Submitted:
08 February 2026
Posted:
10 February 2026
You are already at the latest version
Abstract
Understanding how lifestyle factors influence the dynamic organization of intrinsic brain networks in young adulthood is critical for identifying mechanisms that support cognitive health during a formative developmental period. In this study, we examined whether an 8-week physical activity and cognitive training intervention altered dynamic functional network connectivity (dFNC) patterns in undergraduate students and how these neural dynamics related to physical activity levels, sedentary behavior, and cognitive performance. Resting-state fMRI data were decomposed using a constrained ICA framework to extract 53 intrinsic connectivity networks, from which 10 dynamic connectivity states were identified and individualized via constrained dynamic double functional independent primitives (c-ddFIPs). We quantified state occupancy, convergence, and divergence to characterize network flexibility. Occupancy analyses showed modest but consistent associations linking greater physical activity with increased time in integrative, higher-order states (especially states 6 and 7) and reduced time in segregated or sensory-weighted states. Convergence and divergence analyses further revealed that physically active individuals demonstrated stronger differentiation between integrative and low-engagement states, whereas sedentary behavior corresponded to greater similarity among segregated configurations. Cognitive measures—particularly working memory—showed parallel relationships, aligning improved performance with more flexible and well-differentiated dynamic patterns. Together, these findings suggest that physical activity in young adults is associated with enhanced neural flexibility, characterized by greater engagement and differentiation of integrative connectivity states that support executive and other cognitive functions.
Keywords:
dynamic functional connectivity (dFNC)
; independent component analysis (ICA)
; physical activity
1. Introduction
The human brain exhibits fluctuating patterns of temporal synchronicity across regions that underpin essential processes such as cognition, perception, and action. In young adults, fluctuations in synchronous activity, also known as dynamic functional connectivity, play a pivotal role in supporting neural flexibility, enabling adaptive responses to environmental demands, working memory maintenance, and decision-making [1,2,3]. Emerging evidence suggests that lifestyle interventions, including physical exercise, can modulate functional connectivity to enhance cognitive performance and overall brain health in young adults [4,5,6,7]. Young adulthood is a critical window for lifestyle and habit formation; ; thus, targeted strategies to promote engagement in brain healthy activities during this critical developmental window could promote lifelong maintenance. Given that, young adults, particularly minority undergraduate students, report levels of stress and anxiety that interfere with daily activities, implementation of such strategies may also offset stress-related decrements in functional connectivity within brain networks supporting cognition and stress regulation. Neuroimaging offers one way to measure the degree to which physical activity levels and self-reported stress relate to neurobiological profiles and to gage whether brain health interventions have measurable impacts on brain regions implicated in modulating stress responses.
Dynamic functional network connectivity (dFNC) captures the time-varying interactions between brain regions during resting-state functional magnetic resonance imaging (rs-fMRI). Data obtained using dFNC can provide insights into neural flexibility and the brain’s ability to adapt to changing cognitive demands in young adults [3]. Unlike static connectivity measures, dFNC reveals transient states of brain organization that correlate with cognitive processes such as attention, working memory, and executive function, which are critical during emerging adulthood, when neural plasticity peaks [8].
Constrained dynamic double functional independent primitives (c-ddFIPs) represent a novel analytical framework for dFNC data that quantifies individualized dynamic convergence and divergence in functional networks by extracting subject-specific connectivity patterns from group-level priors, enabling the identification of discrete states (there are 10 in the current study) that reflect differential engagement and transitions linked to behavioral outcomes [9,10,11]. These measures are particularly valuable for assessing how interventions might enhance neural flexibility in healthy young populations. This approach was selected over more traditional dynamic connectivity methods because it provides individualized dynamic connectivity estimates while maintaining consistency with group-level templates. In doing so, c-ddFIPs improve sensitivity to subtle, person-specific shifts in network dynamics—an essential feature when evaluating intervention-related changes in young adult populations, where variability in neural flexibility is a key outcome of interest.
Lifestyle interventions, such as physical exercise and cognitive training, can induce plasticity in brain networks, leading to enhanced functional connectivity and improved cognitive outcomes in healthy young adults. Physical exercise, in particular, has been shown to modulate functional connectivity [12], promoting greater integration within networks involved in attention and executive function [13,14], while also supporting overall brain health through increased cerebral blood flow [15] and neurotrophic factors [16,17].
Furthermore, recent research links brain flexibility to cognitive performance and physical activity levels. Divergence measures, which capture differential engagement patterns across dFNC states, have been associated with enhanced cognitive functions such as attention and working memory [10,18], often showing focused effects in specific state pairs that reflect task-related neural adaptability [19]. In contrast, physical activity has been linked to broader, distributed changes in dynamic functional connectivity, with higher levels of exercise associated with increased network reconfiguration and flexibility, promoting greater state differentiation and integration across large-scale brain systems [20]. Occupancy analyses in the literature indicate that time spent in adaptive, integrative dFNC states correlates positively with better cognitive outcomes on tasks like attention control and working memory [21], while excessive occupancy in segregated states is tied to reduced cognition [22]. These dynamic patterns illustrate how exercise may optimize neural connectivity to bridge physical activity and cognitive performance, offering insights for interventions targeting healthy young adults.
We investigated the effects of exercise, as well as cognitive interventions, on dynamic brain network states in a cohort of young adults, with a focus on associations with cognitive performance (e.g., working memory and attention), physical activity levels, and perceived stress. Drawing from longitudinal data, including rs-fMRI scans at baseline (T1) and post-intervention (T2), cognitive performance (dimensional card sorting task, Flanker task, list sorting working memory) and dynamic functional connectivity metrics, the study examined whether and how exercise interventions influence dFNC divergence and state occupancy. This approach is particularly relevant for young adults, such as college students, who often face high stress and sedentary lifestyles that can impair neural dynamics and cognitive performance, essential for academic success. We hypothesized that greater physical activity levels during the intervention period would relate to greater positive divergences in dFNC states linked to cognitive measures and increased occupancy in adaptive/integrative states (e.g., 4–6, 9).
2. Materials and Methods
2.1. Participants and Behavioral Measures
Participants were drawn from the Healthy Student Brain (HSB) project, a longitudinal intervention study at Georgia State University examining how aerobic exercise and online cognitive training influence neural dynamics, cognition, and well-being in sedentary undergraduate students. The full sample consisted of 103 students (ages 17–33), and a neuroimaging subsample of 32 individuals (ages 18–33) completed MRI scanning at baseline and following the 8-week intervention. All participants provided informed consent, and study procedures were approved by the Georgia State University Institutional Review Board.
Table 1 summarizes the demographic characteristics of both the full study cohort and the MRI subsample. Consistent with the university population, the sample was racially and ethnically diverse and predominantly female. The MRI subsample was comparable to the full sample in age and gender distribution, supporting its representativeness for the current neuroimaging analyses.
2.2. Exercise and Brain Training Intervention
Participants were pseudo-randomly assigned to one of 4 intervention groups; exercise, online brain health training, Exercise + online training, and control condition balanced for gender and ethnicity. To maximize observed effect of the interventions, only the exercise + online training group and controls underwent an MRI scan. In the exercise group, participants were asked to perform 30 minutes of moderate to vigorous physical activity at least three times per week. Participants were not restricted to particular exercise but instead were informed about types of exercises that increase heart rate, heart rate zones and ways to gauge moderate and vigorous activity based on perceived breathlessness and measures collected by the FitBit. In the brain health training condition, participants underwent brain health training created by the UT Dallas Center for BrainHealth. The program consists of short, self-paced digital modules that teach evidence-informed cognitive strategies aimed at strengthening attention, reasoning, mental flexibility, and overall brain wellness. No data from this training were analyzed in the present study, only NIH toolbox cognitive measure taken pre-and post intervention. For the purposes of this analysis we were interested in minutes of exercise and sedentary behavior across the study and we collapsed across intervention groups.
2.2.1. Measures
Participants were continuously monitored using a Fitbit Inspire 2 device worn throughout the 8-week study, yielding objective estimates of daily moderate-to-vigorous physical activity , sleep, and heart rate. Data were collected and managed using Fitabase (citation). Physical activity was measured continuously using Fitbit wearable devices, which classify each minute of movement into standardized intensity categories using triaxial accelerometer data and validated proprietary algorithms. For each participant, we extracted daily totals of Sedentary, Lightly Active, Fairly Active, and Very Active Minutes. Sedentary Minutes represent minimal movement consistent with sitting or reclining. Lightly Active Minutes reflect low-intensity movement (e.g., slow walking or routine daily tasks). Fairly Active Minutes correspond to moderate-intensity activity (approximately 3–6 METs; e.g., brisk walking), and Very Active Minutes represent vigorous-intensity movement (> 6 METs; e.g., running or high-effort exercise). When heart rate was available, Fitbit integrates optical heart-rate data to refine the activity-intensity classification. These intensity minutes are widely used in physical activity research and have demonstrated acceptable validity for estimating free-living activity in young adults. We summed the failry+very active minutes as both categories represented acceptable heart rate zones that we defined as exercise in our instruction to participants. We summed across each participant for the duration of these study and used these outcome measures (total sedentary minutes, and fairly+very active minutes) in our statistical analyses. Initial descriptive analysis of our cohort indicated that controls did in fact exercise even when they were instructed not to. When investigating how many weeks participants met our recommended three times per week of moderate to vigorous exercise, all controls exceeded this recommendation for more than 1 week during the intervention period. Thus, as previously mentioned, we collapsed across all intervention groups to investigate a dose dependent relationship between sedentary behavior and connectivity metrics, and physical activity and connectivity metrics.
Behavioral assessments were collected both online and in person pre and post intervention. Participants completed validated self-report measures of stress, depression, and anxiety, including the Perceived Stress Scale (PSS) and Patient-reported outcomes measurement information system (PROMIS) emotional health domains. Cognitive performance was assessed using the NIH Toolbox Dimensional Change Card Sort, Flanker, and List Sorting Working Memory tasks, providing measures of cognitive flexibility, inhibitory control, and working memory capacity, respectively.
2.3. MRI Data Acquisition and Preprocessing
Resting-state functional magnetic resonance imaging (fMRI) data were collected at the Georgia State/Georgia Tech Center for Advanced Brain Imaging (CABI) and the GSU Urban Life Building during baseline and post-intervention visits. Each scanning session included a high-resolution anatomical scan and a 4D resting-state fMRI acquisition. Standard MRI safety procedures were followed, and participants were included in scanning only if they met all MRI eligibility criteria.
Preprocessing followed the standardized NeuroMark pipeline, which uses adaptive spatially constrained independent component analysis (ICA) to extract reproducible intrinsic connectivity networks (ICNs). For each participant, 53 ICNs were identified and categorized into seven functional domains: subcortical, auditory, sensorimotor, visual, cognitive control, default mode, and cerebellar networks. The pipeline included motion correction, spatial normalization, artifact reduction, and template-guided ICN estimation to maximize cross-subject comparability. The resulting ICN time courses were used to compute both static and dynamic functional network connectivity metrics, forming the basis for subsequent analyses of time-varying network organization.
2.4. Extraction of Intrinsic Connectivity Networks Using Spatially Constrained ICA
To characterize large-scale functional brain networks consistently across participants, we employed a spatially constrained independent component analysis (ICA) approach guided by the Neuromark_fMRI_1.0 template [23]. Spatially constrained ICA extends traditional ICA by incorporating robust spatial priors that reduce component-order ambiguity and enhance cross-subject correspondence. This approach embeds data-driven decomposition within a framework of independently derived network templates, enabling extraction of individualized intrinsic connectivity networks (ICNs) while retaining stability and comparability across participants.
All analyses were implemented using the GIFT toolbox at http://trendscenter.org/software/gift. For each resting-state fMRI scan, 53 ICNs were reconstructed based on NeuroMark’s standardized atlas, which has been validated for detecting reproducible functional networks across diverse cohorts and clinical populations. These networks were grouped into seven functional domains– subcortical (SC), auditory (AUD), visual (VIS), sensorimotor (SM), cognitive control (CC), default mode (DM), and cerebellar networks (CB)– providing broad coverage of large-scale functional architecture.
2.5. Dynamic Functional Network Connectivity Estimation
Temporal variations in connectivity were quantified using a dFNC framework. For each participant and session (baseline and follow-up), the 53 ICN time courses were segmented using a 45 TR sliding window (rectangular window, 1 TR step size). Pearson correlation coefficients were calculated for each window, producing a sequence of connectivity matrices that capture ongoing fluctuations in inter-network coupling.
To focus on dynamic fluctuations rather than static baseline connectivity, each windowed FNC matrix was demeaned. This normalization step removes the static global connectivity pattern, thereby enhancing sensitivity to transient shifts in network organization. The participant-specific dFNC matrices from all individuals and visits were concatenated to form a single group-level dataset representing population-wide temporal variability.
2.6. Identification of Group-Level Dynamic States (ddFIPs)
The aggregated dFNC dataset was decomposed using blind ICA to derive a set of group-level dynamic states, referred to as dynamic double functional independent primitives (ddFIPs). This analysis yielded 10 partially overlapping connectivity states, each reflecting a recurring pattern of network interactions commonly expressed across individuals as shown in Figure 1. These ddFIPs serve as functional connectivity "templates" that summarize population-level dynamic configurations and form the basis for individualized state reconstruction.
These states encapsulate distinct modes of large-scale network integration and segregation and have previously been linked to cognitive performance and clinical characteristics in both healthy and psychiatric populations.
2.7. Subject-Level Reconstruction of Constrained dFNC States (c-ddFIPs)
To obtain individualized estimates of dynamic connectivity patterns, the 10 ddFIPs were used as spatial priors in a constrained ICA back-reconstruction procedure. This approach enforces alignment between individual-level connectivity states and the group-derived templates while preserving subject-specific variability in amplitude and expression.
For each participant, the constrained decomposition yielded time-resolved state contributions– referred to as constrained dynamic double functional independent primitives (c-ddFIPs)– which provide individualized fingerprints of dynamic connectivity. This strategy enhances interpretability by ensuring that the connectivity patterns for each participant map onto a shared set of neurobiologically meaningful states.
A final calibration step was applied to each participant’s reconstructed dFNC time series. Specifically, individual connectivity patterns were regressed onto predictors formed by the outer products of participant-specific state-mixing profiles, producing beta-weighted, variance-scaled time courses. These calibrated trajectories represent standardized measures of state engagement that are directly comparable across individuals and groups.
Following reconstruction, visit 1 profiles were regressed out from visit 2 for each participant to isolate intervention-related changes. Only participants with complete second-visit data were included in downstream analyses.
2.8. Quantifying Dynamic Brain Properties
We computed multiple dynamic connectivity metrics to assess how variations in cognitive performance and exercise engagement were associated with time-varying patterns of brain network activity as follows:
2.8.1. State Occupancy
State occupancy quantifies the proportion of time each dynamic connectivity state predominates within a state. Let denote the calibrated activation of state i at time window t, with states and windows. To determine which state is most influential at a given moment, we computed each state’s relative amplitude by normalizing its absolute activation against the total activity across all states:
The dominant state at time t is then defined as:
For each state i, the occupancy count is obtained by summing the number of windows in which that state is dominant:
where is the indicator function. The corresponding occupancy percentage is given by:
This formulation yields a distribution describing how frequently each dynamic state emerges as the leading contributor to ongoing connectivity patterns, providing a summary of the temporal prevalence and stability of specific modes of brain network coordination.
2.8.2. Dynamic Convergence and Divergence
To further characterize temporal fluctuations in state dominance, we developed a dynamic convergence/divergence analysis, which examines how similarly or differently states contribute to connectivity at each time window.
For each participant, the calibrated mixing matrix
(with T windows and states) represents the time-varying amplitudes of all dynamic states. At each window t, the state contribution vector
captures the instantaneous prominence of each state.
Pairwise distances between states were computed using the Euclidean metric:
Distances below a predefined threshold were classified as convergent, reflecting similar state amplitudes, while distances exceeding a threshold were classified as divergent, indicating distinct dominance patterns. Thresholds were selected using empirical inspection of distance distributions across participants and time.
This framework distinguishes moments of coordinated state engagement (convergence) from periods of heightened differentiation (divergence), yielding complementary measures of neural flexibility and coherence.
2.9. Statistical Analysis
Associations among dynamic connectivity metrics, cognitive performance, and exercise minutes were examined using general linear models. Predictors included exercise and mood data collected before and after the intervention period, such as daily sedentary minutes (Sedentary Minutes-total), light-intensity activity (Lightly Active Minutes-total), moderate-intensity activity (Fairly Active Minutes-total), vigorous-intensity activity (Very Active Minutes-total), and combined moderate-to-vigorous activity minutes (Fairly-plus-Very-total). Weekly adherence to physical activity guidelines (weeks-adherent-to-PA-guideline), defined as the number of weeks participants achieved minutes of moderate or vigorous activity on at least three days (possible range: 0–8), was also included.
Cognitive predictors consisted of age-corrected NIH Toolbox scores at second visit, including Flanker Inhibitory Control and Attention accuracy (Flanker-t2-AC), List Sorting Working Memory accuracy (LSWM-T2-AC), and Dimensional Change Card Sort accuracy (DCS-T2-AC), capturing executive control, working memory capacity, and cognitive flexibility, respectively.
To control for pre-intervention cognition, stress, and connectivity, we regressed time 1 stress, cognition, and connectivity from time 2 (post intervention) data. Outcome variables consisted of state occupancy measures, convergence/divergence metrics, and calibrated state amplitudes. Models were adjusted for gender and motion where appropriate, and significance thresholds were corrected for multiple comparisons when necessary.
3. Results
3.1. Dynamic State Expression Across Participants
Across participants, the ten dynamic connectivity states exhibited heterogeneous prevalence. Figure 2 shows a subset of highly integrated states– most notably State 3, followed by States 6 and 8– accounted for the greatest proportion of occupancy, reflecting configurations with strong cross-network coordination. In contrast, lower-occupancy states (e.g., States 1, 4, 5, 9, and 10) displayed more modular or segregated connectivity patterns. However, Figure 3 indicated substantial inter-individual variability, consistent with known differences in neural flexibility among young adults.
3.2. Associations Between State Occupancy and Physical Activity
Relationships between state occupancy and physical activity levels were generally modest, yet several systematic patterns emerged as shown in Figure 4. Higher engagement in physical activity–particularly vigorous activity and weeks meeting physical activity guidelines–showed weak positive associations with States 6 and 7. These states are characterized by integrative, cognitively supportive network profiles involving coordinated interactions among cognitive control, default mode, and sensory systems. Individuals with greater physical activity, therefore, tended to spend more time in flexible, high-integration dynamic states.
Conversely, increased occupancy of States 9 and 10, which reflect weaker integration and partially segregated sensory–cognitive patterns, was associated with lower physical activity levels. These findings align with evidence that physical fitness supports enhanced neural flexibility and reduced time spent in low-integration configurations. A small positive association between adherence to activity guidelines and State 1, a low-amplitude baseline-like state, may reflect short-lived recovery periods following exertion.
3.3. Associations Between State Occupancy and Cognitive Outcomes
Cognitive performance also demonstrated selective associations with state occupancy as shown in Figure 5. Working memory accuracy (LSWM) showed positive relationships with States 6 and 7, , aligning with evidence that integrated CC–DM and globally coordinated network states play a role in supporting higher-order cognition at rest. Weak negative trends appeared for States 8 and 9, consistent with their more constrained sensory–motor or cerebellar-weighted connectivity patterns. Flanker accuracy showed generally weak associations, though slight positive trends again emerged for globally integrated states. Overall, the findings indicate that cognitive performance is more closely linked to time spent in flexible, integrated dynamic configurations than to occupancy of segregated or sensory-dominated states.
3.4. Threshold-Free Distance Distribution
Prior to defining convergence and divergence, we examined the empirical distribution of all pairwise Euclidean distances between state amplitudes across subjects and time windows. The resulting histogram shown in Figure 6 exhibited a unimodal, right-skewed profile, with the majority of distances falling within a relatively narrow range around a small value (on the order of ) and a gradual tail extending toward larger distances. This pattern suggests that, at any given moment, most state pairs differ only modestly in amplitude and that very large separations between states are comparatively rare.
On the basis of this observed pattern, the convergence () and divergence () thresholds were chosen with reference to the empirical distribution: was placed near the lower shoulder of the density to capture genuinely tight similarity, and was placed in the upper tail to identify only the most pronounced separations. Thus, convergent and divergent events reflect meaningful deviations from typical state differences rather than arbitrary cutoffs.
3.5. Convergence-Based Dynamic Metrics
We next examined convergence, quantifying how often pairs of states expressed similar amplitudes over time. The mean convergence matrix indicated that convergence was not uniform across the dynamic repertoire: a cluster of mid-range states (States 4–7) showed relatively higher mutual convergence than early (States 1–2) or late segregated states (e.g., State 10). This pattern is consistent with the interpretation of States 4–7 as partially overlapping, integrative configurations that frequently co-fluctuate, whereas more modular states tend to remain dynamically distinct.
Behaviorally, convergence effects were generally modest but revealed structure that was not apparent from occupancy alone. For physical activity as shown in Figure 7, convergence involving integrative states showed opposite patterns for active versus sedentary behavior. Participants with more sedentary showed more convergence among lower-integration states (e.g., pairs involving States 1, 2, and 10) and with convergence between these segregated configurations and the CC–DM state (State 6). In contrast, more active individuals tended to show reduced convergence between segregated and integrative states, suggesting greater differentiation between low-engagement and higher-order network patterns in those with less sedentary time.
Cognitive performance showed the clearest relationships with convergence (Figure 8). For working memory (LSWM), the strongest positive associations appeared for convergence among later, more integrative states (roughly within the State 5–9 cluster), whereas higher LSWM accuracy was associated with reduced convergence for specific pairs involving the CC–DM state (State 6). This pattern suggests that better working memory is linked to coordinated expression of several higher-order network configurations, while maintaining differentiation between certain executive-related states. Flanker accuracy showed weaker but directionally similar trends, with better performance associated with convergence among integrative states and reduced convergence involving State 6 and low-integration configurations.
Taken together, the convergence analysis complements the occupancy findings by indicating that it is not only the amount of time spent in a given state that matters, but also how similarly or distinctly states are expressed relative to one another. Higher physical activity and better cognitive performance are associated with a dynamic system in which integrative states co-fluctuate, while segregated states remain relatively differentiated, consistent with the notion that neural flexibility depends on both state prevalence and the degree of separation or blending between states over time.
3.6. Divergence-Based Dynamic Metrics
Divergence reflects the degree to which pairs of dynamic states differ in their temporal activation patterns, providing a complementary lens through which to view the distinctiveness or separation of functional configurations. Across participants, divergence matrices demonstrated clearer structure than convergence results, with several state pairs showing consistent and behaviorally meaningful differentiation.
A central observation was that State 6, characterized by strong CC–DM integration, functioned as a major hub of divergence across multiple behavioral outcomes. Divergence between State 6 and several other states (notably States 4, 5, 7, 9, and 10) was reliably associated with both physical activity and cognitive performance. These findings parallel the convergence results by indicating that individuals with higher physical activity levels or better cognitive scores maintain State 6 as a more distinct, high-integration attractor, differentiating it from more modular or sensory-weighted configurations.
Physical activity measures showed a consistent pattern in which higher levels of moderate and vigorous activity were associated with increased divergence among mid- to high-integration states, particularly those centered on States 4, 5, and 7. This suggests that more active individuals sustain greater functional separation between higher-order integrative states, in line with theories linking physical fitness to enhanced neural flexibility. Conversely, sedentary behavior was associated with reduced divergence among these states, implying diminished dynamic differentiation.
Cognitive outcomes exhibited similar trends. Working memory accuracy (LSWM) was positively related to divergence among several higher-order states, especially within the State 4–8 range, indicating that successful cognitive performance is supported not only by coordinated activity among integrative states but also by preservation of clear boundaries between these configurations and lower-integration states. Flanker accuracy showed weaker but directionally similar effects, again illustrating the role of divergence around State 6.
Collectively, the divergence findings complement occupancy and convergence metrics by demonstrating that neural flexibility is supported not only by time spent in particular states or their degree of co-fluctuation, but also by the system’s ability to maintain functional specialization across its dynamic repertoire. Greater behavioral health is therefore associated with a dynamic landscape in which high-integration states remain both well utilized and well differentiated from more modular configurations.
4. Discussion
The present study leveraged advanced dynamic functional connectivity analyses to demonstrate that higher levels of physical activity in healthy young adults are associated with more adaptive patterns of intrinsic brain network organization at rest. Specifically, greater physical activity corresponded to increased occupancy in integrative, higher-order connectivity states (particularly states 6 and 7), reduced dwell time in segregated or sensory-weighted states, and enhanced divergence which reflects sharper differentiation between high-engagement integrative configurations and low-engagement or segregated ones. Conversely, higher sedentary behavior was linked to greater convergence among segregated states, suggesting a less differentiated, more rigid dynamic profile.
The preferential occupancy of integrative states in physically active participants aligns with reports that aerobic fitness enhances integration across frontoparietal, default mode, and salience networks—systems critical for cognitive control and goal-directed behavior [24,25]. Similarly, the divergence patterns observed here echo recent evidence that higher cardiorespiratory fitness promotes greater state differentiation, particularly between strongly connected, task-positive states and weakly connected, low-engagement states [4,26]. Our use of constrained dynamic double functional independent primitives (c-ddFIPs) adds novel individualized precision to these observations, confirming that physically active young adults exhibit dynamic profiles that more closely resemble those seen during successful cognitive task performance in prior literature [15,27].
These neural patterns showed parallels with cognitive performance, most robustly working memory, where superior scores aligned with greater state flexibility and differentiation.
The parallel associations with working memory performance are especially compelling. Working memory relies on dynamic reconfiguration between segregated processing and integrative control [28], and our results suggest that physical activity may optimize exactly this flexibility. Individuals with higher working memory capacity spent more time in, and showed sharper differentiation of, integrative states, mirroring the pattern seen in active participants. This convergence implies a shared mechanism: exercise-induced increases in BDNF, cerebral blood flow, and hippocampal–prefrontal coupling may stabilize the neural infrastructure required for rapid, adaptive state switching [29,30]. Notably, these relationships held after controlling for perceived stress (PSS scores), suggesting the effects are not merely secondary to reduced psychological distress, though stress remained a significant negative predictor of working memory improvement in exploratory models.
From a developmental perspective, young adulthood represents a window of both peak plasticity and vulnerability to sedentary lifestyles and chronic stress. The fact that even modest variations in habitual physical activity within this age group corresponded to measurable differences in dynamic network flexibility underscores the potential for low-burden interventions to yield meaningful neurocognitive benefits. College students, who often face competing academic demands and high sedentary time, may derive particular advantage from programs that prioritize actual engagement over mere enrollment.
Several limitations warrant consideration. First, levels of fairly and very vigorous physical activity did not exhibit the same variability as levels of sedentary activity. Second, intervention adherence varied substantially. Participants assigned to the control group exercised more than instructed, limiting between group comparisons and defining an intervention effect. Wearing a FitBit may have inadvertently acted as an intervention such that all participants became more aware of their physical activity. Third, while c-ddFIPs offer excellent individualization, the derivation of 10 states from group-level priors may constrain detection of highly idiosyncratic dynamics. Finally, the sample was predominantly female minority undergraduate students from a single institution, limiting generalizability.
Future studies should employ longer interventions, objective activity monitoring, and multimodal imaging (e.g., combining rs-fMRI with diffusion MRI) to clarify causal pathways. Investigating dose–response relationships and potential moderators (sex, baseline fitness, genetic factors such as BDNF Val66Met) will also be valuable. Ultimately, these findings reinforce physical activity as a powerful, accessible modulator of brain network dynamics during a critical life stage, with direct implications for cognitive resilience and academic success. Finally, higher physical activity in young adults is associated with enhanced temporal flexibility and differentiation of intrinsic connectivity states which as patterns that support executive function and parallel superior working memory performance. These results highlight dynamic functional network reconfiguration as a key neurobiological mechanism linking lifestyle to cognitive health, and strongly advocate for prioritizing physical activity promotion in university wellness programs.
References
- Dautricourt, S.; Gonneaud, J.; Landeau, B.; Calhoun, V.D.; de Flores, R.; Poisnel, G.; Bougacha, S.; Ourry, V.; Touron, E.; Kuhn, E.; et al. Dynamic functional connectivity patterns associated with dementia risk. Alzheimer’s research & therapy 2022, 14, 72. [Google Scholar]
- Beaty, R.E.; Chen, Q.; Christensen, A.P.; Qiu, J.; Silvia, P.J.; Schacter, D.L. Brain networks of the imaginative mind: Dynamic functional connectivity of default and cognitive control networks relates to openness to experience. Human brain mapping 2018, 39, 811–821. [Google Scholar] [CrossRef]
- Fu, Z.; Sui, J.; Iraji, A.; Liu, J.; Calhoun, V.D. Cognitive and psychiatric relevance of dynamic functional connectivity states in a large (N> 10,000) children population. Molecular Psychiatry 2025, 30, 402–413. [Google Scholar] [CrossRef]
- Alfini, A.J.; Won, J.; Weiss, L.R.; Nyhuis, C.C.; Shackman, A.J.; Spira, A.P.; Smith, J.C. Impact of exercise on older adults’ mood is moderated by sleep and mediated by altered brain connectivity. Social Cognitive and Affective Neuroscience 2020, 15, 1238–1251. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.W.; Pieruccini-Faria, F.; Bartha, R.; Doherty, T.J.; Nagamatsu, L.S.; Montero-Odasso, M. The effect of physical exercise on functional brain network connectivity in older adults with and without cognitive impairment. A systematic review. Mechanisms of Ageing and Development 2021, 196, 111493. [Google Scholar] [CrossRef]
- Chirles, T.J.; Reiter, K.; Weiss, L.R.; Alfini, A.J.; Nielson, K.A.; Smith, J.C. Exercise training and functional connectivity changes in mild cognitive impairment and healthy elders. Journal of Alzheimer’s Disease 2017, 57, 845–856. [Google Scholar] [CrossRef] [PubMed]
- van Balkom, T.D.; van den Heuvel, O.A.; Berendse, H.W.; van der Werf, Y.D.; Vriend, C. The effects of cognitive training on brain network activity and connectivity in aging and neurodegenerative diseases: a systematic review. Neuropsychology Review 2020, 30, 267–286. [Google Scholar] [CrossRef]
- Arain, M.; Haque, M.; Johal, L.; Mathur, P.; Nel, W.; Rais, A.; Sandhu, R.; Sharma, S. Maturation of the adolescent brain. Neuropsychiatric disease and treatment 2013, 449–461. [Google Scholar]
- Sadeghi, N.; van der Velpen, I.F.; Baker, B.T.; Batta, I.; Cahill, K.J.; Genon, S.; McCormick, E.; Michel, L.C.; Moraczewski, D.; Seraji, M.; et al. The interplay between brain and behavior during development: A multisite effort to generate and share simulated datasets. Scientific Data 2025, 12, 473. [Google Scholar] [CrossRef]
- Soleimani, N.; Wiafe, S.L.; Iraji, A.; Pearlson, G.; Calhoun, V.D. A novel framework to quantify dynamic convergence and divergence of overlapping brain states characterizing four psychiatric disorders. Network Neuroscience 2025, 1–25. [Google Scholar] [CrossRef]
- Soleimani, N.; Iraji, A.; Pearlson, G.; Preda, A.; Calhoun, V.D. Unraveling the Neural Landscape of Mental Disorders using Double Functional Independent Primitives (dFIPs). Biological Psychiatry: Cognitive Neuroscience and Neuroimaging 2025. In press. [Google Scholar] [CrossRef]
- Moore, D.; Jung, M.; Hillman, C.H.; Kang, M.; Loprinzi, P.D. Interrelationships between exercise, functional connectivity, and cognition among healthy adults: A systematic review. Psychophysiology 2022, 59, e14014. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Bennett, H.; Miatke, A.; Dumuid, D.; Curtis, R.; Ferguson, T.; Brinsley, J.; Szeto, K.; Petersen, J.M.; Gough, C.; et al. Effectiveness of exercise for improving cognition, memory and executive function: a systematic umbrella review and meta-meta-analysis. British journal of sports medicine 2025, 59, 866–876. [Google Scholar] [CrossRef]
- Gogniat, M.A.; Robinson, T.L.; Jean, K.R.; Miller, L.S. Physical activity moderates the association between executive function and functional connectivity in older adults. Aging Brain 2022, 2, 100036. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Min, L.; Liu, R.; Zhang, X.; Wu, M.; Di, Q.; Ma, X. The effect of exercise on cerebral blood flow and executive function among young adults: a double-blinded randomized controlled trial. Scientific Reports 2023, 13, 8269. [Google Scholar] [CrossRef]
- Rawliuk, T.; Thrones, M.; Cordingley, D.M.; Cornish, S.M.; Greening, S.G. Promoting brain health and resilience: The effect of three types of acute exercise on affect, brain-derived neurotrophic factor and heart rate variability. Behavioural Brain Research 2025, 115675. [Google Scholar] [CrossRef] [PubMed]
- Sleiman, S.F.; Henry, J.; Al-Haddad, R.; El Hayek, L.; Abou Haidar, E.; Stringer, T.; Ulja, D.; Karuppagounder, S.S.; Holson, E.B.; Ratan, R.R.; et al. Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body β-hydroxybutyrate. elife 2016, 5, e15092. [Google Scholar] [CrossRef]
- Tessadori, J.; Galazzo, I.B.; Storti, S.F.; Pini, L.; Brusini, L.; Cruciani, F.; Sona, D.; Menegaz, G.; Murino, V. Linking dynamic connectivity states to cognitive decline and anatomical changes in Alzheimer’s disease. NeuroImage 2025, 121448. [Google Scholar] [CrossRef]
- Douw, L.; Wakeman, D.G.; Tanaka, N.; Liu, H.; Stufflebeam, S.M. State-dependent variability of dynamic functional connectivity between frontoparietal and default networks relates to cognitive flexibility. Neuroscience 2016, 339, 12–21. [Google Scholar] [CrossRef]
- Sinha, N.; Berg, C.N.; Yassa, M.A.; Gluck, M.A. Increased dynamic flexibility in the medial temporal lobe network following an exercise intervention mediates generalization of prior learning. Neurobiology of Learning and Memory 2021, 177, 107340. [Google Scholar] [CrossRef]
- Cai, W.; Taghia, J.; Menon, V. A multi-demand operating system underlying diverse cognitive tasks. Nature Communications 2024, 15, 2185. [Google Scholar] [CrossRef]
- Ingwersen, T.; Mayer, C.; Petersen, M.; Frey, B.M.; Fiehler, J.; Hanning, U.; Kühn, S.; Gallinat, J.; Twerenbold, R.; Gerloff, C.; et al. Functional MRI brain state occupancy in the presence of cerebral small vessel disease—a pre-registered replication analysis of the Hamburg City Health Study. Imaging Neuroscience 2024, 2, 1–17. [Google Scholar] [CrossRef]
- Du, Y.; Fu, Z.; Sui, J.; Gao, S.; Xing, Y.; Lin, D.; Salman, M.; Abrol, A.; Rahaman, M.A.; Chen, J.; et al. NeuroMark: An automated and adaptive ICA based pipeline to identify reproducible fMRI markers of brain disorders. NeuroImage: Clinical 2020, 28, 102375. [Google Scholar] [CrossRef] [PubMed]
- Voss, M.W.; Prakash, R.S.; Erickson, K.I.; Basak, C.; Chaddock, L.; Kim, J.S.; Alves, H.; Heo, S.; Szabo, A.; White, S.M.; et al. Plasticity of brain networks in a randomized intervention trial of exercise training in older adults. Frontiers in aging neuroscience 2010, 2, 1803. [Google Scholar] [CrossRef]
- Stillman, C.M.; Donofry, S.D.; Erickson, K.I. Exercise, fitness and the aging brain: a review of functional connectivity in aging. Archives of Psychology 2019, 3. [Google Scholar] [CrossRef]
- Ge, L.K.; Hu, Z.; Wang, W.; Siu, P.M.; Wei, G.X. Aerobic exercise decreases negative affect by modulating orbitofrontal-amygdala connectivity in adolescents. Life 2021, 11, 577. [Google Scholar] [CrossRef]
- Pindus, D.M.; Zwilling, C.E.; Jarrett, J.S.; Talukdar, T.; Schwarb, H.; Anderson, E.; Cohen, N.J.; Barbey, A.K.; Kramer, A.F.; Hillman, C.H. Opposing associations between sedentary time and decision-making competence in young adults revealed by functional connectivity in the dorsal attention network. Scientific reports 2020, 10, 13993. [Google Scholar] [CrossRef]
- Cohen, J.R.; D’Esposito, M. The segregation and integration of distinct brain networks and their relationship to cognition. Journal of Neuroscience 2016, 36, 12083–12094. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proceedings of the national academy of sciences 2011, 108, 3017–3022. [Google Scholar] [CrossRef]
- Erickson, K.I.; Hillman, C.; Stillman, C.M.; Ballard, R.M.; Bloodgood, B.; Conroy, D.E.; Macko, R.; Marquez, D.X.; Petruzzello, S.J.; Powell, K.E. Physical activity, cognition, and brain outcomes: a review of the 2018 physical activity guidelines. Medicine and science in sports and exercise 2019, 51, 1242. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
the ddFIP (1–10) demonstrating the variability and shared structure of dynamic functional interaction patterns across canonical brain networks. Warmer colors indicate stronger positive coupling, and cooler colors indicate weaker or negative coupling. Rows and columns correspond to predefined brain networks (SC, AUD, SM, VS, CC, DM, CB).
Figure 1.
the ddFIP (1–10) demonstrating the variability and shared structure of dynamic functional interaction patterns across canonical brain networks. Warmer colors indicate stronger positive coupling, and cooler colors indicate weaker or negative coupling. Rows and columns correspond to predefined brain networks (SC, AUD, SM, VS, CC, DM, CB).
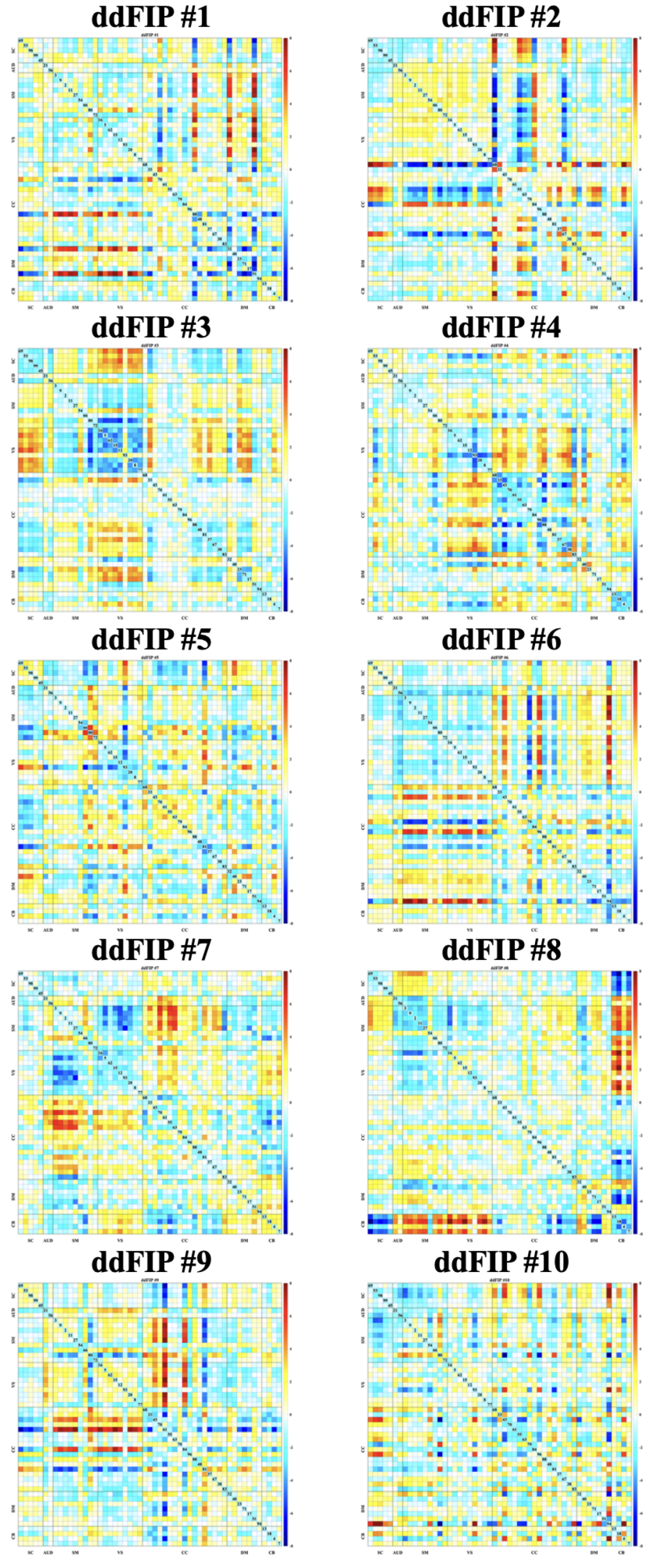
Figure 2.
Average occupancy of the ten dynamic double functional independent primitive states, showing higher prevalence for integrated states (especially State 3, followed by States 6 and 8) and lower occupancy for more modular states.
Figure 2.
Average occupancy of the ten dynamic double functional independent primitive states, showing higher prevalence for integrated states (especially State 3, followed by States 6 and 8) and lower occupancy for more modular states.
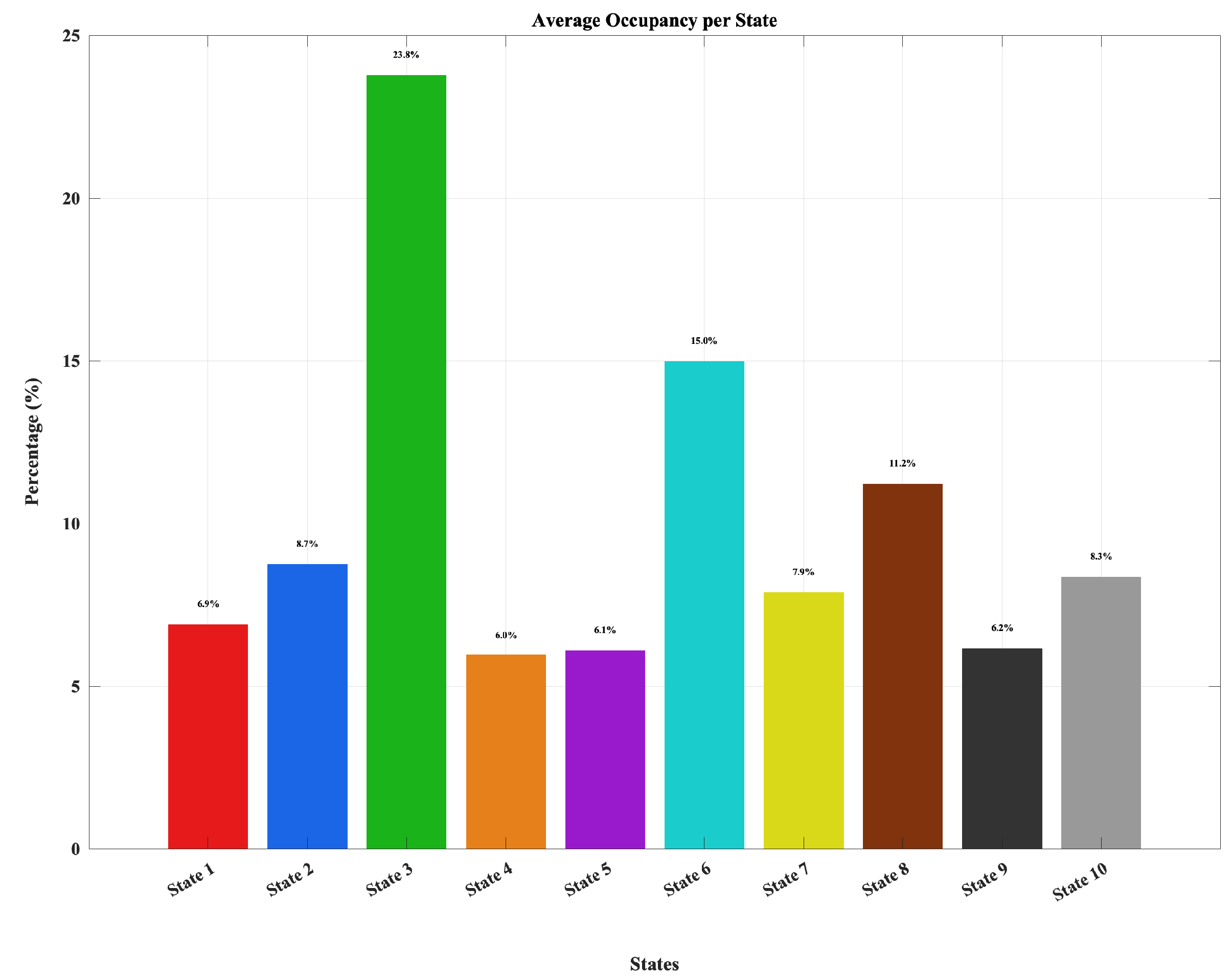
Figure 3.
Average percentage occupancy across the ten dynamic connectivity states, showing highest prevalence for State 3 and moderate occupancy for States 6 and 8, with the remaining states occurring less frequently. Boxplots illustrate inter-individual variability in state expression.
Figure 3.
Average percentage occupancy across the ten dynamic connectivity states, showing highest prevalence for State 3 and moderate occupancy for States 6 and 8, with the remaining states occurring less frequently. Boxplots illustrate inter-individual variability in state expression.
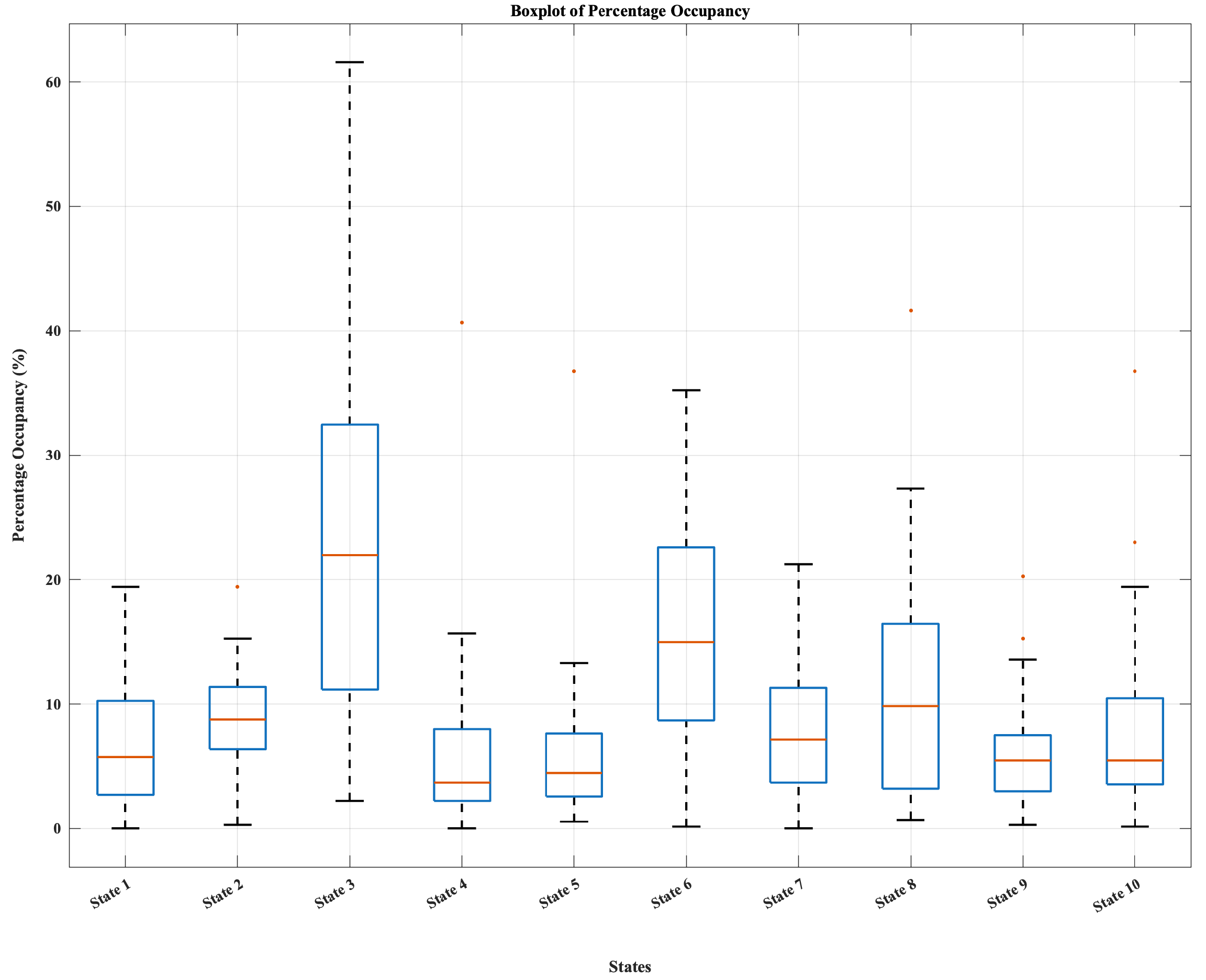
Figure 4.
Associations between dynamic state occupancy and physical activity or behavioral measures. Each panel depicts the relationship between occupancy of the ten dynamic connectivity states and a behavioral outcome, with individual data points shown across participants and linear fits overlaid. (a)Very active minutes; (b)Combined fairly active and very active minutes; (c)Fairly active minutes; (d)Lightly active minutes; (e) Sedentary minutes; (f) Number of weeks adherent to recommended physical activity guidelines. Across metrics, effect sizes were small, though several trends indicate that higher physical activity and better adherence were associated with greater occupancy of integrative states and reduced occupancy of segregated or sensory-weighted states.
Figure 4.
Associations between dynamic state occupancy and physical activity or behavioral measures. Each panel depicts the relationship between occupancy of the ten dynamic connectivity states and a behavioral outcome, with individual data points shown across participants and linear fits overlaid. (a)Very active minutes; (b)Combined fairly active and very active minutes; (c)Fairly active minutes; (d)Lightly active minutes; (e) Sedentary minutes; (f) Number of weeks adherent to recommended physical activity guidelines. Across metrics, effect sizes were small, though several trends indicate that higher physical activity and better adherence were associated with greater occupancy of integrative states and reduced occupancy of segregated or sensory-weighted states.

Figure 5.
Associations between dynamic state occupancy and cognitive performance outcomes. Each panel shows linear associations between occupancy of the ten dynamic connectivity states and a cognitive metric, with individual participants represented as points and regression lines overlaid. (a)Age corrected dimensional change card sort accuracy score at Time 2; (b) List Sorting Working Memory (LSWM) accuracy at Time 2 (c) Flanker inhibitory control and attention accuracy at Time 2; Across measures, effect sizes were small, with modest positive trends appearing for several integrative states (e.g., States 6 and 7), suggesting that greater time spent in flexible, higher-order network configurations may modestly support executive and working memory performance.
Figure 5.
Associations between dynamic state occupancy and cognitive performance outcomes. Each panel shows linear associations between occupancy of the ten dynamic connectivity states and a cognitive metric, with individual participants represented as points and regression lines overlaid. (a)Age corrected dimensional change card sort accuracy score at Time 2; (b) List Sorting Working Memory (LSWM) accuracy at Time 2 (c) Flanker inhibitory control and attention accuracy at Time 2; Across measures, effect sizes were small, with modest positive trends appearing for several integrative states (e.g., States 6 and 7), suggesting that greater time spent in flexible, higher-order network configurations may modestly support executive and working memory performance.
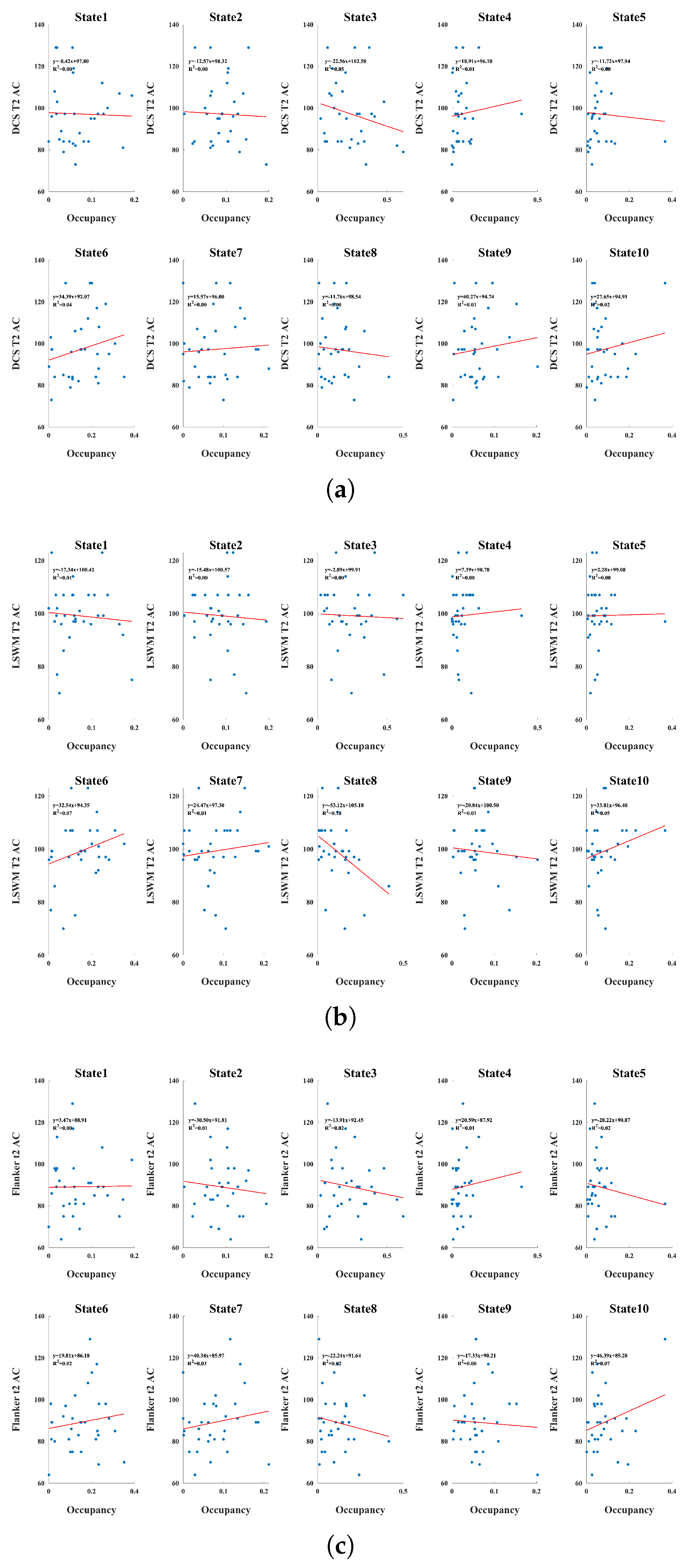
Figure 6.
Distribution of pairwise Euclidean distances between state amplitudes across all subjects and time windows. The unimodal, right-skewed distribution indicates that most state pairs differ only modestly in amplitude at any given moment, with larger separations occurring less frequently. This empirical distribution was used to guide the selection of convergence and divergence thresholds, with lower-bound thresholds capturing genuinely close similarity and upper-bound thresholds capturing only the most pronounced state separations.
Figure 6.
Distribution of pairwise Euclidean distances between state amplitudes across all subjects and time windows. The unimodal, right-skewed distribution indicates that most state pairs differ only modestly in amplitude at any given moment, with larger separations occurring less frequently. This empirical distribution was used to guide the selection of convergence and divergence thresholds, with lower-bound thresholds capturing genuinely close similarity and upper-bound thresholds capturing only the most pronounced state separations.
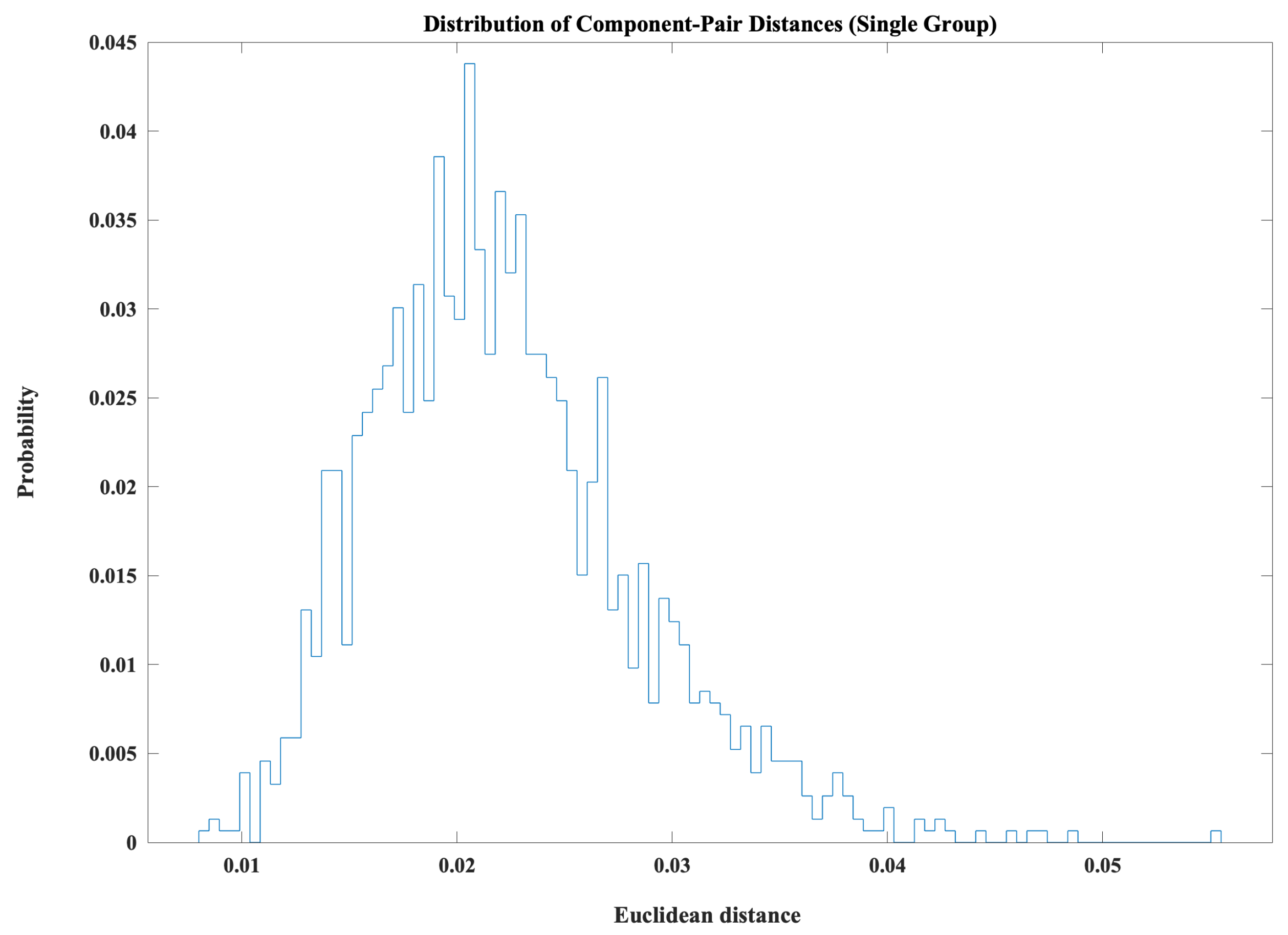
Figure 7.
Convergence-based associations between physical activity measures and dynamic functional connectivity states, quantified using . Each panel displays the statistical strength and direction of associations between physical activity and pairwise state convergence, reflecting how often pairs of dynamic states exhibit similar amplitudes over time. Across measures, a consistent pattern emerged in which mid-range integrative states (States 4–7) showed stronger mutual convergence than early (States 1–2) or late segregated states (e.g., State 10). Higher sedentary behavior was associated with increased convergence among lower-integration states and with convergence between these segregated configurations and the CC–DM integrative state (State 6), whereas greater physical activity tended to correspond to reduced convergence between segregated and integrative states, indicating enhanced differentiation of higher-order network dynamics in more active individuals. (a) Very active minutes; (b) Combined fairly active and very active minutes; (c) Fairly active minutes; (d) Lightly active minutes; (e) Sedentary minutes; (f) Number of weeks adherent to recommended physical activity guidelines.
Figure 7.
Convergence-based associations between physical activity measures and dynamic functional connectivity states, quantified using . Each panel displays the statistical strength and direction of associations between physical activity and pairwise state convergence, reflecting how often pairs of dynamic states exhibit similar amplitudes over time. Across measures, a consistent pattern emerged in which mid-range integrative states (States 4–7) showed stronger mutual convergence than early (States 1–2) or late segregated states (e.g., State 10). Higher sedentary behavior was associated with increased convergence among lower-integration states and with convergence between these segregated configurations and the CC–DM integrative state (State 6), whereas greater physical activity tended to correspond to reduced convergence between segregated and integrative states, indicating enhanced differentiation of higher-order network dynamics in more active individuals. (a) Very active minutes; (b) Combined fairly active and very active minutes; (c) Fairly active minutes; (d) Lightly active minutes; (e) Sedentary minutes; (f) Number of weeks adherent to recommended physical activity guidelines.
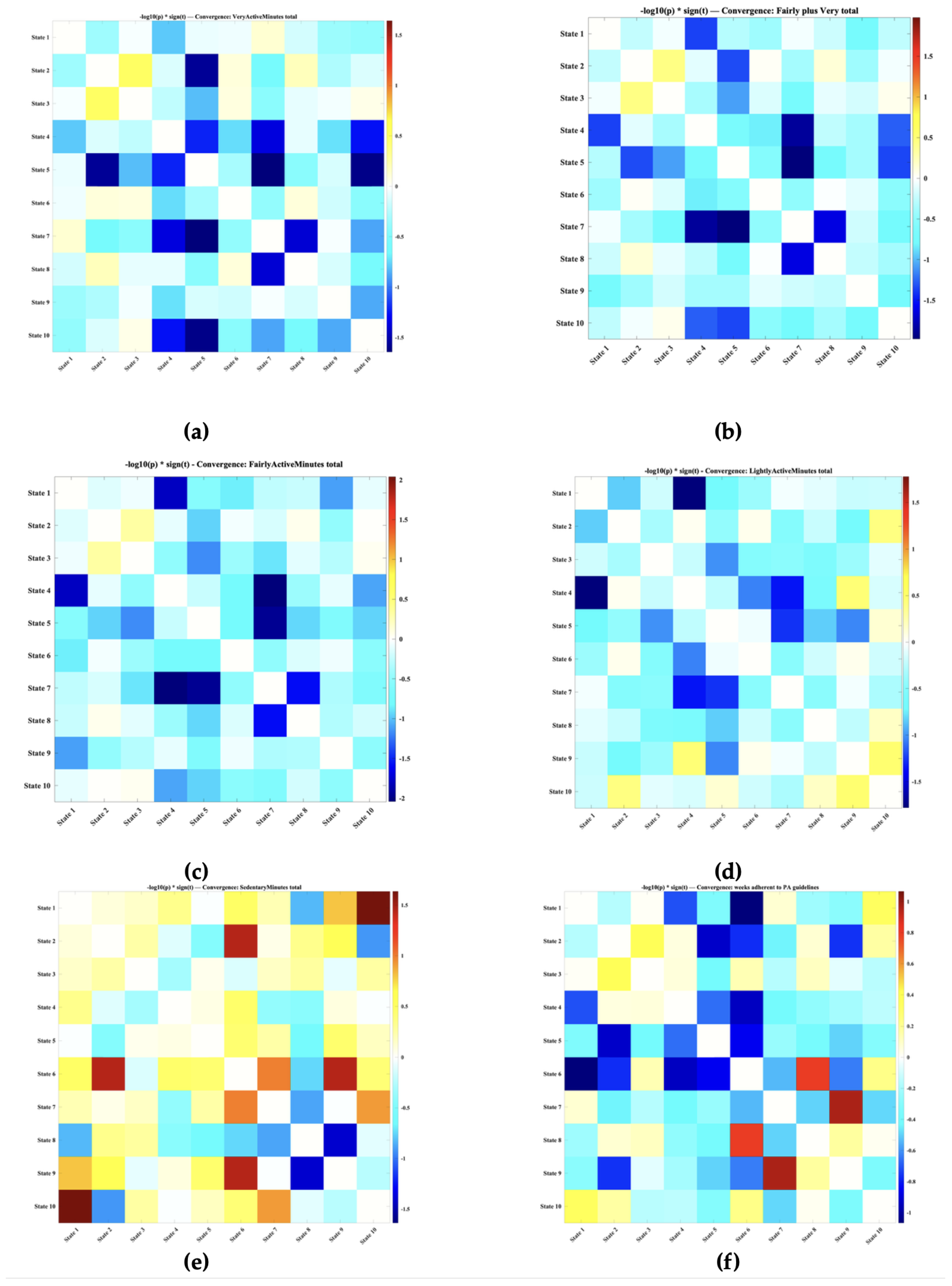
Figure 8.
Convergence-based associations between cognitive performance and dynamic functional connectivity states, quantified using . Each panel presents the statistical associations between cognitive outcomes and pairwise state convergence, reflecting the extent to which pairs of dynamic states exhibit similar amplitudes over time. Higher cognitive performance was generally associated with greater convergence among integrative, mid-range states, whereas lower performance tended to correspond to stronger convergence between segregated or sensory-weighted states and executive–default configurations. (a) Age corrected dimensional change card sort accuracy score at Time 2; (b) List Sorting Working Memory (LSWM) accuracy at Time 2 (c) Flanker inhibitory control and attention accuracy at Time 2;
Figure 8.
Convergence-based associations between cognitive performance and dynamic functional connectivity states, quantified using . Each panel presents the statistical associations between cognitive outcomes and pairwise state convergence, reflecting the extent to which pairs of dynamic states exhibit similar amplitudes over time. Higher cognitive performance was generally associated with greater convergence among integrative, mid-range states, whereas lower performance tended to correspond to stronger convergence between segregated or sensory-weighted states and executive–default configurations. (a) Age corrected dimensional change card sort accuracy score at Time 2; (b) List Sorting Working Memory (LSWM) accuracy at Time 2 (c) Flanker inhibitory control and attention accuracy at Time 2;
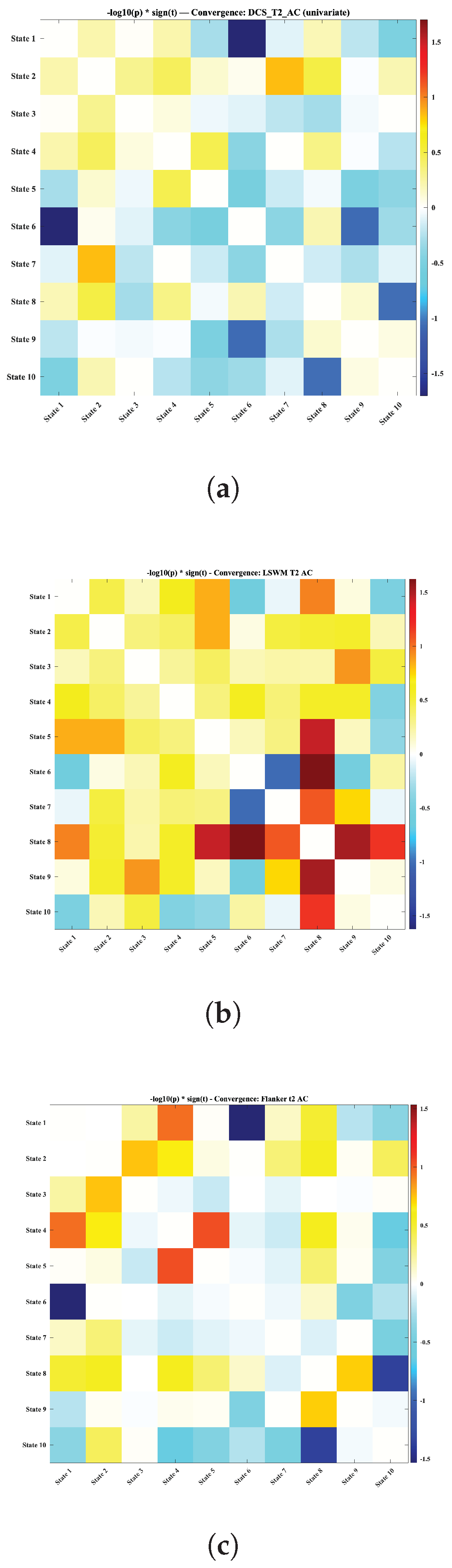

Table 1.
Demographic Characteristics of the Full Sample and MRI Subsample.
| Characteristic | Full Sample (N = 103) | MRI Subsample (N = 32) |
|---|---|---|
| Age (years) | ||
| Mean (SD) | 20.81 (3.13) | 21.22 (3.21) |
| Range | 17–33 | 18–33 |
| Gender | ||
| Female | 77 (74.8%) | 24 (75.0%) |
| Male | 23 (22.3%) | 8 (25.0%) |
| Nonbinary/Other | 3 (2.9%) | 0 (0%) |
| Race/Ethnicity | ||
| Asian | 27 (26.2%) | 10 (31.3%) |
| Black/African American | 42 (40.8%) | 10 (31.3%) |
| White/Caucasian | 14 (13.6%) | 5 (15.6%) |
| Hispanic/Latino | 9 (8.7%) | 6 (18.8%) |
| Biracial (Black–White) | 2 (1.9%) | 1 (3.1%) |
| Other/Multiracial | 9 (8.7%) | 0 (0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



