Submitted:
01 February 2026
Posted:
03 February 2026
You are already at the latest version
Abstract
Oxidative stress arises from an imbalance in redox homeostasis, leading to the accumulation of reactive oxygen species (ROS). This condition is associated with premature aging, as well as the progression of several chronic non-communicable diseases. Among the natural products, geopropolis stands out as a source of molecules with different biological properties. Despite reports of its therapeutic potential, data on effects on biomolecules and lifespan remains unexplored. In this context, we investigated the effects of hydroethanolic extracts of geopropolis from Melipona orbignyi (HGMO) and Melipona quadrifasciata anthidioides (HGMQ) on in vitro and in vivo protection against oxidative stress, as well as their toxicity and effects on lifespan. Firstly, we assessed the effect on protein integrity under AAPH-induced oxidative stress and on DNA stability following exposure to hydrogen peroxide and UV radiation. Furthermore, we evaluated the extracts toxicity, protection against juglone-induced oxidative stress and thermal stress, and effects on longevity in a Caenorhabditis elegans preclinical model. Our results demonstrated that both extracts reduced protein oxidation and DNA fragmentation. No signs of toxicity of the extracts were observed in the C. elegans. In the in vivo assays, both HGMO and HGMQ enhanced resistance to oxidative and thermal stress, and consistently increased the lifespan of nematodes across all tested concentrations. These findings indicate that geopropolis extracts from M. orbignyi and M. q. anthidioides protect biomolecules from oxidative damage, enhance stress resistance, and extend C. elegans lifespan. The characterization of these effects supports the development of geopropolis-based products for preventing or treating oxidative stress-related diseases
Keywords:
bee product
; biotechnological product
; protective
; oxidative stress
; lifespan
; Caenorhabditis elegans
1. Introduction
Reactive oxygen species (ROS) are highly reactive molecules produced naturally as byproducts of cellular metabolism. At physiological levels, ROS act as beneficial signaling molecules that regulate cellular processes; however, as their concentration increases, they may trigger adaptive responses and ultimately exert deleterious effects [1]. The balance between the generation and neutralization of ROS in the organism is responsible for ensuring cellular redox homeostasis [2]. However, when the production of ROS exceeds the capacity of the endogenous antioxidant defense system, a redox imbalance occurs, triggering oxidative stress.
Oxidative stress is defined by elevated levels of ROS in the body, loss of redox homeostasis, and cytotoxic effects on cellular and tissue function [3,4]. At the molecular level, oxidative stress causes damage to lipids, proteins, and nucleic acids, events associated with the acceleration of cellular aging and the onset of chronic noncommunicable diseases (CNCDs) [5].
CNCDs include cardiovascular, respiratory, diabetes, neurodegenerative, and cancer diseases, which together account for approximately 74% of all deaths worldwide [6]. There is a consensus that oxidative stress is associated with the onset, progression, or worsening of these diseases [7]. During aging, ROS generates cumulative and irreversible damage to cells, affecting proper cellular functioning and increasing functional deficits in the body [8,9]. From this perspective, obtaining antioxidant molecules of exogenous origin is essential for health, contributing to the reduction of the harmful effects of oxidative stress and the prevention of diseases related to this condition.
Natural products are an immeasurable source of chemical compounds that can bring benefits to human health, particularly due to their antioxidant action, capable of neutralizing the harmful effects of ROS and preventing diseases [2]. Among natural products, geopropolis is a substance uniquely produced by certain species of stingless bees [10] from diverse natural materials, including plant exudates, pollen, wax, mandibular secretions, and soil [11].
This bee product has gained prominence in recent years, where its emerging biological potential has been demonstrated through its antimicrobial [12], anti-inflammatory [13], antiviral [14], wound healing [15], anti-leishmaniasis [16], gastroprotective [17], anticancer [18], and antioxidant [19,20] properties. In previous studies, we demonstrated the antimicrobial, anti-inflammatory, and antimutagenic properties, as well as a potent antioxidant action against free radicals and lipid peroxidation, of geopropolis of the species M. orbignyi and M. q. anthidioides [12,19]. However, there is a lack of studies assessing the protective action on macromolecules in vitro and their antioxidant effects in vivo.
In this context, we investigated the protective effect of hydroethanolic extracts of geopropolis from M. orbignyi (HGMO) and M. q. anthidioides (HGMQ) against oxidative damage to proteins and DNA. Additionally, we evaluated the effects of the extracts on protection against oxidative and thermal stress, as well as their influence on the modulation of Caenorhabditis elegans lifespan.
2. Results
2.1. Protein Oxidation
The control AAPH reduced BSA integrity to about 35% when compared to the negative control. In contrast, both HGMO and HGMQ extracts exhibited protective effects by reducing BSA oxidation at all tested concentrations, maintaining BSA integrity with values comparable to the negative control at the highest concentrations. Figure 1A,B shows a concentration-dependent increase in band intensity. HGMO was able to maintain BSA integrity at 39.75 ± 0,51 and 97.18 ± 0,18% at concentrations of 12.5 and 250 µg/mL, respectively (Figure 1A). HGMQ maintained BSA integrity at 44.33 ± 0.56 and 90.89 ± 0.86% at the same concentrations, respectively (Figure 1B).
2.2. DNA-Induced Oxidative Damage
The extracts were assessed for their ability to prevent plasmid DNA fragmentation caused by ultraviolet radiation and hydrogen peroxide (H2O2). As a result, we demonstrated that both geopropolis extracts, HGMO and HGMQ, presented protective action against oxidative fragmentation of nucleic acid, as shown in Figure 2A,B. In the positive control exposed to UV radiation and H2O2, a total loss of DNA integrity was observed. However, in the presence of the extracts, DNA integrity was protected in a concentration-dependent manner, being more effective than the positive controls quercetin and gallic acid at the concentrations evaluated. At the lowest concentrations, up to 50 µg/mL, HGMO preserved DNA integrity at levels of 18.5 ± 0.07, 32.6 ± 3.04, and 66.3 ± 3.11%. HGMQ maintained DNA integrity at levels of 15.8 ± 1.27, 81.9 ± 8.41, and 83.8 ± 3.46%. On the other hand, at the highest concentrations (100–250 µg/mL) of both extracts, DNA integrity was fully preserved, with 100% protection against fragmentation.
2.3. In Vivo Tests Using the Caenorhabditis Elegans Model
2.3.1. Toxicity
In the in vivo studies, the first parameter evaluated was toxicity. In this test, the nematodes were treated with HGMO or HGMQ. As shown in Figure 3A–D, after 24 and 48 hours, no toxic effects were observed in any of the tested concentrations (ranging from 12.5 to 250 µg/mL). Once the safe dosage for use was determined, subsequent tests were conducted based on the observed responses.
2.3.2. Protection Against Oxidative Stress Induced by Juglone
The protective effects of HGMO and HGMQ against oxidative stress were evaluated by assessing the survival of C. elegans exposed to Juglone (80 µM) over 24 hours. As shown in Figure 4A,B, the oxidizing agent reduced the viability of the control group by approximately 70%. In contrast, nematodes treated with concentrations of 200 µg/mL and 250 µg/mL of HGMO exhibited an increase in survival of 22.3% and 37.3%, respectively. Additionally, nematodes treated with 250 µg/mL of HGMQ showed a 23.9% increase in survival, as illustrated in Figure 4A,B.
2.3.3. Protection Against Thermal Stress
In the heat stress assay, the increase in temperature led to a progressive reduction in the viability of control nematodes, reaching a reduction of approximately 80% after 6 h at 37 °C (Figures 5 A–F and 6 A–F). However, the results show that both geopropolis extracts exhibited protective action against heat stress. As shown in Figure 5C, at concentrations of 12.5 – 100 µg/mL, HGMO reduced nematode mortality in the 3-h period. Moreover, this extract also demonstrated protection in the 4-h period, at concentrations of 100, 200, and 250 µg/mL (Figure 5D). In Figure 6C, it was observed that HGMQ reduced mortality in the 3-h period in all concentrations evaluated.
2.3.4. Longevity Assay
In this assay, our results showed that nematodes treated with HGMO or HGMQ showed an increase in their lifespan, as observed in Figure 7A,B. Nematodes in the control group had a maximum lifespan of 24 days. On the other hand, C. elegans treated with HGMO at concentrations of 12.5 and 50 µg/mL showed a lifespan of 30 days (25% increase), while those treated at 200 and 250 µg/mL showed a lifespan of 32 days (33% increase). Animals treated with HGMQ showed similar results to those observed in the HGMO treatment.
Additionally, the highest concentration of both extracts was able to increase the average lifespan of C. elegans by approximately 11%, when compared to the control group, as shown in Table 1.
3. Discussion
Natural products have a unique structural diversity of chemical components, which can confer different biological activities [22]. Among these, geopropolis is a product of beekeeping origin, produced and used by stingless bees in their nests as a construction material, as it is a mixture of propolis and soil [10]. Geopropolis has been reported in ethnomedicinal practices for the treatment of infectious, respiratory, and ocular conditions [23]. Despite its use for medicinal purposes, studies about geopropolis are still scarce, especially on its pharmacological properties.
Previous studies from our group showed that geopropolis produced by the stingless bees Melipona orbignyi and Melipona quadrifasciata anthidioides exhibits antimicrobial, anti-inflammatory, antimutagenic, and antioxidant activities [12,19]. We also reported the chemical characterization of both hydroalcoholic extracts [12,19]. A summary of the major compounds identified in each extract is provided in the Supplementary Material (Table S1) to facilitate comparison. Building on these findings, and given that the pharmacological properties of this bee product remain incompletely characterized, we show here that HGMO and HGMQ attenuate AAPH-induced protein oxidative damage.
Proteins are macromolecules that are fundamental to cell biology and are very sensitive to oxidative damage, which can alter their structure and cause loss of function [24]. Oxidative damage to proteins alters the activity of enzymes and receptors, signal transduction mechanisms, and susceptibility to proteolysis [25]. ROS promotes damage to different amino acids and triggers oxidative structural changes in proteins, such as fragmentation and/or aggregate formation [26,27]. Thus, the deleterious effects resulting from oxidative stress are significant factors in neurodegenerative diseases [28].
In addition to the protective effect on proteins, HGMO and HGMQ were also able to protect DNA against oxidative damage caused by hydroxyl radicals, generated through the photolysis of H2O2 induced by UV light [29]. DNA is a macromolecule extremely sensitive to oxidative damage. Hydroxyl radicals react with DNA bases, abstracting electrons or hydrogen atoms, and promoting the cleavage and breakage of the DNA strand [30,31].
Oxidative damage to DNA can accumulate within its structure and trigger genomic instability, increasing the likelihood of mutations, a scenario related to the decline of cellular function, aging, and the carcinogenic process [32,33]. The extracts also had a more efficient protective action compared to the standard controls of gallic acid and quercetin. This effect may be due to the synergism between the bioactive molecules present in the extracts.
It is possible to find a diverse presence of chemical components in geopropolis from different stingless bees, such as phenolic acids (gallic, caffeic, syringic, benzoic) [34], flavonoids as vanillic acid, rutin, aromadendrin, quercetin [35] naringerin [14], ferreirin and dihydrokaempferide [18], hydrolyzable tannins (gallotannins and ellagitannins) [16], alkaloids [36], and terpenes [18,19]. The literature reports that phenolic compounds such as flavonoids exhibit antioxidant activity through different mechanisms, such as: (I) donating electrons and hydrogen atoms to stabilize free radicals, (II) chelating ROS-catalyzing metal ions, and (III) regulating oxidases and antioxidant enzymes [37]. In addition, some studies have reported the ability of certain molecules to bind to DNA and prevent oxidative damage [38].
From this perspective, the protection promoted by the extracts on proteins and DNA is possibly related to the presence of bioactive substances previously identified in the geopropolis of M. orbignyi and M. q. anthidioides, such as flavonoids naringin, aromadendrin, and methyl aromadendrin [12,19]. Additionally, these previous studies demonstrated that one of the antioxidant mechanisms of HGMO/HGMQ is the donation of electrons for the stabilization of free radicals, whether hydrophilic or lipophilic [12,19].
After confirming the protective effects of the extracts on biomolecules, we proceeded to evaluate their activity in vivo using the nematode C. elegans. This organism is genetically well characterized [39] and is widely used as a preclinical toxicity model due to its strong correlation with mammalian toxicity outcomes [40], as well as the presence of numerous genes homologous to those in humans [41]. Geopropolis extracts did not induce toxicity at any of the concentrations evaluated in C. elegans. Absence of toxicity was also observed in the propolis extract (2–8 mg/mL) of the species Heterotrigona itama in C. elegans [42], as well as for propolis of Plebeia catamarcensis and Tetragonisca fiebrigi, which showed no toxicity up to concentrations of 1500 µg/mL [43]. Additionally, no toxicity was detected for the coumarin isolated from the geopropolis of Melipona scutellaris, Cinnamoyloxy-mammeisin (2–20 mg/kg) in the Galleria mellonella model [44].
In the absence of toxicity, the pharmacological potential of the extracts becomes even more relevant. Regarding protection against oxidative stress, the extracts were able to increase the viability of C. elegans exposed to Juglone, a substance that promotes the generation of intracellular ROS H2O2 and O2• [45] and reduces nematode survival. The phenolic acid and flavonoid derivatives found in HGMO/HGMQ may be responsible for this protective effect against oxidative damage induced by Juglone. These compounds may act either directly on the ROS produced or indirectly by regulating the endogenous antioxidant defense system of C. elegans. Previous studies have reported that phenolic compounds can protect cells from oxidative damage by modulating signaling pathways associated with antioxidant action [37]. Thus, the modulation of antioxidant pathways in C. elegans is a potential mechanism of action for geopropolis extracts.
In this sense, an important pathway related to the response to oxidative stress (homologous in mammals) is the insulin/IGF-1 pathway [46]. This pathway is initiated by the daf-2 receptor (IGFR in humans), which negatively regulates the transcriptional factors daf-16 and Skn-1, responsible for the expression of genes associated with longevity and transcriptional control of antioxidant enzymes [41,47]. Daf-16 and Skn-1 are functional orthologs of the transcriptional factors FoxO-1 and Nrf2 in mammals [48]. In the literature, it is reported that phenolic compounds present in propolis and other natural products are capable of positively modulating daf-16 and Snk-1 [49].
In addition to chemical induction, heat stress is a condition that can induce oxidative stress by increasing the temperature to which the animal is subjected [50]. During this type of stress, C. elegans cells activate the heat shock response mechanism regulated by the transcription factor HSF-1 (Heat Shock factor-1), which in turn regulates the expression levels of specific defense proteins, known as chaperones or Heat Shock Proteins (HSPs) [51,52]. Animals treated with both extracts exhibited increased viability under heat stress, indicating enhanced thermotolerance.
It is worth noting that the protection of the extracts occurred only after 3 h or after 3 and 4 h, which suggests that after a certain period of heat stress, the extracts are unable to improve the recovery of the animals due to the severity of the damage generated during the last hours of stress. In the study by Jovic et al. [53], exposure to heat stress for more than 4 hours irreversibly compromised the survival, reproduction, and movement of the animals.
During heat stress, proteins can become misfolded, altering their conformation and potentially leading to the formation of protein aggregates [39,52]. One key pathway involved in cellular responses to various stress factors, such as inflammatory cytokines, UV radiation, oxidative stress, and heat stress, is the c-Jun N-terminal kinase (JNK) signaling pathway [54]. Upon detecting stressful stimuli, the JNK-1 protein is able to activate daf-16 and promote its translocation to the nucleus, where it will induce the expression of target genes responsible for avoiding harmful stresses [47,54]. Heat stress is able to drive the translocation of daf-16 to the nucleus [55], which in turn activates the heat shock response, promoting adaptation and survival in nematodes [52].
After demonstrating that geopropolis extracts increased the survival of C. elegans under abiotic stress (oxidative and thermal), we examined the effects on the longevity of nematodes treated daily with HGMO and HGMQ until all were dead. Notably, both extracts were able to extend the lifespan of the nematodes at all evaluated concentrations.
This study is the first to evaluate the effects of geopropolis on the C. elegans model, including its impact on longevity. This effect on lifespan may be attributed to the antioxidant properties of geopropolis extracts throughout their life. Additionally, this effect may be associated with the modulation of daf-16 and skn-1 transcription factor expression by specific bioactive compounds present in the extracts.
The flavonoid naringenin (present in our extracts) was investigated by Ge et al. [56], finding an increase in lifespan in C. elegans under normal and oxidative stress conditions. In this study, naringenin improved locomotion, delayed paralysis, reversed defective chemotaxis induced by beta-amyloid protein, increased the activity of antioxidant enzymes, and reduced ROS and MDA content. Furthermore, it positively modulated the transcription factors daf-16 and Skn-1 while negatively modulating daf-2, indicating that it promotes longevity through the insulin/IGF-1 pathway and MAPK signaling pathways.
The insulin/IGF-1 receptor and JNK (MAPK) signaling pathways are important signaling cascades involved in cellular responses to stress [57]. Positive signaling of the daf-2 pathway inhibits the activity of daf-16 and Skn-1, while positive signaling of the JNK pathway leads to the activation of daf-16. Once activated, daf-16 induces the expression of antioxidant enzyme genes and heat shock proteins, promoting increased stress resistance and longevity [39,41,47].
4. Materials and Methods
4.1. Chemicals and Reagents
All reagents used in the experiments were of analytical grade and purchased from Sigma-Aldrich/Merck (São Paulo, Brazil).
4.2. Obtaining Samples and Preparing Hydroethanolic Extracts of Geopropolis
Samples of geopropolis produced by the stingless bees Melipona orbignyi and Melipona quadrifasciata anthidioides (SISGEN Registration No. AF5168A) were identified and collected by biologist Prof. Dr. José Benedito Perrela Balestieri at a meliponary in Dourados, Mato Grosso do Sul state, Brazil (22°13’12’’S and 54°49’2’’W).
Crucially, the specific hydroethanolic extracts evaluated in the present study (HGMO and HGMQ) are the identical batches previously prepared and comprehensively chemically characterized by our research group. The full chemical profiles, including antioxidant and antimicrobial markers for these specific samples, were established and reported in the studies titled “Chemical Profile and Antioxidant, Anti-Inflammatory, Antimutagenic and Antimicrobial Activities of Geopropolis from the Stingless Bee Melipona orbignyi” [19] and “Chemical Composition and Pharmacological Effects of Geopropolis Produced by Melipona quadrifasciata anthidioides” [12]. Consequently, the chemical composition data detailed in these prior works directly apply to the bioassays performed herein.
Briefly, the extraction procedure followed the previously validated methodology. The geopropolis samples were macerated using a mortar and pestle and homogenized with 70% ethanol at a ratio of 1:3 (w/v). The resulting solutions were incubated in a shaker under moderate agitation, in the dark, at room temperature for 24 hours. Subsequently, the solutions were filtered through 80 g/m² qualitative filter paper (ProLab, São Paulo, Brazil), concentrated using a rotary vacuum evaporator (Gehaka, São Paulo, Brazil), and lyophilized to yield the final hydroethanolic extracts. The extraction yields were 14.9% for M. orbignyi (HGMO) and 6.0% for M. q. anthidioides (HGMQ), calculated using the formula:
Extraction yield (%) = (lyophilized sample x 100) / (original sample)
The extracts obtained were kept at −20 °C and protected from light until the analyses were performed.
4.3. Macromolecule Protection Assays
4.3.1. Protein Oxidation
The effect of the extracts on protein oxidation was evaluated using an oxidative induction assay with bovine serum albumin (BSA) and AAPH (2,2′-azobis(2-amidinopropane) dihydrochloride) as the oxidizing agent. In microtubes, 15 µL of BSA (1.5 mg/mL) were incubated together with 15 µL of HGMO or HGMQ, at different concentrations (12.5–250 µg/mL) for 30 min. Subsequently, 15 µL of the AAPH solution (75 mM) was added to each microtube, followed by incubation for 120 min at 37 ºC. The samples were then mixed with 75 µL of sample buffer (40% glycerol, 8% SDS, 0.25 mM Tris-HCl pH 6.8; 20% β-mercaptoethanol, 0.01% bromophenol blue) and heated at 100 ºC for 3 min. After this, the samples were applied to agarose gels and subjected to electrophoresis (SDS-PAGE 12%) using the Mini-PROTEAN Tetra Cell system (Bio-Rad Laboratories, CA, USA) at 200 V for 60 min. The gels were scanned in the Gel Doc EZ Imager photodocumenter (Bio-Rad Laboratories). The data were calculated from the average of triplicate measurements across four gels obtained from one independent experiment.
4.3.2. DNA-Induced Oxidative Damage
The effect of HGMO and HGMQ against DNA-induced oxidative damage was investigated by hydrogen peroxide (H2O2) and ultraviolet (UV) light-induced oxidative fragmentation. For this, plasmid DNA (pcDNA 3.1) (50 ng/µL) in 1x PBS solution was incubated with different concentrations of HGMO or HGMQ (12.5–250 µg/mL) and 30% H2O2 for 30 min. Then, the samples were incubated in a UVT-312 transilluminator at 302 nm, at room temperature for 5 min, followed by application and electrophoresis in 2% agarose gel containing ethidium bromide (10 ng/mL). As controls, plasmid DNA samples were also incubated with the standard antioxidants quercetin (Q) and gallic acid (GA) (250 µg/mL) and subjected to the same treatment. Subsequently, the gel was scanned in a Gel Doc EZ Imager (Bio-Rad Laboratories), and the analysis was performed using the Image Lab software. The average of the duplicates of two independent experiments was performed to obtain the data.
4.4. In Vivo Assays Using the Caenorhabditis Elegans Model
4.4.1. Cultivation and Maintenance of C. elegans
To perform the in vivo assays, wild-type N2 Bistrol nematodes obtained from the Caenorhabditis Genetics Center (CGC) (Minnesota), USA, were used. The nematodes were kept in an incubator at 20 ºC, cultivated in Petri dishes containing Nematode Growth Medium (NGM) and fed with Escherichia coli (OP50) bacteria. The bacteria used as food for the animals were inactivated with the antibiotic kanamycin (10 mM). To perform the experiments, the nematode culture was synchronized using 2% sodium hypochlorite and 5 M NaOH. The eggs resistant to alkaline lysis were transferred to new Petri dishes containing NGM medium and E. coli.
4.4.2. Acute Toxicity Assessment
To assess the toxic effect of acute exposure to HGMO and HGMQ, an average of 10 nematodes, synchronized in the L4 phase, were transferred to 96-well microplates containing 100 µL of M9 buffer and 100 µL of the respective extracts at different concentrations (12.5 to 250 µg/mL). Subsequently, the nematodes were incubated at 20 °C for 24 and 48 h. As a negative control, the nematodes were incubated with 200 µL of M9 buffer only. Additionally, an ethanol control was performed using the same concentration of the solvent present in the extracts. After the incubation period, viability was assessed by touch sensitivity using a platinum wire. Three independent experiments were performed in triplicate.
4.4.3. Protection Against Oxidative Stress Induced by Juglone
To evaluate the effects of HGMO and HGMQ against oxidative stress, wild-type C. elegans nematodes were exposed to the oxidizing agent Juglone (5-hydroxy-1,4-naphthoquinone) [21]. After synchronization, an average of 10 L4-stage nematodes were placed in wells containing 100 µL of M9 culture medium. The nematodes were then pre-incubated with 100 µL of different concentrations (12.5 to 250 µg/mL) of HGMO/HGMQ for 1 h. Subsequently, the oxidative stress inducer Juglone (80 µM) was added to the wells containing the nematodes and treatments, followed by incubation at 20 °C for 24 h. Nematodes incubated only with M9 buffer (250 µL) and with juglone (200 µL of M9 buffer + 50 µL of juglone) were used as negative and positive controls for oxidative damage, respectively. After the incubation period, the viability of the nematodes was assessed by touch sensitivity using a platinum wire. Three independent experiments were performed in triplicate.
4.4.4. Protection Against Thermal Stress
To evaluate the effect of heat stress, N2 nematodes were synchronized at the L4 stage and treated with HGMO or HGMQ (12.5 - 250 μg/mL). For this purpose, 10 nematodes were transferred to new plates containing NGM medium, E. coli OP50, and the respective treatments. Heat stress was induced by increasing the culture temperature from 20 to 37 °C for 6 h, with evaluations performed at 1 h intervals. After the incubation period, the nematodes were kept at 20 °C for 16 h. This procedure allows viable nematodes to recover and be counted by touch with the aid of a platinum wire. Three independent experiments were performed in triplicate.
4.4.5. Longevity Assay
To evaluate the effect on lifespan, synchronized N2 nematodes at the L4 stage were treated with HGMO/HGMQ. On the first day of the adult phase, 10 nematodes per group were transferred to new Petri dishes containing E. coli OP50 with or without the presence of HGMO/HGMQ at different concentrations (12.5, 50, 200, and 250 µg/mL), which was considered the first day of the experiment. During the first six days, when the animals were in the reproductive stage, the nematodes were transferred daily to new dishes containing their respective treatments. From the sixth day onwards, the nematodes were transferred from one plate to another every 2 days, and survival was assessed by classifying the animals as dead/alive daily, until all C. elegans died. Animals that did not move after slight stimulation with a platinum wire were considered dead. Nematodes with internally hatched eggs or not found were excluded from the analysis. Two independent experiments were performed in triplicate.
4.5. Statistical Analysis
All data were expressed as the mean ± standard error of the mean (SEM). Significant differences between groups were determined using Student’s t-test for comparison between two groups and analysis of variance (ANOVA), followed by Dunnett’s test for comparison of three or more groups using GraphPad Prism 5 software (San Diego, CA, USA). Results were considered significant when p < 0.05.
5. Conclusions
Taken together, these data broaden the antioxidant perspectives and beneficial effects of geopropolis extracts from M. orbignyi and M. q. anthidioides, evidenced by their protective activities against oxidative damage induced in proteins and DNA, protection against oxidative and thermal stress, and increased life expectancy in C. elegans, in addition to demonstrating the absence of toxicity. Although little known in the scientific literature, geopropolis is a product of native stingless bees used in traditional medicine, which demonstrates the potential for the development of biotechnological products aimed at the prevention and/or treatment of diseases associated with oxidative stress. The rapid anthropogenic transformation of biomes makes it urgent to understand the pharmacological potential of still unknown bee products from stingless bees, especially geopropolis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization H.F.d.S., E.Ld.S., and K.d.P.S.; methodology and validation H.F.d.S.; D.F.L., A.S.O.; W.H.B.; D.d.S.B.; formal analysis H.F.d.S., J.F.C.; P.d.S.R.; S.O.B; K.d.P.S. and E.L.d.S.; investigation, H.F.d.S.; D.F.L., A.S.O.; W.H.B.; resources J.F.C.; J.B.P.B.; K.d.P.S. and E.L.d.S.; data curation H.F.d.S.; A.S.O.; and E.L.d.S.; writing—original draft preparation, H.F.d.S.; and E.L.d.S.; writing—review and editing, D.d.S.B., J.F.C.; P.d.S.R.; S.O.B.; K.d.P.S. and E.L.d.S.; Conceptualization and supervision, K.d.P.S. and E.L.d.S.; project administration K.d.P.S. and E.L.d.S.; funding acquisition K.d.P.S. and E.L.d.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Foundation for the Support of the Development of Education, Science and Technology of the State of Mato Grosso do Sul (FUNDECT), the Coordination for the Improvement of Higher Education Personnel (CAPES), and the National Council for Scientific and Technological Development (CNPq).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kozlov, A.V.; Javadov, S.; Sommer, N. Cellular ROS and antioxidants: physiological and pathological role. Antioxidants 2024, 13(5), 602. [CrossRef]
- Fatma, H.; Jameel, M.; Siddique, H.R. An update on phytochemicals in redox homeostasis: “Virtuous or Evil” in cancer chemoprevention? Chemistry 2023, 5(1), 201–222. [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative stress: a key modulator in neurodegenerative diseases. Molecules 2019, 24(8), 1583. [CrossRef]
- Remigante, A.; Morabito, R. Cellular and molecular mechanisms in oxidative stress-related diseases. Int. J. Mol. Sci. 2022, 23(14), 8017. [CrossRef]
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; et al. Oxidative stress in human pathology and aging: molecular mechanisms and perspectives. Cells 2022, 11(3), 552. [CrossRef]
- World Health Organization—Noncommunicable Diseases. Available online: https://www.who.int/health-topics/noncommunicable-diseases#tab=tab_1 (accessed on 20 August 2025).
- Nediani, C.; Ruzzolini, J.; Dinu, M. Oxidative stress and inflammation as targets for novel preventive and therapeutic approaches in non-communicable diseases III. Antioxidants 2024, 13(11), 1404. [CrossRef]
- Chiang, J.C.; Chen, W.M.; Newman, C.; Chen, B.P.; Lee, H. Lysophosphatidic acid receptor 3 promotes mitochondrial homeostasis against oxidative stress: potential therapeutic approaches for Hutchinson–Gilford progeria syndrome. Antioxidants 2022, 11(2), 351. [CrossRef]
- Goh, J.; Wong, E.; Soh, J.; Maier, A.B.; Kennedy, B.K. Targeting the molecular & cellular pillars of human aging with exercise. FEBS J. 2023, 290(3), 649–668. [CrossRef]
- Nogueira-Neto, P. Vida e criação de abelhas indígenas sem ferrão, 1ª ed.; Edição Nogueirapis: São Paulo, Brazil, 1997; 446 p.
- Oliveira, L.P.G.; Conte, F.L.; de Oliveira Cardoso, E.; Conti, B.J.; Santiago, K.B.; de Assis Golim, M.; Sforcin, J.M. A new chemotherapeutic approach using doxorubicin simultaneously with geopropolis favoring monocyte functions. Life Sci. 2019, 217, 81–90. [CrossRef]
- Santos, C.M.; Campos, J.F.; dos Santos, H.F.; Balestieri, J.B.P.; Silva, D.B.; de Picoli Souza, K.; et al. Chemical composition and pharmacological effects of geopropolis produced by Melipona quadrifasciata anthidioides. Oxid. Med. Cell. Longev. 2017, 2017, 1–12. [CrossRef]
- Honorio, M.S.; Sartori, A.A.; Ripari, N.; Santiago, K.B.; Sforcin, J.M. Anti-inflammatory action of geopropolis produced by stingless bees on human peripheral blood mononuclear cells. Hum. Immunol. 2024, 85(4), 110825. [CrossRef]
- Silva, P.G.D.; Chaves, E.J.F.; Silva, T.M.S.; Rocha, G.B.; Dantas, W.M.; Oliveira, R.N.D.; Pena, L.J. Antiviral activity of flavonoids from geopropolis of the Brazilian Jandaira bee against Zika and dengue viruses. Pharmaceutics 2023, 15(10), 2494. [CrossRef]
- Sousa-Fontoura, D.M.; Olinda, R.G.; Viana, G.A.; De F. M. Costa, K.M.; Batista, J.S.; Serrano, R.M.; et al. Wound healing activity and chemical composition of geopropolis from Melipona subnitida. Rev. Bras. Farmacogn. 2020, 30, 367–373. [CrossRef]
- Dutra, R.P.; Bezerra, J.L.; Silva, M.C.P.D.; Batista, M.C.A.; Patrício, F.J.B.; et al. Antileishmanial activity and chemical composition from Brazilian geopropolis produced by stingless bee Melipona fasciculata. Rev. Bras. Farmacogn. 2019, 29, 287–293. [CrossRef]
- Batista, J.S.; da Silva Teofilo, T.; de Araujo Junior, H.N.; Felix, N.S.; de Paiva, K.A.R.; et al. Gastroprotective effect of the hydroethanolic extract of geopropolis produced by Melipona subnitida (Meliponinae, Apidae) in Wistar rats. Vet. Med. 2025, 70(4), 134. [CrossRef]
- Paz, M.M.D.; Sette, K.M.; dos Santos, R.E.; Barbosa e Vasconcelos, A.L.; Costa, D.C.F.D.; Amaral, A.C.F.; et al. Brazilian stingless bee geopropolis exhibits antioxidant properties and anticancer potential against hepatocellular carcinoma cells. Antioxidants 2025, 14(2), 141. [CrossRef]
- Santos, H.F.D.; Campos, J.F.; Santos, C.M.D.; Balestieri, J.B.P.; Silva, D.B.; Carollo, C.A.; et al. Chemical profile and antioxidant, anti-inflammatory, antimutagenic and antimicrobial activities of geopropolis from the stingless bee Melipona orbignyi. Int. J. Mol. Sci. 2017, 18(5), 953. [CrossRef]
- Gabriel, M.B.; Carneiro, M.J.; de Camargo, R.C.; Sawaya, A.C. The chemical composition and antioxidant activity of mandaçaia (Melipona quadrifasciata) geopropolis varies more due to region than month of collection. Nat. Prod. Res. 2022, 36(6), 1626–1630. [CrossRef]
- Sobeh, M.; Mahmoud, M.F.; Abdelfattah, M.A.; Cheng, H.; El-Shazly, A.M.; Wink, M. A proanthocyanidin-rich extract from Cassia abbreviata exhibits antioxidant and hepatoprotective activities in vivo. J. Ethnopharmacol. 2018, 213, 38–47. [CrossRef]
- Sorokina, M.; Steinbeck, C. Review on natural products databases: where to find data in 2020. J. Cheminform. 2020, 12(1), 20. [CrossRef]
- Souza, R.C.D.S.; Yuyama, L.K.O.; Aguiar, J.P.L.; Oliveira, F.P.M. Valor nutricional do mel e pólen de abelhas sem ferrão da região amazônica. Acta Amazon. 2004, 34, 333–336. [CrossRef]
- Tramutola, A.; Lanzillotta, C.; Perluigi, M.; Butterfield, D.A. Oxidative stress, protein modification and Alzheimer disease. Brain Res. Bull. 2017, 133, 88–96. [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: impact on human health. Pharmacogn. Rev. 2010, 4(8), 118. [CrossRef]
- Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Lopez-Burillo, S.; Reiter, R.J. Oxidative damage to catalase induced by peroxyl radicals: functional protection by melatonin and other antioxidants. Free Radic. Res. 2003, 37(5), 543–553. [CrossRef]
- Cecarini, V.; Gee, J.; Fioretti, E.; Amici, M.; Angeletti, M.; et al. Protein oxidation and cellular homeostasis: emphasis on metabolism. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773(2), 93–104. [CrossRef]
- Selvaraj, N.R.; Nandan, D.; Nair, B.G.; Nair, V.A.; Venugopal, P.; et al. Oxidative stress and redox imbalance: common mechanisms in cancer stem cells and neurodegenerative diseases. Cells 2025, 14(7), 511. [CrossRef]
- Rajan, I.; Narayanan, N.; Rabindran, R.; Jayasree, P.R.; Manish Kumar, P.R. Zingerone protects against stannous chloride-induced and hydrogen peroxide-induced oxidative DNA damage in vitro. Biol. Trace Elem. Res. 2013, 155, 455–459. [CrossRef]
- Collin, F. Chemical basis of reactive oxygen species reactivity and involvement in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20(10), 2407. [CrossRef]
- Wu, X.; Dhanasekaran, S. Protective effect of leaf extract of Abutilon indicum on DNA damage and peripheral blood lymphocytes in combating the oxidative stress. Saudi Pharm. J. 2020, 28(8), 943–950. [CrossRef]
- Savina, N.V.; Nikitchenko, N.V.; Kuzhir, T.D.; Rolevich, A.I.; Krasny, S.A.; Goncharova, R.I. The cellular response to oxidatively induced DNA damage and polymorphism of some DNA repair genes associated with clinicopathological features of bladder cancer. Oxid. Med. Cell. Longev. 2016, 2016, 1–13. [CrossRef]
- Schumacher, B.; Pothof, J.; Vijg, J.; Hoeijmakers, J.H. The central role of DNA damage in the ageing process. Nature 2021, 592(7856), 695–703. [CrossRef]
- Zhao, L.; Yu, M.; Sun, M.; Xue, X.; Wang, T.; Cao, W.; Sun, L. Rapid determination of major compounds in the ethanol extract of geopropolis from Malaysian stingless bees, Heterotrigona itama, by UHPLC-Q-TOF/MS and NMR. Molecules 2017, 22(11), 1935. [CrossRef]
- Ferreira, B.L.; Gonzaga, L.V.; Vitali, L.; Micke, G.A.; Maltez, H.F.; et al. Southern-Brazilian geopropolis: A potential source of polyphenolic compounds and assessment of mineral composition. Food Res. Int. 2019, 126, 108683. [CrossRef]
- Coelho, G.R.; Mendonça, R.Z.; Vilar, K.D.S.; Figueiredo, C.A.; Badari, J.C.; et al. Antiviral action of hydromethanolic extract of geopropolis from Scaptotrigona postica against antiherpes simplex virus (HSV-1). Evid. Based Complement. Alternat. Med. 2015, 2015, 287206. [CrossRef]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6(2), 115–123. [CrossRef]
- Sjakste, N.; Djelić, N.; Dzintare, M.; Živković, L. DNA-binding and DNA-protecting activities of small natural organic molecules and food extracts. Chem.-Biol. Interact. 2020, 323, 109030. [CrossRef]
- Kim, D.K.; Kim, T.H.; Lee, S.J. Mechanisms of aging-related proteinopathies in Caenorhabditis elegans. Exp. Mol. Med. 2016, 48(10), e263. [CrossRef]
- Hunt, P.R. The C. elegans model in toxicity testing. J. Appl. Toxicol. 2017, 37(1), 50–59. [CrossRef]
- Ayuda-Durán, B.; González-Manzano, S.; González-Paramás, M.A.; Santos-Buelga, C. Caenorhabditis elegans as a model organism to evaluate the antioxidant effects of phytochemicals. Molecules 2020, 25(14), 3194. [CrossRef]
- Abdullah, N.A.; Ja’afar, F.; Yasin, H.M.; Taha, H.; Petalcorin, M.I.; et al. Physicochemical analyses, antioxidant, antibacterial, and toxicity of propolis particles produced by the stingless bee Heterotrigona itama found in Brunei Darussalam. Heliyon 2019, 5(9), e02476. [CrossRef]
- Arteman, K.D.S.; da Rocha, P.D.S.; Leite, D.F.; Oliveira, A.S.; da Silva, I.V.; et al. Plebeia catamarcensis and Tetragonisca fiebrigi (Hymenoptera, Apidae) propolis promotes longevity and anti-Alzheimer effects in Caenorhabditis elegans. PLoS One 2025, 20(6), e0321487. [CrossRef]
- Cunha, M.G.; Sardi, J.D.C.O.; Freires, I.A.; Franchin, M.; Rosalen, P.L. Antimicrobial, anti-adherence and antibiofilm activity against Staphylococcus aureus of a 4-phenyl coumarin derivative isolated from Brazilian geopropolis. Microb. Pathog. 2020, 139, 103855. [CrossRef]
- Wang, P.; Zhang, S.D.; Jiao, J.; Wang, W.; Yu, L.; Zhao, X.L.; Fu, Y.J. ROS-mediated p53 activation by juglone enhances apoptosis and autophagy in vivo and in vitro. Toxicol. Appl. Pharmacol. 2019, 379, 114647. [CrossRef]
- Back, P.; Braeckman, B.P.; Matthijssens, F. ROS in aging Caenorhabditis elegans: damage or signaling? Oxid. Med. Cell. Longev. 2012, 2012, 608478. [CrossRef]
- Mathew, R.; Bhadra, M.P.; Bhadra, U. Insulin/insulin-like growth factor-1 signalling (IIS) based regulation of lifespan across species. Biogerontology 2017, 18(1), 35–53. [CrossRef]
- Tullet, J.M.; Green, J.W.; Au, C.; Benedetto, A.; Thompson, M.A.; Clark, E.; et al. The SKN-1/Nrf2 transcription factor can protect against oxidative stress and increase lifespan in Caenorhabditis elegans by distinct mechanisms. Aging Cell 2017, 16(5), 1191–1194. [CrossRef]
- Martel, J.; Wu, C.Y.; Peng, H.H.; Ko, Y.F.; Yang, H.C.; et al. Plant and fungal products that extend lifespan in Caenorhabditis elegans. Microb. Cell 2020, 7(10), 255. [CrossRef]
- Emami, N.K.; Jung, U.; Voy, B.; Dridi, S. Radical response: effects of heat stress-induced oxidative stress on lipid metabolism in the avian liver. Antioxidants 2021, 10(1), 35. [CrossRef]
- Brunquell, J.; Morris, S.; Lu, Y.; Cheng, F.; Westerheide, S.D. The genome-wide role of HSF-1 in the regulation of gene expression in Caenorhabditis elegans. BMC Genomics 2016, 17(1), 1–18. [CrossRef]
- Kyriakou, E.; Taouktsi, E.; Syntichaki, P. The thermal stress coping network of the nematode Caenorhabditis elegans. Int. J. Mol. Sci. 2022, 23(23), 14907. [CrossRef]
- Jovic, K.; Sterken, M.G.; Grilli, J.; Bevers, R.P.; Rodriguez, M.; et al. Temporal dynamics of gene expression in heat-stressed Caenorhabditis elegans. PLoS One 2017, 12(12), e0189445. [CrossRef]
- Oh, S.W.; Mukhopadhyay, A.; Svrzikapa, N.; Jiang, F.; Davis, R.J.; Tissenbaum, H.A. JNK regulates lifespan in Caenorhabditis elegans by modulating nuclear translocation of forkhead transcription factor/DAF-16. Proc. Natl. Acad. Sci. U.S.A. 2005, 102(12), 4494–4499. [CrossRef]
- Tepper, R.G.; Ashraf, J.; Kaletsky, R.; Kleemann, G.; Murphy, C.T.; Bussemaker, H.J. PQM-1 complements DAF-16 as a key transcriptional regulator of DAF-2-mediated development and longevity. Cell 2013, 154(3), 676–690. [CrossRef]
- Ge, Y.; Chen, H.; Wang, J.; Liu, G.; Cui, S.W.; et al. Naringenin prolongs lifespan and delays aging mediated by IIS and MAPK in Caenorhabditis elegans. Food Funct. 2021, 12(23), 12127–12141. [CrossRef]
- Wolf, M.; Nunes, F.; Henkel, A.; Heinick, A.; Paul, R.J. The MAP kinase JNK-1 of Caenorhabditis elegans: location, activation, and influences over temperature-dependent insulin-like signaling, stress responses, and fitness. J. Cell. Physiol. 2008, 214(3), 721–729. [CrossRef]
Figure 1.
Protection against AAPH-induced protein oxidation (75 mM) promoted by HGMO (A) and HGMQ (B) at different concentrations. Values were expressed as mean ± SEM. * vs. CT; # vs. AAPH. ***, ### p < 0,001.
Figure 1.
Protection against AAPH-induced protein oxidation (75 mM) promoted by HGMO (A) and HGMQ (B) at different concentrations. Values were expressed as mean ± SEM. * vs. CT; # vs. AAPH. ***, ### p < 0,001.
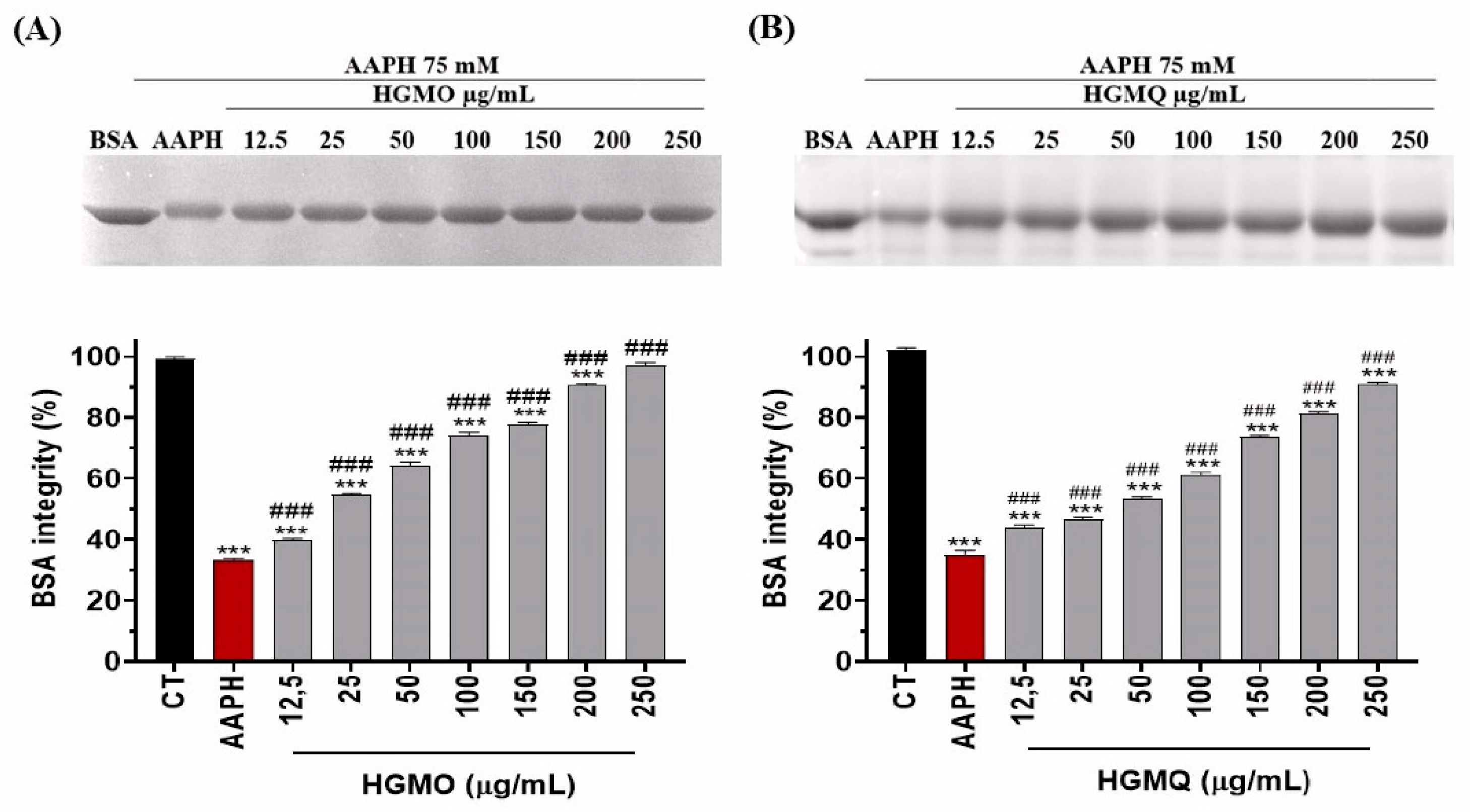
Figure 2.
depicts the protection against oxidative fragmentation in plasmid DNA induced by UV radiation and H2O2 (30%) promoted by HGMO/HGMQ at different concentrations. (A) Representative image of agarose gel with the respective treatments. (B) DNA fragmentation (%). Values were expressed as mean ± SEM. # vs. CT; *vs. UV/H2O2; ***, ###p < 0.001. .
Figure 2.
depicts the protection against oxidative fragmentation in plasmid DNA induced by UV radiation and H2O2 (30%) promoted by HGMO/HGMQ at different concentrations. (A) Representative image of agarose gel with the respective treatments. (B) DNA fragmentation (%). Values were expressed as mean ± SEM. # vs. CT; *vs. UV/H2O2; ***, ###p < 0.001. .
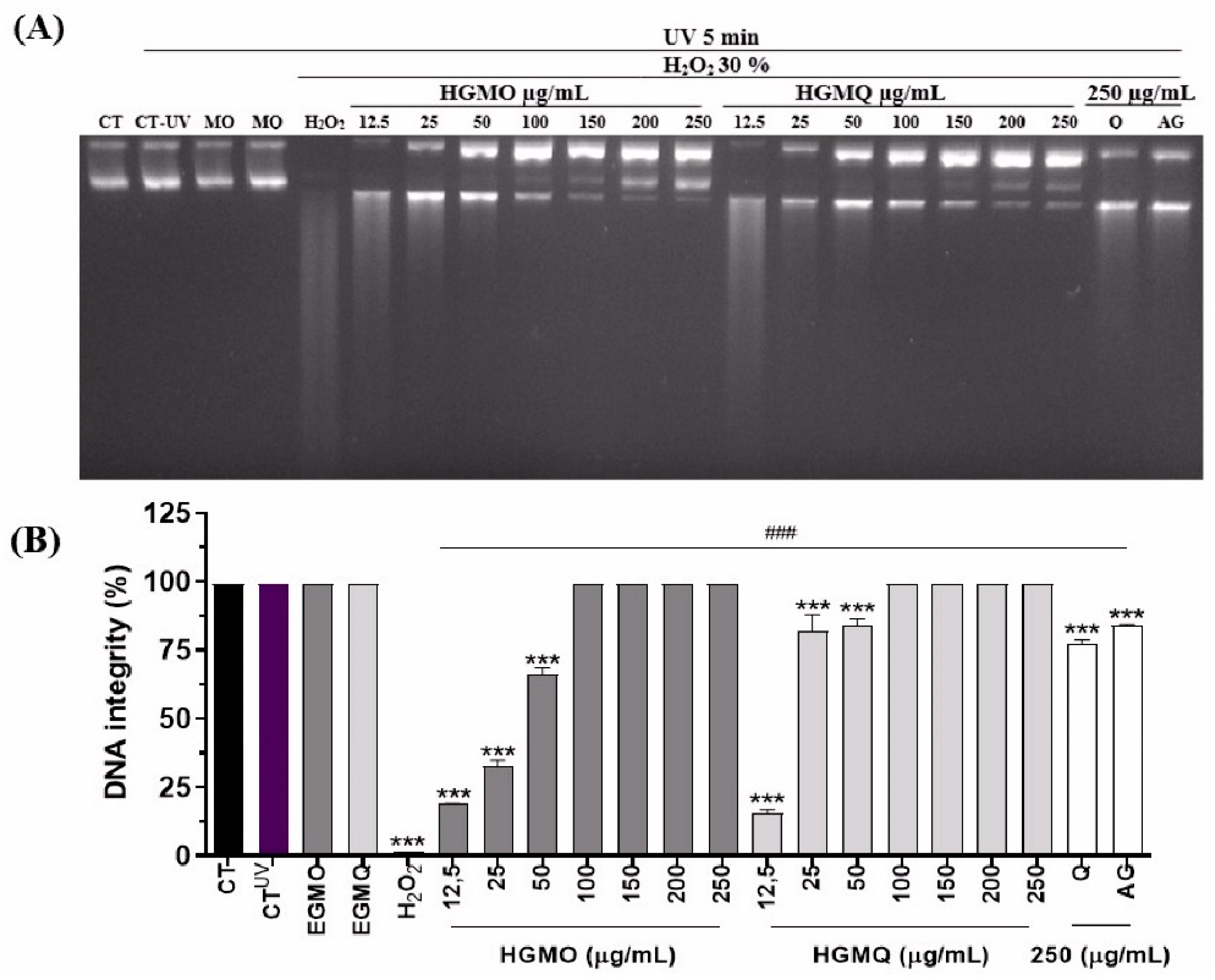
Figure 3.
Toxicity of geopropolis extracts on C. elegans in the periods of (A) 24 h and (B) 48 h for HGMO and (C) 24 h and (D) 48 h for HGMQ. Values were expressed as mean ± SEM. EC: ethanol control.
Figure 3.
Toxicity of geopropolis extracts on C. elegans in the periods of (A) 24 h and (B) 48 h for HGMO and (C) 24 h and (D) 48 h for HGMQ. Values were expressed as mean ± SEM. EC: ethanol control.
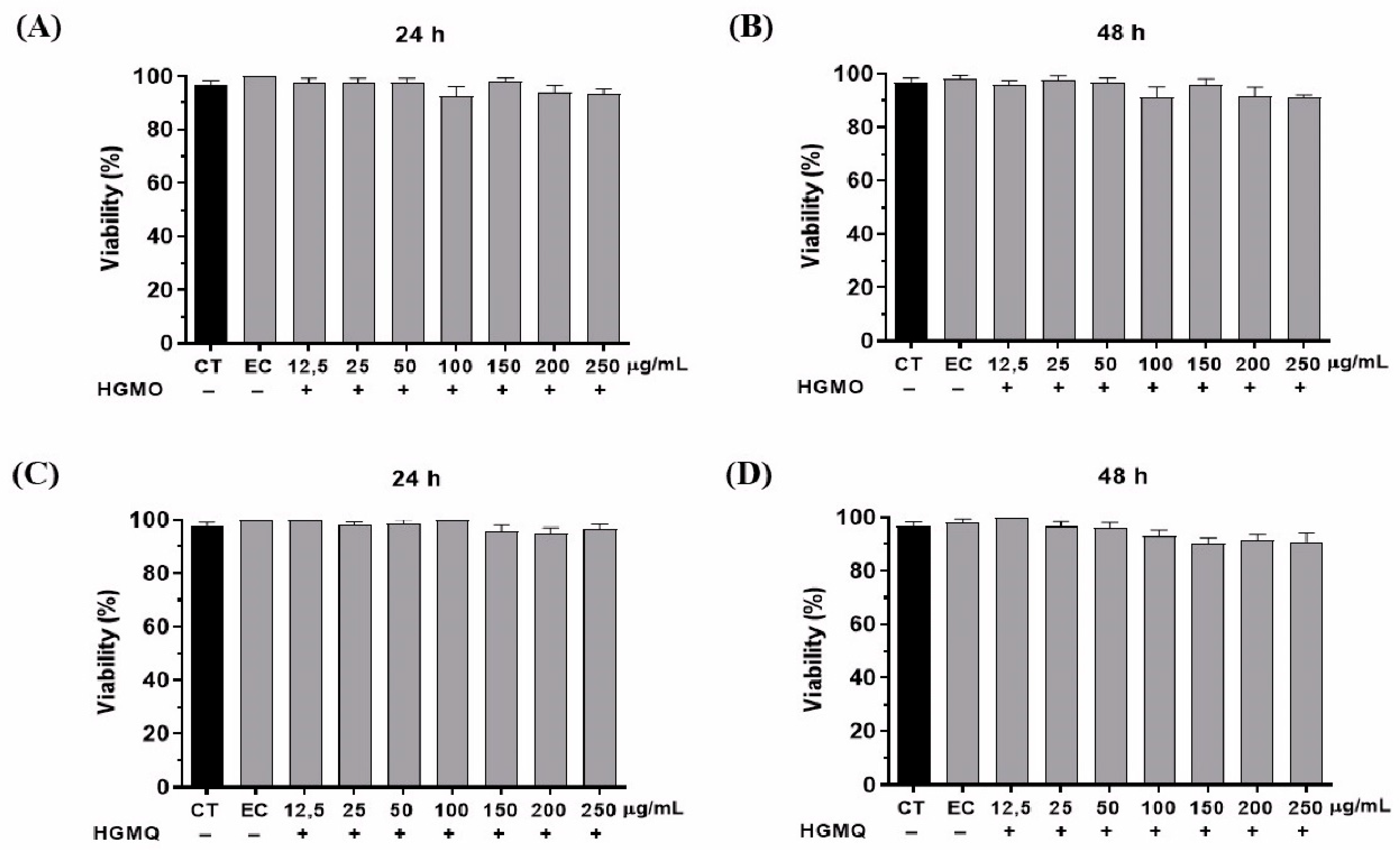
Figure 4.
Protective effect of HGMO (A) and HGMQ (B) against oxidative stress induced by juglone after 24 hours in C.elegans. EC: ethanol control, JC: juglone control. Values were expressed as mean ± SEM. * vs. CT; # vs. CJ; *, # p < 0.05; **, ## p < 0,01; ***, ### p < 0,001.
Figure 4.
Protective effect of HGMO (A) and HGMQ (B) against oxidative stress induced by juglone after 24 hours in C.elegans. EC: ethanol control, JC: juglone control. Values were expressed as mean ± SEM. * vs. CT; # vs. CJ; *, # p < 0.05; **, ## p < 0,01; ***, ### p < 0,001.
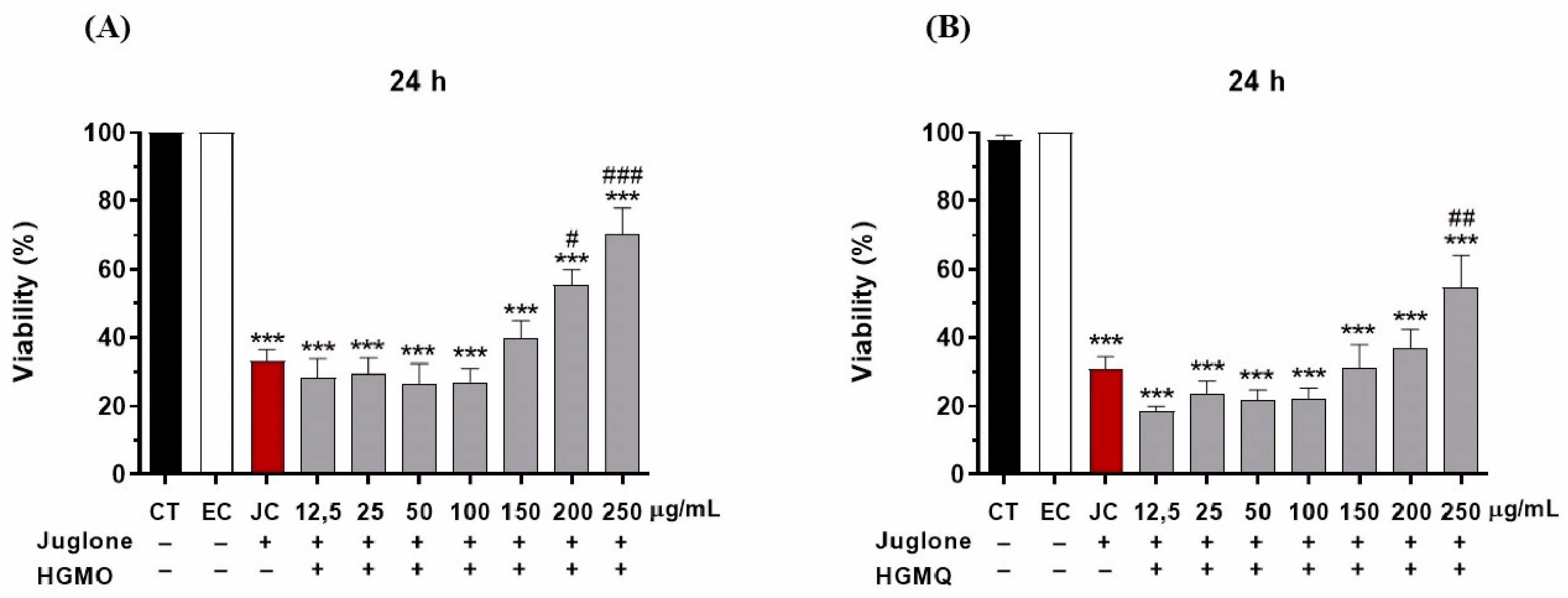
Figure 5.
Effects of HGMO on C. elegans viability after 1 (A), 2 (B), 3 (C), 4 (D), 5 (E), and 6 h (F) of heat stress induction. Values were expressed as mean ± SEM. * vs. CT20 ºC; # vs. CT37 ºC; *, # p < 0.05; **, ## p < 0.01; ***, ### p < 0.001.
Figure 5.
Effects of HGMO on C. elegans viability after 1 (A), 2 (B), 3 (C), 4 (D), 5 (E), and 6 h (F) of heat stress induction. Values were expressed as mean ± SEM. * vs. CT20 ºC; # vs. CT37 ºC; *, # p < 0.05; **, ## p < 0.01; ***, ### p < 0.001.
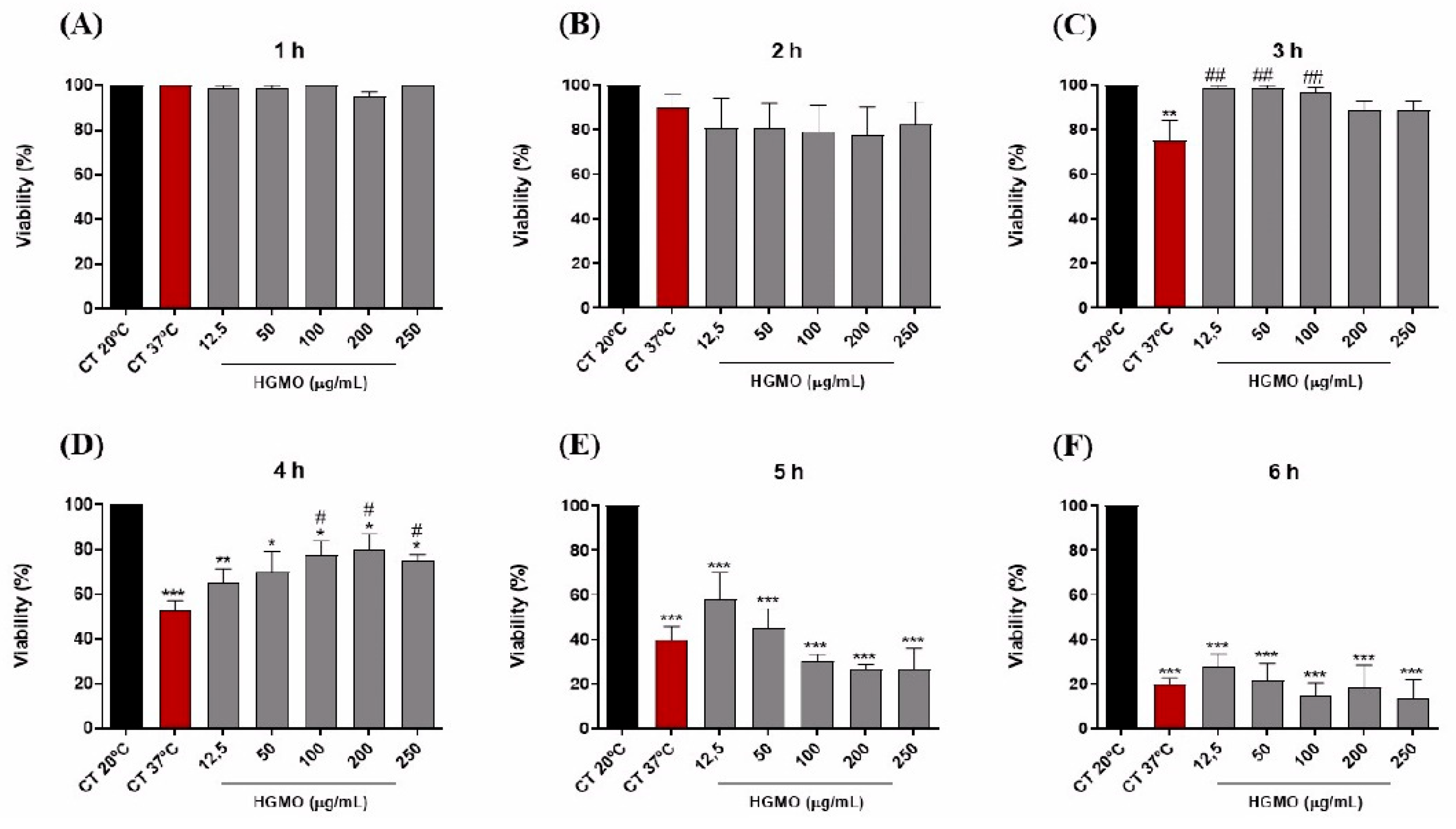
Figure 6.
Effects of HGMQ on C. elegans viability after 1 (A), 2 (B), 3 (C), 4 (D), 5 (E), and 6 h (F) of heat stress induction. Values were expressed as mean ± SEM. * vs. CT20 ºC; # vs. CT37 ºC; *, # p < 0.05; **, ## p < 0.01; ***, ### p < 0.001.
Figure 6.
Effects of HGMQ on C. elegans viability after 1 (A), 2 (B), 3 (C), 4 (D), 5 (E), and 6 h (F) of heat stress induction. Values were expressed as mean ± SEM. * vs. CT20 ºC; # vs. CT37 ºC; *, # p < 0.05; **, ## p < 0.01; ***, ### p < 0.001.
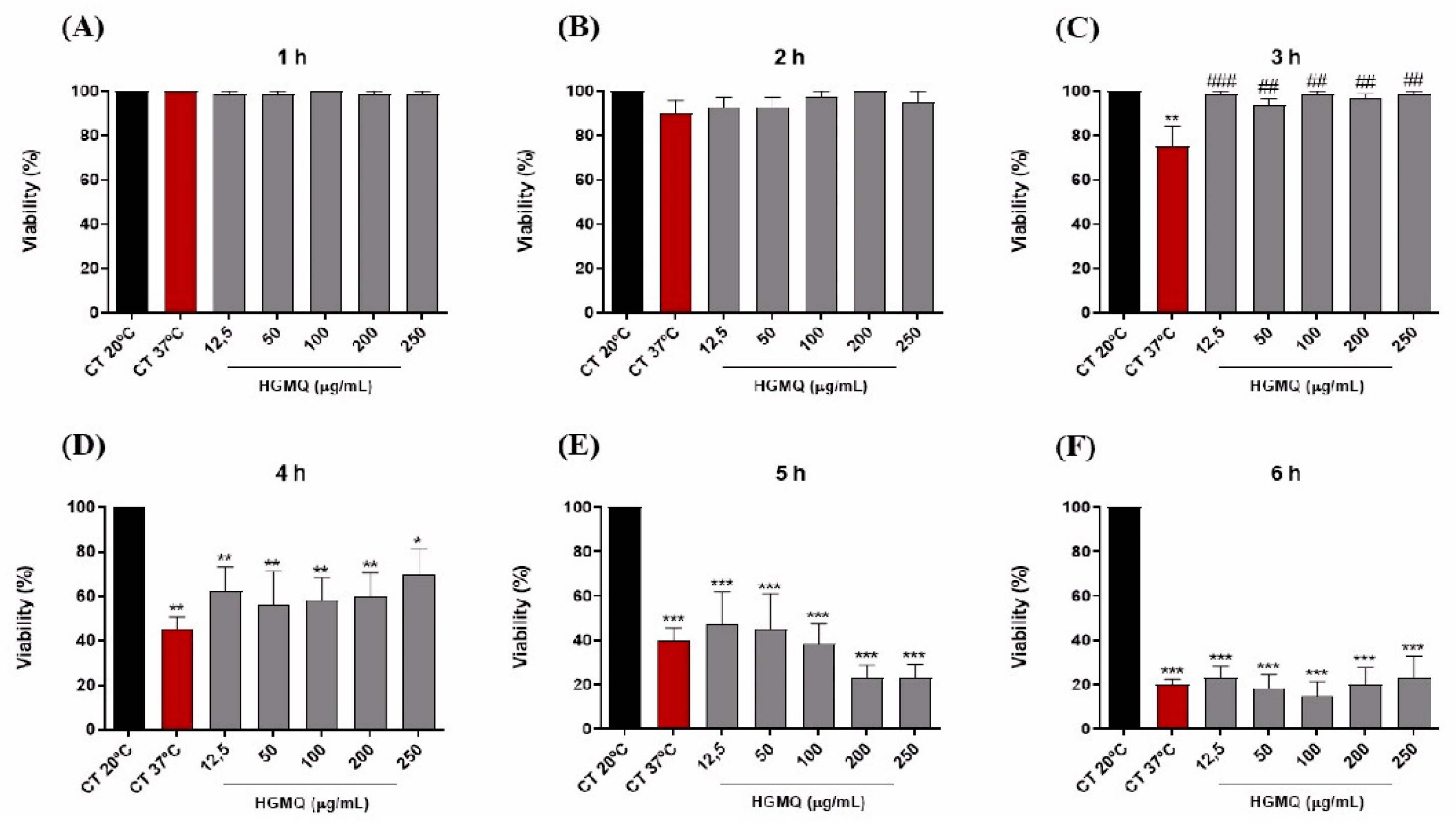
Figure 7.
Effects of geopropolis extracts on C. elegans lifespan. (A) Nematodes treated with different concentrations of HGMO, and (B) HGMQ. Values were expressed as mean ± SEM.
Figure 7.
Effects of geopropolis extracts on C. elegans lifespan. (A) Nematodes treated with different concentrations of HGMO, and (B) HGMQ. Values were expressed as mean ± SEM.
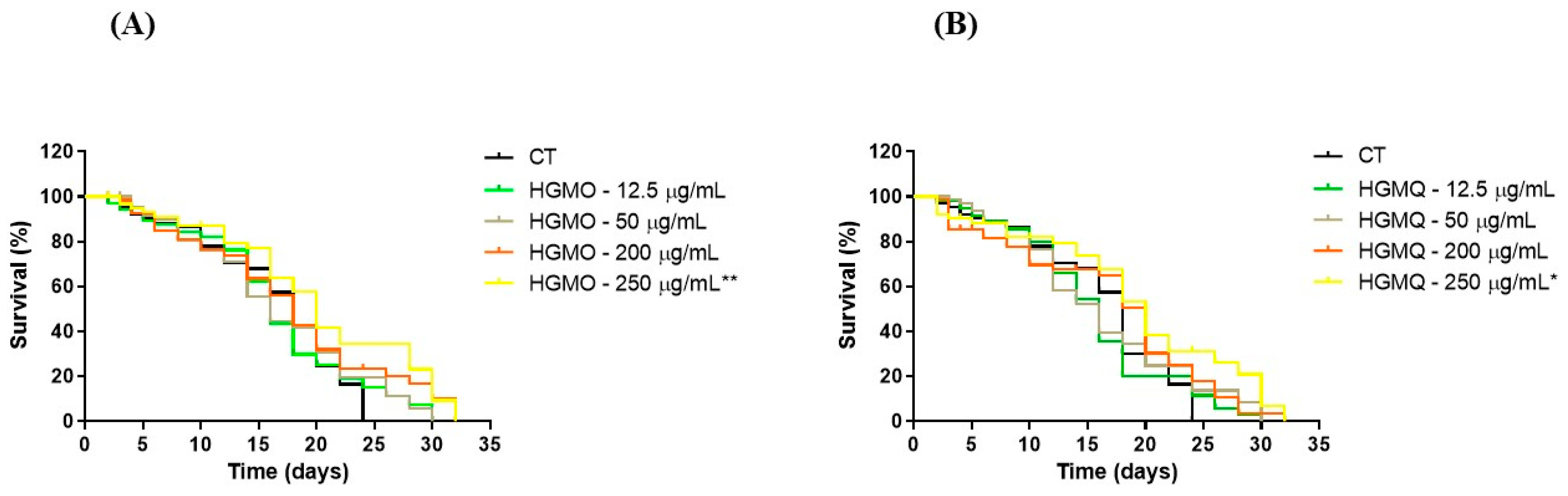

Table 1.
Effects of geopropolis extracts on the mean life expectancy of wild C. elegans under standard conditions.
Table 1.
Effects of geopropolis extracts on the mean life expectancy of wild C. elegans under standard conditions.
| Longevity | Average life expectancya | P value (log rank) vs. Controlb | Nºc |
|---|---|---|---|
| Control | 18 | 60 (2) | |
| HGMO 12,5 µg/mL | 16 | 0.7563 | 60 (2) |
| HGMO 50 µg/mL | 16 | 0.3390 | 60 (2) |
| HGMO 200 µg/mL | 18 | 0.1725 | 60 (2) |
| HGMO 250 µg/mL | 20 | 0.0068** | 60 (2) |
| HGMQ 12.5 µg/mL | 16 | 0.8620 | 60 (2) |
| HGMQ 50 µg/mL | 16 | 0.7451 | 60 (2) |
| HGMQ 200 µg/mL | 20 | 0.2352 | 60 (2) |
| HGMQ 250 µg/mL | 20 | 0.0285* | 60 (2) |
*Represents statistically significant results when the treated group was compared with the control group. * p < 0.05 and ** p < 0.01. aAverage life expectancy measured in days. bComparisons were performed using the log-rank (Mantel-Cox) test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



