1. Introduction
Dysentery, predominantly caused by pathogens such as
Shigella, is a gastrointestinal infectious disease that often occurs in regions with inadequate sanitary conditions and among individuals with compromised immunity, especially children and the elderly. It is primarily transmitted through the fecal-oral route [
1,
2]. The incidence of dysentery in China is notably higher compared to developed nations, with children aged 0 to 10 years constituting over 40% of the total cases. Health statistics indicate that large cities like Beijing and Wuhan in China have a higher incidence rate than remote areas, where sanitary conditions are considerably poorer. This discrepancy raises questions and leads us to consider other factors that might be contributing to the occurrence of dysentery [
3,
4,
5].
Previous studies have shown that pathogens can be transmitted naturally through direct or indirect contact between animals and humans. This includes animal scratches, contact with animal secretions, consumption of undercooked meat, and ingestion of food and water contaminated by animals [
6,
7]. However, urban dwellers have limited exposure to animals, with the exception of pets, which are often kept in close proximity. This proximity prompts us to consider whether pets could be linked to dysentery [
8,
9]. Our surveys have revealed that diarrhea in pets can be attributed to parasites, viruses, bacterial infections, and dietary factors. Although pets are vaccinated, bacterial vaccines are less common, suggesting that bacterial infections are the main cause of diarrhea in pets [
10], particularly in those with limited mobility, such as cats. Household cats seldom exchange microbes with the external environment, making their microbiota more susceptible to disruption and increasing the likelihood of diarrhea. However, this type of bacterial diarrhea is often acute and tends to resolve spontaneously or result in only soft stools, which may not be taken seriously by pet owners [
11,
12]. Nonetheless, pathogens can persist in cats and their feces. For instance,
Shigella can survive in feces for 11 days at room temperature and up to 90 days in colder conditions, posing a significant risk to cat owners [
13,
14].
Shigella is a strict human intestinal pathogen and does not typically cause symptoms in cats. When we vaccinate cats with Shigella, they do not exhibit symptoms of diarrhea, which is why it is not commonly associated dysentery with feline [
15,
16]. However, it is unknown whether cats can be carriers of
Shigella.
In light of these considerations, this study collected feces from cats with diarrhea and utilized C57BL/6J mice as a model. Through fecal microbiota transplantation, we evaluated the potential of cat feces to serve as a source of infection. Our findings indicated that Shigella and Escherichia coli had the highest detection rates in the feces, while Salmonella, often implicated in cat diarrhea, was not detected.
2. Materials and Methods
2.1. Experimental Animals and Design
We utilized six-week-old male C57BL/6J mice, sourced from Huafu Kang in Beijing, China. After a 7-day acclimation period in the animal housing facility, the mice were housed individually in ventilated cages, with a maximum of five mice per cage. They had ad libitum access to food and water, and were maintained in an environment with a temperature range of 18-23 °C, 40-60% humidity, and a 12-hour light/dark cycle. The mice selected for the study were of similar body weight to ensure consistency. The experimental protocols were approved by the Institutional Animal Care and Use Committee of China Agricultural University. The mice received a 7-day pre-treatment with a combination of quadruple antibiotic to fecal microbiota transplantation (ampicillin 1 mg/ml, metronidazole 1 mg/ml, kanamycin sulfate 1 mg/ml, and vancomycin 0.5 mg/ml).
2.2. Fecal Microbiota Transplantation (FMT)
At the China Agricultural University Veterinary Teaching Hospital, feces were collected from 27 cats with diarrhea and resuspended in physiological saline. The mixture was then filtered through gauze, and the filtrate was centrifuged at 1000×g for 3 minutes. The supernatant was subsequently preserved in glycerol and stored at -80 °C. Mice that had been treated with antibiotics were administered 0.2 ml of the liquid via gavage once a day for a period of 15 days.
2.3. Sample Collection
Mice were euthanized by cervical dislocation, and their livers, spleens, and kidneys were harvested and weighed. The colon was also harvested and its length measured. After removing the contents, the colon was divided into two sections; one part was placed in 4% paraformaldehyde, and the remaining part was quickly stored in a -80 °C freezer.
2.4. RNA Extraction and RT-PCR
Total RNA was extracted using TRIzol reagent according to the manufacturer’s instructions, and RNA was reverse transcribed into cDNA using a reverse transcription kit following the instructions provided. RT-PCR reactions were performed according to the manufacturer’s protocol. Primers were designed based on the principles of PCR primer design, with β-actin used as the internal reference gene. All primers were purchased from Shanghai Sangon Biotechnology Co., Ltd., and detailed information is listed in
Table 1.
2.5. Tissue Staining and Imaging
Samples in paraformaldehyde were embedded in paraffin and sectioned at 5μm. Standard HE staining was performed, and the stained tissues were imaged using a transmission microscope with ×10 or ×20 objectives. A TUNEL staining kit from Solaibao was used for the staining, and all steps were carried out strictly according to the kit instructions. For the immunofluorescence procedure, the dewaxed tissue sections underwent antigen retrieval to expose the antigenic sites. Subsequently, the sections were incubated with the primary antibody solution for 24 hours to allow for antibody binding to the target antigens. Afterward, the sections were washed to remove any unbound primary antibody. They were then incubated with the secondary antibody solution for 30 minutes, which binds to the primary antibody, enhancing the signal for detection. Following this, the sections were stained with DAPI for 5 minutes to visualize the nuclei. Finally, the sections were mounted using a fluorescent mounting medium to preserve the fluorescence and allow for imaging under a fluorescence microscope.
2.6. Microbial Community Analysis Based on 16S rRNA Sequencing
Fecal samples from healthy cats and cats with diarrhea were subjected to 16S high-throughput rDNA amplicon sequencing, commissioned to Shanghai Meiji Bio-Pharm Technology Co., Ltd. Operational taxonomic units (OTUs) were clustered and species classified based on effective data to form OTUs and other species classification registers for species richness. Based on the normalized OTU species richness, abundance, diversity indices, and other analyses were performed on the OTUs, and community structure was analyzed at the species level.
2.7. Agarose Gel Electrophoresis
DNA was extracted from feces using a kit. Specific primers were designed based on the sequence of the target DNA fragment. PCR was used to amplify the target DNA fragment with specific primers, and the products were electrophoresed in a 10% agarose gel to ensure effective separation of the DNA molecules. The gels were then visualized under ultraviolet light.
2.8. Statistical Analysis
All values are presented as the mean ± SEM, and Student’s t-test was used to compare differences between groups. All statistical analyses were performed using SPSS 9.0 software, and graphs were generated using GraphPad Prism 9.3 software. A p-value of less than 0.05 was considered to indicate a significant difference.
3. Results
3.1. The Impact of FMT on Mice Growth Performance and Organ Levels
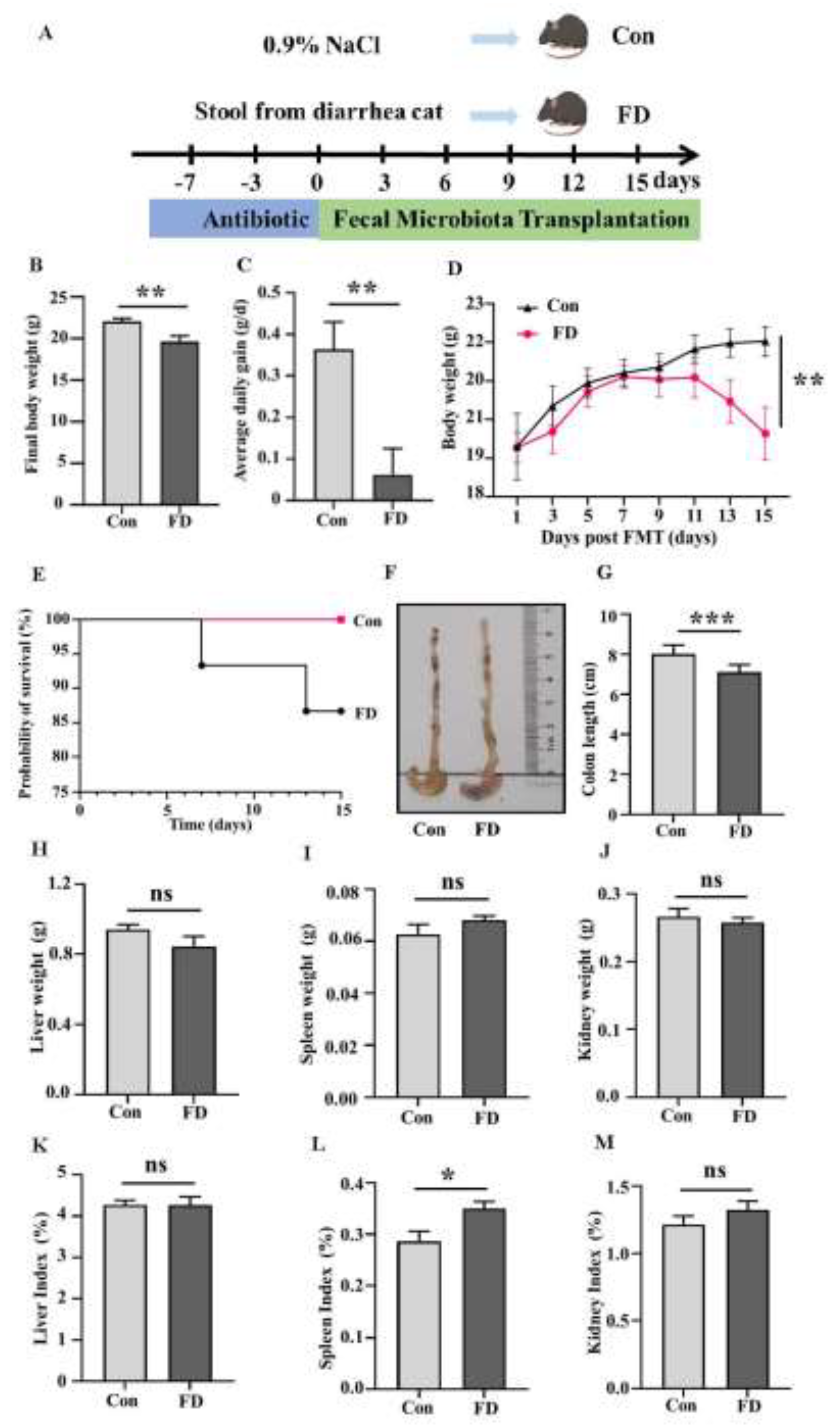
The experimental animal design is shown in
Figure 1A. When we transplanted microbes from the feces of cats with diarrhea into germ-free mice, their final body weight (
Figure 1B) and average daily weight gain (
Figure 1C) were significantly lower than those of the Con group at the end of the experiment. The body weight of mice in the FD group showed a significant difference from that in the Con group on day 9 (
Figure 1D). At the end of the experiment, we found that the microorganism from cats with diarrhea significantly affected the survival rate of the mice (
Figure 1E). Subsequently, we observed the colon, and after FMT, the mice’s colonic and cecal contents were not well-filled (
Figure 1F), and the length of the colon was significantly reduced (
Figure 1G). There were no significant changes in the weights of the liver, spleen, and kidneys (
Figure 1H-J). The liver index and kidney index showed no significant change (
Figure 1K, M), the spleen index increased significantly (
Figure 1L).
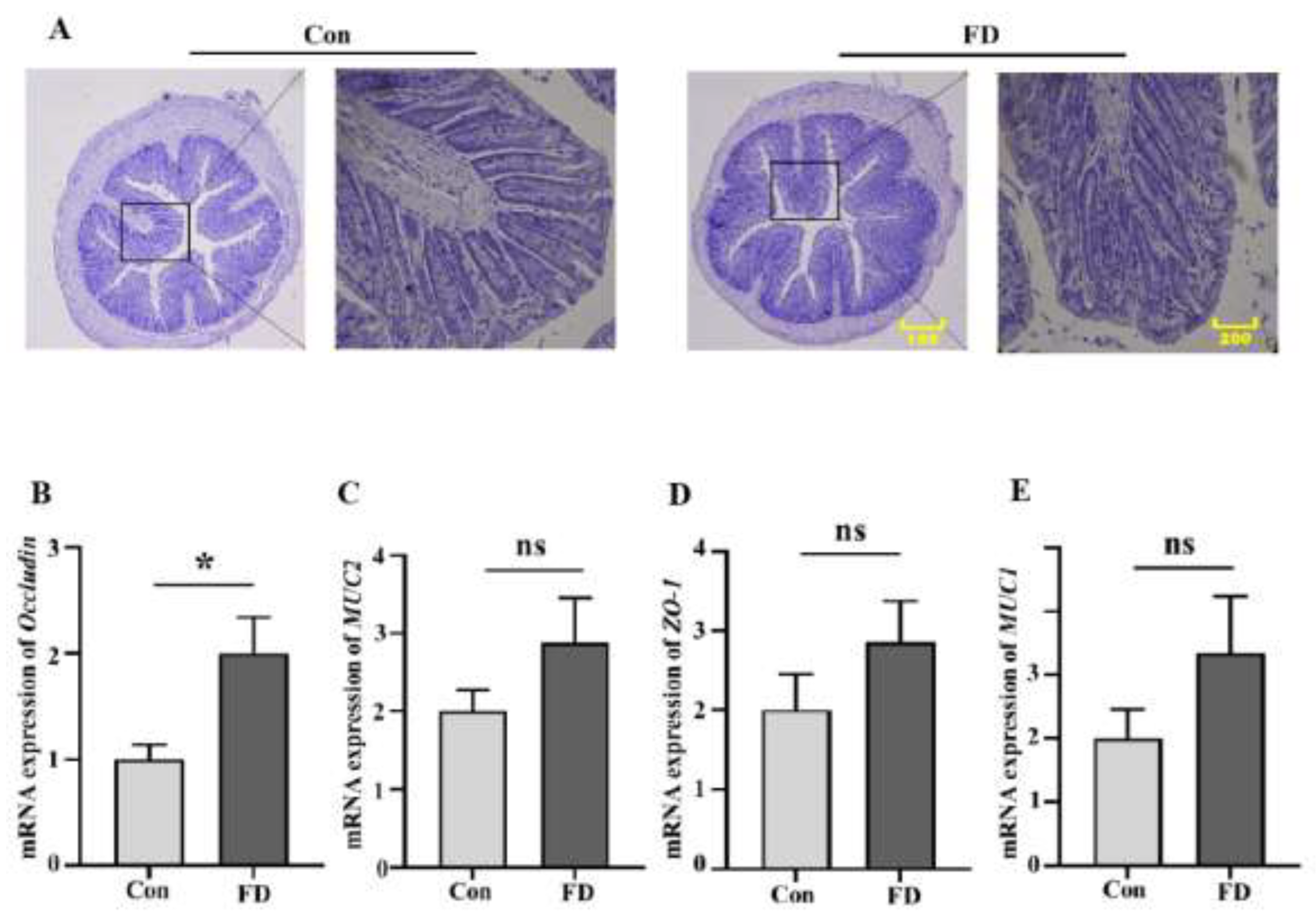
3.2. The Impact of FMT on the Intestinal Barrier of Mice
According to H&E staining, we found obvious lesions in the colon of mice after FMT (
Figure 2A). We then detected the mRNA expression of the intestinal barrier in the mice, and the expression of the Occludin gene increased significantly (
Figure 2B), while the expression of ZO-1, MUC1, and MUC2 genes increased to varying degrees (
Figure 2C-E).
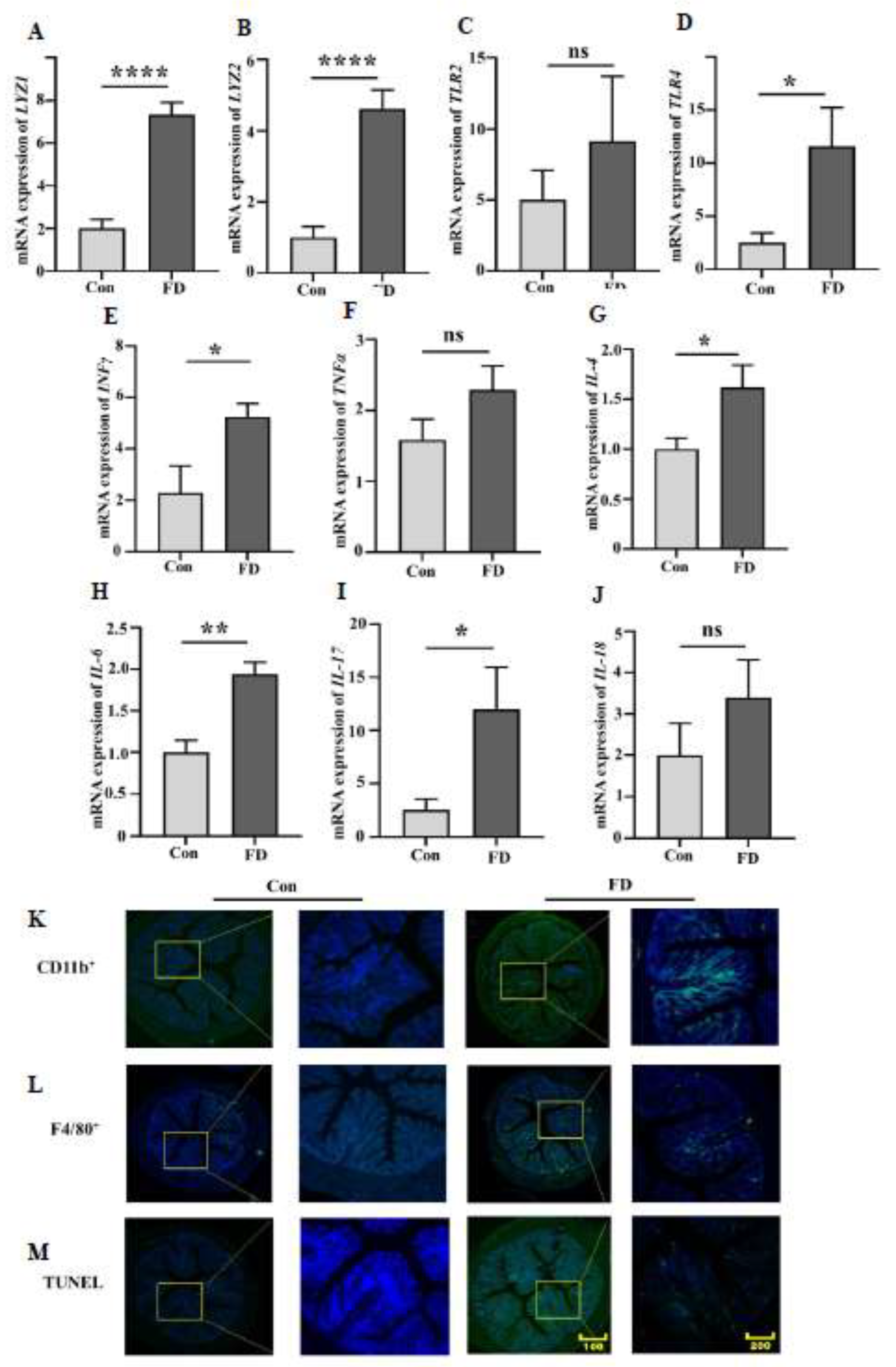
3.3. FMT Leads to Macrophage Aggregation and Increased Inflammatory Response in the Colon of Mice
When the body is invaded by bacteria, the number of lysozymes in the intestine increases significantly. The expression of LYZ1 and LYZ2 genes in mice after FMT increased significantly (
Figure 3A, B). Bacterial infection leads to an increase in the content of lipopolysaccharides (LPS) and Peptidoglycan (PG) in the intestine, and TLR2 and TLR4 are the receptors that intestinal cells use to sense LPS and PG. The expression of TLR2 and TLR4 genes in the colon of mice after fecal microbiota transplantation increased significantly (
Figure 3C, D). Macrophages, upon stimulation by LPS, also aggregate in the colon (
Figure 3K, L), and the aggregated macrophages release more inflammatory factors. Therefore, we found that the expression of inflammatory factors in the colon of mice increased to varying degrees (
Figure 3E-J). Inflammatory factors, in turn, promote apoptosis, and the number of apoptotic cells in the colon of mice after FMT increased significantly (
Figure 3N).
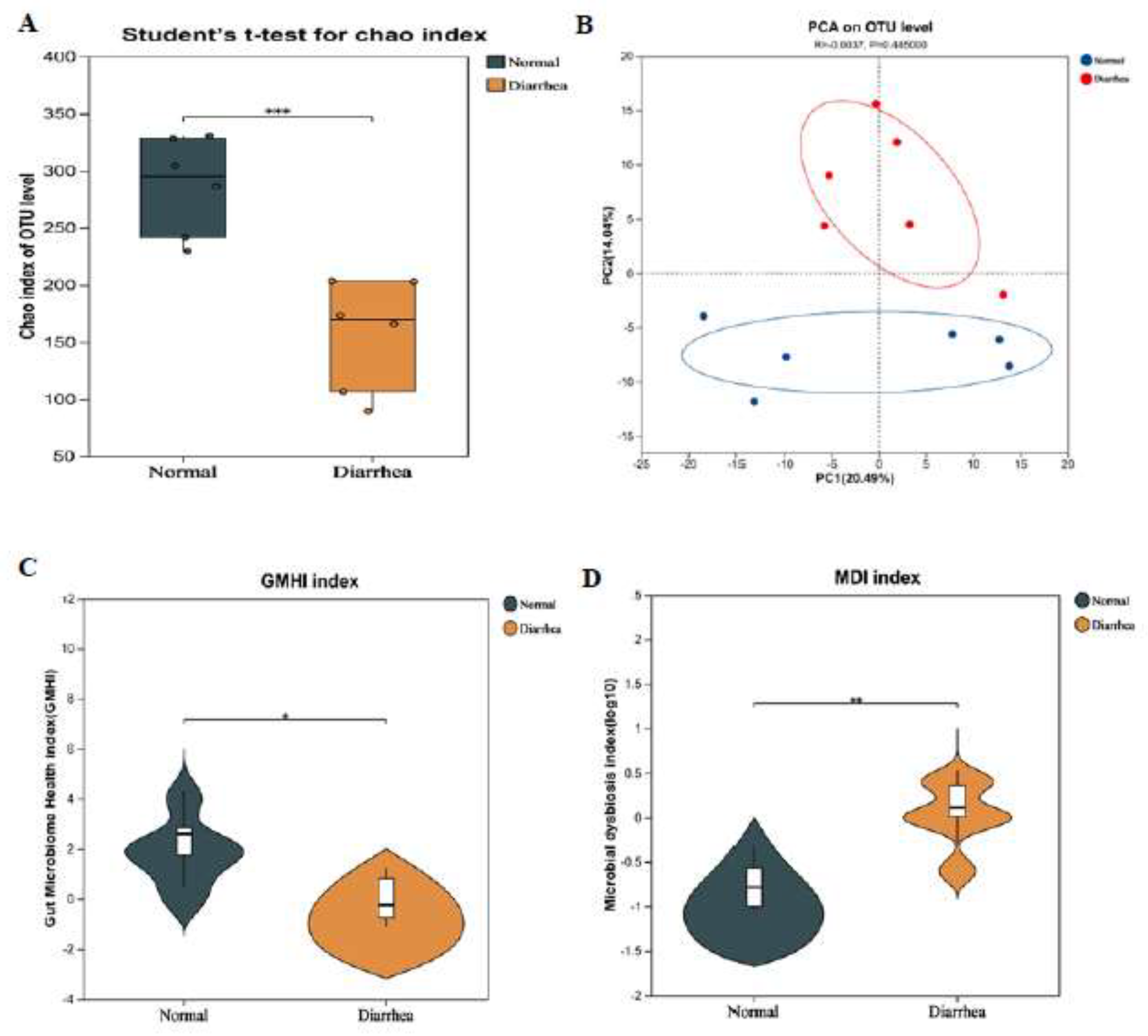
3.4. Differences in Microbiota Between the Feces of Healthy Cats and Cats with Diarrhea
The Chao index is mainly used to assess the richness of items in a sample. The Chao index of microbial flora in the feces of healthy cats was significantly higher than that in the feces of cats with diarrhea (
Figure 4A). In principal component analysis (PCA), the community structure of the fecal microbiota of healthy cats and cats with diarrhea was significantly different (
Figure 4B). The health index (GHMI) of the gut microbiota of healthy cats was significantly higher than that of cats with diarrhea (
Figure 4C), while the microbial dysbiosis index (MDI) showed the opposite trend (
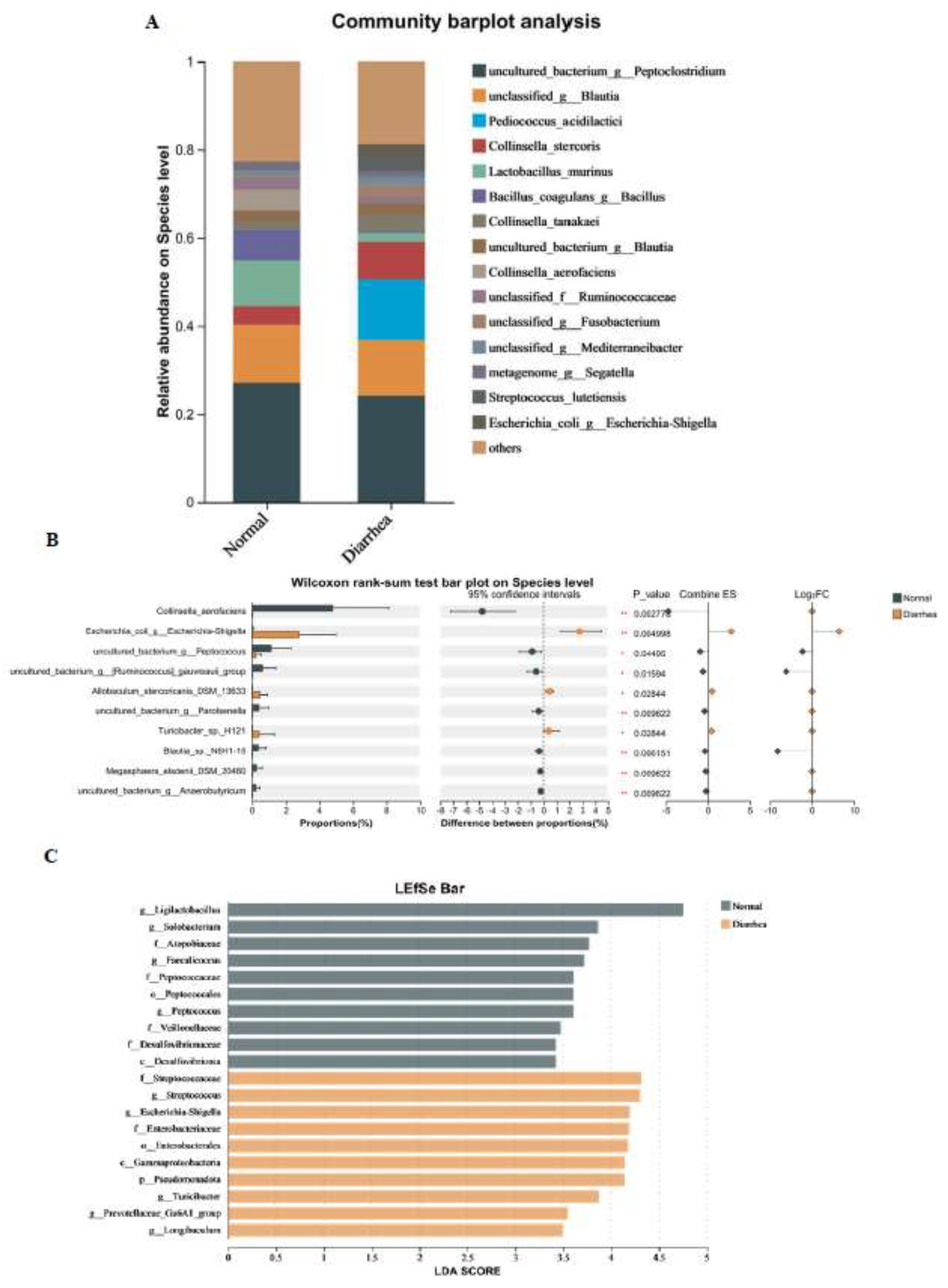
Figure 4D). At the species level, the gut microbiota of both healthy cats and cats with diarrhea were mainly dominated by
Peptoclostridium and
Blautia, with
Lactobacillus murinus and
Bacillus coagulans also playing a major role in the gut of healthy cats, while
Collinsella stercoris and
Shigella played a major role in the gut of cats with diarrhea (
Figure 5A). In LEfSe analysis,
Ligilactobacillus played a major role in healthy cats, while
Streptococcaceae,
Streptococcus, and
Escherichia Shigella played a major role in cats with diarrhea (
Figure 5B). The Wilcoxon rank sum test showed that the content of
Shigella in feces of cats with diarrhea was significantly higher than that of healthy cats (
Figure 5C). We can infer that the microbes in the gut of healthy cats may enhance intestinal resistance due to the high content of lactic acid bacteria, while the increased content of
Shigella in the gut of cats with diarrhea may directly or indirectly damage the intestinal health of cats.
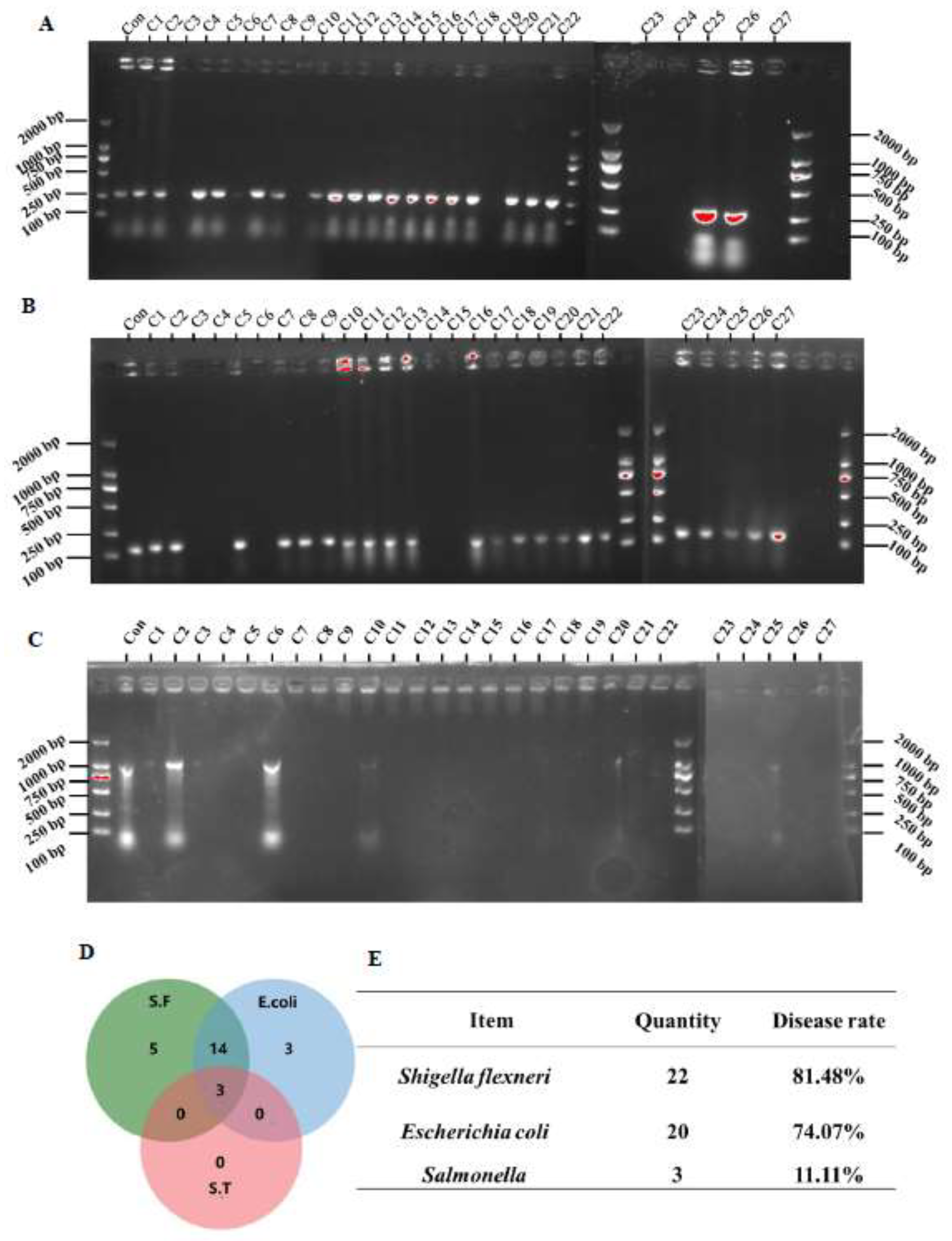
3.5. Detection Rate of Pathogenic Bacteria in Feces
Subsequently, we used specific primers of
Shigella,
Escherichia coli, and
Salmonella to detect whether these microbes were present in the feces.
Escherichia coli was detected in the feces of 20 cats with diarrhea (
Figure 6A).
Shigella was detected in the feces of 22 cats with diarrhea (
Figure 6B), and
Salmonella was detected in the feces of only 3 cats (
Figure 6C). The Venn diagram showed that the feces of 3 cats with diarrhea were simultaneously positive for these three bacteria, and the feces of 14 cats were simultaneously positive for
Shigella and
Escherichia coli (
Figure 6D). We then calculated the detection rates of the three bacteria, which were 81.48% for
Shigella, 74.07% for
Escherichia coli, and 11.11% for
Salmonella (
Figure 6E).
Figure 6.
Detection rates of pathogenic bacteria in the feces of cats with diarrhea. (A-C) Detection levels of Shigella, Escherichia coli, and Salmonella in feces (n=27); (D) Venn diagram distribution of Shigella, Escherichia coli, and Salmonella; (E) Detection rates of Shigella, Escherichia coli, and Salmonella in feces.
Figure 6.
Detection rates of pathogenic bacteria in the feces of cats with diarrhea. (A-C) Detection levels of Shigella, Escherichia coli, and Salmonella in feces (n=27); (D) Venn diagram distribution of Shigella, Escherichia coli, and Salmonella; (E) Detection rates of Shigella, Escherichia coli, and Salmonella in feces.
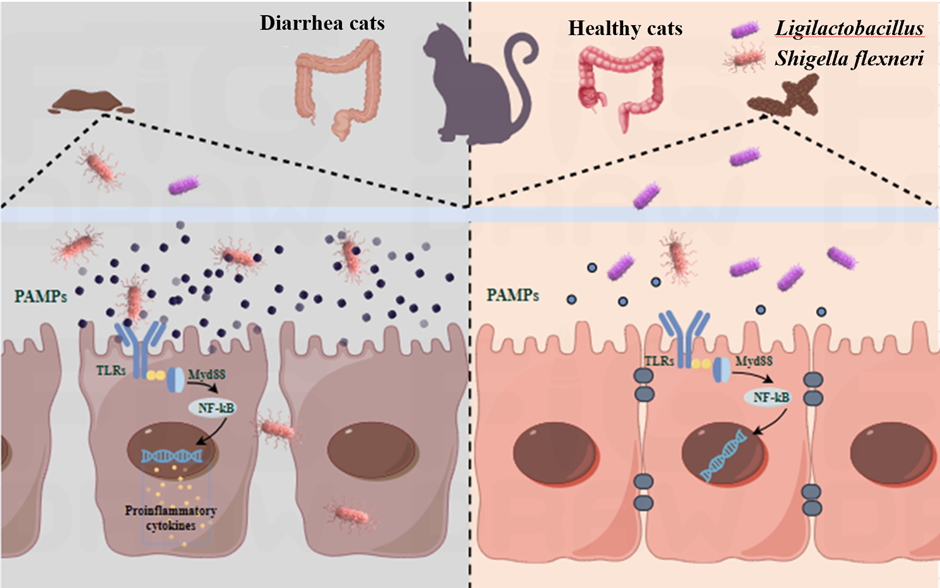
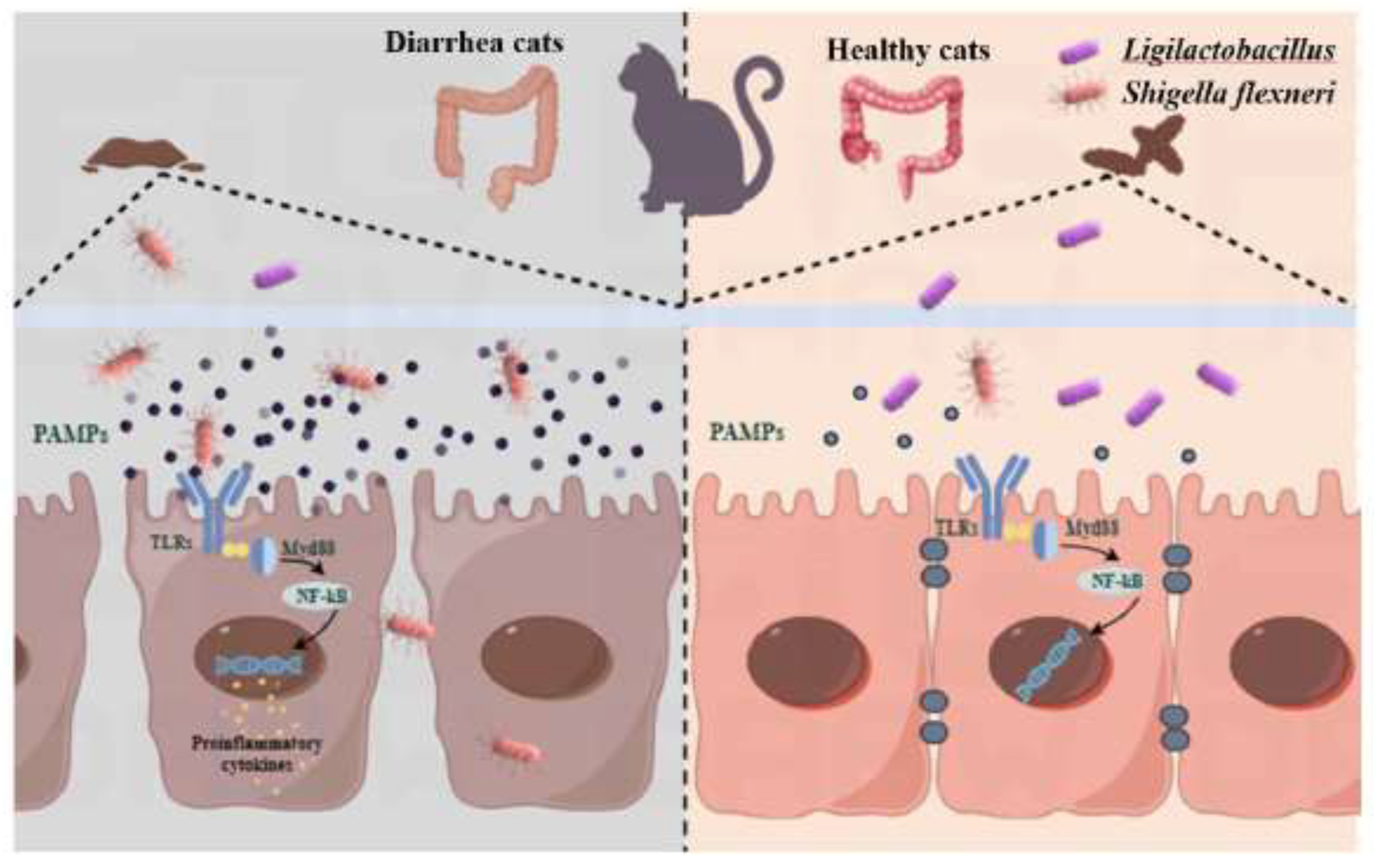
Figure 7.
Ligilactobacillus can prevent the spread of Shigella, and reducing the occurrence of dysentery.
Figure 7.
Ligilactobacillus can prevent the spread of Shigella, and reducing the occurrence of dysentery.
4. Discussion
Fecal Microbiota Transplantation (FMT) is an innovative therapeutic approach that entails transferring the gut microbiota from a healthy donor to a recipient in order to restore microbial equilibrium. Over recent years, FMT has demonstrated considerable promise in addressing a spectrum of medical conditions. Notably, in cases of recurrent Clostridium difficile infections, FMT has achieved cure rates of up to 90%, markedly surpassing the efficacy of vancomycin. Additionally, FMT has proven beneficial in managing inflammatory bowel disease (IBD) and irritable bowel syndrome (IBS), with a notable increase in
Faecalibacterium prausnitzii observed within the recipient’s gut following the procedure [
17,
18,
19]. FMT can effectively transfer microbes from the donor to the recipient. Cats have a digestive structure that is quite different from that of omnivorous animals like dogs. As obligate carnivores, cats have a gut structure and microbiome that are uniquely adapted to their high-protein, high-fat dietary needs. This adaptation results in a considerably lower ratio of intestinal length to body length in cats compared to omnivores, with a less developed cecum and a comparatively simple microbial community in the colon [
20]. Therefore, we infer that feline intestinal health issues may not be directly related to the microbes in their gut, which has led to less focus on changes in the feline gut microbiome. However, in domestic settings, cats are frequently treated with deworming medications and vaccinations, which primarily safeguard against parasitic and viral infections and offer minimal protection against bacterial infections. Although the gut microbial community in cats is relatively straightforward, the impact of microbial structural changes on feline intestinal health remains to be fully understood [
13,
21]. In this experiment, microbes from the feces of cats with diarrhea were transplanted into germ-free mice, and it was found that the mice’s body weight significantly decreased, and the splenic index increased. Bacterial infections in mice typically lead to a substantial decrease in body weight, and the spleen, being the body’s largest secondary lymphoid organ and central to immune responses, tends to enlarge noticeably as a result of immune cell proliferation [
7,
22,
23]. Similar phenomena were observed in the mice after FMT, proving that the gut of cats with diarrhea contains certain pathogenic bacteria that cause pathological changes in mice.
The cecum and colon serve as the primary habitats for microbial aggregation. Upon bacterial infection, the colon’s length is typically shortened, its contents diminished, and the intestinal barrier compromised [
24]. In our experiment, we observed analogous alterations in the mice, accompanied by signs of inflammatory infiltration within their colons. The intestinal barrier’s destruction prompted an upregulation in the expression of genes associated with intestinal integrity, an adaptive response aimed at mending the damage inflicted by bacteria. Lysozyme is mainly secreted by Paneth cells in the gut and is an important antimicrobial peptide. It plays a pivotal role in augmenting the gut’s defenses and preserving microbial equilibrium when faced with overgrowth of certain pathogenic bacteria [
25,
26]. Post-FMT, there was a marked escalation in lysozyme expression within the colonic tissue of the mice, suggesting that the fecal matter of cats with diarrhea is teeming with pathogenic bacteria. These bacteria not only undermine the colon’s health but also trigger an increase in lysozyme production, indicative of the body’s struggle to combat the microbial onslaught.
TLR4 mainly recognizes LPS from Gram-negative bacteria, and TLR2 mainly recognizes components such as peptidoglycan from Gram-positive bacteria, thereby initiating an immune response against pathogenic bacteria. These recognition events trigger an immune response against pathogenic bacteria. In prior studies, invasion of the body by pathogenic bacteria led to the activation of MyD88-dependent signaling cascades, including NF-κB and MAPK pathways, within the gut-associated TLR4 and TLR2 receptors upon their binding with LPS and peptidoglycan. This activation process fosters inflammatory responses [
24,
27,
28]. In our experiment, the expression levels of both TLR4 and TLR2 were observed to increase, with TLR4 showing a particularly significant rise. These receptors also serve as principal sensors for macrophages. Once there, macrophages, stimulated by LPS, differentiate into M1-type inflammatory cells that further release pro-inflammatory mediators. These mediators not only initiate defensive mechanisms but also recruit other immune cells, facilitating the swift assembly of neutrophils and their subsequent removal through apoptosis [
29,
30,
31,
32]. In this experiment, macrophages in the colon significantly aggregated, inflammatory factors were elevated to varying degrees, and the aggregation of apoptotic proteins also significantly increased. LPS is the main component of the outer membrane of Gram-negative bacteria and is a potent immune activator. TLR4 is the main recognition receptor for LPS, indicating that the main pathogenic bacteria in the feces of cats are Gram-negative bacteria, which are the main cause of infection in mice.
In the results above, it was found that there may be a Gram-negative bacterium in the feces of cats with diarrhea, which may have certain infectivity. Subsequently, we examined the microbial composition in the feces of healthy cats and cats with diarrhea and found that the only microorganism detected was
Shigella, which is pathogenic and a Gram-negative bacterium.
Shigella infection is known to induce bacterial dysentery, primarily harming the colonic epithelium. This not only compromises the intestinal epithelial cell barrier but also triggers inflammation. In addition,
Shigella also induces pyroptosis in macrophages and releases the cytokine IL-1β [
33,
34,
35,
36]. This is consistent with the results observed in FMT mice. Based on these findings, we deduce that Shigella is likely responsible for the infections observed in mice. Subsequently, we employed
Shigella-specific primers to examine the prevalence of
Shigella in fecal samples and discovered its frequent presence in the feces of cats with diarrhea. Prior research has predominantly associated
Shigella with causing bacterial dysentery in humans, with scarce accounts of animals serving as hosts. When cats are exposed to
Shigella, it may not always elicit adverse reactions, and they can often recover from diarrhea spontaneously without medical treatment. Nevertheless, the possibility of cats being asymptomatic carriers remains uncertain [
13,
37]. Our experiment revealed that
Shigella is commonly found in the feces of cats with diarrhea, suggesting that while
Shigella may not cause disease in cats, they could act as potential carriers. During episodes of diarrhea in cats, which can be triggered by various factors,
Shigella may contribute to worsening the condition. This insight raises concerns that feline feces could be a potential reservoir for
Shigella infection, highlighting the need for vigilance in this regard.
5. Conclusion
In summary, our research results indicate that although the gut microbiota structure in cats is relatively simple, changes in microbial composition can also contribute to feline diarrhea. The microbes in the diarrheal feces can disrupt the intestinal barrier, increase pro-inflammatory factors, and lead to the aggregation of macrophages. Among the pathogens identified, Shigella stands out as a likely culprit. While Shigella may not be the primary cause of feline diarrhea, it represents a possible vector for the transmission of this pathogen. Our findings enhance the understanding of the various factors that may lead to dysentery in children. In future research, our research will concentrate on exploring the relationship between pets and the development of dysentery. This will facilitate the development of additional preventive strategies, help to mitigate potential risks, and ultimately contribute to a decrease in the prevalence of dysentery.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Jing Wang: Writing – review & editing, Writing – original draft, Investigation, Data curation, Conceptualization, Funding acquisition. Xinhui Zhang: Investigation, Conceptualization. Xinran Song, Yan He, Wuying Lang and Hanlu Liu: review & editing. Xinyu Zhang: Conceptualization, Supervision, Writing.
Ethics approval
All animals were purchased from Hufukang (Beijing, China) and this study was approved by the ethical committee of the China Agricultural University (No. AW02504202-1-11).
Data availability statement
Acknowledgments
This work was supported by grants from Shaanxi Provincial Department of Education Special Scientific Research Program in 2024 (24JK0425); Shangluo University Natural Science Research Project (23SKY008).
Declaration of competing interest
The authors declare no conflicts of interest.
Availability of data and materials
All data needed to assess the conclusions in this study are provided in the manuscript and/or the
Supplementary Materials.
Abbreviations
Hematoxylin-eosin, H&E; Interleukin, IL; Interferon-gamma, INF-γ; Toll-like receptor, TLR; Tumor Necrosis Factor-alpha, TNF-α; Pathogen-Associated Molecular Patterns, PAMPs;
References
- Collaborators GBDDD: Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: a systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect Dis 2017, 17(9):909–948.
- Kotloff KL, Riddle MS, Platts-Mills JA, Pavlinac P, Zaidi AKM: Shigellosis. The Lancet 2018, 391(10122):801–812.
- Qiu S, Xu X, Yang C, Wang J, Liang B, Li P, Li H, Yi S, Liu H, Cui X et al: Shift in serotype distribution of Shigella species in China, 2003–2013. Clinical Microbiology and Infection 2015, 21(3):252.e255–252.e258.
- Chang Z, Zhang J, Ran L, Sun J, Liu F, Luo L, Zeng L, Wang L, Li Z, Yu H et al: The changing epidemiology of bacillary dysentery and characteristics of antimicrobial resistance of Shigella isolated in China from 2004-2014. BMC Infect Dis 2016, 16(1):685. [CrossRef]
- Chang Z, Lu S, Chen L, Jin Q, Yang J: Causative species and serotypes of shigellosis in mainland China: systematic review and meta-analysis. PLoS One 2012, 7(12):e52515. [CrossRef]
- Rahman MT, Sobur MA, Islam MS, Ievy S, Hossain MJ, El Zowalaty ME, Rahman AT, Ashour HM: Zoonotic Diseases: Etiology, Impact, and Control. Microorganisms 2020, 8(9).
- Lambertini E, Buchanan RL, Narrod C, Pradhan AK: Transmission of Bacterial Zoonotic Pathogens between Pets and Humans: The Role of Pet Food. Crit Rev Food Sci Nutr 2016, 56(3):364–418. [CrossRef]
- Damborg P, Broens EM, Chomel BB, Guenther S, Pasmans F, Wagenaar JA, Weese JS, Wieler LH, Windahl U, Vanrompay D et al: Bacterial Zoonoses Transmitted by Household Pets: State-of-the-Art and Future Perspectives for Targeted Research and Policy Actions. J Comp Pathol 2016, 155(1 Suppl 1):S27–40.
- Zhang T, Nickerson R, Zhang W, Peng X, Shang Y, Zhou Y, Luo Q, Wen G, Cheng Z: The impacts of animal agriculture on One Health-Bacterial zoonosis, antimicrobial resistance, and beyond. One Health 2024, 18:100748. [CrossRef]
- Sung CH, Marsilio S, Chow B, Zornow KA, Slovak JE, Pilla R, Lidbury JA, Steiner JM, Park SY, Hong MP et al: Dysbiosis index to evaluate the fecal microbiota in healthy cats and cats with chronic enteropathies. J Feline Med Surg 2022, 24(6):e1–e12. [CrossRef]
- Wang F, Mei X, Wang Q, Zhao P, Zhou Y, Tang L, Wang B, Xu S, Li X, Jin Q et al: Compound Bacillus alleviates diarrhea by regulating gut microbes, metabolites, and inflammatory responses in pet cats. Anim Microbiome 2023, 5(1):49. [CrossRef]
- Smith KA, Kruth S, Hammermueller J, Gyles C, Wilson JB: A case-control study of verocytotoxigenic Escherichia coli infection in cats with diarrhea. Can J Vet Res 1998, 62(2):87–92.
- Bai H, Liu T, Wang S, Gong W, Shen L, Zhang S, Wang Z: Identification of Gut Microbiome and Metabolites Associated with Acute Diarrhea in Cats. Microbiol Spectr 2023, 11(4):e0059023. [CrossRef]
- Jang J, Hur HG, Sadowsky MJ, Byappanahalli MN, Yan T, Ishii S: Environmental Escherichia coli: ecology and public health implications-a review. J Appl Microbiol 2017, 123(3):570–581. [CrossRef]
- Kim Y, Carrai M, Leung MHY, Chin J, Li J, Lee PKH, Beatty JA, Pfeiffer DU, Barrs VR: Dysbiosis of the Urinary Bladder Microbiome in Cats with Chronic Kidney Disease. mSystems 2021, 6(4):e0051021. [CrossRef]
- Santos JPF, Aquino AA, Gloria MBA, Avila-Campos MJ, Oba PM, Santos KM, Vendramini THA, Carciofi AC, Junior AR, Brunetto MA: Effects of dietary yeast cell wall on faecal bacteria and fermentation products in adult cats. J Anim Physiol Anim Nutr (Berl) 2018, 102(4):1091–1101. [CrossRef]
- Khoruts A, Sadowsky MJ: Understanding the mechanisms of faecal microbiota transplantation. Nat Rev Gastroenterol Hepatol 2016, 13(9):508–516. [CrossRef]
- Kump PK, Grochenig HP, Lackner S, Trajanoski S, Reicht G, Hoffmann KM, Deutschmann A, Wenzl HH, Petritsch W, Krejs GJ et al: Alteration of intestinal dysbiosis by fecal microbiota transplantation does not induce remission in patients with chronic active ulcerative colitis. Inflamm Bowel Dis 2013, 19(10):2155–2165. [CrossRef]
- Khoruts A, Dicksved J, Jansson JK, Sadowsky MJ: Changes in the composition of the human fecal microbiome after bacteriotherapy for recurrent Clostridium difficile-associated diarrhea. J Clin Gastroenterol 2010, 44(5):354–360. [CrossRef]
- Pilla R, Suchodolski JS: The Gut Microbiome of Dogs and Cats, and the Influence of Diet. Vet Clin North Am Small Anim Pract 2021, 51(3):605–621. [CrossRef]
- Honneffer JB, Minamoto Y, Suchodolski JS: Microbiota alterations in acute and chronic gastrointestinal inflammation of cats and dogs. World J Gastroenterol 2014, 20(44):16489–16497. [CrossRef]
- Salaheen S, Jaiswal E, Joo J, Peng M, Ho R, D OC, Adlerz K, Aranda-Espinoza JH, Biswas D: Bioactive extracts from berry byproducts on the pathogenicity of Salmonella Typhimurium. Int J Food Microbiol 2016, 237:128–135. [CrossRef]
- Peng M, Biswas D: Short chain and polyunsaturated fatty acids in host gut health and foodborne bacterial pathogen inhibition. Crit Rev Food Sci Nutr 2017, 57(18):3987–4002. [CrossRef]
- Duan Y, Prasad R, Feng D, Beli E, Li Calzi S, Longhini ALF, Lamendella R, Floyd JL, Dupont M, Noothi SK et al: Bone Marrow-Derived Cells Restore Functional Integrity of the Gut Epithelial and Vascular Barriers in a Model of Diabetes and ACE2 Deficiency. Circ Res 2019, 125(11):969–988. [CrossRef]
- Kaser A, Blumberg RS: The road to Crohn’s disease. Science 2017, 357(6355):976–977.
- Zhang Q, Pan Y, Yan R, Zeng B, Wang H, Zhang X, Li W, Wei H, Liu Z: Commensal bacteria direct selective cargo sorting to promote symbiosis. Nat Immunol 2015, 16(9):918–926. [CrossRef]
- Kim SJ, Howe C, Mitchell J, Choo J, Powers A, Oikonomopoulos A, Pothoulakis C, Hommes DW, Im E, Rhee SH: Autotaxin loss accelerates intestinal inflammation by suppressing TLR4-mediated immune responses. EMBO Rep 2020, 21(10):e49332. [CrossRef]
- Liu Y, Yang M, Tang L, Wang F, Huang S, Liu S, Lei Y, Wang S, Xie Z, Wang W et al: TLR4 regulates RORgammat(+) regulatory T-cell responses and susceptibility to colon inflammation through interaction with Akkermansia muciniphila. Microbiome 2022, 10(1):98. [CrossRef]
- Sharma G, Sharma A, Kim I, Cha DG, Kim S, Park ES, Noh JG, Lee J, Ku JH, Choi YH et al: A dietary commensal microbe enhances antitumor immunity by activating tumor macrophages to sequester iron. Nat Immunol 2024, 25(5):790–801. [CrossRef]
- Fan L, Xu C, Ge Q, Lin Y, Wong CC, Qi Y, Ye B, Lian Q, Zhuo W, Si J et al: A. Muciniphila Suppresses Colorectal Tumorigenesis by Inducing TLR2/NLRP3-Mediated M1-Like TAMs. Cancer Immunol Res 2021, 9(10):1111–1124. [CrossRef]
- Carpino G, Del Ben M, Pastori D, Carnevale R, Baratta F, Overi D, Francis H, Cardinale V, Onori P, Safarikia S et al: Increased Liver Localization of Lipopolysaccharides in Human and Experimental NAFLD. Hepatology 2020, 72(2):470–485. [CrossRef]
- Li T, Li Q, Liu S, Cao J, Mei J, Gong J, Chen J, Wang X, Zhang R, Li X et al: Targeted V-type peptide-decorated nanoparticles prevent colitis by inhibiting endosomal TLR signaling and modulating intestinal macrophage polarization. Biomaterials 2025, 314:122843. [CrossRef]
- Zychlinsky A, Prevost MC, Sansonetti PJ: Shigella flexneri induces apoptosis in infected macrophages. Nature 1992, 358(6382):167–169. [CrossRef]
- Li LH, Chen TL, Chiu HW, Hsu CH, Wang CC, Tai TT, Ju TC, Chen FH, Chernikov OV, Tsai WC et al: Critical Role for the NLRP3 Inflammasome in Mediating IL-1beta Production in Shigella sonnei-Infected Macrophages. Front Immunol 2020, 11:1115. [CrossRef]
- Hua KF, Lin YB, Chiu HW, Wong WT, Ka SM, Wu CH, Lin WY, Wang CC, Hsu CH, Hsu HT et al: Cinnamaldehyde inhibits the NLRP3 inflammasome by preserving mitochondrial integrity and augmenting autophagy in Shigella sonnei-infected macrophages. J Inflamm (Lond) 2024, 21(1):18. [CrossRef]
- Koestler BJ, Ward CM, Fisher CR, Rajan A, Maresso AW, Payne SM: Human Intestinal Enteroids as a Model System of Shigella Pathogenesis. Infect Immun 2019, 87(4). [CrossRef]
- Yu YY, Liang L, Xiao HB: Comparative study on fecal flora and blood biochemical indexes in normal and diarrhea British Shorthair cats. Arch Microbiol 2022, 204(5):257. [CrossRef]
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).