Submitted:
12 February 2026
Posted:
12 February 2026
You are already at the latest version
Abstract
Many phenomena in resistance training are often attributed to intrinsic changes within the muscles targeted for hypertrophy. Examples include rapid “newbie gains” among novices, individual differences in training outcomes, and blunted anabolic responses to protein intake in experienced lifters. These are typically interpreted as reflecting diminishing responsiveness of muscles to repeated training stimuli. This article explores an alternative framework predicated on the fact that both performance and adaptation are seldom, if ever, limited only by the target muscle but also by supporting factors, including tendons, posture-specific musculature, neural coordination, skill, etc. Because these constraints often adapt at different rates than the target muscle, disparities in adaptation can create apparent stagnation even when muscle growth potential remains. This framework explains why alternating complementary exercises can sustain progress, why trainees respond differently to the same program, and why modality comparisons often yield null or mixed results. Practically, it underscores the importance of strategic variation, complementary exercise sequencing, individualized programming, and management of facilitating factors such as nutrition and recovery.
Keywords:
resistance training
; hypertrophy
; adaptive resistance
; individual differences
; progressive overload
; periodization
; fatigue management
1. Introduction
Conventional wisdom in resistance training holds
that novices experience rapid “newbie gains,” followed by progressive
stagnation as muscles develop intrinsic changes rendering repeated training
stimuli progressively less effective. Similarly, individual differences in
training outcomes, inconsistencies across training modes and blunted anabolic
responses to dietary protein in experienced lifters are often ascribed to the
intrinsic properties of the target muscle. While such mechanisms are possible
and potentially important, they are usually inferred from the very outcomes
that they are invoked to explain, rather than directly demonstrated. For
example, Coffey et al. (2006) compared signalling responses in the quadriceps
following eight sets of five maximal leg extensions in experienced versus
novice participants. The reduced response observed in experienced lifters has
often been taken as evidence of diminished plasticity within the muscle itself.
Yet, if experienced trainees were limited in how effectively they could use
this exercise to stimulate the quadriceps, then the study measured a reduced stimulus,
not reduced responsiveness. This would be like concluding from a dim
room that a lightbulb had weakened, when in fact the dimmer switch had simply
reached its maximum setting. From this perspective, alternative explanations
deserve consideration—especially if they imply different training strategies.
The present article proposes that it is not necessary to assume changing
resistance to overload inherent within the muscle to explain adaptive
resistance. Because traditional training lore shapes the assumptions that guide
research design and interpretation, a strong prior commitment to within-muscle
mechanisms may cause us to overlook more parsimonious accounts based on
familiar training principles.
Nowhere is this clearer than in the case of
“adaptive resistance”: the observation that adaptations induced by training a
certain way or the use of a particular exercise incrementally vary over time
(Israetel, Hoffman, & Smith, 2015). When training is changed – such as by
shifting to a different exercise, range of repetitions or number of working
sets – rates of improvements in the target muscle will then return to a higher
level, at least for a while, until adaptive resistance begins to rear its ugly
head again. Additionally, if the trainee then returns to the original way of
training following this interlude, improvements may again be larger for some
time (Smith, 2022). Both the tendency of diminishing returns to the same
training stimulus is well documented (Lopez et al., 2020) and its remedy
through training variation have been discovered in various times and places.
For example, Schwarzeneggar (1999. p. 188):
“…if you always put the same kind of stress on the
body, in the same way, it gets used to this, and even very intense training
will yield less response than you expected. You can shock it by training with
more weight than usual; doing more reps and/or sets’ speeding up your training…
unfamiliar exercises, doing your exercises in an unfamiliar order…Change by
itself tends to shock the body, even if the unfamiliar workout is no more
demanding than the one you are used to. But you’ll get to a point where you’ll
find it difficult to make additional progress without shocking your muscles...”
“Shocking” the muscle and other metaphors such as
“muscle confusion” (regularly changing training regimes) and “the instinctive
principle” (altering intensity, exercises order, and other variables based on
variation in internal cues) popularized by Wieder (1989) and others abound that
share the common theme of introducing variation to break through training
plateaus. Notice as well that another commonality is the assumption that it is
changes within the muscle targeted by the training that causes the adaptive
resistance effect. However, such interpretations may be overlooking another
critical factor equally familiar to anyone with experience in resistance
training. Namely, the role of constraints, such as regions and fibres of
the same muscles less targeted by the same exercise, tendons, posture-specific
musculature, neural coordination, and so on, which interact with target muscles
and adapt at different rates. This as an important aspect of training can be
made obvious when we observe strength adaptations induced by training with one
exercise not fully generalizing to others even if dependant on the same muscle,
as unique constraining factors can limit the expression of muscle capacity
(Buckner et al., 2019). In other words, because performance and
cumulative stimulus are determined not only by the target muscle but also by
its interactions with these constraints, disparities in their dynamic responses
over time may provide more comprehensive explanation of adaptive resistance.
Viewed in this light, we will see how many widely
observed phenomena can be reinterpreted. Rapid early gains in novices may
reflect relatively unconstrained pathways rather than anything about the
targeted muscles being somehow “primed for growth”. Observed individual
differences in responsiveness to training may emerge from variation in
constraint profiles rather than inherent muscle potential. This suggests that
strategic variation (combining or altering exercises within a program that
target the same muscle or performance), complementary exercise sequences, and
individualized programming can produce more sustained growth and help explain
away phenomena that might have previously been misattributed to altered
functioning intrinsic to trained muscles In short, rather than invoking
complex, intracellular “brakes” on hypertrophy, we propose that stagnation is
an emergent property of the unequal adaptation rates of the various tissues
required to produce training stimulus that, in principle, can be substantially
circumvented.
2. The Interdependence of Muscle and Constraints
No exercise isolates a muscle completely.
Performance in any lift depends not only on the target muscle but also on
numerous supporting constraints such as joint stabilizers, posture-specific
musculature, connective tissues, and the technical skill needed to execute the
movement effectively. Nor is any single training stimulus likely to influence
the many properties of a muscle evenly. Performance of an exercise can be
conceptualized as the joint product of the muscle and its constraints: if
either is underdeveloped, overall performance is limited. Importantly,
constraints and muscles do not adapt at the same rate: some supporting tissues
and skills improve quickly, while others may plateau earlier, or even become
worn out over time. Logically, this means that training progress is often
limited not by the growth potential of the target muscle, but by whichever
constraint is lagging.
Figure 1
graphically illustrates this with the familiar Stimulus-Recovery-Adaptation
(SRA) framework. After intense training, performance of an exercise can be
thought of as having an SRA cycle: performance of the exercise is greatly
diminished for a while, inducing recovery processes that gradually drives it
back towards the level that it was capable of before training, which it
temporarily overshoots (i.e., supercompensation) before returning to the
baseline (Figure 1a). The key to
successful “progressive overload” training is to then train the exercise again
before this temporary boost decays. Train slightly too early or too late, and
no progress ensues, or performance is driven downward as recovery capacities
become overwhelmed (Figure 1b).
Figure 1.
Implications of hypothetical SRA curves with different periodicity for long-run performance. (a) General shape of the SRA curve, with performance being depicted on the y-axis and time on the x-axis. Performance declines due to fatigue, then recovers to a temporary peak above baseline, measuring the super-compensation effect. (b) When workouts are repeated such that each workout is timed during a period of supercompensation, performance can gradually improve (curve i); if workouts are too infrequent for the trained characteristic, the trainee simply “treads water” or fails to progress (ii); if workouts are too frequent and are undertaken before recovery can return to baseline, performance capability progressively declines (i.e., “overtraining”) (iii) (c) Stimulus provided from an exercise (S) are charted across workouts (t) under different scenarios. If SRA curves for both muscle and constraint are aligned and timed to allow progressive overload (i) progress is sustained. If training is optimized for progressive overload of the muscle but the SRA timing means the constraint is treading water, there is a rise in how stimulative the exercise can be, but this converges on a “ceiling” due to the constraint (ii). If the constraint is progressively overtrained, then even if initially stimulative of the muscle, workouts will begin to negatively impact the ability of the exercise to stimulate growth (iii). Modified from Figure posted by OX Fitness (2021).
Figure 1.
Implications of hypothetical SRA curves with different periodicity for long-run performance. (a) General shape of the SRA curve, with performance being depicted on the y-axis and time on the x-axis. Performance declines due to fatigue, then recovers to a temporary peak above baseline, measuring the super-compensation effect. (b) When workouts are repeated such that each workout is timed during a period of supercompensation, performance can gradually improve (curve i); if workouts are too infrequent for the trained characteristic, the trainee simply “treads water” or fails to progress (ii); if workouts are too frequent and are undertaken before recovery can return to baseline, performance capability progressively declines (i.e., “overtraining”) (iii) (c) Stimulus provided from an exercise (S) are charted across workouts (t) under different scenarios. If SRA curves for both muscle and constraint are aligned and timed to allow progressive overload (i) progress is sustained. If training is optimized for progressive overload of the muscle but the SRA timing means the constraint is treading water, there is a rise in how stimulative the exercise can be, but this converges on a “ceiling” due to the constraint (ii). If the constraint is progressively overtrained, then even if initially stimulative of the muscle, workouts will begin to negatively impact the ability of the exercise to stimulate growth (iii). Modified from Figure posted by OX Fitness (2021).
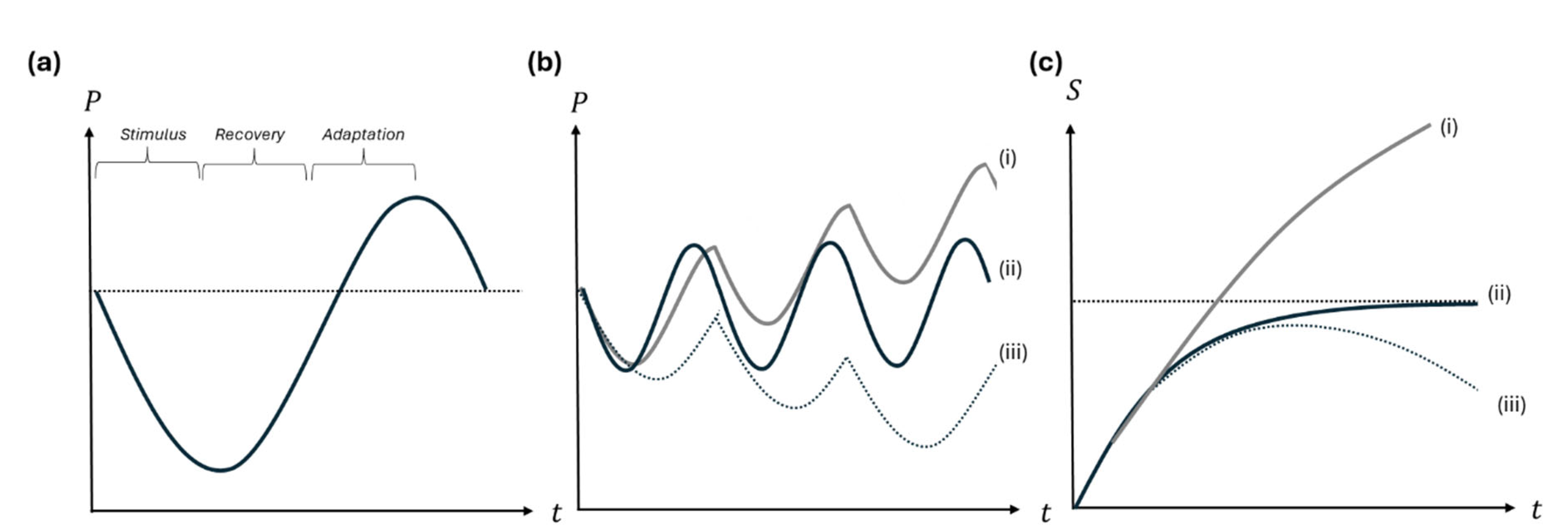
The SRA curve for an exercise can be conceptually
decomposed into SRA curves for the different components contributing to its
performance. For present purposes, we may consider the SRA curve of the muscle
or other fitness quality targeted by the exercise on the one hand and the SRA
curve of what we are calling “constraints” on the other. Implications of
disparities in how SRA curves are aligned for how stimulative an exercise is
across workouts are depicted in Figure 1c.
If the muscle and constraint SRA curves differ, both the SRA curves for
constraints and the targeted muscle will diverge from the overall SRA curve of
the exercise. It follows that the optimal inter-workout interval for the
targeted muscle will be different to that of both the constraints and probably
the overall exercise. If the constraint curve recovers more rapidly, then
workouts timed to maximise the supercompensation of the target muscle risk
training only when super-compensatory effects of the constraints have
dissipated. Conversely, if it is slower to adapt than the muscle, then it means
training again too soon for the constraints. In either scenario (both may be
simultaneously true for different constraints), it is easy to see that
progressive overload of the target muscle will at some point be choked off by
the constraints failing to keep pace. Likewise, training optimised for the
overall SRA curve will eventually fail to maximise growth of the target muscle
because of the constraints. In either case, when only one type of exercise
stimulus is being applied, it is inevitable that the constraints will be key.
2.1. A Basic Model of Training Stagnation
The idea can be captured
formally with the following simple model.
For
brevity, we refer to this as the “motor-clutch model,” drawing on the metaphor
of a power drill. The motor represents the target muscle, while the clutch
represents supporting constraints that limit how much torque the motor can
deliver. Strengthening the motor alone cannot guarantee greater output if the
clutch slips or is locked in a low setting; likewise, raising the clutch
setting without a stronger motor is ineffective. Progress requires advancing
both together, or at least alternating settings to keep drilling deeper into
the material. Thus, the point here is not that muscle adaptation is never the
limiting factor: poor training technique may fail to stimulate the muscle
effectively, and different properties of a muscle may respond at different rates,
such that one becomes a constraint. Nor do we deny that intrinsic muscular
factors could, in principle, impose diminishing returns. Rather, the claim is
that we do not need to assume intrinsic resistance to explain adaptive
resistance. Apparent stagnation can be accounted for more simply by recognizing
the role of constraints.
Let represent the state of the target muscle at time , and the state of the relevant constraint. Suppose that
overall performance, , is a function of the capacity of the muscle
multiplied by the constraint:
This means that low levels of adaptation in either
the target muscle or constraints reduce performance. Training generates a
stimulus, , proportional to performance on the exercise:
where measures how efficiently performance translates
into stimulus for further adaptation (a more efficient exercise means more
performance leads to greater stimulus per unit of performance). Both the muscle
and the constraint adapt in response to this stimulus, but at different rates:
where represent adaptation efficiencies and as natural rates of decay or recovery for muscle
and constraint respectively. If , the muscle adapts faster than the constraint, and
the constraint eventually becomes the bottleneck limiting further improvements.
Notice that even if the muscle’s growth potential is otherwise unlimited
(indeed, muscle may theoretically be improving at linear or constant rates of
return to training if constraints would allow it), performance (and therefore
overload stimulus) stagnates once falls behind. Once the constraint dwindles too
much, the stimulus generated by the increasingly compromised performance is
insufficient to overcome natural decay rate and the muscle will begin to
atrophy.
This framework also explains why alternating
between complementary exercises can be advantageous. Suppose exercise and stimulate the same muscle but rely on different
constraints and :
Alternating between and would allow the muscle to continue progressing
while each constraint has time to recover and adapt in turn. Over time, this
strategy can sustain growth more effectively than focusing exclusively on
either exercise. The point here is that apparent plateaus or blunted responses
may simply reflect constraint limitations rather than intrinsically diminishing
responses of the muscle, and in fact may mask constant or even rising
potential. For if we think of “skill” as the capacity to stimulate the target
muscle, growth in this capacity may lead more quickly to stagnation as the
muscle’s adaptations vastly outstrip that of the constraint. This provides an
added, albeit subtle, rationale for strategic exercise variation in training
programs [1],
skilful management of complementary constraints, and perhaps individualized
programming (if people vary in their particular constellation of constraints
relative to a training goal) to optimize long-term adaptation.
This perspective points to some counter-intuitive
implications for training. First, exercises that highly optimize stimulus to a
target muscle relative to constraints may lead to earlier plateaus, since
constraints will become limiting sooner than when stimulus is more evenly
distributed (they are essentially outrunning the muscle’s support system).
Second, the complementarity of exercises may be far subtler than appearances
suggest: two seemingly similar movements, such as front squats and high-bar back
squats, may be highly complementary and work well in alternation because they
crucially shift which constraints are stressed despite having very similar
movement and loading patterns vis-à-vis the targeted musculature (e.g., the
quadriceps). Meanwhile, two superficially different exercises, like leg
extensions and leg press, may provide little complementary benefit if they fail
to engage the primary target muscles as effectively or stress crucial
constraints. Third, the optimal sequencing and variation of exercises may not
be obvious. Because constraints adapt at different rates, the order, timing,
and even small execution differences can substantially influence long-term
progress. Seemingly minor variations in technique may therefore accumulate into
large effects over time.
2.2. How Skills and Constraints Squeeze Muscle Potential
Above we dichotomized the tissues of interest into
the target muscle and everything else that could be a constraint. Here we
complicate things slightly by identifying two key constraints differing in
their response to training stimuli. One is denoted as , representing constraints that adapt slowly and so
a prone to overuse, such as perhaps tendons and joints, and the other is
characterised as adapting more rapidly but prone to rapid decay or
under-training at low training frequencies, which we will think of as skill or . We use these labels “tendon” and “skill”
hereinafter as an intuitive shorthand for slow-adapting structural constraints
and fast-adapting enabling factors respectively [2].
We again denote the state variable representing the target muscle as . The goal is to maximise cumulative growth in the
target muscle over time:
We now introduce a set of exercises, indexed by:
where each exercise stimulates the same target
muscle but differs in how it loads the tendon and the skill component. To keep
things simple, we characterise each exercise by three, fixed coefficients and define: (1) : effectiveness of exercise at stimulating the target muscle; (2) : degree to which exercise loads the tendon / constraint, and (3) : degree to which exercise depends on and trains skill. For example, a barbel
back squat might have high , high , and high , whereas a machine hack squat might have similar , lower , lower .
When exercise is performed at time , performance is determined jointly by the current
state of muscle, tendon, and skill:
This means that if any component is weak,
performance is limited, and therefore stimulus to the target muscle is limited.
In this model, exercises differ only in how strongly they rely on each
component. This captures the general truisms that some exercises are more
tendon/joint limited than others and some require more skill to execute
effectively. As with before, we have the constraints (skill and tendons) acting
as potential choke points that are both endogenously determined: If is low, exercises with high become especially ineffective even if muscle is
capable of more growth. Meanwhile, if decays between exposures, exercises with high lose effectiveness unless repeated frequently. As
a simplifying assumption, we set for all exercises [3],
so that exercises only differ in which bottlenecks they activate and to what
degree. Performance then reduces to:
We next specify how training with a given exercises
produces adaptation signals in each component, and how these components evolve
over time.
Each component adapts according to a simple law:
where , is the stimulus received at time , is responsiveness, governs decay toward baseline . We assume that each exercise delivers a
component-weighted stimulus. For muscle
For the tendon constraint
and for the skill constraint
This means that exercises that load the constraint
more heavily generate more tendon stimulus and fatigue, that exercises which
are more technically demanding generate more skill stimulus, and all stimulus
scales with realized performance. We now impose the key structure:
This formally means that skills adapt quickly and
decay quickly, muscle adapts at an intermediate rate, and the tendons adapt
slowly and decay slowly. This assumption is key to generating the interesting
results that follow.
First, let’s consider the repeated use of a single
exercise. As before, the constraints are high relative to demands, rises rapidly, all components adapt positively. This
looks like rapid muscle responsiveness. But, over time, tendon adaptation lags
repeated loading making become the binding constraint. Performance growth
slows or halts:
Crucially, muscle capacity may still be capable of increasing at the point it
becomes choked off because realised stimulus no longer rises. The produces the
illusory adaptive resistance we mentioned earlier. Now, suppose we alternate
two exercises, and , such that both target the same muscle but and . Then, when training , tendon components stressed by recover, and vice versa, while muscle stimulus
remains high each training session. Formally. never becomes binding for both exercises
simultaneously, allowing for continued growth in the target muscle without
increasing total “fatigue”.
Now, what happens with the fast-adjusting skill
component? If and differ substantially, alternation is too wide,
then skill supercompensation decays between exposures, depressing performance
and again choking off muscle stimulus. This adds the insight from the previous
section that exercises should differ strongly in constraints but minimally in
skill demands. In other words, because skill adapts and decays quickly,
exercises targeting them must be practiced frequently enough or else falls below its peak muscle stimulus is reduced (a
direct result of ). A corollary of this is that both types of
stagnation can occur simultaneously, with overtrained tendon capping
performance from “above”, undertrained skill can cap performance from below,
squeezing muscle in between. The moral of this model is that sustained muscle
growth is only possible in the “Goldilocks” where tendons are not overtrained
and skill is sufficiently practiced, otherwise these both act as constraints
that sandwich performance and hence effective stimulation of the target muscle.
This may correspond to real world reports where
training feels hard, effort is high, felt fatigue is high, and yet progress
stalls. Some examples might include when barbell-based lifts regress after time
off despite muscle mass retention, athletes feeling “uncoordinated” or “out of
groove”, machines or cables producing better short-term hypertrophy for some
individuals, and instances where more complex lifts produce plateaus that
vanish with slight technical variation. These are all consistent with our model
suggesting skill may act as a fast-decaying enabling factor for muscle
stimulus, based simply on differential adaptation rates and multiplicative
performance (rather than invoking mysterious neural adaptations or overloading,
muscle “confusion”, or exercise-specific hypertrophic mechanisms).
We suppose that sustained growth of the target
muscle is possible if:
Or, more weakly (and perhaps more realistically),
that muscle can continues to increase for an arbitrarily long time without
converging to a finite ceiling. Importantly, this does not require monotonic
growth while it allows for plateaus and oscillations. It means that, even if
muscle is still capable of adapting, and skill is perfectly maintained (best
case), tendon must eventually become binding unless it adapts at least as fast
as muscle, by assumption:
So, over time, tendon load accumulates faster than
tendon capacity converges to a steady state or declines and
performance ceases to increase Formally:
Since muscle stimulus is proportional to
performance:
which implies:
This gives us the following result. With one
exercise, sustained muscle growth is generically impossible unless constraints
adapt at least as fast as muscle. Now suppose we alternate exercises and , each session focusing on one. Assuming both
target the same muscle, but differ in constraint loading, this means, formally
that . Assuming skill demands are similar (), When training , tendon components heavily loaded by recover and skill remains near peak (because
exercises are similar) When training the reverse holds. Thus, no single constraint is
loaded at maximal intensity every session, and recovery time is endogenously
introduced by alternation This means that the muscle still receives
near-maximal stimulus every session. Two exercises are sufficient for sustained
growth if and only if no single constraint is essential and maximally loaded by
both exercises. In other words, IFF
for every binding constraint component, then
constraints alternate between loading and recovery, does not converge to a binding ceiling, and
performance can continue to increase with . This is a key prediction of our model and
embodied conceptually in Figure 2: two
exercises are sufficient to permit sustained growth of the target muscle
provided their constraint profiles differ sufficiently and that their skill
demands overlap enough. Put another way, skills practiced by each exercise
should transfer to the other, so that the skill “floor” on performance does not
drop while letting the “ceiling” imposed by the tendon to recover.
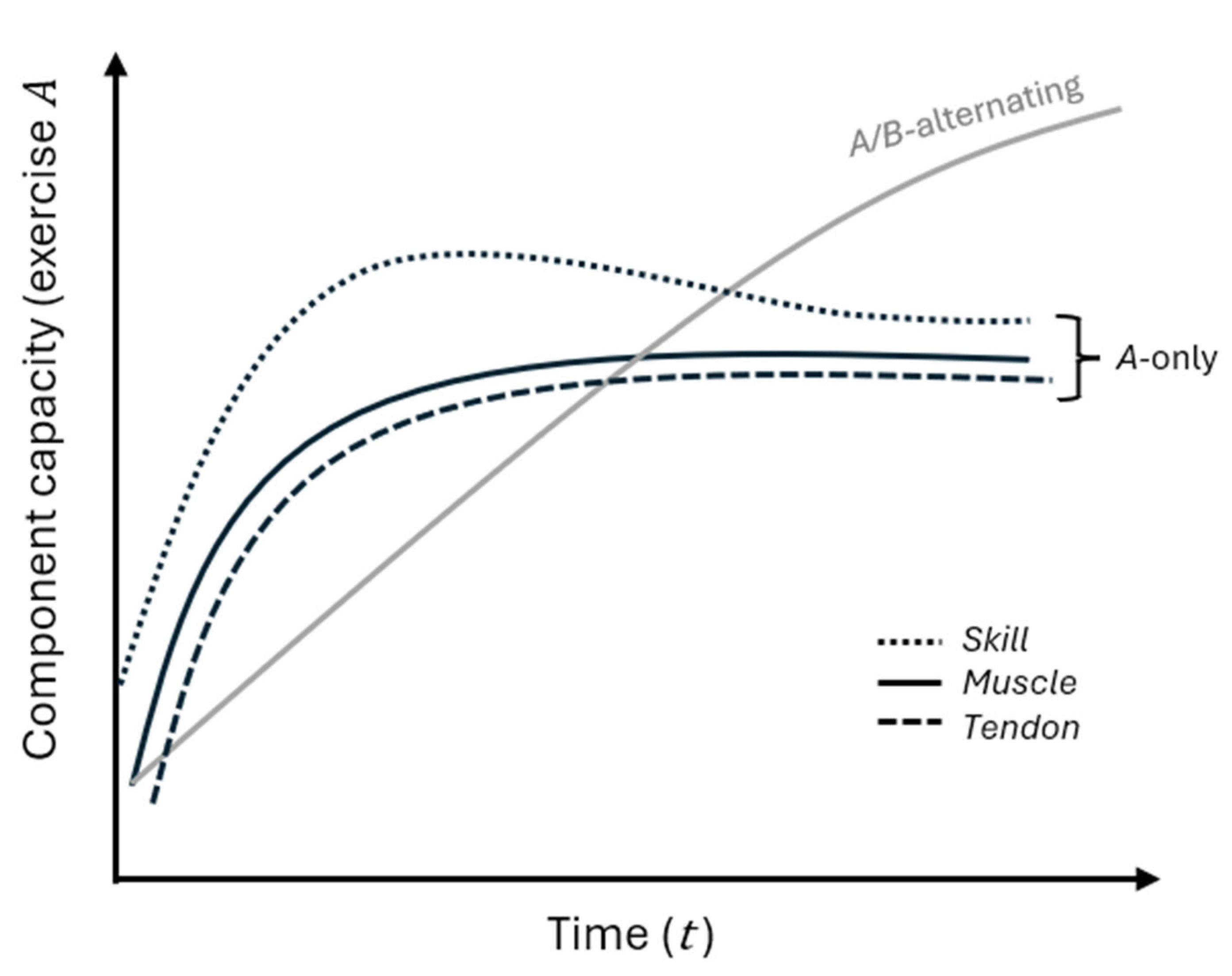
), the dotted line skill
(
), and the dashed line
tendon (
) under a single-exercise
(A-only) program. All plateau over time, with tendon lagging and limiting
further muscle gains. The grey line shows muscle under an alternating A/B
program, continuing to rise as tendon constraints recover, illustrating how
strategic exercise variation sustains long-term growth.
Figure 2.
Muscle, tendon, and skill under single versus alternating exercises. Time is on the x-axis, and component capacity relative to exercise A is on the y-axis. The solid black line shows muscle (
Figure 2.
Muscle, tendon, and skill under single versus alternating exercises. Time is on the x-axis, and component capacity relative to exercise A is on the y-axis. The solid black line shows muscle (
2.3. Short Versus Long Run Implications for Exercise Programming
Nevertheless, two exercises or variations may not
be enough. The above establishes possibility, not optimality. It assumes
near-perfect constraint complementarity. It ignores skill decay from infrequent
exposure. Which raises the next question: What happens when constraint overlap
is imperfect, or skill decays too quickly? We now relax the knife-edge
assumptions that made two exercises sufficient and ask: Under realistic
conditions, can alternation between just two exercises still lead to
stagnation?
The answer will be yes, for two independent
reasons: (1) Imperfect constraint complementarity, and (2) Skill decay due to
infrequent exposure.
Previously, we assumed exercises and differed enough in how they loaded the tendon that
each allowed meaningful recovery from the other. Now suppose instead that both
exercises load the same constraint to some degree but with different
intensities. Formally:
with:
What changes? Even though alternation reduces peak
loading, the constraint is never fully unloaded. Over time, tendon stimulus
accumulates, recovery is incomplete and so converges to a finite steady state. As before:
So, for continuous improvements, constraint relief
must be qualitative, not merely quantitative. Put another way: reducing how much
a constraint is loaded is not enough; what matters is whether the identity of
the binding constraint changes. Small differences in loading tendon constraints
merely slow stagnation, but do not eliminate it (assuming constraint recovery
exhibits diminishing returns and does not fully reset under partial unloading).
Also, only rough overlap in skill acquisition can likewise lead to stagnation.
Let each exercise require exercise-specific skill:
When training exercise , increases and decays. When training , the reverse occurs with strict alternation each
skill is practiced only every other session. As exercise-specific skills may
partially overlap here denotes the exercise-relevant component of skill.
If is large relative to training frequency, then:
for all , where denotes peak attainable skill. Performance becomes:
Even if tendon recovery is adequate and muscle capacity is rising, performance is suppressed by submaximal skill. Muscle stimulus is therefore reduced.
Now let’s introduce a third exercise , such that its constraint profile differs from both and while its skill demands overlap strongly with them
Formally:
Now, each constraint gets longer recovery windows and skill remains reinforced frequently across exercises. This would lead us to expect performance rebounds. This is the first point where the number of exercises matters structurally, not heuristically. When constraint overlap is non-trivial and skill decays rapidly, more than two exercises may be required to permit sustained muscle growth. The minimum number depends on (1) degree of constraint overlap, (2) rate of skill decay, and (3) similarity of skill demands across exercises. This immediately helps explains why some lifters stagnate on “A/B” programs and why adding one carefully chosen movement unlocks progress. It also helps to say why random variety for its own sake often fails: optimal variation reduces tendon overlap and maximises skill transfer while random variation will necessarily be inferior to this proportional to the total number of exercises selected.
We now have opposing forces affecting what the optimal number of exercises is likely to be. The force encouraging more exercises to be included is the improvement to constraint recovery this can provide along with the reduced cumulative overload. Against this, favouring fewer exercises, is that this may help to preserve or promote skill, increasing the per-exercise stimulus. This guarantees an interior optimum.
This optimum likely differs depending on the time horizon over which the lifter is concerned. We now consider a set of exercises
that are rotated in a fixed cycle, one per session. The question is how does increasing affect long-run growth of the target muscle. Adding an exercise affects the system in two structurally opposite ways.
First, as increases, each individual constraint is loaded less frequently, recovery windows lengthens and so average constraint capacity rises. In terms of our model, cumulative tendon load per unit time is “spread out” across exercises, loosening it as a binding constraint. This effect is initially strong but strictly diminishing in returns, as once constraints are no longer binding, further relief from them has little value.
Second, working in the opposite direction is the effect of exercise number on skill dilution. If each exercise is practiced only once every number of sessions, exercise-specific skill decays between exposures. Given:
we have:
This reduces realized performance and hence effective muscle stimulus. Importantly, this effect is presumably convex: skill loss accelerates as spacing grows, and spacing grows with number of exercises. Let average realized muscle stimulus per session be:
Then structurally, constraint relief makes increase in ; skill dilution makes decrease in . With standard regularity assumptions, this implies that has a singular peak. Thus, there exists an optimal finite number of exercises that maximizes long-run muscle growth. What can we say about such an optimum? From above, we can say it must be more than one exercise, possibly greater than 2, but strictly finite. This may mean adding one more exercise at a time will often work, until it suddenly does not. The optimal number depends endogenously on constraint overlap, skill decay rate and similarity across exercises, and training frequency (if a greater frequency is possible, there is a higher feasible ). In short, although there is no universally “right” amount of variation, there is a principled way to think about it beyond variety for variety’s sake. Crucially, in practice, exercises are not interchangeable, variation must be structured, and random novelty is likely worse than repetition (especially in the short term). The model therefore predicts that the best exercise sets differ in constraints, overlap in skill, and converge on the same targeted muscle.
We now distinguish between two optimization problems: (1) short-run optimization that maximises muscle growth over a finite horizon , and (2) long-run optimization, that maximizes sustainable growth as . The key result here concerns whether these generally select different exercise strategies.
Based on the foregoing, we can already see that, in the short run, progress is less constraint-sensitive (constraint capacity is high relative to demands). As constraints are not yet binding, performance is initially driven and limited by muscle responsiveness. As a result, repeating a small set of exercises (maybe just one) maximises muscle stimulus intensity while minimizing skill dilution. Formally:
is small when is small. This reproduces the “newbie gains” phenomenon and predicts greatest early success from minimalist programs[4]. Meanwhile, as increases, slow-to-recover constraints accumulate load as recovery becomes incomplete. Eventually the same strategy that maximized early gains produces stagnation. In the long run:
is strictly larger than the short-run optimum. In other words, as depicted conceptually in Figure 3, our tortoise versus the hare comparison yields a tortoise versus the hare result: exercise strategies that are optimal in the short run are generically suboptimal in the long run. This is a “dynamic inconsistency” result” in the sense that the training strategy that produces the fastest results in the first, say, 24 weeks is, by definition, a strategy that will assuredly cause a plateau by week 52 (whereas different programs may enable greater overall progress by the 52nd week despite initially being outpaced).
This may help explain mismatches between various self-reinforcing beliefs. For example, the notion that “simple programs work best” or “basics are best” versus “you need novelty to break plateaus”. Both are partially correct, but because the mechanisms is misunderstood their implementation may be misapplied. More generally, what is commonly interpreted as adaptive resistance of muscle is often the predictable outcome of optimizing training over too short a horizon in the presence of heterogeneous adaptation rates. This holds without invoking either muscle-specific novelty effects, exercise-specific hypertrophy, intrinsic responsiveness decline or “bad genetics”.
3. Discussion
Although the motor-clutch model may be simple, much follows from it and allows several widely observed and otherwise separate phenomena to be interpreted through a single approach. As mentioned, so-called “newbie gains” may emerge not from the absence of “adaptive resistance” that training builds into the muscle, but because both the muscles targeted by exercise and the factors that potentially constrain them are initially unchallenged. This allows components to respond positively to training for a while without being choked off by constraints failing to keep pace or be driven into an overtrained state. Stagnation then occurs when the constraining components fall behind in improvement relative to the targeted muscle. This agrees with the general observation that trainees who appear to plateau under one exercise regime tend to resume progress when switched to another (Israetel, Hoffman, & Smith, 2015; Schwarzeneggar, 1998). Relevant here is the review by Kassiano and colleagues (2022) which found evidence supports the view that some variation in exercise selection promotes some hypertrophic and strength adaptations in muscles targeted compared to performing simply performing the same exercises, although very large or random variations in exercises hinders muscular adaptations. The framework offers a further testable prediction in the following general form: alternating complementary exercises or other modes of training a targeted muscle will result in more sustained progress for that muscle than training focused on the same exercise or regime alone. Designing experiments to test this would require careful consideration of what the relevant constraints are and so what exercises count as distinct but complementary. For now, this hypothetical point can be illustrated with the example already given: a training regime that alternates between a workout focused on front squats with one focused on back squats should produce more sustained progress in the quadriceps than a regime focused on either alone (or one that selects less relevant exercise combinations). A corollary is that, if the relevant constraints can be identified, training specialized at enhancing the capacity of those constraints could also play a potentially powerful role in supporting continued progress.
3.2. “Anabolic Resistance” Versus Diminished Training Stimulus?
The same logic can be used to explain the apparently paradoxical finding of blunted anabolic responses to high protein meals following training in experienced lifters (Morton et al., 2018). Ordinarily, a high quantity of quality protein intake can induce muscle protein synthesis in people even in the absence of resistance training, while resistance training can likewise be a potent stimulus for muscle protein turnover. Combining the two (i.e., ingesting a high protein meal post-workout) has been found to be highly synergistic (Joanisse et al., 2021), as resistance training fundamentally “sensitizes” muscle to utilizing dietary protein (Moore, 2019). However, this multiplicative effect has not detected as being so substantial in participants who identified as being “experienced” (Atherton & Smith, 2012; Burd et al., 2011; van Vliet, Burd, & van Loon, 2018). The implication, then, is that protein intake and hypertrophic training are not as complementary with each other once people have been training for a while. In contrast, we would argue that many such lifters may be better described as “stagnated” than experienced: their training no longer delivers sufficient stimulus to the target muscle because constraints have limited their ability to do so. Alternatively, their training outside of the study may continue to progress, but the same training regime used in the research protocol may simply not be as stimulative for these lifters due to the relevant constraints as compared to novices. In either case, the blunted anabolic response may simply be measuring the effect of less stimulation from the workout, rather than indicating something deficient within the muscle. Incorporating some independent measure of how effective the training stimulus is in such studies would be useful to resolve the ambiguity.
If elevating muscle protein synthesis is a key factor in promoting exercise adaptation, then the current analysis implies that experienced lifters should not neglect nutrition as protein dosing and timing may be just as important, providing that they are training in ways that allow both their constraining and targeted components to improve. An empirical prediction is that the training protein intake interaction effect on muscle protein synthesis should be significant in participants regardless of their years of training experience provided that their workouts are providing effective stimulus[5]. Parallel predictions may be made for other complementary inputs that may have little direct effect if the training stimulus is bottlenecked by constraints but become potent when constraints are managed in a way that allows training to produce effective hypertrophic stimulus.
3.3. Implications for Research
To explore the sorts of dynamics alluded to here, constraining aspects of an exercise of interest could be identified, measured, and tracked alongside those of the muscle targeted over the course of a training program in future studies. For example, measures of ankle mobility and strength, spinal erectors, cardiac output, and other potentially limiting factors could be monitored to see if changes in these measures predict progress in quadriceps hypertrophy as targeted by a program based on squat variations. Consideration of constraints also means caution should be used when interpreting ostensibly straightforward comparisons of training modes and techniques. A recent review of resistance training variables (Bernárdez-Vázquez et al., 2022) highlights that while exercise variation and training frequency are key variables, empirical results remain inconsistent or yield null effects when volume is equated. From the perspective of the Motor-Clutch model, these inconsistencies are predictable. If experimental protocols do not account for the complementarity of constraints — specifically how different exercises load slow-adapting tissues versus fast-decaying skills — the benefits of variation may be masked, as variation for variety’s sake may often be counterproductive on our analysis. Research that treats all exercises as interchangeable 'units of volume' ignores the underlying divergence in constraint loading that our model suggests is the primary driver of long-term progress.
To give another example, research comparing training using machines versus free weights typically report no overall difference in outcomes, leading to the conclusion that either tool is equally effective (Haegen et al., 2023). From our perspective, this null finding is unsurprising. The same observations could be accounted for by a variety of factors, including different constraint profiles across exercises. For instance, suppose that the ratio how well the target muscle is stressed relative to the constraint by exercise X is mirrored by the how the muscle and constraint is affected by exercise Y (that is ). Both therefore could converge on a similar asymptote over time despite having different effects, with one perhaps being vastly more effective at stimulating the muscle than the other. How constraints adjust (or fail to adjust) in response to training could mask meaningful differences in how different techniques stimulate a target muscle, and it is not unreasonable to suppose free weight exercises differ dramatically from machines in both regards. Future research should therefore consider constraint-specific effects, rather than relying solely on group aggregates and explore whether findings based on a single type of training are robust to more complex programming factors that may affect muscle/constraints dynamics, such as periodization or exercise variation.
Nowhere would we expect constraints to conceal the relationship between important variables and training more than when individual differences are concerned. If individuals differ in their particular constellation of constraints (even if only initially), then for some individuals restricted to a particular training regime, constraints may cap their progress earlier, while others flourish for longer under the same conditions[6]. Individual differences in studies are normally treated as noise, as athletes often appear to differ greatly in their capacity to build muscle under the same program. This is again commonly taken as an indication of inherent differences in the responsiveness of the targeted muscles. Yet, if different exercises place different demands upon constraints that also vary among individuals, then individual variation in responsiveness to the same exercise regime may reflect constraint profiles more than intrinsic muscle properties. For instance, a trainee whose spinal erectors or ankle flexibility are disproportionately taxed by barbell squats may stagnate sooner when squatting is used as the primary exercise for quadriceps development, even though their quadriceps may respond well under a different program. Using only one exercise as a test of potential could therefore be very misleading and discourage people prematurely from pursuing resistance training (especially if the constraints themselves can be subject to training effects). This is a key corollary of our model, and supported by findings recently summarized by Saeterbakken (2025) regarding low transferability underscoring the danger of interpreting a plateau in a specific lift as a plateau in muscular adaptation.
3.4. Comparison with Existing Frameworks
It is instructive to compare our approach with some contemporary training philosophies that recognize the usefulness of exercise variation for balancing training stimulus with fatigue. A popular example is Israetel’s concept of the Stimulus-to-Fatigue Ratio (SFR) as a suggested criterion for exercise selection and program design (Israetel, Hoffman, & Smith, 2015), and can be compared with the present approach in Table 1. On this view, decisions are made based on how much hypertrophic stimulus is generated for a given amount of fatigue. Smith (2022) and others have similarly highlighted the role of “strategic variation” or “the principle of variation’ in exercises and how they are performed as ways of managing fatigue over time. Both approaches have intuitive appeal and may have a lot going for them and so should not be gainsaid too much here. It is sufficient for our purposes to note that they may nevertheless be limited from the present standpoint in the following ways. First, the notion of “fatigue” is difficult to specify, much less measure. It can refer to systemic tiredness, local muscle soreness, or technique breakdown under load. Because the term can be slippery and its operational meaning is hard to pin down, decision-making and theories based on this can easily become arbitrary. An advantage of the Motor Clutch Model is that it implies specific constraints should be identified and addressed, and variation is a practical tool for both these purposes. Moreover, fatigue minimization may lead to long-term stagnation if it means failing to address limiting constraints on exercises and modes of training that would otherwise be very effective. To make this example intuitive, consider again barbell squats. These can be demanding on the cardiovascular system, which can become a significant constraint at higher repetition and set ranges. Strict SFR considerations would seek to train in a way that maximizes hypertrophic stimulus relative to cardiovascular-related fatigue, which might involve some set range of repetitions, sets, workout frequency, etc. If we assume that this configuration of parameters is a subset of a wider one that would include potentially more stimulating ways of training the squat – if only the cardiovascular constraint was alleviated – then the SFR is clearly a poor strategy in the longer term. This is because the cardiovascular components likely show a more elastic response to training than most other characteristics determining squat performance (e.g., as compared to bones, tendons, and muscles), corresponding the “skill” parameter in our model, because they can be improved by training quite rapidly over time (Hughes et al. 2017). In other words, the cardiovascular constraint may be severe in the very short-term and so would be avoided by a pure SFR maximisation program. But improving it might be comparatively easy, which in turn could vastly expand the potential for progression in the target muscle over the medium to longer term, depending on what level of advantage there might be from unlocking training at these ranges of higher repetitions or sets.
Focus on fatigue as the key denominator additionally risks obscuring the issue that the more fundamental trade-off in training is not between stimulus and fatigue but between the marginal benefit from competing alternatives. On this basis, an alternative rationale is to see exercise selection and programming as a problem of allocating time and effort according to the overall improvements induced in the target muscle expected from training one way versus another[7]. Our framework sharpens this idea further. Because hypertrophy is produced by the joint product of the target muscle and its supporting constraints, what matters is not avoiding the constraints but enabling them to improve them in addition to the targeted muscle. Two exercises, such as the back squat and front squat, may appear redundant based on SFR as both may arguably produce higher overall “fatigue” than various isolation movements while targeting a common muscle. Yet they may be complementary with each other precisely because they similarly stress overlapping muscles while subtly but sufficiently differing in how they engage the constraints. If this complementarity thereby raises the long-run ceiling for muscle growth, then the SFR point is moot. The real question, then, is not how some fatigue is affected in the general or abstract sense, but how exercises affect the stimulus-response-adaptation of constraints. Exercises that target the same muscle but are very different in other ways may fail to complement one another if they neither tax constraints productively to raise that ceiling nor generate strong enough stimulus for the target muscle. In short, whereas SFR-based thinking encourages athletes to minimize constraint stress – effectively treating it as static or a negative influence – the motor clutch model mindset suggests training in ways that enable both constraints and muscle to adapt, hand in glove.
If variation along the lines recommended here are effective, it should be that the same principle has been repeatedly discovered in various guises. Some aspects or versions of training periodization likely reflect the differential impact of different phases on constraints as well as their influence on muscles targeted. Nevertheless, exercise variation may be a crucial but underappreciated variable in periodization planning (Fisher et al., 2018), and the Motor Clutch Model may be useful for research and program design in this regard. It may also be noted that many training methods, such as “pre-exhaust” techniques that combine movements in orders or agonist supersets to target a common muscle, such as nautilus pullovers with T-bar rows to train the latissimus, likely have utility because they get around constraints unique to each exercise (e.g., biceps and forearms). These combinations often stress using an “isolation” or single joint movement either before or after a compound or multi-joint movement. Based on the present model, we would not expect such pairings to be superior to straight set training or other more conventions approaches because the constraints will still begin to bite, although mixing them in may be a way to allow the constraint ceiling to rise. Additionally, our approach would suggest that the compound/isolation distinction is not important but rather may be useful as a heuristic for selecting exercises that likely differ in their constraint profiles (Rosa et al., 2023), and there may be compound-compound or isolation-isolation exercise pairings that work as well or even better.
Likewise, the use of “accessory movements” in strength sports (e.g., powerlifting and Olympic lifting), have been justified based on their role in addressing weak points in a competition lift, building robustness to injury, and for adding training volume without excessive stress. From the standpoint of SFR, accessories may be selected because they promote localized stimulus at lower levels of general fatigue. Our model adds a potentially stronger rationale: exercises function best as accessory movements not because they are less fatiguing in the abstract but because they alter the balance between the muscle targeted for competition and its constraints. A well-chosen accessory lift provides a stimulus in a manner that either (1) reduces the constraint bottleneck, such as so-called Romanian deadlifts to strengthen the posterior chain in ways that prevent the lower back from capping progress in conventional deadlifts (directly raising the constraint ceiling), or (2) stresses the target muscle more than the constraints relevant to the competition lift, such as belt squats providing a strong and similar stimulus for lower body muscles that are important for competition back squats while giving constraints (e.g., shoulders and lower back) time to recover. The present thesis may help clarify the role of accessories not as peripheral “add-ons” or redundant duplication but as central to raising the long-run ceiling of the main lift by managing the joint development of the targeted muscles and constraints. As alluded to for hypertrophy, this may help explain why some accessories traditionally seen as “very similar” to the main lift (e.g., front squats for a back squat specialist) can be more complementary than exercises that seem superficially different (e.g., leg extensions). The key is not how much the accessory departs from the main lift in general, but how it reshapes the constraint parameters in particular.
4. Conclusions
What is often attributed to adaptive resistance or other mysterious malaise located within the muscles targeted by training may instead reflect constraint-related bottlenecks. Recognizing constraints clarifies phenomena such as newbie gains, individual variation, and blunted responses to protein rich meals in experienced trainees. Strategic exercise variation, complementary constraint management, and individualized programming can produce more sustained growth than traditionally assumed. For both practitioners and researchers, considering adaptation as much a constraint-limited rather as it is a muscle-limited phenomena offers actionable insights and testable predictions. If this paper encourages exploration of new avenues for improving training outcomes and interpreting resistance training research, then it has achieved its purpose.
Ethics Approval Statement: no new data was generated in this article and so not ethics approval was necessary (IRB statement not applicable).
Conflicts of Interest
The author has no conflict of interests to declare and received no funding to support the writing of this article.
References
- Atherton, P. J.; Smith, K. Muscle protein synthesis in response to nutrition and exercise. The Journal of Physiology 2012, 590(5), 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Bernárdez-Vázquez, R.; Raya-González, J.; Castillo, D.; Beato, M. Resistance training variables for optimization of muscle hypertrophy: An umbrella review. Frontiers in sports and active living 2022, 4, 949021. [Google Scholar] [CrossRef]
- Buckner, S. L.; Kuehne, T. E.; Yitzchaki, N.; Zhu, W. G.; Humphries, M. N.; Loenneke, J. P. The generality of strength adaptation. Journal of Trainology 2019, 8(1), 5–8. [Google Scholar] [CrossRef] [PubMed]
- Burd, N. A.; et al. Enhanced amino acid sensitivity of myofibrillar protein synthesis persists for up to 24 h after resistance exercise in young men. Journal of Nutrition 2011, 141(4), 568–573. [Google Scholar] [CrossRef] [PubMed]
- Coffey, V. G.; Zhong, Z.; Shield, A.; Canny, B. J.; Chibalin, A. V.; Zierath, J. R.; Hawley, J. A. Early signaling responses to divergent exercise stimuli in skeletal muscle from well-trained humans. The FASEB journal 2006, 20(1), 190–192. [Google Scholar] [CrossRef] [PubMed]
- Columbu, F. Winning Bodybuilding [Audiobook]; Creators Publishing. Audible, 2016. [Google Scholar]
- Fisher, J. P.; Steele, J.; Smith, D.; Gentil, P. Periodization for optimizing strength and hypertrophy; the forgotten variables. Journal of Trainology 2018, 7(1), 10–15. [Google Scholar] [CrossRef] [PubMed]
- Goldman, A. L.; Pope, H. G., Jr.; Bhasin, S. The health threat posed by the hidden epidemic of anabolic steroid use and body image disorders among young men. The Journal of Clinical Endocrinology & Metabolism 2019, 104(4), 1069–1074. [Google Scholar]
- Haugen, M. E.; Vårvik, F. T.; Larsen, S.; Haugen, A. S.; van den Tillaar, R.; Bjørnsen, T. Effect of free-weight vs. machine-based strength training on maximal strength, hypertrophy and jump performance–a systematic review and meta-analysis. BMC Sports Science, Medicine and Rehabilitation 2023, 15(1), 103. [Google Scholar] [CrossRef] [PubMed]
- Hoseini, R.; Hoseini, Z. Exploring the prevalence of anabolic steroid use among men and women resistance training practitioners after the COVID-19 pandemic. BMC Public Health 2024, 24(1), 798. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D. C.; et al. Adaptations to endurance and strength training. Frontiers in Physiology 2017, 8, 537. [Google Scholar] [CrossRef] [PubMed]
- Israetel, M.; Hoffmann, J.; Smith, C. W. Scientific Principles of Strength Training. In Juggernaut Training Systems; 2015. [Google Scholar]
- Joanisse, S.; McKendry, J.; Lim, C.; Nunes, E. A.; Stokes, T.; Mcleod, J. C.; Phillips, S. M. Understanding the effects of nutrition and post-exercise nutrition on skeletal muscle protein turnover: Insights from stable isotope studies. Clinical Nutrition Open Science 2021, 36, 56–77. [Google Scholar] [CrossRef]
- Kassiano, W.; Nunes, J. P.; Costa, B.; Ribeiro, A. S.; Schoenfeld, B. J.; Cyrino, E. S. Does varying resistance exercises promote superior muscle hypertrophy and strength gains? A systematic review. The Journal of Strength & Conditioning Research 2022, 36(6), 1753–1762. [Google Scholar]
- Lopez, P.; Radaelli, R.; Taaffe, D. R.; Newton, R. U.; Galvão, D. A.; Trajano, G. S.; Pinto, R. S. Resistance training load effects on muscle hypertrophy and strength gain: systematic review and network meta-analysis. Medicine and science in sports and exercise 2020, 53(6), 1206. [Google Scholar] [CrossRef] [PubMed]
- Fitness, OX. Stimulus-Recovery-Adaptation (SRA) principle explains how training frequency can affect our progression [Image embedded in Facebook Post]. Facebook. 2021. Available online: https://www.google.com/url?sa=i&url=https%3A%2F%2Fwww.facebook.com%2FOxFitnessAu%2Fposts%2Fstimulus-recovery-adaptation-sra-principle-explains-how-training-frequency-can-a%2F486189512775757%2F&psig=AOvVaw37XRvVIEYjSn4PHYEg9GBI&ust=1758850324827000&source=images&cd=vfe&opi=89978449&ved=0CBkQjhxqFwoTCJCx9Zji8o8DFQAAAAAdAAAAABAM.
- Moore, D. R. Maximizing post-exercise anabolism: the case for relative protein intakes. Frontiers in nutrition 2019, 6, 485822. [Google Scholar] [CrossRef] [PubMed]
- Morton, R. W.; Murphy, K. T.; McKellar, S. R.; Schoenfeld, B. J.; Henselmans, M.; Helms, E.; Phillips, S. M. A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults. British journal of sports medicine 2018, 52(6), 376–384. [Google Scholar] [CrossRef] [PubMed]
- Rosa, A.; Vazquez, G.; Grgic, J.; Balachandran, A. T.; Orazem, J.; Schoenfeld, B. J. Hypertrophic effects of single-versus multi-joint exercise of the limb muscles: a systematic review and meta-analysis. Strength & Conditioning Journal 2023, 45(1), 49–57. [Google Scholar]
- Saeterbakken, A. H.; Stien, N.; Paulsen, G.; Behm, D. G.; Andersen, V.; Solstad, T. E. J.; Prieske, O. Task specificity of dynamic resistance training and its transferability to non-trained isometric muscle strength: a systematic review with meta-analysis. Sports Medicine 2025, 1–26. [Google Scholar] [CrossRef]
- Sale, D. G. Neural adaptation to resistance training. Medicine and science in sports and exercise 1988, 20((5) Suppl, S135–45. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenegger, A. The New Encyclopedia of Modern Bodybuilding; Simon & Schuster, 1999. [Google Scholar]
- Smith, C. W. The Powerlifting Program Design Manual . In Juggernaut Training Systems, Kindle ed.; 2022; Available online: https://www.amazon.com/Powerlifting-Program-Design-Manual-Kindle-ebook/dp/B09TSH226P.
- van Vliet, S.; Burd, N. A.; van Loon, L. J. C. Achieving optimal post-exercise muscle protein remodeling in physically active adults through whole food protein ingestion. Frontiers in Nutrition 5 2018, 60. [Google Scholar]
- Weider, J. Joe Weider’s Ultimate Bodybuilding . In Contemporary Books; 1989. [Google Scholar]
Figure 3.
Short- versus long-run exercise number optimization in the presence of heterogeneous adaptation rates. (a) underlying forces determining effective per-session muscle stimulus as a function of the number of exercises. The downward-sloping curve represents skill dilution: as N increases, each exercise is practiced less frequently, reducing performance due to undertrained skill. The upward-sloping curves represent tendon/constraint relief: spreading training across more exercises allows each tendon to recover more fully between sessions. The intersection of each tendon relief curve with the skill dilution curve indicates the exercise number that maximizes per-session muscle stimulus for the short run (SR, rapid gains, lower N) versus the long run (LR, sustainable gains, higher N). (b) the net outcome in terms of average realized muscle stimulus per session. Each curve corresponds to the short-run and long-run scenarios, with peaks occurring at different exercise numbers depending on the training horizon.
Figure 3.
Short- versus long-run exercise number optimization in the presence of heterogeneous adaptation rates. (a) underlying forces determining effective per-session muscle stimulus as a function of the number of exercises. The downward-sloping curve represents skill dilution: as N increases, each exercise is practiced less frequently, reducing performance due to undertrained skill. The upward-sloping curves represent tendon/constraint relief: spreading training across more exercises allows each tendon to recover more fully between sessions. The intersection of each tendon relief curve with the skill dilution curve indicates the exercise number that maximizes per-session muscle stimulus for the short run (SR, rapid gains, lower N) versus the long run (LR, sustainable gains, higher N). (b) the net outcome in terms of average realized muscle stimulus per session. Each curve corresponds to the short-run and long-run scenarios, with peaks occurring at different exercise numbers depending on the training horizon.
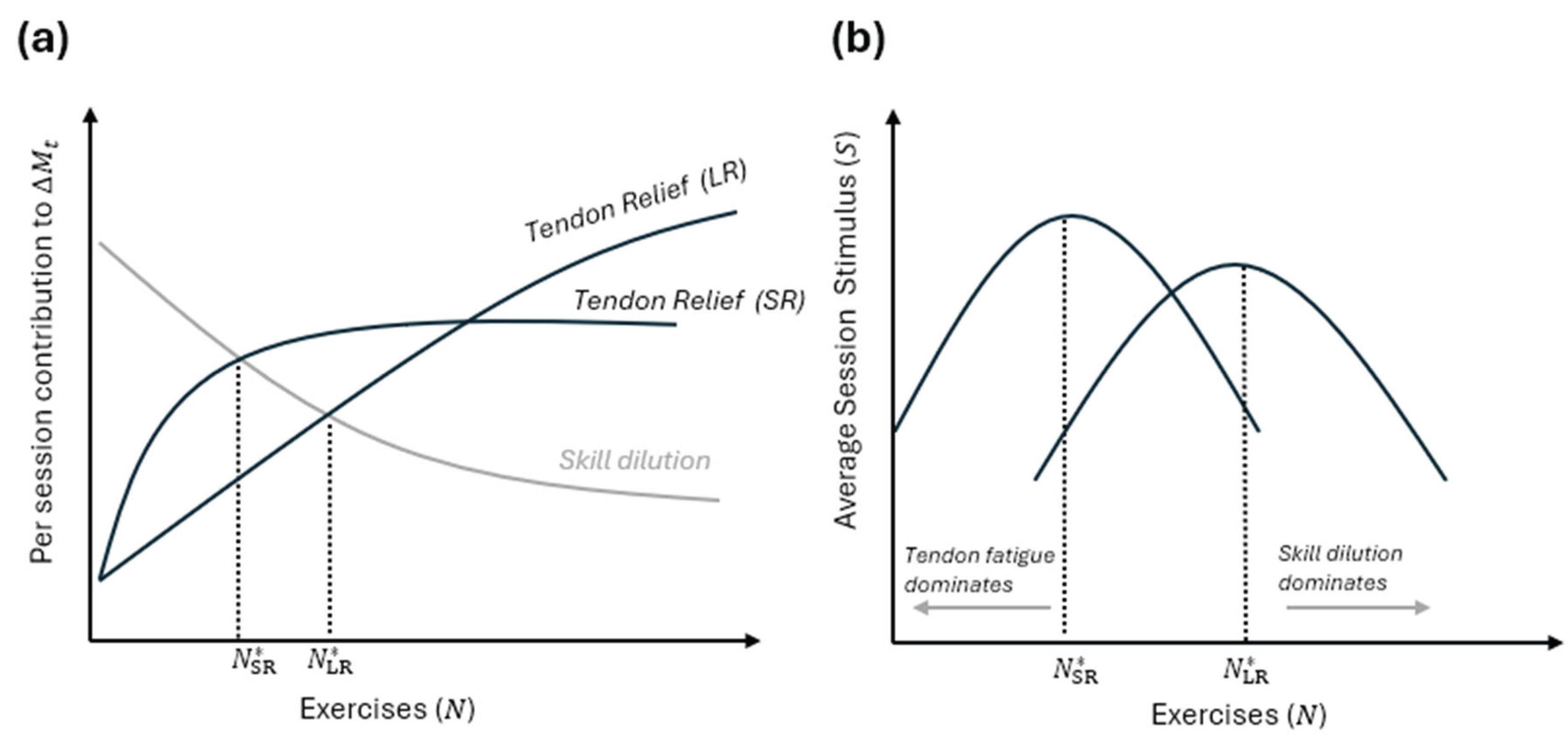

Table 1.
caption.
| Feature | Stimulus-to-Fatigue Ratio | Motor-Clutch |
|---|---|---|
| Role of Constraints | Static Cost (general fatigue) to be minimized | Specific dynamic variables to be managed/trained |
| Strategy | Select exercises with the lowest fatigue per stimulus ratio | Select exercises that complement each other by alternating in their effects on constraints |
| Long-Term Goal | Stay within current recovery limits | Raise the ceiling by training constraints (progressive overload rather than over or under-training them) |
| Variation Logic | To reduce generalized or systemic fatigue | To bypass technical and connective bottlenecks |
Note. High level summary contrasting the Motor Clutch Model with contemporary and popular alternative.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



