attachment, even if it is grounded in universal biological foundations, can be expressed through culture-specific pathways with meaningful implications for cognitive, emotional, and social development. Recognizing such variations helps to avoid ethnocentric interpretations and supports the development of clinical interventions protocols and preventive practices that take care more of cultural differences.
Recently, some studies have highlighted the crucial role of genetic foundations and gene–environment interactions in the development of the attachment. Specific genetic polymorphisms and epigenetic mechanisms help explain the reason for which individuals differ in environmental sensitivity and attachment trajectories. The Table 3 [21,22,23,34,48,49] summarizes the main findings.
These findings confirm that attachment is not only the outcome of relational experiences but quite develops from the dynamic interplay between biological inclinations and environmental contexts. Genetic variability represents an evolutionary strategy that maintains diversity and enables rapid adaptation to changing environments.
According to an integrated perspective model, attachment could be assumed as the complex outcome of multiple interacting dimensions rather than the product of a single factor.
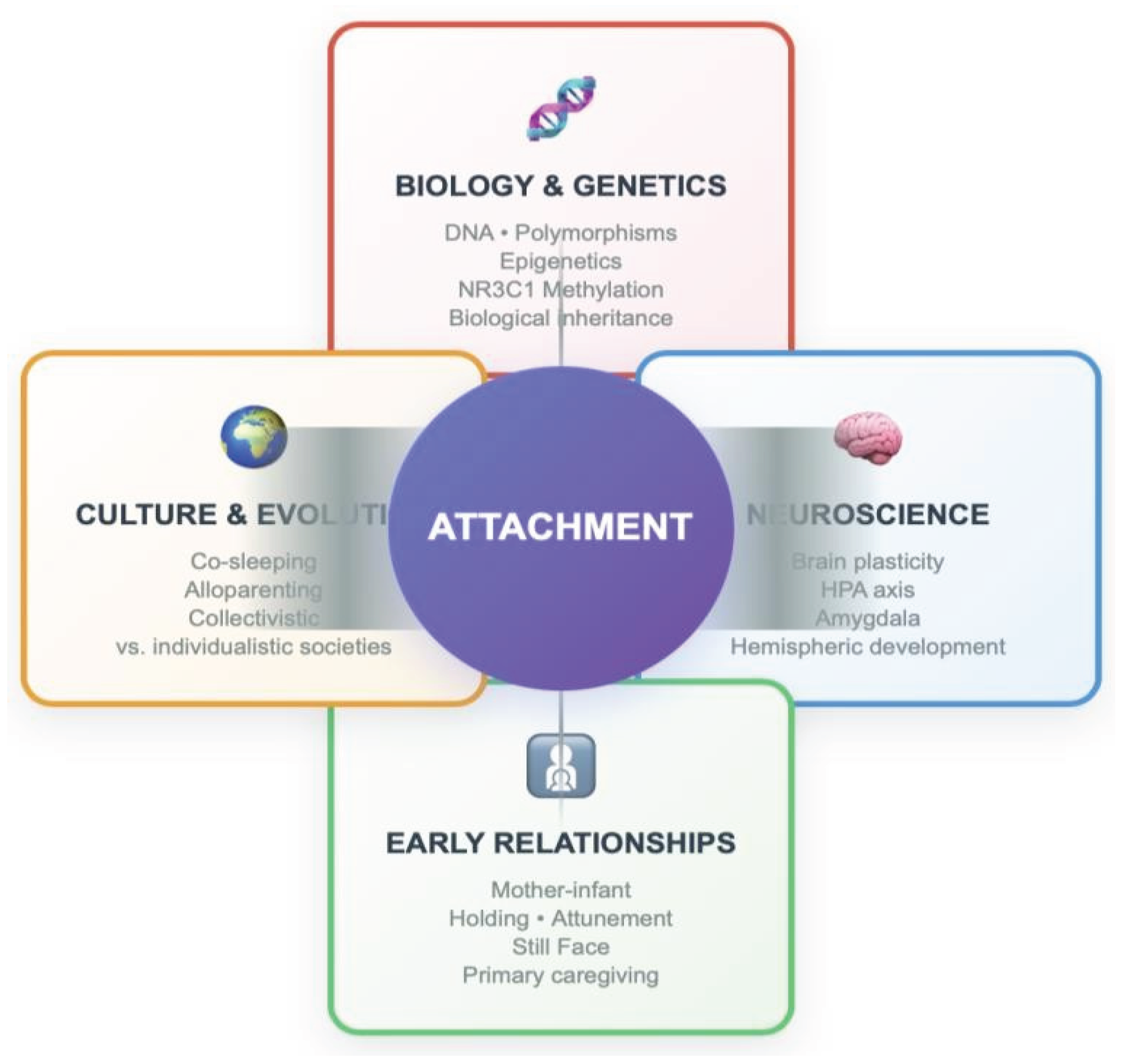
Contemporary research highlights the converging contributions of biological and genetic mechanisms (e.g., gene polymorphisms and epigenetic processes such as NR3C1 methylation), neuroscientific evidence (brain plasticity, HPA axis functioning, amygdala activation, hemispheric specialization), early relational experiences (mother–infant attunement, holding, and the Still Face paradigm) and cultural as well as evolutionary contexts (e.g., co-sleeping, alloparenting, and differences between collectivistic and individualistic societies). As summarized in Figure 2, attachment emerges from the dynamic integration of these factors, which together form developmental trajectories and individual differences in socio-relational-emotional functioning [20,23,40,50,51].
Figure 1.
Four-Domain Integrative Framework of Early Attachment. The Four-Domain Integrative Framework describes early attachment as the emergent result of dynamic, bidirectional interactions among Biology/Genetics, Neuroscience, Relational Experiences, and Cultural Contexts. Unlike traditional biopsychosocial models, this framework emphasizes (a) temporal cascades and differential susceptibility, (b) recursive causality across levels, and (c) evolutionary-adaptive plasticity. Genetic predispositions (e.g., 5-HTTLPR) shape periods of heightened neural plasticity; relational experiences such as maternal attunement modulate oxytocin and HPA-axis activity; cultural practices influence physiological regulation; and these domains mutually co-regulate each other across development.
Figure 1.
Four-Domain Integrative Framework of Early Attachment. The Four-Domain Integrative Framework describes early attachment as the emergent result of dynamic, bidirectional interactions among Biology/Genetics, Neuroscience, Relational Experiences, and Cultural Contexts. Unlike traditional biopsychosocial models, this framework emphasizes (a) temporal cascades and differential susceptibility, (b) recursive causality across levels, and (c) evolutionary-adaptive plasticity. Genetic predispositions (e.g., 5-HTTLPR) shape periods of heightened neural plasticity; relational experiences such as maternal attunement modulate oxytocin and HPA-axis activity; cultural practices influence physiological regulation; and these domains mutually co-regulate each other across development.
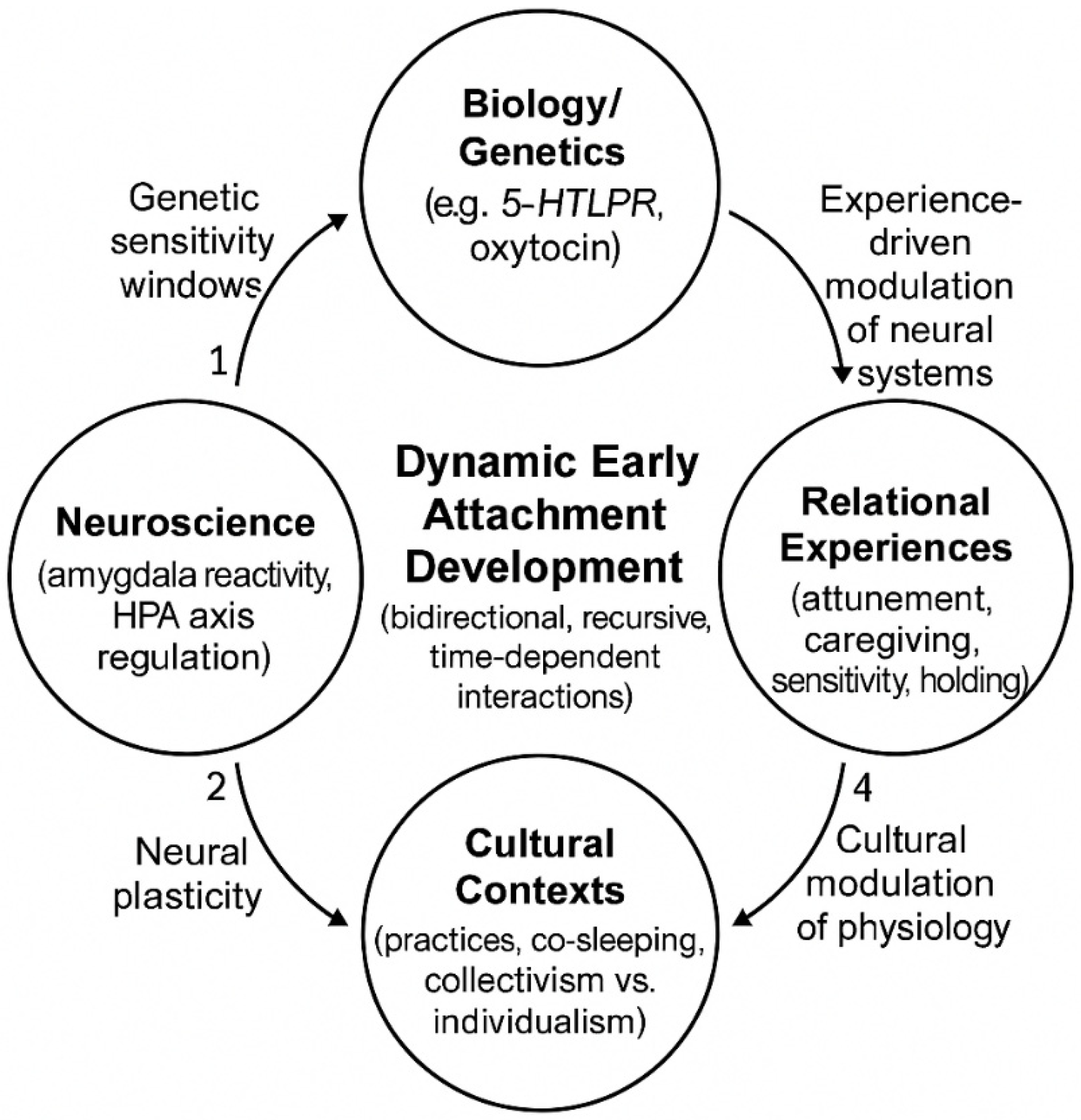
Figure 2.
Attachment as a complex result of the interactionbetween biological/genetic, neuroscientific, relational and cultural factors (adapted from Cassidy & Shaver, 2016; Keller, 2018; Meaney, 2010; Schore, 2019; Tronick, 2007). The model illustrates how Attachment emerges from the dynamic integration of four fundamental domains: Biology (genetic predispositions, polymorphic variants, and epigenetic modifications); Neuroscience (brain development, stress systems, and neural plasticity); Relationships (quality of early interactions and caregiving patterns); Culture (developmental practices, social contexts, and cross-cultural differences). Each individual develops their own attachment style through the complex and ongoing interaction of these factors.
Figure 2.
Attachment as a complex result of the interactionbetween biological/genetic, neuroscientific, relational and cultural factors (adapted from Cassidy & Shaver, 2016; Keller, 2018; Meaney, 2010; Schore, 2019; Tronick, 2007). The model illustrates how Attachment emerges from the dynamic integration of four fundamental domains: Biology (genetic predispositions, polymorphic variants, and epigenetic modifications); Neuroscience (brain development, stress systems, and neural plasticity); Relationships (quality of early interactions and caregiving patterns); Culture (developmental practices, social contexts, and cross-cultural differences). Each individual develops their own attachment style through the complex and ongoing interaction of these factors.
5. Clinical and Therapeutic Implications
Several factors can interfere with the quality of early bonding, including postpartum depression, traumatic experiences, relationship difficulties, and disadvantaged socioeconomic conditions [40]. Chronic maternal stress can alter epigenetic transmission and negatively affect the child’s neurobiological development. In these cases, early intervention by perinatal health professionals is essential to support the mother in her parenting role and promote a supportive relational environment.
Understanding early attachment has profound implications for psychotherapeutic Perinatal psychotherapy, for example, is an effective clinical tool for supporting the mother-child dyad, promoting secure attachment and healthy emotional regulation. Early intervention, even during pregnancy, can prevent the transgenerational transmission of trauma and promote the child’s harmonious development.
In the clinical setting, it is essential to recognize the signs of early relational distress—such as breastfeeding difficulties, sleep disturbances, irritability—and to intervene with sensitivity and competence [41]. The training of practitioners must include neuroscientific knowledge, empathic listening skills, and relational observation tools to promote a culture of affective care.
From this perspective, considering attachment as an adaptive survival function allows us to reinterpret early difficulties not only as indicators of risk, but as defensive strategies that are functional to the context. This guides therapeutic intervention toward recognizing and transforming these mechanisms into resources for growth and resilience.
Furthermore, the interdisciplinary approach suggests the need for an integrated network of psychologists, pediatricians, midwives, and psychiatrists, capable of offering comprehensive support for parenting and children’s mental health [54,55,56,57].
The data summarized in the tables are not only of theoretical and evolutionary value, but also offer relevant insights for clinical practice. In fact, from a clinical point of view, recognizing that early behaviors (physical contact, attunement, holding) transform into personality structures and styles of affective regulation allows the therapist to interpret symptoms not as mere deficits, but as adaptive trategies that made sense in a specific relational context. In therapy, this means valuing the patient’s implicit skills and accompanying them in the transformation of rigid strategies into more flexible resources.
In clinical practice, it is essential for the therapist to consider cultural models too and avoid ethnocentric interpretations (e.g., considering an attachment that is functional in another culture to be “insecure”), with the aim to develop a transcultural competence that recognizes the plurality of patterns felt as security and safe. In this sense, therapeutic process doesn’t consist of “normalising” the patient according to Western standards, but rather recognising and reinforcing attachment patterns that are adaptive in the context to which they belong.
In particular, we can underline that the evidences on genetic polymorphisms and epigenetic modifications open up clinical perspectives roughly in two directions. The first paves the way for psychoeducational interventions aimed at helping patients (and parents) to understand that vulnerability is not a definitive label, but rather a greater sensitivity to environmental stimuli. On the other hand, the second direction offer opportunities for targeted prevention, explicitly the awareness that, in genetically more susceptible individuals (“orchids”), early interventions and supportive environments have a particularly transformative effect. Clinical practice, therefore, is not limited to “curing” but becomes a laboratory of resilience that deeds the plasticity offered by the underlying predispositions.
6. Discussion
The Four-Domain Integrative Framework presented here advances beyond traditional models by explicating how mechanisms operate across levels simultaneously. Rather than viewing attachment as the sum of independent influences, our model demonstrates recursive causality: cultural practices modulate physiological systems, which shape neural development, which in turn influences how children engage with cultural practices. This perspective has practical implications: interventions targeting a single domain (e.g., psychoeducation alone) may be less effective than coordinated approaches that recognize how changes in one domain cascade through others. For example, supporting maternal sensitivity (relational domain) may be enhanced by addressing cultural beliefs about caregiving (cultural domain) while considering individual genetic sensitivity profiles (biological domain) and providing psychoeducation about neural plasticity (neuroscience domain).
As highlighted in the summary in Table 2, basic behaviors (seeking proximity, emotional attunement, intrauterine experiences) are not only immediate adaptive responses, but also matrices of individual differences that endure long life. It is interesting to note how elements that are apparently “survival” (e.g., physical contact and maternal protection) progressively become complex paradigms linked to trust, motivation, and exploration [58,59,60,61,62]. This last date confirms the connection between primary biological needs and long life term psychological trajectories, in particular it shows that the quality of the bond has a deep impact on individual processes of resilience, self-regulation, and psychopathological risk. As shown in Table 2, each characteristic of early attachment operates simultaneously across the four domains of the integrative framework. For example, seeking proximity is not only a relational behavior, but involves the activation of the oxytocin system (biology), modulates amygdala-prefrontal connectivity (neuroscience), responds to maternal sensitivity (relationships), and is calibrated according to cultural norms about appropriate interpersonal distance (culture), demonstrating the recursive causality of the model.
Cross-cultural evidences (Table 3) clearly shows that, although attachment is universally biological, the ways in which it is expressed and regulated vary according to ecological environments and cultural systems. This broadens clinical reflection: “secure” attachment in Western contexts does not necessarily coincide with the same indicators in collectivist cultures or traditional societies. In this sense, it is important that psychotherapy research takes these cross-cultural peculiarities into account in order to adopt a model that is also ethnocentric [26,33,63]. In reality, caregiving practices (cosleeping, alloparenting, reduced social stimulation) show that attachment security can emerge in different ways, all of which are functional to adaptation. Table 3 illustrates how cultural caregiving practices function as ‘scaffolds’ that simultaneously modulate genetic expression (e.g., OXTR variants), neural development (e.g., cortisol rhythms in co-sleeping), relational patterns (e.g., alloparenting in ! Kung) and adaptive outcomes, demonstrating that secure attachment emerges through culturally specific trajectories rather than following a single universal model.
Individual differences emerge in a complex dynamic and cannot be reduced solely to the environment or relationships, as highlighted in the summary of genetic polymorphisms and genetic mechanisms (Table 4). An integtated model emerges in which biology, environment, and culture intertwine and produce different but functional results. Table 4 demonstrates how genetic and epigenetic mechanisms do not operate in a deterministic manner, but create windows of differential susceptibility during which neural systems are most plastic and responsive to relational inputs (e.g., NR3C1 and maternal care), which are in turn embedded in cultural scaffolds (e.g., OXTR variants and collectivist vs. individualistic contexts), exemplifying the cascades of development from the molecular to the behavioral level described in the integrative framework.
In this sense, the variability observed is not an “error” but rather an evolutionary strategy aimed at ensuring flexibility and adaptation. The metaphor of “orchid children” and “dandelion children” becomes particularly powerful in communicating, even at the clinical level, that sensitivity and vulnerability are not deficits but potentials that depend on context [21].
The data show that:
1. Universality and diversity coexist: although attachment is a universal mechanism, its developmental trajectories differ depending on culture, environment, and genetic predispositions.
2. Prevention and early intervention: recognizing attachment as an adaptive function implies that even initial difficulties should be interpreted as signs of adaptation to an adverse context. This has clinical implications: early intervention must aim to transform defensive mechanisms into resources for resilience.
3. Transcultural clinical practice: the data highlight the necessity for therapeutic practices that rate forms of security other than those traditionally accepted in the West.
4. Bio-psycho-social integration: the complexity of early attachment can only be understood through an integrated approach that combines neuroscience, psychoanalysis, and transcultural evidence.
5. Evolutionary perspective: attachment represents a psychological construct and an evolutionary survival strategy.
The individual’s first emotional experiences are described in the literature on the subject as preparation for social and relational life. From an evolutionary perspective, these experiences constitute adaptive mechanisms which, through the modulation of brain plasticity and emotional regulation [27,46,64], generate individual differences in resilience, motivation, and personality expression.
Selective processes aimed at adaptation to different environmental and social contexts shape the variability observed in the temperaments and developmental trajectories of fetuses and children. Considering attachment as an adaptive function of survival, as well as a psychological construct, allows us to reinterpret the vulnerabilities associated with early bonding dysfunctions as functional mechanisms [64,65,66]: this has significant implications for prevention and clinical practice.
Early emotional bonding is a multidimensional construct that profoundly influences an individual’s mental health, cognitive development, and relational capacity. The integration of neuroscience and psychoanalysis provides a useful theoretical and clinical perspective for understanding the genesis of the mind and promoting preventive therapeutic interventions in the first thousand days of a child’s life.
Contemporary psychotherapy must recognize and value the relational dimension as the foundation of subjectivity. A culture of early emotional care is an investment in future generations’ mental health.
7. Conclusions
This bio-psycho-socio-evolutionary clinical model, integrating evolutionary, transcultural, and genetic perspectives, provides a framework for psychotherapists to facilitate adaptive processes across individuals.
Author Contributions
Marina Cafaro and Laura Ambrosecchia conceptualized and wrote the sections related to the psychoanalytic framework of early emotional bonds and the clinical implications for parent–infant dynamics. Valeria Cioffi and Enrica Tortora developed the cross-cultural analysis and contributed to the integrative synthesis connecting psychoanalytic, neuroscientific, and anthropological perspectives. Raffaele Sperandeo contributed to the sections on developmental neuroscience, neurobiological mechanisms, and epigenetic processes involved in early attachment. Daniela Cantone provided overall scientific supervision, contributed to the conceptual integration across domains, and ensured the methodological and theoretical coherence of the manuscript. All authors participated in the critical revision of the final manuscript and approved it for publication.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki. Ethical review and approval were not applicable, because it did not involve humans or animals.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Di Leva, A. (2023). Being in the world “between” psychotherapy and neuroscience. Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 5(1), 21–33. [CrossRef]
- Freud, S. (1926/1959). Inhibitions, Symptoms and Anxiety (A. Strachey, Trans.). New York, NY: Norton. (Original work published 1926).
- Piontelli A., From Fetus to Child, Tavistock/Routledge, London, 1992.
- Piontelli, A. (1987). Infant observation from before birth. The International Journal of Psycho-analysis, 68, 453-463.
- Mychasiuk, R., Harker, A., Ilnytskyy, S., & Gibb, R. (2013). Paternal stress prior to conception alters DNA methylation and behaviour of developing rat offspring. Neuroscience, 241, 100-105.
- Gainotti G. (2006). Unconscious emotional memories and the right hemisphere. In Mancia M. (a cura di), Psychoanalysis e Neuroscience, Milano, Springer.
- Phelps, E. A. (2004). Human emotion and memory Interactions of the amygdala and hippocampal complex. Current Opinion in Neurobiology, 14, 198-202. [CrossRef]
- Schore, A. N. (2000). Attachment and the regulation of the right brain. Attachment & Human Development, 2(1), 23–47. [CrossRef]
- Schore, A. N. (2001). Affect Regulation and the Repair of the Self. New York, NY: Norton.
- Schore, A. N. (2003). Affect Regulation and the Origin of the Self: The Neurobiology of Emotional Development. New York, NY: Norton.
- Schore, A. N. (2017). The development of the right brain across the life span: What’s love got to do with it? Journal of Research in Medicine and Nursing, 1(1), 23–39. [Conference paper, Oslo].
- Negri, R. (2018). The newborn in the intensive care unit: A neuropsychoanalytic prevention model.
- Walsh, N. D., Dalgleish, T., Dunn, V. J., Abbott, R., St Clair, M. C., Owens, M., … Goodyer, I. M. (2012). 5 HTTLPR–environment interplay and its effects on neural reactivity in adolescents. NeuroImage, 63(3), 1670 1680. [CrossRef]
- Landoni, M., Missaglia, S., Tavian, D., Ionio, C., & Di Blasio, P. (2022). Influence of 5-HTTLPR polymorphism on postpartum depressive and posttraumatic symptoms. Psychiatric Genetics, 32(1), 9-14.
- Pluess, M., & Belsky, J. (2012). Vantage sensitivity: Individual differences in response to positive experiences. Psychological Bulletin. Advance online publication. [CrossRef]
- McKenna, J. J., & McDade, T. (2005). Why babies should never sleep alone: A review of co-sleeping, breastfeeding and SIDS. Paediatric Respiratory Reviews, 6(2), 134–152. [CrossRef]
- Markova, G., & Siposová, B. (2019). The role of oxytocin in early mother infant interactions: Variations in maternal affect attunement. Infant Behavior and Development, 55, 58 68. [CrossRef]
- Feldman R. The Neurobiology of Human Attachments. Trends Cogn Sci. 2017 Feb;21(2):80-99. Epub 2016 Dec 30. [CrossRef] [PubMed]
- LeVine, R. A., LeVine, S., Dixon, S., Richman, A., Leiderman, P. H., Keefer, C. H., & Brazelton, T. B. (1994). Child care and culture: Lessons from Africa. New York, NY: Cambridge University Press.
- Keller, H. (2018). Universality claim of attachment theory: Children’s socioemotional development across cultures. Proceedings of the National Academy of Sciences, 115(45), 11414 11419. [CrossRef]
- Belsky, J., & Pluess, M. (2009). The nature (and nurture?) of plasticity in early human development. Perspectives on Psychological Science, 4(4), 345–351. [CrossRef]
- Weaver, I. C., Cervoni, N., Champagne, F. A., D’Alessio, A. C., Sharma, S., Seckl, J. R., ... & Meaney, M. J. (2004). Epigenetic programming by maternal behavior. Nature Neuroscience, 7(8), 847–854. [CrossRef]
- Meaney, M. J. (2010). Epigenetics and the biological definition of gene × environment interactions. Child Development, 81(1), 41–79. [CrossRef] [PubMed]
- Provenzi, L., Grumi, S., Altieri, L., Bensi, G., Biasucci, G., Bertazzoli, E., ... & Montirosso, R. (2022). Prenatal maternal stress during the COVID-19 pandemic and infant regulatory capacity at 3 months: The role of epigenetic signatures at birth. Development and Psychopathology, 34(5), 1972–1984. [CrossRef]
- Oberlander, T. F., Weinberg, J., Papsdorf, M., Grunau, R., Misri, S., & Devlin, A. M. (2008). Prenatal exposure to maternal depression, neonatal methylation of NR3C1 and infant cortisol stress responses. Epigenetics, 3(2), 97–106. [CrossRef]
- Riem, M. M., Lotz, A. M., Horstman, L. I., Cima, M., Verhees, M. W., Alyousefi-van Dijk, K., ... & Bakermans- Kranenburg, M. J. (2021). A soft baby carrier intervention enhances amygdala responses to infant crying in fathers: a randomized controlled trial. Psychoneuroendocrinology, 132, 105380.
- Kim, P., Strathearn, L., & Swain, J. E. (2016). The maternal brain and its plasticity in humans. Hormones and Behavior, 77, 113–123. [CrossRef]
- Kohlhoff, J., Karlov, L., Dadds, M., Barnett, B., Silove, D., & Eapen, V. (2022). The contributions of maternal oxytocin and maternal sensitivity to infant attachment security. Attachment & Human Development, 24(4), 525-540.
- Kohlhoff, J., Karlov, L., Dadds, M., Barnett, B., Silove, D., Mendoza Diaz, A., & Eapen, V. (2024). Preschool Behavioral Problems: Links with Maternal Oxytocin and Caregiving Sensitivity in the Postnatal Period, and Concurrent Maternal Psychopathology and Attachment State-of-Mind. Child psychiatry and human development, 55(6), 1736–1746. [CrossRef]
- Tost, H., Kolachana, B., Hakimi, S., Lemaitre, H., Verchinski, B. A., Mattay, V. S., ... & Meyer-Lindenberg, A. (2010). A common allele in the oxytocin receptor gene (OXTR) impacts prosocial temperament and human hypothalamic-limbic structure and function. Proceedings of the National Academy of Sciences, 107(31), 13936-13941. [CrossRef]
- Li, J., Zhao, Y., Li, R., Broster, L. S., Zhou, C., & Yang, S. Li, J., Zhao, Y., Li, R., Broster, L. S., Zhou, C., & Yang, (2015). Association of oxytocin receptor gene (OXTR) rs53576 polymorphism with sociality: a meta-analysis. PLOS One, 10(6), e0131820. [CrossRef]
- Bakermans-Kranenburg, M. J., & van IJzendoorn, M. H. (2008). Oxytocin receptor (OXTR) and serotonin transporter (5-HTTLPR) genes associated with observed parenting. Social Cognitive and Affective Neuroscience, 3(2), 128–134. [CrossRef]
- Konner, M. (2005). Hunter-gatherer infancy and childhood: The !Kung and others. In B. S. Hewlett & M. E. Lamb (Eds.), Hunter-gatherer childhoods: Evolutionary, developmental, and cultural perspectives (pp. 19–64). New Brunswick, NJ: Aldine Transaction.
- Caspi, A., Sugden, K., Moffitt, T. E., Taylor, A., Craig, I. W., Harrington, H., ... & Poulton, R. (2003). Influence of life stress on depression: Moderation by a polymorphism in the 5-HTT gene. Science, 301(5631), 386–389. [CrossRef]
- Di Sarno A. D., Costa, V., Di Gennaro, R., Di Leva, G., Fabbricino, I., Iennaco, D., … Mosca, L. L. (2019). At the roots of the sense of self: Proposals for a study on the emergence of body awareness in early childhood. Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 1(2), 37–46. [CrossRef]
- Winnicott, D. W. (2016). The collected works of DW Winnicott (Vol. 12). Oxford University Press.
- Bion, W. R. (1962). Learning from Experience. London: Heinemann.
- Francesetti, G. (2024). The phenomenal field: the origin of the self and the world. Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 6(1), 1–5. (Original work published March 1, 2024). [CrossRef]
- Bowlby, J. (1969). Attachment and Loss: Vol. 1. Attachment. New York, NY: Basic Books.
- Tronick, E. Z. (2007). The Neurobehavioral and Social-Emotional Development of Infants and Children. New York, NY: Norton.
- Imbasciati, A. (2001). Psicoanalisi e Neuroscienze. FrancoAngeli.
- Imbasciati, A. (2014). Our Future Mind: Epochal Developments of Perinatal Clinical Psychology. International Journal of Child Health and Nutrition, 3(3), 148–155. [CrossRef]
- Cena, L., & Imbasciati, A. (2010). PERINATAL CLINICAL PSYCHOLOGY: PARENT-CHILD INTERACTIONS IN THE PRIMARY “CARE”. Journal of Prenatal and Perinatal Psychology and Health, 25(2/2010), 67-88.
- Frasch, M. G., Lobmaier, S., Stampalija, T., Pallarés, M. E., Pastor, V., Brocco, M., … Antonelli, M. (2018). Non- invasive biomarkers of fetal brain development reflecting prenatal stress: An integrative multi-scale multi-species perspective. Developmental Psychobiology, 60(7), 783–801. [CrossRef]
- Hofer, M. A. (2005). The psychobiology of early attachment. Clinical Neuroscience Research, 4(5–6), 291–300. [CrossRef]
- Barba-Müller, E., Craddock, S., Carmona, S., Hoekzema, E., & Desco, M. (2018). Brain plasticity in pregnancy and the postpartum period. Neuroscience & Biobehavioral Reviews, 92, 179–189. [CrossRef]
- Jin, M. K., Jacobvitz, D., Hazen, N. L., & Jung, S. H. (2012). Maternal sensitivity and infant attachment security in Korea: Cross-cultural validation of the Strange Situation. Attachment & Human Development, 14(1), 33–44. [CrossRef]
- Bakermans-Kranenburg, M. J., & Van IJzendoorn, M. H. (2006). Gene–environment interaction of the dopamine D4 receptor (DRD4) and observed maternal insensitivity predicting externalizing behavior in preschoolers. Developmental Psychobiology, 48(5), 406–409. [CrossRef]
- Gluckman, P. D., & Hanson, M. A. (2004). Living with the past: Evolution, development, and patterns of disease. Science, 305(5691), 1733–1736. [CrossRef]
- Cassidy, J., & Shaver, P. R. (Eds.). (2016). Handbook of attachment: Theory, research, and clinical applications (3rd ed.). New York: Guilford Press.
- Schore, A. N. (2019). Right brain psychotherapy. New York: Norton.
- Zampi, R. (2023). Missing Twin Syndrome: life after perinatal mourning. Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 5(1), 88–96. [CrossRef]
- Guerriera, C., & Cantone, D. (2019). The current clinic in psychoanalysis of childhood and adolescence: questions and heuristic hypotheses. Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 1(2), 69–75. [CrossRef]
- Di Leva, G., Nascivera, N., & Di Sarno, A. D. (2022). Parenting practices of students’ parents in a high school in the province of Salerno. An exploratory research. Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 4(1), 77–84. [CrossRef]
- Marino, F., Gautiero, G., Glorioso, A., Lauro, G., Zampi, R., & Perrone, M. (2024). A Gestalt approach at couples psychotherapy: From attachment theory to emotion-focused therapy (EFT). Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 6(2), 29–39. [CrossRef]
- Negri (1996), Il neonato in terapia intensiva. Un modelo neuropsicoanalitico di prevenzione, Raffaello Cortina Milano.
- Fornari (1977). Codice materno e disturbi della femminilità. Cerutti M., Ginecologia psicosomatica e profilassi ostetrica, Padova.
- Siegel, D. J. (2012). The Developing Mind: How Relationships and the Brain Interact to Shape Who We Are (2nd ed.). New York, NY: Guilford Press.
- Stern, D. N. (1985). The Interpersonal World of the Infant. New York, NY: Basic Books.
- Stern, D. N., Sander, L. W., Nahum, J. P., Harrison, A. M., Lyons-Ruth, K., Morgan, A. C., … Tronick, E. Z. (1998). Non-interpretive mechanisms in psychoanalytic therapy: The “something more” than interpretation. International Journal of Psychoanalysis, 79(5), 903–921.
- Trevarthen, C., & Aitken, K. J. (2001). Infant intersubjectivity: Research, theory and clinical applications. Journal of Child Psychology and Psychiatry, 42(1), 3–48.
- Trevarthen, C. (1979). Communication and cooperation in early infancy: A description of primary intersubjectivity. In M. Bullowa (Ed.), Before Speech: The Beginning of Human Communication (pp. 321–347). Cambridge: Cambridge University Press.
- Mazzara, M. (2020). Feed me Stories: Taking care through cultural mediums. Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 2(1), 54–61. [CrossRef]
- Capodilupo, A. (2022). Woman’s sentimental attachment style and depressive states in pregnancy: observational study on pregnant women in childbirth preparation courses. Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 4(2), 31–49. [CrossRef]
- Guerriera, C., & Cantone, D. (2019). Psychopathology and Psychoanalysis between knowledge, diagnosis and treatment. Phenomena Journal—International Journal of Psychopathology, Neuroscience and Psychotherapy, 1(1), 37–43. [CrossRef]
- Van IJzendoorn, M. H., & Bakermans-Kranenburg, M. J. (2008). The distribution of adult attachment representations in clinical groups: A meta-analytic search for patterns of attachment in 105 AAI studies.
- Mancia, M. (2007). Psychoanalysis and Neuroscience. Milan: Springer.

Table 1.
Terminological Framework.
| TERM | DEFINITION | KEY CHARACTERISTICS / DISTINCTIONS |
|---|---|---|
| ATTACHMENT | The enduring emotional bond between an infant and a primary caregiver. | Behaviors include proximity seeking, separation protest, and use of the caregiver as a secure base. Patterns may be secure or insecure. Associated with specific neurobiological correlates. |
| BONDING | The process through which the parent (particularly the mother) develops an emotional bond with the newborn, typically in the first hours or days. | Emotional movement from parent to child. Includes hormonal, behavioral, and psychological components. Differs from attachment, which concerns the infant’s bond to the caregiver. |
| EARLY EMOTIONAL BOND | An umbrella term encompassing both attachment and bonding. | Highlights the bidirectional, co-constructed nature of the primary relationship. |
| PRIMARY RELATIONSHIP | The set of mother–infant interactions and interpersonal dynamics, with references to psychoanalytic constructs. | Includes holding, containment, and alpha function. |
| ATTUNEMENT | The caregiver’s ability to perceive, interpret, and respond contingently and appropriately to the infant’s emotional states and needs. | Prerequisite for secure attachment. Essential for the development of emotional regulation. |
| USAGE IN THE TEXT | — | The terms “attachment” and “early emotional bond/bonding” are sometimes used interchangeably when referring to the overall phenomenon addressed by the integrative model. |
Table 2.
Individual differences generated by early attachment.
| CHARACTERISTIC OF EARLY ATTACHMENT | EVOLUTIONARY FUNCTION | INDIVIDUAL DIFFERENCES GENERATED | FOUR-DOMAIN INTEGRATION | REFERENCES |
|---|---|---|---|---|
| PROXIMITY SEEKING TO THE CAREGIVER | Ensuring protection and survival of the infant in threatening contexts | Levels of secure vs. insecure attachment; predisposition to relational trust | Biology: Oxytocin system activation; Neuroscience: Amygdala-prefrontal connectivity; Relationships: Maternal responsiveness; Culture: Acceptable proximity distance varies | Bowlby, 1969 |
| MOTHER-INFANT EMOTIONAL ATTUNEMENT | Promoting affect regulation and social cohesion | Differential capacities for self-regulation, resilience, and stress management | Biology: Serotonin transporter variants modulate sensitivity; Neuroscience: Right hemisphere limbic circuits; Relationships: Quality of interactive synchrony; Culture: Display rules for emotions | Schore, 2000 |
| EARLY INTRAUTERINE EXPERIENCES (MATERNAL VOICE, HEARTBEAT, HORMONES) | Preparing the fetus for the postnatal environment and anticipating social interaction | Early temperamental differences (reactivity, sensitivity to stimuli, neurocognitive plasticity) | Biology: Epigenetic programming via NR3C1 methylation; Neuroscience: Prenatal neural connectivity patterns; Relationships: Maternal stress transmission; Culture: Pregnancy beliefs affect maternal physiology | Frasch et al., 2018 |
| MATERNAL HOLDING AND CONTAINMENT | Creating a safe environment for the development of the Self | Differences in the development of basic trust and exploratory motivation | Biology: HPA axis calibration; Neuroscience: Insula and somatosensory cortex development; Relationships: Physical contact frequency; Culture: Bodily contact norms (high-contact vs. low-contact cultures) | Hofer, 2005 |
| SECURE VS. INSECURE ATTACHMENT | Fostering cooperation and social adaptation | Divergent trajectories in personality, motivation, and relational capacities | Biology: DRD4 variants influence exploration; Neuroscience: Prefrontal-limbic integration patterns; Relationships: Caregiver sensitivity and availability; Culture: Security defined differently (autonomy vs. interdependence) | Bowlby, 1969; Schore, 2000 |
| BRAIN PLASTICITY IN THE FIRST 1000 DAYS | Optimizing adaptation to the environment | Individual variability in memory, attention, and emotional regulation | Biology: Synaptic pruning guided by experience; Neuroscience: Critical period plasticity mechanisms; Relationships: Relational experiences shape neural architecture; Culture: Culturally-valued skills receive more neural resources | Kim et al., 2016; Barba-Müller et al., 2018 |
| EARLY MATERNAL TRAUMA OR STRESS | Signaling adverse environmental conditions and "preparing" for difficult contexts | Differential vulnerability to anxiety disorders, depression, and affect dysregulation | Biology: Glucocorticoid receptor methylation; Neuroscience: Altered amygdala reactivity and HPA axis; Relationships: Intergenerational transmission patterns; Culture: Social support networks buffer or amplify effects | Schore, 2000; Frasch et al., 2018 |
Table 3.
Cross-cultural variations in attachment development.
| CULTURAL CONTEXT | CAREGIVING PRACTICES | ATTACHMENT / DEVELOPMENTAL PATTERNS | FOUR-DOMAIN INTEGRATION | REFERENCES |
|---|---|---|---|---|
| !KUNG (HUNTER-GATHERERS, BOTSWANA) | - Constant physical contact - Immediate responsiveness - Extended nursing until 3-4 years - Extensive alloparenting | - Rapid motor development - Secure attachment through continuous proximity - Low infant crying | Biology: Sustained oxytocin levels from prolonged contact; Neuroscience: Enhanced vestibular-motor integration; Relationships: Multiple attachment figures (alloparenting); Culture: Egalitarian social structure supports shared caregiving | Konner, 2005 |
| GUSII (KENYA, RURAL SOCIETY) | - Limited face-to-face interaction - Focus on physical protection and nutrition | - Secure attachment despite different interaction styles | Biology: Attachment security achieved via tactile rather than visual modality; Neuroscience: Different sensory pathway dominance; Relationships: Instrumental care as attachment communication; Culture: Agricultural demands shape caregiving priorities | LeVine et al., 1994 |
| COLLECTIVISTIC SOCIETIES (E.G., EAST ASIA) | - Emphasis on interdependence and emotional control - Group-oriented socialization | - "Culturally modulated secure attachment" - Group social competencies before individual ones | Biology: Same oxytocin system, different phenotypic expression; Neuroscience: Enhanced social brain network connectivity; Relationships: Extended family involvement; Culture: Interdependent self-construal shapes secure base function | Jin et al., 2012 |
| INDIVIDUALISTIC SOCIETIES (E.G., WESTERN CULTURES) | - Promotion of autonomy and early self-regulation - Valorization of independent exploration | - Attachment patterns oriented toward separation and individuation | Biology: Genetic variants (DRD4-7R) may confer advantage in exploration-oriented contexts; Neuroscience: Prefrontal systems for self-regulation emphasized; Relationships: Dyadic mother-infant focus; Culture: Independence valued as developmental milestone | Jin et al., 2012 |
| CO-SLEEPING PRACTICES | - Shared mother-infant sleep | - Different REM sleep development - Variations in HPA axis maturation - Differences in nocturnal cortisol levels | Biology: Synchronized circadian rhythms; Neuroscience: Modified cortisol rhythms affect HPA development; Relationships: Continuous nighttime proximity; Culture: Co-sleeping as normative practice maintains physiological synchrony | McKenna & McDade, 2005 |
| PROLONGED CARRYING PRACTICES | - Constant infant carrying | - Accelerated vestibular development - Enhanced spatial coordination - Different amygdala activation in response to movement | Biology: Increased oxytocin release in both parent and infant; Neuroscience: Enhanced vestibular-cerebellar maturation; Relationships: Continuous sensorimotor interaction; Culture: Babywearing as culturally-specific scaffold for development | Riem et al., 2021 |
Table 4.
Genetic foundations and gene–environment interactions in attachment.
| SYSTEM / GENE | KEY VARIANTS | EFFECTS ON ATTACHMENT / DEVELOPMENT | FOUR-DOMAIN INTEGRATION | REFERENCES |
|---|---|---|---|---|
| OXYTOCIN RECEPTOR (OXTR) | rs53576 (A/G) | G-allele: ↑ social sensitivity, secure attachment, better stress regulation; A-allele: ↓ social sensitivity, ↑ vulnerability to early trauma; Strong gene × environment effects (caregiving quality) | Biology: Genetic variants create differential susceptibility windows; Neuroscience: Modulates amygdala-prefrontal coupling; Relationships: Effect depends on caregiving quality; Culture: Phenotypic expression varies (G-allele more advantageous in collectivist contexts) | Bakermans-Kranenburg & van IJzendoorn, 2006; Tost et al., 2010; Li, 2015 |
| SEROTONIN TRANSPORTER (5-HTTLPR) | Short (s) vs. Long (l) allele | s-allele: high environmental sensitivity; secure in positive contexts, vulnerable in adverse ones (differential susceptibility model); l-allele: low sensitivity; stable development regardless of environment | Biology: "Orchid" (s-allele) vs. "dandelion" (l-allele) strategies; Neuroscience: s-allele linked to increased amygdala reactivity; Relationships: s-carriers benefit more from high-quality caregiving; Culture: Prevalence varies across populations, suggesting adaptive polymorphism | Caspi et al., 2003 |
| DOPAMINE RECEPTOR (DRD4) | 7-repeat variant | Associated with novelty seeking; Modulates sensitivity to parenting quality; Interacts with disciplinary style → predicts behavioral problems | Biology: DRD4-7R confers exploration advantage in novel environments; Neuroscience: Affects reward circuitry and executive function; Relationships: Interaction with parental warmth vs. harshness; Culture: 7R frequency higher in migratory populations, suggesting selection for exploration | Bakermans-Kranenburg & Van IJzendoorn, 2006 |
| EPIGENETICS (NR3C1) | Glucocorticoid receptor methylation | Maternal stress alters fetal methylation patterns; Cross-generational transmission; Shapes HPA-axis reactivity lifelong | Biology: Epigenetic "memory" of relational context; Neuroscience: Programs stress system sensitivity; Relationships: Quality of maternal care modulates methylation; Culture: Cultural stressors (e.g., discrimination, poverty) transmitted epigenetically | Weaver et al., 2004; Meaney, 2010 |
| FETAL PROGRAMMING HYPOTHESIS | Prenatal stress vs. protective environment | Stressful intrauterine conditions: ↑ reactivity, survival-oriented metabolism; Supportive conditions: ↑ regulation, growth, learning, long-term strategy | Biology: Predictive adaptive response to expected postnatal environment; Neuroscience: Calibrates stress systems prenatally; Relationships: Maternal-fetal physiological communication; Culture: Societal conditions (war, famine) produce cohort effects | Gluckman & Hanson, 2004 |
| DIFFERENTIAL SUSCEPTIBILITY MODEL | "Orchid" vs. "Dandelion" children | Orchid: carriers of sensitivity alleles (5-HTTLPR-s, DRD4-7R); thrive in good contexts, vulnerable in bad ones; Dandelion: resilient/stable across environments, less plasticity | Biology: Evolutionary hedging strategy maintains population diversity; Neuroscience: Susceptibility alleles linked to greater neural plasticity; Relationships: Orchids require high-quality caregiving; Culture: Different ecologies favor different strategies (stable vs. variable environments) | Belsky & Pluess, 2009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



