
Submitted:
06 February 2026
Posted:
09 February 2026
You are already at the latest version
Abstract
miRNA has been studied for over 30 years, but the interaction of miRNA with mRNA has not been sufficiently studied to date. Problems in this area of research include the lack of adequate methods for predicting the characteristics of miRNA-mRNA interactions, and wet lab experiments are many times more labor-intensive and time-consuming. The aim of this study was to determine the characteristics of fully complementary interactions of miRNAs from the 2568 miRNAs of the NCBI database with mRNAs of 17508 protein-coding genes using the MirTarget program. It was found that mRNA from 384 genes have binding sites (BS) with 89 miRNAs. miR-619-5p has 220 BSs in mRNA from 201 genes. mRNA from 17 genes has two BSs of miR-619-5p and mRNA of the CACNG8 gene has three BSs. Three mRNAs contained miR-619-5p BSs in the 5'UTR, only one gene had a CDS, and the remaining BSs were in the 3'UTR. miR-5096 binds to the mRNAs of 45 genes, 12 of which also bound miR-619-5p. IL18 mRNA bound two miR-5096s. MiR-1273f targeted 12 genes, miR-1273g-3p targeted nine genes, and miR-1273h-5p targeted four genes. MiR-1548ap-3p, miR-1285ap-5p, and miR-4478 each had four target genes. Several pairs of miRNA-5p and miRNA-3p were identified that bind to mRNAs of the RTL, CCDC42B, FOXF2, GLYCTK, KAA2026, and LPPR3 genes. The highest free energy of interaction was found for miR-4665-3p (-159 kJ/mole), miR-1-356-5p (-146 kJ/mole), and miR-1-155-3p (-138 kJ/mole). miRNAs from the NCBI database can significantly suppress the translation of many mRNAs.
Keywords:
miRNA
; mRNA
; human genes
; nucleotides
; complementary interactions
1. Introduction
miRNA (mRNA inhibiting RNA) that bind to mRNA of genes and suppress protein synthesis were discovered more than 30 years ago [1]. Nanosized miRNAs, ranging in length from 18 nucleotides (5.8 nanometers) to 25 nucleotides (8.5 nanometers), are involved in the regulation of the synthesis of many proteins and likely regulate the expression of most cellular genes [2,3,4,5,6,7]. Identifying miRNA target genes in wet assays is costly and time-consuming. Many computer programs have been proposed that, unfortunately, predict false-positive miRNA target genes. Therefore, many publications have essentially only established correlations between changes in miRNA concentration and the amount of synthesized protein or disease development. Since many diseases are associated with disrupted expression of protein-coding genes, the involvement of miRNA in these processes requires studying the direct interaction of miRNA with mRNA, preferably with quantitative characteristics of this interaction. The use of inadequate methods for establishing specific miRNA-mRNA relationships involved in these processes has not led to the development of reliable diagnostic and therapeutic methods using miRNA. The goal of this study is to demonstrate that many miRNAs can interact fully complementarily with mRNA and, therefore, can play an important role in regulating human genome expression.
2. Materials and Methods
The nucleotide (nt) sequence of 17,508 genes were downloaded from National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov, 2020). The nucleotide sequences of 2567 miRNAs were taken from NCBI (http://mirbase.org). The miRNA binding sites (BSs) in mRNA were predicted using the MirTarget program [8]. This program predicts the following features of miRNA binding to mRNA: (a) the initiation of miRNA binding to the mRNA from the first nucleotide of the mRNA; (b) the localization of the miRNA BSs in the 5′-untranslated region (5′UTR), coding domain sequence (CDS), and 3′-untranslated region (3′UTR) of the mRNAs; (c) the schemes of nucleotide interactions between miRNA with mRNA, which clearly demonstrate the interactions of all nucleotides between miRNA with mRNA; (d) the free energy Gibbs (ΔG, kJ/mol) of the interaction between miRNA and the mRNA; and the ratio ΔG/ΔGm (%) is determined for each site. ΔGm equals the free energy of miRNA binding with its fully complementary canonical nucleotide sequence. Only miRNAs whose nucleotides interacted with mRNA using canonical (G-C and A-U) nucleotides with a given ΔG value were selected from the calculated data. The MirTarget program finds hydrogen bonds between miRNAs with mRNA according to the physicochemical characteristics of nucleotide interactions [9,10,11,12] MirTarget differs from other programs in terms of finding miRNA BSs on mRNA in the following: it takes into account the interaction of miRNA with mRNA over the entire miRNA nucleotides sequence; and it calculates the free energy of the interaction of the miRNA with mRNA. Note that the G, A, C, and U nucleotides, which comprise the RNA structure of microorganisms, plants, and animals, interact identically under equal conditions. Therefore, the physicochemical properties of canonical nucleotide pairs given above do not require additional proof of the previously established physicochemical characteristics of their interaction. The reliability of translation suppression by miRNAs that are fully complementary to mRNAs was proven by A.Z. Fire et al. [13]. A.Z. Fire and C.C. Mello were awarded the Nobel Prize in 2006 for this research.
3. Results
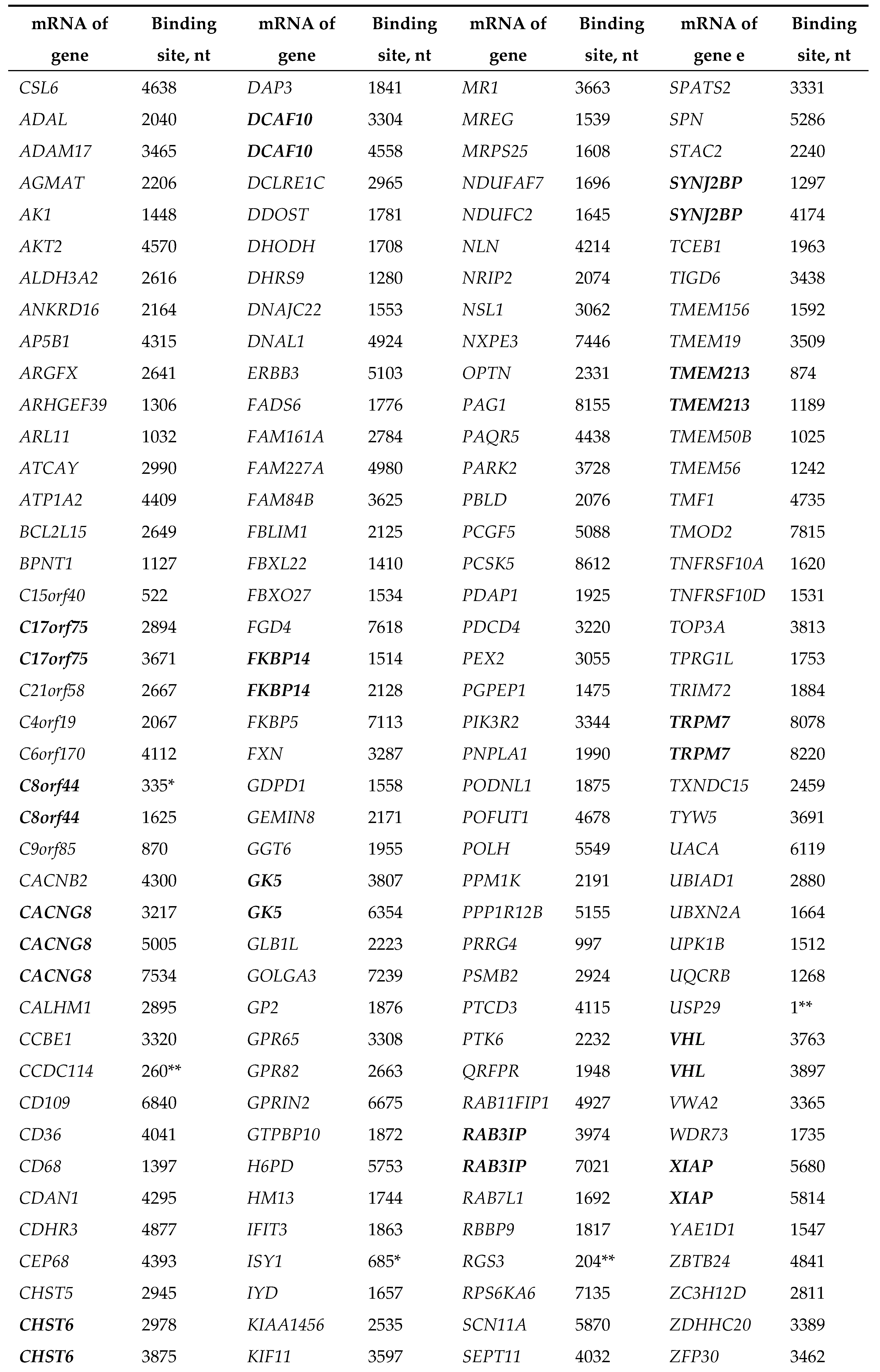
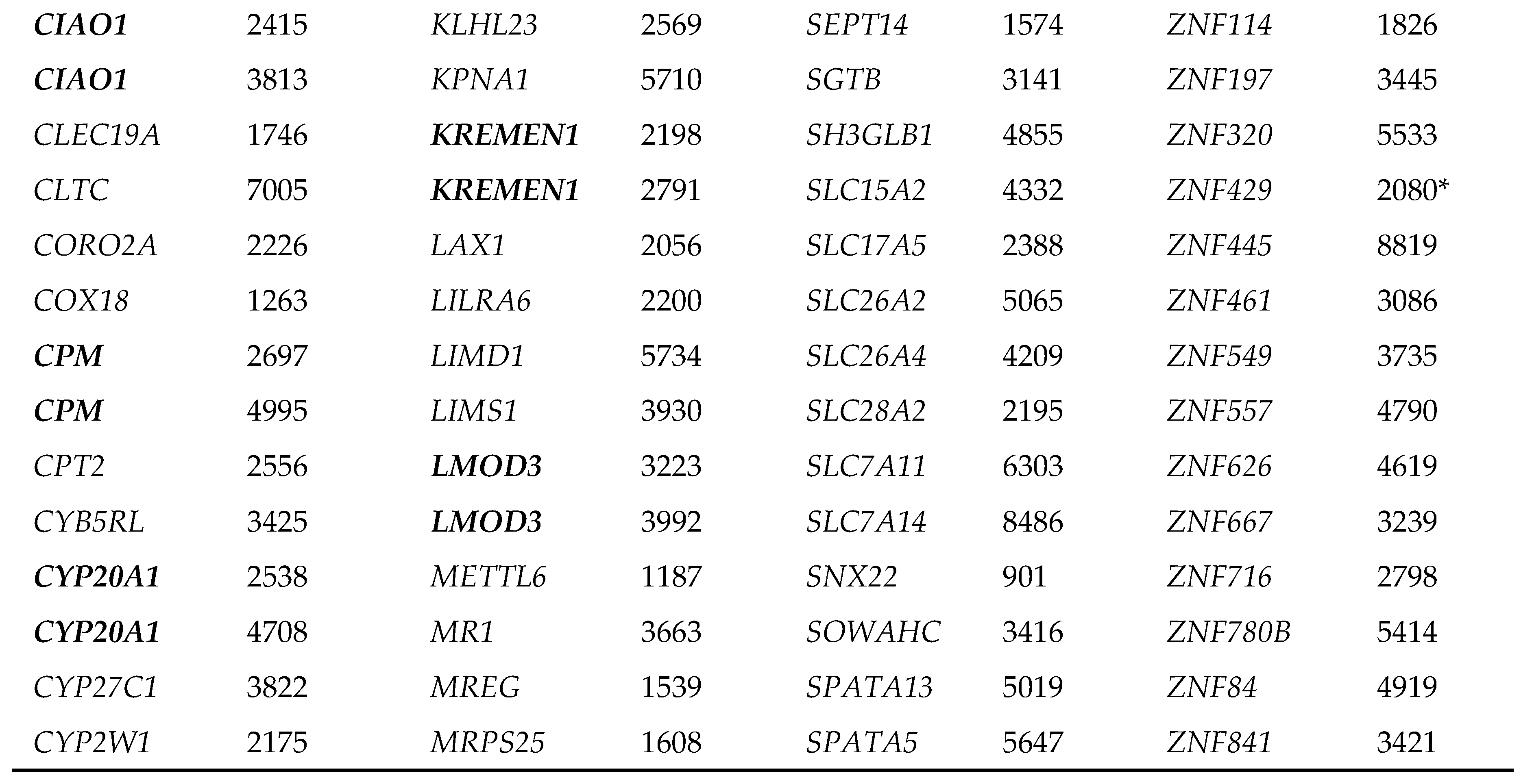
In this study, we identified miRNAs that bind fully complementarily to mRNAs from 17,508 genes in the NCBI database. The largest number of target genes was identified for miR-619-5p, which had 220 binding sites in 201 genes (Table 1). The mRNA of the CACNG8 gene had three miR-619-5p binding sites located in the 3'UTR. The CACNG8 gene is weakly expressed in the stomach, heart, and kidney, and therefore is under strong control of miR-619-5p. Seventeen genes had two miR-619-5p binding sites each: C17orf75, C8orf44, CHST6, CIAO1, CPM, CYP20A1, DCAF10, FKBP14, GK5, KREMEN1, LMOD3, RAB3IP, SYNJ2BP, TMEM213, TRPM7, VHL, XIAP, and ZNF667. miR-619-5p has an intronic origin from the pre-miRNA of the SSH1 gene, which is expressed in many tissues, and therefore miR-619-5p may also be present in many tissues and influence the expression of its target genes in them. Many of these target genes are expressed to a higher degree than the SSH1 gene, and therefore miR-619-5p cannot exert significant control over the synthesis of the corresponding proteins. However, due to the general dependence on miR-619-5p, a balance is established between the protein synthesis of the target genes of this miRNA. A large number of miR-619-5p target genes have been identified for a long time [14], but this miRNA has not attracted much interest from researchers. Clearly, controlling the influence of miR-619-5p on such a large number of genes is very difficult, but it is necessary, since this unique phenomenon is highly likely not random and can explain many physiological properties of humans, given the wide range of functions of the target genes.
MiR-5096 has 45 target genes, of which only the mRNA of the IL18 gene had two BSs (Table 2). All miR-5096 BSs were localized in the 3'UTR. The expression of the 45 miR-5096 target genes may likely be linked via this miRNA, since increased expression of one gene will lead to increased binding of miR-5096 to its mRNA, and the expression of other target genes will increase. Twelve miR-5096 target genes have BSs for miR-619-5p, which increases the control of these genes' expression by miRNA. Of these, in the mRNAs of the C15orf40, LMOD3, PNPLA1, and ZNF197 genes, the beginning of the miR-5096 BS is located 74 nt before the beginning of the miR-619-5p BS. This arrangement of BS miR-5096 and miR-619-5p is not random and reflects the relationship between these miRNAs and their target genes.
Expression of several genes depends on pairs of miRNAs (miRNA-5p and miRNA-3p) that originate from the same pre-miRNA hairpin. We have previously identified quantitative characteristics of their interactions with target genes [15]. This surprising property is possessed by mRNA of the RTL1 gene, containing five pairs of binding sites for miR-127-3p and miR-127-5p, miR-136-3p and miR-136-5p, miR-431-3p and miR-431-5p, miR-432-3p and miR-432-5p, miR-433-3p and miR-433-5p. Schemes of interaction of these miRNAs with quantitative characteristics are shown in Figure 1. All binding sites for these miRNAs were located within the CDS of the RTL1 gene mRNA at intervals of 34 nt in the miR-127-3p and miR-127-5p pair, 33 nt in the miR-136-3p and miR-136-5p pair, 44 nt in the miR-431-3p and miR-431-5p pair, 46 nt in the miR-432-3p and miR-432-5p pair, and 52 nt in the miR-433-3p and miR-433-5p pair. Importantly, the miRNA nucleotide sequences were identical in animals that diverged millions of years ago.
Each of the mRNAs from the genes CCDC42B, FOXF2, GLYCTK, KAA2026, and LPPR3 bound a pair of miRNA-5p and miRNA-3p (Figure 2). The data show that the binding sites for these miRNA pairs were localized in the CDS, 3’UTR, and 5’UTR. For these genes, experiments showed that the presented results of computer analysis were fully confirmed. Our results add to them the free energy of interaction between miRNA and mRNA. The beginnings of miRNA binding sites in the pairs miR-7106-5p and miR-7106-3p, miR-6720-5p and miR-6720-3p, miR-135a-5p and miR-135a-3p, miR-4665-5p and miR-4665-3p, miR-3187-5p and miR-3187-3p were located at 40 nt, 35 nt, 38 nt, 39 nt, and 34 nt, respectively.
miRNA-1-155-3p has ten target genes (Table 3). The mRNAs of six genes have binding sites in the 5'UTR and four in the CDS, which is caused by the increased GC content of miRNA-1-155-3p and the mRNAs of the target genes. Naturally, this is reflected in the free energy of interaction of –138 kJ/mole. miRNA-1-356-5p has three BSs in the 5'UTR of the mRNAs of two genes, including the BMP8B gene with two BSs with a binding onset interval of one nucleotide. This mRNA-miRNA pair has an even higher free energy of interaction of –146 kJ/mole. It is worth noting the effect of miR-762 on the BCL7C and TBCID10B genes with an interaction free energy of –136 kJ/mole. This interaction also ensures strong suppression of the synthesis of the corresponding proteins.
The miR-548 family has binding sites in the mRNA of five genes (Table 4). The mRNAs of the ALG10 and SMU1 genes each had one miR-548 binding site. The mRNAs of the ANKRD5 and TCEANC2 genes each had three miR-548 binding sites, and the mRNA of the KATNAL1 gene had six miR-548 binding sites. All binding sites for these miRNAs are located in the 3'UTR. The free energy of interaction of these miRNAs with mRNA varied from -89 kJ/mole to -123 kJ/mole, which depended on the GC content and the length of the miRNA. If the miRNA binding sites in mRNA were located with overlapping nucleotides, we called such mRNA regions binding site clusters. In such clusters, competition between miRNAs for binding to mRNA occurs. Three miRNAs from the miR-548 family bound in a cluster at 3534 nt, and three miRNAs at 3495 nt. Of note is the effect of miR-762 on the BCL7C and TBCID10B genes, with a free energy of interaction of -136 kJ/mole. This interaction ensures strong suppression of the synthesis of the corresponding proteins.
Members of the miR-1273 family play a significant role in the regulation of gene expression (Table 5). miR-1273f affects 12 target genes, each of whose mRNAs contains one BS of this miRNA. In all but one of these mRNAs, the BSs are located in the 3'UTR. miR-1273f consists of 19 nucleotides, and the free energy of interaction with the mRNA of the target genes is -104 kJ/mole (Table 5). BSs of miR-1273g-3p are located in the 3'UTR of the mRNA of nine genes. mRNA of four genes is the target of miR-1273h-5p (Table 5). The free energy of interaction of miR-1273g-3p and miR-1273h-5p with the mRNA of the target genes is -117 kJ/mole at a length of 21 nucleotides. In total, the miR-1273 family targets 25 different genes, making it difficult to elucidate the biological role of these miRNAs.
miR-1184 bound to mRNA of three different genes, and miR-1285-5p bound to mRNA of four genes (Table 6). The family of five miR-1260s bound to mRNA of five genes in the CDS and 3'UTR. miRNA-3178 and other miRNAs had binding sites in individual genes. Table 6 highlights miRNA-4478, which has four target genes, and miRNA-4508 and miRNA-5095, which each have three target genes. The total number of miRNAs identified in this study between 18 and 24 nucleotides in length was 310, of which 21 and 22 nucleotides miRNAs accounted for 82%, ensuring highly selective miRNA and mRNA interactions.
For identical miRNA lengths, the free energy of interaction (ΔG) between miRNA and mRNA depends on the GC content, and for identical miRNA GC content, the free energy of interaction depends on the miRNA length.
4. Discussion
A previous publication demonstrated that 201 target genes with miR-619-5p binding sites are involved in numerous metabolic reactions, including those associated with various diseases [14]. Therefore, the proposed biological role of miR-619-5p and other miRNAs with multiple target genes is to maintain a balance between the expression of the target genes in this association. Increased mRNA synthesis for one gene will result in preferential miRNA binding, and other genes will show a slight increase in expression while the amount of this miRNA remains constant. We found that not only one miRNA can bind to an mRNA, but other miRNAs can bind to the mRNA in a fully complementary manner. It has been established that multiple miRNAs can bind to a single region (cluster of binding sites) of mRNA, competing with each other, indicating robust control of gene expression. In addition to the interactions of canonical nucleotide pairs A-U and G-C, non-canonical nucleotide pairs A-C and G-U can be formed. It has been shown that mRNAs of different genes contain identical binding clusters for identical miRNAs. The use of a specific miRNA for diagnosis or therapy necessarily requires identifying the effect of this miRNA on other genes to avoid side effects of the miRNA used. A miRNA can suppress oncogenesis if it acts on an oncogene and can stimulate oncogenesis if the miRNA acts on a tumor suppressor. The provided list of properties of miRNA-mRNA interactions demonstrates the need to consider the above-mentioned features of miRNA-mRNA interaction. Since many diseases are associated with impaired expression of protein-coding genes, the involvement of miRNA in these processes requires confirmation of the interaction of miRNA with mRNA, preferably with specific characteristics of this interaction. The use of inadequate methods for establishing specific miRNA and mRNA relationships involved in these processes has not led to the development of reliable diagnostic and therapeutic methods using miRNA. The present studies have shown that many miRNAs can interact fully complementarily with mRNA and, therefore, can play an important role in the regulation of human genome expression.
Author Contributions
Conceptualization, A.I. and A.P.; methodology, R.N.; software, S.O.; validation, A.I., R.N. and A.P.; formal analysis, A.P; investigation, S.O.; resources, R.N.; data curation, A.I.; writing—original draft preparation, R.N.; writing—review and editing, A.I.; visualization, S.O.; supervision, A.P.; project administration, A.P.; funding acquisition, A.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Committee of Science of the Ministry of Science and Higher Education of the Republic of Kazakhstan, grant number AP26102055 “Design of a System for Monitoring and Analyzing Health Indicators of Patients with Metabolic Syndrome.
Data Availability Statement
The nucleotide sequences of 17,508 genes were downloaded from National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov, 2020). The nucleotide sequences of 2567 miRNAs were taken from NCBI (http://mirbase.org).
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Lее, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Mehanna, E.T; Ghattas, M.H.; Mesbah, N.M.; Saleh, S.M.; Abo-Elmatty, D.M. Association of MicroRNA-146a rs2910164 Gene Polymorphism with Metabolic Syndrome. Folia Biol. 2015, 61(1), 43–48. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yan, Y.; Xv, W.; Qian, G.; Li, C.; Zou, H.; Li, Y. A New Insight into the Roles of MiRNAs in Metabolic Syndrome. Biomed Res Int. 2018, 2018, 7372636. [Google Scholar] [CrossRef] [PubMed]
- Vorobeva, E.V.; Kyyaly, M.A.; Sones, C.L.; He, P.J.W.; Arshad, S.H.; Sanchez-Elsner, T.; Kurukulaaratchy, R.J. Circulating microRNAs as Potential Diagnostic Tools for Asthma and for Indicating Severe Asthma Risk. Int. J. Mol. Sci. 2025, 26(14), 6676. [Google Scholar] [CrossRef] [PubMed]
- Stevanovic, J.; Mitic, N.; Penezic, A.; Radojicic, O.; Ardalic, D.; Mandic, M.; Mandic-Markovic, V.; Mikovic, Ž.; Brkušanin, M.; Nedic, O.; et al. Upregulation of the Antioxidant Response-Related microRNAs miR-146a-5p and miR-21-5p in Gestational Diabetes: An Analysis of Matched Samples of Extracellular Vesicles and PBMCs. Int. J. Mol. Sci. 2025, 26(14), 6902. [Google Scholar] [CrossRef] [PubMed]
- Khalyfa, A.; Verma, M.; Alexander, M.M.; Qiao, Z.; Rood, T.; Kapoor, R.; Joshi, T.; Gozal, D.; Francisco, B.D. Childhood Asthma Biomarkers Derived from Plasma and Saliva Exosomal miRNAs. Int. J. Mol. Sci. 2025, 26(15), 7043. [Google Scholar] [CrossRef] [PubMed]
- Dobre, M.; Manuc, T.E.; Manuc, M.; Matei, I.-C.; Dobre, A.-M.; Dragne, A.-D.; Maffioletti, E.; Pelisenco, I.A.; Milanesi, E. Circulating miRNA Profile in Inflammatory Bowel Disease Patients with Stress, Anxiety, and Depression. Int. J. Mol. Sci. 2025, 26(15), 7321. [Google Scholar] [CrossRef] [PubMed]
- Ivashchenko, A.; Berillo, O.; Pyrkova, A.; Niyazova, R.; Atambayeva, S. MiR-3960 binding sites with mRNA of human genes. Bioinformation 2014, 10(7), 423–427. [Google Scholar] [CrossRef] [PubMed]
- Kool, E.T. Hydrogen bonding, base stacking, and steric effects in DNA replication. Annu. Rev. Biophys. Biomol. Struct. 2001, 30, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Leontis, N.B.; Stombaugh, J.; Westhof, E. The Non-watson-crick Base Pairs and their associated isostericity matrices. Nucleic Acids Res. 2002, 30(16), 3497–3531. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.; Caiment, F.; Tordoir, X.; Cavaillé, J.; Ferguson-Smith, A.; Cockett, N.; Georges, M.; Charlier, C. RNAi-mediated allelic trans-interaction at the imprinted Rtl1/ Peg11 locus. Curr. Biol. 2005, 15(8), 743–749. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Heinemann, U.A. Novel form of RNA double helix based on G·U and C·A+ wobble base pairing. RNA 2018, 24(2), 209–218. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S. A.; Driver, S. E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391(6669), 806–811. [Google Scholar] [CrossRef] [PubMed]
- Atambayeva, S.; Niyazova, R.; Ivashchenko, A.; Pyrkova, A.; Pinsky, I.; Akimniyazova, A.; Labeit, S. The binding sites of miR-619-5p in the mRNAs of human and orthologous genes. BMC Genomics 2017, 18, 428. [Google Scholar] [CrossRef] [PubMed]
- Yurikova, O.Yu.; Aisina, D.E.; Niyazova, R.E.; Atambayeva, S.A.; Labeit, S.; Ivashchenko, A.T. The interaction of miR-5p and miR-3p with the mRNAs of orthologous genes. Mol. Biol. (Mosk) 2019, 53(4), 692–704. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schemes of interaction of miRNA nucleotides with mRNA of the RTL1 gene. The mRNA nucleotides are shown in red, and the miRNA nucleotides are shown in purple.
Figure 1.
Schemes of interaction of miRNA nucleotides with mRNA of the RTL1 gene. The mRNA nucleotides are shown in red, and the miRNA nucleotides are shown in purple.
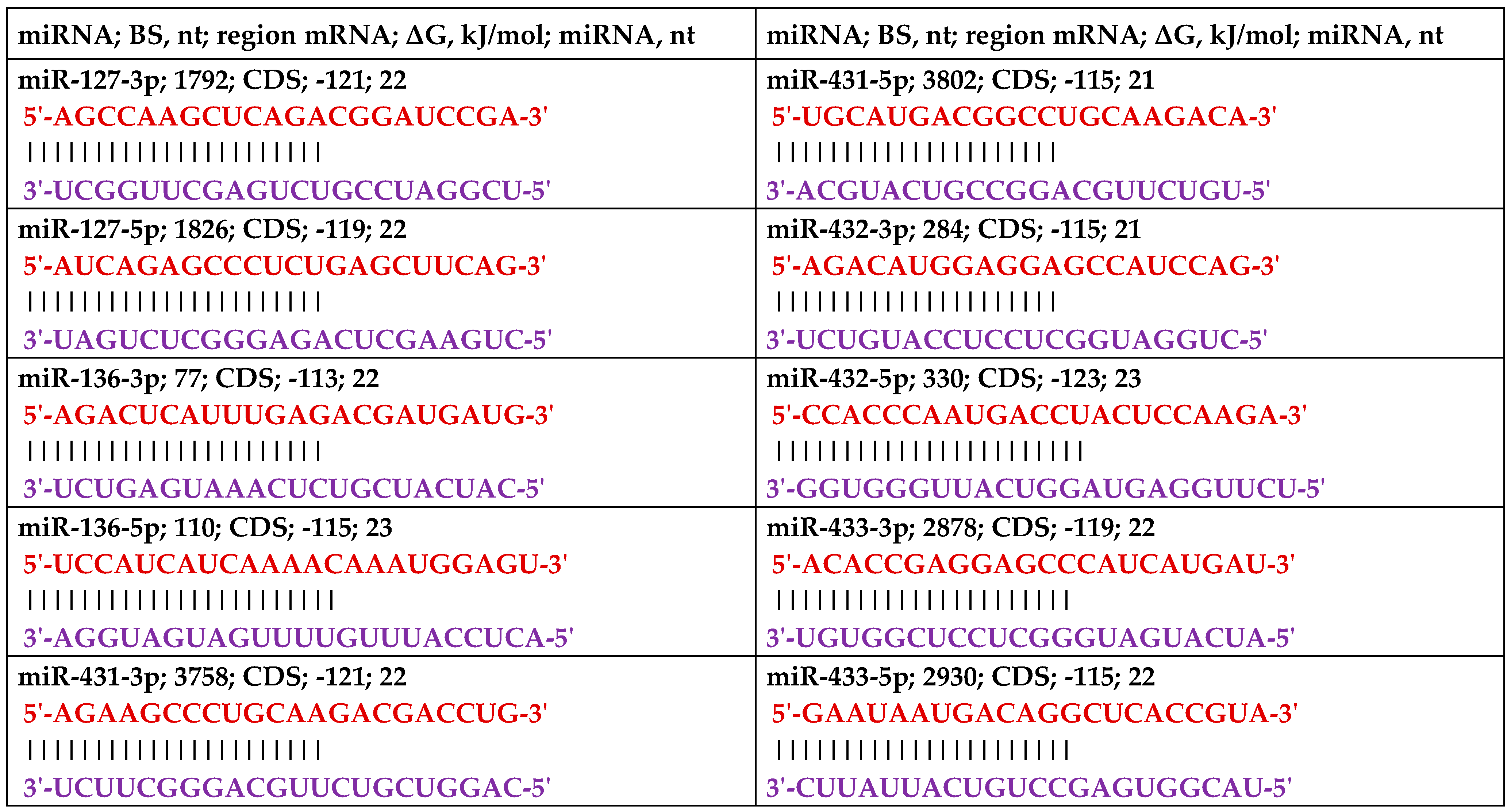
Figure 2.
Schemes of interaction of miRNA nucleotides with mRNA of the CCDC42B, FOXF2, GLYCTK, KAA2026 and LPPR3 genes. The mRNA nucleotides are shown in red, and the miRNA nucleotides are shown in purple.
Figure 2.
Schemes of interaction of miRNA nucleotides with mRNA of the CCDC42B, FOXF2, GLYCTK, KAA2026 and LPPR3 genes. The mRNA nucleotides are shown in red, and the miRNA nucleotides are shown in purple.
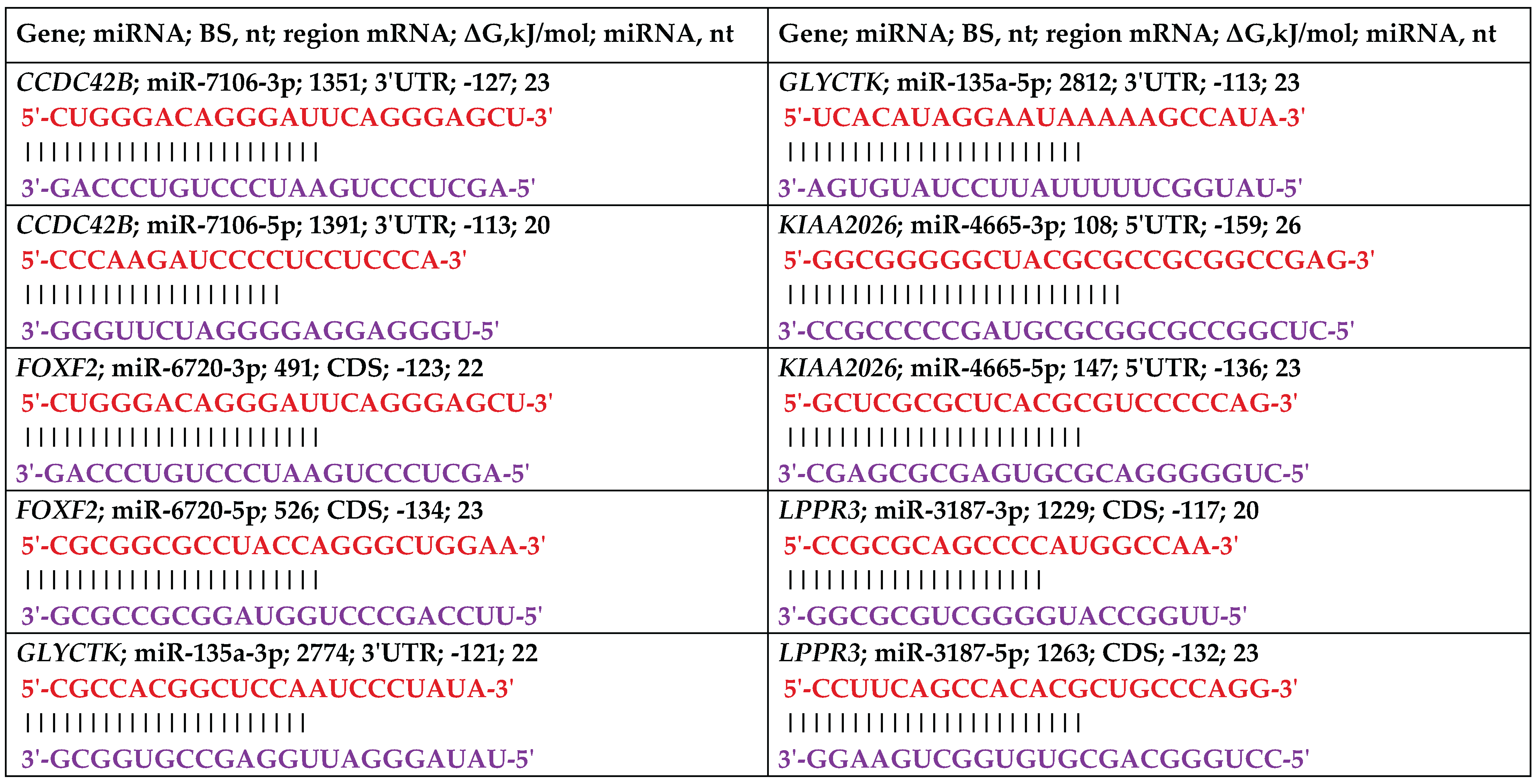

Table 1.
Binding sites miR-619-5p in mRNA target genes.
|
Genes with two miR-619-5p binding sites are shown in bold.
Table 2.
Binding sites miR-5096 in mRNA target genes.
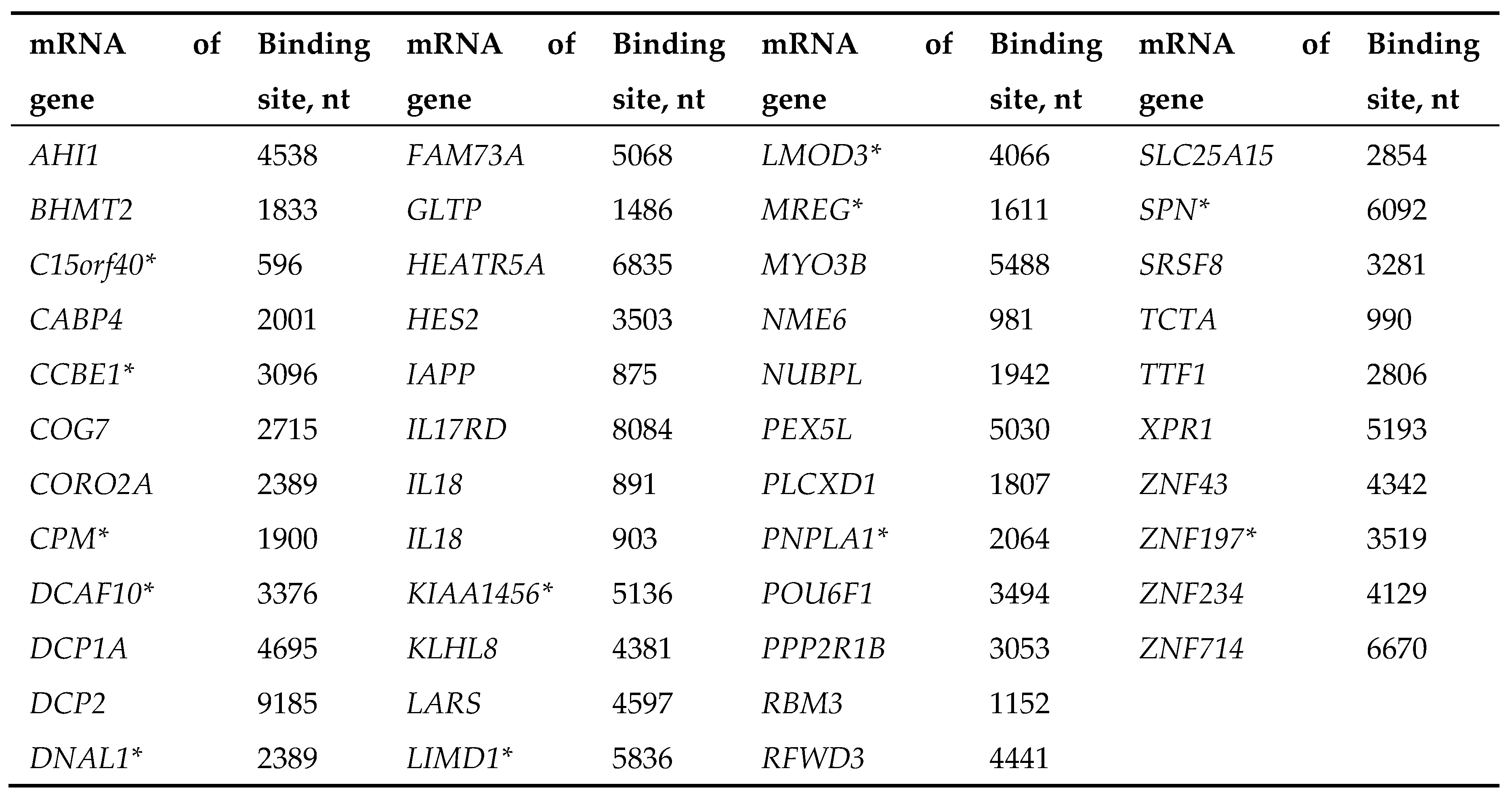 |
* - target gene of miR-619-5p.
Table 3.
Characteristics of the interaction of mRNA of target genes with miRNAs of the miR-1-155-3p and miR-1-356-5p families.
Table 3.
Characteristics of the interaction of mRNA of target genes with miRNAs of the miR-1-155-3p and miR-1-356-5p families.
| mRNA of gene | miRNA | Binding site, nt | ΔG,kJ/mole | Region of mRNA |
| ASXL1-v-1 | miR-1-155-3p | 340 | -138 | 5'UTR |
| CPT1A-v-2 | miR-1-155-3p | 97 | -138 | 5'UTR |
| CXXC4-v-1 | miR-1-155-3p | 742 | -138 | CDS |
| DISP2-v-1 | miR-1-155-3p | 20 | -138 | 5'UTR |
| DNAJC21- | miR-1-155-3p | 62 | -138 | 5'UTR |
| ESPN-v-1 | miR-1-155-3p | 1966 | -138 | CDS |
| FAM131C- | miR-1-155-3p | 1 | -138 | 5'UTR |
| 1HTT-v-1 | miR-1-155-3p | 263 | -138 | CDS |
| >IRX4-v-1 | miR-1-155-3p | 1243 | -138 | CDS |
| USP25-v-1 | miR-1-155-3p | 183 | -138 | 5'UTR |
| ACTN2 | miR-1-356-5p | 99 | -146 | 5'UTR |
| BMP8B | miR-1-356-5p | 250 | -146 | 5'UTR |
| BMP8B | miR-1-356-5p | 251 | -146 | 5'UTR |
Table 4.
Characteristics of the interaction of mRNA of target genes with miRNAs of the miR-548 family.
Table 4.
Characteristics of the interaction of mRNA of target genes with miRNAs of the miR-548 family.
| mRNA of gene | miRNA | Binding site, nt | ΔG,kJ/mole | Region of mRNA |
| KATNAL1 | miR-548aa | 3945 | -123 | 3'UTR |
| TCEANC2 | miR-548aa | 2658 | -123 | 3'UTR |
| KATNAL1 | miR-548am-5p | 3534 | -110 | 3'UTR |
| LG10 | miR-548ap-3p | 2116 | -89 | 3'UTR |
| ANKRD5 | miR-548ap-3p | 3406 | -89 | 3'UTR |
| KATNAL1 | miR-548ap-3p | 3501 | -89 | 3'UTR |
| TCEANC2 | miR-548ap-3p | 2664 | -89 | 3'UTR |
| ANKRD5 | miR-548au-5p | 3444 | -104 | 3'UTR |
| KATNAL1 | miR-548au-5p | 3535 | -104 | 3'UTR |
| KATNAL1 | miR-548c-5p | 3534 | -110 | 3'UTR |
| SMU1 | miR-548h-3p | 6756 | -115 | 3'UTR |
| ANKRD5 | miR-548t-3p | 3400 | -123 | 3'UTR |
| KATNAL1 | miR-548t-3p | 3495 | -123 | 3'UTR |
| TCEANC2 | miR-548t-3p | 2658 | -123 | 3'UTR |
Table 5.
Characteristics of the interaction of miR-1273 families with mRNAs of target genes.
| mRNA of gene | miRNA | Binding sate, nt | ΔG,kJ/mole | Region of mRNA |
| ACACB | miR-1273f | 7996 | -104 | 3'UTR |
| ATF7IP2 | miR-1273f | 2817 | -104 | 3'UTR |
| CA13 | miR-1273f | 2779 | -104 | 3'UTR |
| COX18 | miR-1273f | 1851 | -104 | 3'UTR |
| GTPBP3 | miR-1273f | 2143 | -104 | 3'UTR |
| KRAS | miR-1273f | 3208 | -104 | 3'UTR |
| LOC100996605 | miR-1273f | 238 | -104 | CDS |
| MAP3K13 | miR-1273f | 4636 | -104 | 3'UTR |
| RNF125 | miR-1273f | 2810 | -104 | 3'UTR |
| SLA2 | miR-1273f | 1910 | -104 | 3'UTR |
| SPTSSA | miR-1273f | 1564 | -104 | 3'UTR |
| XPNPEP3 | miR-1273f | 5392 | -104 | 3'UTR |
| AP3S2 | miR-1273g-3p | 4111 | -117 | 3'UTR |
| C15orf38 | miR-1273g-3p | 4433 | -117 | 3'UTR |
| C20orf203 | miR-1273g-3p | 1938 | -117 | 3'UTR |
| CRLF3 | miR-1273g-3p | 2030 | -117 | 3'UTR |
| EIF1AD | miR-1273g-3p | 2165 | -117 | 3'UTR |
| EVI5 | miR-1273g-3p | 4673 | -117 | 3'UTR |
| GJC1 | miR-1273g-3p | 4830 | -117 | 3'UTR |
| TRAF6 | miR-1273g-3p | 4719 | -117 | 3'UTR |
| ZNF621 | miR-1273g-3p | 4815 | -117 | 3'UTR |
| CASP10 | miR-1273h-5p | 2622 | -117 | 3'UTR |
| GINS3 | miR-1273h-5p | 442 | -117 | CDS |
| MXRA7 | miR-1273h-5p | 1379 | -117 | 3'UTR |
| ZNF445 | miR-1273h-5p | 5930 | -117 | 3'UTR |
Table 6.
Characteristics of the interaction of several mRNA target genes with one miRNA.
| mRNA of gene | miRNA | Binding sate, nt | ΔG,kJ/mole | Region of mRNA |
| FBA1 | miR-1184 | 1016 | -127 | CDS |
| FBA2 | miR-1184 | 1001 | -127 | CDS |
| FBA3 | miR-1184 | 1001 | -127 | CDS |
| ERP29 | miR-1260a | 377 | -100 | CDS |
| NGB | miR-1260a | 1118 | -100 | 5'UTR |
| MAML2 | miR-1260a | 1715 | -104 | CDS |
| SC5DL | miR-1260a | 4212 | -108 | 3'UTR |
| MAML2 | miR-1260b | 1715 | -104 | CDS |
| CENPL | miR-1285-5p | 2316 | -113 | 3'UTR |
| FAM73A | miR-1285-5p | 2974 | -113 | 3'UTR |
| LRAT | miR-1285-5p | 3475 | -113 | 3'UTR |
| TSN | miR-1285-5p | 1964 | -113 | 3'UTR |
| GPR179 | miR-4478 | 2280 | -96 | CDS |
| KHSRP | miR-4478 | 66 | -96 | 5'UTR |
| PRR12 | miR-4478 | 4396 | -96 | CDS |
| RNF111 | miR-4478 | 2436 | -96 | CDS |
| KANK3 | miR-4508 | 510 | -106 | CDS |
| OXR1 | miR-4508 | 103 | -106 | 5'UTR |
| PXDNL | miR-4508 | 2686 | -106 | CDS |
| ALG1 | miR-4526 | 864 | -127 | CDS |
| ALG1L2 | miR-4526 | 400 | -127 | CDS |
| ITGAL | miR-5095 | 4708 | -117 | 3'UTR |
| KANK4 | miR-5095 | 4753 | -117 | 3'UTR |
| RBBP4 | miR-5095 | 4279 | -117 | 3'UTR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



