Submitted:
07 February 2026
Posted:
09 February 2026
You are already at the latest version
Abstract
Hydra vulgaris, an ancient cnidarian, exhibits extraordinary regenerative and neurogenic abilities mediated by morphogenetic peptides, particularly the head activator. This neuropeptide regulates cell proliferation, differentiation, and nerve net maintenance in Hydra, and surprisingly, exserts similar mitogenic and neurogenic effects in mammalian systems. Despite early enthusiasm, research on the head activator waned due to controversies about its gene of origin and receptor identity. Experimental evidence implicates the sortilin-related receptor (SorLA) as a primary receptor in mammals, with potential modulatory roles for the G-protein-coupled receptor GPR37. This review consolidates the head activator’s evolutionary, molecular, and functional landscape, highlighting its pleiotropic functions across species. Given its neuroprotective, regenerative, and immunomodulatory properties, the head activator warrants reconsideration as a therapeutic candidate for neurodegenerative disorders and regenerative medicine.
Keywords:
demyelination
; neurorepair
; small‐molecule therapeutics
; neurodegeneration
1. Introduction
Some of the earliest organisms to develop body symmetry and a simple neuronal net belong to the ancient clade Cnidaria that has branched away from the bilaterians 850-635 million years ago [1]. One representative, Hydra vulgaris or simply Hydra, has been studied as a classical model organism for morphogenesis, neurogenesis, and regeneration [2,3]. Hydra has a nearly unlimited regenerative and stem cell renewal capacity [4]. At the same time, Hydra possesses one of the first primitive forms of a neuronal net to evolve on Earth, yet strikingly intricate in its complexity [3].
Hydra is a small predatory fresh-water polyp that reproduces mostly asexually. It is primitively organised with a tubular body subdivided into the basal disc (foot), body column (gastric region), and head (Figure 1). The body can reach up to 30 mm in length. A hypostome with an oral opening, surrounded by a radially symmetrical crown of tentacles form the head region. It is settled on the top of the body column with the gastric cavity, where the microbiome resides and prey digestion occurs. The body is composed of two layers: the outer epidermis, and the inner gastrodermis, separated by mesoglea, a layer of extracellular matrix. Three main cell types shape the histological architecture of Hydra: epithelial cells, interstitial cells, and neurons (Figure 1).
The epithelial cells outline the epi- and gastrodermal layers. These cells have epitheliomuscular features, hence they are alternatively denominated as myoepithelial cells. They follow a circular arrangement in the gastroderm, and a circular one in the ectoderm, providing body movements [6]. These cells can proliferate upon injury and differentiate into more specialised entities called “cnidocytes” harbouring stinging organelles (nematocysts) [7]. The cnidocytes densely populate the tentacle areas of Hydra, being enriched in neurotoxic substances, ready for a rapid injection to paralyse different kinds of prey. Captured by the tentacles, the immobilised prey is propelled towards the hypostome. Alternatively, the epithelial cells outlining the endoderm can transfigure into secretory gland cells to release mucus and digestive enzymes [6].
The interstitial cells of Hydra represent an equivalent of stem cells with a high mitosis turnover to maintain a pool of pluripotent cells and replenish epithelia or neurons through terminal differentiation (Figure 2). This allows a full recovery from even the most severe injuries such as decapitation or a complete disaggregation into a suspension of cells. Driven by its interstitial cells into a continuous state of cell renewal and adolescence, Hydra escapes senescence to maintain biological longevity [8]. Strikingly, some molecular mechanisms underlying longevity in Hydra are shared by vertebrates and specifically mammals. In particular, the telomeric sequences are similar in cnidarians and mammals [9,10], also orthologous aging-related genes such as FoxO, myc, and PIWI were found in both cnidarians and mammals [8].
In simple terms, the neurons of Hydra build two major functional groups: sensory neurons and ganglion cells. Deeper molecular analyses have revealed a highly complex architecture encompassing at least 11 neuronal subtypes with specific biomarkers, localisation and unique developmental trajectories [11]. For instance, Hym-176 positive neurons single-handedly form the motor network [12]. These cells build a diffuse nerve net scattered across the entire body of Hydra. In fact, there are two separate nerve nets without apparent synaptic connections between each other – one in the epidermal layer and another in the gastrodermal layer [13,14]. In addition, there are hypostomal and peduncle nerve ring regions, which are densely populated by neurons [15]. It should be emphasised that due to the common origin of bilaterian and cnidarian nervous systems a high number of key neurogenesis-regulator genes have shared orthologs, e.g. CREB, POU, LIM, Sox/TCF, Notch, Wnt, Hedgehog, and JAK/STAT [16].
The epithelial and neuronal cells secrete a wide range of signaling molecules, most of which are peptides [17]. These peptides form a rich neuroendocrine system, covering every aspect of Hydra’s life – they control development and regeneration, neurotransmission, behavioural responses, and by far pathogen defense. Previous research efforts have been mainly made to study antibacterial peptides, cytotoxic compounds for cancer treatment, or the use of cnidarian neurotoxins to modulate ion channels in context of neurological disorders such as neuropathic pain and epilepsy [18,19]. Peculiarly, one morphogenic peptide that is deeply anchored in cnidarian neurogenesis, Hydra’s head activator, is the only cnidarian molecule that has remained conserved through eons of evolution up to Homo sapiens [20]. The head activator has been found to alter neural differentiation and regeneration in mammalian tissues and cell lines, indicating its potential application as a therapeutic compound in human disease conditions [20]. This wealth of scientific evidence awakened our interest in the potential of cnidarian morphogenic peptides for application in preclinical models and human patients to stimulate regeneration and neuroprotection.
Between the years 1970 and 1990, pioneering research on the head activator peptide yielded essential insights into neurogenesis and neuropathology in both cnidarians and mammals. However, after the pinnacle of this research, some contradicting findings and viewpoints in the literature caused the head activator to soon fade into obscurity and disappear from the scope of the scientific community. Major concerns have been raised towards the enigmatic gene of origin of the head activator. In addition, the signaling cascade governing proliferation via GPR37 receptor activation has been highly questioned. Despite these controversies, multiple studies in preclinical mammalian models have assessed the head activator’s mitogenic effect and deemed the molecule to be credible as a regenerative therapeutic.
We assert that the primary factors contributing to research stagnation in this field are not only the identified inconsistencies in literature but also the selection of unsuitable testing models. Although controversially disputed, we conclude that the head activator may represent a significant entity for both cnidarian and foundational research, as well as for therapeutic applications in human diseases. Here, we present a review of head activator’s functions and implications that are relevant to regeneration and neuroprotection.
2. Peptidergic Signaling in Hydra
Hicklin and colleagues have identified head and foot organisers in Hydra as central hubs of development and maintenance of the body and neuronal net, mediated through intrinsically secreted morphogenic factors [21]. Pioneering works of Chica Schaller, Toshio Takahashi, Toshitaka Fujisawa and their colleagues have discovered the underlying signaling peptides of morphogenesis in Hydra [22,23,24]. The predominantly peptidergic signaling in Hydra consists of a wide palette of peptides with different and sometimes antagonistic functions [24]. These peptides are classified into epithelio- [25] and neuropeptides [26] according to their cellular origin, and further subdivided in respect to their structural and functional hallmarks.
In Schaller’s laboratory, chromatographic fractions of Hydra homogenates have displayed morphogenic activity, and further separation techniques led to the isolation of the head activator neuropeptide [27] and foot formation factors, such as the epitheliopeptides pedin and pedibin [28]. These peptides have been shown to control axial body patterning by shaping the head and foot regions in Hydra as well as regulate stem cell proliferation and neuronal differentiation [28,29].
Subsequent large-scale analysis within the scope of The Hydra Peptide Project identified nearly 800 peptides, out of which only 55 were functionally characterised [24]. Different peptide families have been discovered: morphogenic, neurogenic, myoactive, and behavioural regulators [24]. We have summarised some morphogenic and neurogenic peptides in Table 1.
The main morphogenic peptides in Hydra can be classified into head- and foot-specific factors. Some peptides have a dual function and participate both in morpho- and neurogenesis. The head-specific peptides are the head activator and HEADY. The head activator neuropeptide, being the most prominent head stimulator, induces development of the head region and promotes mitotic expansion of the interstitial cells, followed by an enhancement of their differentiation into epithelial and neuronal cells [29].
Another morphogenic entity, the foot-formation factor was identified to contain the pedin and pedibin peptides [28]. Both peptides originate from epithelial cells and have a similar function: they induce development and regeneration of the foot region [28]. Additionally, pedibin participates in head and tentacle organisation [33], while pedin can support axial body patterning and asexual reproduction by budding [34].
Maintenance and formation of neural networks in Hydra is controlled by stimulatory peptides such as pedin, head activator, and Hym-355 [26,35], and also by inhibitory peptides such as Hym-33H, Hym-35, Hym-37, and Hym-310 [26,35]. The stimulatory peptides induce proliferation of the interstitial stem-like cells and their differentiation into a neuronal phenotype, while the inhibitory peptides strive to keep a conserved state by showing an inverse effect [24]. The interaction between these two systems is an equilibrium, which can however adaptively be shifted, e.g. upon injury. Particularly, the inhibitory Hym-33H and the stimulatory Hym-355 are antagonistic peptides that form a regulatory feedback loop depending on the density of the secreting cells and corresponding relative peptide concentrations, thus allowing tight surveillance of the neuronal differentiation [31,36].
The most prominent stimulatory peptide is Hym-355, while other neuronal stimulators such as head activator and pedin likely modify the neuronal differentiation response. Hym-355 is a neuropeptide expressed in head and foot region and endodermal nerve net [31], notably it contains a C-terminal proline-arginine-glycine-amide (PRGamide) motif that corresponds to mammalian Arg-vasopressin [37]. The epitheliopeptide pedin is present in both endo- and ectoderm, having a dual mitogen and morphogen function, it participates in axial body patterning and stimulates proliferation and neuronal differentiation [28]. Of note, pedin is related to an actin-binding protein thymosin-β₄ [34]. The head activator peptide exerts a dualistic functional nature as a mitogen and neuromorphogen, and is predominantly expressed in the ectodermal neurons [38].
The peptides inhibiting neuronal differentiation belong to the proline-tryptophan-family (PW-family) and contain an eponymous C-terminal proline-tryptophan residue [24]. An example of these peptides is Hym-33H, which is mainly expressed in the ectodermal epithelium of the body column and understood to inhibit the neuronal differentiation across the entire nerve net by antagonising Hym-355 [36]. However, prolonged exposure to Hym-33H neutralises this inhibitory effect and suggests therefore an autoregulatory loop [36].
Such a complex interplay between the peptides allows for a targeted control of morphogenesis, neurogenesis, and regeneration. We hypothesise that cnidarian peptides such as the head activator can exert in mammalian tissues similar neuromorphogenic effects as seen in Hydra.
3. Context-Dependent Signaling of the Head Activator
In Hydra, the head activator regulates formation of the head region, cell division, and neuronal differentiation [29,39,40,41,42]. The molecule has been postulated to act as a growth factor for early development of the nervous and neuroendocrine systems in mammals [20]. The head activator seems to have two temporally dynamic and context-dependent modes of action controlled by different pathways in Hydra (Table 2 and Figure 3). As an early ontogenetic cue, the molecule stimulates proliferation of mitotically active cells [43,44]. As a later ontogenetically occurring signal in post-mitotic cells, it allows to set the course of terminal differentiation into neurons or head-specific epithelial cells to form the nerve net and head organiser [43,44]. It turns out that the border between differentiation and proliferation signals lies within the concentration of the head activator: under low dosage (10⁻¹³ M), the interstitial and epithelial cells begin to proliferate, whereas when the concentration rises up to 10⁻¹¹ – 10⁻⁹ M, the cells initiate head-specific epithelial and neuronal differentiation [41,45]. Several kinetic studies have determined that Hydra possesses at least two types of head activator receptors, one with a high affinity responsible for the mitogenic effect, and another with a lower affinity which triggers cell differentiation [46,47,48], thus explaining the dose-effect relationship through different receptor affinities. Interestingly, Schaller and colleagues could show more pronounced binding of the head activator to the surface of immature cells rather than of terminally differentiated cells, thus favouring cell cycle phase and differentiation status-dependent receptor expression patterns [49]. Moreover, an autocrine growth factor function of the head activator has been proposed to control the G2/S transition during mitosis [49]. The early pro-mitotic activation inhibits the adenylyl cyclase and depletes cAMP, while later occurring neuronal differentiation requires cAMP upregulation [44,45,50]. Of note, similar temporal fluctuation patterns of cAMP concentration after head activator treatment could be observed in rodent livers [51]. Radically opposing effects of the head activator on cAMP concentration indicate that both a stimulatory and an inhibitory G-protein coupled receptor (GPCR) are likely involved. However, if both receptors were simultaneously active, they would possibly cancel each other out. We assume therefore that their activity or expression must be controlled in a temporally dynamic manner, allowing one single receptor class to dominate during the corresponding phase of cell cycle or differentiation, as Schaller and colleagues have previously shown [49].
4. Signaling Pathways of the Head Activator Peptide in Hydra
Upon secretion from neurons, the head activator can become inhibited through dimerisation and proteolytic degradation [52], or bind to a carrier molecule to escape proteolysis and preserve its activity [53]. However, an NMR-spectroscopic analysis has questioned the dimerisation as an inactivation mechanism [54]. Furthermore, the dimer has also been reported to be a potent mitogen [55]. The carrier molecule of the head activator peptide, the head activator binding protein (HAB), has been identified as a high-affinity receptor that exists in both membrane-anchored and soluble forms [55,56]. Interestingly, HAB is related to the mammalian sortilin-related receptor, SorLA, also known as SORL1 [56,57], an intracellular trafficking protein located in the Golgi apparatus [58]. Furthermore, the mammalian homolog SorLA has been shown to interact with the head activator and facilitate its signaling [59]. HAB/SorLA seems to be the central entity and very first step in the head activator signaling cascade. We believe that this protein could context-dependently engage in a complex with either a Gs or a Gi protein, considering the downstream regulation of the cAMP as a second messenger [50]. Indeed, the existence of such complex seems convenient, since SorLA alone would not be sufficient to trigger cAMP concentration changes due to SorLA lacking association to any G proteins. Rezgaoui and colleagues have proposed a receptor complex consisting of SorLA and inhibitory GPR37 to facilitate mitotic signaling in mammalian cells [60]. This could indicate that a similar receptor complex may be present in Hydra. However, there are no known GPR37 orthologs in Hydra, thus leaving the identity of the cnidarian head activator mitotic co-receptor obscure. Another supporting thesis about a receptor complex has been delivered by Jacobsen and colleagues [59]. They have shown that monomeric SorLA binds the head activator with a lower affinity in a purified SorLA/head activator system [59]. Earlier, Hampe and colleagues have observed a high affinity binding between the dimerised head activator and SorLA in a pull-down experiment with NT2 cell lysates [55]. The authors have shown that this discrepancy cannot be attributed to the different monomer and dimer affinities, since both conformations are nearly equipotent [59]. They have suggested that SorLA requires a partner protein such as a co-receptor to reach high affinity interaction with the head activator [59]. In a pull-down fraction, a wide range of proteins is present, including the putative parter of SorLA, leading to high affinity binding, while in a purified system with only SorLA present, the receptor partner must be missing, resulting in a low affinity.
Although the mechanism of controlling neuronal differentiation by the head activator has remained enigmatic, there are some data indicating involvement of a stimulatory GPCR. It appears very likely that this pathway induces activation of the adenylyl cyclase (AC) and adenylyl cyclase associated-protein (CAP), causing the intracellular concentration of the cyclic adenosine monophosphate (cAMP) to rise [50]. Of note, the CAP proteins can regulate actin filament dynamics [61], and possess a high degree of homology between cnidarians and mammals [50]. Upregulation of cAMP results in activation of the cAMP response element binding protein (CREB) transcription factor, likely through the protein kinase A (PKA), and the activated CREB then modulates the transcriptional activity of differentiation-associated target genes [45]. Of note, CREB is evolutionary conserved between Hydra and mammals [45]. Synthesis of the presented data implies that the head activator signaling cascade seems to be heavily dependent on the regulation of the AC-cAMP-PKA axis, which is indeed one of central entities in regenerative processes across multiple species and cell types. This pathway has been reported to be involved in head regeneration [62], neuronal determination and differentiation [45] in Hydra. HAB or SorLA is likely the primary receptor that facilitates the head activator signaling, however the resulting effect depends on the previously described dynamic conditions and differential co-receptor expression. The identity of the head activator G-protein coupled co-receptors that could regulate this pathway have however remained unknown.
5. Effects of Exogenous Head Activator Applied to Mammals
Strikingly, mitogenic and neurogenic responses that were observed in Hydra could be reproduced in mammals through application of the head activator both in cell culture and in vivo. Mammalian cell culture experiments have shown that addition of the head activator to the culturing medium elevated mitosis rates of different cell lines, in particular the P19 embryonal carcinoma cells [63], NH-15-CA2 neuroblastoma-glioma cells [38,64], NT2 neuronal precursor cell line [55], BON neuroendocrine cell line [55,65], COS-7, HEK-293 [60], and AtT20 cells [29].
Furthermore, exogenous application of the head activator peptide induced neuronal differentiation and stimulated neurite formation in murine neuroblastoma Neuro-2A cells [66] and in P19 embryonal carcinoma cells [63]. Supplemented head activator promoted survival of dorsal root ganglion neurons and paravertebral sympathetic ganglion neurons isolated from chicken embryos in a culture devoid of neurotrophic factors that would have otherwise not remained viable for long time [66]. Similarly, the head activator stimulated growth of cultured embryonal chicken brain cells [67]. The neurotrophic activity in culture and the cerebral expression of the head activator in vivo led Schaller and Bodenmüller to a conclusion that the head activator may have a similar function in the human nervous system as observed in Hydra, namely to act as a regulator of neural development [20].
A large row of experiments with in vivo application of the head activator in rodents and cats showed its rapid systemic distribution [68] and differential tissue- and dose-dependent pleiotropic effects across multiple organs (Table 3).
The native head activator peptide is hydrophobic with an estimated half-life time less than 30 seconds, and after administration, the peptide is quickly absorbed by various organs and transformed into hydrophilic derivatives [68]. The effects of the head activator could be observed at dosages of 10-100 μg/kg, furthermore some effects were dose-dependent, and the responses varied based on the administered dosage (Table 3), favouring different context-dependent signaling mechanisms akin to cnidarian signaling.
Notably, application of the head activator to pregnant rats has elevated the number of full-term pregnancies from 66.3% in control to 100% in treated animal groups [76]. Sakura and colleagues could show that the head activator has been detected in the human placenta, specifically in the trophoblasts, and in the maternal bloodstream, additionally the head activator concentration has increased over the course of gestation [84]. This suggests that the head activator may support the placenta in maintaining fetal development. We assume that this effect could be used to counteract placenta failure and prolong the intrauterine period sufficiently for fetus maturation.
The head activator has been shown to stimulate epithelial proliferation in tongue, trachea, cornea, and pylorus, but has not affected the smooth myocytes in the respiratory and gastrointestinal tract [69,70,72]. These results imply that the head activator could stimulate wound epithelialisation both for surface wounds and mucous membrane lesions. Additionally, the head activator has attenuated tracheal tissue alterations in rats subjected to hypoxia [71]. Lebed’ko and colleagues have proposed that normalisation of the respiratory epithelial and myocyte proliferation under the head activator treatment may be used to modify the pathologic tissue remodelling observed in bronchopulmonary disorders such as asthma and chronic obstructive pulmonary disease [68]. These disorders are associated with epithelial abnormalities and hypertrophy of the smooth muscles of the bronchi [85]; the head activator has displayed exactly opposite effects which could alleviate these anomalies [68].
The gastrointestinal system seems to be also affected by the head activator. The peptide has led to an increase of the proliferation of gastric epithelium [70] and secretion of amylase in cultured pancreatic cells [82]. Interestingly, the duodenal epithelium did not respond to the head activator [81]. Under prenatal hypoxia, administered head activator has normalised the rate of hepatocyte proliferation [71]. Furthermore, normalisation of the hepatic protein content and dose-dependent modulation of the hepatic ornithine decarboxylase, an enzyme that synthesises polyamines and reflects the hepatocyte metabolic and mitotic activity [78,79], could be observed. These findings point to hepatoprotective properties, which may be interesting for treatment of liver disorders.
There are also cardiovascular implications of the head activator peptide. Fedoseev and colleagues could demonstrate remodelling of the myocardial tissue upon compensatory left ventricular hypertrophy induced by an artificial coarctation of the aorta treated with the head activator [73,74]. The structural changes in the myocardium showed spatial and temporal dynamics, in particular the histological alterations were most prominent in the subepicardial layer; the peptide has influenced the cardiomyocyte size, extent of fibrosis, and capillary area [73,74], as well as increased proliferation in the rat myocardium [75]. These interesting findings would demand further analysis of the head activator effects in the scenario of a heart failure and cardiomyopathies.
Moreover, the morphogen has reduced stress response in animals by regulating the hormonal and immune axes and attenuated lipid peroxidation. The head activator has normalised glutathione peroxidase (GPO) activity, elevated α-tocopherol and total lipid levels [71,72]. Additionally, the peptide has increased thymocyte proliferation rate and number of peripheral blood lymphocytes to normal reference in rats, suggesting an immunomodulatory activity [76]. In rats subjected to stress by immobilisation, the head activator has normalised the thyrotropic stimulating hormone (TSH), triiodthyronine (T3), thyroxine (T4), and corticosterone levels [70]. The morphogen has attenuated lipid peroxidation by reducing the lipoperoxide content in blood [70]. These combined studies reveal a cytoprotective and pro-regenerative potential of the head activator.
Administration of the head activator peptide to rats, not subjected to any kind of stress, revealed dose-dependent changes in hormone levels. At a lower dosage (10 μg/kg), β-endorphin was downregulated, while noradrenaline and corticosterone were upregulated; and at a higher dosage (100 μg/kg) β-endorphin, T3 and insulin were upregulated, while corticosterone and noradrenalin were downregulated [77]. These hormonal alterations seem to follow a logical pattern in a sense of a fight-or-flight reaction, the low-dose effect activates the stress-related catabolic hormones, while the high-dose effect does the opposite steady-state anabolic activation.
The mechanism that underlies hormonal alterations induced by the head activator has remained unclear. Kalenikova and colleagues could show that radiolabelled head activator is enriched in the pituitary gland and the pineal body after intravenous administration [68]. The pituitary gland, being a circumventricular organ, is supplied by fenestrated capillaries that allow hormones to be secreted into the blood stream, in contrast to the mostly impenetrable blood-brain-barrier in other brain regions [86]. The pituitary enrichment of the head activator is likely attributed to the special characteristics of the blood-brain barrier in this region. We do not rule out that this might allow the head activator to penetrate the barrier and regulate the secretion of pituitary hormones, thus resulting in the described hormonal disturbances. Our assumption also refers to the results of Drozd and colleagues, who have discovered an antiemetic effect of the head activator in a cat model [80]. Area postrema, the emesis centre located in the brainstem, is also a circumventricular organ, which contains an incomplete leaky blood-brain barrier [87]. The antiemetic effect may be dependent on the modulation of this brain region.
The great body of scientific literature on the head activator points towards a multifunctional nature. Due to its multiple effects and pleiotropic signaling the head activator peptide can be considered as an appealing therapeutic molecule.
Small bioactive molecules can trigger pleiotropic biological responses with high affinity and specificity, while tending to have low toxicity, immunogenicity and production costs, compared to classic antibody-based biologicals [88]. The therapeutic peptides cover a wide range of applications, reaching from simple hormone mimetics such as insulin analogues to more complex compounds such as the recently booming GLP-1 receptor agonists in diabetes and obesity therapy, an osteoanabolic fragment of parathormone teriparatide, somatostatin mimetics, anabolic ghrelin mimetics used to treat cachexia in cancer patients, and glatiramer acetate for multiple sclerosis [88]. However, there is a huge demand for therapeutic molecules to counteract nervous system disorders. Cnidarian peptides could set new frontiers in neuroregeneration, due to their morphogenic nature, which enables cell proliferation and de novo formation of new tissues.
6. Evolutionary Conservation of the Head Activator
Since exogenously applied head activator is biologically active in mammals, a central question that arises is whether the head activator itself or its signaling cascade is evolutionary conserved. In this respect, it is important to mention that detecting homologs or orthologs in phylogenetically distant taxa such as Cnidaria and Mammalia is an untrivial task, since such extreme phylogenetic divergence may result in a loss of evolutionary information. The dominating academic opinion is that the peptidergic systems of Cnidaria and Bilateria evolved mostly independently from each other [17]. Despite this fact, we can still see parallels in the basic principles of cnidarian and bilaterian peptidergic signaling. The peptides are usually synthesised as preprohormones containing multiple copies of a peptide or even multiple different peptides that are liberated via proteolytic processing [17]. Some basic structural features are shared by Cnidaria and Bilateria, e.g. the N-terminal pyroglutamylation and the C-terminal amidation [17]. The neuropeptidergic signaling in Cnidaria and Bilateria is usually dependent on G-protein coupled receptors (GPCRs), however only the leucine-rich repeat-containing GPCRs have been identified to be conserved between Cnidaria and Bilateria [17].
Regardless of the tremendously divergent sequences of orthologous proteins, it seems that the 3D structure of the orthologues can be significantly preserved between distantly related species. For instance, the tissue inhibitors of the matrix metalloproteases (TIMPs) are unified by a universal prototype structure that is significantly congruent even in distant taxa [89]. We can deduce that the structural resemblance may allow the interaction of the head activator with the mammalian receptors according to the “keyhole-lock-key” model without the necessity of the sequence preservation. This fact could explain the functional activity of the head activator in mammals.
A peptide identical to the cnidarian head activator has been detected in the human, bovine and rodent hypothalamus, placenta, intestine and blood [32,84,90,91,92], as well as in biopsies and serum samples from patients with neural and neuroendocrine tumours [93,94]. In fact, the head activator is the only known morphogenic peptide that is conserved between Cnidaria and Mammalia, since no other homologs could be detected in cnidarian and bilaterian neuropeptides and their receptors [17]. Furthermore, endogenous expression of the head activator was confirmed in P19 embryonal carcinoma cells [63], NH-15-CA2 neuroblastoma-glioma cells [38,64], NT2 neuronal precursor cell line [55], and in BON neuroendocrine cell line [55,65]. Interestingly, the head activator is structurally similar to bradykinin, but different in functions, since bradykinin failed to reproduce the neurotrophic effects of the head activator [66].
It should be emphasised that the head activator has been detected in mammals only at the protein level, and attempts to clone a corresponding gene have failed. Major criticism has been raised towards the detection of the peptide in mammals due to reproducibility difficulties, but little attempt has been made to identify the peptide via alternative methods such as mass-spectrometry. The referenced studies on peptide detection in multiple cell lines and tissues have used immunocyto- and histochemical stainings, direct chemical isolation, and sequencing so far. The generated antibodies might detect similar epitopes of related peptides. Furthermore, the short half-life of the head activator that has been estimated to be less than 30 seconds [68] has to be taken under consideration too. The question of the evolutionary conservation remains still unresolved.
7. Signaling Pathway of the Head Activator Peptide in Mammals
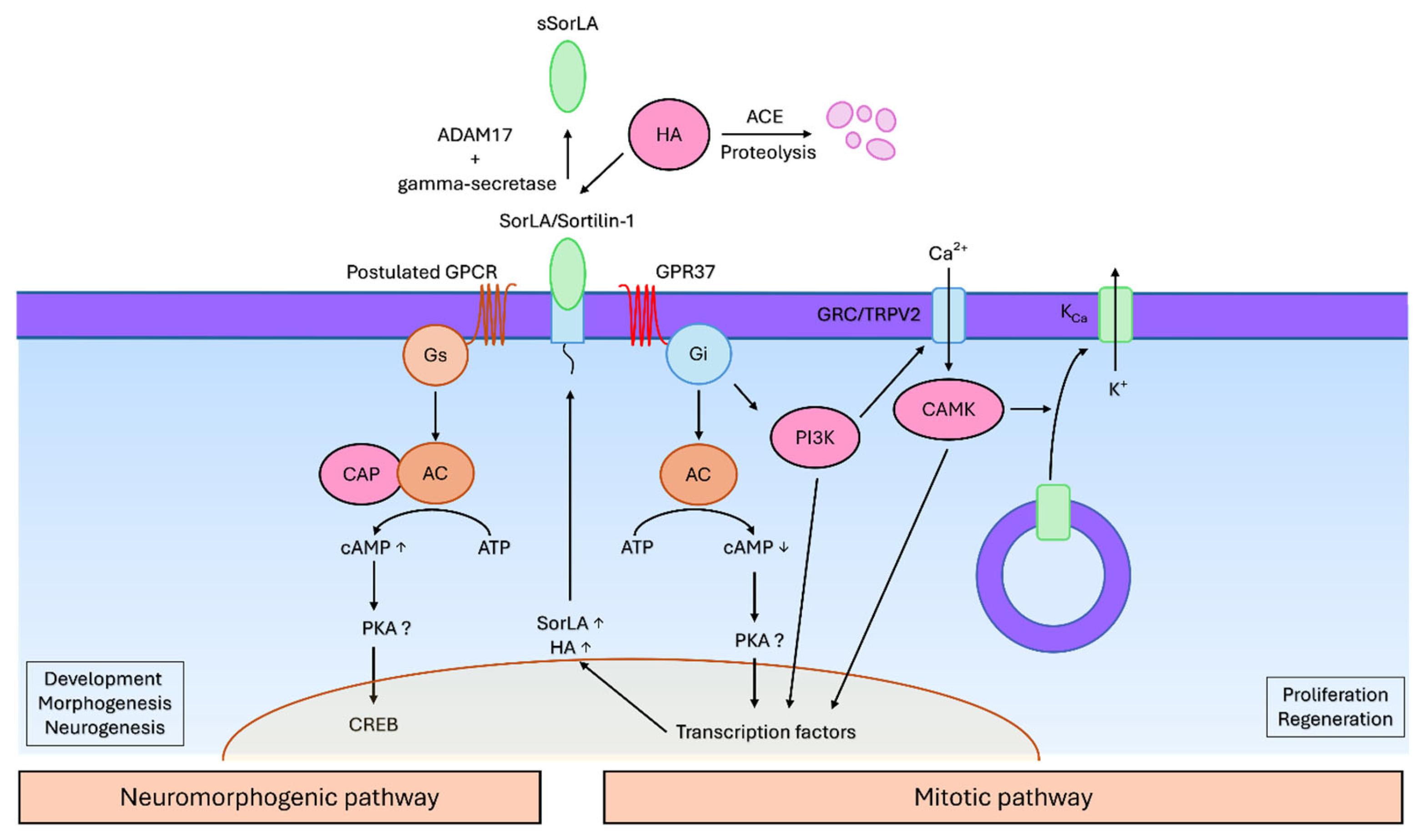
The signaling pathway of the head activator in mammalian cells seems to occur in a similar manner compared to the cnidarian pathway (Figure 4). The first step is the binding of the head activator to the VPS10 domain [59,95] of SorLA (SORL1, sortilin related protein) [55]. This transmembrane receptor can exist in a membrane-bound and in an extracellular soluble form generated by metalloprotease shedding [55], analogously to the head activator binding protein in the cnidarian pathway [56]. The proteolytic processing occurs through ADAM17 (a disintegrin and metalloprotease 17, also known as TACE or tumour necrosis factor-α-converting enzyme) and gamma secretase [96]. Interestingly, a structurally and functionally related receptor sortilin-1 has also a low affinity to the head activator [59]. It becomes thus clear that SorLA and sortilin-1 are involved in the head activators’ mechanism of action. The unbound head activator is rapidly degraded through proteolytic processing by angiotensin converting enzyme [92]. Blocking SorLA with antisense oligonucleotides has neutralised the mitogenic effect of the head activator in NT2 cells, favouring its mitogenic signaling through SorLA [55]. The subsequent molecular response that controls mitosis occurs in a pertussis toxin-sensitive Gi-protein-dependent manner [64,65], in contrast to the cAMP response that controls neurogenesis, which would require a Gs protein [50]. This is consistent with the cnidarian pathway, where the cAMP axis is also being regulated, which then points to the likelihood of a forming SorLA/GPCR complex. Additionally, previously described findings of Jacobsen and colleagues also support the receptor complex as a signaling mediator [59].
Rezgaoui and colleagues have reported that the involved inhibitory GPCR is the orphan GPR37 that seems to exist in a complex with SorLA and internalises upon head activator treatment [60]. Furthermore, a direct ligand-receptor interaction between the head activator and GPR37 could be observed [60]. Another work has also detected clustering and internalisation of the GPR37 and inhibition of the adenylyl cyclase upon treatment with head activator [97]. Meyer and colleagues have noted that the head activator and prosaptide (another putative ligand of the GPR37) have a similar C-terminal region, favouring the head activator/GPR37 interaction [98]. Controversially, Dunham and colleagues could not reproduce such an interaction [99]. In an attempt to deorphanise some GPCRs, a high-throughput β-arrestin screening assay for potential ligands has also failed to show any association between the head activator and GPR37 [100]. Another author has also expressed concerns regarding the interpretation of the results obtained by Rezgaoui and Gandía, since the presented images were not entirely consistent with the receptor internalisation [101].
After binding to its receptors, the head activator/SorLA-complex (and possibly GPR37) becomes internalised [55,60]. In parallel, the expression of SorLA rises in a sense of an autocatalytic feedback loop, and the unbound pool of SorLA in the intracellular compartment translocates to the cell surface [55].
This culminates in a calcium-influx and calcium-dependent (gardos-type) potassium efflux, leading to a hyperpolarisation of the cells [64,65]. The initial calcium-influx is provided by a transient receptor potential cation channel subfamily V member 2 (TRPV2), previously known as growth-factor-regulated calcium-channel (GRC), which externalises from the intracellular compartment to the cell surface [102]. This translocation requires the phosphoinositide-3-kinase and calmodulin-kinase [60,102]. The following steps of the cascade have not yet been characterised. A reasonable assumption is that the shifted ion levels and activated kinases might affect transcription factors, such as CREB, to regulate the expression of developmental and morphogenic genes.
The signaling cascade of the head activator that controls neuronal differentiation in higher animals has not been extensively studied, however there are indications that it may occur in a similar way to the previously described cnidarian cAMP-PKA-CREB pathway. While working on cultured embryonal chicken brain cells, Kajiwara and Sato have found that head activator peptide administration has led to a rise in cAMP, total protein concentrations, and incorporation of radiolabelled nucleotides [67], as seen also in Hydra. The studied culture has been chosen to be a mixture of all cell types present in the brain, and due to technical limitations, a distinction between neuronal and glial or precursor cell response was not possible. The incorporation of the nucleotides has been attributed to the proliferation of dividing cells, and rising cAMP concentration has been related to the development of post-mitotic differentiating cells [67]. Similarly, in injured hepatic tissue from rats, the head activator has been shown to induce a biphasic response with initially elevated cAMP concentration attributed to the inhibition of the repair processes, and later on, with cAMP reduction related to the activation of tissue repair [51]. Both phases have been accompanied by inversely oriented cyclic guanosine monophosphate (cGMP) concentration changes [51]. This corroborates the hypothesis, that the head activator shifts the GPCR activity between inhibitory and stimulatory G-proteins, and thus temporally changes its mode of action dependent on the differentiation status of the target cells, similarly to the cnidarian pathway.
Whether the head activator can act as a ligand for the GPR37 receptor has been a subject of a heated discussion in the scientific community. This conflict is likely the reason why research on the head activator was not pursued further. Experiments on GPR37 are methodically a challenging undertaking, since its expression is mainly restricted to the cytosol in heterologously overexpressing cells, and little number of receptor molecules are exposed on the cell surface, leading to no apparent activation or ligand binding [103]. Southern and colleagues did not address this factor. To overcome this difficulty Dunham and colleagues have ensured that their construct had sufficient surface presentation of GPR37 through several different approaches: N-terminal truncation of the receptor or its co-expression with syntenin-1 [99]. This approach however did not yield any head activator-induced response, presenting strong evidence against the head activator/GPR37 interaction. However, we have to mention that neither Dunham nor Southern have tested their cell constructs for the expression of SorLA, the putative component of the head activator receptor complex, hence one component of the signaling cascade could have been missing. This could mean that GPR37 alone is not enough to induce a response and may only signal in complex with SorLA. Another point of criticism around Dunhams’ work is the absence of any control condition to compare with the head activator treatment. It is unclear whether GPR37 truncation or syntenin-1 co-expression, performed by Dunham and colleagues, could completely abolish the receptor activity, and therefore no head activator-induced response has been observed. An alternative way to increase the receptor surface recruitment without potentially interfering with the receptor activity could be the treatment with chemical chaperones such as 4-PBA [104]. These results need to be reviewed with caution. We believe that more appropriate (cell) models need to be established to unravel the functions of the head activator. In particular, considering the primarily neuronal origin and activity of the head activator, the choice of neuronal cells/cell lines seems to be more appropriate, furthermore the membrane localisation of the GPR37 receptor and its co-expression with SorLA must be ensured. Confirmation studies using primary neuronal cells and/or their precursors cells rather than immortalised cell lines as well as in vivo testing would help to find an answer. Altogether, according to the available literature there is stronger evidence for SorLA than GPR37 as a component of the head activators’ signaling pathway, the GPR37 involvement has yet to be confirmed, but it shall not be dismissed. We believe that a broader approach without focusing on a single receptor would be a fitting way to study the head activator signaling. For instance, qPCR analysis of a broad receptor panel or a mass spectrometric analysis of a pull-down fraction against an immobilised head activator could shed more light on the specific signaling cascade.
Figure 4.
Signaling pathways of the head activator. The head activator peptide context-dependently activates two functionally different signaling pathways – a mitotic and a neuromorphogenic cascade. The central receptor for both cascades is the SorLA transmembrane protein. It forms a complex with an inhibitory GPCR to initiate mitosis, or a stimulatory GPCR (potentially GPR37) to induce neuronal differentiation. In the neuromorphogenic pathway, a Gs protein activates the adenylyl cyclase (AC) to synthesise cyclic adenosine monophosphate (cAMP) as a second messenger, which in turn modulates the expression of target genes through the CREB transcription factor. In the mitotic pathway a Gi protein is activated, leading to a depletion of the cAMP concentration. In parallel the Gi protein activates the phosphoinositide 3 kinase (PI3K), that activates the GRC/TRPV2 calcium channel. The resulting calcium influx activates the calmodulin kinase (CAMK), which induces an externalisation of the calcium dependent potassium channel (KCa) and consequently a calcium dependent potassium efflux and hyperpolarisation. Following the ion rearrangements and kinase activations, mitosis occurs, likely accompanied by transcription factors regulating the necessary genes.
Figure 4.
Signaling pathways of the head activator. The head activator peptide context-dependently activates two functionally different signaling pathways – a mitotic and a neuromorphogenic cascade. The central receptor for both cascades is the SorLA transmembrane protein. It forms a complex with an inhibitory GPCR to initiate mitosis, or a stimulatory GPCR (potentially GPR37) to induce neuronal differentiation. In the neuromorphogenic pathway, a Gs protein activates the adenylyl cyclase (AC) to synthesise cyclic adenosine monophosphate (cAMP) as a second messenger, which in turn modulates the expression of target genes through the CREB transcription factor. In the mitotic pathway a Gi protein is activated, leading to a depletion of the cAMP concentration. In parallel the Gi protein activates the phosphoinositide 3 kinase (PI3K), that activates the GRC/TRPV2 calcium channel. The resulting calcium influx activates the calmodulin kinase (CAMK), which induces an externalisation of the calcium dependent potassium channel (KCa) and consequently a calcium dependent potassium efflux and hyperpolarisation. Following the ion rearrangements and kinase activations, mitosis occurs, likely accompanied by transcription factors regulating the necessary genes.
8. Potential Role of the Head Activator in Human Neuropathology
SorLA has been reported to participate in the pathogenesis of Parkinson’s [105] and Alzheimer’s disease, atherosclerosis, type 2 diabetes, obesity [58], HER-2 driven cancers such as breast and ovarian cancer [106,107,108], and in immune-mediated processes such as neuroinflammation and pathogen defence [109]. SorLA belongs to the group of the VPS10 domain receptors; and is mostly associated to the Golgi apparatus where it regulates intracellular trafficking of various proteins [58]. The receptor molecule is structurally similar to low-density lipoprotein receptor and can also internalise lipoproteins into cells [58]. Furthermore, SorLA can facilitate intracellular transport of glial derived neurotrophic factor and its receptors [110]. Under inflammatory conditions, SorLA regulates cytokine secretion, and thus the activity of microglia, macrophages, and lymphocytes as observed in neuroinflammatory, neurodegenerative and even psychiatric disorders [109]. Truncated variants of SorLA are associated with development of Alzheimer’s disease [111]. Wild-type SorLA is a protective factor that binds amyloid precursor protein and prevents its proteolytic processing into peptides that can nucleate into cytotoxic aggregates [112,113,114]. Parkinson’s disease is a major pathology that is associated with GPR37 abnormalities [115]. GPR37, also known as rhodopsin-like orphan G-protein coupled receptor 37 or parkin-associated endothelin receptor-like receptor, is hypothesised to act as a neuropeptide receptor [115,116]. Its ligand is currently unknown, and the head activator has not been recognised as such. GPR37 is expressed in neurons, microglia and oligodendrocytes and coupled to a Gi/o protein that signals through the MAPK/Erk pathway [115]. In Parkinson’s disease, the misfolded GPR37 receptor forms cytotoxic aggregates responsible for the degeneration of the dopaminergic neurons. GPR37 fragments in the cerebrospinal fluid were suggested to be used as a biomarker of disease’s progression – interestingly, different fragment patterns of the receptor were detected in the idiopathic and atypic variants of Parkinson’s disease, allowing to differentiate between these phenotypes [117]. Under normal conditions, the native GPR37 has a neuroprotective role against oxidative stress and ischemia [115,118]. GPR37 has been linked to epilepsy, autism, neuroinflammation, and demyelination [115]. Loss of GPR37 has been associated with premature hypermyelination [119] and a higher likelihood of demyelination in a cuprizone model of multiple sclerosis [120], while preventing neurodegeneration in Parkinson’s disease models [121]. Apart from the nervous system, GPR37 is involved also in spermatogenesis and carcinogenesis [115].
As to the head activator, it is important to mention in this respect that endogenously expressed head activator has been found in immortalised brain tumour cell lines [64,66], and in human material of meningeomas, astrocytomas, and glioblastomas [93]. The patients also had displayed elevated systemic levels of the head activator peptide in their blood which normalised after the tumour resection, thus highlighting the head activator as a potential neurooncological biomarker [93,94]. If the head activator is indeed endogenously expressed and supports neurogenesis, it is reasonable to assume that different growth factors, including the molecule itself, might be expected to be elevated in tumorous brain tissue. However, one study has but found reduced levels of the head activator in astrocytoma in comparison to the normal tissue [122]. To explain these discrepancies, Ekman and colleagues have postulated different specificity of the antibodies used in both studies. Another important fact to be mentioned is that the head activator is exclusively a biomarker for neuronal and not glial tumours, as this neuropeptide is produced and stored in neurosecretory granula. In addition to that, we have to also consider the high heterogeneity of brain tumours where the expression of the head activator could vary significantly depending on type, characteristics and grading.
In summary, the head activator’s signaling pathway and its interconnection with different pathologic conditions, particularly those of the nervous system, are evident. SorLA and GPR37 are interesting targets that put the head activator at the core of many neurological diseases (Figure 5).
Addressing the head activator and its putative receptors may provide an opportunity to obtain useful insights into neuropathology and maybe identify novel therapeutic drugs. Alone, the interaction of the head activator peptide with SorLA is a significant framework applicable to neurological disorders. Furthermore, the association of the head activator with GPR37 is valuable for the neuropathology field.
9. Conclusions
In this work we focused on one cnidarian morphogen, the head activator neuropeptide, to revisit and discuss its roles in neurogenesis and regeneration. In Hydra, the head activator regulates regeneration and development of the head region, and concurrently nervous net maintenance by determining the neuronal fate of interstitial cells. Intriguingly, this peptide is active in both cnidarian and mammal tissues with similar effects. Multiple preclinical studies using rodents could demonstrate mitogenic effects of exogenously administered head activator across various organs. Beside mitosis, the peptide can also influence myocardial structure, hormonal axis, lipid and liver metabolism. Indirectly, cell culture experiments on various neuronal cells could assess neurogenic activity of the head activator as similarly as observed in Hydra. All of these findings strongly suggest the head activator to be a potent multifunctional molecule. Despite such compelling evidence, the head activator neuropeptide has been largely forgotten since its last publication in the early 2000s. The reason for this is likely due to several controversial findings on its evolution and signaling. Firstly, the gene of origin could never be identified and some findings reporting the head activator detection in mammalian tissues could not be reproduced. Furthermore, one of the head activator putative receptors, the orphan GPR37, has been heavily disputed. The question of the head activators’ evolutionary conservation requires further experimental testing to give a definitive answer bridging the primordial nervous nets of cnidarians and the complex nervous system of mammals. The experimental evidence for the peptide to act as an exogenous ligand on mammalian systems is intriguing. We have summarised reported findings on the signaling cascades controlled by the head activator: the peptide employs at least two different context-dependent pathways that are activated based on the mitotic activity and differentiation status of the targeted cells. The central receptor of the head activator seems to be the SorLA protein, which most likely engages in a complex with both stimulatory and inhibitory G proteins to facilitate further signaling through regulation of the cAMP axis. We have also collected and discussed in depth critical aspects around the head activator/GPR37 interaction. We also hypothesise that the head activator may be a central link between multiple neuropathologies ranging from neurodegeneration to myelination disorders including Alzheimer’s and Parkinson’s diseases, as well as multiple sclerosis.
Author Contributions
Conceptualization, A.K. and D.L.; methodology, A.K and D.L..; writing—original draft preparation, A.K. and D.L.; writing—review and editing, A.K. and D.L.; supervision, D.L.; funding acquisition, D.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the NextGenerationEU via the Bulgarian National Recovery and Resilience Plan, Project #BG-RRP-2.004-0009-C03 (to D.L.), and by the National Scientific Program “Excellent Research and People for the Development of European Science” 2021 (VIHREN) of the Bulgarian National Science Fund, contract #KP-06-DV/7 from 17.12.2024 (to D.L.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Van Iten, H.; Leme, J.M.; Pacheco, Mírian L. A. F.; Simões, M.G.; Fairchild, T.R.; Rodrigues, F.; Galante, D.; Boggiani, P.C.; Marques, A.C. Origin and Early Diversification of Phylum Cnidaria: Key Macrofossils from the Ediacaran System of North and South America. Palaeontology 2014, 677–690. [CrossRef]
- Reddy, P.C.; Gungi, A.; Unni, M. Cellular and Molecular Mechanisms of Hydra Regeneration. Results and problems in cell differentiation 2019, 68, 259–290. [CrossRef]
- Yuste, R. Breaking the neural code of a cnidarian: Learning principles of neuroscience from the "vulgar" Hydra. Current Opinion in Neurobiology 2024, 86, 102869. [CrossRef]
- Tomczyk, S.; Fischer, K.; Austad, S.; Galliot, B. Hydra, a powerful model for aging studies. Invertebrate Reproduction & Development 2015, 59, 11–16. [CrossRef]
- Lucia Mattera. Functionalization, bio-conjugation and toxicity studies of quantum dots, 2016.
- Buzgariu, W.; Al Haddad, S.; Tomczyk, S.; Wenger, Y.; Galliot, B. Multi-functionality and plasticity characterize epithelial cells in Hydra. Tissue Barriers 2015, 3, e1068908. [CrossRef]
- Plachetzki, D.C.; Fong, C.R.; Oakley, T.H. Cnidocyte discharge is regulated by light and opsin-mediated phototransduction. BMC Biology 2012, 10, 17. [CrossRef]
- Bellantuono, A.J.; Bridge, D.; Martínez, D.E. Hydra as a tractable, long-lived model system for senescence. Invertebrate Reproduction & Development 2015, 59, 39–44. [CrossRef]
- Ojimi, M.C.; Isomura, N.; Hidaka, M. Telomerase activity is not related to life history stage in the jellyfish Cassiopea sp. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 152, 240–244. [CrossRef]
- Pascual-Torner, M.; Carrero, D.; Pérez-Silva, J.G.; Álvarez-Puente, D.; Roiz-Valle, D.; Bretones, G.; Rodríguez, D.; Maeso, D.; Mateo-González, E.; Español, Y.; et al. Comparative genomics of mortal and immortal cnidarians unveils novel keys behind rejuvenation. Proc. Natl. Acad. Sci. U. S. A. 2022, 119, e2118763119. [CrossRef]
- Primack, A.S.; Cazet, J.F.; Little, H.M.; Mühlbauer, S.; Cox, B.D.; David, C.N.; Farrell, J.A.; Juliano, C.E. Differentiation trajectories of the Hydra nervous system reveal transcriptional regulators of neuronal fate. bioRxiv 2023, 2023.03.15.531610. [CrossRef]
- Noro, Y.; Shimizu, H.; Mineta, K.; Gojobori, T. A single neuron subset governs a single coactive neuron circuit in Hydra vulgaris, representing a possible ancestral feature of neural evolution. Sci Rep 2021, 11, 10828. [CrossRef]
- Dupre, C.; Yuste, R. Non-overlapping Neural Networks in Hydra vulgaris. Current biology : CB 2017, 27, 1085–1097. [CrossRef]
- Keramidioti, A.; Schneid, S.; Busse, C.; Laue, C.C. von; Bertulat, B.; Salvenmoser, W.; Heß, M.; Alexandrova, O.; Glauber, K.M.; Steele, R.E.; et al. A new look at the architecture and dynamics of the Hydra nerve net. eLife 2024, 12. [CrossRef]
- Badhiwala, K.N.; Primack, A.S.; Juliano, C.E.; Robinson, J.T. Multiple neuronal networks coordinate Hydra mechanosensory behavior. eLife Sciences Publications, Ltd [Online], July 30, 2021. Available online: https://elifesciences.org/articles/64108#bib64 (accessed on 25 June 2025).
- Rentzsch, F.; Layden, M.; Manuel, M. The cellular and molecular basis of cnidarian neurogenesis. Wiley Interdisciplinary Reviews. Developmental Biology 2017, 6, e257. [CrossRef]
- Hauser, F.; Koch, T.L.; Grimmelikhuijzen, C.J.P. Review: The evolution of peptidergic signaling in Cnidaria and Placozoa, including a comparison with Bilateria. Frontiers in endocrinology 2022, 13, 973862. [CrossRef]
- Liao, Q.; Feng, Y.; Yang, B.; Lee, S.M.-Y. Cnidarian peptide neurotoxins: a new source of various ion channel modulators or blockers against central nervous systems disease. Drug Discovery Today 2019, 24, 189–197. [CrossRef]
- Jayathilake, J.M.N.J.; Gunathilake, K.V.K. Cnidarian toxins: recent evidences for potential therapeutic uses. The European Zoological Journal 2020, 87, 708–713. [CrossRef]
- Schaller, H.C.; Bodenmüller, H. Structure and Function of the Head Activator in Hydra and in Mammals. In Advances in Metabolic Disorders : Gastrointestinal Hormones; Mutt, V., Ed.; Elsevier, 1988; pp 519–526, ISBN 0065-2903.
- Hicklin, J.; Hornbruch, A.; Wolpert, L.; Clarke, M. Positional information and pattern regulation in hydra: the formation of boundary regions following axial grafts. Development (Cambridge, England) 1973, 30, 701–725. [CrossRef]
- Chica Schaller, H. Neuropeptides in Hydra. Trends in Neurosciences 1979, 2, 120–122. [CrossRef]
- Takahashi, T.; Muneoka, Y.; Lohmann, J.; Lopez de Haro, M.S.; Solleder, G.; Bosch, T.C.; David, C.N.; Bode, H.R.; Koizumi, O.; Shimizu, H.; et al. Systematic isolation of peptide signal molecules regulating development in hydra: LWamide and PW families. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 1241–1246. [CrossRef]
- Fujisawa, T. Hydra peptide project 1993-2007. Development, growth & differentiation 2008, 50 Suppl 1, S257-68. [CrossRef]
- Takahashi, T.; Fujisawa, T. Important roles for epithelial cell peptides in hydra development. BioEssays : news and reviews in molecular, cellular and developmental biology 2009, 31, 610–619. [CrossRef]
- Takahashi, T.; Hayakawa, E.; Koizumi, O.; Fujisawa, T. Neuropeptides and their functions in Hydra. BIOLOGIA FUTURA 2008, 59 Suppl, 227–235. [CrossRef]
- Schaller, H.C.; Bodenmüller, H. Isolation and amino acid sequence of a morphogenetic peptide from hydra. Proc. Natl. Acad. Sci. U. S. A. 1981, 78, 7000–7004. [CrossRef]
- Hoffmeister, S.A. Isolation and characterization of two new morphogenetically active peptides from Hydra vulgaris. Development (Cambridge, England) 1996, 122, 1941–1948. [CrossRef]
- Schaller, H.C.; Hoffmeister, S.A.; Dübel, S. Role of the neuropeptide head activator for growth and development in hydra and mammals. Development (Cambridge, England) 1989, 107 Suppl, 99–107. [CrossRef]
- Lohmann, J.U.; Bosch, T.C. The novel peptide HEADY specifies apical fate in a simple radially symmetric metazoan. Genes & Development 2000, 14, 2771–2777. [CrossRef]
- Takahashi, T.; Koizumi, O.; Ariura, Y.; Romanovitch, A.; Bosch, T.C.; Kobayakawa, Y.; Mohri, S.; Bode, H.R.; Yum, S.; Hatta, M.; et al. A novel neuropeptide, Hym-355, positively regulates neuron differentiation in Hydra. Development (Cambridge, England) 2000, 127, 997–1005. [CrossRef]
- Bodenmüller, H.; Schaller, H.C. Conserved amino acid sequence of a neuropeptide, the head activator, from coelenterates to humans. Nature 1981, 293, 579–580. [CrossRef]
- Hoffmeister-Ullerich, S.A. The foot formation stimulating peptide pedibin is also involved in patterning of the head in hydra. Mechanisms of Development 2001, 106, 37–45. [CrossRef]
- Herrmann, D.; Hatta, M.; Hoffmeister-Ullerich, S.A.H. Thypedin, the multi copy precursor for the hydra peptide pedin, is a beta-thymosin repeat-like domain containing protein. Mechanisms of Development 2005, 122, 1183–1193. [CrossRef]
- Takahashi, T.; Takeda, N. Insight into the molecular and functional diversity of cnidarian neuropeptides. International journal of molecular sciences 2015, 16, 2610–2625. [CrossRef]
- Takahashi, T.; Koizumi, O.; Hayakawa, E.; Minobe, S.; Suetsugu, R.; Kobayakawa, Y.; Bosch, T.C.G.; David, C.N.; Fujisawa, T. Further characterization of the PW peptide family that inhibits neuron differentiation in Hydra. Archiv f mikr Anat u Entwicklungsmechanik 2009, 219, 119–129. [CrossRef]
- Morishita, F.; Nitagai, Y.; Furukawa, Y.; Matsushima, O.; Takahashi, T.; Hatta, M.; Fujisawa, T.; Tunamoto, S.; Koizumi, O. Identification of a vasopressin-like immunoreactive substance in hydra. Peptides 2003, 24, 17–26. [CrossRef]
- Schawaller, M.; Schenck, K.; Hoffmeister, S.A.; Schaller, H.; Schaller, H.C. Production and characterization of monoclonal antibodies recognizing head activator in precursor form and immunocytochemical localization of head activator precursor and head activator peptide in the neural cell line NH15-CA2 and in hydra. Differentiation; research in biological diversity 1988, 38, 149–160. [CrossRef]
- Schaller, H.C. Action of the head activator as a growth hormone in hydra. Cell Differentiation 1976, 5, 1–11. [CrossRef]
- Schaller, H.C. Action of the head activator on the determination of interstitial cells in hydra. Cell Differentiation 1976, 5, 13–25. [CrossRef]
- Schaller, H.C.; Hofmann, M.; Javois, L.C. Effect of head activator on proliferation, head-specific determination and differentiation of epithelial cells in hydra. Differentiation; research in biological diversity 1990, 43, 157–164. [CrossRef]
- Hobmayer, B.; Holstein, T.W.; David, C.N. Stimulation of tentacle and bud formation by the neuropeptide head activator in Hydra magnipapillata. Developmental biology 1997, 183, 1–8. [CrossRef]
- H. Schaller; I. Hermans-Borgmeyer; S. Hoffmeister. Neuronal control of development in hydra. International Journal of Developmental Biology 1996.
- Hampe, W.; Hermans-Borgmeyer, I.; Schaller, H.C. Function of the neuropeptide head activator for early neural and neuroendocrine development. Results and problems in cell differentiation 1999, 26, 323–337. [CrossRef]
- Galliot, B.; Welschof, M.; Schuckert, O.; Hoffmeister, S.; Schaller, H.C. The cAMP response element binding protein is involved in hydra regeneration. Development (Cambridge, England) 1995, 121, 1205–1216. [CrossRef]
- Neubauer, K.H.; Christians, S.; Hoffmeister, S.A.; Kreger, S.; Schaller, H.C. Characterisation of two types of head activator receptor on hydra cells. Mechanisms of Development 1990, 33, 39–47. [CrossRef]
- Christians, S.; Neubauer, K.H.; Ulrich, H. Purification and characterization of the head-activator receptor from a multi-headed mutant of Chlorohydra viridissima. FEBS Letters 1993, 316, 141–146. [CrossRef]
- Franke, I.; Buck, F.; Hampe, W. Purification of a head-activator receptor from hydra. Eur. J. Biochem. 1997, 244, 940–945. [CrossRef]
- Schaller, H.C.; Druffel-Augustin, S.; Dübel, S. Head activator acts as an autocrine growth factor for NH15-CA2 cells in the G2/mitosis transition. The EMBO journal 1989, 8, 3311–3318. [CrossRef]
- Fenger, U.; Hofmann, M.; Galliot, B.; Schaller, H.C. The role of the cAMP pathway in mediating the effect of head activator on nerve-cell determination and differentiation in hydra. Mechanisms of Development 1994, 47, 115–125. [CrossRef]
- v. d. Slepushkin; Prum, I.A.; Vinogradov, V.A.; Pavlov, V.A.; Titov, M.I.; Vasil'ev, N.V. Effect of hydra peptide morphogen on cyclic nucleotide levels in injured tissues. Bull Exp Biol Med 1989, 108, 1491–1493. [CrossRef]
- Bodenmüller, H.; Schilling, E.; Zachmann, B.; Schaller, H.C. The neuropeptide head activator loses its biological acitivity by dimerization. The EMBO journal 1986, 5, 1825–1829. [CrossRef]
- Schaller, H.C.; Roberge, M.; Zachmann, B.; Hoffmeister, S.; Schilling, E.; Bodenmüller, H. The head activator is released from regenerating Hydra bound to a carrier molecule. The EMBO journal 1986, 5, 1821–1824. [CrossRef]
- Lai, J.R.; Gellman, S.H. Reinvestigation of the proposed folding and self-association of the Neuropeptide Head Activator. Protein science : a publication of the Protein Society 2003, 12, 560–566. [CrossRef]
- Hampe, W.; Riedel, I.B.; Lintzel, J.; Bader, C.O.; Franke, I.; Schaller, H.C. Ectodomain shedding, translocation and synthesis of SorLA are stimulated by its ligand head activator. J Cell Sci 2000, 113 Pt 24, 4475–4485. [CrossRef]
- Hampe, W.; Urny, J.; Franke, I.; Hoffmeister-Ullerich, S.A.; Herrmann, D.; Petersen, C.M.; Lohmann, J.; Schaller, H.C. A head-activator binding protein is present in hydra in a soluble and a membrane-anchored form. Development (Cambridge, England) 1999, 126, 4077–4086. [CrossRef]
- Hermans-Borgmeyer, I.; Hampe, W.; Schinke, B.; Methner, A.; Nykjaer, A.; Süsens, U.; Fenger, U.; Herbarth, B.; Schaller, H.C. Unique expression pattern of a novel mosaic receptor in the developing cerebral cortex. Mechanisms of Development 1998, 70, 65–76. [CrossRef]
- Schmidt, V.; Subkhangulova, A.; Willnow, T.E. Sorting receptor SORLA: cellular mechanisms and implications for disease. Cell. Mol. Life Sci. 2017, 74, 1475–1483. [CrossRef]
- Jacobsen, L.; Madsen, P.; Jacobsen, C.; Nielsen, M.S.; Gliemann, J.; Petersen, C.M. Activation and functional characterization of the mosaic receptor SorLA/LR11. Journal of Biological Chemistry 2001, 276, 22788–22796. [CrossRef]
- Rezgaoui, M.; Süsens, U.; Ignatov, A.; Gelderblom, M.; Glassmeier, G.; Franke, I.; Urny, J.; Imai, Y.; Takahashi, R.; Schaller, H.C. The neuropeptide head activator is a high-affinity ligand for the orphan G-protein-coupled receptor GPR37. J Cell Sci 2006, 119, 542–549. [CrossRef]
- Rust, M.B.; Khudayberdiev, S.; Pelucchi, S.; Marcello, E. CAPt'n of Actin Dynamics: Recent Advances in the Molecular, Developmental and Physiological Functions of Cyclase-Associated Protein (CAP). Frontiers in cell and developmental biology 2020, 8, 586631. [CrossRef]
- Kaloulis, K.; Chera, S.; Hassel, M.; Gauchat, D.; Galliot, B. Reactivation of developmental programs: the cAMP-response element-binding protein pathway is involved in hydra head regeneration. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 2363–2368. [CrossRef]
- Niemann, S.; Schaller, H.C. Head-activator and the neuroectodermal differentiation of P19 mouse embryonal carcinoma cells. Neuroscience letters 1996, 207, 49–52. [CrossRef]
- Ulrich, H.; Tárnok, A.; Schaller, H.C. Head-activator induced mitosis of NH15-CA2 cells requires calcium influx and hyperpolarization. Journal of Physiology-Paris 1996, 90, 85–94. [CrossRef]
- Kayser, S.T.; Ulrich, H.; Schaller, H.C. Involvement of a Gardos-type potassium channel in head activator-induced mitosis of BON cells. European Journal of Cell Biology 1998, 76, 119–124. [CrossRef]
- Quach, T.T.; Duchemin, A.M.; Oliver, A.P.; Schrier, B.K.; Wyatt, R.J. Hydra head activator peptide has trophic activity for eukaryotic neurons. Developmental Brain Research 1992, 68, 97–102. [CrossRef]
- Kajiwara, S.; Sato, T. Growth promoting effect of head activator in cultured chick embryo brain cells. Acta Endocrinologica 1986, 113, 604–608. [CrossRef]
- Kalenikova, E.I.; Tishchenko, V.A.; Rubina, A.Y.; Sokolov, A.S.; Blidchenko, Y.A.; Kuznetsova, T.V.; Bespalova, Z.D. Distribution of a morphogenic peptide activating the head growth of hydra in rat organs following intravascular administration of3H-labeled peptide. Bull Exp Biol Med 1994, 118, 1162–1165. [CrossRef]
- Khomichuk, A.Y.; Timoshin, S.S. Participation of hydra peptide morphogen in regulation of proliferation of various albino rat tissues. Bull Exp Biol Med 1991, 112, 1332–1334. [CrossRef]
- Timoshin, S.S.; Yakovenko, I.G.; Berezina, G.P.; Obukhova, G.G.; Rubina, A.Y.; Murzina, N.B. Hydra peptide morphogen weakens poststress disturbances in albino rats. Bull Exp Biol Med 1997, 124, 965–967. [CrossRef]
- Lebed'ko, O.A.; Yatsenko, T.V.; Timoshin, S.S.; Rubina, A.Y. Effect of the hydra peptide morphogen on posthypoxic disorders in rats exposed to prenatal hypoxia. Bull Exp Biol Med 1997, 123, 232–234. [CrossRef]
- Lebed'ko, O.A.; Timoshin, S.S.; Rubina, A.Y. Effects of hydra peptide morphogen and its analogue and fragments on DNA synthesis in tracheal epithelium and smooth muscle cells in newborn albino rats. Bull Exp Biol Med 2000, 129, 550–552. [CrossRef]
- Fedoseev, V.A.; Kazimirskii, A.N.; Ionina, I.A.; Lomakina, E.B.; Rubina, A.Y.; Pylaev, A.S. Effect of hydra morphogen peptide on the dynamics of tissue components of the myocardial layers in compensatory hypertrophy of the heart. Bull Exp Biol Med 1993, 116, 1162–1164. [CrossRef]
- Fedoseev, V.A.; Kazimirskii, A.N.; Lomakina, E.B.; Ionina, I.A.; Rubina, A.Y.; Pylaev, A.S. Effect of hydra morphogen peptide on the structure of the tissue components of the myocardium layers in the early development of heart hypertrophy in rats. Bull Exp Biol Med 1993, 115, 339–341. [CrossRef]
- Sazonova, E.N.; Yakovenko, I.G.; Kryzhanovskaya, S.Y.; Budylev, A.A.; Timoshin, S.S. Effects of analogues of hydra peptide morphogen on DNA synthesis in the myocardium of newborn albino rats. Bull Exp Biol Med 2012, 152, 304–306. [CrossRef]
- Gan'cheva, E.A.; Kozlov, V.K.; Timoshin, S.S. Effect of the hydra peptide morphogen on cell proliferation in the thymus of newborn rats exposed to prenatal hypoxia. Bull Exp Biol Med 1997, 124, 929–931. [CrossRef]
- Murzina, N.B.; Khomichuk, A.Y.; Timoshin, S.S.; Obukhova, G.G.; Anosova, O.A.; Berezina, G.P. Effect of hydra peptide morphogen on levels of?-endorphin and certain hormones in albino rat blood and adrenals. Bull Exp Biol Med 1991, 112, 1510–1512. [CrossRef]
- Yarygin, K.N.; Kazimirskii, A.N.; Kositskii, G.I.; Rubina, A.Y.; Vinogradov, V.A.; Pylaev, A.S. Modulation of ornithine decarboxylase activity in the normal and regenerating rat liver by various doses of the peptide morphogen ofHydra. Bull Exp Biol Med 1986, 101, 755–757. [CrossRef]
- Kazimirskii, A.N.; Yarygin, K.N.; Kositskii, G.I.; Rubina, A.Y.; Vinogradova, V.A.; Pylaev, A.S. Effect of low doses of hydra peptide morphogen on protein synthesis in the intact and regenerating rat liver. Bull Exp Biol Med 1988, 105, 503–506. [CrossRef]
- Drozd, Y.V.; Yasnetsov, V.V.; Shashkov, V.S. Vestibuloprotective properties of some regulatory peptides. Bull Exp Biol Med 1988, 106, 971–973. [CrossRef]
- Sazonova, E.N.; Zhivotova, E.Y.; Sazonov, O.A.; Fleishman, M.Y.; Timoshin, S.S. Effects of regulatory peptides on DNA synthesis in duodenal smooth muscle tissues of albino rats during the early postnatal period. Bull Exp Biol Med 1999, 127, 593–595. [CrossRef]
- Feurle, G.E.; Bodenmüller, H.; Baća, I. The neuropeptide head activator stimulates amylase release from rat pancreas in vitro. Neuroscience letters 1983, 38, 287–289. [CrossRef]
- Khomichuk, A.Y.; Timoshin, S.S.; Orlov, S.N.; Pokudin, N.I.; Kubatiev, A.A. Hydra peptide morphogen activates Na/H exchange in human erythrocytes. Bull Exp Biol Med 1991, 111, 763–764. [CrossRef]
- Sakura, H.; Aoki, S.; Ozawa, T.; Hashimoto, T.; Sakura, N. The neuropeptide, head activator, in human placenta and serum from pregnant women. Acta Endocrinologica 1991, 125, 454–458. [CrossRef]
- Kume, H. Role of Airway Smooth Muscle in Inflammation Related to Asthma and COPD. Advances in experimental medicine and biology 2021, 1303, 139–172. [CrossRef]
- Miyata, S. New aspects in fenestrated capillary and tissue dynamics in the sensory circumventricular organs of adult brains. Frontiers in Neuroscience 2015, 9, 390. [CrossRef]
- Zhang, C.; Kaye, J.A.; Cai, Z.; Wang, Y.; Prescott, S.L.; Liberles, S.D. Area Postrema Cell Types that Mediate Nausea-Associated Behaviors. Neuron 2021, 109, 461-472.e5. [CrossRef]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic peptides: current applications and future directions. Sig Transduct Target Ther 2022, 7, 48. [CrossRef]
- Nicosia, A.; Maggio, T.; Costa, S.; Salamone, M.; Tagliavia, M.; Mazzola, S.; Gianguzza, F.; Cuttitta, A. Maintenance of a Protein Structure in the Dynamic Evolution of TIMPs over 600 Million Years. Genome Biol Evol 2016, 8, 1056–1071. [CrossRef]
- Schaller, H.C. A neurohormone from hydra is also present in the rat brain. Journal of Neurochemistry 1975, 25, 187–188. [CrossRef]
- Bodenmüller, H.; Schaller, H.C.; Darai, G. Human hypothalamus and intestine contain a hydra-neuropeptide. Neuroscience letters 1980, 16, 71–74. [CrossRef]
- Roberge, M.; Escher, E.; Schaller, H.C.; Bodenmüller, H. The hydra head activator in human blood circulation. Degradation of the synthetic peptide by plasma angiotensin-converting enzyme. FEBS Letters 1984, 173, 307–313. [CrossRef]
- Schaller, H.C.; Schilling, E.; Theilmann, L.; Bodenmüller, H.; Sachsenheimer, W. Elevated levels of head activator in human brain tumors and in serum of patients with brain and other neurally derived tumors. J Neuro-Oncol 1988, 6, 251–258. [CrossRef]
- Winnikes, M.; Schaller, H.C.; Sachsenheimer, W. Head activator as a potential serum marker for brain tumour analysis. European journal of cancer (Oxford, England : 1990) 1992, 28, 421–424. [CrossRef]
- Lintzel, J.; Franke, I.; Riedel, I.B.; Schaller, H.C.; Hampe, W. Characterization of the VPS10 domain of SorLA/LR11 as binding site for the neuropeptide HA. Biological chemistry 2002, 383, 1727–1733. [CrossRef]
- Böhm, C.; Seibel, N.M.; Henkel, B.; Steiner, H.; Haass, C.; Hampe, W. SorLA signaling by regulated intramembrane proteolysis. Journal of Biological Chemistry 2006, 281, 14547–14553. [CrossRef]
- Gandía, J.; Fernández-Dueñas, V.; Morató, X.; Caltabiano, G.; González-Muñiz, R.; Pardo, L.; Stagljar, I.; Ciruela, F. The Parkinson's disease-associated GPR37 receptor-mediated cytotoxicity is controlled by its intracellular cysteine-rich domain. Journal of Neurochemistry 2013, 125, 362–372. [CrossRef]
- Meyer, R.C.; Giddens, M.M.; Schaefer, S.A.; Hall, R.A. GPR37 and GPR37L1 are receptors for the neuroprotective and glioprotective factors prosaptide and prosaposin. Proc. Natl. Acad. Sci. U. S. A. 2013, 110, 9529–9534. [CrossRef]
- Dunham, J.H.; Meyer, R.C.; Garcia, E.L.; Hall, R.A. GPR37 surface expression enhancement via N-terminal truncation or protein-protein interactions. Biochemistry 2009, 48, 10286–10297. [CrossRef]
- Southern, C.; Cook, J.M.; Neetoo-Isseljee, Z.; Taylor, D.L.; Kettleborough, C.A.; Merritt, A.; Bassoni, D.L.; Raab, W.J.; Quinn, E.; Wehrman, T.S.; et al. Screening β-arrestin recruitment for the identification of natural ligands for orphan G-protein-coupled receptors. Journal of biomolecular screening 2013, 18, 599–609. [CrossRef]
- Smith, N.J. Drug Discovery Opportunities at the Endothelin B Receptor-Related Orphan G Protein-Coupled Receptors, GPR37 and GPR37L1. Front. Pharmacol. 2015, 6, 275. [CrossRef]
- Boels, K.; Glassmeier, G.; Herrmann, D.; Riedel, I.B.; Hampe, W.; Kojima, I.; Schwarz, J.R.; Schaller, H.C. The neuropeptide head activator induces activation and translocation of the growth-factor-regulated Ca(2+)-permeable channel GRC. J Cell Sci 2001, 114, 3599–3606. [CrossRef]
- Hertz, E.; Terenius, L.; Vukojević, V.; Svenningsson, P. GPR37 and GPR37L1 differently interact with dopamine 2 receptors in live cells. Neuropharmacology 2019, 152, 51–57. [CrossRef]
- Kubota, K.; Niinuma, Y.; Kaneko, M.; Okuma, Y.; Sugai, M.; Omura, T.; Uesugi, M.; Uehara, T.; Hosoi, T.; Nomura, Y. Suppressive effects of 4-phenylbutyrate on the aggregation of Pael receptors and endoplasmic reticulum stress. Journal of Neurochemistry 2006, 97, 1259–1268. [CrossRef]
- Xiromerisiou, G.; Bourinaris, T.; Houlden, H.; Lewis, P.A.; Senkevich, K.; Hammer, M.; Federoff, M.; Khan, A.; Spanaki, C.; Hadjigeorgiou, G.M.; et al. SORL1 mutation in a Greek family with Parkinson's disease and dementia. Annals of Clinical and Translational Neurology 2021, 8, 1961–1969. [CrossRef]
- Pietilä, M.; Sahgal, P.; Peuhu, E.; Jäntti, N.Z.; Paatero, I.; Närvä, E.; Al-Akhrass, H.; Lilja, J.; Georgiadou, M.; Andersen, O.M.; et al. SORLA regulates endosomal trafficking and oncogenic fitness of HER2. Nat Commun 2019, 10, 2340. [CrossRef]
- Al-Akhrass, H.; Pietilä, M.; Lilja, J.; Vesilahti, E.-M.; Anttila, J.M.; Haikala, H.M.; Munne, P.M.; Klefström, J.; Peuhu, E.; Ivaska, J. Sortilin-related receptor is a druggable therapeutic target in breast cancer. Molecular oncology 2022, 16, 116–129. [CrossRef]
- Jiang, Z.; Bi, F.; Ge, Z.; Mansolf, M.; Hartwich, T.M.P.; Kolesnyk, V.; Yang, K.; Park, W.; Kim, D.; Grechukhina, O.; et al. SORL1-Mediated EGFR and FGFR4 Regulation Enhances Chemoresistance in Ovarian Cancer. Cancers 2025, 17, 244. [CrossRef]
- Talbot, H.; Saada, S.; Naves, T.; Gallet, P.-F.; Fauchais, A.-L.; Jauberteau, M.-O. Regulatory Roles of Sortilin and SorLA in Immune-Related Processes. Frontiers in pharmacology 2018, 9, 1507. [CrossRef]
- Glerup, S.; Lume, M.; Olsen, D.; Nyengaard, J.R.; Vaegter, C.B.; Gustafsen, C.; Christensen, E.I.; Kjolby, M.; Hay-Schmidt, A.; Bender, D.; et al. SorLA controls neurotrophic activity by sorting of GDNF and its receptors GFRα1 and RET. Cell reports 2013, 3, 186–199. [CrossRef]
- Rovelet-Lecrux, A.; Feuillette, S.; Miguel, L.; Schramm, C.; Pernet, S.; Quenez, O.; Ségalas-Milazzo, I.; Guilhaudis, L.; Rousseau, S.; Riou, G.; et al. Impaired SorLA maturation and trafficking as a new mechanism for SORL1 missense variants in Alzheimer disease. Acta Neuropathologica Communications 2021, 9, 196. [CrossRef]
- Caglayan, S.; Takagi-Niidome, S.; Liao, F.; Carlo, A.-S.; Schmidt, V.; Burgert, T.; Kitago, Y.; Füchtbauer, E.-M.; Füchtbauer, A.; Holtzman, D.M.; et al. Lysosomal sorting of amyloid-β by the SORLA receptor is impaired by a familial Alzheimer's disease mutation. Sci. Transl. Med. 2014, 6, 223ra20. [CrossRef]
- Mehmedbasic, A.; Christensen, S.K.; Nilsson, J.; Rüetschi, U.; Gustafsen, C.; Poulsen, A.S.A.; Rasmussen, R.W.; Fjorback, A.N.; Larson, G.; Andersen, O.M. SorLA complement-type repeat domains protect the amyloid precursor protein against processing. The Journal of biological chemistry 2015, 290, 3359–3376. [CrossRef]
- Shih, A.Z.L.; Chen, Y.-C.; Speckmann, T.; Søndergaard, E.; Schürmann, A.; Verchere, C.B.; Willnow, T.E. SORLA mediates endocytic uptake of proIAPP and protects against islet amyloid deposition. Molecular metabolism 2022, 65, 101585. [CrossRef]
- Bolinger, A.A.; Frazier, A.; La, J.-H.; Allen, J.A.; Zhou, J. Orphan G Protein-Coupled Receptor GPR37 as an Emerging Therapeutic Target. ACS chemical neuroscience 2023, 14, 3318–3334. [CrossRef]
- Wang, X.; Ju, J.; Xie, Y.; Hang, L. Emerging roles of the G-protein-coupled receptor 37 in neurological diseases and pain. Neuroscience 2024, 559, 199–208. [CrossRef]
- Argerich, J.; Garma, L.D.; López-Cano, M.; Álvarez-Montoya, P.; Gómez-Acero, L.; Fernández-Dueñas, V.; Muñoz-Manchado, A.B.; Aso, E.; Boxer, A.; Andres-Benito, P.; et al. GPR37 processing in neurodegeneration: a potential marker for Parkinson's Disease progression rate. npj Parkinsons Dis. 2024, 10, 172. [CrossRef]
- Bian, X.; Wang, Y.; Zhang, W.; Ye, C.; Li, J. GPR37 and its neuroprotective mechanisms: bridging osteocalcin signaling and brain function. Frontiers in cell and developmental biology 2024, 12, 1510666. [CrossRef]
- Yang, H.-J.; Vainshtein, A.; Maik-Rachline, G.; Peles, E. G protein-coupled receptor 37 is a negative regulator of oligodendrocyte differentiation and myelination. Nat Commun 2016, 7, 10884. [CrossRef]
- Smith, B.M.; Giddens, M.M.; Neil, J.; Owino, S.; Nguyen, T.T.; Duong, D.; Li, F.; Hall, R.A. Mice lacking Gpr37 exhibit decreased expression of the myelin-associated glycoprotein MAG and increased susceptibility to demyelination. Neuroscience 2017, 358, 49–57. [CrossRef]
- Ma, Q.; Tian, J.-L.; Lou, Y.; Guo, R.; Ma, X.-R.; Wu, J.-B.; Yang, J.; Tang, B.-J.; Li, S.; Qiu, M.; et al. Oligodendrocytes drive neuroinflammation and neurodegeneration in Parkinson's disease via the prosaposin-GPR37-IL-6 axis. Cell reports 2025, 44, 115266. [CrossRef]
- Ekman, R.; Salford, L.; Brun, A.; Larsson, I. Hydra head activator-like immunoreactivity in human brain astrocytomas grade III-IV and the surrounding brain tissue. Peptides 1990, 11, 271–275. [CrossRef]
Figure 1.
Anatomy of Hydra. The body column of Hydra is composed of a peduncle (foot region), gastric region, and head region with the oral opening called hypostome, surrounded by tentacles. The reproduction of Hydra occurs primarily asexually through budding, resulting in genetically identical clones. The body consists of endoderm (gastroderm), mesoglea, and ectoderm. The main cell types are interstitial, epithelial, and neuronal. A pool of pluripotent interstitial cells gives rise to all cell lineages. The epithelial cells can differentiate into secretory gland cells or into armed nematocytes, while ganglion and sensory neurons form the gastrodermal and ectodermal nerve nets. Adapted from [5].
Figure 1.
Anatomy of Hydra. The body column of Hydra is composed of a peduncle (foot region), gastric region, and head region with the oral opening called hypostome, surrounded by tentacles. The reproduction of Hydra occurs primarily asexually through budding, resulting in genetically identical clones. The body consists of endoderm (gastroderm), mesoglea, and ectoderm. The main cell types are interstitial, epithelial, and neuronal. A pool of pluripotent interstitial cells gives rise to all cell lineages. The epithelial cells can differentiate into secretory gland cells or into armed nematocytes, while ganglion and sensory neurons form the gastrodermal and ectodermal nerve nets. Adapted from [5].
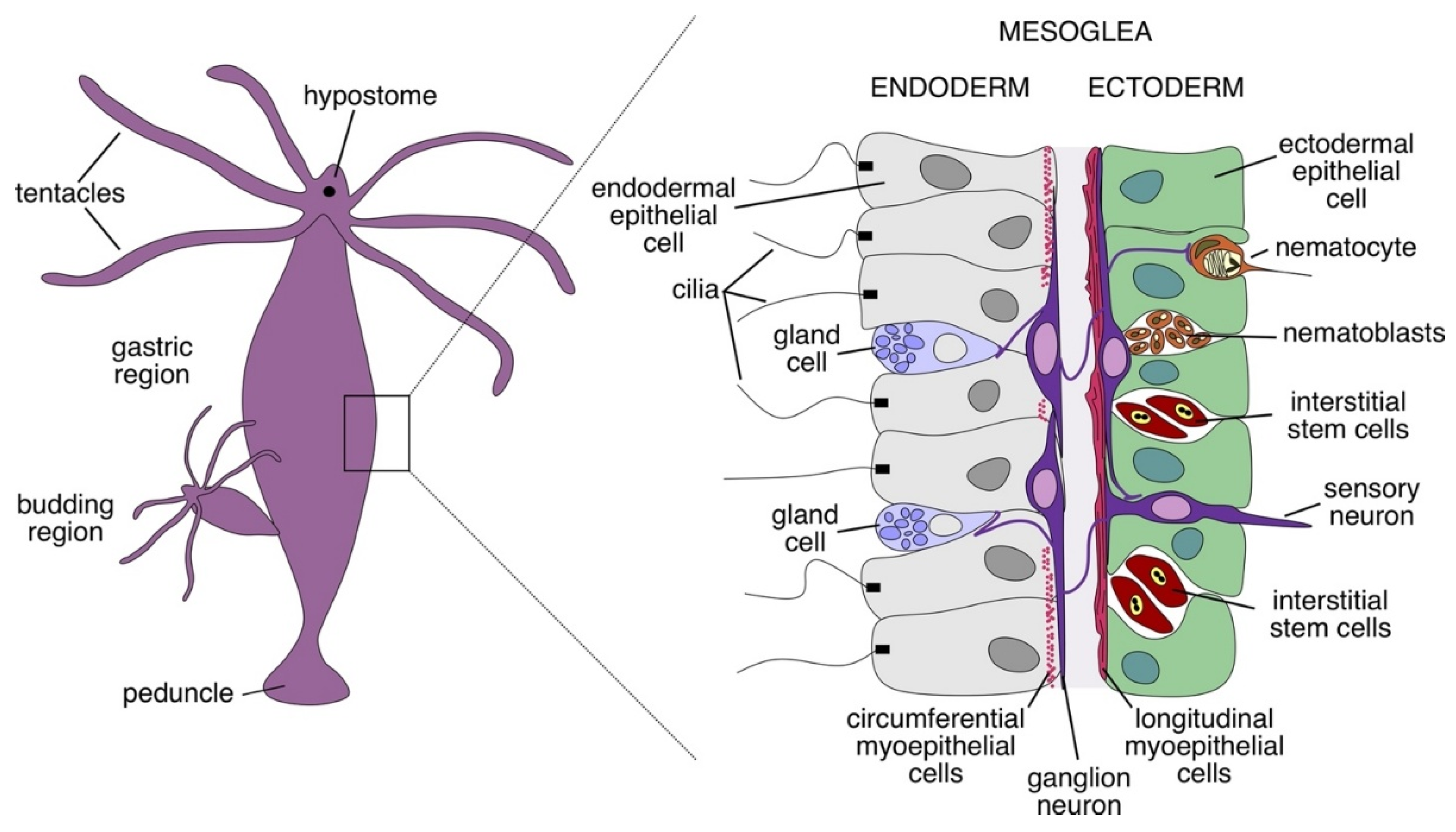
Figure 2.
Regeneration and differentiation cycles in Hydra. Hydra resides in a perpetual state of cell renewal driven by a pool of pluripotent interstitial cells. These cells can give rise to all lineages: epithelial, neuronal and interstitial cells themselves. Proliferating and differentiating cells continuously replenish the endo- and ectodermal layers, allowing regeneration and budding. Adapted from [5].
Figure 2.
Regeneration and differentiation cycles in Hydra. Hydra resides in a perpetual state of cell renewal driven by a pool of pluripotent interstitial cells. These cells can give rise to all lineages: epithelial, neuronal and interstitial cells themselves. Proliferating and differentiating cells continuously replenish the endo- and ectodermal layers, allowing regeneration and budding. Adapted from [5].
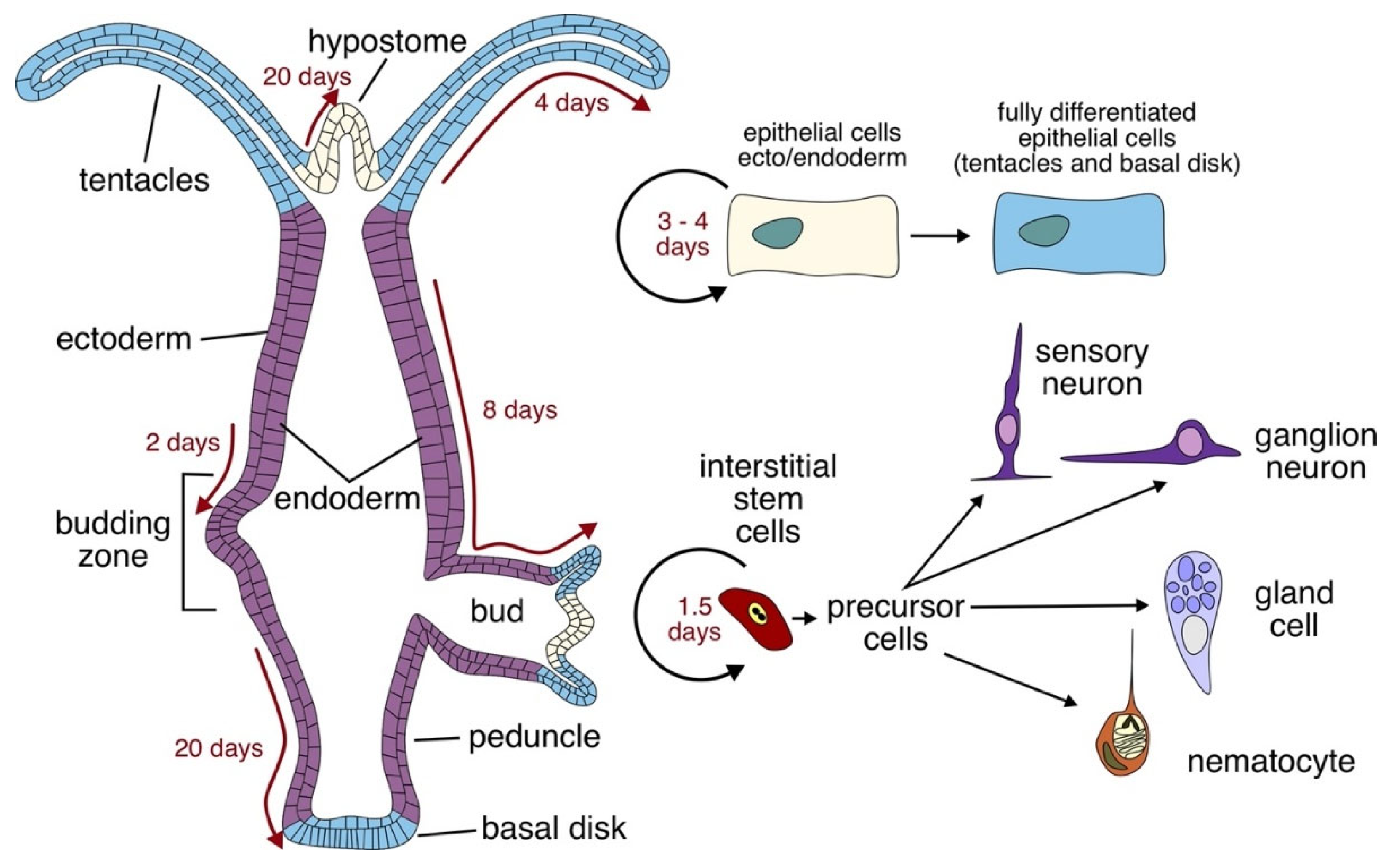
Figure 3.
Determinants of the context-dependent mode of action of the head activator peptide. The effects of the head activator on the cells are temporally dynamic and depend on the differentiation status and mitotic activity of the cells. The main difference between immature and mature cells lies within the expressed head activator receptors. The immature cells express the high affinity receptor, which requires little concentration of the head activator peptide to become activated. It is a Gi-coupled receptor, which inhibits the adenylyl cyclase and depletes the concentration of cAMP – this signaling cue induces proliferation of the cells. In maturating cells that lose the ability to divide, a low-affinity Gs-coupled receptor dominates, which induces cAMP upregulation to assist in the differentiation process.
Figure 3.
Determinants of the context-dependent mode of action of the head activator peptide. The effects of the head activator on the cells are temporally dynamic and depend on the differentiation status and mitotic activity of the cells. The main difference between immature and mature cells lies within the expressed head activator receptors. The immature cells express the high affinity receptor, which requires little concentration of the head activator peptide to become activated. It is a Gi-coupled receptor, which inhibits the adenylyl cyclase and depletes the concentration of cAMP – this signaling cue induces proliferation of the cells. In maturating cells that lose the ability to divide, a low-affinity Gs-coupled receptor dominates, which induces cAMP upregulation to assist in the differentiation process.
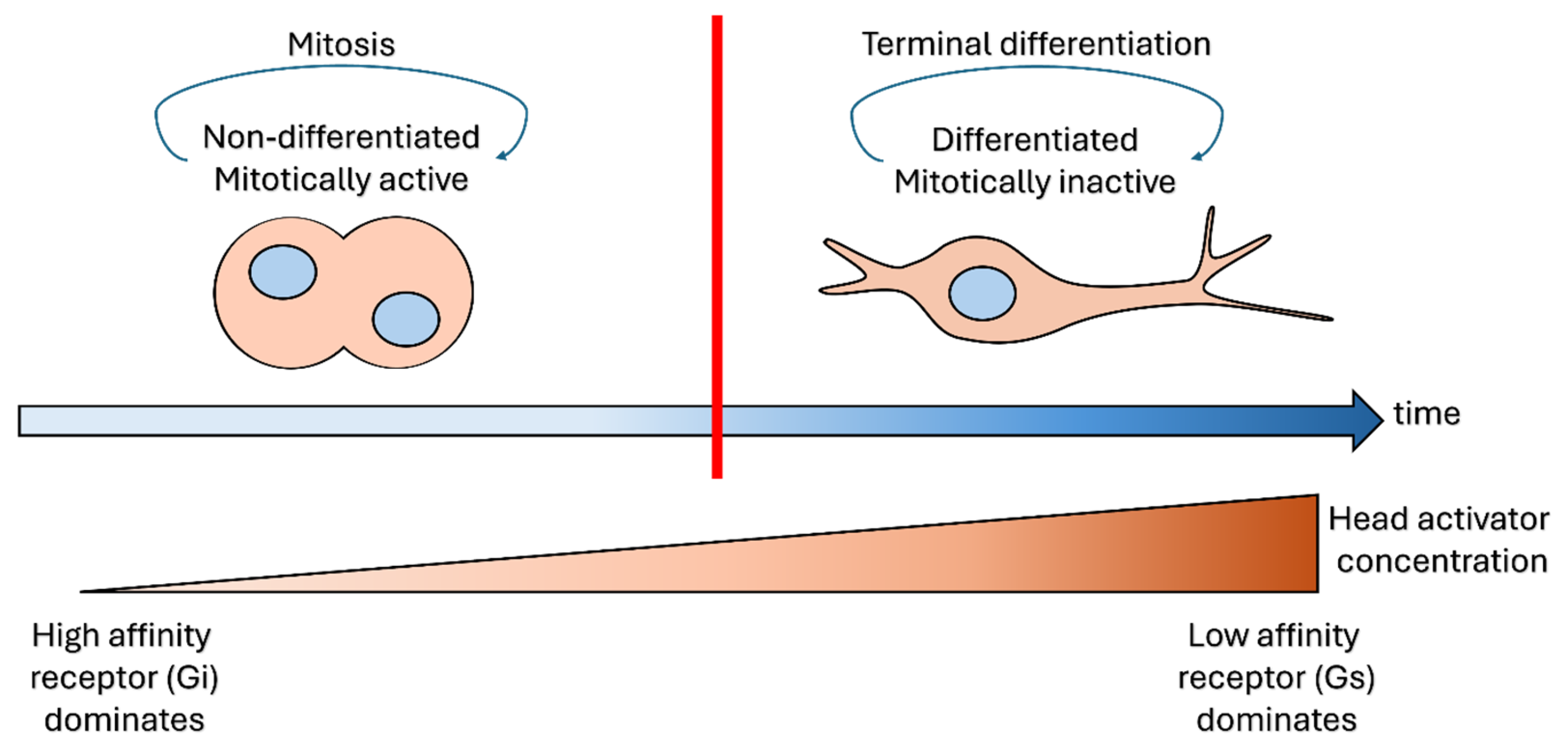
Figure 5.
SorLA and GPR37 in human neuropathology. Two major putative receptors involved in the head activator signaling are the SorLA and GPR37. SorLA is mostly an intracellular protein of the Golgi apparatus, with pleiotropic functions ranging from intracellular transport of neurotrophic factors and cytokines to lipoprotein-receptor like ability to bind and internalise lipoproteins, and amyloid precursor protein processing. This rich functional variability is also mirrored in the pathogenetic involvement of SorLA in multiple disorders. Similarly, GPR37 homeostasis was also shown to participate in pathogenesis of neurological disorders. Its overexpression results in formation of neurotoxic aggregates, observed in Parkinson disease, likewise, GPR37 deficiency is also pathological, and can lead to demyelination.
Figure 5.
SorLA and GPR37 in human neuropathology. Two major putative receptors involved in the head activator signaling are the SorLA and GPR37. SorLA is mostly an intracellular protein of the Golgi apparatus, with pleiotropic functions ranging from intracellular transport of neurotrophic factors and cytokines to lipoprotein-receptor like ability to bind and internalise lipoproteins, and amyloid precursor protein processing. This rich functional variability is also mirrored in the pathogenetic involvement of SorLA in multiple disorders. Similarly, GPR37 homeostasis was also shown to participate in pathogenesis of neurological disorders. Its overexpression results in formation of neurotoxic aggregates, observed in Parkinson disease, likewise, GPR37 deficiency is also pathological, and can lead to demyelination.
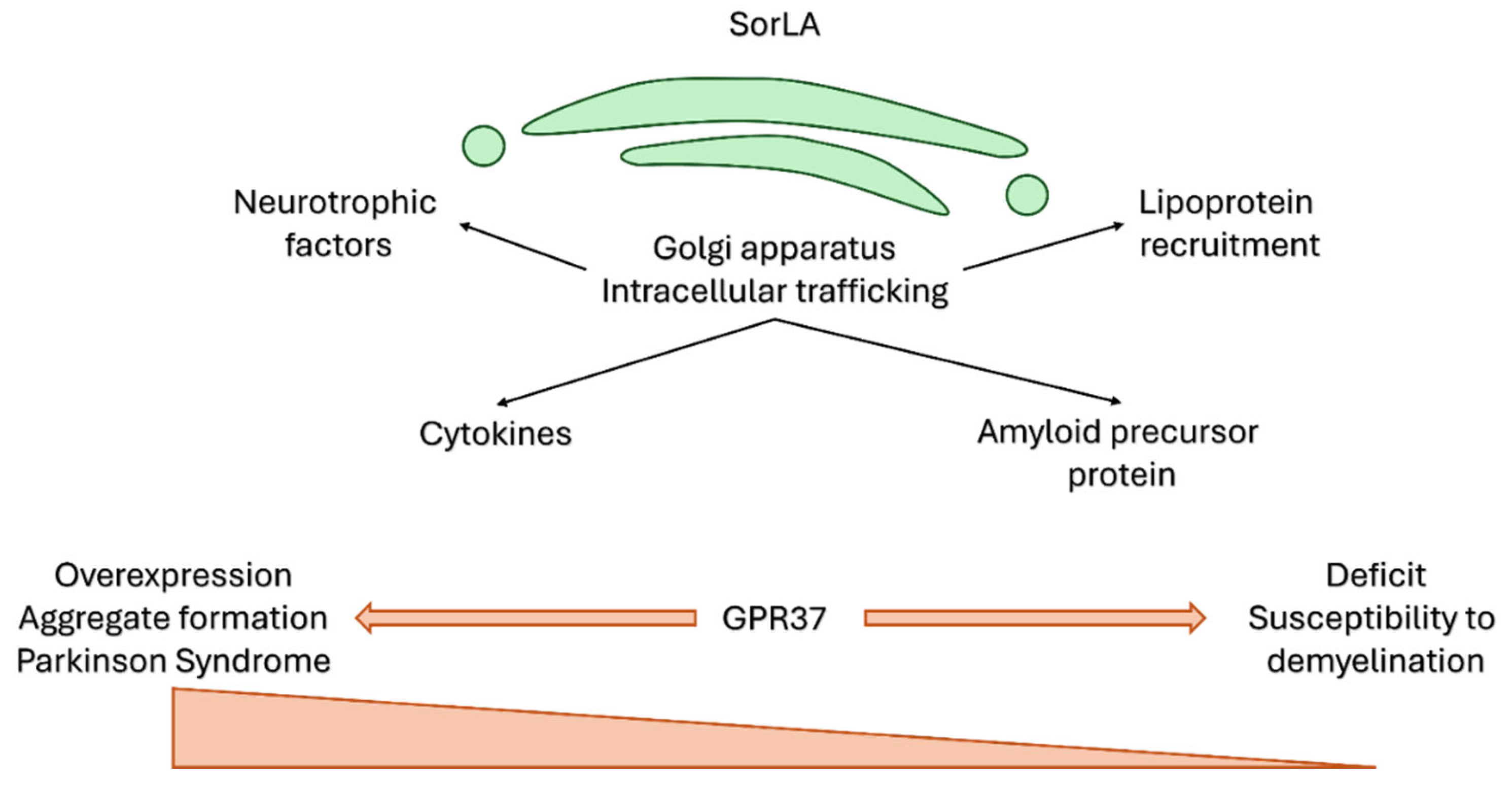

Table 1.
Morphogenic and neurogenic cnidarian peptides.
| peptide | effect | reference |
|---|---|---|
| morphogenesis | ||
| pedibin (Hym-346) | foot formation and regeneration head patterning |
[28] |
| HEADY | head formation budding |
[30] |
| neurogenesis | ||
| Hym-355 | stem cell proliferation stimulation of neuronal differentiation |
[31] |
| Hym-33H | inhibition of neuronal differentiation | [23] |
| Hym-35 | ||
| Hym-37 | ||
| Hym-310 | ||
| morphogenesis and neurogenesis | ||
| head activator | head formation and regeneration stem cell proliferation stimulation of neuronal differentiation |
[32] |
| pedin (Hym-323) | axial body patterning foot formation and regeneration budding stem cell proliferation stimulation of neuronal differentiation |
[28] |
Table 2.
Context-dependent modes of action of the head activator peptide.
| condition | effect | |
|---|---|---|
| cell type | mitotic | mitosis and differentiation |
| post-mitotic | differentiation | |
| time point | early | mitosis |
| late | differentiation | |
| concentration (receptor affinity) | low (high) | mitosis |
| high (low) | differentiation | |
| pathway (AC-cAMP-PKA axis) | AC inhibition | mitosis |
| AC activation | differentiation, neurogenesis | |
Table 3.
Effects of the head activator in mammals.
| tissue/cell type/cell line | effect | reference |
|---|---|---|
| rat corneal epithelium | proliferation ↑ | [69,70] |
| rat lingual epithelium | proliferation ↑ | |
| rat tracheal epithelium | proliferation ↑ | [71,72] |
| rat tracheal smooth muscle cells | proliferation ↓ | |
| lipid peroxidation system in hypoxic rats | inhibition in lungs | |
| stress response in rats | reduction of lipid peroxidation, corticosterone, normalised thyrotropic hormone, T3, T4, and thymus weight | [70] |
| rat myocardium upon a left ventricular hypertrophy | myocardial remodelling | [73,74] |
| rat myocardium | proliferation ↑ | [75] |
| rats exposed to prenatal hypoxia | normalisation of thymocyte proliferation | [76] |
| rats exposed to prenatal hypoxia | normalisation of hepatocyte and tracheal epithelium proliferation, reduction of pulmonary lipid peroxidation |
[71] |
| rat β-endorphine | low dose: ↓ high dose: ↑ |
[77] |
| rat corticosterone | low dose: ↑ high dose: ↓ |
[77] [78,79] |
| rat noradrenaline | low dose: ↑ high dose: ↓ |
|
| rat thyroid hormones | low dose: = high dose: Τ3 ↑ |
|
| rat insulin | low dose: = high dose: ↑ |
|
| liver function in partially hepatectomised rats | ornithine decarboxylase activity in low/medium/high dose: =/↑/↓ protein content: ↑ |
|
| cat model of motion sickness | antiemesis | [80] |
| rat duodenal smooth myocytes | proliferation = | [81] |
| injured rat liver | 4 h after injury: cAMP ↑, cGMP = 24 h after injury: cAMP ↓, cGMP ↑ |
[51] |
| injured rat striated muscle | cAMP =, cGMP = | |
| rat cultured pancreatic cells | amylase secretion ↑ | [82] |
| human erythrocytes | activation of Na/H exchange | [83] |
↑: upregulation, ↓: downregulation, =: unchanged.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



