
Submitted:
02 February 2026
Posted:
05 February 2026
You are already at the latest version
Abstract
This study investigates the application of soybean peroxidase (SBP), an enzyme extracted from a soybean processing byproduct, for the decolourization and oxidative treatment of three industrial azo dyes: Acid Orange 7 (AO7), Acid Orange 20 (AO20), and Reactive Black 5 (RB5), each at a concentration of 50 µM. These dyes are widely used in textile, paper, and leather industries and persist in wastewater. Optimization experiments were conducted at room temperature (approximately 22°C) to examine the effects of pH, SBP activity, and hydrogen peroxide (H2O2) concentration. Optimal degradation conditions were identified as: for RB5, pH 3.5, 0.075 U/mL SBP, and 0.0375 mM H2O2; for AO7, pH 3.0, 0.5 U/mL SBP, and 0.0375 mM H2O2; and for AO20, pH 3.0, 0.0025 U/mL SBP (200-fold less than for the isomeric AO7) and 0.0625 mM H2O2. Under these conditions, dye removal kinetics followed pseudo-first-order behaviour during the initial stages, assuming constant SBP and H2O2 concentrations. However, accumulation of oligomeric products, depletion of H2O2 over time, and enzyme inactivation caused deviation from first-order kinetics. The initial rate constants and half-lives were 10.7 min-1 and 0.065 min (AO7), 7.3 min-1 and 0.095 min (AO20), and 8.5 min-1 and 0.081 min (RB5). When normalized to enzyme activity, AO7 showed the highest catalytic efficiency. More than 95% decolourization was achieved in under 30 seconds. These findings support the use of SBP as a low-cost, eco-friendly, and effective biocatalyst for the rapid treatment of dye-containing industrial wastewater.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Buffer Preparation
2.3. Enzyme Stock Solutions
2.4. Soybean Peroxidase Activity Assay
2.5. Dye Reactions
2.6. HPLC Analysis
3. Results and Dicussion
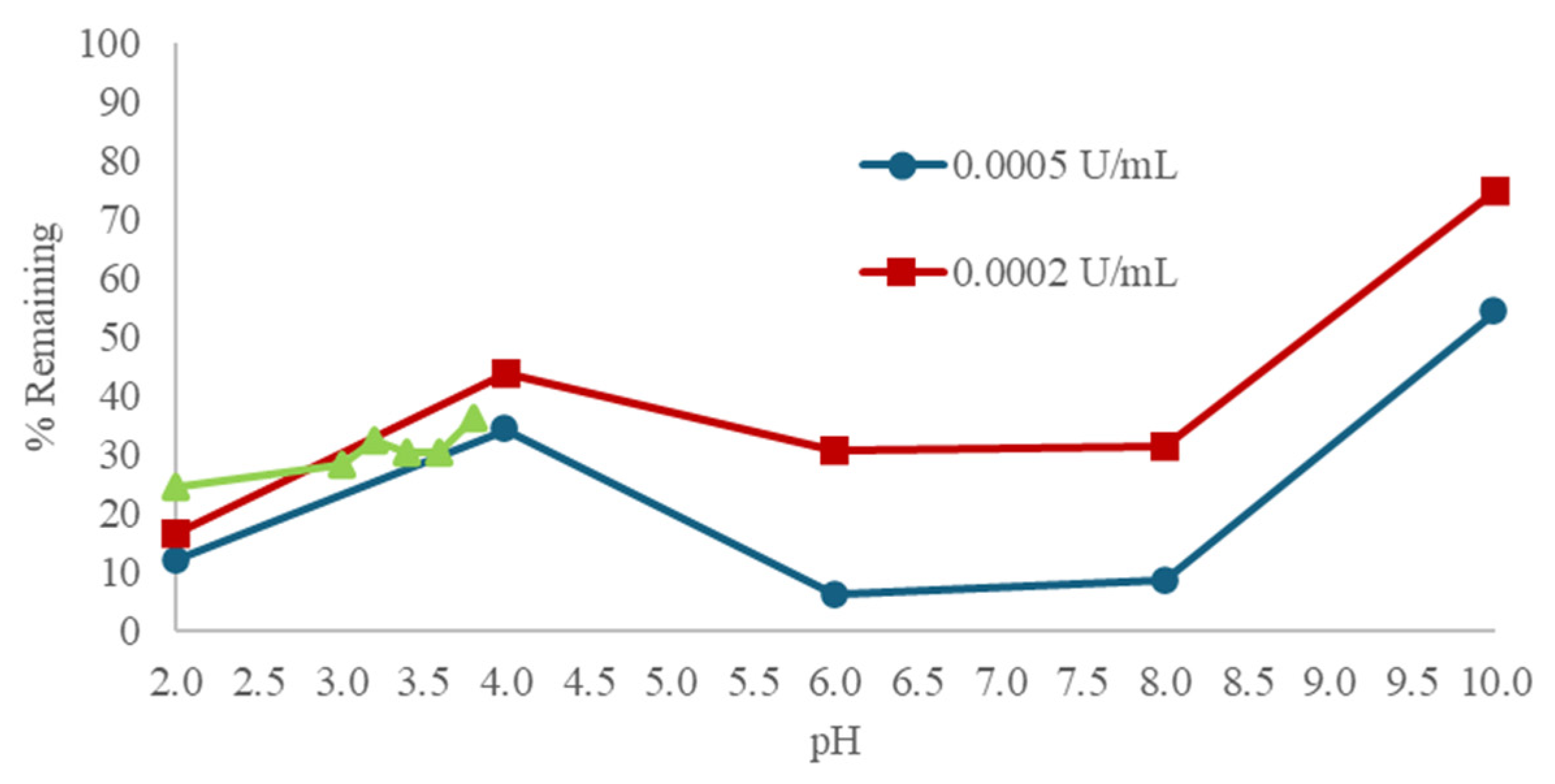
3.1. Effect of pH
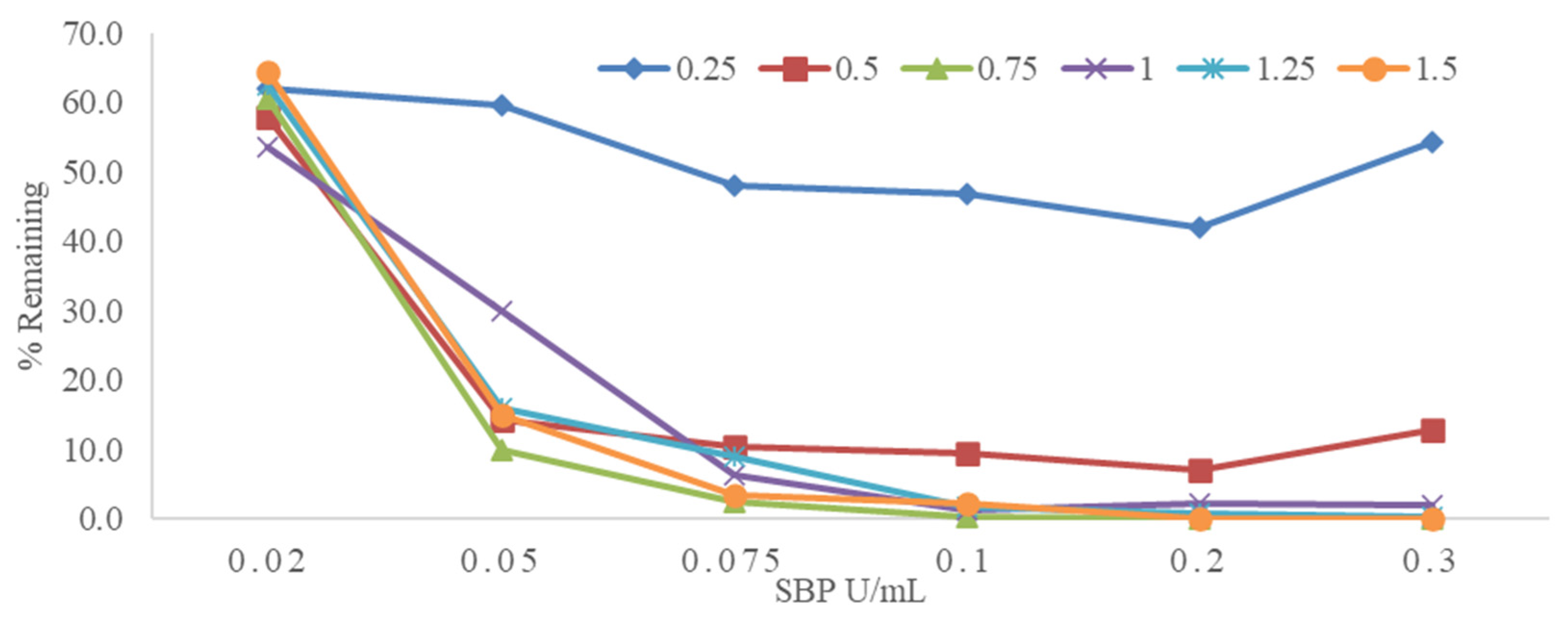
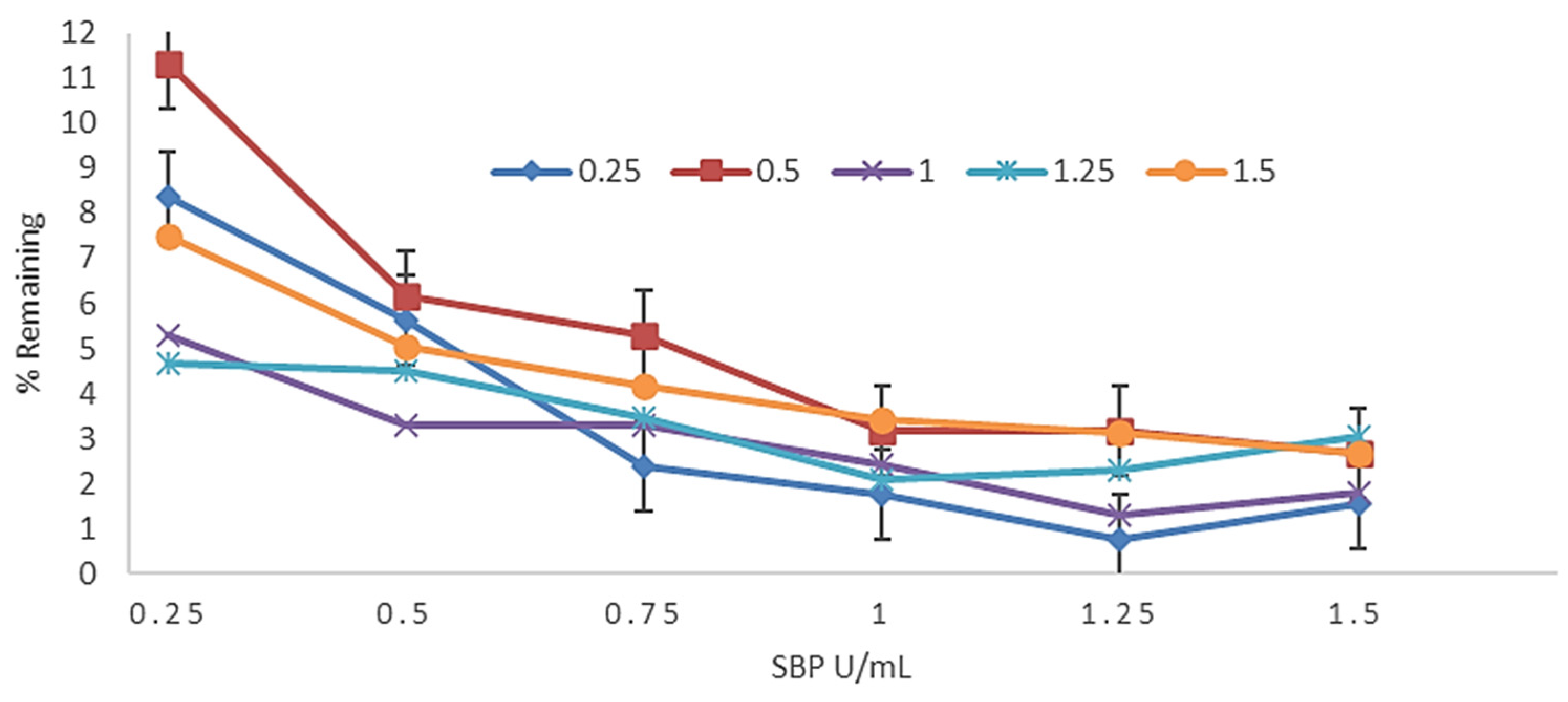
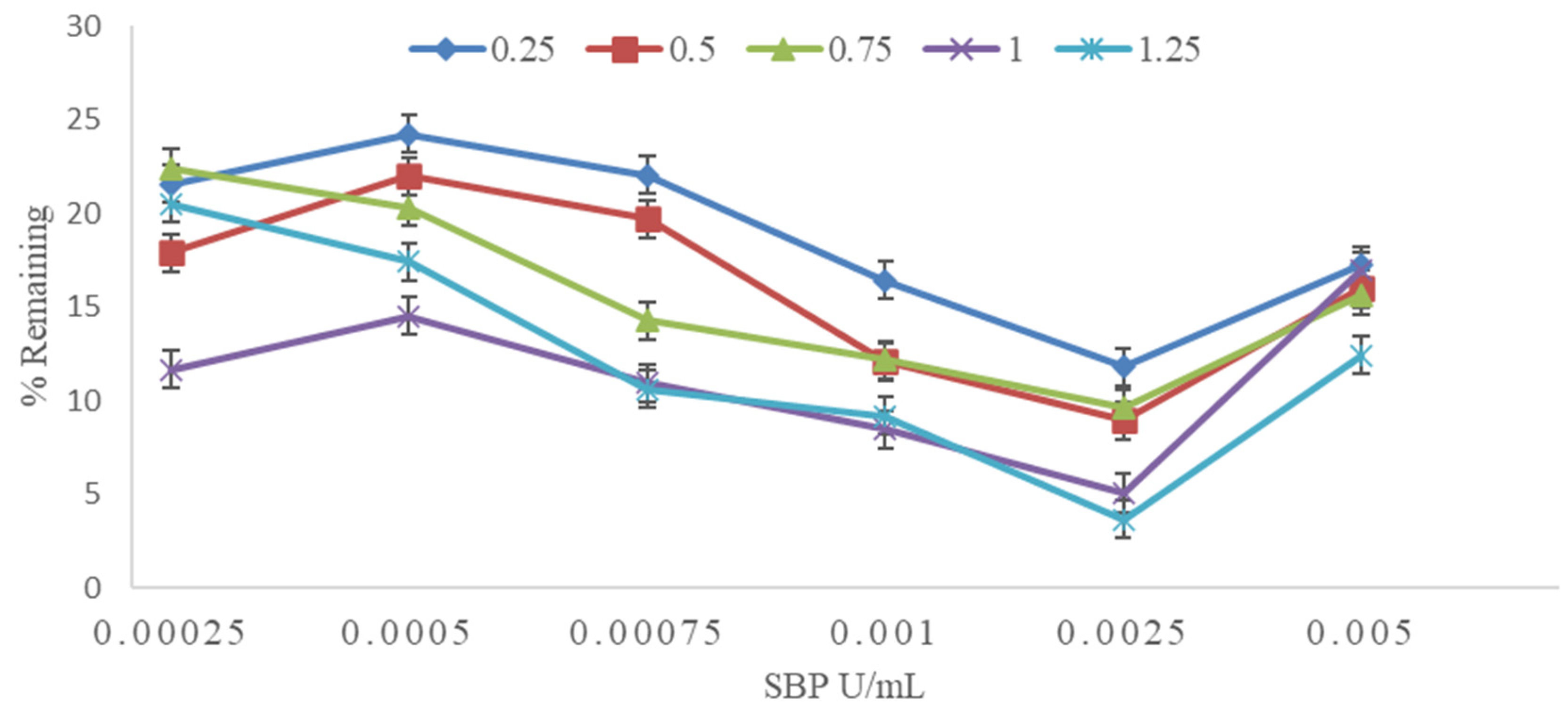
3.2. Effect of SBP and H2O2
3.3. SBP Efficiency in Dye Removal
3.4. Kinetic Measurements
3.5. Products
4. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Compliance With Ethical Standards
Conflicts of Interest
References
- Allen, N., Colour Chemistry, by H. Zollinger, Weinheim: VCH Verlagsgesellschaft, 1987, 367 pp. J. Photochem. Photobiol. A: Chem., 1987, vol. 41, no. 1, p. 145.2.
- Lewis, D.M., Developments in the chemistry of reactive dyes and their application processes, Color. Technol., 2014, vol. 130, no. 6, pp. 382–412.
- Saleem, M.A., Pei, L., Saleem, M.F., Shahid, S., and Wang, J., Sustainable dyeing of nylon fabric with acid dyes in decamethylcyclopentasiloxane (D5) solvent for improving dye uptake and reducing raw material consumption, J. Clean. Prod., 2021, vol. 279, p. 123480.
- Singh, S.N., Microbial Degradation of Xenobiotics (Springer, Berlin–Heidelberg, 2012).
- Ghaly, A., Ananthashankar, R., Alhattab, M., and Ramakrishnan, V.V., Production, characterization and treatment of textile effluents: a critical review, J. Chem. Eng. Process Technol., 2014, vol. 5, no. 1, pp. 1–19.
- Piaskowski, K., Świderska-Dąbrowska, R., and Zarzycki, P.K., Dye removal from water and wastewater using various physical, chemical, and biological processes, J. AOAC Int., 2018, vol. 101, no. 5, pp. 1371–1384.
- Katheresan, V., Kansedo, J., and Lau, S.Y., Efficiency of various recent wastewater dye removal methods: A review, J. Environ. Chem. Eng., 2018, vol. 6, no. 4, pp. 4676–4697.
- Slama, H.B., Chenari Bouket, A., Pourhassan, Z., Alenezi, F.N., Silini, A., Cherif-Silini, H., et al., Diversity of synthetic dyes from textile industries, discharge impacts and treatment methods, Appl. Sci., 2021, vol. 11, no. 14, 6255.
- Das, S. and Mahalingam, H., Multiphase reactors in photocatalytic treatment of dye wastewaters: design and scale-up considerations, in Advanced Oxidation Processes in Dye-Containing Wastewater, Sharma, Y.C. and Sanghi, R., Eds., Singapore: Springer, 2022, vol. 1, pp. 219–250.
- Salehi, S., Abdollahi, K., Panahi, R., Rahmanian, N., Shakeri, M., Mokhtarani, B. Applications of biocatalysts for sustainable oxidation of phenolic pollutants: A review. Sustainability, 2021, 13, 8620.
- Jun, L.Y., Yon, L.S., Mubarak, N., Bing, C.H., Pan, S., Danquah, M.K., et al., An overview of immobilized enzyme technologies for dye and phenolic removal from wastewater, J. Environ. Chem. Eng., 2019, vol. 7, no. 2, 102961.
- Šekuljica, N.Ž., Prlainović, N.Ž., Stefanović, A.B., Žuža, M.G., Čičkarić, D.Z., Mijin, D.Ž., et al., Decolorization of anthraquinonic dyes from textile effluent using horseradish peroxidase: optimization and kinetic study, Sci. World J., 2015, vol. 2015, no. 1, pp. 371625.
- Barreiro, D.S., Oliveira, R.N., and Pauleta, S.R., Bacterial peroxidases–Multivalent enzymes that enable the use of hydrogen peroxide for microaerobic and anaerobic proliferation, Coord. Chem. Rev., 2023, vol. 485, 215114.
- Freitas, C.D., Costa, J.H., Germano, T.A., Rocha, R.d.O., Ramos, M.V., and Bezerra, L.P., Class III plant peroxidases: From classification to physiological functions, Int. J. Biol. Macromol., 2024, vol. 263, 130306.
- ur Rahman, M., Ullah, M. W., Shah, J. A., Sethupathy, S., Bilal, H., Abdikakharovich, S. A., Khan, A. U., Khan, K. A., Elboughdiri, N., Zhu, D. Harnessing the power of bacterial laccases for xenobiotic degradation in water: A 10-year overview. Sci. Tot. Environ., 2024, 918, 170498.
- Sellami, K., Couvert, A., Nasrallah, N.,Maachi, R., Abouseoud, M.,Amrane, A. Peroxidase enzymes as green catalysts for bioremediation and biotechnological applications: A review. Sci, Tot. Environ., 2022, 806, 150500.
- Pishyar, S., Narimannejad, S., Taylor, K.E., and Biswas, N., Enzymatic removal of diclofenac and aceclofenac from water by soybean peroxidase, Molecules, 2025, vol. 30, no. 8, pp. 1817.
- Kamal, J.A. and Behere, D.V., Kinetic stabilities of soybean and horseradish peroxidases, Biochem. Eng. J., 2008, vol. 38, no. 1, pp. 110–114.
- Gogliettino, M., Cocca, E., Apone, F., Del Prete, S., Balestrieri, M., Mirino, S., et al., A new versatile peroxidase with extremophilic traits over-produced in MicroTom cell cultures, Sci. Rep., 2023, vol. 13, no. 1, p. 15338.
- Chen, Z., Wu, J., Huang, W., Li, Y., Mao, Y., Han, J., et al., Preparation and catalytic degradation of phenol achieved by utilizing pH and upper critical solution temperature dual responsive intelligent enzyme catalysts, J. Clean. Prod., 2023, vol. 423, 138579.
- Dunford, H.B., Heme Peroxidase Kinetics, in Heme Peroxidases, Dunford, H.B. and Raven, E.,, Ed., Cambridge: Cambridge University Press, 2015, pp. 99–112.
- Uyama, H., Synthesis of poly(aromatics). I: Oxidoreductase as catalyst, in Enzymatic Polymerization Towards Green Polymer Chemistry, 2019, pp. 267–305.
- Urlacher, V., Peroxidases and Catalases: Biochemistry, Biophysics, Biotechnology and Physiology, Dunford, H.B., Ed., Wiley, Hoboken,2010.
- Wright, H., Nicell, J. A. Characterization of soybean peroxidase for the treatment of aqueous phenols. Bioresour. Technol., 1999, 70, 69-79.
- Ortiz de Montellano, P.R., Hydrocarbon hydroxylation by cytochrome P450 enzymes, Chem. Rev., 2010, vol. 110, no. 2, pp. 932–948.
- Feng, W., Taylor, K., Biswas, N., and Bewtra, J.K., Phenolic precipitates from soybean peroxidase–catalyzed wastewater treatment: concentrated waste serves to concentrate its progenitor, J. Hazard. Toxic Radioact. Waste, 2016, vol. 20, no. 2, p. 04015023.
- Arnao, M., Acosta, M., del Río, J., Varón, R., and García-Cánovas, F., A kinetic study on the suicide inactivation of peroxidase by hydrogen peroxide, Biochim. Biophys. Acta, Protein Struct. Mol. Enzymol., 1990, vol. 1041, no. 1, pp. 43–47.
- Gomori, G., Preparation of buffers for use in enzyme studies, in Methods in Enzymology, Colowick, S.P. and Kaplan, N.O., Eds., New York: Academic Press, 1955, vol. 1, part C, pp. 138–146.
- Baumer, J., Valério, A., Souza, S., Guelli, S., Erzinger, G., and Furigo, A., Jr., Remazol black dye (reactive black 5) decolorization by horseradish peroxidase enzyme, J. Textile Eng. Fashion Technol., 2018, vol. 4, no. 2, pp. 147–149.
- Yousefi, V. and Kariminia, H.-R., Statistical analysis for enzymatic decolorization of acid orange 7 by Coprinus cinereus peroxidase, Int. Biodeterior. Biodegrad., 2010, vol. 64, no. 3, pp. 245–252.
- Saroyan, H., Ntagiou, D., Rekos, K., and Deliyanni, E., Reactive black 5 degradation on manganese oxides supported on sodium hydroxide modified graphene oxide, Appl. Sci., 2019, vol. 9, no. 10, 2167.
- Murugesan, S., Sasibabu, V., Jegadeesan, G.B., and Venkatachalam, P., Photocatalytic degradation of Reactive Black dye using ZnO–CeO2 nanocomposites, Environ. Sci. Pollut. Res., 2023, vol. 30, no. 15, pp. 42713–42727.
- Hsueh, C.-C., Chen, B.-Y., and Yen, C.-Y., Understanding effects of chemical structure on azo dye decolorization characteristics by Aeromonas hydrophila, J. Hazard. Mater., 2009, vol. 167, nos. 1–3, pp. 995–1001.
- Zimmermann, T., Kulla, H.G., and Leisinger, T., Properties of purified Orange II azoreductase, the enzyme initiating azo dye degradation by Pseudomonas KF46, Eur. J. Biochem., 1982, vol. 129, no. 1, pp. 197–203.
- Gholami-Borujeni, F., Mahvi, A.H., Nasseri, S., Faramarzi, M.A., Nabizadeh, R., and Alimohammadi, M., Enzymatic treatment and detoxification of acid orange 7 from textile wastewater, Appl. Biochem. Biotechnol., 2011, vol. 165, no. 5, pp. 1274–1284.
- Zhang, A., Fang, L., Wang, J., and Liu, W., Enzymatic decolorization of Orange II: optimization by response surface methodology and pathway, Environ. Prog. Sustain. Energy, 2013, vol. 32, no. 2, pp. 294–301.
- Morales Urrea, D.A., Haure, P.M., García Einschlag, F.S., and Contreras, E.M., Horseradish peroxidase-mediated decolourization of Orange II: modelling hydrogen peroxide utilization efficiency at different pH values, Environ. Sci. Pollut. Res., 2018, vol. 25, no. 20, pp. 19989–20002.
- Morales Urrea, D.A., Haure, P.M., and Contreras, E.M., Orange II removal by a horseradish peroxidase immobilized onto chemically modified diatomites is a combination of adsorption and oxidation processes, J. Appl. Res. Technol., 2022, vol. 20, no. 2, pp. 203–220.
- López, C., Valade, A.-G., Combourieu, B., Mielgo, I., Bouchon, B., and Lema, J.M., Mechanism of enzymatic degradation of the azo dye Orange II determined by ex situ ¹H nuclear magnetic resonance and electrospray ionization-ion trap mass spectrometry, Anal. Biochem., 2004, vol. 335, no. 1, pp. 135–149.
- Asgher, M., Kausar, S., Bhatti, H.N., Shah, S.A.H., and Ali, M., Optimization of medium for decolorization of Solar golden yellow R direct textile dye by Schizophyllum commune IBL-06, Int. Biodeterior. Biodegrad., 2008, vol. 61, no. 2, pp. 189–193.
- Zhu, Y., Liu, Y., Guo, X., Wu, Q., and Xu, X. Decolorization of Color Index Acid Orange 20 buffer solution using horseradish peroxidase immobilized on modified PAN-beads, RSC Adv., 2017, vol. 7, no. 31, pp. 18976–18986.
- Haghighatnama, M., Narimannejad, S., Biswas, N., and Taylor, K.E. Biocatalytic treatment of cresols in aqueous solution with soybean peroxidase, RSC Adv., 2026, 16, pp. 5079–5087.
- Altahir, B.M., Feng, W., Jasim, H.H., Taylor, K.E., Biswas, N., Bewtra, J.K., et al., Soybean peroxidase-catalysed removal of benzidines from water, J. Environ. Eng. Sci., 2016, vol. 10, no. 4, pp. 73–80.
- Mukherjee, D., Bhattacharya, S., Taylor, K.E., and Biswas, N., Enzymatic treatment for removal of hazardous aqueous arylamines, 4,4′-methylenedianiline and 4,4′-thiodianiline, Chemosphere, 2019, vol. 235, pp. 365–372.
- Sharifzadeh, M., Narimannejad, S., Taylor, K.E., and Biswas, N., Enzymatic removal of the sulfa drugs sulfamethoxazole and sulfamerazine from synthetic wastewater by soybean peroxidase, Environ. Sci. Pollut. Res., 2024, vol. 31, no. 56, pp. 64760–64771.
- Yarkwan, B., Soybean peroxidase catalyzed oligomerization of acetaminophen and selected parabens for wastewater treatment. Ph. D. Thesis, Windsor: University of Windsor, 2021.
- Cordova Villegas, L.G., Mazloum, S., Taylor, K.E., and Biswas, N., Soybean peroxidase-catalyzed treatment of azo dyes with or without FeO pretreatment, Water Environ. Res., 2018, vol. 90, no. 8, pp. 675–684.
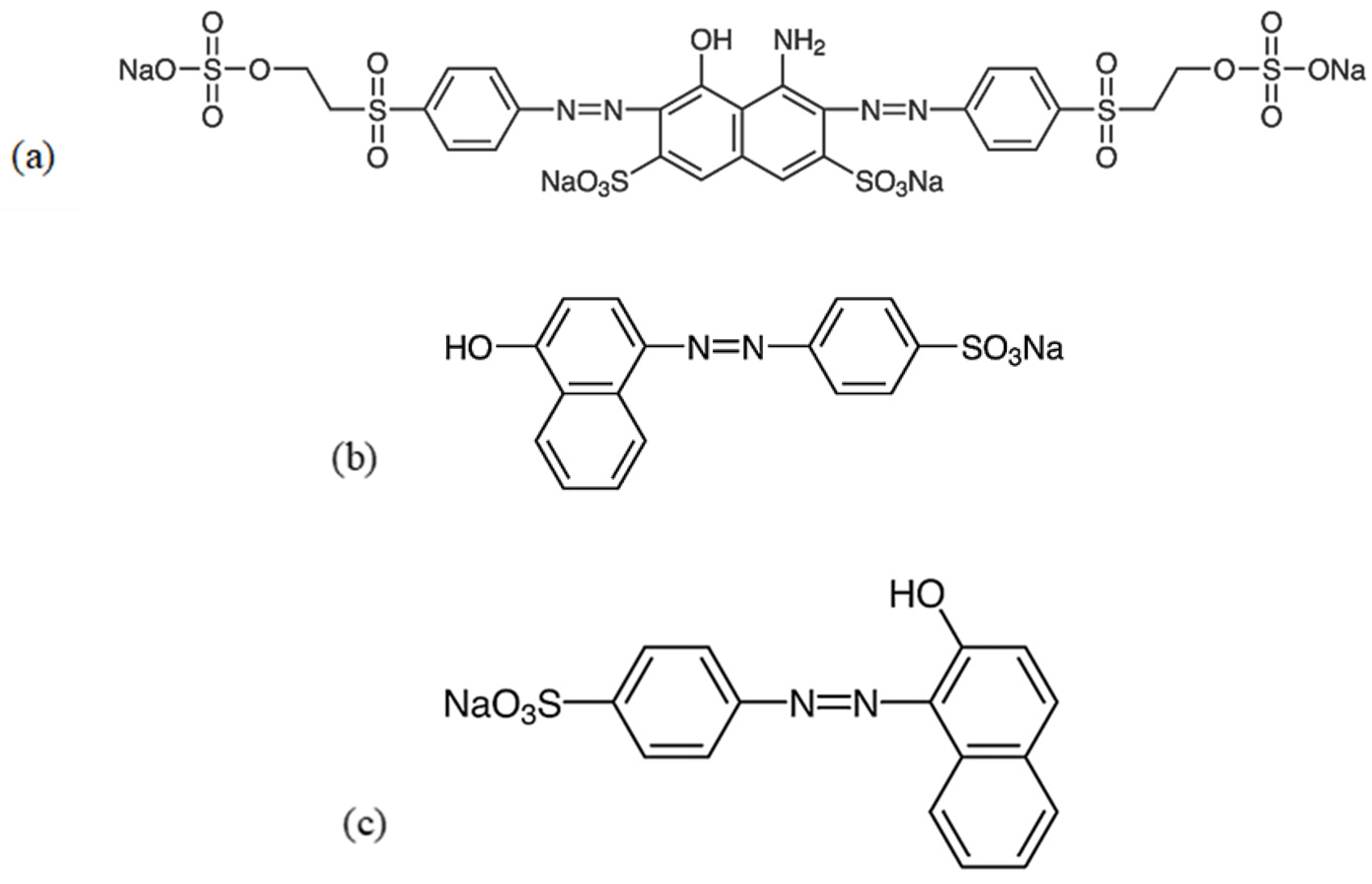
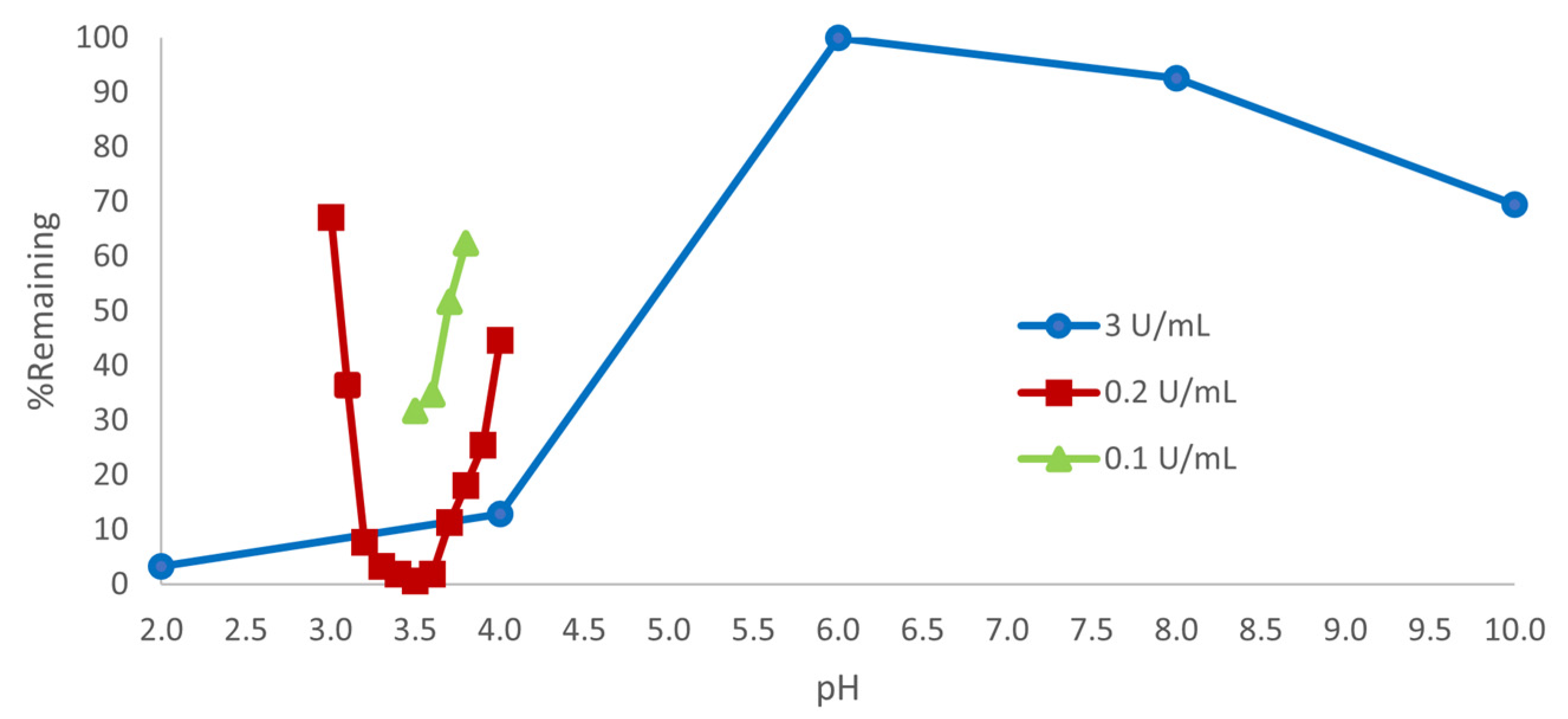
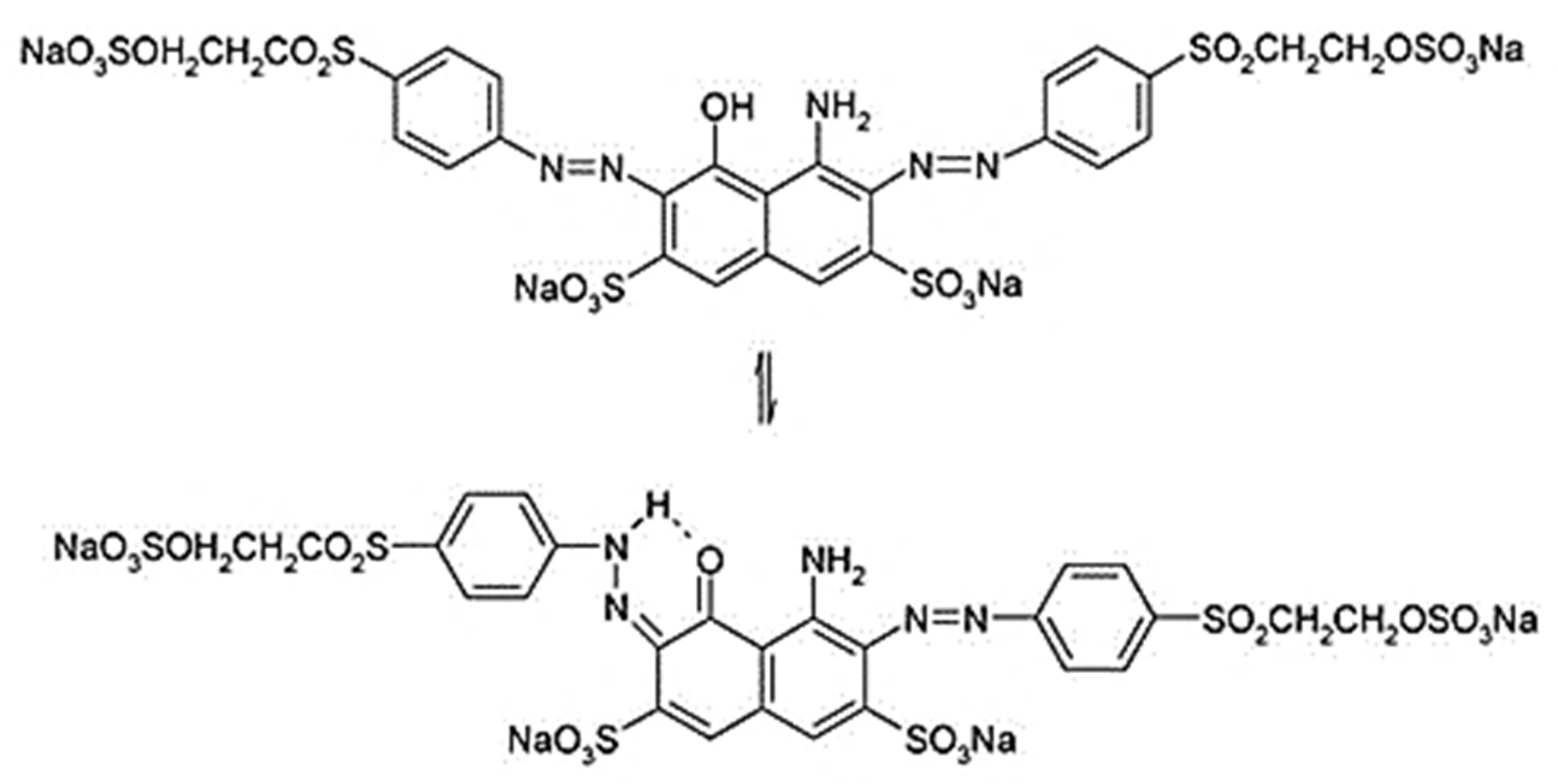
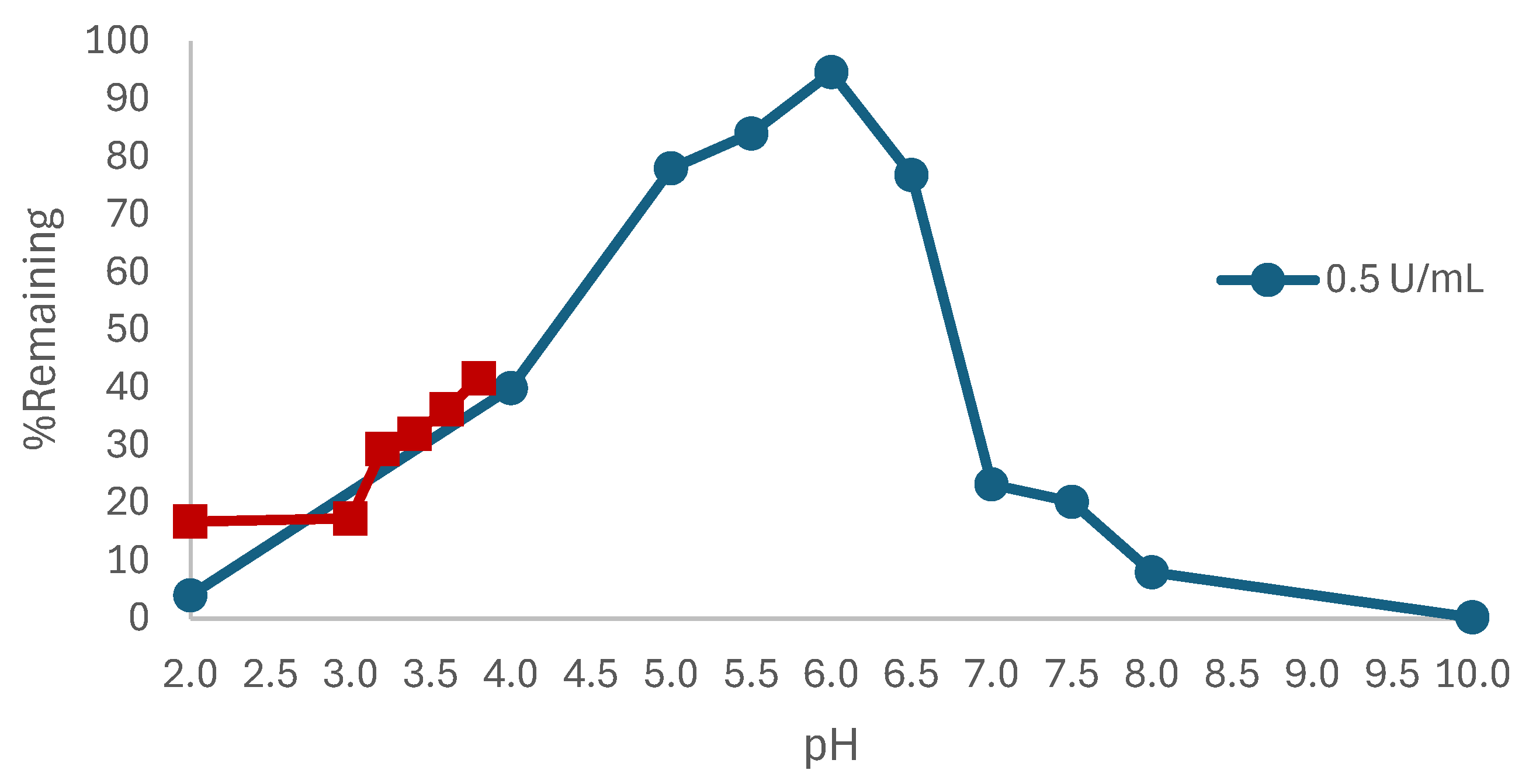

|
Dye |
Aqueous Phase A |
Organic Phase B |
Mobile Phase composition (%) | Wavelength (λmax) | |
| A | B | ||||
| RB5 | Ammonium Formate (5 mM) |
Methanol | 70 | 30 | 596 nm |
| AO7 | Ammonium Formate (5 mM) |
Methanol | 40 | 60 | 485 nm |
| AO20 | Ammonium Formate (5 mM) |
Methanol | 40 | 60 | 485 nm |
| Dye | pH | SBP Activity (U/mL) |
H₂O₂ (mM) |
Normalized SBP* (U/mL/mM) |
Removal % |
| RB5 | 3.5 | 0.075 | 0.0375 | 1.5 | 98.6 |
| AO7 | 3.0 | 0.5 | 0.0375 | 10 | 99.5 |
| AO20 | 3.0 | 0.0025 | 0.0625 | 0.05 | 97.4 |
| Substrate |
Rate constant (min-1) |
Half-life (min) |
Normalized rate constant* (min-1) |
Normalized half-life (min) |
| RB5 | ≥8.5 | ≤0.081 | ≥0.64 | ≤0.0061 |
| AO7 | ≥10.7 | ≤0.065 | ≥5.3 | ≤0.033 |
| AO20 | ≥7.3 | ≤0.095 | ≥0.018 | ≤0.00024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



