
Submitted:
03 February 2026
Posted:
03 February 2026
You are already at the latest version
Abstract
Glypican-2 (GPC2) is a heparan sulfate proteoglycan that plays essential roles in enhancing re-ceptor signaling by recruiting heparan sulfate-binding ligands. Overexpression of GPC2 is ob-served in pediatric cancers, including neuroblastoma and medulloblastoma, where it supports oncogenic signaling and tumor progression. High-affinity GPC2 binders have been developed as effective immunotherapies targeting tumors with GPC2 overexpression. In this study, we gener-ated new anti-GPC2 monoclonal antibodies using a flow cytometry–based high-throughput screening. Among the clones, G2Mab-7 (IgG1, κ) selectively bound GPC2-overexpressing Chinese hamster ovary cells (CHO/GPC2), with no reactivity toward parental CHO-K1 cells in flow cy-tometry. Notably, G2Mab-7 also recognized endogenous GPC2 in the human neuroblastoma cell line IMR-32. Specificity testing showed that G2Mab-7 reacted with CHO/GPC2 and did not crossreact with other GPCs overexpressed in CHO-K1. The dissociation constant (KD) for G2Mab-7 was 2.3 × 10⁻⁸ M for CHO/GPC2 and 4.2 × 10⁻⁹ M for IMR-32, indicating moderate binding affinity. Additionally, G2Mab-7 can be used in western blotting and immunohistochem-istry. Overall, these findings show that G2Mab-7 is a versatile tool for basic research and a promising candidate for antibody-based therapies.
Keywords:
glypican-2
; monoclonal antibody
; flow cytometry
; western blotting
; immunohistochemistry
1. Introduction
Glypicans constitute a major class of extracellular matrix–associated heparan sulfate (HS) proteoglycans [1,2]. In mammals, the glypican family comprises six members, glypican-1 (GPC1) through glypican-6 (GPC6), which exhibit a high degree of amino acid sequence conservation across species [3]. Despite this evolutionary conservation, individual glypicans exhibit distinct structural features and exert nonredundant biological functions [4]. Owing to their ability to modulate multiple signaling pathways, glypicans have garnered considerable attention for their critical roles in development [4] and disease, particularly in tumor [5].
GPC2 acts as a context-dependent signaling co-receptor through its heparan sulfate (HS) chains. GPC2 increases Wnt ligand availability and signaling strength, promoting cell proliferation and survival [6]. GPC2 can influence the distribution and gradient formation of hedgehog ligands, affecting developmental patterning [7]. Physiologically, GPC2 is mainly expressed during embryonic and early postnatal development, particularly in the nervous system. GPC2 is involved in neural progenitor proliferation, neuronal differentiation, axon guidance, and neural circuit formation [8,9,10]. Therefore, GPC2 plays a role in early neural development, and its expression is significantly decreased in most adult tissues [5,11].
High-risk neuroblastoma remains a highly aggressive pediatric cancer, with long-term survival rates below 50% despite intensive multimodal chemoradiotherapy. This highlights a critical unmet need for innovative treatment approaches [12]. GPC2 promotes tumor growth in high-risk neuroblastoma and is transcriptionally controlled by MYCN [13]. GPC2 maintains oncogenic Wnt signaling and encourages neuroblastoma cell growth and progression [14]. Therefore, GPC2 is an immunotherapeutic target in high-risk neuroblastoma [13]. GPC2-directed immunotherapeutic modalities, including chimeric antigen receptor (CAR) T cells, have been developed. Second-generation GPC2 CAR T cell therapies demonstrate potent antitumor activity and a favorable safety profile in preclinical models [15,16,17,18,19]. Notably, one of the GPC2 CAR T cell products is currently being evaluated in a first-in-human phase I clinical trial (NCT05650749). In pediatric neuroblastoma and other disialoganglioside 2 (GD2)-positive tumors, GD2 CAR T cells are also being evaluated for safety and efficacy in several clinical trials [20,21,22]. These efforts have recently yielded the first compelling clinical evidence supporting the therapeutic efficacy of CAR T cells in this pediatric embryonal tumor [21]. Nevertheless, as observed with other highly potent CAR constructs and across solid tumor indications, therapeutic resistance frequently emerges due to antigen downregulation under CAR-mediated selective pressure [18,23], limited CAR T cell persistence, and the profoundly immunosuppressive tumor microenvironment (TME) [24,25].
The single-chain variable fragment (scFv) derived from a monoclonal antibody (mAb) has been a central focus of CAR design, as it plays a decisive role in determining antigen specificity and therapeutic efficacy [26]. The scFv binding affinity critically influences the functional potency of CAR T cells. In general, higher-affinity scFvs enhance CAR signaling and antitumor activity [27]. However, excessively high antigen-binding affinity can impair CAR T cell serial killing capacity and persistence [28]. Moreover, high-affinity scFvs may increase on-target, off-tumor toxicity by enabling recognition of low-level antigen expression in normal tissues [29,30]. In addition to affinity, epitope selection is a critical determinant of CAR T cell performance [31]. Therefore, the development of various mAbs is essential for fine-tuning scFv to achieve an optimal balance between efficacy and safety.
Using the Cell-Based Immunization and Screening (CBIS) method, various mAbs against membrane proteins, such as GPC5 [32], cadherins [33,34], and receptor tyrosine kinases [35,36] have been developed. The CBIS method involves immunizing antigen-overexpressing cells and performing high-throughput screening by flow cytometry. This study employed the CBIS method to develop versatile anti-GPC2 mAbs.
2. Materials and Methods
2.1. Cell Lines
IMR-32 was obtained from the Japanese Collection of Research Bioresources (Osaka, Japan). Chinese hamster ovary (CHO)-K1, mouse myeloma P3X63Ag8U.1 (P3U1), and human glioblastoma LN229 were cultured as described previously [34].
2.2. Plasmid Construction and Establishment of Stable Transfectants
The cDNA of human GPC2 (NM_152742) was obtained from RIKEN RBC (Ibaraki, Japan). The human GPC1-6 overexpressed CHO-K1 were established as described previously [32]. The mouse GPC2 (NM_172412.2) cDNA was obtained from OriGene Technologies, Inc. (Rockville, MD, USA). The mouse GPC2 cDNA was cloned into a pCAG-Ble-ssnPA16 vector. The plasmid was transfected into CHO-K1, and stable transfectants were established as described previously [32].
2.3. Production of Hybridomas
The female BALB/cAJcl mice (CLEA Japan, Tokyo, Japan) were immunized intraperitoneally with LN229/GPC2 cells (1 × 108 cells/injection) and 2% Alhydrogel adjuvant (InvivoGen). After three additional immunizations of LN229/GPC2 per week, a booster injection was administered two days before harvesting the spleen cells from immunized mice. The hybridomas were generated as previously described [32].
2.4. Flow Cytometry and Determination of Dissociation Constant Values
The cells were treated with primary mAbs in blocking buffer [0.1% bovine serum albumin (BSA) in phosphate-buffered saline]. The cells were then stained with anti-mouse IgG conjugated with Alexa Fluor 488 (Cell Signaling Technology, Inc.). The data were collected using an SA3800 Cell Analyzer (Sony Corporation, Tokyo, Japan) and analyzed using FlowJo software (BD Biosciences, Franklin Lakes, NJ, USA).
The fitted binding isotherms were used to determine dissociation constant (KD) values, which were used to build one-sided binding models in GraphPad Prism 6 (GraphPad Software, Inc., La Jolla, CA, USA).
2.5. Western Blotting
Western blotting was performed using G2Mab-7 (0.1 μg/mL), CT3 (0.1 μg/mL), or a β-actin mAb (clone AC-15, Sigma-Aldrich Corp., St. Louis, MO, 1 μg/mL) as described previously [34].
2.6. IHC Using Cell Blocks
All procedures of IHC were performed using VENTANA BenchMark ULTRA PLUS (Roche Diagnostics, Indianapolis, IN, USA). The formalin-fixed paraffin-embedded (FFPE) cell sections were stained with G2Mab-7 (1 μg/mL) or CT3 (0.1 μg/mL) using the ultraView Universal DAB Detection Kit and BenchMark ULTRA PLUS.
3. Results
3.1. Development of anti-GPC2 mAbs
LN229/GPC2 was used as an antigen to develop anti-GPC2 mAbs (Figure 1A). Hybridomas were generated by fusing LN229/GPC2-immunized splenocytes with P3U1 cells (Figure 1B). After colony formation, hybridoma supernatants were screened for reactivity that was positive for CHO/GPC2 and negative for CHO-K1 (Figure 1C). Subsequently, anti-GPC2 mAb-producing hybridomas were cloned by limiting dilution. A total of 16 clones were finally established. After screening for the availability to flow cytometry, western blotting, and immunohistochemistry, a clone G2Mab-7 (IgG1, κ) was finally selected (Figure 1D).
3.2. Specificity of G2Mab-7 Against GPC Family Members
We previously established GPC1, GPC3, GPC4, GPC5, and PA16-GPC6-overexpressed CHO-K1 cells (named as CHO/GPC1, CHO/GPC3, CHO/GPC4, CHO/GPC5, and CHO/GPC6) [32]. We next investigated the specificity of G2Mab-7 to the cell lines used. As shown in Figure 2A, G2Mab-7 recognized CHO/GPC2 and did not react with other GPC-overexpressed CHO-K1. All GPC expression was detected using the specific mAbs (Figure 2B). These results confirmed the specificity of G2Mab-7 to GPC2 among GPC family members.
3.3. Flow Cytometry Using G2Mab-7 and CT3
We performed flow cytometry using the G2Mab-7 and a commercially available anti-GPC2 mAb (clone CT3) against CHO-K1 and CHO/GPC2. The G2Mab-7 and CT3 recognized CHO/GPC2 dose-dependently from 1 to 0.001 μg/mL (Figure 3A), but did not recognize CHO-K1 even at 1 μg/mL (Figure 3B). We next examined the cross-reactivity of G2Mab-7 to mouse GPC2. As shown in Figure 3C, G2Mab-7 showed cross-reactivity to mouse GPC2. CT3 did not recognize mouse GPC2 as described previously [11]. We further investigated the reactivity of G2Mab-7 and CT3 against an endogenous GPC2-expressing cell line, IMR-32. G2Mab-7 and CT3 showed similar dose-dependent reactivity to IMR-32 (Figure 4). These results indicate that G2Mab-7 recognizes human and mouse GPC2 in flow cytometry.
3.4. Determination of KD Values of G2Mab-7 and CT3 by Flow Cytometry
The KD values of G2Mab-7 were next determined using a flow cytometry-based assay. The KD values of G2Mab-7 and CT3 for CHO/GPC2 were 2.3 × 10⁻8 M and 1.3 × 10⁻8 M, respectively (Figure 5). The KD value of G2Mab-7 for IMR-32 was also determined as 4.2 × 10⁻9 M (Supplementary Figure 1). These results indicated that both G2Mab-7 and CT3 have similar moderate affinity to GPC2-positive cells.
3.5. Western Blotting Using G2Mab-7 and CT3
We next examined whether G2Mab-7 is suitable for western blotting of whole-cell lysates from CHO-K1, CHO/GPC2, and IMR-32. G2Mab-7 and CT3 detected a major 63 kDa and lower bands in CHO/GPC2, but not in CHO-K1 (Figure 6A, B upper panels). CT3 was detected with high sensitivity under the same exposure conditions. In a long exposure, G2Mab-7 could detect an endogenous 63 kDa band in IMR-32 (Figure 6A, B, lower panels). Smear bands were also detected at more than 63 kDa in CHO/GPC2 and IMR-32. β-actin detected by AC-15 was used as an internal control (Figure 6C). These results indicate that G2Mab-7 can detect overexpressed and endogenous GPC2 in western blotting.
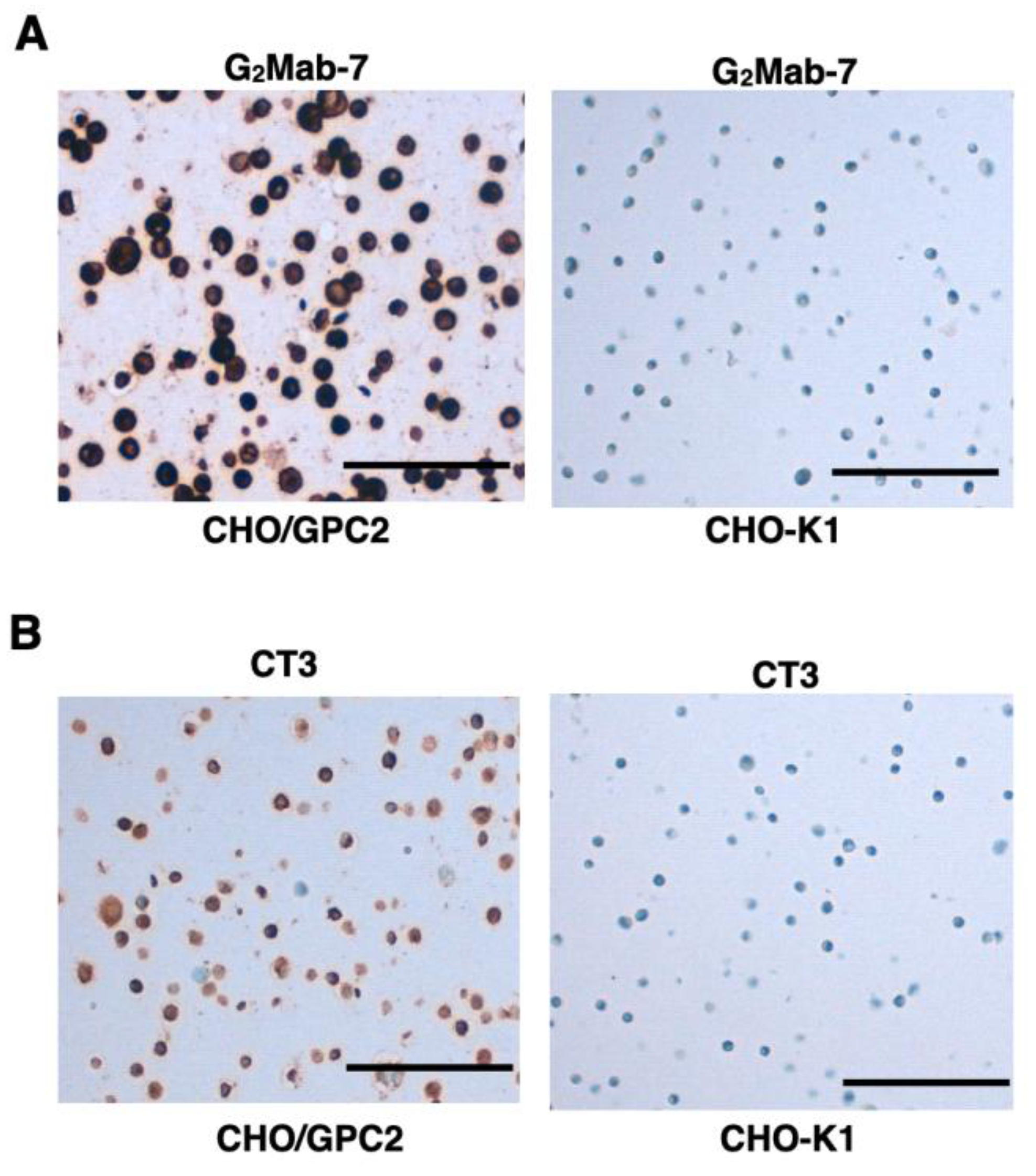
3.6. IHC Using G2Mab-7 and CT3 in FFPE Cell Sections
We examined whether G2Mab-7 is suitable for the IHC of CHO-K1 and CHO/GPC2 sections. G2Mab-7 showed membranous and cytoplasmic staining in CHO/GPC2 but not in CHO-K1 at 1 µg/mL (Figure 7A). CT3 could detect the membranous and cytoplasmic staining in CHO/GPC2 but not in CHO-K1 at 0.1 µg/mL (Figure 7B), although 0.1 µg/mL of G2Mab-7 could not detect GPC2, indicating that the sensitivity of CT3 is much better in IHC than G2Mab-7.
4. Discussion
This study demonstrated the property of a novel anti-GPC2 mAb, clone G2Mab-7, established by the CBIS method. In flow cytometry, G2Mab-7 exhibited specificity to GPC2 among GPC family members (Figure 2) and similar reactivity/affinity to GPC2-positive cells compared to CT3 (Figure 3, Figure 4 and Figure 5). Furthermore, G2Mab-7 can detect GPC2 in western blotting (Figure 6) and GPC2-positive cells in IHC (Figure 7). Although we obtained 16 clones of anti-GPC2 mAbs, G2Mab-7 is a highly versatile mAb, but others are only suitable for flow cytometry (http://www.med-tohoku-antibody.com/topics/001_paper_antibody_PDIS.htm). Some of G2Mabs, including G2Mab-7, exhibit cross-reactivity to mouse GPC2 (Figure 3C), which is useful to assess the on-target, off-tumor toxicity in mouse models. We will determine their binding epitope and affinity, which will be useful for developing novel CARs. Since we have determined the epitope of various transmembrane (TM) proteins including single TM [37], 2TM [38], 4TM [39], and 7TM [40] receptors, these strategies will be applied to the determination of G2Mabs’ epitopes.
An anti-GPC2 mAb (CT3) used in this study was previously developed by immunization of the membrane-proximal C-terminal region (exon 10) of GPC2 [11]. Surprisingly, CT3 was shown to bind not only to the C-terminal region (immunogen) but also to the exon 3 fragment of GPC2 [11]. Negative stain electron microscopy analysis revealed that CT3 formed a rigid and stable complex with GPC2 through interaction with exon 3 and exon 10 epitopes, which show close localization based on the 3D structure [11]. The GPC2 exon 10-encoded region possesses two serine residues modified with HS. CT3 was generated by the immunization of the C-terminal 50 amino acids of the GPC2 peptide [11]. Since the HS-binding ligands such as FGF, Wnt, and vascular endothelial growth factor ligands are present in TME [41], the HS-modifications and ligand binding may influence the CT3 binding in vivo. Therefore, the preparation of anti-GPC2 mAbs targeting different epitopes is considered important for therapeutic applications.
CT3 has been developed for CAR T cells and has suppressed neuroblastoma growth in preclinical mouse orthotopic and metastasis models [11,42]. Furthermore, the CT3 CAR T also demonstrated significant tumor regression in a patient-derived medulloblastoma xenograft, the most common pediatric malignant brain tumor [43]. A study arm for pediatric GPC2-positive medulloblastoma is planned in an upcoming clinical trial using the CT3 CAR T cells [44]. Additionally, CT3 is also applicable to IHC for GPC2-positive neuroblastoma, retinoblastoma, and medulloblastoma [11]. Further study is essential to determine whether G2Mab-7 can stain tumor tissues with the automated slide-staining system used in this study, thereby ensuring standardized, reproducible staining conditions.
Author Contributions
Kenichiro Ishikawa: Investigation, Writing – original draft; Hiroyuki Suzuki: Writing – original draft; Akira Ohkoshi: Conceptualization; Mika K. Kaneko: Conceptualization; Yukio Katori: Conceptualization; Yukinari Kato: Conceptualization, Funding acquisition, Project administration, Writing – review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP25am0521010 (to Yukinari Kato), JP25ama121008 (to Yukinari Kato), JP25ama221153 (to Yukinari Kato), JP25ama221339 (to Yukinari Kato), and JP25bm1123027 (to Yukinari Kato), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant no. 25K10553 (to Yukinari Kato).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- Filmus, J. Glypicans, 35 years later. Proteoglycan Research 2023, 1, e5. [Google Scholar] [CrossRef]
- Filmus, J.; Capurro, M.; Rast, J. Glypicans. Genome Biol 2008, 9, 224. [Google Scholar] [CrossRef]
- Filmus, J. Glypicans in growth control and cancer. Glycobiology 2001, 11, 19r–23r. [Google Scholar] [CrossRef]
- Filmus, J. The function of glypicans in the mammalian embryo. Am J Physiol Cell Physiol 2022, 322, C694–C698. [Google Scholar] [CrossRef]
- Li, N.; Spetz, M.R.; Ho, M. The Role of Glypicans in Cancer Progression and Therapy. J Histochem Cytochem 2020, 68, 841–862. [Google Scholar] [CrossRef]
- Li, N.; Fu, H.; Hewitt, S.M.; Dimitrov, D.S.; Ho, M. Therapeutically targeting glypican-2 via single-domain antibody-based chimeric antigen receptors and immunotoxins in neuroblastoma. Proc Natl Acad Sci U S A 2017, 114, E6623–E6631. [Google Scholar] [CrossRef]
- Filmus, J.; Capurro, M. The role of glypicans in Hedgehog signaling. Matrix Biol 2014, 35, 248–252. [Google Scholar] [CrossRef]
- Ouchida, J.; Ozaki, T.; Segi, N.; Suzuki, Y.; Imagama, S.; Kadomatsu, K.; Sakamoto, K. Glypican-2 defines age-dependent axonal response to chondroitin sulfate. Exp Neurol 2023, 366, 114444. [Google Scholar] [CrossRef] [PubMed]
- Lugert, S.; Kremer, T.; Jagasia, R.; Herrmann, A.; Aigner, S.; Giachino, C.; Mendez-David, I.; Gardier, A.M.; Carralot, J.P.; Meistermann, H.; et al. Glypican-2 levels in cerebrospinal fluid predict the status of adult hippocampal neurogenesis. Sci Rep 2017, 7, 46543. [Google Scholar] [CrossRef]
- Ivins, J.K.; Litwack, E.D.; Kumbasar, A.; Stipp, C.S.; Lander, A.D. Cerebroglycan, a developmentally regulated cell-surface heparan sulfate proteoglycan, is expressed on developing axons and growth cones. Dev Biol 1997, 184, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Torres, M.B.; Spetz, M.R.; Wang, R.; Peng, L.; Tian, M.; Dower, C.M.; Nguyen, R.; Sun, M.; Tai, C.H.; et al. CAR T cells targeting tumor-associated exons of glypican 2 regress neuroblastoma in mice. Cell Rep Med 2021, 2, 100297. [Google Scholar] [CrossRef]
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat Rev Dis Primers 2016, 2, 16078. [Google Scholar] [CrossRef]
- Bosse, K.R.; Raman, P.; Zhu, Z.; Lane, M.; Martinez, D.; Heitzeneder, S.; Rathi, K.S.; Kendsersky, N.M.; Randall, M.; Donovan, L.; et al. Identification of GPC2 as an Oncoprotein and Candidate Immunotherapeutic Target in High-Risk Neuroblastoma. Cancer Cell 2017, 32, 295–309.e212. [Google Scholar] [CrossRef] [PubMed]
- Tsao, H.E.; Ho, M. Structural Features of Glypicans and their Impact on Wnt Signaling in Cancer. Proteoglycan Res 2025, 3. [Google Scholar] [CrossRef]
- Pascual-Pasto, G.; McIntyre, B.; Hines, M.G.; Giudice, A.M.; Garcia-Gerique, L.; Hoffmann, J.; Mishra, P.; Matlaga, S.; Lombardi, S.; Shraim, R.; et al. CAR T-cell-mediated delivery of bispecific innate immune cell engagers for neuroblastoma. Nat Commun 2024, 15, 7141. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Pasto, G.; McIntyre, B.; Giudice, A.M.; Alikarami, F.; Morrissey, A.; Matlaga, S.; Hofmann, T.J.; Burgueño, V.; Harvey, K.; Martinez, D.; et al. Targeting GPC2 on Intraocular and CNS Metastatic Retinoblastomas with Local and Systemic Delivery of CAR T Cells. Clin Cancer Res 2024, 30, 3578–3591. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Cao, Y.; Okada, R.; Reyes-González, J.M.; Stack, H.G.; Qin, H.; Li, N.; Seibert, C.; Kelly, M.C.; Ruppin, E.; et al. Preclinical optimization of a GPC2-targeting CAR T-cell therapy for neuroblastoma. J Immunother Cancer 2023, 11. [Google Scholar] [CrossRef]
- Heitzeneder, S.; Bosse, K.R.; Zhu, Z.; Zhelev, D.; Majzner, R.G.; Radosevich, M.T.; Dhingra, S.; Sotillo, E.; Buongervino, S.; Pascual-Pasto, G.; et al. GPC2-CAR T cells tuned for low antigen density mediate potent activity against neuroblastoma without toxicity. Cancer Cell 2022, 40, 53–69.e59. [Google Scholar] [CrossRef]
- Foster, J.B.; Griffin, C.; Rokita, J.L.; Stern, A.; Brimley, C.; Rathi, K.; Lane, M.V.; Buongervino, S.N.; Smith, T.; Madsen, P.J.; et al. Development of GPC2-directed chimeric antigen receptors using mRNA for pediatric brain tumors. J Immunother Cancer 2022, 10. [Google Scholar] [CrossRef]
- Li, C.H.; Sharma, S.; Heczey, A.A.; Woods, M.L.; Steffin, D.H.M.; Louis, C.U.; Grilley, B.J.; Thakkar, S.G.; Wu, M.; Wang, T.; et al. Long-term outcomes of GD2-directed CAR-T cell therapy in patients with neuroblastoma. Nat Med 2025, 31, 1125–1129. [Google Scholar] [CrossRef]
- Del Bufalo, F.; De Angelis, B.; Caruana, I.; Del Baldo, G.; De Ioris, M.A.; Serra, A.; Mastronuzzi, A.; Cefalo, M.G.; Pagliara, D.; Amicucci, M.; et al. GD2-CART01 for Relapsed or Refractory High-Risk Neuroblastoma. N Engl J Med 2023, 388, 1284–1295. [Google Scholar] [CrossRef]
- Majzner, R.G.; Ramakrishna, S.; Yeom, K.W.; Patel, S.; Chinnasamy, H.; Schultz, L.M.; Richards, R.M.; Jiang, L.; Barsan, V.; Mancusi, R.; et al. GD2-CAR T cell therapy for H3K27M-mutated diffuse midline gliomas. Nature 2022, 603, 934–941. [Google Scholar] [CrossRef]
- Majzner, R.G.; Mackall, C.L. Tumor Antigen Escape from CAR T-cell Therapy. Cancer Discov 2018, 8, 1219–1226. [Google Scholar] [CrossRef]
- Yan, T.; Zhu, L.; Chen, J. Current advances and challenges in CAR T-Cell therapy for solid tumors: tumor-associated antigens and the tumor microenvironment. Exp Hematol Oncol 2023, 12, 14. [Google Scholar] [CrossRef]
- Tumino, N.; Weber, G.; Besi, F.; Del Bufalo, F.; Bertaina, V.; Paci, P.; Quatrini, L.; Antonucci, L.; Sinibaldi, M.; Quintarelli, C.; et al. Polymorphonuclear myeloid-derived suppressor cells impair the anti-tumor efficacy of GD2.CAR T-cells in patients with neuroblastoma. J Hematol Oncol 2021, 14, 191. [Google Scholar] [CrossRef] [PubMed]
- Dotti, G.; Gottschalk, S.; Savoldo, B.; Brenner, M.K. Design and development of therapies using chimeric antigen receptor-expressing T cells. Immunol Rev 2014, 257, 107–126. [Google Scholar] [CrossRef]
- Chang, Z.L.; Chen, Y.Y. CARs: Synthetic Immunoreceptors for Cancer Therapy and Beyond. Trends Mol Med 2017, 23, 430–450. [Google Scholar] [CrossRef]
- Ma, P.; Ren, P.; Zhang, C.; Tang, J.; Yu, Z.; Zhu, X.; Fan, K.; Li, G.; Zhu, W.; Sang, W.; et al. Avidity-Based Selection of Tissue-Specific CAR-T Cells from a Combinatorial Cellular Library of CARs. Adv Sci (Weinh) 2021, 8, 2003091. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Chen, R.; Huang, Y.; Meng, X.; Chen, J.; Liao, C.; Tang, Y.; Zhou, C.; Gao, X.; Sun, J. Tuning the ignition of CAR: optimizing the affinity of scFv to improve CAR-T therapy. Cell Mol Life Sci 2021, 79, 14. [Google Scholar] [CrossRef]
- Richman, S.A.; Nunez-Cruz, S.; Moghimi, B.; Li, L.Z.; Gershenson, Z.T.; Mourelatos, Z.; Barrett, D.M.; Grupp, S.A.; Milone, M.C. High-Affinity GD2-Specific CAR T Cells Induce Fatal Encephalitis in a Preclinical Neuroblastoma Model. Cancer Immunol Res 2018, 6, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Yoo, K.H.; Szymura, S.; Dong, Z.; Kandel, A.; Cha, S.C.; Kwak, L.W. Predictive markers for the efficacy of CAR T-cell therapy: the interplay between CAR T-cell fitness and systemic immunity. Blood Adv 2025, 9, 6432–6442. [Google Scholar] [CrossRef]
- Kaneko, Y.; Tanaka, T.; Fujisawa, S.; Li, G.; Satofuka, H.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Development of a novel anti-human glypican 5 monoclonal antibody (G(5)Mab-1) for multiple applications. Biochem Biophys Rep 2025, 43, 102140. [Google Scholar] [CrossRef]
- Ubukata, R.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of novel anti-CDH1/E-cadherin monoclonal antibodies for versatile applications. Biochem Biophys Rep 2026, 45, 102401. [Google Scholar] [CrossRef]
- Ubukata, R.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of an anti-CDH15/M-cadherin monoclonal antibody Ca(15)Mab-1 for flow cytometry, immunoblotting, and immunohistochemistry. Biochem Biophys Rep 2025, 43, 102138. [Google Scholar] [CrossRef]
- Satofuka, H.; Suzuki, H.; Tanaka, T.; Li, G.; Kaneko, M.K.; Kato, Y. Development of an anti-human EphA2 monoclonal antibody Ea(2)Mab-7 for multiple applications. Biochem Biophys Rep 2025, 42, 101998. [Google Scholar] [CrossRef]
- Itai, S.; Yamada, S.; Kaneko, M.K.; Chang, Y.W.; Harada, H.; Kato, Y. Establishment of EMab-134, a Sensitive and Specific Anti-Epidermal Growth Factor Receptor Monoclonal Antibody for Detecting Squamous Cell Carcinoma Cells of the Oral Cavity. Monoclon Antib Immunodiagn Immunother 2017, 36, 272–281. [Google Scholar] [CrossRef]
- Sano, M.; Kaneko, M.K.; Aasano, T.; Kato, Y. Epitope Mapping of an Antihuman EGFR Monoclonal Antibody (EMab-134) Using the REMAP Method. Monoclon Antib Immunodiagn Immunother 2021, 40, 191–195. [Google Scholar] [CrossRef]
- Okada, Y.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Epitope Mapping of an Anti-Mouse CD39 Monoclonal Antibody Using PA Scanning and RIEDL Scanning. Monoclon Antib Immunodiagn Immunother 2024, 43, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Takei, J.; Furusawa, Y.; Saito, M.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Epitope Mapping of an Anti-CD20 Monoclonal Antibody (C(20)Mab-60) Using the HisMAP Method. Monoclon Antib Immunodiagn Immunother 2021, 40, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Epitope Mapping of an Anti-Mouse CCR8 Monoclonal Antibody C(8)Mab-2 Using Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2024, 43, 101–107. [Google Scholar] [CrossRef] [PubMed]
- de Visser, K.E.; Joyce, J.A. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell 2023, 41, 374–403. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Nguyen, R.; Thiele, C.J.; Ho, M. Preclinical testing of chimeric antigen receptor T cells in neuroblastoma mouse models. STAR Protoc 2021, 2, 100942. [Google Scholar] [CrossRef]
- Okada, R.; Fanuzzi, M.; Rodriguez, C.; Oh, J.; Sreenivasan, A.; Stack, H.G.; Puebla, M.; Phadke, I.; Cole, A.P.; Bradford, S.; et al. GPC2-CAR T cells have potent preclinical activity against orthotopic medulloblastoma xenografts. Mol Ther Oncol 2025, 33, 201067. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Foster, J.B.; Bosse, K.R. Scratching the surface of targeting GPC2 on pediatric CNS tumors. Mol Ther Oncol 2025, 33, 201102. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Procedure of anti-GPC2 mAbs production. (A) LN229/GPC2 was intraperitoneally injected into BALB/cAJcl mice. (B) The splenocytes were fused with P3U1 after five immunizations per week. (C) Selection of the CHO/GPC2-positive and CHO-K1-negative supernatants of hybridomas. (D) Anti-GPC2-specific mAb-producing hybridoma clones were established by limiting dilution.
Figure 1.
Procedure of anti-GPC2 mAbs production. (A) LN229/GPC2 was intraperitoneally injected into BALB/cAJcl mice. (B) The splenocytes were fused with P3U1 after five immunizations per week. (C) Selection of the CHO/GPC2-positive and CHO-K1-negative supernatants of hybridomas. (D) Anti-GPC2-specific mAb-producing hybridoma clones were established by limiting dilution.
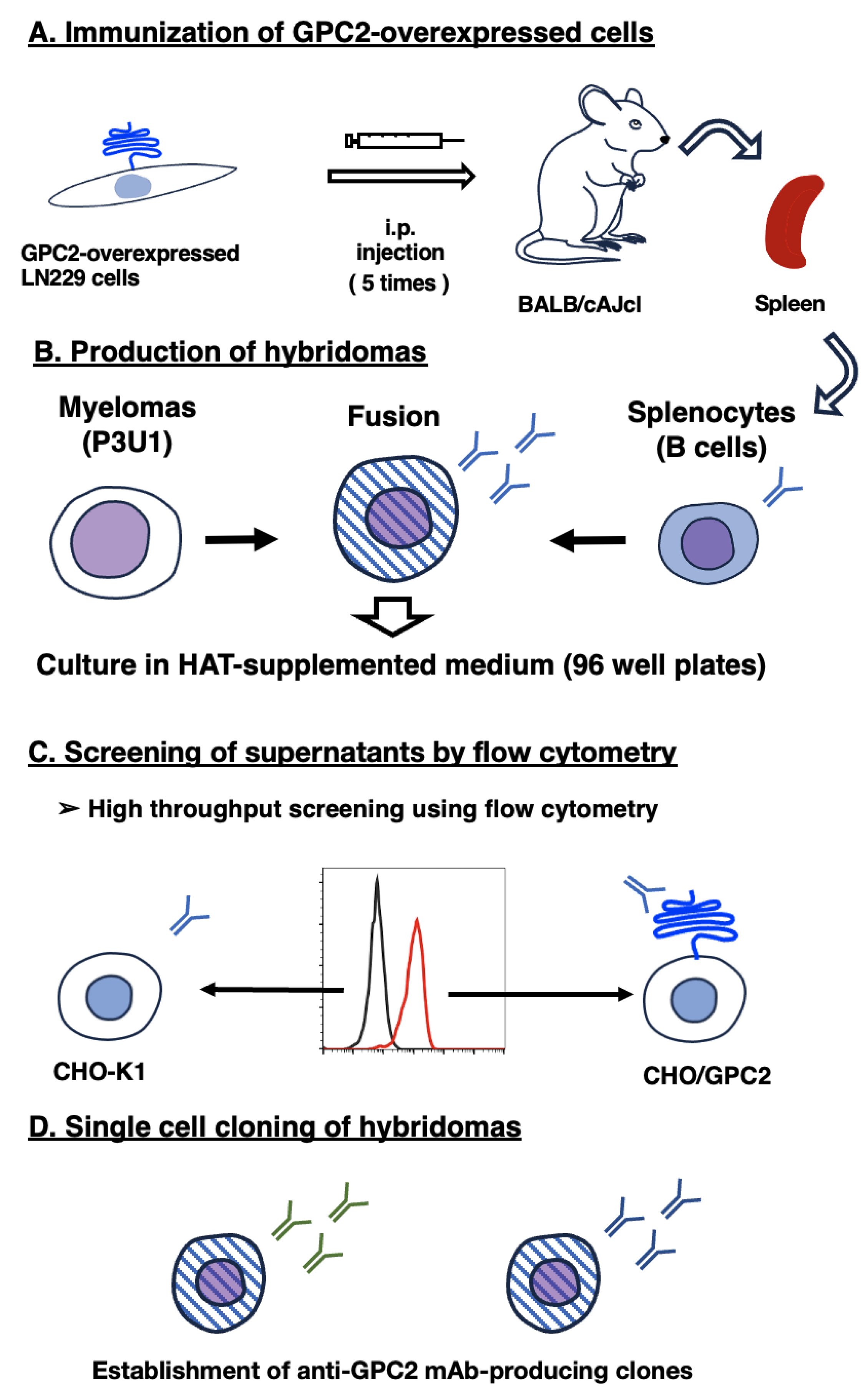
Figure 2.
Specificity of G2Mab-7 in GPCs-overexpressed CHO-K1. (A) The GPCs (GPC1, GPC2, GPC3, GPC4, GPC5, and PA16-GPC6)-overexpressed CHO-K1 cells were treated with 1 µg/mL of G2Mab-7 (red) or control blocking buffer (black), followed by treatment with anti-mouse IgG conjugated with Alexa Fluor 488. (B) The expression of each GPC was confirmed by using 1 µg/mL of an anti-GPC1 mAb (1019718), 1 µg/mL of an anti-GPC2 mAb (CT3), 1 µg/mL of an anti-GPC3 mAb (SP86), 1 µg/mL of an anti-GPC4 mAb (A21050B), 1 µg/mL of an anti-GPC5 mAb (297716), and 1 µg/mL of an anti-PA16 mAb, NZ-1.
Figure 2.
Specificity of G2Mab-7 in GPCs-overexpressed CHO-K1. (A) The GPCs (GPC1, GPC2, GPC3, GPC4, GPC5, and PA16-GPC6)-overexpressed CHO-K1 cells were treated with 1 µg/mL of G2Mab-7 (red) or control blocking buffer (black), followed by treatment with anti-mouse IgG conjugated with Alexa Fluor 488. (B) The expression of each GPC was confirmed by using 1 µg/mL of an anti-GPC1 mAb (1019718), 1 µg/mL of an anti-GPC2 mAb (CT3), 1 µg/mL of an anti-GPC3 mAb (SP86), 1 µg/mL of an anti-GPC4 mAb (A21050B), 1 µg/mL of an anti-GPC5 mAb (297716), and 1 µg/mL of an anti-PA16 mAb, NZ-1.
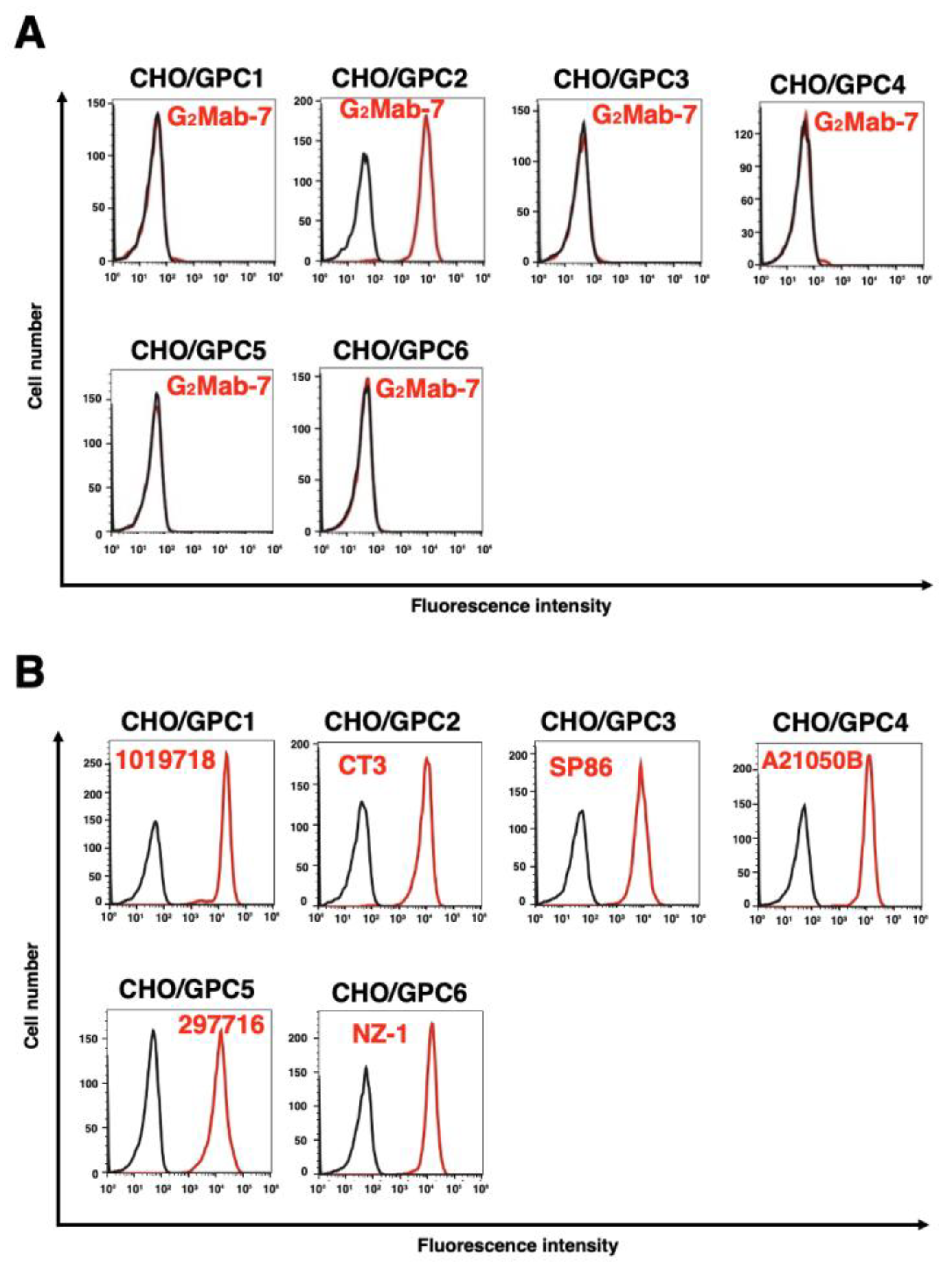
Figure 3.
Flow cytometry analysis of G2Mab-7 and CT3 against GPC2-overexpressed CHO-K1. (A and B) CHO/GPC2 (A) and CHO-K1 (B) were treated with G2Mab-7 or CT3 at the indicated concentrations (red) or blocking buffer (black). (C) CHO-K1, CHO/GPC2, and CHO/mouse GPC2 were treated with G2Mab-7 (10 µg/mL, blue) or blocking buffer (black). The cells were incubated with anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were collected using the SA3800 Cell Analyzer.
Figure 3.
Flow cytometry analysis of G2Mab-7 and CT3 against GPC2-overexpressed CHO-K1. (A and B) CHO/GPC2 (A) and CHO-K1 (B) were treated with G2Mab-7 or CT3 at the indicated concentrations (red) or blocking buffer (black). (C) CHO-K1, CHO/GPC2, and CHO/mouse GPC2 were treated with G2Mab-7 (10 µg/mL, blue) or blocking buffer (black). The cells were incubated with anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were collected using the SA3800 Cell Analyzer.
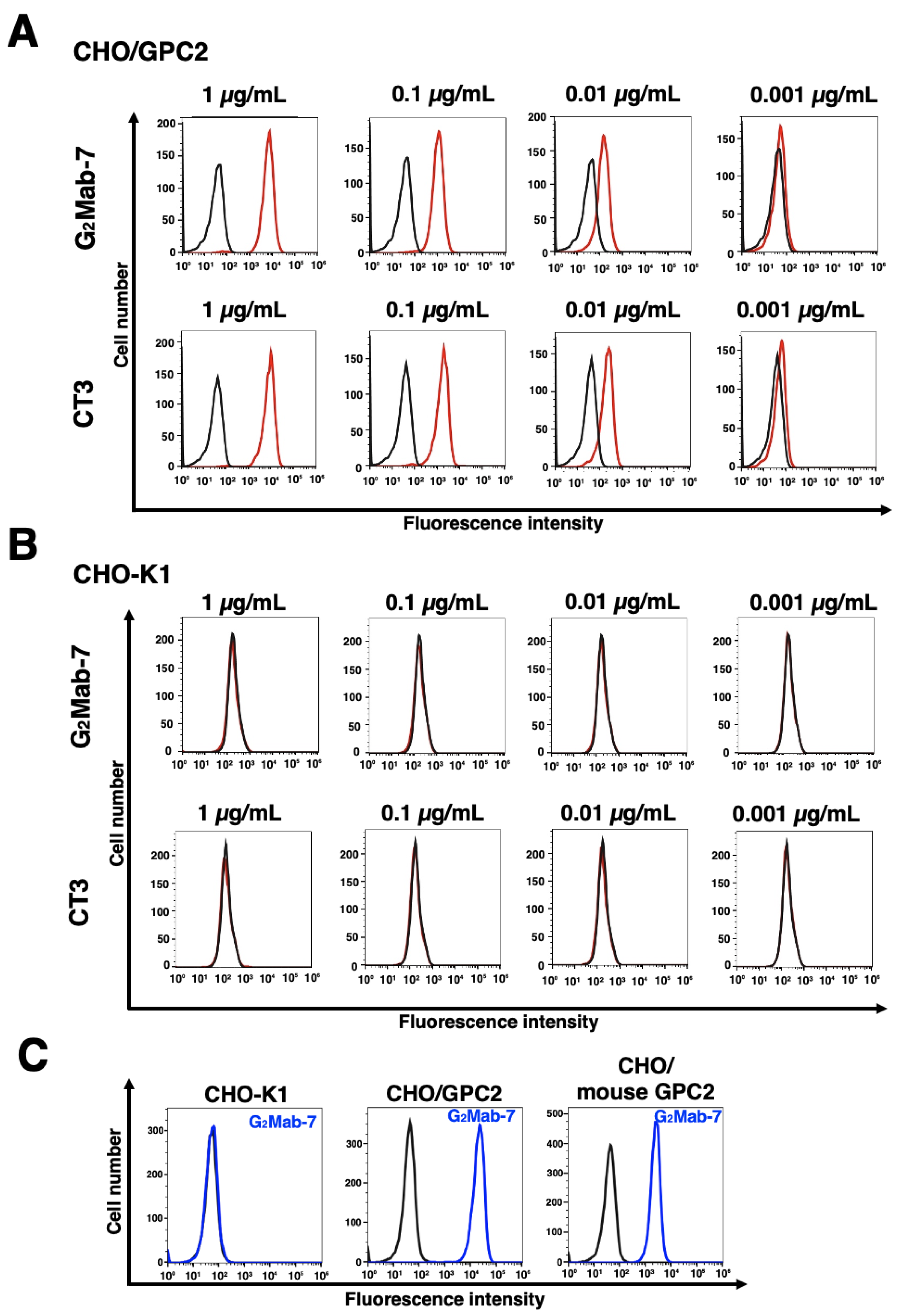
Figure 4.
Flow cytometry analysis of G2Mab-7 and CT3 against endogenous GPC2-positive neuroblastoma IMR-32. IMR-32 was treated with G2Mab-7 or CT3 at the indicated concentrations (red) or blocking buffer (black). The cells were incubated with anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were collected using the SA3800 Cell Analyzer.
Figure 4.
Flow cytometry analysis of G2Mab-7 and CT3 against endogenous GPC2-positive neuroblastoma IMR-32. IMR-32 was treated with G2Mab-7 or CT3 at the indicated concentrations (red) or blocking buffer (black). The cells were incubated with anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were collected using the SA3800 Cell Analyzer.
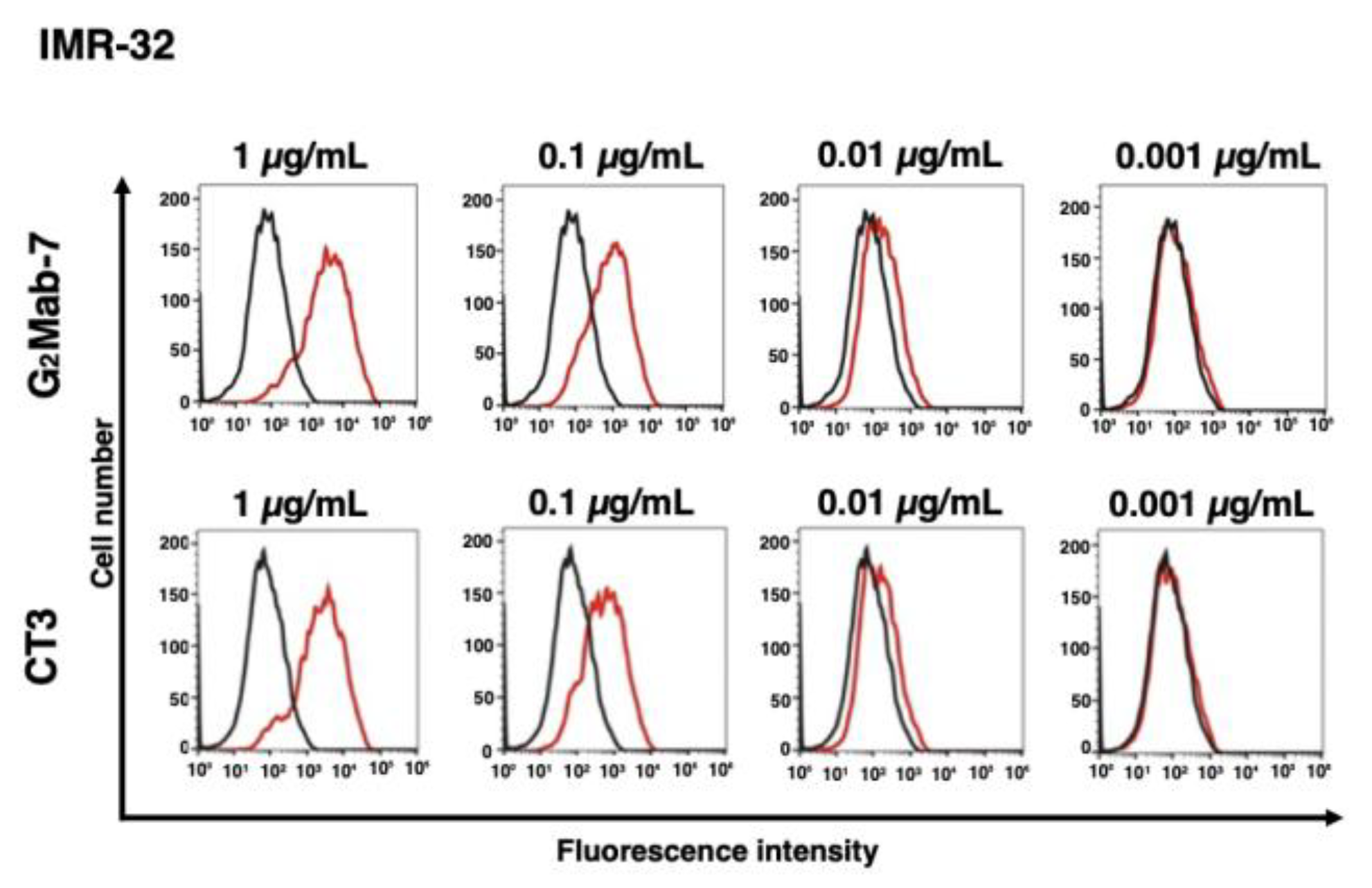
Figure 5.
Measurement of the binding affinity of G2Mab-7 and CT3. CHO/GPC2 was treated with serially diluted G2Mab-7 or CT3, followed by anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were analyzed using the SA3800 Cell Analyzer. The KD values were determined using GraphPad PRISM 6.
Figure 5.
Measurement of the binding affinity of G2Mab-7 and CT3. CHO/GPC2 was treated with serially diluted G2Mab-7 or CT3, followed by anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were analyzed using the SA3800 Cell Analyzer. The KD values were determined using GraphPad PRISM 6.
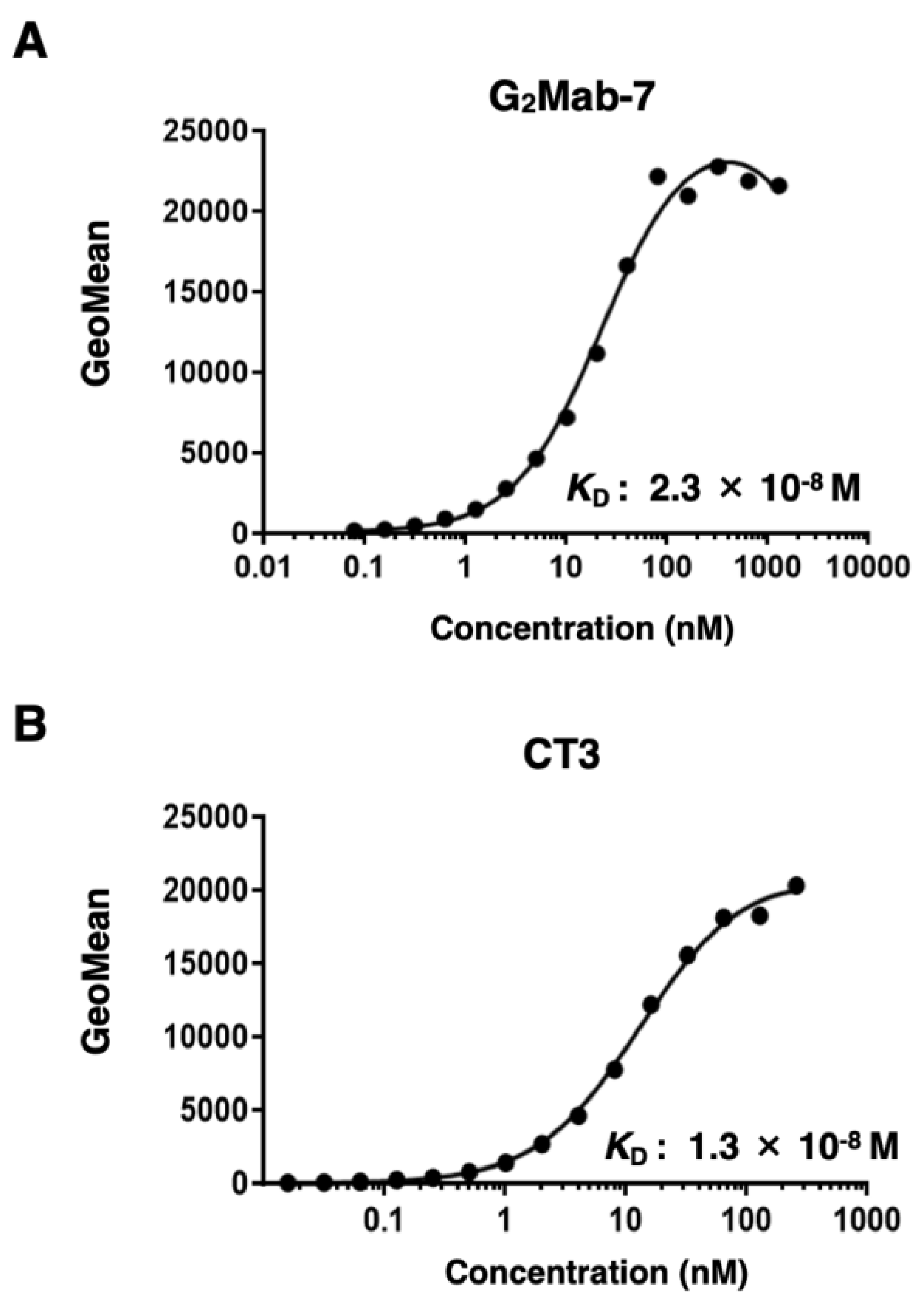
Figure 6.
Western blotting using G2Mab-7. The cell lysates of CHO-K1, CHO/GPC2, and IMR-32 were electrophoresed and transferred onto polyvinylidene difluoride membranes. The membranes were incubated with 1 μg/mL of G2Mab-7 (A), 1 μg/mL of CT3 (B), or 1 μg/mL of AC-15 (an anti-β-actin, C), followed by treatment with anti-mouse IgG conjugated with horseradish peroxidase.
Figure 6.
Western blotting using G2Mab-7. The cell lysates of CHO-K1, CHO/GPC2, and IMR-32 were electrophoresed and transferred onto polyvinylidene difluoride membranes. The membranes were incubated with 1 μg/mL of G2Mab-7 (A), 1 μg/mL of CT3 (B), or 1 μg/mL of AC-15 (an anti-β-actin, C), followed by treatment with anti-mouse IgG conjugated with horseradish peroxidase.
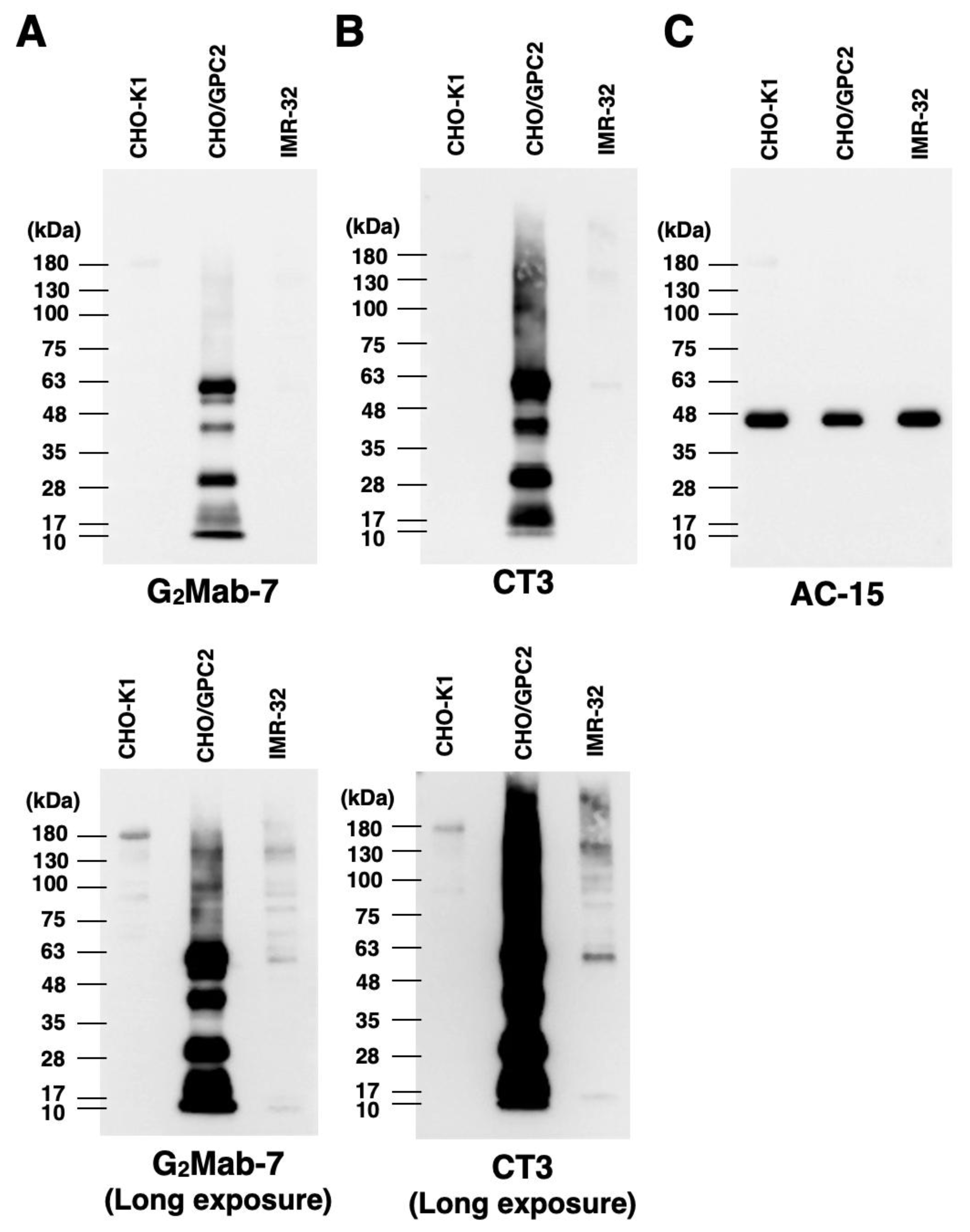
Figure 7.
Immunohistochemistry using G2Mab-7 and CT3 in formalin-fixed paraffin-embedded cell blocks. CHO/GPC2 and CHO-K1 sections were treated with 1 μg/mL of G2Mab-7 (A) or 0.1 μg/mL of CT3 (B). All procedures of immunohistochemistry were performed using VENTANA BenchMark ULTRA PLUS. Scale bar = 100 μm.
Figure 7.
Immunohistochemistry using G2Mab-7 and CT3 in formalin-fixed paraffin-embedded cell blocks. CHO/GPC2 and CHO-K1 sections were treated with 1 μg/mL of G2Mab-7 (A) or 0.1 μg/mL of CT3 (B). All procedures of immunohistochemistry were performed using VENTANA BenchMark ULTRA PLUS. Scale bar = 100 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



