Submitted:
10 February 2026
Posted:
11 February 2026
You are already at the latest version
Abstract
Demodex mites are common commensals of mammalian skin, but under certain conditions, they can cause severe skin diseases. This study analyzed the presence, diversity, and phylogenetic relationships of Demodex species in two wolf subspecies from southern Europe to determine whether species-level differences exist between wild and domestic canids after thousands of years of divergence. A total of 1,400 hair samples from 140 wolves were analyzed using qPCR targeting mitochondrial 16S rRNA and nuclear 18S rRNA genes. Overall, 37% of wolves were positive for Demodex DNA, with a higher prevalence in Italian (46%) than in Iberian (36%) wolves. The lip and chin areas were the most reliable sampling sites. Four Demodex species were identified in wolves: D. injai and D. canis (associated with dogs), and D. folliculorum and D. brevis (associated with humans). Co-infestations involving multiple Demodex species were recorded for the first time in wild canids. These results challenge the long-held belief of strict host specificity in Demodex mites. The discovery of Demodex species associated with both humans and dogs in wolves supports the idea that host-switching and ecological interactions have occurred throughout the evolution of canids and humans. Such cross-species transfers may have taken place during the early domestication of dogs. However, given the isolated history of the two southern wolf populations, it is more probable that these findings result from recent interspecific transmission events, likely facilitated by ecological overlap with domestic animals and human environments. Future genomic studies will be essential for clarifying the evolutionary relationships within the genus Demodex and its host associations.
Keywords:
Demodex
; Canis lupus
; wolf
; qPCR
; 18S rRNA gene
; 16S rRNA gene
1. Introduction
Demodex mites are considered normal inhabitants in mammalian skin [1,2,3]. Currently, more than 80 species of Demodex spp. have been described, genetically and/or morphologically, in more than 80 wild and domestic mammals, including humans, dogs, cats, deer, ferrets, otters, and mice [2,4,5,6,7,8,9,10,11,12,13,14]. Most mammals harbor Demodex mites on the skin without developing lesions or any other clinical signs [1,15]. Multiple pieces of evidence and studies indicate that the immune system controls Demodex populations on the skin [1,3,16]. Although Demodex mites are considered commensals in small numbers, when they proliferate, they may cause severe dermatitis, for instance, rosacea in humans or demodicosis in dogs and cats [17,18,19]. In almost all cases, proliferation results from a compromised immune system, as occurs in transgenic mice [16,20] or in dogs or humans receiving immunosuppressive medical treatment [3,21,22].
Genetic sequencing has proven highly effective for detecting and identifying Demodex mites in both healthy and diseased mammals [5,8,10,15]. Because of the sensitivity and specificity of genetic techniques, mites can be detected where non-molecular methods, such as microscopy or trichoscopy, fail to yield results [23,24]. Demodex mites have been detected, identified, and classified in humans, dogs, cats, mice, and ferrets using different genetic approaches such as the mitochondrial 16S rRNA gene region [5,7,8,25], and the nuclear 18S rRNA gene region [2,10,26,27]. Ravera et al. (2013) [15], using a fragment of the chitin synthase gene, concluded that Demodex DNA can be detected in the skin of any dog if 20 or more skin sites are sampled. In wolves, Demodex mites have been detected in fecal samples [28,29,30]. Since Demodex spp. are hair follicle mites, their presence in feces could be explained by accidental ingestion during grooming or from the prey itself. Nevertheless, it remains unknown which Demodex species are present in wolves' skin.
Research on Demodex mites is relevant not only to medicine but also to evolutionary biology. Their parasitic association with mammals may date back approximately 220 million years, coinciding with the emergence of hair follicles [9,31]. Roughly 100 million years ago, the clade Boreoeutheria diverged into two sister lineages: Euarchontoglires, which includes primates, and Laurasiatheria, which includes canids [32]. Demodex mites have been documented, both morphologically and genetically, in species from both groups, suggesting either a long-term stability of the hair follicle niche or a high capacity for mite transmission across and within placental mammals [5,6,8,10,27,33,34]. The genus Demodex therefore represents an exceptional case of parasitic parallelism [2,9,15,35].
The domestic dog originated from the gray wolf (Canis lupus) and is associated with Neolithic humans on most continents [36,37,38]. Axelsson et al. (2012) [39] suggest that dogs descended from scavenger wolves adapted to a starch-rich diet during the dawn of the agricultural revolution. However, it remains unclear where domestication occurred and whether it occurred once or multiple times worldwide. Archaeological records suggest different origins and domestication times: fossilized early dog remains have been found in Belgium (36,000 years ago; [40]), eastern Siberia (33,500 years ago; [41]) , and northern Israel (12,000 years ago; [42]). Nevertheless, specimens older than 14,000 years do not appear related to modern dogs [41]. Genetic studies also yield different results. Autosomal single nucleotide polymorphism (SNP) data indicate contributions from Middle Eastern and European wolves [43], while shotgun sequencing data suggest an origin in Eurasia, possibly southern Asia [44]. Wang et al. (2015) [45] propose that domestication began around 33,000 years ago in East Asia, followed by migrations toward the Middle East and Europe. More recently, Bergström et al. (2022) [46] found that modern dogs are genetically closer to ancient wolves from eastern Eurasia than to those from western Eurasia. However, Freedman et al. (2014) [47] found that no extant wolf lineage is more closely related to dogs, implying that an extinct wolf lineage was their ancestor. In any case, dog domestication can be traced back at least 15,000 years.
The gray wolf (Canis lupus) is listed on the IUCN Red List (2018) as “Least Concern” and is included in Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) (except for the populations of Bhutan, India, Nepal, and Pakistan, which are listed in Appendix I). Currently, five subspecies of Canis lupus are recognized in North America and seven in Eurasia. This classification is primarily based on genetic and morphological differences among populations, reflecting adaptations to distinct climates, latitudes, or habitats [48,49,50,51]. In Western Europe, the differentiation between the subspecies C. l. signatus of the Iberian Peninsula and C. l. italicus of the Italian Peninsula is the result of prolonged geographic isolation, which continues to this day [52,53,54,55].
Our study aims to determine whether there are species-level differences in Demodex mites between wild and domestic canids after this extensive period of divergence. For this purpose, we analyzed 1,400 hair samples from 140 wolves belonging to the two southern European subspecies, using two gene fragments, 16S rDNA and 18S rDNA, widely used for the molecular identification of Demodex mites.
2. Materials and Methods
2.1. Sampling
Hair samples from Iberian and Italian wolves were obtained from northern Spain, including Cantabria (126 samples), Aragón (1), Castilla y León (1), and Catalunya (1), and from the Piemonte region of northern Italy (11) (Table 1). One hundred twenty-seven wolves (61 females and 66 males) from Cantabria and Castilla y León were morphologically and genetically identified as Canis lupus signatus, and 13 wolves (6 females and 7 males) from Piemonte, Catalunya, and Aragón were identified as Canis lupus italicus (Table 1). Between 2017 and 2025, we collected a total of 1,400 hair samples from 140 wolf carcasses, with 10 samples per carcass obtained during necropsy examinations (Table 1). Five sites were from the face: periocular, lips, nose, chin, and entrance to the external ear canal, and five sites were from the rest of the body: dorsum, lumbar, abdomen, forelimb, and hindlimb. Using gloved hands and surgical mosquito forceps, hair was plucked in the direction of growth to include the hair bulb (root) for DNA extraction. Hair samples were stored at -20ºC until DNA extraction.
2.2. DNA Extraction and Real-Time PCR (qPCR) Amplification
Over the years, DNA from hair bulbs was extracted using two different methods (Table 1): Tris HCl approach, following Ravera et al. 2011 [4]; and PerkinElmer automated extraction, Chemagic 360 Instrument (Baesweiler, Germany), following the manufacturer’s protocol, including an incubation step at 56°C overnight and a final elution volume of 50 µL DNA. All extracted DNA was diluted 1:10 for qPCR amplification.
16S and 18S primers were used to amplify approximately 300-bp and 500-bp fragments of the mitochondrial 16S rRNA gene and the nuclear 18S rRNA gene, respectively [10]. All DNA samples were amplified by real-time qPCR and prepared under a laminar flow hood. Positive qPCR controls were obtained from known Demodex mite DNA [10]. Duplicates were amplified for each sample, and blank DNA extraction and negative qPCR controls were included to detect any qPCR exogenous DNA contamination. qPCR amplifications were performed in a QuantStudioTM 12K Flex Real-Time PCR System (Thermo Fisher Scientific). Amplicons were sequenced and purified when melting curves showed Tm = 75 ± 1 °C for the 16S rRNA gene and Tm = 83 ± 2 °C for the 18S rRNA gene, and amplification cycles had Cp < 37. Sequences were separated on an ABI PRISM 3730 automated sequencer (Thermo Fisher Scientific) according to the manufacturer’s instructions.
2.3. Genetic Variability and Phylogenetic Analysis
Genetic diversity in Demodex mites was analyzed using DNASP 5.10 [56]. For comparisons and phylogenetic analysis, we used mite sequences from wolves (those ending in UAB) and those available in GenBank. Sequences were analyzed using SEQSCAPE 2.1.1 software (Thermo Fisher Scientific) and were compared with the GenBank database (www.ncbi.nlm.nih.gov/BLAST). Phylogenetic analysis for the mitochondrial 16S rRNA gene was carried out using 44 sequences and 302 bp (gaps included). Phylogenetic analysis for the 18S rRNA gene was carried out using 45 sequences and 491 bp (gaps included). The trees were rooted using outgroups from the class Pycnogonida, Achelia hispida (FJ862845) and Ammothea sp. (FJ862841) for the 16S rRNA gene, and Achelia echinata (AF005438) and Callipallene sp. (AF005439) for the 18S rRNA gene. MODELTEST 3.7 [57] was applied to select the best evolutionary model among 56 models of evolution by the Akaike information criterion. The Bayesian program MrBayes v3.2.7 [58] was implemented to generate the phylogenetic tree with 1,000,000 Markov Chain Monte Carlo iterations and a burn-in rate of 25%. The bootstrap values show the repeatability and accuracy of the three [59]. To display the phylogenies, we used the program FigTree (http://tree.bio.ed.ac.uk/software/figtree/).
3. Results
3.1. Mite Prevalence in Wolves
We screened 1,400 hair samples from 140 wild wolves for mite DNA targeting the 16S rRNA and 18S rRNA genes. qPCR samples were considered positive when the melting curves (Tm) were close to Demodex control values (Tm = 75 ºC ± 0.5 ºC, 16S rDNA; Tm = 81.5 ºC ± 0.5 ºC, 18S rDNA), and at least one gene was successfully sequenced. The prevalence of Demodex mites in the hairs of 140 wolves was 37% (52/140). 46% of the Italian wolves (6/13) were positive for Demodex, while 36% were positive among Iberian wolves (46/127) (Table 1). No bias toward females and males (χ2 (1, N=140) = 1.02, p > 0.05) was observed among Demodex-positive wolves.
Sampling 10 sites, 101 out of 1,400 hair samples tested positive for Demodex by qPCR (Table 1). Twenty-three wolves (44%) were positive at one site, 19 wolves (36%) at two sites, 6 wolves (12%) at three sites, two wolves (4%) at four sites, and two wolves (2%) at five and nine sites, respectively. Demodex positivity was significantly higher in face samples (71%) compared to body samples (χ2 (1, N=1,400) = 19.73, p < 0.001). The most likely site to detect Demodex mites was the lip area (20%), followed by the chin area (18%). The least common sites were the lumbar (4%) and the dorsal (3%) areas.
Regarding DNA extraction methods, the PerkinElmer automated extraction was significantly more efficient (χ2 (1, N=1400) = 8.52, p < 0.05), with 71 positives out of 790 extractions (9%), compared to the This-HCL approach, which had 30 positives out of 610 extractions (5%). A significant disadvantage of the This-HCL method is that DNA is not purified, leading to fewer successful qPCRs due to increased inhibitor levels. However, its main advantages are speed and low cost.
3.2. Identification of Demodex spp.
Using the 18S rRNA gene, 83 samples were positive for Demodex spp., 21 samples could not be identified due to different mite types in the same sample (double sequence), and 88 samples were positive for other acarid mites (Table 1). Among the positive Demodex samples, we identified Demodex injai (52), Demodex canis (26), and Demodex folliculorum (5), but not Demodex brevis (Table 1 and Table 2). Using the 16S rRNA gene, we identified 101 positive samples, including D. injai (54), D. canis (30), D. folliculorum (12), and D. brevis (5) (Table 1 and Table 2).
Interestingly, the Demodex species commonly reported in dogs, such as D. canis and D. injai, were not detected in any of the Italian wolves examined. Instead, two species commonly associated with humans, D. folliculorum and D. brevis, were identified. Although differential sampling effort (13 Italian wolves versus 127 Iberian wolves) could potentially bias detection probabilities, the absence of D. canis and D. injai in Italian wolves was statistically significant (Fisher’s exact test, p = 0.015), suggesting that this absence is unlikely to be attributable solely to sampling effort. One possible explanation would have been putative reduced contact rates between Italian wolves and domestic dogs compared with Iberian wolves, but this is inconsistent with a recent study that found evidence of extensive wolf-dog hybridization in peninsular Italy [60].
In contrast, all four Demodex species, those usually reported in both dogs and humans, were identified in Iberian wolves. D. injai showed the highest prevalence, present in more than half of the individuals (64%), followed by D. canis (24%), while D. brevis and D. folliculorum had lower rates (8% and 4%, respectively). Additionally, co-infestation with D. canis and D. injai and D. canis and D. brevis were observed in three and one Iberian wolves, respectively, and D. brevis and D. folliculorum in one Italian wolf (Table 1 and Table 2).
3.3. 16S rDNA Sequence Variability and Phylogenetic Relationships
We identified 11 Demodex sequences (haplotypes) among 101 sequences of the mitochondrial 16S rRNA gene from 52 wolves (Table 1 and Table 2). The 11 Demodex haplotypes were submitted to GenBank because they had not been previously described in wolves and, in most cases, represented new records (Table 2).
They were identified as follows: 1) Demodex injai (three haplotypes): D.injai_UABW1 (N=19 sequences, identical to the sequence KT259449 from GenBank), D.injai_UABW2 (N=29 sequences), and D.injai_UABW3 (N=6 sequences); 2) Demodex canis (two haplotypes): D.canis_UABW1 (variant cornei) (N=27 sequences, identical to sequences JX390979 and MN161404 from GenBank), and D.canis_UABW2 (N=3 sequences, identical to several sequences from GenBank such as JF784000); 3) Demodex folliculorum (three haplotypes): D.folliculorum_UABW1 (N=7 sequences, identical to sequences HQ844221, FN424245, and FN424246 from GenBank), D.folliculorum_UABW2 (N=3 sequences), and D.folliculorum_UABW3 (N=2 sequences); and 4) Demodex brevis (three haplotypes): D.brevis_UABW1 (N=2 sequences), D.brevis_UABW2 (N=2 sequences), and D.brevis_UABW3 (N=1 sequence). Haplotypes from D. injai and D. canis were exclusive to the Iberian wolves, as well as haplotypes D.folliculorum_UABW3, D.brevis_UABW1, and D.brevis_UABW3 (Table 2). The exclusive haplotype found in Italian wolves was D.folliculorum_UABW2. Finally, haplotypes D.folliculorum_UABW1 and D.brevis_UABW2 were shared between wolf populations.
We conducted a phylogenetic analysis that included the 11 Demodex sequences, 31 sequences from the order Trombidiformes, and two outgroups obtained from GenBank (Fig 1). In total, we aligned 44 fragments of 302 bp each (gaps included). The Tamura-Nei substitution model, which assumes equal base frequencies and accounts for among-site rate heterogeneity using a gamma distribution (TrNef+G) was identified as the best-fit model for the 16S rRNA gene.
The 16S-Bayesian tree (Figure 1) showed two major splits among the Trombidiformes order (bootstrap value 100%). All our samples belong to the Anystina cohort, which includes the family Demodecidae. However, the Demodecidae sequences did not cluster by host species. For instance, D. brevis (host: human) was sister taxa with D. gatoi (host: cat), and D. folliculorum clade (host: human) was sister to the D. injai clade (host: dog) with strong bootstrap support (100%). These results suggest that the evolutionary relationships among Demodex mites do not strictly follow host phylogeny, indicating possible host-switching events over evolutionary time.
3.4. 18S rDNA Sequence Variability and Phylogenetic Relationship
We identified three Demodex sequences (haplotypes) among 83 sequences of the nuclear 18S rRNA gene from 40 wolves (Table 1 and Table 2). The three Demodex haplotypes were submitted to GenBank because they had not been previously described in wolves (Table 2). They were identified as Demodex injai, Demodex canis and Demodex folliculorum, and they have been described previously in dogs and humans (Table 1).
In addition to Demodex-positive samples, we sequenced 49 samples that amplified at Cp < 37 and melting curves Tm = 82 ºC + 3 ºC (Table 1). We amplified DNA from 11 mites belonging to the order Oribatida, 3 to the order Prostigmata, and one to the order Ixodida, the only hematophagous mite detected in an Iberian wolf (Table 1). The remaining mites were most likely soil-dwelling and not host-associated. Table 1 shows the sequences of the 15 mites, along with their percentage identity relative to the reference sequence in GenBank. The genus names used in the phylogenetic tree should be considered tentative, as the order-level classification is reliable, but species-level identification is not possible with the available data. These findings indicate that the 18S rDNA fragment is not an optimal marker for detecting Demodex in wolves.
The 18S-Bayesian tree (Figure 2) showed a clear split (bootstrap value 100%) between the Parasitiformes (Ixodida) and the Sarcoptiformes (Oribatida) and the Trombidiformes (Prostigmata) Orders. However, Oribatida did not form a distinct cluster, whereas Prostigmata split into several clusters, including the Eupodina and Anystina cohorts. The Demodecidae family constitutes a monophyletic group of the Anystina cohort that includes four large clusters: 1) the D. folliculorum cluster (host: human), 2) the D. canis cluster (host: dog), 3) the D. gatoi, D. cati, and an unnamed Demodex species in a cat (here referred to as D. felis) cluster (host: cat), and 4) the D. brevis cluster (host: human). D. injai appears to be more closely related to D. folliculorum than to D. canis; however, the bootstrap value (<70%) should be considered with caution.
4. Discussion
4.1. Detection, Prevalence, and Identification of Demodex spp.
Demodex mites were detected in over one-third of the wolves, with a slightly higher prevalence in Italian than in Iberian wolves and no sex bias, indicating widespread infestation in both populations. It is important to emphasize that the wolves in this study were wild. In a previous study, in which we examined five body regions from 14 Mexican (Canis lupus baileyi) and 8 Alaskan (Canis lupus occidentalis) captive wolves, we were unable to detect any positive cases with confidence [10]. The fact that these captive animals were vaccinated annually, fed regularly, and kept in sheltered conditions possibly reduced mite proliferation and, consequently, the likelihood of detection, despite our certainty that all wolves harbor Demodex mites in their pilosebaceous complexes. Notably, to detect 37% of wolves positive for Demodex, each wolf had to be sampled at 10 different skin sites, yielding a total of 1,400 samples, of which only 7% tested positive. Therefore, it is more effective to examine fewer wolves across multiple sites than to explore a larger number of wolves at a single site [1,10,15]. Based on our results, the number of sampling sites for detecting Demodex mites can be limited to the face, with the most suitable sites being the lips and chin. The dorsal and lumbar regions can be excluded. Reducing the number of sites decreases laboratory workload and overall costs. The resulting time and resources are recommended for use in a DNA extraction method incorporating a purification step, despite its higher cost relative to non-purified DNA extraction. Regarding the genetic markers, our results suggest that the 16S rRNA gene is more suitable for Demodex detection and phylogenetic inference than the 18S rRNA gene. Amplification of the 18S marker may lead to the unintended detection of non-target mites, such as soil mites, or, in some cases, to the failure to detect Demodex mites. The mitochondrial 16S marker, instead, showed greater sequence variability and, consequently, higher resolution at short evolutionary distances than the nuclear 18S marker. However, despite this increased variability, bootstrap support for the nodes separating the major Demodex lineages remained low, generally failing to reach the 0.9–0.95 threshold, except for the folliculorum–injai clade. A similar lack of strong support was observed in the 18S-based phylogeny. These results indicate that some uncertainty persists in the inferred relationships among Demodex species, highlighting the need for additional loci and increased sampling to robustly resolve species boundaries and evolutionary relationships within the genus.
4.2. Co-Infestations and Host Specificity Concerns
To date, no peer-reviewed studies have explicitly documented co-infestations of D. canis and D. injai, and D. canis and D. brevis, within canids. Most reports describe these species in isolation, as dogs exhibit clinical signs of demodicosis [1,61,62]. Similarly, studies conducted in healthy dogs have not reported co-infestations [15,23,24]. In our study, no known diseases were present in either population at the time, which supports previous findings that Demodex mites can be carried asymptomatically across various mammalian hosts [10,15,63]. The absence of confirmed co-occurrence in dogs may indicate underdiagnosis due to limitations in sampling strategies, as co-infestations of D. canis with D. folliculorum and D. brevis have been documented in two healthy cats [8]. Co-infestation of D. brevis and D. folliculorum has also been documented in humans, particularly in relation to dermatological and ophthalmological conditions, such as chronic blepharitis and rosacea [64,65,66]. These two species usually inhabit different ecological niches: D. folliculorum in hair follicles and D. brevis in sebaceous or meibomian glands, but they can coexist in the same individual [66], and their combined presence has been suggested as a potential factor in the development of these diseases [67,68].
The observation of interspecific co-infestations in some wolves, along with findings that the Iberian wolf harbours Demodex species found in dogs and humans, suggests substantial ecological plasticity and raises important questions about the strict host specificity paradigm. Previous studies have raised similar concerns by reporting the presence of D. canis not only in dogs and wolves, but also in a variety of hosts, including bats, ferrets, cats, and mice, and, exceptionally, in a single bear and a single human [8,10,13,34,69]. These findings support a revision of the host-parasite model for Demodex, proposing that at least some species, especially those infecting carnivores, may function as opportunistic generalists rather than obligate specialists. Molecular studies across a broader range of wild and domestic hosts are needed to further assess the extent of host specificity versus host plasticity in this group.
4.3. Inference on the Process of Dog Domestication
The identification of D. canis, D. injai, D. folliculorum, and D. brevis in wild wolves is a remarkable finding that challenges the traditional view of strict host specificity in Demodex mites. While D. canis and D. injai are typically associated with domestic dogs, and D. folliculorum and D. brevis with humans, their concurrent presence in wolves raises several questions regarding their evolutionary history and host associations. The detection of D. canis and D. injai in Iberian wolves is fully expected, whereas their absence from the skin of Italian wolves is unexpected. Although these mites should have been detected statistically, further analysis of samples would be necessary to confirm their presence in this wolf population. And even more surprising is the detection of human-associated Demodex in wolves. Two possible explanations can be proposed for this unexpected detection: 1, wolves served as a reservoir and potential source of transmission to humans, occurring through early interactions between canids and hominids during the process of dog domestication [2], and 2, a more plausible scenario involving ecological contact and host-switching events, given the recent isolation history of these two wolf populations. The Iberian and Italian wolf populations have been geographically isolated since the early 20th century, when wolves became extinct in France [55]. Since then, the population sizes of Spanish and Italian wolves have also declined dramatically, with minima reached in both countries during the 70s of the 20th century [54,55,70,71]. The presence of human-associated Demodex species in wolves may result from cross-species transmission, potentially facilitated by overlapping habitats, direct contact (e.g., in captivity or rehabilitation centers), or indirect contact through domestic dogs. However, to date, no reports have been published of dogs harboring human-associated Demodex on their skin. Future studies employing longer or more variable genetic markers alongside broader host and geographic sampling will be crucial to disentangle the evolutionary relationships of Demodex mites, clarify potential host-switching events, and enhance our understanding of their ecology and evolution in both wild and domestic hosts.
Author Contributions
Conceptualization, N.S., O.F. and L.F.; methodology, N.S., S.RO.; validation, N.S., O.F., S.RO., and L.F.; formal analysis, N.S., S.RO.; resources, M.F., L.R., R.V.; writing—original draft preparation, N.S.; writing—review and editing, N.S., M.F., L.R., O.F., R.V., S.RO., L.F.; supervision, O.F, L.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by the LoupO project (EFA354/19) of the European Interreg Program V-A Spain-France-Andorra (POCTEFA 2014-2020). This work was also supported by grants SEV-2015-0533 and by the CERCA Programme, Generalitat de Catalunya.
Institutional Review Board Statement
In Cantabria, wolves were classified as a hunting species under Annex I of the Cantabrian Law, 12/2006. From 2017 to 2020, wolves were removed under a species-control program implemented by “Subdirección General del Medio Natural” of the Government of Cantabria, in accordance with Laws 2/2017, 5/2018, and MED/5/2019, under expedient numbers CVE-2017-1827, CVE-2018-17138, and CVE-2019-2950, to reduce wolf attacks on livestock in conflict areas. From 2021 to 2025, wolves in the Iberian Peninsula were protected, and hunting was banned. Individuals necropsied from 2021 onwards died accidentally. Natural Environment agents transported the carcasses to the Wildlife Rescue Center of Cantabria for necropsies. The two wolves from Castilla y León and Catalunya died after being hit by a car. Both carcasses were transported by Natural Environment agents to the UAB for necropsy. Individuals from Italy (where wolf hunting is banned) died accidentally. Technicians from the European project Life12 NAT/IT/000807 WolfAlps moved carcasses to the Department of Veterinary Science (Università degli Studi di Torino) for necropsies. All procedures involving animal carcasses complied with national and regional regulations and did not involve the deliberate killing of animals for research purposes.
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data are within the paper. All new sequences are available from the GenBank database (accession numbers: PX232547, PX232548, PX232549, PX232550, PX232551, PX232552, PX232553, PX232554, PX232555, PX232556, PX232557, PX421029, PX421030, PX421031).
Acknowledgments
We thank Francesca Marucco, coordinator of the European Life WolfAlps, Gabriel Lampreave, and Ricard Casanovas from the Government of Catalonia (Generalitat de Catalunya), Santiago Lavin from the UAB, and Servicio de Conservación de la Naturaleza - Dirección General de Montes y Biodiversidad, Consejería de Desarrollo Rural, Ganadería, Pesca y Alimentación - from the Government of Cantabria for providing and authorizing the use of wolf carcasses for this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.:
Abbreviations
The following abbreviations are used in this manuscript:
| qPCR | Quantitative Polymerase Chain Reaction |
| Tm | Melting curve |
| Cp | Amplification cycle |
| SNP | Single Nucleotide Polymorphism |
| UAB | Universitat Autònoma de Barcelona |
| IUCN | International Union for Conservation of Nature |
| CITES | Convention on International Trade in Endangered Species of Wild Fauna and Flora |
References
- Ferrer, L.; Ravera, I.; Silbermayr, K. Immunology and Pathogenesis of Canine Demodicosis. Veterinary Dermatology 2014, 25, 427–e65. [Google Scholar] [CrossRef]
- Thoemmes, M.S.; Fergus, D.J.; Urban, J.; Trautwein, M.; Dunn, R.R. Ubiquity and Diversity of Human-Associated Demodex Mites. PLoS ONE 2014, 9, e106265. [Google Scholar] [CrossRef]
- El-Moamly, A.; El-Swify, O. Raising Awareness of Demodex Mites: A Neglected Cause of Skin Disease. Infection 2025, 53, 1273–1298. [Google Scholar] [CrossRef] [PubMed]
- Ravera, I.; Altet, L.; Francino, O.; Bardagí, M.; Sánchez, A.; Ferrer, L. Development of a Real-Time PCR to Detect Demodex Canis DNA in Different Tissue Samples. Parasitology Research 2011, 108, 305–308. [Google Scholar] [CrossRef]
- Zhao, Y.-E.; Wu, L.-P. Phylogenetic Relationships in Demodex Mites (Acari: Demodicidae) Based on Mitochondrial 16S rDNA Partial Sequences. Parasitology Research 2012, 111, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Izdebska, J.N.; Rolbiecki, L. Demodex Lutrae n. Sp. (Acari) in European Otter Lutra Lutra (Carnivora: Mustelidae) with Data from Other Demodecid Mites in Carnivores. Journal of Parasitology 2014, 100, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Silbermayr, K.; Horvath-Ungerboeck, C.; Eigner, B.; Joachim, A.; Ferrer, L. Phylogenetic Relationships and New Genetic Tools for the Detection and Discrimination of the Three Feline Demodex Mites. Parasitology Research 2014, 114, 747–752. [Google Scholar] [CrossRef]
- Ferreira, D.; Sastre, N.; Ravera, I.; Altet, L.; Francino, O.; Bardagí, M.; Ferrer, L. Identification of a Third Feline Demodex Species through Partial Sequencing of the 16S rDNA and Frequency of Demodex Species in 74 Cats Using a PCR Assay. Veterinary Dermatology 2015, 26, 239–e53. [Google Scholar] [CrossRef]
- Palopoli, M.F.; Fergus, D.J.; Minot, S.; Pei, D.T.; Simison, W.B.; Fernandez-Silva, I.; Thoemmes, M.S.; Dunn, R.R.; Trautwein, M. Global Divergence of the Human Follicle Mite Demodex Folliculorum: Persistent Associations between Host Ancestry and Mite Lineages. Proceedings of the National Academy of Sciences 2015, 201512609. [Google Scholar] [CrossRef]
- Sastre, N.; Francino, O.; Curti, J.N.; Armenta, T.C.; Fraser, D.L.; Kelly, R.M.; Hunt, E.; Silbermayr, K.; Zewe, C.; Sánchez, A.; et al. Detection, Prevalence and Phylogenetic Relationships of Demodex Spp and Further Skin Prostigmata Mites (Acari, Arachnida) in Wild and Domestic Mammals. PLoS ONE 2016, 11, e0165765. [Google Scholar] [CrossRef]
- Nashat, M.A.; Luchins, K.R.; Lepherd, M.L.; Riedel, E.R.; Izdebska, J.N.; Lipman, N.S. Characterization of Demodex Musculi Infestation, Associated Comorbidities, and Topographic Distribution in a Mouse Strain with Defective Adaptive Immunity. Comparative Medicine 2017, 67, 315–329. [Google Scholar]
- Ollé, J.; Ventura, J.; Feliu, C.; Foronda, P.; Francino, O.; Sastre, N. First Record of the Presence of Skin Mites (Demodex Musculi) in Wild House Mice from the Canary Islands (Spain). International Journal of Acarology 2020, 46, 283–286. [Google Scholar] [CrossRef]
- Esenkaya Taşbent, F.; Dik, B. A dog related Demodex spp. infestation in a student: a rare Demodex case. Mikrobiyol Bul 2018, 52, 214–220. [Google Scholar] [CrossRef]
- Ilie, M.S.; Imre, M.; Giubega, S.; Luca, I.; Florea, T.; Morariu, S. Feline Demodicosis Case Report—First Molecular Characterization of Demodex Mites in Romania. Pathogens 2021, 10, 1474. [Google Scholar] [CrossRef] [PubMed]
- Ravera, I.; Altet, L.; Francino, O.; Sánchez, A.; Roldán, W.; Villanueva, S.; Bardagí, M.; Ferrer, L. Small Demodex Populations Colonize Most Parts of the Skin of Healthy Dogs. Veterinary Dermatology 2013, 24, 168–172.e37. [Google Scholar] [CrossRef]
- Ricardo-Gonzalez, R.R.; Kotas, M.E.; O’Leary, C.E.; Singh, K.; Damsky, W.; Liao, C.; Arouge, E.; Tenvooren, I.; Marquez, D.M.; Schroeder, A.W.; et al. Innate Type 2 Immunity Controls Hair Follicle Commensalism by Demodex Mites. Immunity 2022, 55, 1891–1908.e12. [Google Scholar] [CrossRef] [PubMed]
- Crawford, G.H.; Pelle, M.T.; James, W.D. Rosacea: I. Etiology, Pathogenesis, and Subtype Classification. J. Am. Acad. Dermatol. 2004, 51, 327–341. [Google Scholar] [CrossRef]
- Chen, W.; Plewig, G. Human Demodicosis: Revisit and a Proposed Classification. Br J Dermatol 2014, 170, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Mueller, R.S.; Rosenkrantz, W.; Bensignor, E.; Karaś-Tęcza, J.; Paterson, T.; Shipstone, M.A. Diagnosis and Treatment of Demodicosis in Dogs and Cats: Clinical Consensus Guidelines of the World Association for Veterinary Dermatology. Vet Dermatol 2020, 31, 5–27. [Google Scholar] [CrossRef]
- Hill, L.R.; Kille, P.S.; Weiss, D.A.; Craig, T.M.; Coghlan, L.G. Demodex Musculi in the Skin of Transgenic Mice. Contemporary Topics in Laboratory Animal Science 1999, 38, 13–18. [Google Scholar]
- Gazi, U.; Taylan-Ozkan, A.; Mumcuoglu, K.Y. Immune Mechanisms in Human and Canine Demodicosis: A Review. Parasite Immunology 2019, 41, e12673. [Google Scholar] [CrossRef]
- Marquardt-Feszler, A.; Cekała, K.; Dębska-Ślizień, A.; Imko-Walczuk, B. Demodicosis among Immunocompromised Patients: A Review. Postepy Dermatol Alergol 2022, 39, 829–837. [Google Scholar] [CrossRef]
- Fondati, A.; De Lucia, M.; Furiani, N.; Monaco, M.; Ordeix, L.; Scarampella, F. Prevalence of Demodex Canis-Positive Healthy Dogs at Trichoscopic Examination. Vet. Dermatol. 2010, 21, 146–151. [Google Scholar] [CrossRef]
- Izdebska, J.N.; Fryderyk, S. Diversity of Three Species of the Genus Demodex (Acari, Demodecidae) Parasitizing Dogs in Poland. Polish J. of Environ. Stud 2011, 20, 565–569. [Google Scholar]
- Sastre, N.; Ravera, I.; Ferreira, D.; Altet, L.; Sánchez, A.; Bardagí, M.; Francino, O.; Ferrer, L. Development of a PCR Technique Specific for Demodex Injai in Biological Specimens. Parasitology Research 2013, 112, 3369–3372. [Google Scholar] [CrossRef]
- Zhao, Y.-E.; Wu, L.-P.; Hu, L.; Xu, Y.; Wang, Z.-H.; Liu, W.-Y. Sequencing for Complete rDNA Sequences (18S, ITS1, 5.8S, ITS2, and 28S rDNA) of Demodex and Phylogenetic Analysis of Acari Based on 18S and 28S rDNA. Parasitol Res 2012, 111, 2109–2114. [Google Scholar] [CrossRef] [PubMed]
- Yabsley, M.J.; Clay, S.E.; Gibbs, S.E.J.; Cunningham, M.W.; Austel, M.G. Morphologic and Molecular Characterization of a Demodex (Acari: Demodicidae) Species from White-Tailed Deer (Odocoileus Virginianus). ISRN Parasitology 2013, 2013, 1–7. [Google Scholar] [CrossRef]
- Sallows, T.A. Diet Preferences and Parasites of Grey Wolves in Riding Mountain National Park of Canada. Master of Environment Thesis, University of Manitoba, Winnipeg, Manitoba, 2007. [Google Scholar]
- Stronen, A.V.; Sallows, T.A.; Forbes, G.J.; Wagner, B.; Paquet, P.C. Diseases and Parasites in Wolves of the Riding Mountain National Park Region, Manitoba, Canada. J. Wildl. Dis. 2011, 47, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Cafiero, S.A.; Petroni, L.; Natucci, L.; Casale, L.; Raffaelli, M.; Baldacci, D.; Di Rosso, A.; Rossi, C.; Casulli, A.; Massolo, A.; et al. Parasite Diversity in Grey Wolves (Canis Lupus) from Tuscany, Central Italy: A Copromicroscopical Investigation. International Journal for Parasitology: Parasites and Wildlife 2025, 27, 101092. [Google Scholar] [CrossRef]
- Zhou, C.-F.; Wu, S.; Martin, T.; Luo, Z.-X. A Jurassic Mammaliaform and the Earliest Mammalian Evolutionary Adaptations. Nature 2013, 500, 163–167. [Google Scholar] [CrossRef]
- Murphy, W.J.; Eizirik, E.; O’Brien, S.J.; Madsen, O.; Scally, M.; Douady, C.J.; Teeling, E.; Ryder, O.A.; Stanhope, M.J.; de Jong, W.W.; et al. Resolution of the Early Placental Mammal Radiation Using Bayesian Phylogenetics. Science 2001, 294, 2348–2351. [Google Scholar] [CrossRef] [PubMed]
- Frank, L.A.; Kania, S.A.; Chung, K.; Brahmbhatt, R. A Molecular Technique for the Detection and Differentiation of Demodex Mites on Cats. Vet Dermatol 2013, 24, 367–e83. [Google Scholar] [CrossRef]
- Lankton, J.S.; Chapman, A.; Ramsay, E.C.; Kania, S.A.; Newkirk, K.M. Preputial Demodex Species in Big Brown Bats (Eptesicus Fuscus) in Eastern Tennessee. J. Zoo Wildl. Med. 2013, 44, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Palopoli, M.F.; Minot, S.; Pei, D.; Satterly, A.; Endrizzi, J. Complete Mitochondrial Genomes of the Human Follicle Mites Demodex Brevis and D. Folliculorum: Novel Gene Arrangement, Truncated tRNA Genes, and Ancient Divergence between Species. BMC Genomics 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.K.; Pedersen, N.C.; Jafarishorijeh, S.; Bannasch, D.L.; Ahrens, K.D.; Wu, J.-T.; Okon, M.; Sacks, B.N. Phylogenetic Distinctiveness of Middle Eastern and Southeast Asian Village Dog Y Chromosomes Illuminates Dog Origins. PLoS ONE 2011, 6, e28496. [Google Scholar] [CrossRef]
- Leonard, J.A.; Wayne, R.K.; Wheeler, J.; Valadez, R.; Guillén, S.; Vilà, C. Ancient DNA Evidence for Old World Origin of New World Dogs. Science 2002, 298, 1613–1616. [Google Scholar] [CrossRef]
- Savolainen, P.; Zhang, Y.; Luo, J.; Lundeberg, J.; Leitner, T. Genetic Evidence for an East Asian Origin of Domestic Dogs. Science 2002, 298, 1610–1613. [Google Scholar] [CrossRef]
- Axelsson, E.; Ratnakumar, A.; Arendt, M.-L.; Maqbool, K.; Webster, M.T.; Perloski, M.; Liberg, O.; Arnemo, J.M.; Hedhammar, Å.; Lindblad-Toh, K. The Genomic Signature of Dog Domestication Reveals Adaptation to a Starch-Rich Diet. Nature 2012, 495, 360–364. [Google Scholar] [CrossRef]
- Germonpré, M.; Sablin, M.V.; Stevens, R.E.; Hedges, R.E.M.; Hofreiter, M.; Stiller, M.; Després, V.R. Fossil Dogs and Wolves from Palaeolithic Sites in Belgium, the Ukraine and Russia: Osteometry, Ancient DNA and Stable Isotopes. Journal of Archaeological Science 2009, 36, 473–490. [Google Scholar] [CrossRef]
- Ovodov, N.D.; Crockford, S.J.; Kuzmin, Y.V.; Higham, T.F.G.; Hodgins, G.W.L.; van der Plicht, J. A 33,000-Year-Old Incipient Dog from the Altai Mountains of Siberia: Evidence of the Earliest Domestication Disrupted by the Last Glacial Maximum. PLoS ONE 2011, 6, e22821. [Google Scholar] [CrossRef]
- Davis, S.J.M.; Valla, F.R. Evidence for Domestication of the Dog 12,000 Years Ago in the Natufian of Israel. Nature 1978, 276, 608–610. [Google Scholar] [CrossRef]
- vonHoldt, B.M.; Pollinger, J.P.; Lohmueller, K.E.; Han, E.; Parker, H.G.; Quignon, P.; Degenhardt, J.D.; Boyko, A.R.; Earl, D.A.; Auton, A.; et al. Genome-Wide SNP and Haplotype Analyses Reveal a Rich History Underlying Dog Domestication. Nature 2010, 464, 898–902. [Google Scholar] [CrossRef]
- Skoglund, P.; Götherström, A.; Jakobsson, M. Estimation of Population Divergence Times from Non-Overlapping Genomic Sequences: Examples from Dogs and Wolves. Mol. Biol. Evol. 2011, 28, 1505–1517. [Google Scholar] [CrossRef]
- Wang, G.-D.; Zhai, W.; Yang, H.-C.; Wang, L.; Zhong, L.; Liu, Y.-H.; Fan, R.-X.; Yin, T.-T.; Zhu, C.-L.; Poyarkov, A.D.; et al. Out of Southern East Asia: The Natural History of Domestic Dogs across the World. Cell Res 2015. [Google Scholar] [CrossRef]
- Bergström, A.; Stanton, D.W.G.; Taron, U.H.; Frantz, L.; Sinding, M.-H.S.; Ersmark, E.; Pfrengle, S.; Cassatt-Johnstone, M.; Lebrasseur, O.; Girdland-Flink, L.; et al. Grey Wolf Genomic History Reveals a Dual Ancestry of Dogs. Nature 2022, 607, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Freedman, A.H.; Gronau, I.; Schweizer, R.M.; Ortega-Del Vecchyo, D.; Han, E.; Silva, P.M.; Galaverni, M.; Fan, Z.; Marx, P.; Lorente-Galdos, B.; et al. Genome Sequencing Highlights the Dynamic Early History of Dogs. PLoS Genet 2014, 10, e1004016. [Google Scholar] [CrossRef] [PubMed]
- Crandall, K.A.; Bininda-Emonds, O.R.; Mace, G.M.; Wayne, R.K. Considering Evolutionary Processes in Conservation Biology. Trends Ecol Evol 2000, 15, 290–295. [Google Scholar] [CrossRef]
- Carmichael, L.E.; Nagy, J.A.; Larter, N.C.; Strobeck, C. Prey Specialization May Influence Patterns of Gene Flow in Wolves of the Canadian Northwest. Mol Ecol 2001, 10, 2787–2798. [Google Scholar] [CrossRef]
- Geffen, E.; Anderson, M.J.; Wayne, R.K. Climate and Habitat Barriers to Dispersal in the Highly Mobile Grey Wolf. Mol Ecol 2004, 13, 2481–2490. [Google Scholar] [CrossRef] [PubMed]
- Musiani, M.; Leonard, J.A.; Cluff, H.D.; Gates, C.C.; Mariani, S.; Paquet, P.C.; Vilà, C.; Wayne, R.K. Differentiation of Tundra/Taiga and Boreal Coniferous Forest Wolves: Genetics, Coat Colour and Association with Migratory Caribou. Mol Ecol 2007, 16, 4149–4170. [Google Scholar] [CrossRef]
- Lucchini, V.; Galov, A.; Randi, E. Evidence of Genetic Distinction and Long-Term Population Decline in Wolves (Canis Lupus) in the Italian Apennines. Mol Ecol 2004, 13, 523–536. [Google Scholar] [CrossRef]
- Pilot, M.; Branicki, W.; Jędrzejewski, W.; Goszczyński, J.; Jędrzejewska, B.; Dykyy, I.; Shkvyrya, M.; Tsingarska, E. Phylogeographic History of Grey Wolves in Europe. BMC Evolutionary Biology 2010, 10, 104. [Google Scholar] [CrossRef]
- Sastre, N.; Vilà, C.; Salinas, M.; Bologov, V.V.; Urios, V.; Sánchez, A.; Francino, O.; Ramírez, O. Signatures of Demographic Bottlenecks in European Wolf Populations. Conserv Genet 2011, 12, 701–712. [Google Scholar] [CrossRef]
- Valiere, N.; Fumagalli, L.; Gielly, L.; Miquel, C.; Lequette, B.; Poulle, M.L.; Weber, J.M.; Arlettaz, R.; Taberlet, P. Long-Distance Wolf Recolonization of France and Switzerland Inferred from Non-Invasive Genetic Sampling over a Period of 10 Years. Animal Conservation 2003, 6, 83–92. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A Software for Comprehensive Analysis of DNA Polymorphism Data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the Model of DNA Substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Systematic biology 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Bull, J.J. An Empirical Test of Bootstrapping as a Method for Assessing Confidence in Phylogenetic Analysis. Systematic Biology 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Lorenzini, R.; Pizzarelli, A.; Attili, L.; Biagetti, M.; Sebastiani, C.; Ciucci, P. Genetic Evidence Reveals Extensive Wolf-Dog Hybridisation in Peninsular Italy: Warnings against Ineffective Management. Biological Conservation 2026, 313, 111615. [Google Scholar] [CrossRef]
- Saridomichelakis, M.; Koutinas, A.; Papadogiannakis, E.; Papazachariadou, M.; Liapi, M.; Trakas, D. Adult-Onset Demodicosis in Two Dogs Due to Demodex Canis and a Short-Tailed Demodectic Mite. J Small Anim Pract 1999, 40, 529–532. [Google Scholar] [CrossRef]
- Mueller, R.S.; Bensignor, E.; Ferrer, L.; Holm, B.; Lemarie, S.; Paradis, M.; Shipstone, M.A. Treatment of Demodicosis in Dogs: 2011 Clinical Practice Guidelines. Vet. Dermatol. 2012, 23, 86–96, e20-21. [Google Scholar] [CrossRef] [PubMed]
- Fryderyk, S.; Izdebska, J.N. Demodex Phylloides (Acari, Demodecidae) as a Specific Parasite of Sus Scrofa (Mammalia, Artiodactyla). Wiad Parazytol 2001, 47, 797–800. [Google Scholar]
- Biernat, M.M.; Rusiecka-Ziółkowska, J.; Piątkowska, E.; Helemejko, I.; Biernat, P.; Gościniak, G. Occurrence of Demodex Species in Patients with Blepharitis and in Healthy Individuals: A 10-Year Observational Study. Jpn J Ophthalmol 2018, 62, 628–633. [Google Scholar] [CrossRef]
- Pyzia, J.; Mańkowska, K.; Czepita, M.; Kot, K.; Łanocha-Arendarczyk, N.; Czepita, D.; Kosik-Bogacka, D.I. Demodex Species and Culturable Microorganism Co-Infestations in Patients with Blepharitis. Life 2023, 13, 1827. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.; Łanocha-Arendarczyk, N.; Pilarczyk, R.; Schneider-Matyka, D.; Kot, K.; Grzeszczak, K.; Pyzia, J.; Grochans, E. Ocular Symptoms in Pre- and Perimenopausal Woman Infected with Demodex Spp. Pathogens 2025, 14, 297. [Google Scholar] [CrossRef]
- Fromstein, S.R.; Harthan, J.S.; Patel, J.; Opitz, D.L. Demodex Blepharitis: Clinical Perspectives. Clin Optom (Auckl) 2018, 10, 57–63. [Google Scholar] [CrossRef]
- Zeytun, E.; Yazici, M. Human Demodex Mites (Acari: Demodicidae) as a Possible Etiological Factor in Rosacea: A Cross-Sectional Study from Turkey. Systematic and Applied Acarology 2024, 527–538. [Google Scholar] [CrossRef]
- Oleaga, Á.; Fayos, M.; Balseiro, A.; Borragán, S.; de Pedro, G.; Armenteros, J.Á.; Balsera, R.; Moreiro, M.; Sastre, N.; Ferrer, L. Demodicosis in a Free-Ranging Eurasian Brown Bear (Ursus Arctos Arctos) Cub in the Endangered Cantabrian Population, Spain. J Wildl Dis 2024, 60, 786–791. [Google Scholar] [CrossRef]
- Valverde, J. El Lobo Español. Montes 1971, 228–241. [Google Scholar]
- Zimen, E.; Boitani, L. Number and Distribution of Wolves in Italy. Zeitschrift für Säugetierkunde: im Auftrage der Deutschen Gesellschaft für Säugetierkunde e.V 1974, 40, 102--112. [Google Scholar]
Figure 1.
Phylogenetic analyses of Demodex spp. The tree was estimated using MrBayes based on aligned fragments of the 16S rRNA gene. Branch support is based on 10,000 bootstrap replications. The scale at the bottom represents genetic distances, expressed as the number of nucleotide substitutions per site.
Figure 1.
Phylogenetic analyses of Demodex spp. The tree was estimated using MrBayes based on aligned fragments of the 16S rRNA gene. Branch support is based on 10,000 bootstrap replications. The scale at the bottom represents genetic distances, expressed as the number of nucleotide substitutions per site.
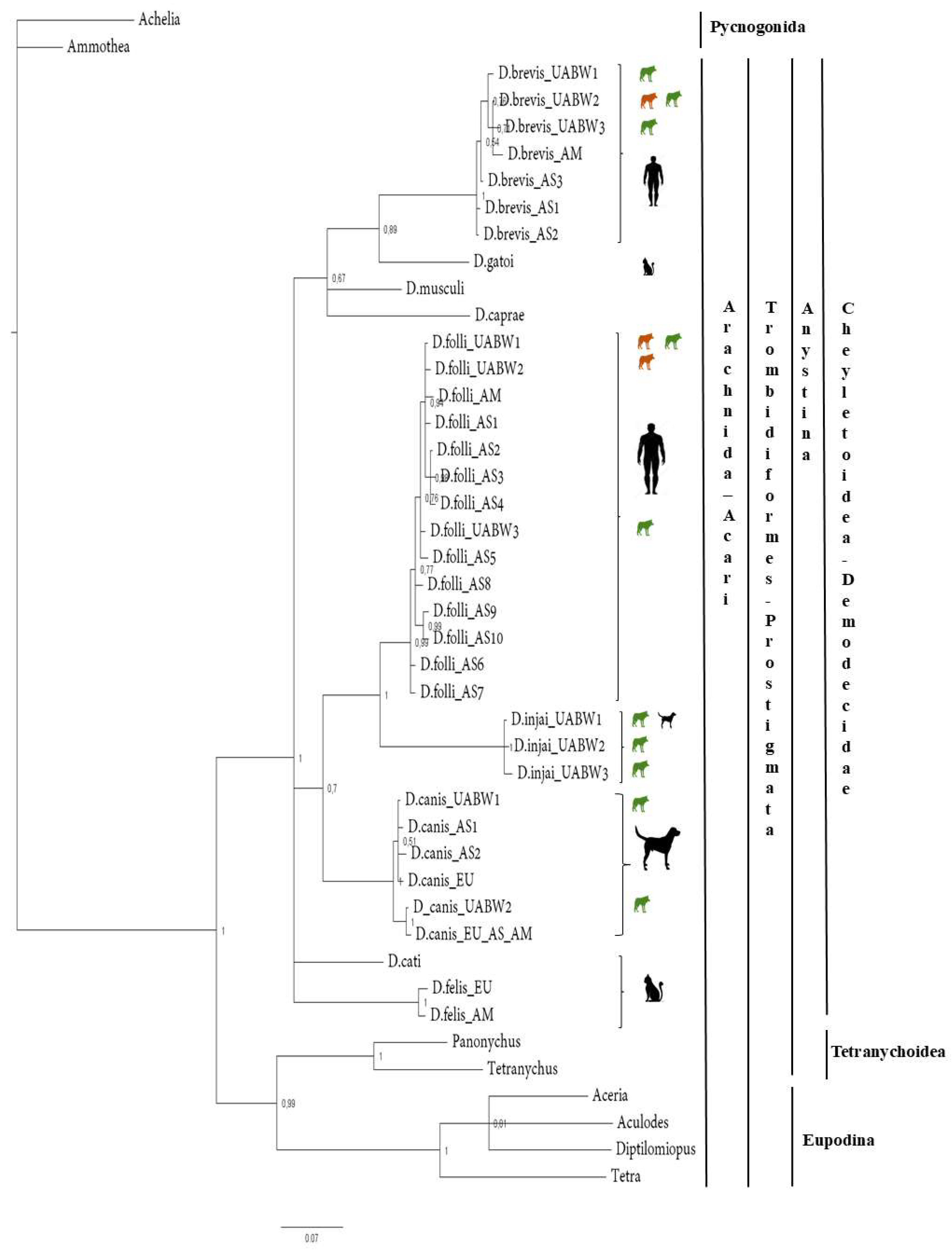
Figure 2.
Phylogenetic analyses of Demodex spp. The tree was estimated using MrBayes based on aligned fragments of the 18S rRNA gene. Branch support is based on 10,000 bootstrap replications. The scale at the bottom represents genetic distances, expressed as the number of nucleotide substitutions per site.
Figure 2.
Phylogenetic analyses of Demodex spp. The tree was estimated using MrBayes based on aligned fragments of the 18S rRNA gene. Branch support is based on 10,000 bootstrap replications. The scale at the bottom represents genetic distances, expressed as the number of nucleotide substitutions per site.
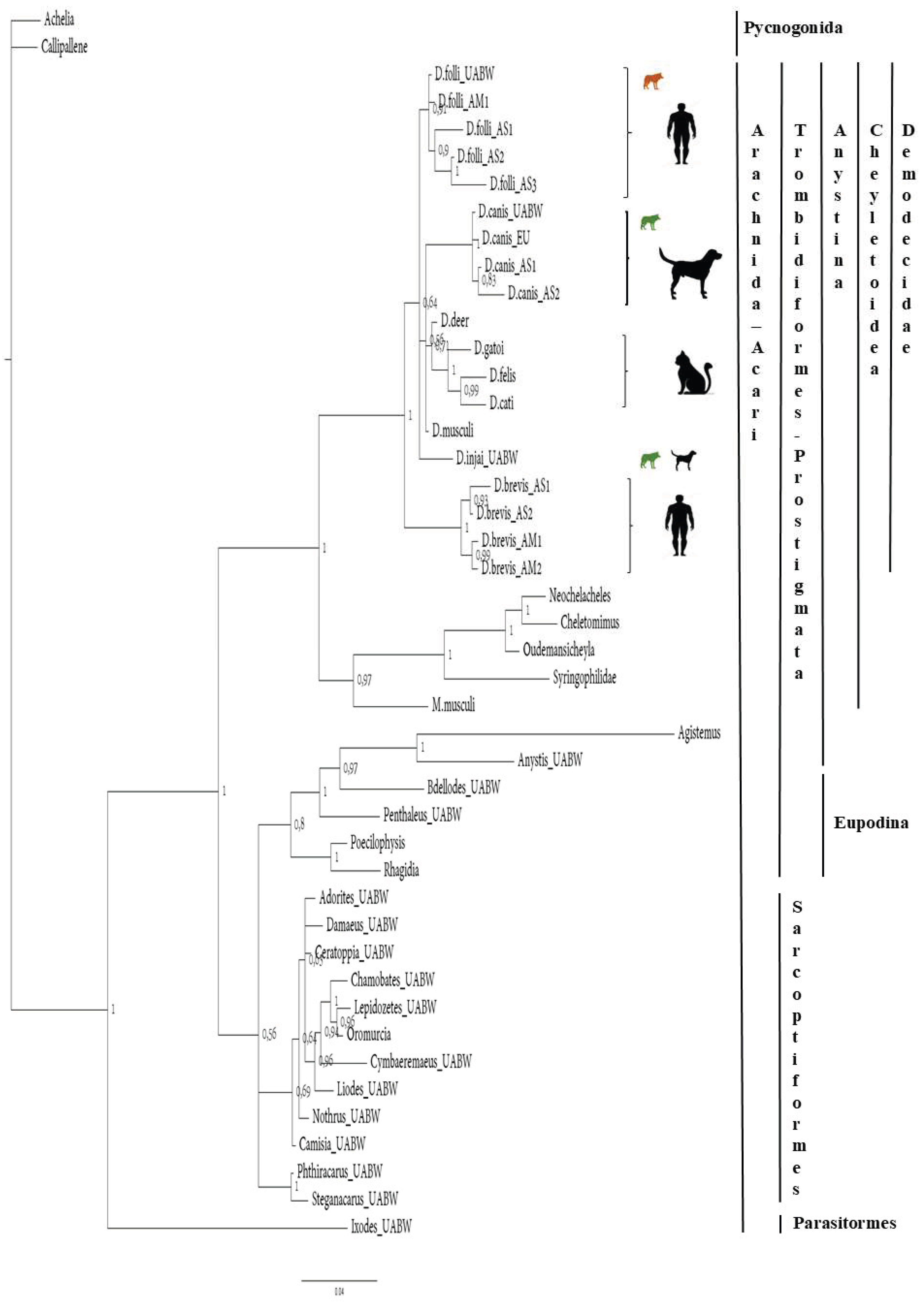

Table 1.
Sample collection
| Host | Sample | Coll. Year | Sex | Collection Place | Positive areas | Nº sites | Nº sites (no DMX) | Nº sites (DMX) | DNA extraction | 16S qPCR Detection | 18S qPCR Detection | 16S Tree name | GenBank (GB) number 18S | 18S GB Identity | 18S Tree name |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Wolf A | CL01 | 2017 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL02 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL03 | 2017 | M | Ca-Spain | N/Li/C/A | 4 | 4 | Tris HCL | + | + | D.canis_UABW1 | KC010484 | 100% | D.canis_UABW | |
| Wolf A | CL04 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL05 | 2017 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL06 | 2017 | M | Ca-Spain | P/H | 2 | 2 | Tris HCL | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL07 | 2017 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL08 | 2017 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL09 | 2017 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL10 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL11 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL12 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL13 | 2017 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL14 | 2017 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL15 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL16 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL17 | 2017 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL18 | 2017 | F | Ca-Spain | P/N/L | 3 | 3 | Tris HCL | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL19 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL20 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL21 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL22 | 2017 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL23 | 2017 | M | Ca-Spain | Li | 1 | 1 | Tris HCL | - | - | OR820194 | 100% | Phthiracarus_UABW | ||
| Wolf A | CL24 | 2017 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL25 | 2018 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL26 | 2018 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL27 | 2018 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL28 | 2018 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL29 | 2018 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL30 | 2018 | F | Ca-Spain | Li | 1 | 1 | Tris HCL | - | - | GQ864286 | 100% | Adoristes_UABW | ||
| Wolf A | CL31 | 2018 | M | Ca-Spain | Li | 1 | 1 | Tris HCL | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL32 | 2018 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL33 | 2018 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL34 | 2018 | M | Ca-Spain | Li | 1 | 1 | Tris HCL | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL35 | 2018 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL36 | 2018 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL37 | 2018 | M | Ca-Spain | Li | 1 | 1 | Tris HCL | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL38 | 2018 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL39 | 2018 | F | Ca-Spain | P/Fo | 2 | 2 | Tris HCL | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL40 | 2018 | F | Ca-Spain | Li | 2 | 1 | Tris HCL | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| CL40 | C | 1 | + | + | D.canis_UABW1 | KC010484 | 100% | D.canis_UABW | |||||||
| Wolf A | CL41 | 2018 | F | Ca-Spain | Li | 1 | 1 | Tris HCL | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL42 | 2018 | F | Ca-Spain | P | 1 | 1 | Tris HCL | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL43 | 2018 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL44 | 2018 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL45 | 2018 | M | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL46 | 2018 | M | Ca-Spain | P | 1 | 1 | Tris HCL | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL47 | 2018 | M | Ca-Spain | C | 2 | 1 | 1 | Tris HCL | + | - | D.canis_UABW1 | Z74479 | 99,81% | Ixodes_UABW |
| CL47 | Fo | 1 | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||||||||
| Wolf A | CL48 | 2018 | F | Ca-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL49 | 2018 | M | Ca-Spain | P | 2 | 1 | Perkins | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| CL49 | C | 1 | Perkins | - | - | HM070358 | 97,37% | Bdellodes_UABW | |||||||
| Wolf A | CL50 | 2018 | F | Ca-Spain | N | 1 | 1 | Perkins | + | + | D.injai_UABW3 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL51 | 2018 | F | CyL-Spain | None | Tris HCL | - | - | |||||||
| Wolf A | CL52 | 2019 | F | Ca-Spain | C | 2 | 1 | Perkins | + | + | D.injai_UABW3 | KU253789 | 100% | D.injai_UABW | |
| CL52 | P | 1 | Perkins | - | - | MK014972 | 100% | Camisia_UABW | |||||||
| Wolf A | CL53 | 2019 | F | Ca-Spain | Li | 2 | 1 | Perkins | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| CL53 | N | 1 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | |||||||
| Wolf A | CL54 | 2019 | F | Ca-Spain | L | 1 | 1 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||
| Wolf A | CL55 | 2019 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL56 | 2019 | F | Ca-Spain | P | 1 | 1 | Perkins | - | - | KR081607 | 100% | Damaeus_UABW | ||
| Wolf A | CL57 | 2019 | M | Ca-Spain | P | 1 | 1 | Perkins | - | - | EU432193 | 99,34% | Lepidozetes_UABW | ||
| Wolf A | CL58 | 2019 | M | Ca-Spain | D | 1 | 1 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||
| Wolf A | CL59 | 2019 | F | Ca-Spain | N | 1 | 1 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||
| Wolf A | CL60 | 2019 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL61 | 2019 | M | Ca-Spain | D | 3 | 1 | Perkins | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| CL61 | H | 1 | Perkins | + | + | D.canis_UABW1 | KC010484 | 100% | D.canis_UABW | ||||||
| CL61 | L | 1 | Perkins | - | - | KY922216 | 100% | Nothrus_UABW | |||||||
| Wolf A | CL62 | 2019 | M | Ca-Spain | Fo | 1 | 1 | Perkins | - | - | GQ864286 | 100% | Adoristes_UABW | ||
| Wolf A | CL63 | 2019 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL64 | 2019 | M | Ca-Spain | E/D/L | 3 | 3 | Perkins | - | - | OM249668 | 99,54% | Penthaleus_UABW | ||
| Wolf A | CL65 | 2019 | M | Ca-Spain | Li/N | 2 | 2 | Perkins | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL66 | 2019 | F | Ca-Spain | Li/Fo | 2 | 2 | 2 | Perkins | + | - | D.injai_UABW3 | EU432204 | 99,81% | Ceratoppia_UABW |
| Wolf A | CL67 | 2019 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL68 | 2019 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL69 | 2019 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL70 | 2019 | F | Ca-Spain | A | 1 | 1 | Perkins | - | - | GQ864286 | 100% | Adoristes_UABW | ||
| Wolf A | CL71 | 2019 | M | Ca-Spain | Li/N/C/L | 4 | 4 | Perkins | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL72 | 2019 | M | Ca-Spain | E/A | 2 | 2 | 2 | Perkins | + | - | D.folliculorum_UABW3 | Z74479 | 99,81% | Ixodes_UABW |
| Wolf A | CL73 | 2019 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL74 | 2019 | M | Ca-Spain | Fo | 1 | 1 | Perkins | + | + | D.canis_UABW1 | KC010484 | 100% | D.canis_UABW | |
| Wolf A | CL75 | 2019 | F | Ca-Spain | E | 1 | 1 | Perkins | + | - | D.brevis_UABW1 | ||||
| Wolf A | CL76 | 2019 | M | Ca-Spain | A | 2 | 1 | Perkins | + | - | D.brevis_UABW1 | ||||
| Wolf A | CL77 | 2020 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL78 | 2020 | F | Ca-Spain | Li/C/D/Fo | 4 | 4 | Perkins | - | - | GQ864286 | 100% | Adoristes_UABW | ||
| Wolf A | CL79 | 2020 | F | Ca-Spain | C/L | 3 | 2 | Perkins | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| CL79 | Fo | 1 | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||||||||
| Wolf A | CL80 | 2020 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL81 | 2020 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL82 | 2020 | F | Ca-Spain | N/E | 2 | 2 | Perkins | - | - | GQ864301 | 100% | Steganacarus_UABW | ||
| Wolf A | CL83 | 2020 | F | Ca-Spain | P/N | 2 | 2 | Perkins | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL84 | 2020 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL85 | 2020 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL86 | 2020 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL87 | 2020 | F | Ca-Spain | Li/C | 3 | 2 | Perkins | + | + | D.injai_UABW3 | KU253789 | 100% | D.injai_UABW | |
| CL87 | P | 1 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | |||||||
| Wolf A | CL88 | 2020 | F | Ca-Spain | P/Li | 2 | 2 | Perkins | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL89 | 2020 | M | Ca-Spain | P | 1 | 1 | Perkins | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL90 | 2020 | M | Ca-Spain | Li/N | 2 | 2 | Perkins | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL91 | 2020 | M | Ca-Spain | P/N/E/D/A/Fo/H | 7 | 7 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||
| Wolf A | CL92 | 2020 | F | Ca-Spain | P/N/C/E/D/A/Fo/H | 8 | 8 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||
| Wolf A | CL93 | 2020 | M | Ca-Spain | C/D/A | 3 | 3 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||
| Wolf A | CL94 | 2020 | F | Ca-Spain | P/Li/N/C/E/D/A/Fo/H | 9 | 9 | Perkins | + | + | D.canis_UABW1 | KC010484 | 100% | D.canis_UABW | |
| Wolf A | CL95 | 2020 | M | Ca-Spain | C | 8 | 1 | 1 | Perkins | + | - | D.canis_UABW1 | MK014972 | 100% | Camisia_UABW |
| CL95 | Fo | 1 | 1 | Perkins | + | - | D.brevis_UABW3 | MK014972 | 100% | Camisia_UABW | |||||
| CL95 | N/E/D/L/A/H | 6 | - | - | MK014972 | 100% | Camisia_UABW | ||||||||
| Wolf A | CL96 | 2020 | F | Ca-Spain | E | 6 | 1 | 1 | Perkins | + | - | D.folliculorum_UABW1 | EU432204 | 99,81% | Ceratoppia_UABW |
| CL96 | P/Li/C/L/Fo | 5 | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||||||||
| Wolf A | CL97 | 2021 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL98 | 2021 | M | Ca-Spain | E/D/L | 3 | 3 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||
| Wolf A | CL99 | 2021 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL100 | 2021 | M | Ca-Spain | N/C | 2 | 2 | Perkins | + | + | D.canis_UABW1 | KC010484 | 100% | D.canis_UABW | |
| Wolf A | CL101 | 2021 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL102 | 2021 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL103 | 2022 | M | Ca-Spain | P/Li/N/C/H | 5 | 5 | Perkins | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL104 | 2023 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL105 | 2023 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL106 | 2023 | F | Ca-Spain | C | 3 | 1 | Perkins | + | + | D.canis_UABW1 | KC010484 | 100% | D.canis_UABW | |
| CL106 | E/D | 2 | 2 | + | - | D.canis_UABW1 | GQ864286 | 100% | Adoristes_UABW | ||||||
| Wolf A | CL107 | 2023 | M | Ca-Spain | L/A/Fo | 3 | 3 | Perkins | + | + | D.canis_UABW1 | KC010484 | 100% | D.canis_UABW | |
| Wolf A | CL108 | 2023 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL109 | 2024 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL110 | 2024 | M | Ca-Spain | Fo/H | 2 | 2 | Perkins | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL111 | 2024 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL112 | 2025 | F | Ca-Spain | P/Li | 2 | 2 | Perkins | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL113 | 2025 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL114 | 2025 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL115 | 2025 | F | Ca-Spain | C | 1 | 1 | Perkins | + | - | D.brevis_UABW2 | ||||
| Wolf A | CL116 | 2025 | M | Ca-Spain | Li | 2 | 1 | Perkins | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| CL116 | A | 1 | + | + | D.canis_UABW1 | KC010484 | 100% | D.canis_UABW | |||||||
| Wolf A | CL117 | 2025 | F | Ca-Spain | C/D/L/A | 4 | 4 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||
| Wolf A | CL118 | 2025 | M | Ca-Spain | Li/N | 2 | 2 | Perkins | + | + | D.injai_UABW1 | KU253789 | 100% | D.injai_UABW | |
| Wolf A | CL119 | 2025 | M | Ca-Spain | C/Fo/H | 4 | 3 | Perkins | + | + | D.canis_UABW2 | KC010484 | 100% | D.canis_UABW | |
| CL119 | L | 1 | - | - | EU432204 | 99,81% | Ceratoppia_UABW | ||||||||
| Wolf A | CL120 | 2025 | M | Ca-Spain | E | 2 | 1 | Perkins | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| CL120 | N | 1 | Perkins | - | - | OR820194 | 100% | Phthiracarus_UABW | |||||||
| Wolf A | CL121 | 2025 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL122 | 2025 | M | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL123 | 2025 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL124 | 2025 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL125 | 2025 | F | Ca-Spain | None | Perkins | - | - | |||||||
| Wolf A | CL126 | 2025 | F | Ca-Spain | N/C | 3 | 2 | Perkins | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| CL126 | D | 1 | Perkins | - | - | EU432204 | 99,81% | Ceratoppia_UABW | |||||||
| Wolf A | CL127 | 2025 | M | Ca-Spain | H | 2 | 1 | Perkins | + | + | D.injai_UABW2 | KU253789 | 100% | D.injai_UABW | |
| CL127 | Fo | 1 | Perkins | - | - | HM070361 | 100% | Anystis_UABW | |||||||
| Wolf B | CL128 | 2017 | F | T-Italy | N/H | 2 | 2 | Tris HCL | - | - | LC817364 | 99,78% | Cymbaeremaeus_UABW | ||
| Wolf B | CL129 | 2017 | M | Al-Italy | Li | 1 | 1 | Tris HCL | - | - | HM070361 | 100% | Anystis_UABW | ||
| Wolf B | CL130 | 2017 | M | Cu-Italy | Li/N | 3 | 2 | Tris HCL | + | + | D.folliculorum_UABW2 | KF745889 | 100% | D.folliculorum_UABW | |
| CL130 | C | 1 | Tris HCL | + | - | D.brevis_UABW2 | |||||||||
| Wolf B | CL131 | 2017 | M | Cu-Italy | None | Tris HCL | - | - | |||||||
| Wolf B | CL132 | 2017 | F | T-Italy | None | Tris HCL | - | - | |||||||
| Wolf B | CL133 | 2017 | F | Cu-Italy | None | Tris HCL | - | - | |||||||
| Wolf B | CL134 | 2017 | F | T-Italy | N/C/E | 3 | 3 | Tris HCL | + | + | D.folliculorum_UABW1 | KF745889 | 100% | D.folliculorum_UABW | |
| Wolf B | CL135 | 2017 | M | V-Italy | H | 1 | 1 | Tris HCL | + | - | D.folliculorum_UABW1 | ||||
| Wolf B | CL136 | 2017 | M | T-Italy | Li | 1 | 1 | Tris HCL | + | - | D.folliculorum_UABW1 | ||||
| Wolf B | CL137 | 2018 | F | Cu-Italy | E | 2 | 1 | 1 | Tris HCL | + | - | D.folliculorum_UABW2 | EU432204 | 99,81% | Ceratoppia_UABW |
| CL137 | P | 1 | Tris HCL | - | - | AF022035 | 99,75% | Liodes_UABW | |||||||
| Wolf B | CL138 | 2018 | F | Cu-Italy | None | Tris HCL | - | - | |||||||
| Wolf B | CL139 | 2018 | M | C-Spain | C | 8 | 1 | 1 | Tris HCL | + | - | D.folliculorum_UABW1 | EU432190 | 99,41% | Chamobates_UABW |
| CL139 | E/D | 2 | Tris HCL | - | - | EU432204 | 99,81% | Ceratoppia_UABW | |||||||
| CL139 | P/Li/N/L/A | 5 | Tris HCL | - | - | EU432190 | 99,41% | Chamobates_UABW | |||||||
| Wolf B | CL140 | 2023 | M | Ar-Spain | None | Perkins | - | - |
Wolf A (Canis lupus signatus); Wolf B (Canis lupus italicus); F (Female); M (Male); DMX (Demodex). P: Periocular; Li: Lips; N: Nose; C: Chin; E: Ear; D: Dorsum; A: Abdominal; L: Lumbar; Fo: Forelimb; H: Hindlimb. Ar: Aragón; Ca: Cantabria; CyL: Castilla y León; C: Catalunya; Cu: Cuneo; T: Torino; V: Vercelli.
Table 2.
Demodex spp. detected by site using the 18S rRNA and 16S rRNA genes with the corresponding GenBank accession number, and Demodex variants identified, and the number of positive wolves using the 16S rRNA gene.
Table 2.
Demodex spp. detected by site using the 18S rRNA and 16S rRNA genes with the corresponding GenBank accession number, and Demodex variants identified, and the number of positive wolves using the 16S rRNA gene.
| Demodex sp. | N sites - 18S | GENBANK-18S | N Sites - 16S | Variants - 16S | GENBANK- 16S | N Sites - 16S | N Iberian wolf - 16S | N Italian wolf - 16S |
|---|---|---|---|---|---|---|---|---|
| D.brevis | 0 | - | 5 | D.brevis_UABW1 | PX232547 | 2 | 2 | |
| D.brevis_UABW2 | PX232548 | 2 | 1 | 1(5) | ||||
| D.brevis_UABW3 | PX232549 | 1 | 1(3) | |||||
| D.folliculorum | 5 | PX421031 | 12 | D.folliculorum_UABW1 | PX232550 | 7 | 1 | 4 |
| D.folliculorum_UABW2 | PX232551 | 3 | 2(5) | |||||
| D.folliculorum_UABW3 | PX232552 | 2 | 1 | |||||
| D.canis | 26 | PX421029 | 30 | D.canis_UABW1 | PX232553 | 27 | 11(1)(2)(3)(4) | |
| D.canis_UABW2 | PX232554 | 3 | 1 | |||||
| D.injai | 52 | PX421030 | 54 | D.injai_UABW1 | PX232555 | 19 | 12(1)(4) | |
| D.injai_UABW2 | PX232556 | 29 | 16(2) | |||||
| D.injai_UABW3 | PX232557 | 6 | 4 | |||||
| Total | 83 | 101 | 101 | 50 | 7 |
(1-5) co-infested wolves.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



