Submitted:
29 January 2026
Posted:
12 February 2026
Read the latest preprint version here
Abstract
Kin-Selection has traditionally been viewed as a stabilizing force explaining when, and to what degree it can be adaptive for benefits to be shifted among relatives. As such, it is not usually considered a potential accelerant of evolutionary change. Yet benefits conferred on relatives — particularly from parents to offspring — may function more as forward-looking investments than simple transfers. When seen in this light, Kin-Selection emerges as a potential catalyst for rapid phenotypic change. Drawing on concepts from both evolutionary biology and economics, this article introduces the concept of “fitness capital”— traits enhanced by altruistic investment that increase an individual’s capacity to acquire resources for further investment — and shows how overlapping generations and assortative mating can generate cascading, multigenerational effects. These dynamics amplify correlations between parental altruism, offspring ability, lineage-level investment, and traits reflecting fitness capital, creating conditions for runaway increases in both altruism and capacity to invest. By recasting kin-directed altruism as a driver of cumulative social and evolutionary change, the proposed synthesis highlights a novel pathway through which populations may evolve greater investment, social complexity, and phenotypic stratification. It also indicates new perspectives on parent-offspring conflict, costly signalling, sexual selection, and Green-Beard effects.
Keywords:
kin-selection
; human capital
; inclusive fitness
; parental investment
; altruism
; mate choice
; assortative mating
; green beard
1. Introduction
Selfish genes sometimes prosper by promoting compassion. As Hamilton showed, a gene can spread by making its bearer act altruistically toward relatives, provided the cost to the actor is outweighed by the benefit to the recipient — if the benefit is appropriately discounted by relatedness. The simplest version of this logic, known as Hamilton’s rule, established the concept of inclusive fitness (Hamilton, 1963; 1964). This proved to be one of the most powerful extensions of Darwinian thinking in the modern era (Dawkins, 2004), giving rise to the genes-eye-view of evolution (Dawkins, 1996; Hamilton, 1996). Kin-Selection is a major mechanism for explaining why altruism can evolve in a competitive world otherwise red in tooth and claw and has illuminated phenomena ranging from patterns of social organization such as the evolution of eusociality (Bourke, 2011) to intragenomic conflict (Haig, 2001). However, its role is usually cast as stabilizing rather than as a factor in runaway evolutionary dynamics. Explanations for the rapid evolution of extreme traits more often invoke sexual selection (Fisher, 1930), arms races (Dawkins & Krebs, 1979), or a dramatic environmental event such as causing the splitting of populations by geographic isolation (Eldredge & Gould, 1972). The present article reinforces the argument that, under certain conditions that are likely satisfied in many contexts in nature where kin-selected traits have evolved, they can be expected to catalyse rapid phenotypic changes in a certain type of trait (Hawkes, 1983).
That type of trait is one that can be modified by altruistic forms of investment wherever such modification enhances the capacity of the individual to, in turn, invest and thereby improve the same trait in others. Evidence of such investment is widespread. Many varieties of birds, for instance, provide sustained care for their relatives in ways that enhance their future reproductive success, usually in the form of food provisioning (e.g., McKinnon et al., 2024). In many types of mammals, such as dolphins, impressive hunting tactics and techniques are transmitted from parent to offspring, with evidence of deliberate instructed practice (e.g., Krützen et al., 2005). In primates, maternal care, grooming, and coalitionary support create durable advantages for offspring in social hierarchies (Silk, 2007). In humans, parental choices about nutrition, education, and socialization may systematically shape children’s life chances, with effects potentially multiplying across generations (Sowell, 2016). Across these and many other species, altruism is often not an act with transient effect on the recipient’s wellbeing but may be seen as a strategic allocation problem with cumulative consequences.
Below we consider that, under special (but psychobiologically plausible) conditions, investments in kin have the potential to cascade across overlapping generations, accumulating like a form of “fitness capital” within lineages. To explore this, we develop a unified framework linking Hamilton’s Kin-Selection logic with Becker’s theory of parental investment from economics. Here, the present thesis draws heavily on analyses and concepts from Becker (1993; 1996), notes taken from courses he taught at the University of Chicago titled “Lectures on Human Capital” and “Chicago Price Theory”, and accompanying textbooks (Friedman, 1962; Jaffe et al., 2019). This integration suggests how parental allocation decisions can generate strong correlations between ability, altruism, and fitness capital, particularly under assortative mating. Moreover, these forces may produce runaway increases in investment, stratification of lineages, and even set the stage for evolutionary transitions often attributed solely to other mechanisms, such as sexual selection for “good genes” or ornamental traits. By reframing Hamiltonian altruism as a mediator of intergenerational investment, it is concluded that Kin-Selection is not merely a force favouring some calibrated level of beneficence towards relatives but also potentially a powerful accelerant of evolutionary change.
2. Investment and Fitness Capital
We begin with three premises. First, we assume a species in which Hamilton’s logic already applies. That is, one where individuals exist in a system where genes for altruistic behaviours can persist if the benefit they confer on relatives, weighted by relatedness, exceeds the cost to the actor. Second, we extend the notion of “benefit” to include not only direct gains to a recipient’s fitness but also indirect gains, such as improvements in traits that enhance the recipient’s later success, inclusive of the capacity to confer similar benefits on others. This is intended as the evolutionary analogue of the economic concept of human capital — characteristics like skills, knowledge, health, or attractiveness that increase an individual’s productivity (Becker, 2002). Human capital is a sort of “capital” because it can be built up through investment (e.g., training, nutrition, education) and because greater stocks of it tend to yield higher returns, whether in allowing higher wages to be earned or by enhancing wellbeing through richer consumption opportunities (e.g., being able to read allows more benefit to be derived from books). Finally, we assume that there is an appreciable, additive genetic variance directly determining every characteristic described (e.g., altruism) except fitness capital.
For present purposes, we generalize this idea beyond humans. For any species, we may define a stock of fitness capital as characteristics that improve an animal’s ability to secure resources, survive, and reproduce. Just as in economics, fitness capital can be increased by investment and affects the individual’s future wellbeing and fitness. Fitness capital can be manifested through both direct and indirect parental effects. Maternal effects and transgenerational plasticity provide direct pathways: nutrient provisioning, epigenetic modifications, or early-life experiences can enhance offspring survival, growth, and reproductive potential (Marshall & Uller, 2007; Mousseau & Fox, 1998). More broadly, niche construction illustrates how parents and lineages modify environments (physical, social, or cultural) in ways that increase the returns to offspring traits over time. Examples range from cooperative breeding networks in birds and mammals to the transmission of knowledge and tools in humans (Laland et al., 2016; Odling-Smee et al., 2003). Together, these mechanisms show that fitness capital is both genetically and environmentally mediated.
2.1. Fitness Capital and Effective Relatedness
Becker’s models of altruism provide a useful starting point for our present purposes. He assumed that an individual’s utility depends not only on their own consumption but also on the wellbeing of others they care about – especially their own offspring. Consider a simple scenario where each adult has only one child and an income , which can be spent either on their own consumption, , or on investment, , in their child’s fitness capital. Then:
Where is the parent’s utility from consumption, is the benefit to the child from investment, and is an “altruism parameter” approximating the parent’s concern for the child’s wellbeing. Since , the parent’s first-order condition for optimal investment is found by differentiating with respect to :
Using the chain rule for the first term:
The derivative of the second term is:
Setting the first order condition to zero for a maximum
Or more simply
(1)
This means that parents invest until the marginal utility cost to themselves equals the weighted marginal benefit to their child. Hamilton’s rule provides a biological analogue in that it argues altruism may evolve if:
Or, put in marginal terms,
Where is relatedness, is the marginal gain in offspring fitness, and is the marginal cost to the parent’s fitness. These ideas can be unified if we treat Becker’s altruism weight as equivalent to relatedness , if utility is measured in the same “currency” as fitness. To make map in this way, let us introduce as a conversion factor which scales marginal utility into marginal fitness. This acknowledges that not all parental investments that feel rewarding (in utility terms) translate perfectly into genetic returns. Some activities may raise subjective wellbeing without affecting survival or reproduction, while others may have strong fitness consequences. Formally, let the marginal fitness cost of reducing parental consumption be:
Substituting into (1) gives:
.
Thus, the effective relatedness governing altruistic investment is
This captures both the biological fact of genetic relatedness () and the psychological or economic fact of how strongly a given investment in the child’s wellbeing translates into fitness outcomes1.
In sum, the Becker and Hamilton perspectives are likely looking at related phenomena from different perspectives. The two can be bridged with the concept of fitness capital and a scaling factor for translating utility into fitness. This permits us to say that, if there are forms of fitness capital that can be enhanced through investment, then parental altruism need not operate as only a direct transfer of benefits to offspring (or other kin): for example, shifting consumption from the parent to consumption by the child. Instead, parents may weigh against their own consumption the opportunity to invest in ways that improve their children’s underlying traits or capacities. This means that investment decisions should scale with both genetic relatedness and the reliability of wellbeing cues as predictors of fitness outcomes. Put another way, this distinction is the difference between giving a fish to eat and teaching how to fish: parents may not be motivated merely helping offspring now — they could be responding to cues that help them to build the fitness capital in their offspring and yield higher fitness later2. Thus, more altruistic parents will tend to invest a higher fraction of their budget on their children, and this could be favoured if this investment is adjudicated appropriately given effective relatedness.
2.2. Overlapping Generations and the Legacy of Altruism
An important question in economics is “and then what happens?”. This is because the short- and long-run impact of decisions can be quite different, whether in magnitude, direction, or both. Answering this question brings us to the heart of the present thesis, and to do so we follow Becker (reference). Let the child’s fitness capital during the next time period be described by a production function:
(2)
Where is parental investment and is the child’s baseline level of fitness capital. This is to say that describes how an individual’s eventual fitness potential emerges from the interaction between what it starts out with and with how much this is boosted by their parents’ investment. More investment always helps, but there are diminishing returns. Each additional bit of investment matters less than the last bit3. When adult, the child’s “income” then follows from its fitness capital, such as:
Where can be thought of as a “wage rate” that represents the strength with which capital determines income. Or, more generally, income may be thought of as any increasing function of . Either way, we are assuming that differences in the total budget that individuals earn to then spend on their own consumption and investment are only explained by differences in capital. This means that parents with more income are generally going to invest more income in their offspring, even if altruism is equal. So, variation among individuals in income at any given time is here determined by their developed capabilities which in turn flows from differences in the capital of their parents (Figure 1).
We also assume inter-individual differences in how altruistic parents are and that they pass on their altruism to offspring:
Where represents genetic and potentially epigenetic and cultural transmission of altruism, such as with representing the heritability or persistence of altruism across generations, ranging from zero to perfect, .
The parent’s optimization problem at time is to spend their budget across consumption and investment in a way that maximises their utility. So, selecting the amount to investment in offspring is and optimization problem of the form:
We differentiate with respect to Using the chain rule for the first term the derivative of is times the derivative of , which is . For the second term (), we treat as a constant with respect to . Thus, the first order condition is:
Which, if we set equal to zero () to find the optimum gives:
(3)
Equation (3) states that the marginal utility lost from foregoing a unit of personal consumption, , equals the marginal altruism-weighted gain from investing that same unit in the offspring, . At equilibrium, the parent allocates its resources sot that these terms are balance. The key implication is that, at any given period (e.g., ), a parent will invest more if they are more altruistic (), if they have higher capital themselves (), and if their own parents invested more () in them (because that increased their ). Lineages of more altruistic parents therefore invest more, and the more that they invest, the more capital their children accumulate. The more capital their children have, the larger their future incomes and budgets will be for investment in their own offspring (higher leads to greater ). Hence, rising capital of subsequent generations is driven by increases in equilibrium investment and capital of the preceding generation. With equation 2, this dynamic produces a positive feedback loop:
With lineages with higher altruism systematically choose higher investment, generating children with greater human or fitness capital, and transmit above-average altruism (Figure 2). This induces a positive correlation between altruism and capital, both in cross-sectional snapshots and over time. This is a Becker-based insight, but it can be reinterpreted in Hamiltonian terms by translating utility into fitness units and altruism anchored in effective relatedness.
If parental investments produce benefits that can compound across multiple generations (investment in a child increase their capital and so raises the fitness of the grandchildren they produce), then the discounted sum of benefits flowing from investment will be larger than that of any single period. Put in marginal terms, this is equivalent to multiplying the right side of equation 3 by some variable representing the present value of these future benefits. To express this as a symbol, suppose that parent values their child’s fitness with a discount factor or , which we will assume is positive. This discounts the effect on utility of a parent from the fitness of each generation separated from the parent in time when he or she makes their investment decision4. Then parental utility is
And maximised when
So, in “fitness units”, this can be written as
With the Hamilton weight being
Thus, a single-period Hamiltonian calculation may significantly under-predict investment when multi-generational returns are large. Greater overlap between generations (less discounting or higher ) tightens the link from Becker’s economic framework where individuals are concerned about the impact of their current decisions on posterity to Hamilton’s biological one. The more that a parent’s actions today influence their child’s future reproduction and survival, the larger the effective Hamiltonian weight. We may think of this as a parent’s inclusive-fitness horizon: the further ahead they look (e.g., valuing not only their child’s fitness a little but a lot, as well as additionally being concerned with that of their grandchildren, and beyond), the greater the effective overlap and the stronger the incentive to invest becomes. In short, the same first-order conditions subsume Becker’s altruistic investment with Hamilton’s inclusive fitness logic: ,. Overlapping generations, through the discount factor , extend the weight parent’s place on the benefits of current investment for future fitness. This predicts a positive coevolutionary correlation between altruism and fitness capital, given transmission of altruism may also be genetically based and diminishing returns in .
The alert reader may notice that, although this model tracks one cohort per period, it is effectively an overlapping generations system in the causal sense. This is because each parent’s investment in and influences the reproductive capacity of descendants. Terms like “overlapping generations” and “lineage-level feedback” refer to causal persistence across generations, not literal simultaneous co-occurrence of multiple cohorts. The geometric sum in quantifies this cumulative inclusive fitness impact, reconciling simple recursion with biologically interpretable and meaningful multigenerational feedback. For readers preferring the overlapping-generation logic to be fully explicit, a minimal model is presented in Appendix A. This demonstrates the same first order conditions and that intergenerational feedbacks naturally arise through recursive inheritance of capital and altruism.
2.3. Ability, Altruism, and Investment
Becker (1991) extended his framework by considering the possibility that parents invest in their children according to the efficiency with which each child translates that investment into future capital. Even if parents are equally altruistic to each child, they are not blindly egalitarian but may rationally pattern their investments among children in ways calibrated according to how they differ in the rates of return they provide. The same idea can be applied in an evolutionary framework. Sexual reproduction and genetic variation mean that offspring can differ in their intrinsic capacities for translating parental investment into traits related to fitness. Some individuals may grow faster, achieve greater size, be more resistant to parasites, or demonstrate greater capacity to learn skills and use knowledge. If parents can detect and act on cues for these prospective differences, we should expect them to make the necessary adjustments. Although Becker did not originally consider its interaction with altruism, we can extend the production function described in 2.1 like so:
when Where remains parental investment, is the offsprings baseline endowment of fitness at birth (e.g., health, size, immunity), and represents the offspring’s ability to convert investment into fitness (essentially a multiplier of parental efforts). In words, ability acts like an “efficiency weight”. If two siblings, one with ability and the other , each receive the same amount of, say, food (), but , then the sibling with will translate food into more reproductive potential. The utility function of the parent who must choose how they will invest in two children with differing ability, and , is:
Assuming that the altruistic coefficient is the same across all children, and given our assumptions, if parents invest until the rate of returns in each child are the same then that will imply that (unless progressively more investment produces sufficiently drastic diminished or negative returns5). Or, supposing parents have offspring, a fixed budget of investment resources, they must decide how to allocate their resources across children:
all subject to Where is the ability of child . Taking the first order condition with respect to (investment in child ):
Since and , we get:
Which implies that, for every child :
For two children, and , the optimal allocation requires that:
Assuming altruism is constant:
As , if, say, , then to make marginal returns from each child equal implies (unless returns to investment fall so fast that the equality is reversed). This means that the optimal allocation will generally favour children with higher ability because the marginal benefit of investing in them (in terms of future fitness capital) is greater even if parents are equally altruistic towards each of their children (see Figure 3). The important point here is that ability does more than simply arrange patterns of investment within families. It can accelerate the overall process of capital accumulation. It can be expected to do this through two channels. The first may be referred to as “Roy sorting amplification” (after Roy, 1951). Because the marginal return of investing in a higher ability child will remain higher than the cost for longer, parents can achieve higher levels of returns (and smarter children overall) if they invest accordingly. If parents preferentially invest in offspring who appear to have higher ability, then those children will accumulate disproportionately more capital over time. This feedback loop produces a self-reinforcing sorting process, where high-ability individuals become both the most invested-in and the most capital-rich (so have the most to invest).
The second is related to the complementarity between ability and investment. If ability not only raises returns to investment but also amplifies the effectiveness of already accumulated capital, then ability and capital interact multiplicatively. For example, perhaps a child who is cognitively gifted and learns faster than expected also gets taught more, and if they are taught more then they learn faster. This produces accelerating differences in outcomes across siblings within and between families.
Figure 4.
Hypothetical relationship between parent and offspring characteristics highlighting the role of ability. Ability is integrated with the same relationships depicted in Figure 2. Offspring tend to inherit ability from parents, while the child’s ability both enhances the effect of the parental investment and induces more of that investment. Red arrows depict genetic transmission from parent to offspring, solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. Red arrows depict genetic transmission from parent to offspring, solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. .
Figure 4.
Hypothetical relationship between parent and offspring characteristics highlighting the role of ability. Ability is integrated with the same relationships depicted in Figure 2. Offspring tend to inherit ability from parents, while the child’s ability both enhances the effect of the parental investment and induces more of that investment. Red arrows depict genetic transmission from parent to offspring, solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. Red arrows depict genetic transmission from parent to offspring, solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. .
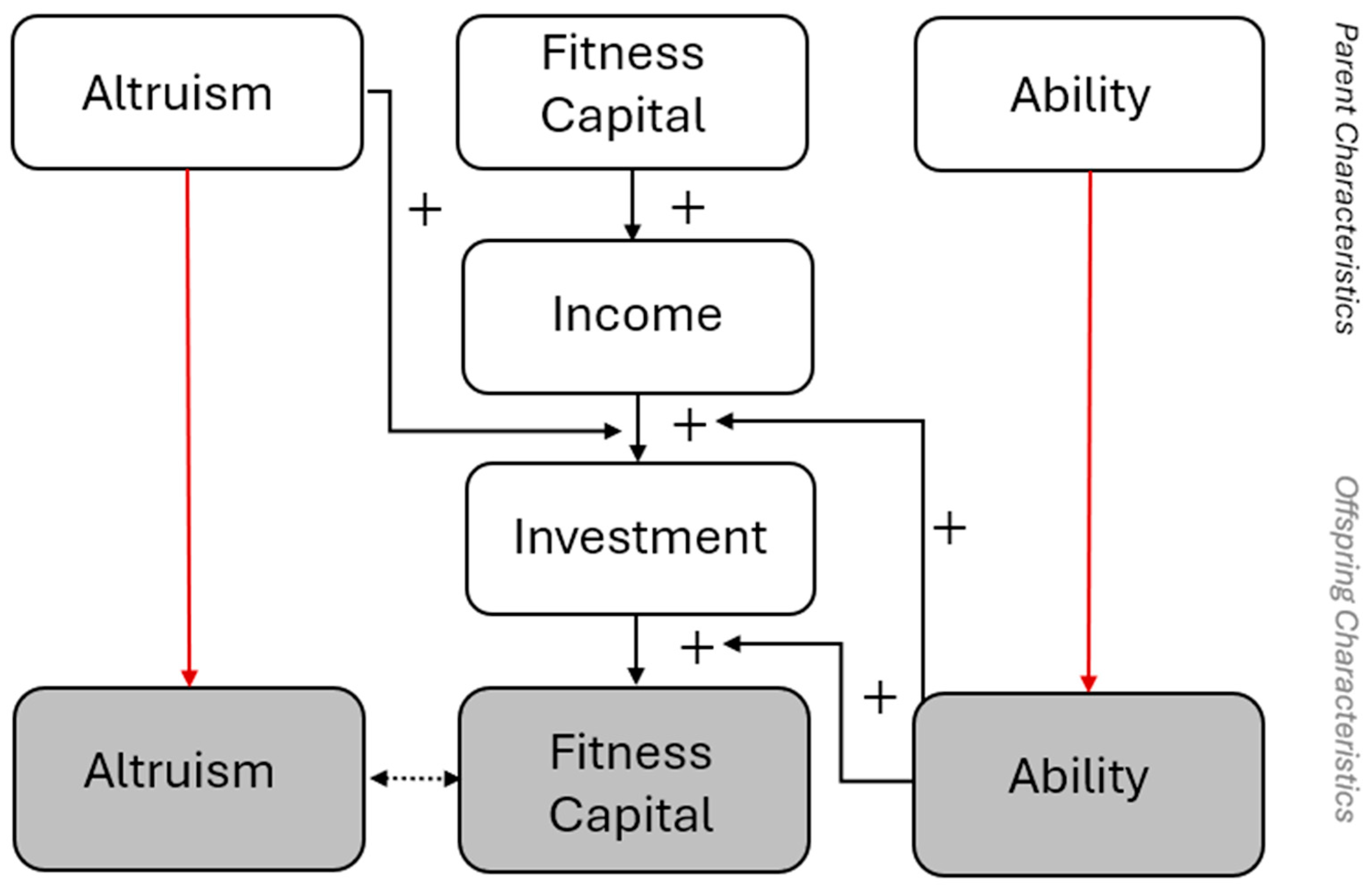
Together, Roy sorting and complementarity mean that ability, altruism, and capital can be expected to become rapidly correlated within the population. In other words, signals of higher ability draw more investment, and the higher ability amplifies the effect of that investment. If ability and altruism are heritable, then parents who are more altruistic towards higher ability children will generate lineages where ability and capital accumulate together, and these lineages will in turn reinforce the apparent payoff to altruism.
In summary, adding ability to the model generates the following predictions: (1) parents should preferentially invest in children who signal higher ability, (2) because ability accelerates the accumulation of capital, even small initial differences may snowball into larger disparities over time, and (3), this creates a natural correlation between ability, capital, and the expression of altruism. The strength of the correlation between ability, investment, and fitness capital can weaken if abilities are poorly detectable by parents, if investment has very strong diminishing returns, or if environmental shocks randomly reduce the translation of investment into fitness. Nevertheless, it seems reasonable to conclude based on reasonable assumptions that families that hit upon this alignment will tend to flourish disproportionately.
2.4. Parenting Synergies and Mate Choice
We have so far examined how altruism and fitness capital can reinforce each other across generations through altruism’s directing of parental investment. A further channel may come from mate choice. The more impactful that decisions become about how to invest in offspring, the weightier the consideration of who to mate with becomes beyond just their genetic contribution. In any species where individuals invest in kin in ways receptive to the sorts of feedback we have described, such as where it is rewarding to see relatives do well, it is unlikely that mates are chosen at random with respect to traits such as altruism or the capacity to obtain and command resources (whether through “good genes”, markers of fitness capital, or both). In a variety of animals, investments in children are also not made solely by one parent in isolation but by both, and these investments plausibly interact to produce the fitness of the offspring that result.
Becker (1973, 1974) analysed how human capital shapes partner choice in marriage markets. We can build on this framework, linking it explicitly to altruism as considered so far. Following Becker’s treatment of marriage markets, consider a model in which each male and female adult has two key traits. Firstly, they vary in their altruism, denoted by for individual (where means male and means female). As before, this measures how much weight they put on their child’s wellbeing relative to their own consumption. Secondly, they have “fitness” capital . A “household” (i.e., marriage or cooperative breeding pair) forms between two adults, () and (), to produce a combined income:
Where is a non-decreasing function reflecting the productive complementarity between reproductive partners.
If we think of the household as the decision-making unit, the effective altruism of it will be:
, For instance, this might be a simple average, or a function that gives extra weight to couples who are both altruistic. The couple then must decide how much of their income to consume themselves and how much to invest in their offspring :
Where is the utility from consumption, which is constrained by the budget , and is the payoff to the offspring from parental investment, which can be measured in the same units as utility or fitness as discussed 2.1. Assuming parents both consume much the same under optimal conditions () for simplicity, the first order conditions are:
This means that the marginal cost of giving up consumption equals the marginal benefit of investing in the child, scaled by the altruism of the breeding pair. Meanwhile, the child’s human capital during the next period will be
where is increased by investment to capture the idea that returns on investing in the child are higher when parents have more income and when they are more altruistic. This implies that, at equilibrium, higher household altruism or parental capital leads to greater investment and higher offspring capital.
We next assume “supermodularity” between male and female characteristics. In other words, their joint efforts of each parent are force multipliers of one another, particularly their fitness capital:
If the household income produced by a breeding pair is greater than the sum of its parts, this makes it rational for males and females to seek each other out and mate based on matching features6. Such positive assortative mating (hereinafter PAM for brevity) on would then raise the correlation between and , hence raising income variation and the average income of the most productive households7. At equilibrium, this generates correlations between partners’ traits and increases the variance and average of household income. Since equilibrium investment rises when income increases, higher- couples invest more, so of the children rises too via this budget pathway.
It is important to note, however, that unless and are already correlated, positive assortative mating on would not directly raise . If how altruistic parents are is independent of their capital, the distribution of altruism throughout the population should remain unchanged. By contrast, suppose partners choose their mates on the basis of altruism (i.e., how much they expect their partners will care about investing in their jointly produced offspring). This is not a difficult assumption to motivate: an individual wanting to spend more on their children than consume themselves would likely want to find a like-minded partner to mate with so that there is less conflict about how the household budget is allocated. Meanwhile, relatively selfish couples would be happier together because that produces more income to spend on themselves. This would directly make and positively correlated, and so there will be a higher average and higher variation in household altruism than random mating. Because the optimal investment in offspring rises with higher household altruism, high- couples invest disproportionately more of their income, raising their children’s even if their incomes remain the same. As we are assuming transmission of altruism (is an increasing function of ), then high- couples produce children with more altruism and more capital. In other words, if couples prefer to mate based on altruism in one generation, then this will create and/or increase the correlation between altruism and capital in the next generation. Alternatively expressed, let the covariance between household altruism and offspring fitness at generation be:
This can be expressed in an equation of the manner pioneered by Price (1970)8 as follows,
Where positive assortative mating based on altruism makes and so increases the correlation (this feedback process is depicted in Figure 5), while positive assortative mating based on capital only affects , whose sign already mirrors whatever correlation already exists between altruism and human capital. If mates select each other on both altruism and human capital (or ability), then this should produce the most rapid stratification of human capital within the population, leading to extreme combinations of all traits.
This feature opens the door to an important dynamic in evolutionary biology, originally described by Fisher (1930). Whereas the Becker model proposes people mate for reasons of supermodularity that make seeking out partners with similarly high levels of ability, capital, or (as we have considered, altruism) because it is optimal from the standpoint of utility, once there is a correlation between altruism and preferences for altruism, this sets the stage for potentially runaway sexual selection. To see how, consider Lande’s (1981) model of the joint evolution of a traits with preferences, e.g., trait and sexual preferences for that trait in partners . From Lande’s analysis, if both have a genetic basis, and these are positively correlated, , positive feedback can ensue when:
Where represents is genetic variance for trait , is the strength with which preference () for trait confers a mating advantage to individuals higher in this trait, and refers to the strength of stabilizing selection pushing back towards some optimal level. So, the larger the genetic covariance is between trait and preference, the more that selection favouring an increase in the trait also increases strength of preference for that trait, and vice versa. Larger makes sexual selection stronger for a given level of covariance between trait and preference (), a larger increases the required needed for runaway to occur. When a trait variance is large relative to covariance with preference, stronger sexual selection pressures are needed to build the coupling. Strong natural selection also resists exaggerated . This builds the covariance between preference and the trait, leading to exaggerated traits and stronger preferences. As with the model so far considered, Lande-like runaway selection does not in itself require new genetic variation to arise: the runaway process comes from restructuring the patterns of correlation (linkage disequilibrium) between preference and trait throughout the population, increasingly concentrating genes for extremes in these features in more extraordinary individuals (Dawkins, 1976).
In summary, if fitness capital, ability, or altruism have a strong genetic component, they may quickly become genetically correlated through the investment and mating decisions of parents. If these mating decisions are made based on supermodularity or utility-maximising considerations (e.g., choosing a wife or husband because they are altruistic), any innate sexual preference for any of these traits (e.g., finding a wife or husband sexually attractive because they are altruistic) could in turn drive runaway evolution of these correlated traits. In other words, mating initially based on supermodularity may be a pump-priming or ignition system that gets the engine of runaway sexual selection of these traits going (Figure 6), and this may extend the evolution of these traits to beyond what would otherwise be optimal. Likewise, a preference for partners based on “good genes” (if a marker for ability), could fan the flames of marriages motivated by supermodularity of altruism or human capital as these would become correlated through the rational investment decisions of preceding generations.
It is worth emphasizing that the present analysis follows the classic approach to runaway sexual selection, as pioneered by Fisher (1930) and formalized by Lande (1981). In this tradition, the focus is on identifying conditions under which traits and preferences can covary positively across generations, generating self-reinforcing feedbacks, rather than on explicitly iterating full generational dynamics. In our model, these feedbacks are captured by the covariance between parental altruism and offspring fitness capital (). PAM on altruism ensures , demonstrating a mechanism by which more altruistic households produce children with both greater altruism and higher fitness capital. Similarly, PAM on fitness capital influences , amplifying variation in household investment potential. As in Fisher and Lande, these equilibrium-based covariances indicate the potential conditions for accelerating correlations between traits and preferences, without asserting that trait divergence is inevitable or unbounded. In this sense, our “runaway” description refers to self-reinforcing increases in correlations across generations, consistent with the logic of the locus classicus of this literature (to see an example of how these conditions could be explicitly formalized for mate choice and altruism complementarity generate covariation, see Appendix B).
To round off this discussion of mate choice, it is worth addressing a point related to the nature versus nurture determination of fitness capital. A perhaps intuitive objection to the above is that parents may often use investment to compensate in children lacking good genes. For example, if there is a sibling that requires more resources to grow to the same size than another, redirecting some of the food provided by the parents from the latter towards the former may be wisest – especially if there are diminishing returns to such investment as intensified by the effect of ability. There might be important implications for mate choice dynamics hinging on whether investment and ability generally function more as substitutes in this way or as complements (i.e., where more ability leads to higher returns on investment, as assumed so far). If and parents match based on altruism, high-θ households make larger investments, and target more of this investment to higher- children. Selection will then favour genotypes carrying both high and high , leading to positive covariance building between altruism, ability, and fitness capital. If instead extra investment helps low- offspring catch up more than it propels high- individuals, potentially weakening or even reversing the correlation between altruism and ability. We may still expect higher levels of fitness capital to accumulate in high altruistic yet low ability lineages, so the correlation between and capital may remain positive, but the correlation between altruism and ability would be lessened or even become negative.
If instead parents match directly based on ability – as many models of mate choice assume they do if this is a stand in for “good genes” (Hamilton, 2001)– then we should find good genes concentrated in high- households. If , such that even large investments earn bigger returns in high- individuals, then it will be rational for even parents with moderate altruism to bias their investment in high- offspring. That selection on returns should pull altruism upward in high-a lineages because high ability offspring benefit most from more altruism. This should generate a positive linkage over time between ability and altruism despite the fact mates are choosing each other solely based on good genes. If instead , high levels of investment are not needed as much in high- lineages to reach high levels of fitness capital. So, selection pressure on altruism is weakened within those lineages. This may still result in a strong correlation between ability and fitness capital, a weak altruism-fitness capital link, and perhaps a negligible or negative correlation between ability and altruism.
Finally, if partners choose based on manifest fitness capital, , then this is effectively mating based indirectly on whatever trait mix produces high fitness capital. If , then, this means there is a straightforward route towards a positive three-way correlation between altruism, ability, and fitness capital. If instead either cues signalling capital function as a direct readout of ability or there are such strong diminishing returns to investment that , then PAM on manifest could weaken or slow linkages between altruism and human capital because altruism is less important or negligible. More realistically, investment and ability are unlikely to be strong substitutes in a species where altruistic investment operates and only compensate for each other partially. Even if is only mildly positive, we would expect selection, then, to favour ability but drive the system towards a positive correlation between ability and altruism through greater returns on investment in higher- offspring.
2.5. Altruism-based mating and Green Beard Effects
While the processes considered so far increase correlations between altruism and investment they do not, by themselves, extend altruistic behaviour to levels beyond what would be considered adaptive based on a sufficiently inclusive definition of Hamilton’s rule. However, they do create conditions whereby specific genes may be favoured to push investment to levels beyond what would be advantageous to the rest of the genome based on relatedness between individuals. This is because PAM on altruism may be especially conducive to the evolution of “supergenes” (Hamilton, 1996) or “Green Beards” (Dawkins, 1976). This refers to how a hypothetical gene can be favoured by selection for directing altruistic behaviour of the organism carrying it towards another individual likely carrying the same gene (regardless of the chances the two individuals are likely to share other genes). To evolve and spread, Green Beards require the gene to (1) produce a recognizable effect (the green beard), (2) make its bearers recognize that trait in others, and (3) cause its bearer to preferentially direct benefit to those individuals (Dawkins, 1982).
We have assumed genetic variation influencing willingness to invest in offspring (altruism) and later supposed that this may become linked to genetic variation for preference to mate with individuals who also display cues of altruism towards offspring. If there is a genetic variant that combines (1) unusually high altruism with (2) desiring of unusually high altruism in a partner, then positive assortative mating on altruism could reinforces their connection and spread. In other words, instead of a literal green beard, such a gene could recognize itself in other bodies through the behavioural or cognitive cues related to altruism and preference for altruistic partners (which we have supposed might exist for reasons already explored). By choosing to mate with an especially altruistic partner and direct a large amount of own investment to the offspring of that, this gene is likely helping itself. Put another way, as far as the result of the genome is concerned, optimal investment in offspring (or the wider kin-network) is determined by . But from the standpoint of the genetic variant for noticeably higher altruism, relatedness with offspring is now approaching one hundred per cent.
Assuming simplified genetics and breeding (haploid and one offspring per mating), we can proceed to illustrate the above point by presuming the qualitative conclusions hold in more complicated models. Consider an allele , which codes for unusually high parental investment in offspring, with an allele , which codes for the non or usual altruistic investment. When an -carrier provides parental help to a recipient offspring, the actor pays the cost (this reduces the actor’s direct reproductive fitness), and the recipient offspring obtains a benefit to their own survival or fecundity9. If we assume baseline random mating in the population gives a frequence of allele. Next, suppose that the offspring inherits the allele of one randomly chosen parent. So, if parents are and , then it has a fifty per cent probability of carrying (and likewise for ); if both parents have , then it certainly inherits , and if both parents have , its probability of inheriting it is zero. This means mating with another -carrier increases the probability of offspring carrying .
Next, imagine parents select mates via preference (the sexual green beard) such that carriers of a preference allele (which we will assume is tightly linked to or comes part of the same package by inheritance) prefer to mate with partners who they judge to also be good parents – that is, parents carrying allele . Let’s say, then, that the probability that an -carrier mates with another -carrier partner is
Where is the how much more probable it is above some baseline frequency that an meets and matches with another (this may be thought of as an assortment coefficient). If mating were random, then .
We may expect the number of -copies in the next generation that are caused by an -carrier’s actions. Spread occurs when the expected gain from altruistic investment exceeds the neutral expectation. We have supposed that the offspring has a baseline fifty per cent chance of inheriting the allele from an -parent, and from this must be subtracted the cost of performing the altruistic act of investment . So, there is a direct lost equal to . The benefit is an indirect increase caused to the offspring’s fitness, . So, ordinarily, the question would be whether this is greater than . But here is the crucial twist: that benefit will more often be delivered to offspring that carry when the actor mates assortatively with another . When both partners have probability of , both parents might have , so the offspring assuredly inherits . When the actor mates with an -partner (), the offspring inherits with a probability of fifty percent. So, the probability that the recipient offspring carries A (conditional on the actor being an -carrier) is:
Therefore, the expected indirect genetic benefit to the actor from increasing his or her offspring’s success by is:
The allele will increase in frequency when the indirect gain is greater than the direct loss, which is:
Which can be multiplied by two to simplify and rearranged to into the Hamilton form:So, the term is the extra inclusive fitness advantage to the allele because carriers preferentially mate with other carriers and invest in joint offspring. This may be thought of as the “sexual green beard bonus”, with the standard Hamilton rule for the rest of the genome applying with . The larger becomes for gene , the easier it becomes for the benefit to outweigh the costs. Even if the benefit of altruistically investing some amount in offspring is insufficient to justify it based on the standard Hamiton rule, may be strong enough to escalate from the standpoint of the altruistic genes if the effect of assortative mating is strong10.
3. Discussion
By connecting Hamilton’s insight with an adjusted Becker’s Human Capital framework, we can see how Kin-Selection may be a more potent driving force of evolutionary change than previously recognized. Central to this proposal are two constructs (1) fitness capital – that is, an adaptive characteristic of an individual that can be built up by investment from others, and which contributes to how much resources the individual can in turn acquire and invest in others – and (2) altruism defined here as the determining willingness to invest in kin. Such investment may constitute anything from simply sourcing and providing infants with more food rather than eating it themselves, to spending time and attention transmitting knowledge (e.g., teaching foraging skills) or even constructing tools or other extended phenotypes to pass on for the benefit their children when they mature. Whether manifest in brain or brawn, the critical feature is that the trait being invested in improves proportional to investment, and that there is potential for the trait to build cumulatively. In a species where parents are motivated to invest altruistically in their offspring, genetic variation promoting altruism will tend to become correlated with prowess in acquiring resources for investment. Lineages that are more altruistic invest more, which causes their descendants to inherit both higher altruism and greater fitness capital. These descendants, in turn, have more resources to invest and a higher propensity to invest in their own offspring, producing cascading investments in inclusive fitness.
Other factors may act as force multipliers of this general effect such as when there the feedback guiding the parental decisions is psychological. Given generations overlap, altruistic parents find it rewarding to observe signs that their children prosper, and so investing efforts can become reinforced accordingly. This extends the time horizons over which investment in a lineage can feedback to affect the motivation of parents, grandparents, and so forth. Another amplifier arises when mate choice is assortative on altruism. A male strongly motivated to invest in his children benefits by pairing with a female who is similarly motivated, and vice versa. Partnerships based on shared altruism increase the value placed on offspring investment and simultaneously command greater resources. As a result, descendants of these pairings inherit both higher altruism and higher fitness capital, further intensifying the cascade.
The decision to match and mate positively based on altruism or capital may initially be purely practical: matching based on either or both likely represent an unbeatable or stable strategy if they are complements in the total “utility” each couple jointly produces because, under such an arrangement, no partner can benefit from splitting and “remarrying” with another willing partner. If there is also genetic variation affecting preferences for partner characteristics indicating or correlated with altruism, then the process or assortative mating would build the correlation between genes for higher altruism and genes for preferring altruistic partners. This produces precisely the conditions that can make mate choice become a self-reinforcing amplifier of the process (i.e., Fisher’s runaway effect), potentially leading to sexual selection for signals of willingness to invest highly in offspring.
Another interesting complication we considered was genes in children affecting how receptive they are to their parents’ investments. We summarized this as “ability” and it also likely accelerates the process of altruism and fitness capital becoming linked. This is because parents should be generally motivated to invest more in abler children even if they value each child the same as this should yield higher overall returns than if all children were invested in evenly or randomly. Although under some assumptions ability and investment may act like substitutes, and ability may act like an eddy current in investment cascades, it seems the most straightforward implication is that ability, altruism, and fitness capital all become positively intercorrelated. This means an increase in selection for or mating advantage of one of these characteristics will induce increase in the other characteristics as well.
The recursiveness of these forces is one of two critical features for understanding the implications of cascading fitness capital. The other is their complementarity in building adaptive fitness. Rank-ordered pairings of the varieties described, such as pairing mates high in the same attributes (whether capital or altruism) or by pairing higher ability with higher investment in the same bodies of offspring, maximizes both the total and average level of the resulting traits, compared to what would be produced from random sorting or negative sorting. Such arranging also increases variation, since pairing large multipliers produces disproportionately large outputs, while pairing smaller multipliers together produces little change. The income is greater disparity between lowest and highest levels of fitness capital across lineages. Thus, although the processes we have described do not assume any genetic change at the loci affecting the traits involved, it can produce large levels of phenotypic change and variation exposed to selection resulting from decisions parents make given the circumstances we have assumed11.
Crucially, the processes described do not presuppose any genetic change at loci underlying the traits. Rather, they generate large amounts of phenotypic change and variation exposed to selection, arising purely from the investment and mating decisions parents make under the assumed conditions. This implies both a higher average level of altruistic effort toward kin and a more unequal distribution of fitness capital than would be predicted by simplistic interpretations of Hamilton’s rule. In nature, how we define “fitness capital” will be decisive for which lineages thrive, and which decline. Thus, although the model shows how phenotypic change can occur without shifts in allele frequencies, in practice genetic evolution will follow wherever greater fitness capital confers a Darwinian advantage. If the advantage is strong, our framework identifies mechanisms by which phenotypic change —whether in fitness capital or parental altruism — could rise explosively, with broad evolutionary consequences.
One such consequence arises from the Fisher-Lande-like dynamics of assortative mating on altruism because these create ideal conditions for a distinctive form of Green Beard effect. When a preference allele is linked to an altruism allele, assortative mating allows carriers to preferentially pair with altruists and invest in those carrying the same gene, with the children carrying the green beard enjoying extra investment from both parents. Mathematically, the impact of this mechanism can be expressed as an additional positive term, added to the usual benefit in Hamilton’s rule. This “assortative mating bonus” allows elevated levels of altruism to spread in situations where conventional pedigree-based relatedness would not be sufficient. In nature, this mechanism may be vulnerable to cheaters, but, unlike other forms of Green Beards, we might expect it to be less vulnerable to recombination splitting the link between the green beard expression and preference or costs of detecting the green beard because, in this case, sexual selection would be continuously acting to mitigate these factors.
Previous research has shown that multi-generational effects, vertical cultural transmission, spatial population structure, and ecological interactions can shape the evolution of social traits and investment behaviours (e.g., Brown et al., 2009; Lehmann, 2008; Mullon & Lehmann, 2017; Van Cleve & Akçay, 2014). For instance, Mullon and Lehmann (2017) examine gene–culture co-evolution in family-structured populations, highlighting how vertically transmitted cultural information interacts with genetically determined learning behaviours. Their analysis demonstrates that vertical transmission can modestly increase adaptive information and constrain evolutionary branching, emphasizing kin structure in cultural evolution. While conceptually related, this framework focuses on cultural knowledge rather than direct fitness capital or altruistic traits, and it does not consider assortative mating or amplification of trait covariance across generations.
Lehmann (2008) investigated the evolution of niche-constructing traits whose phenotypic effects extend beyond the actor’s lifespan. By modelling spatially subdivided populations and the future fitness consequences of environmental modifications, he showed that selection can favour traits influencing subsequent generations. This resonates with our focus on multi-generational feedbacks, but our model emphasizes direct behavioural traits—altruism, parental investment, and mate choice—while avoiding the complexity of environmental or spatial dynamics. This simplification allows clearer predictions regarding trait covariance, amplification of altruism, and potential runaway selection.
Van Cleve and Akçay (2014) explore how behavioral responsiveness, genetic relatedness, and synergistic interactions jointly shape social evolution. Their results illustrate that these factors can produce feedbacks not easily captured by simple assortment indices. While our model shares the goal of understanding interaction-driven trait evolution, it abstracts from ecological or reciprocal dynamics to isolate heritable links among parental investment, mate choice, and offspring fitness capital, providing a tractable mechanism for generating positive correlations and potential runaway dynamics.
The main advantage of our framework is its combination of simplicity, interpretability, and predictive clarity. By linking parental investment, assortative mating, and heritable traits, it predicts: (i) the emergence of positive correlations between altruism and fitness capital, (ii) amplification of these correlations via assortative mating on altruism or ability, and (iii) conditions under which these mechanisms may generate runaway evolution, without requiring complex spatial or ecological modelling. Moreover, by bridging Becker’s economic models of parental investment with biological models of inclusive fitness and sexual selection (Hamilton, Fisher, Lande), it provides a unified perspective on multi-generational feedbacks. In sum, while prior work has explored gene–culture co-evolution, social synergy, and niche construction, our model identifies a parsimonious pathway by which behavioural traits, parental investment, and mate choice interact to generate accelerating evolutionary dynamics. By focusing on heritable variation, investment decisions, and mating preferences, it reproduces qualitative effects observed in previous studies while allowing transparent derivation of covariances, correlations, and potential for runaway evolution. This simplicity facilitates causal interpretation and generates broadly applicable, empirically testable predictions, including positive three-way correlations among altruism, ability, and offspring fitness.
3.1. Evidence and Implications
The dynamics we have considered do not disqualify the classic Hamilton calculus, but it does mean that the variables involved may not be as obvious from observation as a naïve application of Hamilton’s rule would suggest. For instance, the benefit caused by altruistic acts, when seen as investment in fitness capital, may require inclusion of multigenerational and indirect returns – not only on the recipient’s survival or fecundity. The cost to the altruist is not simply the risk or effort involved, but the opportunity costs that can be moderated by the wider effects of the decision on the parent’s economy. This could include complementarities between what the parent is doing and with what the partner, and how this combines to determines their offspring’s fitness. Finally, the relatedness should be understood as effective relatedness, which may be altered by assortative mating. This point is illustrated by the opportunity this system creates for otherwise rare “Green Beard” effects, which means genes that have an evolutionary incentive to invest more in the offspring produced by such a mating than the rest of the genome should prefer12.
In addition to humans, many species present the preconditions for altruistic investment in fitness capital. A useful starting point may be to distinguish between one-off benefits to offspring (e.g., gifts, feedings, discrete acts) and investments that enhance long-term potential (e.g., provisioning, protection, or training). If this distinction is valid, the models predict that parental effort devoted to investment should exceed what would be expected from simple relatedness-based heuristics. Measuring this directly is difficult, since the line between “gift” and “investment” is blurred. What is more tractable is to examine resources that parents are motivated to acquire: when parents are experimentally provided with additional resources, their energy and time for investment should increase even if the precise channels are harder to track. For example, it has been documented that meerkats will sometimes capture and disable (but not kill) prey items and release them close to their infants for them to apparently practice hunting (Thornton & McAuliffe, 2006). Providing meerkat mothers with plenty of dead food to eat might increase the chances she will reserve more of the live prey she encounters for her offspring to build their hunting capacities rather than eat them herself instead. Anecdotes and natural history documentaries have similarly interpreted some behaviours among other mammalian mothers, such as in big cats, where a mother will bring live but injured prey to her cubs to apparently facilitate their learning prey capture skills – although this is difficult to demonstrate experimentally (Thornton & Raihani, 2010).
From this follows the prediction that experimental boosts to parental resources should yield compounding effects over generations. This is because this increases the income of the parents, which should increase reserves available for investment. Children should acquire greater fitness capital, and grandchildren disproportionately so, as the lineage accumulates advantages. Crucially, if the cascade is sustained, these lineages should also evolve to become more altruistic, as we predict that lineages with more to invest and lineages with higher altruism tend to become intertwined13. Conversely, when historical links between resources and genes become abruptly decoupled, the distribution of fitness capital should shift. Historical episodes such as mass migration, famine, epidemic disease, or war may temporarily weaken the link between inherited capital, mate choice, and altruism. For one or more generations, assortative matching based on these traits may be disrupted, leading to less skewed distributions of capital and weaker correlations between altruism and resources.
A further prediction concerns variation among offspring. Higher-ability offspring should attract greater investment, even if parents show no explicit preference in terms of altruism per se. This aligns with widespread observations. For example, in many mammals, “runts” receive less postnatal care (Clutton-Brock, 1991), while in birds, weaker hatchlings are often fed less, with parents apparently being indifferent to them being effectively culled by their stronger siblings (Mock & Forbes, 2022). Experimental manipulations of cues associated with ability could directly test whether such signals alter parental investment decisions14. It should be noted that previous theories already make this prediction (Olson et al., 2008). What may be novel here is the added prediction that this should also be observed across couples: families with higher ability children should also tend to invest more in them than families with lower ability children. Experiments where couples with are effectively made to “adopt” low ability young should invest in them less than if given high ability young to raise.
The classic model of parental investment by Smith and Fretwell (1974) provides foundational insight into the trade-off between offspring size and number, assuming that each unit of investment yields the same marginal fitness return. While touching on similar issues to the present thesis, their framework applies most directly to species producing many small offspring with minimal postnatal care. More recent refinements, such as Johnson et al. (2024), have shown that incorporating indirect reproductive costs that scale nonlinearly with offspring traits can substantially modify predicted investment patterns. Building on these ideas, the present model explicitly considers heterogeneity in offspring ability as a multiplier of parental investment returns, allowing for differential allocation even when parents are equally altruistic toward all offspring. Unlike the previous frameworks, our approach captures intergenerational feedbacks, whereby investment, ability, and capital accumulation reinforce each other over time, and extends naturally into domains of mate choice and signalling. This produces testable predictions that are not addressed by earlier models: for example, parents should preferentially invest in offspring with higher ability, leading to lineages that accumulate disproportionately more capital, and small initial differences in ability or investment can snowball into larger disparities across generations. In this way, our model retains the simplicity of the foundational Smith–Fretwell logic while highlighting novel mechanisms by which parental investment can interact with offspring traits to shape evolutionary trajectories.
3.2. Limitations
Can the processes we have considered here be expected to push investment indefinitely high? In principle, escalation may be rapid, but there are realistic constraints to consider. Ever more extreme levels of parental investment may, for instance, be resisted if they require changes that sacrifice other fitness components. In the same way that the sheer size of egg produced by kiwi bird or the head size of human foetuses are likely near limits balanced against the capacity of mothers to produce such infants, limits imposed by constraints may be less elastic to selection than altruism. Thus, as with other potential drivers of runaway evolution (e.g., Dawkins & Krebs, 1979), there may be direct costs of rising investment in terms of viability, energy, time, or other factors that curtail the process. Another obvious limitation is environmental resources. For example, we can imagine a predator evolving to specialize in hunting ever larger prey because both hunting skills and meat are the sorts of resources that can be concentrated and invested in offspring. But it such a predator may find it increasingly less worthwhile as such prey become scarcer, or if this induces overwhelming counter-adaptations on the part of the prey species, or if it falls victim to its own success if it drives the over-exploited prey species extinct. For these and other reasons, we have assumed diminishing returns on investment in fitness capital.
A yet more interesting theoretical limitation was that we did not consider the role of altruism, ability, or capital in decisions about the quantity of children to have. On the one hand, there may be quantity–quality trade-offs, where higher-ability parents decide to invest more in fewer children (e.g., Becker, 1991), while lower-ability parents produce more children with less investment. This might result in a r–K distribution of traits, as has been suggested to exist in humans (e.g., Eysenck and Gudjonsson, 1989). On the other hand, if having higher-ability children, valuing their fitness more, and having more to invest in those children goes hand in hand, then this may be a pressure in the other direction, motivating high-altruism couples to produce more children15. Modelling variation in family size along these lines would also make it clearer why the parent’s own “consumption” cannot be treated as irrelevant in a Darwinian model: even when parents have only one child at a time, resources reserved for their own well-being may contribute indirectly to the survival of additional children in the future, or serve as a buffer against risk if current offspring die. Thus, modelling a division between “consumption” and “investment” remains defensible (cf. Trivers, 1974; Kaplan, 1996).
We have also not considered how the same predictions may play out within a wider kin-network, (cousins, nephews and nieces, etc.), much less when the genetic relatedness among such networks departs from the standard r=.5 assumed for parents and offspring. Finally, different types of mating structure may be important. For example, systems without monogamy, or where not all of one sex get paired (e.g., more adult males than females are available for mating), or truncated, soft versus hard selection regimes. Any of these aspects may reveal conditions under which altruism is less reliable connected with capital, potentially weakening the recursive dynamic and other conclusions.
3.3. Extensions
The discussion turns here to speculative extensions and future directions. We have so far assumed that parents act based on accurate knowledge about the characteristics of others, such as their partner’s altruism and offspring ability. While the present model did not address deception or signalling, the logic extends naturally into these domains, offering testable hypotheses. Introducing errors or deceptive signalling may mean selection favouring altruism can be expected to weaken or otherwise become more complicated as investment no longer reliably translates into transgenerational returns. In particular, a disconnect between altruism ability and actual relatedness raises opportunities for exploitation and parasitism. Haig’s (1997; 2001) Intergenomic Conflict theory, for example, may make the following prediction in situations where there is paternal uncertainty. Paternally-imprinted genes might be expected to program offspring to inflate their apparent ability — or play up issues of equity — to better extract resources from the mother and her partner, who may or may not be the actual father. Meanwhile, maternally imprinted genes should want optimal allocation in agreement with the mother’s genetic perspective. Exaggerated signals that investment in a particular child are especially effective may work because they hijack parental responsiveness to this as a reward signal. Further thought is required to clarify whether the resulting pressures on parents to be discerning is enough to nullify this exploitation, or if some other result is likely to emerge from an arms race between children obscuring their ability and parents striving to allocate their scarce resources efficiently.
The same logic may explain why parent birds parasitized by cuckoos have often been observed behaving as if strongly motivated to continue feeding cuckoo chicks even when they have grown enormous (Krüger, 2007a). A classic interpretation is that this is despite the cuckoo’s enormous rate of growth, and due to the parents responding slavishly to especially salient stimuli the cuckoo chic has evolved to convey (Rojas Ripari et al., 2021). Based on the present model, it may be partly because the cuckoo bird’s accelerated development mimics high ability that explains the parasitized bird’s behaviour: they may have been selected to respond with more investment to this as a sign that of underlying ability in their own offspring, inadvertently benefiting the cuckoo. Species where parents pattern their investment in this way may be especially vulnerable to parasitism by cuckooing. This may explain why most cuckoos parasitize species that are smaller than or similarly sized to themselves (Krüger, 2006b). In the exceptions where the cuckoo grows to a smaller adult size than the parasitized species, they nonetheless grow at faster rates when young (e.g., Kleven et al., 1999).
The present thesis may have applications to understanding so-called “handicap” signals as well (Zahavi, 1975). Conceivably, animals can be selected to pay high costs to transmit honest signals of their underlying qualities because they advertise their underlying qualities, such as having good genes. A classic example is gazelle stoting in response to predators like leopards, with the idea being that the healthiest can afford to jump the highest in this way, thereby displacing the predator’s efforts to target whichever individuals cannot afford to advertise their vigour in this way – even though all gazelle would be faster at putting distance between themselves and the leopard by simply running away instead (Dawkins, 1976). Crucially, Grafen (1990) showed that such a signalling system is stable only when the cost of signal strength is proportional to the underlying quality it is supposed to indicate. The kind of cascading investment process we have been considering may naturally set up such a set of conditions for costly signalling of either ability or altruism. This is because these characteristics can be expected to correlate over time with earnings on our model, thus making signals costing a given amount of earnings being disproportionately cheaper for more altruistic individuals to afford, and we have seen why there may be advantages to conveying such signals to prospective mates. Our model could be extended to predict, for example, that a highly altruistic predator may signal its quality as a parent by tackling more dangerous or elusive prey than they would otherwise choose (serving as a trophy to impress potential mates or rivals). Obviously, there may be reasons for preferring high ability and capital individuals, but this approach could explain why costly or handicapped signalling of these attributes might be expected to evolve.
Finally, something may be said about other types of altruism to close this survey of potential implications of the general model. Recall our assumption that returns in a child diminish incrementally as more is invested. On Becker’s analysis of human behaviour, this means there is a point at which parents might become motivated to allocate some of their investment in that of the capital of unrelated children. This is because, if combined with the assumption of efficient capital markets, lending investment in this way to non-relatives means that this may yield higher returns later when the non-relatives grow up and effectively repay the loan to the descendants who made the loan. Obviously, the assumption of perfect capital markets does not hold well in most biological systems including the ancestral environments of human beings. However, an extension of the foundational concepts of Trivers (1971; 2002) and Axelrod and Hamilton (1981) on altruism among non-relatives to include fitness capital may be workable provided that the unrelated children invested in are likely to interact cooperatively with the offspring of the parents investing. The age-old problem for altruism based mutual reciprocation is how to reinforce repayment (Ridley, 1997). In Becker’s case, this can be done through contracts, while in the biological case reciprocal altruism relies on effective deterrence of defection. In some cases, it may be plausible that parents who have had a proven history of cooperation with individuals in another lineage might expect their own children and the children of the unrelated lineage to also cooperate, in a way substituting for a contract and the need to directly enforce it16. Further modelling may be needed to develop this point, but it is intuitive that the present value of cooperating now might be enhanced if it is felt that the “shadow of the future” extends across overlapping generations. For example, in addition to having higher than average relatedness to other lionesses within a pride, each mother lioness has some level of certainty that their offspring will grow up to later rely on the skills and cooperation of the offspring of other mothers in the pride. This is one way that altruism for kin might influence the evolution of altruism in other forms.
Another potential implication is, more generally, the effect of practice on competence. If partners in cooperation improve in their capacity to provide benefits through the practice of reciprocal altruism, then this may have implications that have been underappreciated. Previous models have considered what happens with tit-for-tatters find each other and avoid defectors: in the same way that binding altruism with investment leads to higher variance and average fitness capital, non-random association and repeated mutually beneficial interactions by cooperators (another type of altruism) might magnify average levels of cooperation. Simply basing cross lineage investment on track record might be enough to justify an enlightened “Benselfishness”17.
4. Conclusions
If altruism can be understood as motivating investments in fitness capital, then its evolutionary significance may be more profound than previously appreciated. Investment cascades can bind the capacity to acquire resources with the motivation to invest them in kin, thereby increasing both average altruism and fitness capital. Such dynamics may interact with mating systems and other evolutionary forces to shape how individuals cooperate and compete. This preliminary synthesis of ideas from evolutionary biology and economics not only points to novel empirical predictions but is rich with implications that can be further explored to build a richer understanding of how perhaps some of the strongest and intimate social bonds have been, and are being, evolved.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
In the main text, we described parental investment and its intergenerational consequences using a single-period optimization framework. To make the overlapping logic explicit, we can formalize a two-stage life cycle where parents coexist with their offspring for a time and where offspring inherit both fitness capital and parental altruism. This appendix provides a brief illustration of this dynamic.
Let the population be indexed by discrete time steps t. Each generation consists of individuals who receive fitness capital from their parents, earn income , allocate a share of income () to altruistic investment in offspring , and transmit genes for altruism to the next generation. The variable summarizes health, skills, or embodied ability that enhances income and survival, and that can be enhanced by more parental investment, and evolves over time according to the accumulation function:
Where is the rate of depreciation (aging, obsolescence, random mutation), is the parental investment, >0 captures transmission efficiency, and governs diminishing returns. Investment depends on altruism and parental resources . Substituting, we obtain the recursion:
(A1)
Let individual fitness, depend on income retained for own reproduction and survival:
Where represents the direct contribution of fitness capital to reproductive success or longevity. The change in mean altruism across generations follows the standard covariance (from Price equation):
(A2)
Where the first term represents selection on altruism and the second transmission effects. Using A1 and A2, altruism evolves when investment benefits () generate heritable fitness advantages through offspring success. Differentiating A1 with respect to :
(A3)
Thus, altruism increases offspring capital whenever . Iterating A1 forward shows that altruism has compounding effects:
When altruism is heritable, the positive cross-generational covariance is
And so amplifies both traits over time, generating intergenerational feedback between altruism and fitness capital. To find the steady state (), equation A1 produces:
(A4)
This means that the equilibrium amount of fitness capital and heritable altruism that individuals maintain in the population rises when parents invest a larger share of their income in offspring, when investment is more efficiently transmitted, or when existing capital erodes slowly. The more strongly returns diminish (γ < 1), the less sensitive the equilibrium is to changes in investment or efficiency. Higher equilibrium altruism () increases steady state capital () provided that . The system is locally stale if the derivative of the RHS of A1 with respect to satisfies the following:
(A5)
This ensures convergence to the steady state feedback loop, rather than runaway escalation or extinction.
Equations A1 to A5 formalize the model in the text of section 2.3. They show how altruism raises investment, which enhances offspring capital, which in turn raises both income and fitness. Heritable variation in altruism and the investible trait of fitness capital become positively correlated across generations.
During the first period, they are reproductively mature adults, making decisions about their own consumption and investment in their offspring. In the second period, they survive to observe the outcomes of their investment, contributing to overlapping generation effects. Altruism is heritable. Let denote an individual’s fitness capital at time encompassing both heritable traits and the effects of parental investment. Parental utility is assumed to depend on their own consumption and wellbeing of their offspring, weighted by altruism , and a discount factor reflecting the parent’s time horizon over future fitness returns.
Appendix B
In the main text, we showed that assortative mating on altruism, ability or fitness capital can generate intergenerational covariances that are mutually reinforcing. This appendix formalizes that mechanism to clarify conditions under which a runaway process can emerge. Let each individual in generation be characterized by a pair of traits (, ), where represents altruism and is fitness capital. From Appendix A, the offspring’s fitness capital evolves as:
(B1)
Assume that altruism is heritable such that the expected altruism of offspring is an increasing function of the average altruism of the parents:
(B2)
Where measures regression to the population means due to imperfect transmission. The parameter can be interpreted as an inverse measure of heritability (where higher means weaker inheritance). Let mates pair assortative based on altruism, such that covariance between altruism of mating partners is:
(B3)
Where captures the strength of assortative mating ( indicates PAM on altruism). Taking expectations over mating pairs, the average altruism of couples is:
, and the variance among pairs is (B4)
Thus, assortative mating increases between-family variance in altruism, which in turn magnifies the variance in offspring investment in capital. From (B1), the sensitivity of offspring capital to parental altruism is:
(B5)
Hence, the covariance between altruism and offspring capital at is approximately:
(B6)
PAM on altruism () raises , and therefore increases the covariance . This strengthens the cross generational link between altruism and fitness capital. Because altruism is itself heritable (equation B2), this positive covariance feeds back into the next generation’s variance and mean altruism. The approximate recursion for the covariance is:
()( ) ) (B7)
Whenever the second term is positive and sufficiently large relative to the first, decay term )( )), the covariance grows over time:
(B8)
This implies that altruistic, high capital lineages increasingly assort and amplify both traits across generations in a self-reinforcing process. If we define such an amplification factors as equal to the ration:
(B9)
Runaway selection occurs when , that is, when the joint inheritance and assortative pairing on altruism and capital lead to ever-increasing covariances between the traits. From B7, this condition is satisfied when:
(B10)
In other words, runway amplification occurs when the marginal productivity of altruism in generating offspring fitness capital, weighted by the variance in altruism, exceeds the dampening effects of imperfect transmission and capital depreciation. Thus, strong heritability, efficient investment transmission, and positive assortative mating on altruism together favour explosive growth in both altruism and capital. This parallels the Fisher runaway selection idea, but here operates through parental investment and mate choice rather than ornament and preference.
This mechanism predicts the coevolution of altruism, ability, and mate preferences that reinforce one another across generations. Couples high in altruism invest more, producing offspring with higher fitness capital, who are in turn more altruistic and more attractive as mates to other altruists. This creates a feedback loop that raises both mean altruism and capital in the population. Equation (B10) formalizes the threshold condition for this process. When violated (e.g., through weak heritability, strong depreciation, or low assortative mating), the system instead converges to a stable equilibrium as described in Appendix A (Appendix B thereby complements Appendix A by explicitly deriving the conditions under which intergenerational feedback between altruism and capital — mediated by assortative mating — leads to self-reinforcing or “runaway” evolutionary dynamics).
References
- Axelrod, R., & Hamilton, W. D. (1981). The evolution of co-operation. Science, 211, 1390-1396.
- Becker, G.S. A Theory of Marriage: Part I. J. Politi- Econ. 1973, 81, 813–846. [CrossRef]
- Becker, G.S. A Theory of Marriage: Part II. J. Politi- Econ. 1974, 82, S11–S26. [CrossRef]
- Happel, S.K.; Becker, G.S. A Treatise on the Family. J. Marriage Fam. 1983, 45, 231. [CrossRef]
- Becker, G. S. (1996). Accounting for Tastes. Harvard University Press.
- Becker, G. S. (2002). Human capital. The concise encyclopedia of economics, 2, 1-12.
- Bourke, A.F.G. The validity and value of inclusive fitness theory. Proc. R. Soc. B: Biol. Sci. 2011, 278, 3313–3320. [CrossRef]
- Brown, S.P.; West, S.A.; Diggle, S.P.; Griffin, A.S. Social evolution in micro-organisms and a Trojan horse approach to medical intervention strategies. Philos. Trans. R. Soc. B: Biol. Sci. 2009, 364, 3157–3168. [CrossRef]
- Caplan, B. (2011). Selfish Reasons to Have More Kids. Basic Books.
- Class, B., & Dingemanse, N. J. (Eds.). (2022). Assortative mating for quantitative traits: Mechanisms, estimation, and evolutionary consequences [Special issue]. Journal of Evolutionary Biology, 35(4).
- Zuk, M.; Allen, L.; Cupp, L.; Eller, K.; Hamilton, P.; Hazard, L.C.; Jorgenson, M.; Kaufhold, M.; Mirmovitch, V.; Rabitoy, E.; et al. The Evolution of Parental Care. Copeia 1992, 1992, 594. [CrossRef]
- Dawkins, R. (1976). The Selfish Gene. Oxford: Oxford University Press.
- Dawkins, R. (1982). The Extended Phenotype. Oxford: W. H. Freeman.
- Dawkins, R., & Krebs, J. R. (1979). Arms races between and within species. Proceedings of the Royal Society of London. Series B. Biological Sciences, 205(1161), 489-511.
- Dawkins, R. (2004). A Devil's Chaplain. Houghton Mifflin Harcourt.
- nbsp;Eldredge, N.; Gould, S.J. Punctuated equilibria: An alternative to phyletic gradualism. In Essential Readings in Evolutionary Biology; Ayala, F.J., Avise, J.C., Eds.; John Hopkins University Press: Baltimore, MD, USA, 1972; pp. 82–115.
- Eysenck, H.J.; Gudjonsson, G. Concerning the causes and cures of criminality: A reply to Trasler. Contemp. Psychol. A J. Rev. 1992, 37, 724–725. [CrossRef]
- Fisher, R. A. (1930). The Genetical Theory of Natural Selection. Oxford: Clarendon Press.
- Frank, S.A. Natural selection. IV. The Price equation*. J. Evol. Biol. 2012, 25, 1002–1019. [CrossRef]
- Friedman, M. (1962). Price Theory. University of Chicago Press.
- Gould, S. J. (1977). Ever Since Darwin. New York: Norton.
- Grafen, A. (1990). Biological signals as handicaps. Journal of Theoretical Biology, 144(4), 517-546.
- Haig, D. Parental antagonism, relatedness asymmetries, and genomic imprinting. Proc. R. Soc. B: Biol. Sci. 1997, 264, 1657–1662. [CrossRef]
- Haig, D. Genomic Imprinting and Kinship: How Good is the Evidence?. Annu. Rev. Genet. 2004, 38, 553–585. [CrossRef]
- Hamilton, W. D. (1963). The evolution of altruistic behavior. The American Naturalist, 97, 354-356.
- Hamilton, W. The genetical evolution of social behaviour. II. J. Theor. Biol. 1964, 7, 17–52. [CrossRef]
- Badcock, C.; Hamilton, W.D. Narrow Roads of Gene Land: The Collected Papers of W. D. Hamilton, Volume 1: The Evolution of Social Behaviour. Br. J. Sociol. 1997, 48, 332. [CrossRef]
- Badcock, C.; Hamilton, W.D. Narrow Roads of Gene Land: The Collected Papers of W. D. Hamilton, Volume 1: The Evolution of Social Behaviour. Br. J. Sociol. 1997, 48, 332. [CrossRef]
- Hawkes, K. kin selection and culture. Am. Ethnol. 1983, 10, 345–363. [CrossRef]
- Jaffe, S., Minton, R., Mulligan, C. B., & Murphy, K. M. (2019). Chicago Price Theory. Princeton University Press.
- Johnson, D.W.; Chhor, J.T.; E Shelley, C.; Siegfried, E.J. Indirect costs of reproduction and the tradeoff between offspring size and number: a framework illustrated by fitness costs and benefits of ovarian fluid. Evolution 2024, 78, 1248–1260. [CrossRef]
- Kaplan, H. A theory of fertility and parental investment in traditional and modern human societies. Am. J. Phys. Anthr. 1996, 101, 91–135.
- Kleven, O.; Moksnes, A.; Røskaft, E.; Honza, M. Host species affects the growth rate of cuckoo ( Cuculus canorus ) chicks. Behav. Ecol. Sociobiol. 1999, 47, 41–46. [CrossRef]
- Krüger, O. Cuckoos, cowbirds and hosts: adaptations, trade-offs and constraints. Philos. Trans. R. Soc. B: Biol. Sci. 2006, 362, 1873–1886. [CrossRef]
- Krüger, O. Cuckoos, cowbirds and hosts: adaptations, trade-offs and constraints. Philos. Trans. R. Soc. B: Biol. Sci. 2006, 362, 1873–1886. [CrossRef]
- Krützen, M.; Mann, J.; Heithaus, M.R.; Connor, R.C.; Bejder, L.; Sherwin, W.B. Cultural transmission of tool use in bottlenose dolphins. Proc. Natl. Acad. Sci. 2005, 102, 8939–8943. [CrossRef]
- Laland, K. N., & Feldman, M. W. (2003). Niche construction: The neglected process in evolution. Princeton University Press.
- Laland, K.; Odling-Smee, J.; Endler, J. Niche construction, sources of selection and trait coevolution. Interface Focus 2017, 7, 20160147. [CrossRef]
- Lande, R. Models of speciation by sexual selection on polygenic traits. Proc. Natl. Acad. Sci. 1981, 78, 3721–3725. [CrossRef]
- Lehmann, L. THE ADAPTIVE DYNAMICS OF NICHE CONSTRUCTING TRAITS IN SPATIALLY SUBDIVIDED POPULATIONS: EVOLVING POSTHUMOUS EXTENDED PHENOTYPES. Evolution 2008, 62, 549–566. [CrossRef]
- Ligon, R.A.; Hill, G.E. Sex-biased parental investment is correlated with mate ornamentation in eastern bluebirds. Anim. Behav. 2010, 79, 727–734. [CrossRef]
- Marshall, D. J, & Uller, T. (2007). When is a maternal effect adaptive?. Oikos, 116(12), 1957-1963.
- Maynard Smith, J. (1966). Sympatric speciation. American Naturalist, 100(916), 637–650.
- McKinnon, R.A.; Hedlin, E.; Hawkshaw, K.; Mathot, K.J. Food supplementing peregrine falcon ( Falco peregrinus tundrius ) nests increases reproductive success with no change in mean parental provisioning rate. R. Soc. Open Sci. 2024, 11, 240576. [CrossRef]
- Mock, D. W., & Forbes, L. S. (2022). Parental care in birds. Current Biology, 32(1), R42-R53.
- A Mousseau, T.; Fox, C.W. The adaptive significance of maternal effects. Trends Ecol. Evol. 1998, 13, 403–407. [CrossRef]
- Mullon, C.; Lehmann, L. Invasion fitness for gene–culture co-evolution in family-structured populations and an application to cumulative culture under vertical transmission. Theor. Popul. Biol. 2017, 116, 33–46. [CrossRef]
- Olson, V.; Liker, A.; Freckleton, R.; Székely, T. Parental conflict in birds: comparative analyses of offspring development, ecology and mating opportunities. Proc. R. Soc. B: Biol. Sci. 2007, 275, 301–307. [CrossRef]
- Price, G.R. 1970. Selection and covariance. Nature, 227, 520–521.
- Ridley, M. (1997). The Origins of Virtue. Penguin UK.
- Ripari, J.M.R.; Ursino, C.A.; Reboreda, J.C.; De Mársico, M.C. Tricking Parents: A Review of Mechanisms and Signals of Host Manipulation by Brood-Parasitic Young. Front. Ecol. Evol. 2021, 9. [CrossRef]
- Roy, A.D. SOME THOUGHTS ON THE DISTRIBUTION OF EARNINGS 1. Oxf. Econ. Pap. 1951, 3, 135–146. [CrossRef]
- Roy, D.; Roy, B. A red queen model of personality. Evol. Mind Behav. 2018, 16, 1–36. [CrossRef]
- Santos, M.; Monteiro, A.L.; Biz, A.N.; Guerra, A.; Cramer, H.; Canuto, V.; Cruz, L.; Pinto, M.; Viegas, M.; Fernandes, R.; et al. Guidelines for Utility Measurement for Economic Analysis: The Brazilian Policy. Value Heal. Reg. Issues 2022, 31, 67–73. [CrossRef]
- Shine, R.; Brown, G.P.; Phillips, B.L. An evolutionary process that assembles phenotypes through space rather than through time. Proc. Natl. Acad. Sci. 2011, 108, 5708–5711. [CrossRef]
- Shine, R.; Brown, G.P.; Phillips, B.L. Reply to Lee: Spatial sorting, assortative mating, and natural selection. Proc. Natl. Acad. Sci. 2011, 108, E348–E348. [CrossRef]
- Smith, C.C.; Fretwell, S.D. The Optimal Balance between Size and Number of Offspring. Am. Nat. 1974, 108, 499–506. [CrossRef]
- Sowell, T. (2016). Wealth, Poverty and Politics. Hachette UK.
- Thornton, A.; McAuliffe, K. Teaching in Wild Meerkats. Science 2006, 313, 227–229. [CrossRef]
- Thornton, A.; Raihani, N.J. Identifying teaching in wild animals. Learn. Behav. 2010, 38, 297–309. [CrossRef]
- Trivers, R.L. The Evolution of Reciprocal Altruism. Q. Rev. Biol. 1971, 46, 35–57. [CrossRef]
- Trivers, R. L. (1974). Parent-offspring conflict. American Zoologist, 14, 249-264.
- Trivers, R. (2002). Natural Selection and Social Theory: Selected papers of Robert Trivers. Oxford University Press.
- Uller, T. Developmental plasticity and the evolution of parental effects. Trends Ecol. Evol. 2008, 23, 432–438. [CrossRef]
- Van Cleve, J., & Akçay, E. (2014). Pathways to social evolution: reciprocity, relatedness, and synergy. Evolution, 68(8), 2245-2258.
- Zahavi, A. Mate selection—A selection for a handicap. J. Theor. Biol. 1975, 53, 205–214. [CrossRef]
Figure 1.
Income effect on investment. Line CB represents the budget line for a parent choosing between investing on their child or spending on their own consumption (if they spent all their resources on consumption, this would be equal to line AB and zero investment; if all they did was invest, this would result in zero consumption and AC level of investment). The optimal mix is given where the budget line is tangent to the parent’s indifference curve (), corresponding to investment level . Assuming the shape of the indifference curve is unchanged, increasing the parent’s total budget from CB to DE would give a level of optimal investment that is .
Figure 1.
Income effect on investment. Line CB represents the budget line for a parent choosing between investing on their child or spending on their own consumption (if they spent all their resources on consumption, this would be equal to line AB and zero investment; if all they did was invest, this would result in zero consumption and AC level of investment). The optimal mix is given where the budget line is tangent to the parent’s indifference curve (), corresponding to investment level . Assuming the shape of the indifference curve is unchanged, increasing the parent’s total budget from CB to DE would give a level of optimal investment that is .
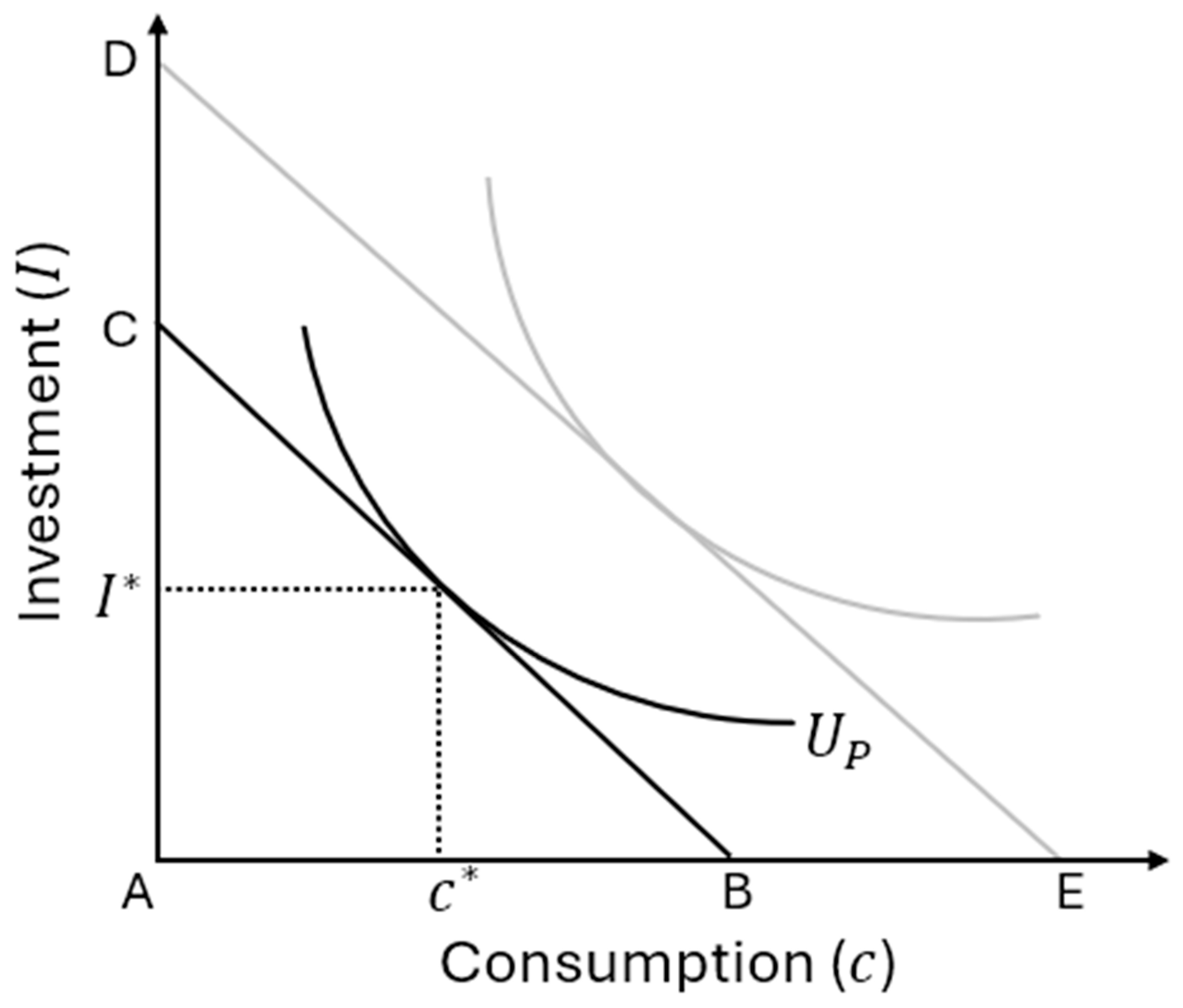
Figure 2.
Hypothetical relationship between parent and offspring characteristics. Parents with higher fitness capital earn more income, which means that they invest more in their offspring’s fitness capital. Higher parental altruism tends to be inherited by the offspring, as well as increases the fraction of income earned going towards investment. Red arrows depict genetic transmission from parent to offspring (measured by its heritability or ), solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. .
Figure 2.
Hypothetical relationship between parent and offspring characteristics. Parents with higher fitness capital earn more income, which means that they invest more in their offspring’s fitness capital. Higher parental altruism tends to be inherited by the offspring, as well as increases the fraction of income earned going towards investment. Red arrows depict genetic transmission from parent to offspring (measured by its heritability or ), solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. .
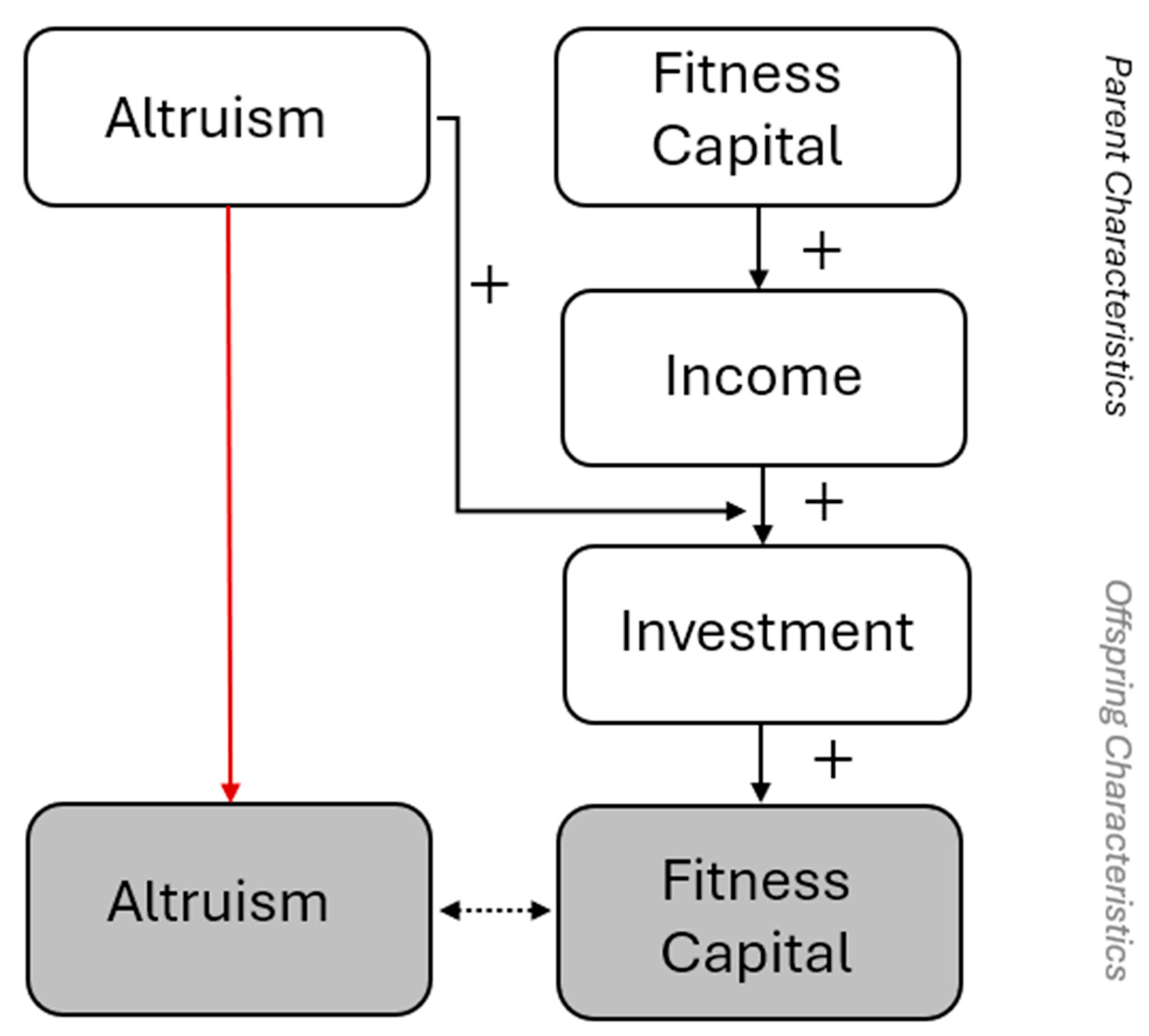
Figure 3.
Different levels of investment in kin depending on individual differences in ability. Lines and show diminishing marginal utility of investment for a high and low ability recipient of investments, respectively. It is rational to invest up until the point until the incremental benefit produced from investing another unit of resources is balanced against the marginal benefit that could be obtained from spending it instead on consumption (). This means it is optimal to invest than in child with ability .
Figure 3.
Different levels of investment in kin depending on individual differences in ability. Lines and show diminishing marginal utility of investment for a high and low ability recipient of investments, respectively. It is rational to invest up until the point until the incremental benefit produced from investing another unit of resources is balanced against the marginal benefit that could be obtained from spending it instead on consumption (). This means it is optimal to invest than in child with ability .
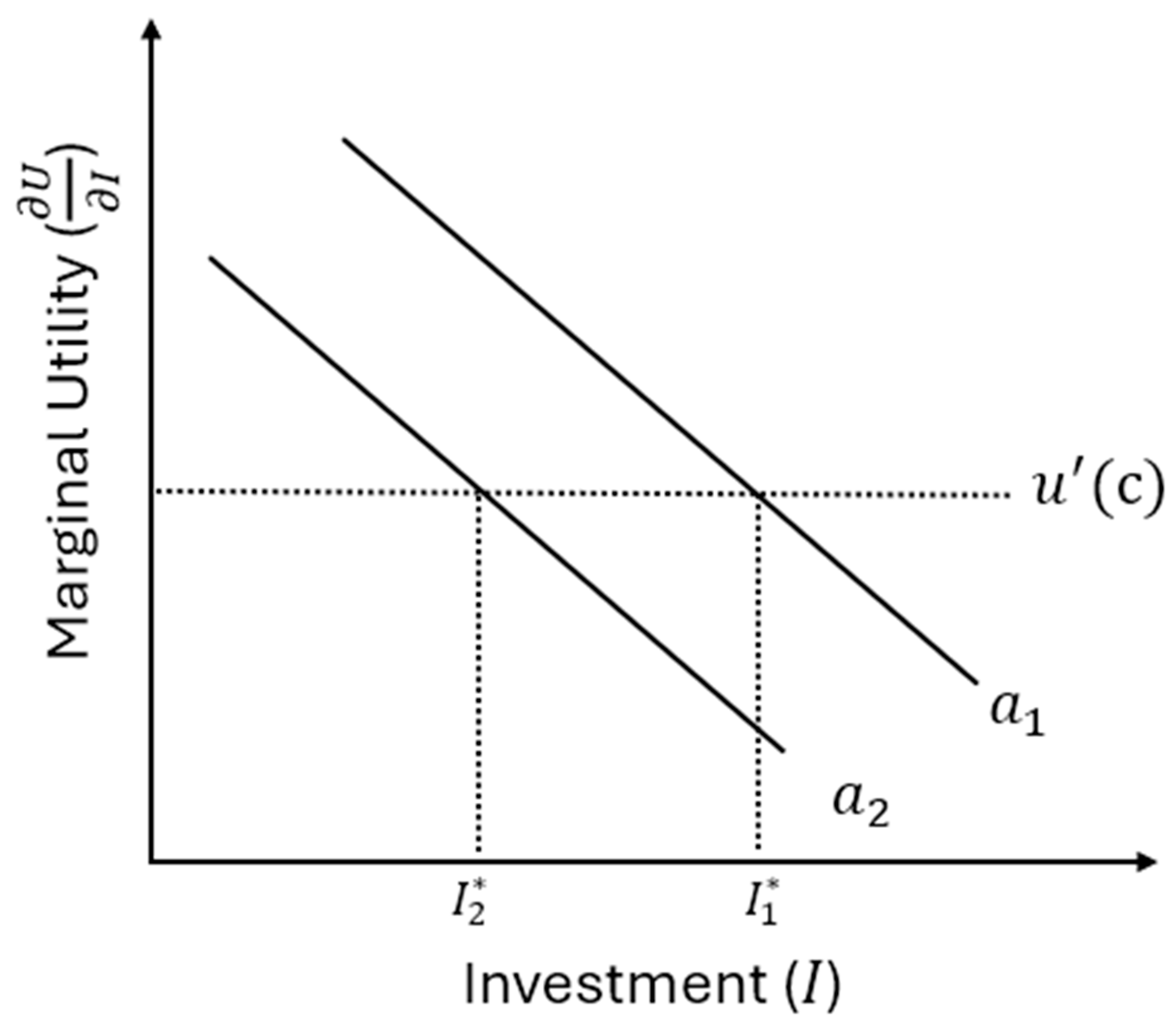
Figure 5.
Positive Assortative Mating on Altruism. Purple and blue boxes represent characteristics of parent A and parent B, who unite to produce a “household” that earns income and invests in their offspring (whose characteristics are shaded in grey). Parents with more fitness capital earn more income and so tend to invest more in the fitness capital of offspring. If such couples are more altruistic, a greater proportion of whatever they earn gets invested. The process tends to increase the correlation between altruism and fitness capital, which implies that, if more altruistic parents choose more altruistic partners, couples will tend to become correlated in altruism and fitness capital. Red arrows depict genetic transmission from parent to offspring, solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. .
Figure 5.
Positive Assortative Mating on Altruism. Purple and blue boxes represent characteristics of parent A and parent B, who unite to produce a “household” that earns income and invests in their offspring (whose characteristics are shaded in grey). Parents with more fitness capital earn more income and so tend to invest more in the fitness capital of offspring. If such couples are more altruistic, a greater proportion of whatever they earn gets invested. The process tends to increase the correlation between altruism and fitness capital, which implies that, if more altruistic parents choose more altruistic partners, couples will tend to become correlated in altruism and fitness capital. Red arrows depict genetic transmission from parent to offspring, solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. .
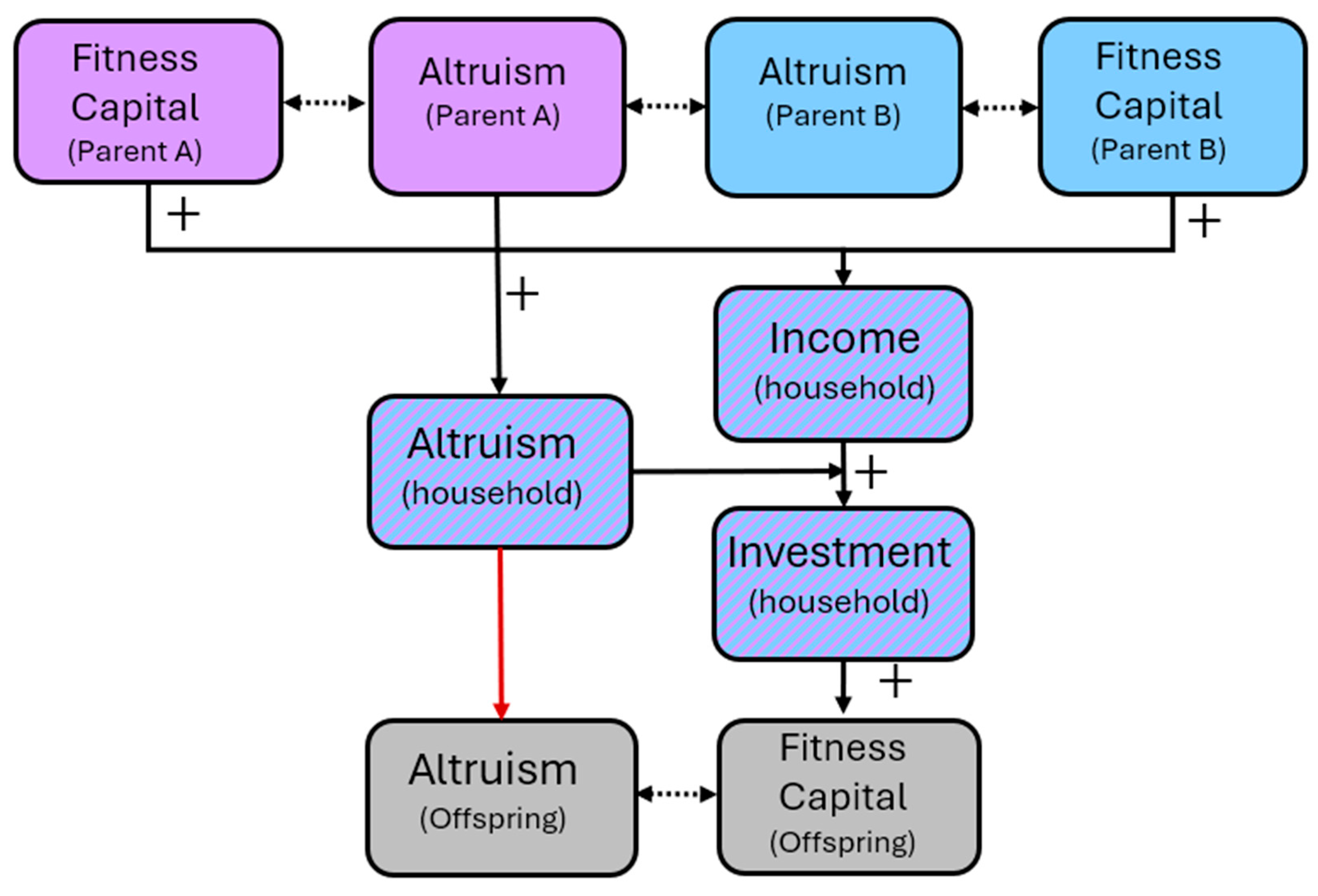
Figure 6.
PAM on altruism and preconditions for runaway sexual selection. Parents sort based on altruism because of similar characteristics have complementarities in household production (e.g., couples earn disproportionately more working together if they are both high on altruism). If there are genetic components affecting this preference () and these traits (), this can increase their correlation in subsequent next generations (linkage disequilibrium) and so raise . Red arrows depict genetic transmission from parent to offspring, solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. .
Figure 6.
PAM on altruism and preconditions for runaway sexual selection. Parents sort based on altruism because of similar characteristics have complementarities in household production (e.g., couples earn disproportionately more working together if they are both high on altruism). If there are genetic components affecting this preference () and these traits (), this can increase their correlation in subsequent next generations (linkage disequilibrium) and so raise . Red arrows depict genetic transmission from parent to offspring, solid arrows with pluses represent positive causal effects of behaviour, and the dotted line represents where these effects, if recuring over generations, produce a correlation. .
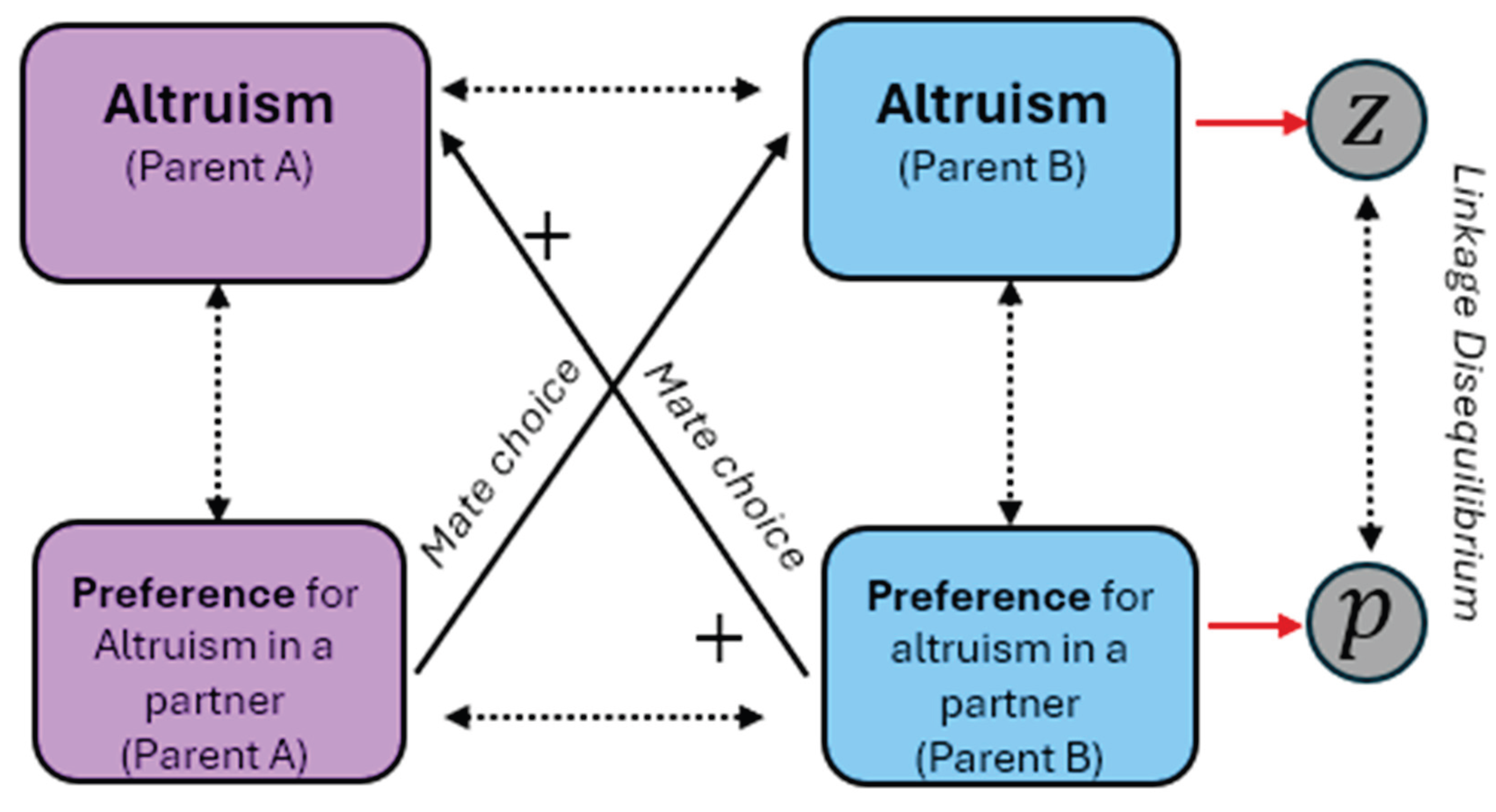
| 1 | In economics, it is standard practice to convert different units of value—such as utility, consumption, or wellbeing—into a common metric so that marginal costs and benefits can be meaningfully compared in optimization problems (Jaffe et al., 2019; Santos et al., 2022). We adopt a similar approach here by translating parental valuation of offspring outcomes into expected effects on fitness. This allows the framework of altruistic parental investment to be formally linked to Hamilton’s rule: the effective weight a parent places on a child’s outcome combines both the genetic relatedness and the degree to which increases in the child’s wellbeing translate into reproductive success. In this way, we can integrate economic and evolutionary perspectives while maintaining a consistent scale for the parent’s decision-making. |
| 2 | In some species of birds, for example, parents can be thought of as doing more than just feeding their chicks for their immediate survival, but also to enhance feather development, because brighter, well-formed plumage can improve future mating prospects (e.g., Ligon & Hill, 2010). We have, of course, oversimplified in the sense that many goods and services parents can provide for their young are likely to affect both children’s current wellbeing and future fitness. Between hypothetical extremes of goods that are purely for the child’s consumption and those that are purely investments in their later potential, we may imagine a continuum that is captured by . Benefits that lean towards the investment extreme will correspond to a higher . |
| 3 | Put another way, encodes the “efficiency curve” by which parental effort is transformed into offspring fitness. Many biological examples may be given of this. For instance, in passerine birds, investment may be the number of food items brought to chicks in the nest, while represents the chick’s size and condition. then represents how these meals translate into growth and fledging success, with this being large at first but making less of a difference when very large amounts of food have already been provided. In mammals, investment might be thought of as lactation. In longer lived organisms, such as elephants, this may take forms including teaching and transmitting skills by setting examples and facilitating learning opportunities. In each case, the key principle is that parents can push their offspring’s ultimate capacity to acquire and invest resources. |
| 4 | While relatedness coefficients also attenuate this contribution (each descendent generation would be less related in sexually reproducing organisms), it can be helpful to distinguish altruism () from discounting () for the following reasons. Altruism here reflects the strength of preference a parent places on descendent outcomes, while the discount factor reflects the fact that the benefit to more distant descendants occurs later in time and passes through additional demographic filters, which may attenuate their effect on current fitness. Thus, its inclusion here may not be redundant with relatedness, capturing causes of temporal decay other than genetic dilution. |
| 5 | In general, Becker’s economic framework imply that higher-ability offspring will attract greater parental investment, since returns to resources tend to rise (or diminish only slowly) with ability. This conclusion can be overturned only under special conditions—for example, if marginal returns to investment decline steeply, if survival thresholds make the weakest offspring the most responsive to aid, or if parents adopt equalization or risk-spreading strategies. Thus, favouritism toward the more able is the default expectation, with reversed patterns requiring additional complications. |
| 6 | Assuming also that males and females are free to choose their partners then, at equilibrium, the male with the highest level of capital will pair with the female with the highest level, the male with the second highest level with the female having the second highest level, and so on, down the line. |
| 7 | Evidence for positive assortative mating (choosing mates because they are similar in some characteristic) is widespread in nature (see Class, B., & Dingemanse, N. J. (Eds.). (2022). Assortative mating for quantitative traits: Mechanisms, estimation, and evolutionary consequences [Special issue]. Journal of Evolutionary Biology, 35(4)). |
| 8 | Admittedly, only as interpreted here based on explanations from Hamilton (1996) and Frank (2012). |
| 9 | All quantities mentioned here are being measured as marginal expected offspring equivalents. |
| 10 | The amount of extra assortative mating required to overcome the “baseline shortfall” is . |
| 11 | In this sense, it is like the “spatial sorting” effect described by Shine, Brown, & Phillips (2011a). An example of this is the invasive population of cane toads in Australia. Because leg length determines their rate of dispersal, genes for long legs tend to find themselves in bodies further out from where they were first introduced, while genes for the shortest legs tend to be left behind. This means that the longest legged toads naturally mate with each other, because that reflects a statistical bias automatically generated by the differences in speed of movement as the invasion proceeds to enter new territory. The result is an extreme skew in the distribution of leg length, much like what is predicted here for fitness capital and altruism. A parallel point about positive assortative mating potentially interacting with spatial sorting to build phenotypic variance has also been appreciated (Shine, Brown, & Phillips, 2011b). |
| 12 | The alert reader may also notice that for the same reasons that Green Bearded genetic elements may be favoured by this process, replicating cultural elements (i.e., “memes”) may evolve and perpetuate by promoting the selection of highly altruistic mates to invest more in offspring than may be optimal from the from the standpoint of the genes involved. All we need assume is that parents with such a “memetic parasite” are also more likely to install that meme in the children they invest so heavily in. More generally, we would expect that cultural and other epigenetic factors promoting investment in kin and relevant genetic aspects to become related and coevolve, as culture represents a significant channel through which much investment in human abilities flows. At minimum, Kin-Selection provides a strong rational for Becker’s assumption of altruism and explains why relatedness should determine its parameters. The present analysis is consistent with evolutionary and cultural influences on altruism are likely mutually reinforcing, rather than mutually exclusive or antagonistic in human society. This contrasts with some earlier criticisms of attempts to apply Kin-Selection to understanding human behaviour. These often proceed as though evidence (or even the logical possibility of) cultural factors favouring an altruistic trait negate or preclude Kin-Selection interpretations (for example, see Gould, 1977, pp. 255-258). |
| 13 | A caveat is that the strength of this effect depends on the shape of the returns-to-investment function: if returns to capital are strongly diminishing, the effect may be modest; if returns are constant or convex, the compounding will be much stronger. |
| 14 | In humans, if ability is operationalized through measures such as IQ or related cognitive tests, the model intersects with findings from behavioural genetics. Psychometric traits often show heritability estimates of roughly 50%, while variation attributable to the shared family environment is small (often 0–10%) (Roy & Roy, 2018). This pattern has sometimes been taken to imply that parental investment has little influence beyond genes (e.g., Caplan, 2011). However, the present model suggests an alternative interpretation: if parental altruism and investment strategies have been under stronger historical selection than cognitive ability genes, then variation in investment behaviour may already be compressed within a narrow range. At equilibrium, most observed variation in fitness capital will then appear to stem from ability, even though investment differences — especially at the tails of the distribution — can have profound consequences for descendants. |
| 15 | It is tempting to speculative that, if high-altruism couples tend to produce fewer offspring with high per-child investment, while low-altruism couples produce many offspring with little investment, such divergence in life-history strategies could resemble a form of disruptive selection within the population. In principle, such dynamics might reduce gene flow between extremes and thereby contribute to conditions favourable for reproductive isolation (cf. Lande, 1981; Maynard Smith, 1966). |
| 16 | On this view, the fitness capital of one individual may be the “extended phenotype” of genes in another who is providing the investment, because it makes the individual a better cooperator with the kin of the individual doing the investing. Notice this potentially opens another avenue facilitative of altruistic Green Beards. |
| 17 | A term meaning cooperative behaviour based on far-sighted self-intertest (Dennett, 2003). More generally, it may be worth further considering how conclusions from traditional models of reciprocal altruism are affected when reasonable assumptions are about fitness capital are included. For instance, when the magnitude of benefits from cooperative exchanges are improved with practice, such where the nature of relationships is not solely a matter of “you scratch my back, and I will scratch yours” but where cooperation makes individuals become better at scratching backs and perhaps more reliant on others so that their own backs get scratched. On this view, individuals may not only be motivated to cooperate by the prospect of punishment for failing to reciprocate, but also from seeing such mutual backscratching as a sort of investment process. In other words, early cooperative exchanges may not only build trust by establishing a track record by also by making each partner more valuable to the other as their skills can be expected to rise. Modelling by Becker on the social effects on activities has found there can be dramatic swings between different steady states of behaviour when many individuals become involved and this could be a rich area for further research to explore (general examples are given in Becker, 1996). Taken together, these extensions suggest that reciprocal altruism, when considered with fitness capital, might not only be about short-term exchanges but about long-run investments that shape both competence and interdependence across generations. This perspective invites new models that may reveal thresholds or tipping points in the evolution of cooperation. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



