Submitted:
30 January 2026
Posted:
02 February 2026
You are already at the latest version
Abstract
Ulcerative Colitis (UC) represents a major global health burden characterized by chronic intestinal inflammation, dysbiosis, and limited therapeutic options. The G pro-tein-coupled receptor 35(GPR35), highly expressed in the gastrointestinal tract and immune cells, has emerged as a promising target for maintaining gut homeostasis and promoting mucosal repair. Here, we identified ellagic acid (EA), a natural polyphenol abundant in fruits and nuts, as a novel dietary agonist of GPR35 through Nanobit assay. Then we utilized molecular docking and molecular dynamics simulations to prove the binding ability of GPR35 and EA, and we further elucidated the binding site via point mutation. In a murine model of experimental colitis, EA administration significantly alleviated disease severity, as evidenced by improved body weight, disease activity index, and colon length. Notably, EA strongly promoted mucosal healing and preserved intestinal barrier integrity. Mechanistically, EA enhanced the migration and prolifera-tion of human colonic epithelial cells, an effect that was specifically abolished by the GPR35 antagonist CID2745687,indicating the key role GPR35 played in the intestinal repair. Our findings reveal a previously unrecognized role for EA as a natural GPR35 agonist with potent protective effects in colitis, highlighting the potential of polyphe-nol-rich diets as adjunctive strategies for IBD management by targeting mucosal repair pathways.
Keywords:
GPR35
; Nanobit assay
; ellagic acid
; ulcerative colitis
; intestinal epithelial repair
1. Introduction
As a chronic and relapsing type of inflammatory bowel disease (IBD), UC is characterized by continuous and diffuse inflammation of the colonic mucosa. Common clinical symptoms primarily include abdominal pain, diarrhea, bloody stools, and so on [1] . Epidemiologically, the incidence of UC has been rising steadily, rendering it a growing global health challenge. And it is worth noting that the westernization of dietary was recognized to contribute to the prevalence of UC [2] . A complex interplay of genetic, immune, gut microenvironmental, infectious, and dietary factors is widely believed to underlie ulcerative colitis. Nevertheless, a complete understanding of its precise etiology and pathogenesis remains elusive. [3] .Current therapeutic strategies of UC, including 5-aminosalicylic acid agents(5-ASA), corticosteroids, immunosuppressants, biologics, and Janus kinase (JAK) inhibitors, are deployed according to disease stage [4] ; but owing to the significant side effects, high cost and drug dependency of these therapies, the alternative and/or complementary therapies become a urgent need for many people suffering from UC [5] .
As the largest and most versatile family of cell surface receptors in the human genome, GPCRs play a fundamental role as signaling conduits, translating extracellular stimuli into intracellular responses [6] . Indeed, GPCRs are currently targeted approximately 37% of approved drugs, making it as an ideal target for natural products [7] . GPR35, an orphan GPCR highly expressed in the gastrointestinal tract, primarily intestinal epithelial cells (IECs), plays an important role in gastrointestinal homeostasis [8] . Evidence indicated that the hyperactive T108M mutation in the GPR35 gene, a coding variant, correlated with a higher susceptibility to UC, leading to the classification of GPR35 as an IBD risk gene [9] . Despite several molecules, such as zaprinast, lodoxamide, pamoic acid and compound1 has been confirmed to be the potential agonist of GPR35, the ligand derived from natural products remains in vein [10] .
There are many methods in drug screening of GPCR, including cAMP assay, calcium assay, FRET, BRET, β-arrestin recruitment assay, NanoBiT assay and so on [11] . Among them, BRET and NanoBiT assay have been widely adopted in GPCR ligand discovery owing to their low background noise and high sensitivity [12] . The specific recruitment and activation of intracellular G proteins and β-arrestins are triggered by the conformational changes that GPCRs undergo upon ligand binding. As previously demonstrated, the complex structure of GPCR recruiting Gα13 has been resolved by cryo-electron microscopy, providing a foundation for further ligand screen of GPR35 [13] Besides, computational biology approaches, most notably molecular docking and molecular dynamics simulations, are also widely applied in binding site prediction and structure-based drug design [14,15] . For a known active molecule, molecular docking can help reveal the microscopic mechanisms of its interaction with the ligand [16] . However, the accuracy of molecular docking is directly tied to the quality of the target protein and the accuracy of the docking algorithm. Therefore, to ensure the reliability and biological relevance of the docking outcomes, subsequent validation is often required by integrating experimental methods, such as site-directed mutations [17,18] . Compared to molecular docking, molecular dynamics simulation can reveal the conformational dynamics of receptor-ligand complexes through nanosecond or longer-scale dynamic modeling. Building based on the results of molecular docking, molecular dynamics simulation are used to verify the stability and accuracy of docked conformations, and enhance the reliability of docking models and molecular recognition mechanisms [19] . This integrated computational strategy dramatically accelerates the development timeline while reducing experimental costs, acting as an increasingly central role in the development of novel therapeutic agents [20] .
Accumulating evidence indicated that dietary intake is a critical role in the disease management of UC by modulating gut microbiota, epithelial barrier function, and mucosal immunity [21] . In a murine colitis model, administration of Porphyra tenera ameliorated disease severity by attenuating intestinal inflammation and improving microbiota dysbiosis [22] . And a case-control study suggested that dietary intake of vitamin C, vegetables, and retinol is linked to a reduced risk of UC [23] . Therefore, dietary intake represents an increasingly vital approach for the prevention and management of UC. Polyphenols are widely distributed in various foods including fruits, vegetables, grains, tea, and coffee, which are common in our daily dietary intake [24] . Owing to their well-documented antioxidant and anti-inflammatory activities, Polyphenols have recently gained increasing attention for their potential in treating UC. For instance, a protective effect on the intestinal tract was exerted by Hydroxytyrosol, a polyphenol abundant in olive oil. This effect was mediated through the activation of the PI3K/Akt-Nrf2 signaling pathway, which attenuated oxidative stress. [25] . Forsythia suspensa polyphenols alleviated symptoms in UC mice, potentially by inhibiting macrophage polarization to the pro-inflammatory M1 phenotype [26] . Besides the antioxidant and anti-inflammation effect of polyphenols, the role of polyphenolic compounds in modulating the gut microbiota has been increasingly reported recently. Epigallocatechin-3-gallate (EGCG), as the major bioactive component of tea polyphenols in green tea, has been confirmed to alleviate symptom of colitis mice via gut microbiota [27] . Despite their broad bioactivity, the precise molecular targets of most polyphenols are poorly defined, which limited the development of their applications in UC therapy.
Herein, we screened out EA as a GPR35 agonist via Nanobit assay from 30 representative phenolic compounds. Then we leveraged computational biology to identify the binding pocket and specific sites for EA with GPR35, and provided comprehensive validation through site-directed mutagenesis. Subsequently, the in vivo studies elucinated that EA could alleviate dextran sulfate sodium (DSS) induced symptoms and promote barrier repair; furthermore, in vitro analyses demonstrated the ability of EA to stimulate intestinal epithelial cell migration and proliferation. Importantly, using the specific GPR35 inhibitor CID2745687 [28] , we confirmed that these protective effects of EA were dependent on GPR35 signaling. In summary, this study integrates cell-based functional assays, computational biology, in vivo and in vitro models to systematically elucidate the molecular mechanism and therapeutic potential of EA as a GPR35 agonist, bridging the gap between dietary compounds and UC therapy.
2. Materials and Methods
2.1. Materials
All phenolic compounds (purity≥98%) was purchased from Nature Standard Co. Ltd(Shanghai, China). The HEK-293T cells were obtained from ATCC (ATCC # CRL-11268), and the normal colonic epithelial cell line NCM460 was provided from Guangzhou Xinyuan Biotechnology Co.Ltd (Guangzhou, China). The Dulbecco’s modified Eagle’s medium (DMEM) was purchased from Thermo Fisher Scientific (Gibco, China), and the fetal bovine serum (FBS) was sourced from Gemini Bio(USA). The liposomal reagent, CTZ-400a and penicillin–streptomycin was provided from Yeasen Biotechnology Co., Ltd(Shanghai, China). The NanoLuc substrate (furimazine) and CID2745687 were provided by Topscience Biochemical Technology Co., Ltd. (Shanghai, China). Dextran sulfate sodium (DSS) was sourced from Dalian Meilun Biotechnology Co.,Ltd (Dalian, China). Hematoxylin and eosin(HE) solution and Carboxymethyl Cellulose-Na(CMC-Na) was obtained from Beijing Solarbio Science & Technology Co., Ltd(Beijing, China). Primary antibody against ZO-1( Zonula Occludens-1) was obtained from Proteintech Group, Inc (Wuhan, China). Sulforhodamine B (SRB) was purchased from Sigma-Aldrich(USA).
2.2. NanoBiT Assay
We engineered a pbit vector encoding full-length human GPR35 (residues 1–309) with an N-terminal FLAG tag and a C-terminal LgBiT, and encoding full-length β-arrestin2 with an N-terminal SmBiT. Eight site-directed mutations (Q93A, L97A, R100A, R151A, F163A, H168A, R240A, and S262A) were introduced into the construct. HEK-293T cells were grown using DMEM plus 10% FBS, under conditions of 37 °C and 5% CO₂ in a humidified environment. For NanoBiT assay, a cell suspension was prepared at a concentration of 3.5 × 10⁵ cells per well and subsequently inoculated into 6-well plates. After 24 h, transfections were performed using 7 μL of liposomal reagent per well, with each well receiving 1.5 μg of either wild-type or mutant GPR35 plasmid together with 1.5 μg of β-arrestin2-encoding plasmid. Following transfection, cells were harvested at the 48-hour time point and then resuspended in Tyrode’s buffer (Coolaber, China). In the screening assay, each well of a 96-well plate received 70 μL of cell suspension, after which 20 μL of ligand solution was added (pre-diluted to fixed concentrations in Tyrode’s buffer) and 10 μL of diluted furimazine. Lodoxamide and phosphate-buffered saline (PBS) served as positive and negative controls, respectively. Luminescence was recorded at 25 °C using a BioTek plate reader, with measurements taken every 30 s over a 15 min interval. For concentration–response experiments, ligands were serially diluted in Tyrode’s buffer and applied as described above. All signals were normalized to vehicle-only controls. Following independent replication in at least three separate experiments, each performed in duplicate, the data (presented as mean ± SEM) were subsequently analyzed with GraphPad Prism 9.0.
2.3. Bret Assay
We engineered a pcDNA vector encoding full-length human GPR35 (residues 1–309) and mouse GPR35 with an N-terminal FLAG tag and a C-terminal LgBiT. HEK-293T cells were grown using DMEM plus 10% FBS, under conditions of 37 °C and 5% CO₂ in a humidified environment. For BRET assays, a cell suspension was prepared at a concentration of 3.5 × 10⁵ cells per well and subse-quently inoculated into 6-well plates. After 24 h, transfections were performed using 7 μL of liposomal reagent per well. Each well received 0.75 μg of wild-type GPR35 plasmid, together with 0.75 μg each of plasmids encoding Gα₁₃, Gβ₁-SmBiT and Gγ₁. After 48 h, cells were collected and resuspended in Tyrode’s buffer (Coolaber, China). For the assay, each well of a 96-well plate received 70 μL of cell suspension, after which 20 μL of ligand solution was added (pre-diluted to fixed concentrations in Tyrode’s buffer) with 10 μL of diluted CTZ-400a. Subsequent steps were performed as described for the NanoBiT assay.
2.3. Molecular Docking
ChemBioDraw Ultra 14.0 was employed to generate the 2D structure of EA, which was subsequently rendered into a 3D model with ChemBio3D Ultra 14.0. AutoDock Tools 1.5.6 was used to convert the resulting structure, which had been saved in mol2 format, into the pdbqt format. The Protein Data Bank (RCSB PDB, http://www.rcsb.org) was the source from which the molecular structure of GPR35 (PDB ID: 8H8J) was obtained in PDB format. The structure was prepared by removing water molecules and existing ligands, adding hydrogen atoms, and performing other necessary adjustments in PyMOL. It was then converted to pdbqt format using AutoDock Tools 1.5.6. In AutoDock Tools 1.5.6, map files were generated and a grid box was defined to encompass the putative binding site, with its center and dimensions adjusted to appropriately cover the active pocket. AutoDock Vina, using the Lamarckian genetic algorithm, was employed to perform the molecular docking. The output docking poses were ranked according to their calculated binding energies, based on the energy scores, the conformation with the most favorable score was designated as representing the optimal binding mode. The highest-scoring pose was visualized using PyMOL.
2.4. Molecular Dynamics Stimulation
Molecular dynamics simulations were performed using GROMACS. Initial structures were prepared by converting protein PDB files to gro format with the AMBER99SB-ILDN force field applied to the protein. Small-molecule ligands were parameterized with the GAFF force field; hydrogen atoms were added and RESP partial charges were derived using Gaussian 16W. Each protein–ligand complex was solvated in a periodic box with TIP3P water molecules, maintaining a minimum distance of 1.2 nm between the protein and the box edges. The system was neutralized by adding Na⁺ ions. Energy minimization was carried out using the steepest-descent algorithm. A 1.0 nm cutoff was used for truncating short-range van der Waals and electrostatic interactions, whereas the particle-mesh Ewald method was employed to handle long-range electrostatics. Following the application of the LINCS algorithm to constrain hydrogen bonds, the system underwent equilibration. This involved a 100 ps NVT simulation prior to a 100 ps NPT simulation conducted at 300 K and 1 bar. Subsequently, a production-phase simulation of 200 ns was carried out under NPT conditions using a 2 fs time step. During this simulation, coordinates, velocities, and energies were recorded at 10 ps intervals for future analysis.
2.5. Animals Experiment
Eighteen male C57BL/6 mice (6 weeks old, 20–22 g) were provided by Beijing Vital River Laboratory Animal Technology Co. Ltd. (Beijing, China). The animal experimental procedure was approved by the Institutional Animal Care and Use Committee (IACUC) of the Zhongshan Institute for Drug Discovery, Shanghai Institute of Materia Medica, Chinese Academy of Sciences (IACUC No. 2024-12-GDA-01). All animals were housed under specific pathogen-free conditions. The environmental parameters were strictly controlled as follows: temperature, 23 ± 3 °C; relative humidity, 55 ± 15%; and a 12-h light/12-h dark photoperiod. The mice were kept three to a cage and provided with food and water ad libitum. After a 7-day acclimation period, they were randomly assigned to the following groups: Control, Model, and EA, with six mice in each group. The Control group received distilled water only. To induce colitis, both the Model and EA groups were administered distilled water containing 3% (w/v) DSS for seven consecutive days [29] . Beginning on the same day as DSS exposure, mice in the EA group were treated daily by oral gavage with EA(60 mg/kg) suspended in 0.5% CMC-Na for 8 days. Control and Model mice received an equal volume of vehicle (0.5% CMC-Na). All animals were maintained on a standard diet. We assessed body weight, stool consistency, and the presence of fecal blood each day, performing these evaluations at a consistent time. Disease activity was assessed using an established disease activity index (DAI) scoring system(Table S1) [30,31] . On day 9, mice were anaesthetized by intraperitoneal injection of Zoletil 50 (30 mg/kg). Following collection from the abdominal aorta into heparin-coated tubes, blood was centrifuged to obtain the plasma component. Following cervical dislocation, the entire colon was removed and its length recorded. The isolated tissue was then gently rinsed with ice-cold saline. Finally, the segment of the distal colon was fixed in 4% paraformaldehyde for subsequent HE-staining and immunofluorescence analysis.
2.6. H&E Staining and Immunofluorescence Analysis
Fresh tissue samples were fixed in fixative solution for a minimum of 24 hours. Following fixation, the specimens were trimmed, dehydrated through a graded ethanol series, cleared in xylene, and embedded in paraffin. Sections were cut at a thickness of 4 μm using a microtome. For histological analysis, deparaffinized sections were stained with H&E staining, mounted with neutral balsam, and scanned in their entirety at 20× magnification using an Olympus SLIDEVIEW VS200 slide scanner. The resulting whole-slide images were evaluated based on a standardized histopathological scoring system shown in Table S2 [32] . For immunofluorescence staining, following pretreatment involving antigen retrieval and blocking with Bovine Serum Albumin (BSA) for 30 minutes, the sections were incubated overnight at 4°C in the dark with an primary antibody (ZO-1,diluted 1:500). Subsequently, a 1-hour incubation with a 488-conjugated secondary antibody, the samples were nuclear-stained with DAPI and the prepared slides were mounted with an anti-fade medium. High-resolution images were acquired using a Leica STELLARIS STED confocal microscope. We performed the quantitative analysis of ZO-1 mean fluorescence density using ImageJ software (ver. 1.5.7 from the National Institutes of Health, USA).
2.7. Wound Healing Assay
Under standard incubator conditions (37 °C, 5% CO₂, humidified), NCM460 cells were cultured in DMEM containing 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin. For the wound healing assay, we seeded cells into 6-well plates at 5×10⁵ cells per well and cultured them until they achieved 90–100% confluence. A sterile 100 µL pipette tip was guided vertically using a straightedge to generate three parallel, uniform scratches in each well. Once detached cells were removed by washing with PBS, the remaining cells received the following treatments in serum-free medium: vehicle control (containing DMSO), 15 µM EA, 10 µM CID2745687, or a combination of both 10 µM CID2745687 and 15 µM EA. Wound images of each group were acquired at 0 h and 24 h using a Nikon microscope (Nikon, Tokyo, Japan) at 20× magnification, and then wound closure was quantified using ImageJ software.
2.8. SRB Assay
At each designated time point (6, 24, 48, and 72 hours), the culture medium was first aspirated. It was then replaced with 50 µL of trichloroacetic acid (TCA) solution at a final concentration of 50% to fix cells. After incubation at 4 °C for 1 h, the fixed cells were washed at least five times with distilled water and allowed to dry completely. For staining, a freshly prepared solution of 0.4% SRB in 1% acetic acid was added to all wells, after which the plates were kept at room temperature, protected from light, for a 15-minute incubation period. Following incubation, the staining solution was removed, and the microtiter plates were washed with 1% acetic acid to eliminate any unbound dye. The protein-bound SRB dye remaining in the wells was then solubilized using a 10 mM Tris base solution. Finally, the absorbance was measured at a wavelength (λ) of 510 nm using an BioTEK plate reader.
2.8. Statistical Analysis
Data analysis was conducted with SPSS software (version 25.0). Post-hoc pairwise comparisons, conducted following one-way ANOVA, employed either the LSD test when the assumption of homogeneity of variance was met, or the Games-Howell test when it was violated. Data are presented as mean ± SEM, with between-group differences quantified using 95% confidence intervals. Statistical significance was assigned to findings where the two-sided p-value fell below 0.05 (SEM).
3. Results
3.1. Agonist Screening of GPR35 from Polyphenols via Nanobit Assay
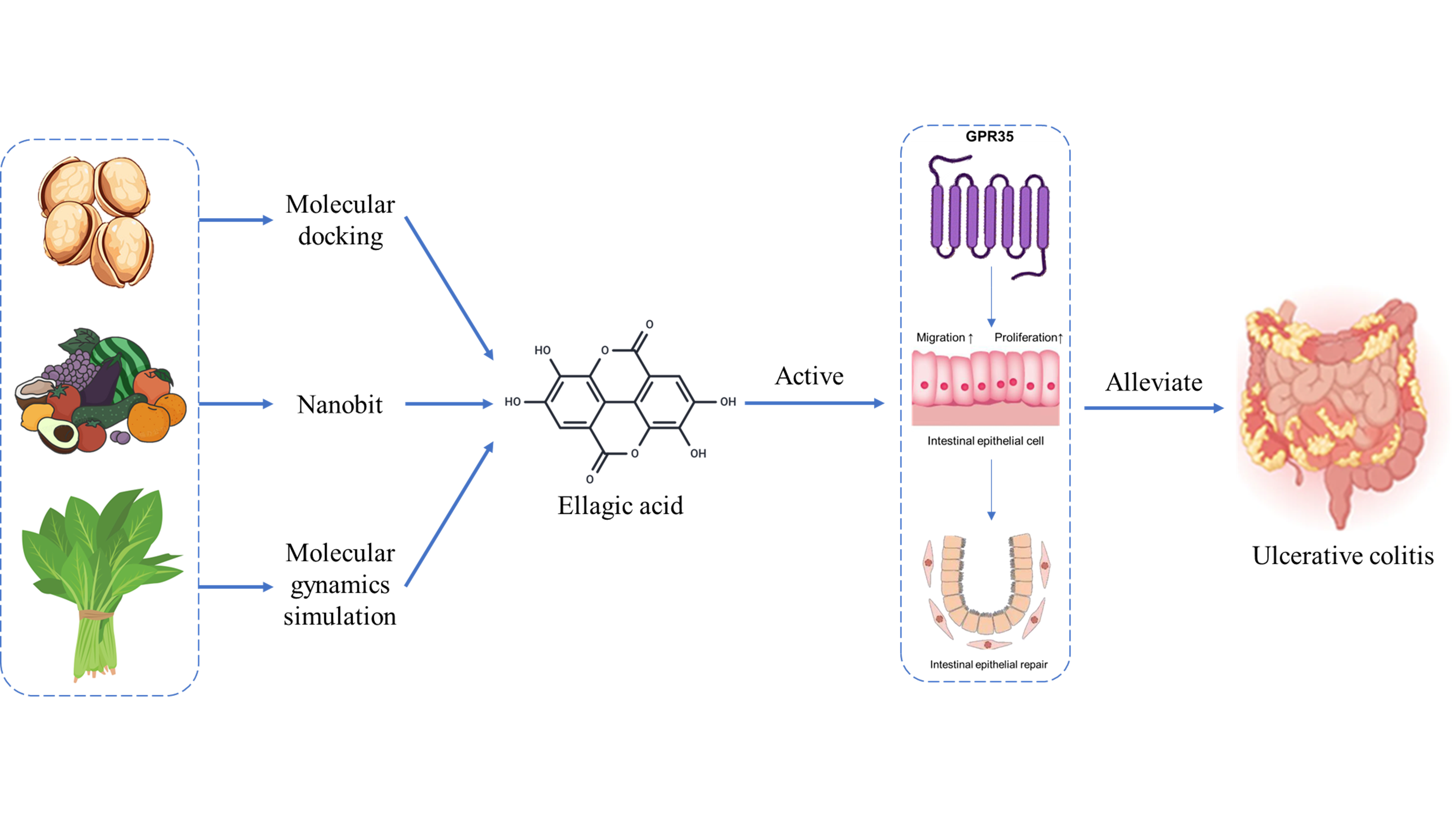
In an effort to discover novel natural agents capable of effectively activating GPR35, we performed a drug screening using the Nanobit assay on 30 common polyphenol primarily sourced from nuts, vegetables, and fruits. β-arrestin proteins and G proteins are major downstream effector proteins of GPCRs, and their recruitment levels can indirectly reflect the activation status of the receptor. Using the Nanobit assay, we examined the recruitment of β-arrestin2 by these 30 compounds and found that EA exhibited the strongest activating effect (Figure.1A). Moreover, through Nanobit experiments, we demonstrated that the activation of hGPR35 by EA was concentration-dependent (Figure.1C). To further confirm the agonist activity of EA, we employed BRET technology, which detects G protein recruitment, and found that EA could recruit Gα13 in a concentration-dependent manner, suggesting its potential as a ligand for hGPR35. Since ligand-induced activation of GPR35 exhibits species-specific effects, we also evaluated the activation of mouse-derived GPR35 by EA. The results indicated that EA can also active mGPR35 in concentration-dependent manner, and the activation of both human and mouse GPR35 exhibited the comparable potency (Figure.1D). Herein, we demonstrated that the dietary polyphenol EA acts as a dose-dependent agonist of GPR35 in both human and murine systems.
Figure 1.
Agonist screening of GPR35 from Polyphenols via Nanobit assay. (A) GPR35 agonist efficacy of 30 polyphenols derived from dietary intake (n = 3). The red bar means the agonist effect of Lodoxamide, a reported potent agonist of GPR35. The purple bar indicates the highest β-arrestin recruitment of EA among the 30 polyphenols.(B) Chemical structures of EA. (C) Dose-dependent β-arrestin2 recruitment activities of EA(n=3). (D) Species-dependent agonist activity at human and mouse GPR35. The red curve indicates the mouse origined GPR35, and the purple curve indicates the human origined GPR35(n=3).
Figure 1.
Agonist screening of GPR35 from Polyphenols via Nanobit assay. (A) GPR35 agonist efficacy of 30 polyphenols derived from dietary intake (n = 3). The red bar means the agonist effect of Lodoxamide, a reported potent agonist of GPR35. The purple bar indicates the highest β-arrestin recruitment of EA among the 30 polyphenols.(B) Chemical structures of EA. (C) Dose-dependent β-arrestin2 recruitment activities of EA(n=3). (D) Species-dependent agonist activity at human and mouse GPR35. The red curve indicates the mouse origined GPR35, and the purple curve indicates the human origined GPR35(n=3).
3.2. Molecular Docking
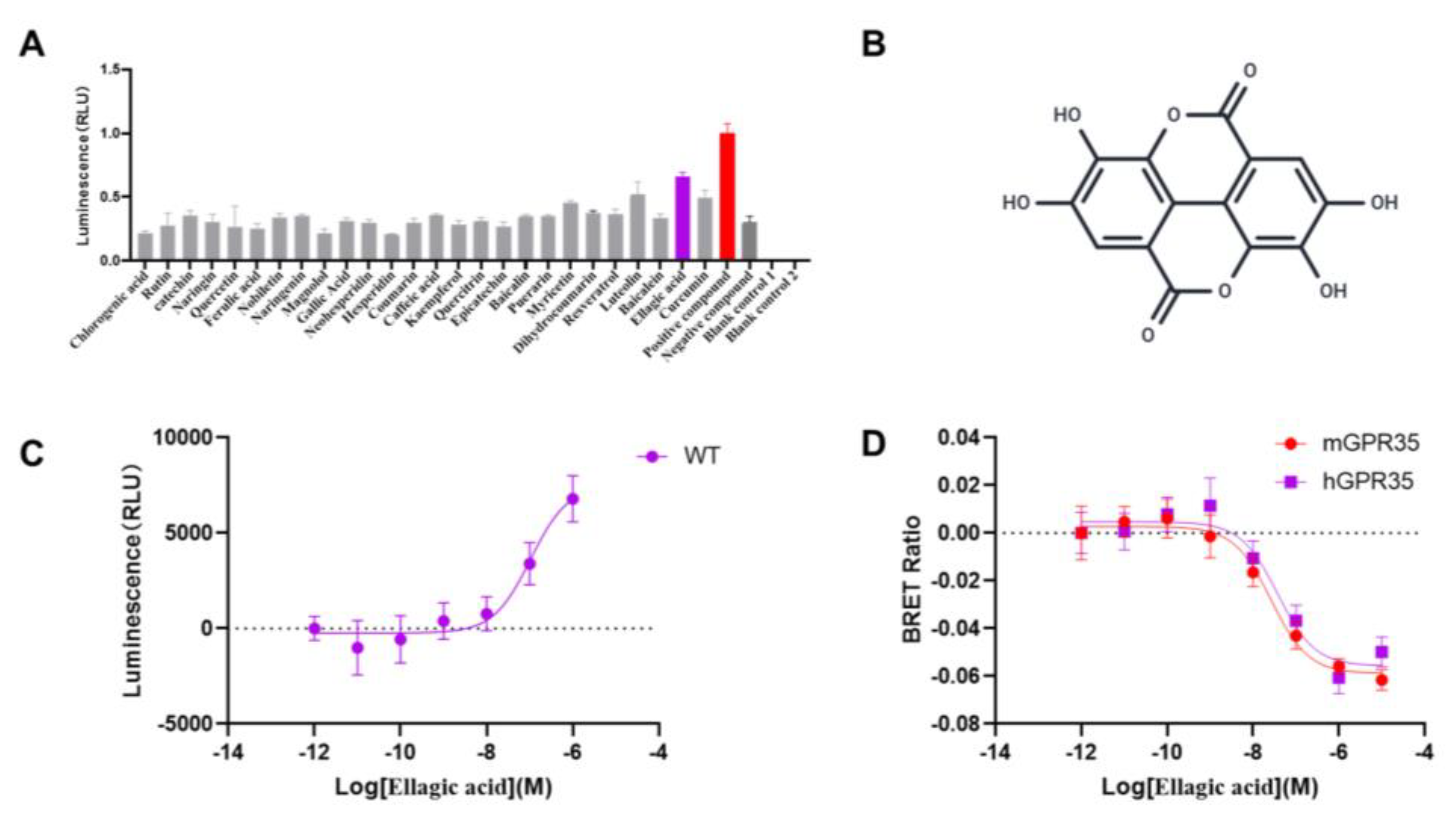
This docking conformation exhibited a strong binding affinity, with a calculated energy of -8.709 kcal/mol. The surface model shown in Figure.2A-B demonstrated that the ligand is snugly embedded within a cavity on the protein surface, a configuration that is complementary to the ligand's structure and favorable for stable binding. As shown in Figure.2C, five hydrogen bonds (including Gln93, Arg100, Phe163, Arg240, Ser262) were formed between the ligand and the protein’s amino acid residues, according to the three-dimensional structural analysis. Specifically, a hydrogen bond with a length of 2.29 Å was observed between the ligand and the residue of Gln93 and Arg100, indicating that these residues constitute key binding sites. The two-dimensional interaction diagram further illustrated that van der Waals forces from residues of Leu97 surround the ligand, which are essential for stabilizing the binding site. Additionally, two π-interactions (e.g., π-cation, π-π stacking, etc.) were identified between the ligand and the residues Arg151 and Phe163, further influencing the binding affinity and specificity. Furthermore, the ligand participates in a stable electrostatic network by forming salt bridges with three key positively charged residues, including Arg151, His168, and Arg240. In summary, a relatively strong and specific interaction between the EA and the GPR35 was indicated by the molecular docking results.
Figure 2.
Molecular docking model of EA in the GPR35-binding site. (A) 3D docking pose of EA within the GPR35 binding site. The GPR35 structure is visualized as a yellow ribbon. An enlarged view illustrates EA's positioning within the binding pocket. (B) The binding pocket of EA occupied in GPR35. (C)Interaction model of GPR35 and EA.
Figure 2.
Molecular docking model of EA in the GPR35-binding site. (A) 3D docking pose of EA within the GPR35 binding site. The GPR35 structure is visualized as a yellow ribbon. An enlarged view illustrates EA's positioning within the binding pocket. (B) The binding pocket of EA occupied in GPR35. (C)Interaction model of GPR35 and EA.
3.3. Functional Analysis of Wild-Type and Mutant Receptors
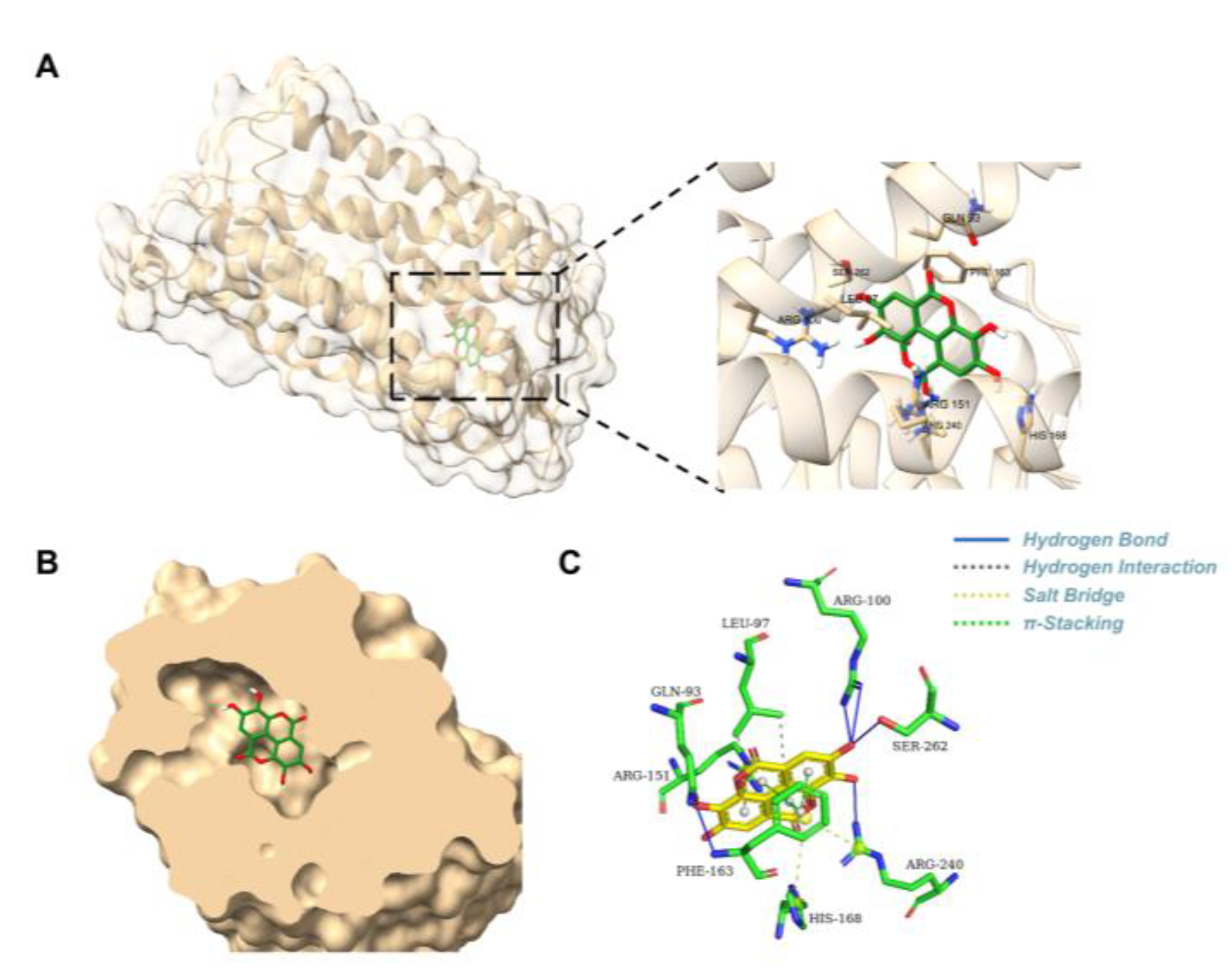
To experimentally validate the predicted complex of EA binding to GPR35, the site-directed mutagenesis was performed followed by molecular docking. In site-directed mutations, the key amino acid residue was mutated to smaller residues (Ala or Gly), resulting in the loss of their original interactions. Therefore the site-directed mutations (Q93A, L97A, R100A, R151A, F163A, H168A, R240A, S262A) were generated and evaluated for receptor activation using a Nanobit assay. Compared with wild-type (WT) GPR35, the Q93A, R100A, R151A, F163A and S262A mutations completely abolished EA-induced receptor activation. The L97A and R240A mutants reduced the maximal activation efficacy (ECmax) of EA by approximately twofold, indicative of substantially diminished binding affinity. In contrast, the H168A mutant exhibited activation levels comparable to WT, suggesting that His168 plays an not critical residue for EA binding to GPR35. Collectively, these results identify Gln93, Arg100, Arg151, Phe163 and Ser262 as critical residues for EA binding and activation of GPR35, while Leu97 and Arg240 appear to play auxiliary roles in the interaction. Our findings provide experimental support for the computationally predicted binding site and mode of action of EA on GPR35.
Figure 3.
Functional analysis of wild-type and mutant receptors. (A)Model showing key hydrophobic residues (Q93, R100, F168 and R240) interact with EA in the binding pocket. (B)Model showing key hydrophobic residues (L97, R151, H163 and S262) interact with EA in the binding pocket. (C-D) Nanobit assay showing the response of wild-type and mutant GPR35 receptors to EA.
Figure 3.
Functional analysis of wild-type and mutant receptors. (A)Model showing key hydrophobic residues (Q93, R100, F168 and R240) interact with EA in the binding pocket. (B)Model showing key hydrophobic residues (L97, R151, H163 and S262) interact with EA in the binding pocket. (C-D) Nanobit assay showing the response of wild-type and mutant GPR35 receptors to EA.
3.4. Molecular Dynamics Simulation of GPR35 and EA
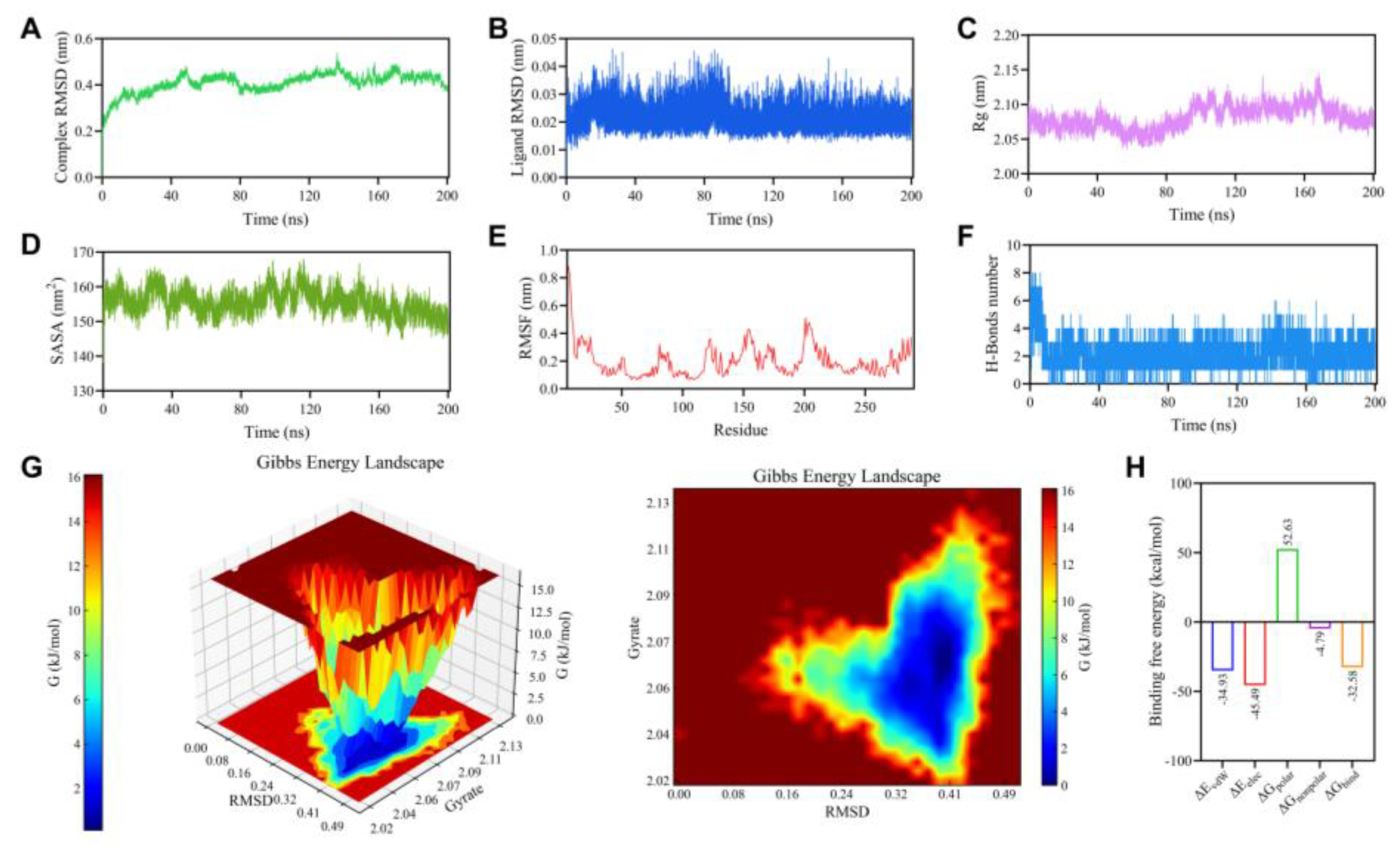
Molecular dynamics simulations provide a means to monitor, in real time, the binding stability and dynamic interactions within small molecule–protein complexes. As shown in Figure. 4A-B, RMSD analysis revealed a rapid increase for the complex in the first 20 ns of simulation, after which the fluctuations diminished, suggesting the attainment of dynamic equilibrium. As shown in Figure. 4C, the Rg value showing only minor oscillations over time without any systematic upward or downward trend, and the SASA values exhibiting slight oscillations over time but no abrupt changes, suggesting that a modest compaction or rearrangement of hydrophobic regions within the protein during the simulation may facilitate tighter ligand binding, thereby contributing to the maintenance of a stable configuration. To investigate the effect of ligand binding on the flexibility of protein residues, we calculated the RMSF of each amino acid over the entire simulation. As illustrated in Figure. 4E, most residues exhibited fluctuations below 0.3 nm. Slightly higher fluctuations were observed in certain regions, predominantly at the N-/C-termini or loop regions, while the core structural regions and potential binding sites displayed restricted mobility. Hydrogen bonds represent strong non-covalent interactions, and their number was therefore used to assess the stability of the protein–ligand complex during the simulation. To further examine hydrogen-bond dynamics, we statistically analyzed intermolecular hydrogen bonds formed between the ligand and the protein over the 200 ns trajectory. As shown in Fig. 4F, the system maintained approximately 2–4 hydrogen bonds throughout the simulation, with occasional higher values at certain time points, and no prolonged periods of complete absence were observed. This consistent presence of multiple hydrogen bonds suggests a stable binding interface. The free-energy landscape (FEL) projected onto the RMSD–Rg reaction coordinates revealed a single, well-defined low free-energy basin for the protein–ligand complex (Figure. 4G). The global minimum was concentrated in the region corresponding to RMSD values of approximately 0.32–0.41 nm and Rg values of approximately 2.04–2.08 nm, indicating that the conformation corresponds to a state of relatively high stability. The binding between the protein and ligand was primarily driven by van der Waals and electrostatic forces, with the latter making a notably dominant contribution, as revealed by MM/PBSA binding free energy decomposition(Figure. 4H). The final calculated binding free energy was −32.58 kcal/mol. All molecular dynamics simulation results collectively demonstrate that the GPR35–EA complex structure predicted by molecular docking exhibits robust stability.
Figure 4.
Analysis of 200 ns molecular dynamics simulations of the complex. (A) RMSD(complex). (B)RMSD (ligand). (C)Rg. (D)SASA. (E)RMSF. (F)Protein-ligand H-bonds. (G)Free energy landscape. (H) Binding free energy.
Figure 4.
Analysis of 200 ns molecular dynamics simulations of the complex. (A) RMSD(complex). (B)RMSD (ligand). (C)Rg. (D)SASA. (E)RMSF. (F)Protein-ligand H-bonds. (G)Free energy landscape. (H) Binding free energy.
3.5. EA Administration Ameliorates DSS-Induced Colitis in Mice
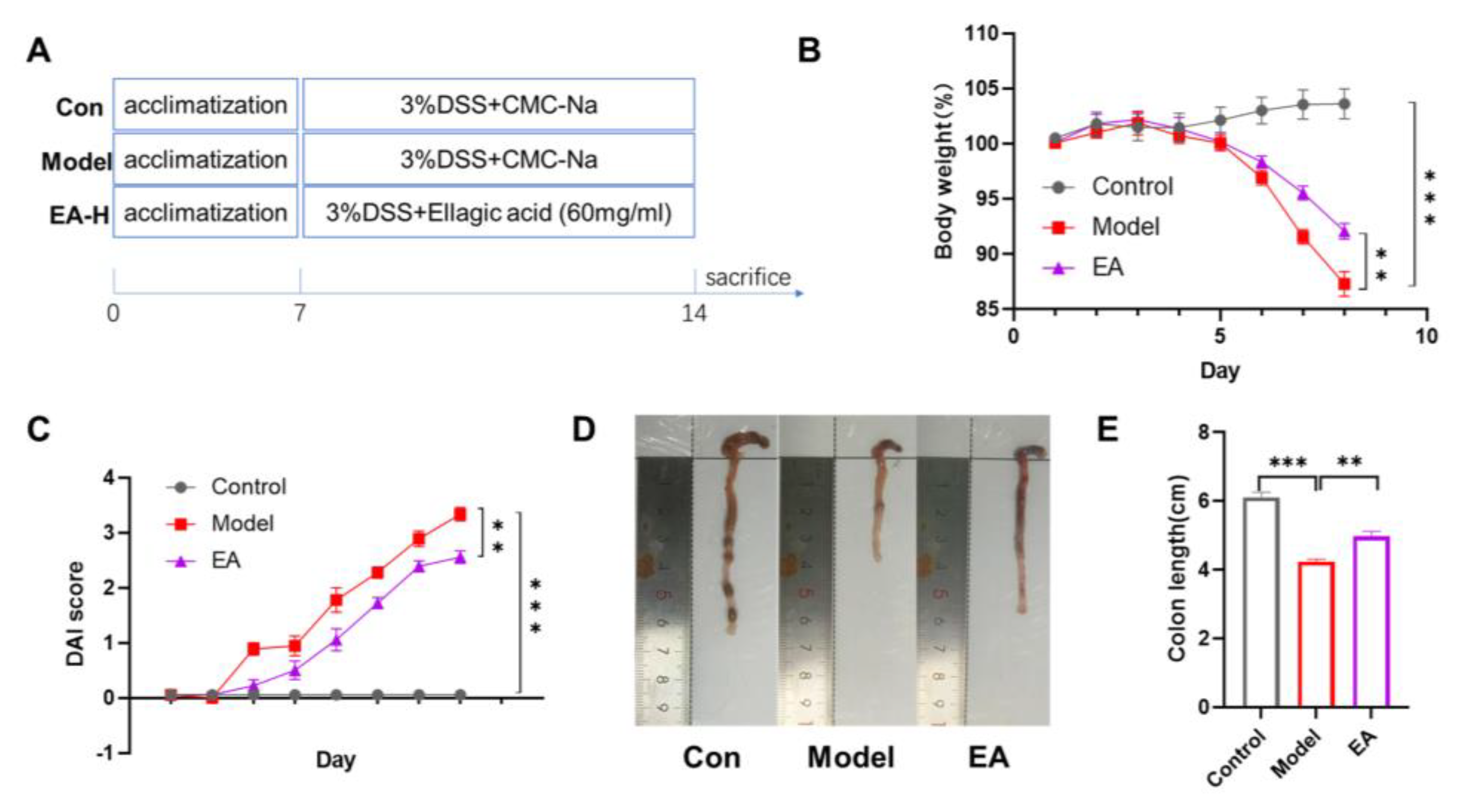
In this study, the therapeutic potential of EA for colitis was evaluated by employing a DSS-induced mouse model. This DSS-induced colitis model is widely employed in drug screening for UC treatment owing to its close pathological resemblance to human UC. Mice in the model group receiving 3% DSS in drinking water exhibited progressive weight loss (p*** < 0.001). Disease severity, assessed by the DAI score, showed a significant increase in the model group (p*** < 0.001). In contrast, EA treatment markedly attenuated DSS-induced intestinal damage, as evidenced by a significant reduction in weight loss and a substantially lower DAI score (p** < 0.01). Beyond these symptoms, colon shortening serves as a hallmark structural alteration in colitis. This phenomenon is associated with DSS-mediated disruption of the intestinal epithelial barrier, sustained inflammatory infiltration, and subsequent tissue fibrosis leading to colon wall contraction. Results demonstrated that colon length was significantly shorter in DSS-treated mice compared to the control group (p*** < 0.001). EA intervention significantly ameliorated this shortening, with colon length notably greater than that in the model group (p** < 0.01). Collectively, our results indicate that EA protects mice from DSS-induced colitis, markedly attenuating disease symptoms.
Figure 5.
EA administration ameliorates DSS-induced colitis in mice. (A)Schematic diagram of the experimental design. (B) Time course of body weight. (C) Change in DAI score. (D) Representative colon images from each group. (E)Colon length of each group. Data are shown as mean±SEM. **p < 0.01, ***p < 0.001 versus Model group.
Figure 5.
EA administration ameliorates DSS-induced colitis in mice. (A)Schematic diagram of the experimental design. (B) Time course of body weight. (C) Change in DAI score. (D) Representative colon images from each group. (E)Colon length of each group. Data are shown as mean±SEM. **p < 0.01, ***p < 0.001 versus Model group.
3.6. EA Preserves Colonic Structure and Barrier Function
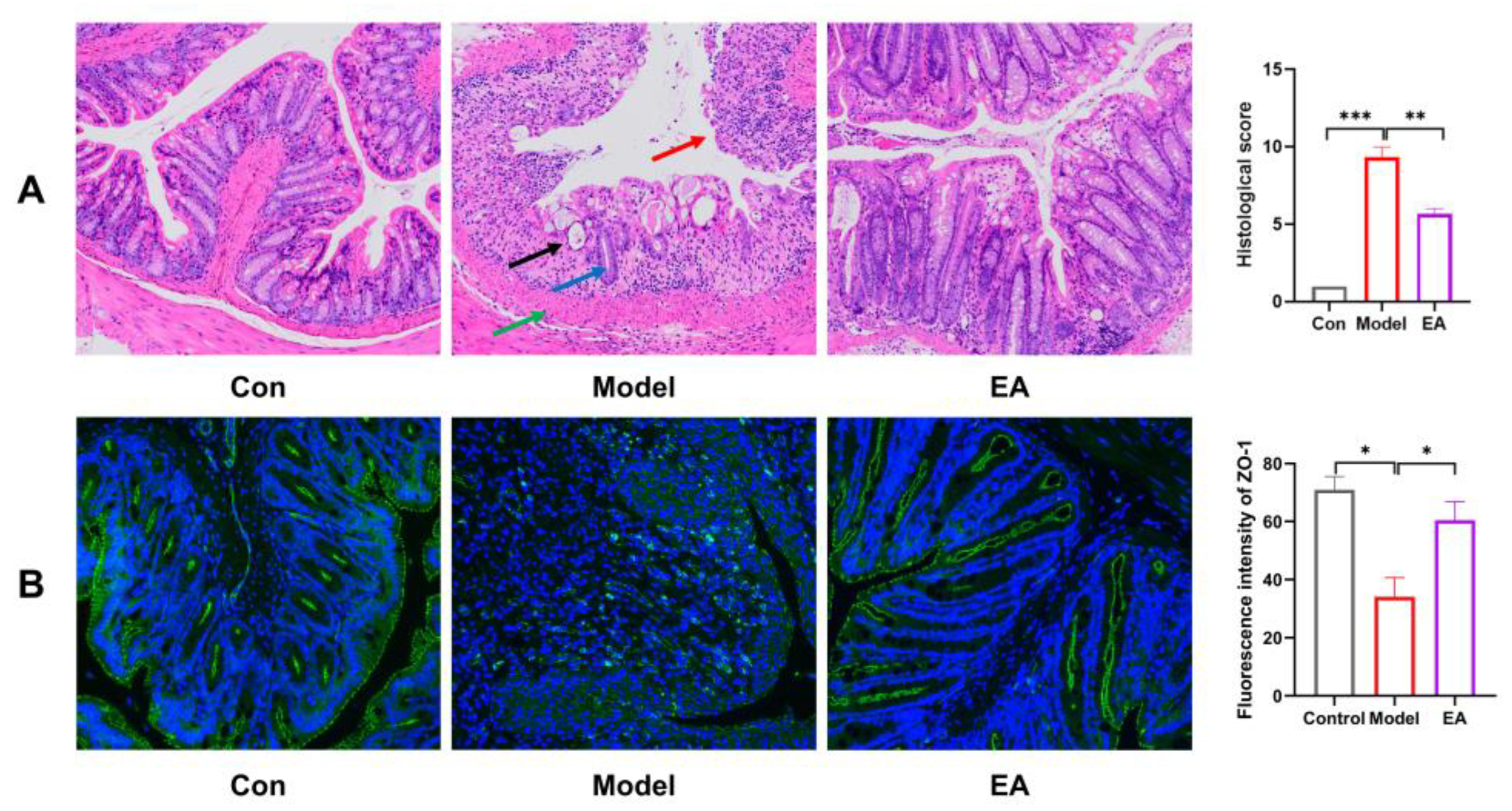
The disruption of intestinal epithelial cells is often considered the primary cause of recurrent colitis, and the function of the biological barrier is closely linked to intestinal epithelial repair and tight junction proteins. First, histological changes in colonic tissue were assessed by H&E staining. As illustrated, mice in the model group displayed pronounced pathological alterations compared with control group, including loss of intestinal epithelial cells (red arrow), disappearance of crypts (black arrow), widespread reduction in goblet cell numbers (blue arrow), and extensive inflammatory cell infiltration into the mucosal layer accompanied by muscularis mucosa thickening (green arrow). In contrast, treatment with EA significantly ameliorated colon damage, particularly promoting repair of the intestinal epithelium. Furthermore, the expression of ZO-1, a crucial tight junction protein that participates in maintaining tight junctions and barrier function between cells, was examined by immunofluorescence staining. The results indicated that A significant decrease in ZO-1 expression was observed in the model group compared to the control group(*P < 0.05). EA treatment markedly restored the downregulated expression of ZO-1(*P < 0.05). Together, these findings suggest that EA contributes to the preservation of intestinal barrier integrity by mitigating epithelial damage and enhancing the expression of key tight-junction proteins in a mouse model of colitis.
Figure 6.
EA preserves colonic structure and barrier function. (A) H&E staining of each group(20×)and Histopathological assessment. (B) Immunofluorescence of ZO-1 in each group(200×). Data were presented as mean ± SEM. *p <0.05, **p < 0.01, ***p < 0.001 versus Model group.
Figure 6.
EA preserves colonic structure and barrier function. (A) H&E staining of each group(20×)and Histopathological assessment. (B) Immunofluorescence of ZO-1 in each group(200×). Data were presented as mean ± SEM. *p <0.05, **p < 0.01, ***p < 0.001 versus Model group.
3.7. EA promotes the Repair of the Intestinal Epithelium via Activating GPR35
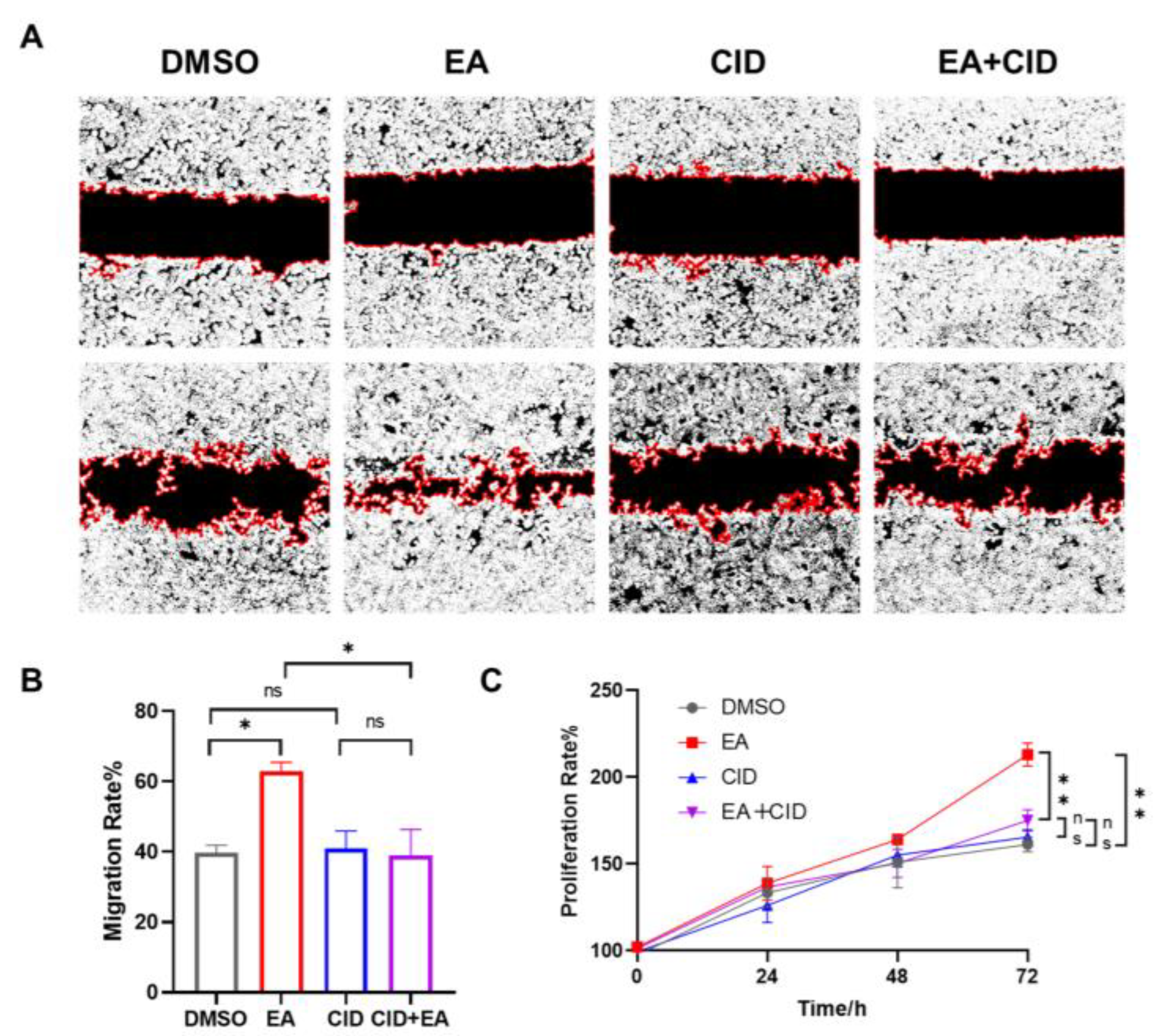
To investigate the role of GPR35 in epithelial repair, we then assessed the effect of EA on wound healing using a scratch assay. As shown in Figure.7B, compared to the DMSO treated control, EA significantly enhanced the migration of NCM460 cells over 24 hours(*p <0.05). The specific GPR35 antagonist CID2745687, when added alone, did not affect basal cell migration compared to the DMSO control. However, the pro-migratory effect of 20 µM EA was completely abolished by co-treatment with CID2745687. Consequently, the migration capacity in the EA+CID2745687 co-treatment group showed no significant difference from the CID2745687-alone group but was significantly lower than that in the EA-only group(*p<0.05). The effect of EA on cell proliferation was further evaluated by using the SRB assay. As illustrated in Figure.7C, treatment with EA significantly increased the proliferation rate of NCM460 cells compared to the DMSO control (**p < 0.01). In contrast, the GPR35 antagonist CID2745687 alone had no measurable effect on proliferation. Notably, co-treatment with EA and CID2745687 resulted in a proliferation level comparable to that of the CID2745687-alone group and was significantly reduced relative to the EA-only group(**p < 0.01). Taken together, these results collectively demonstrate that EA enhances intestinal epithelial cell migration and proliferation through specific activation of GPR35, thereby revealing a key mechanism underlying its promotion of epithelial repair at a functional level.
Figure 7.
EA promotes the repair of the intestinal epithelium via activating GPR35. (A) Representative images of Wound Healing Assay. (B) The effect of EA on promoting migration of intestinal epithelium cells via GPR35. (C) The effect of EA on promoting proliferation of intestinal epithelium cells via GPR35. Data were presented as mean±SEM.*p <0.05, **p < 0.01 versus Model group, ns>0.05.
Figure 7.
EA promotes the repair of the intestinal epithelium via activating GPR35. (A) Representative images of Wound Healing Assay. (B) The effect of EA on promoting migration of intestinal epithelium cells via GPR35. (C) The effect of EA on promoting proliferation of intestinal epithelium cells via GPR35. Data were presented as mean±SEM.*p <0.05, **p < 0.01 versus Model group, ns>0.05.
4. Discussion
With the attribution to the Westernization of lifestyles, the prevalence in Asia, Latin America, and Africa experiencing a 5 to 10-fold surge in UC incidence [33,34] . Unfortunately, current treatments for colitis are often associated with side effects, high costs, and a tendency for recurrence [35] . Consequently, the role of dietary intake in managing UC is attracting growing attention as a potential therapeutic method [36] . Polyphenols are plant-derived bioactive compounds commonly found in foods like vegetables, fruits, and nuts. Their demonstrated anti-inflammatory, antioxidant, and microbiota-regulating effects are driving their growing incorporation into functional food development [37] . Notably, bioactive components in breast milk and infant formula have been shown to activate GPR35, thereby underscoring its potential as a target for dietary interventions [38] . Morever, GPR35 has emerged as a potential therapeutic target for UC treatment, with reported efficacy in promoting intestinal epithelial repair [39] . However, whether dietary components can act as ligands for GPR35 to ameliorate UC remains largely unexplored.
Thus, the NanoBiT assay, a method commonly used for GPCR ligand screening, was applied to screen polyphenolic compounds targeting GPR35 [40] . In this study, EA, a widely available dietary polyphenol, was identified as a novel and potent natural agonist of GPR35 from thirty dietary polyphenolic compounds. Interestingly, EA has found broad application in wound healing and tissue regeneration, but its effects on intestinal epithelial restitution remain unexplored [41,42] . So, the significant protective role of EA in DSS-induced colitis was highlighted in our vivo experiment, which was primarily mediated through the promotion of intestinal epithelial repair. Moreover, this protective effect was further verified to be dependent on GPR35 activation in the vitro studies, leading to enhanced migration and proliferation of intestinal epithelial cells. These findings link dietary polyphenols to GPCR-mediated intestinal epithelial protection, establishing a molecular basis for the health benefits of polyphenol-rich foods.
Owing to the rapid development of structural biology, the computational biology methods were extensively employed to not only drug screen, but also the discovery of binding model of the ligand-protein complex [43] . Studies demonstrate that astragaloside IV alleviated fructose-induced intestinal metabolic ageing by directly inhibiting ketohexokinase (KHK) activity. Mechanistically, molecular docking and site-directed mutagenesis studies revealed that an Asn261 mutation in KHK impairs enzymatic function and diminishes the therapeutic efficacy of astragaloside IV [44] . Hence, to elucidate the binding model of the EA–GPR35 complex, molecular docking was employed to characterize the binding pocket and interaction sites. We found that EA occupied the orthosteric binding pocket of GPR35, with the binding energy of -8.709 kcal/mol . Broadly aligning with the predicted outcomes, the site-directed mutation confirmed the Gln93, Arg100, Arg151, Phe163 and Ser262 was the key binding-site between EA and GPR35, and the residues Leu97 and Arg240 may play an accessory role within the complex. In addition, consistent with the results of the molecular docking, the stability of the complex of GPR35 and EA was further confirmed by 200ns molecular dynamics simulations. These integrated computational approaches enhance the validity of the findings and thereby establish a foundation for subsequent structure-based compound refinement.
EA, a naturally occurring polyphenolic compound abundant in foods such as grape seeds, tea, and nuts, exhibits a wide range of biological activities, demonstrating considerable potential for health-promoting applications [45] . Aligning with the conclusions of Li,X et al [46] , EA alleviated the symptoms of colitis rats, including weight loss, DAI score and shortened colon length. And the damage to the intestinal epithelium and barrier function was significantly ameliorated following EA treatment. Recently, the therapeutic potential of EA in colitis has attracted growing interest. Research indicated that EA alleviates colitis symptoms by suppressing inflammatory responses and inhibiting NLRP3 inflammasome activation [47] . Furthermore, studies also reported that by scavenging reactive oxygen species, hydrogel-encapsulated EA nanoparticles augment their colonic retention and confer protection against DSS-induced colitis [48] . Notably, among its biological activities, the modulation of gut microbiota by EA has been extensively studied. Evidence suggests that Streptococcus abundance is closely associated with intestinal immune responses, and pretreatment with EA reduces the abundance of intestinal Streptococcus [49] . And treatment with EA restored Lactobacillus abundance while reducing the levels of Bacteroides and Escherichia coli, protecting the intestine from potentially harmful gut bacteria and fungi [50] . Although the repair of the intestinal epithelium (epithelial restitution) is a critical therapeutic target in colitis, the role of EA in promoting intestinal epithelium repair remains underexplored.
Interestingly, it has been validated that the activation of GPR35 can promote intestinal epithelial repair. Studies indicate that GPR35 agonists promote intestinal epithelial repair through a mechanism involving Gi protein recruitment, activation of the ERK1/2 signaling pathway, and subsequent upregulation of fibronectin and integrin α5 expression [51] . During intestinal epithelial repair, IECs near the wound lose their polarity, and the microvilli on the cell surface disappear. Subsequently, they form protrusions that extend and migrate toward the damaged area, thereby facilitating wound closure [52] . The rapid proliferation of IECs replenishes the lost epithelial cells and likewise plays a key role in the overall repair process of the intestinal epithelium [53] . Wound healing assay and SRB assays revealed that EA significantly enhanced intestinal epithelial cell migration and proliferation, but these promotive effects were blocked by the GPR35-specific inhibitor CID2745687, whereas the inhibitor alone showed no effect. In vitro studies demonstrate that EA promotes the proliferation and migration of intestinal epithelial cells, which is dependent on GPR35 activation. These findings reveal its potential in promoting epithelial repair and provide a new direction for developing functional foods targeting GPR35.
Nevertheless, several questions and limitations warrant consideration. First, the pharmacokinetics of EA, including its bioavailability and the nature of its bioactive metabolites in the gut, require further investigation to translate these findings into practical dietary recommendations. The observed effects might be mediated not only by EA itself but also by its gut microbiota-derived metabo-lites, such as urolithins, whose activity on GPR35 is currently unknown. Second, to substantiate the presious binding mode, the application of cryo-electron microscopy (cryo-EM) analysis of the EA–GPR35 complex would greatly strengthen the mechanistic interpretation of their interaction, and this high-resolution structure obtained by cryo-EM can provides a precise blueprint for the targeted modification of compounds. Third, employing genetic tools such as GPR35 knockout mice in vivo would provide definitive validation of target engagement and further elucidate the receptor-dependent pathways underlying EA-mediated epithelial repair. This study provides a screening strategy for identifying dietary compounds that target the intestinal epithelial repair protein GPR35. Future studies will screen the metabolites derived from ellagic acid for GPR35 targeting, further employ cryo-electron microscopy to validate its binding site, and utilize gene-knockout models to confirm target engagement and delineate the associated signaling pathways.
5. Conclusions
In summary, EA was identified as a natural agonist of GPR35 from 30 polyphenolic compound via Nanobit assay. EA is demonstrated to activate both human and murine GPR35 with equivalent potency, leading to the recruitment of Gα13 and β-arrestin2. The binding mode and key interaction sites of EA with GPR35 were predicted by molecular docking and substantiated through site-directed mutagenesis. Residues Q93A, R100A, F163A and S262A , which form hydrogen bonds with EA, were shown to play critical roles in receptor activation, along with the hydrophobic contribution of Leu97. Additional support was provided by Arg240 (hydrogen bond) and Arg151 (salt bridge). These predictions were further validated by molecular dynamics simulations, confirming the stable binding of EA to GPR35. Functionally, EA was found to alleviate symptoms in a murine colitis model, consistent with its known anti-inflammatory, antioxidant, and microbiota-modulating properties. Notably, a previously unreported role of EA in promoting intestinal epithelial migration and proliferation was uncovered in vitro. This pro-repair effect was reversed by the GPR35 antagonist CID2745687, underscoring the specificity of EA action through GPR35 activation. Collectively, these results highlight the protective role of EA in colitis and provide a mechanistic rationale for the development of EA-rich foods as potential dietary interventions for intestinal disorders.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Table S1: Disease activity index (DAI) and Table S2: Histopathological Score;.
Author Contributions
Conceptualization, D.A.; methodology, D.W. and X.L.; software, D.W.; validation, L.Y. and X.L.; formal analysis, H.L.; investigation, H.L., L.Y. and X.M.; resources, Z.L.; data curation, G.W.; writing—original draft preparation, L.Y.; writing—review and editing, H.L.; visualization, X.M., G.W. and D.W.; supervision, X.L.; project administration, D.A. and Z.L.; funding acquisition, D.A. and Z.L.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Guangdong Basic and Applied Basic Research Foundation (2023A1515111180), National Natural Science Foundation of China (32141005), and Key Program of National Natural Science Foundation of China (82130111). “.
Institutional Review Board Statement
The animal experimental procedure was approved by the Institutional Animal Care and Use Committee (IACUC) of the Zhongshan Institute for Drug Discovery, Shanghai Institute of Materia Medica, Chinese Academy of Sciences (IACUC No. 2024-12-GDA-01).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviations
The following abbreviations are used in this manuscript:
| UC | Ulcerative Colitis |
| EA | Ellagic acid |
| GPR35 | G Protein-Coupled Receptor 35 |
| ZO-1 | Zonula Occludens-1 |
| GPCRs | G protein-coupled receptors |
| IECs | Intestinal epithelial cells |
| ZO-1 | Zonula Occludens-1 |
References
- Hashash, JG; Limdi, JK; Shapiro, JM; Shah, SA. Medical management of inflammatory bowel diseases. BMJ 2025, 391, e079050. [Google Scholar] [CrossRef]
- Tao, C; Luo, F; Wang, Y; Gao, X; Cao, Y; Wang, K; et al. Mannose-modified Prunus persica kernel protein nanoparticles loading baicalin coated with Lycium barbarum polysaccharide for ulcerative colitis treatment. Carbohydr Polym 2026, 371, 124489. [Google Scholar] [CrossRef]
- Huang, J; Liu, Z; Liu, Y; Liu, X; Tian, Z; Yao, M; et al. Butylchlorogenate from Chaenomeles speciosa alleviates ulcerative colitis via suppressing NLRP3 inflammasome activation and remodeling intestinal homeostasis. Phytomedicine 2025, 148, 157282. [Google Scholar] [CrossRef] [PubMed]
- Fudman, DI; McConnell, RA; Ha, C; Singh, S. Modern Advanced Therapies for Inflammatory Bowel Diseases: Practical Considerations and Positioning. Clin Gastroenterol Hepatol 2025, 23, 454–68. [Google Scholar] [CrossRef]
- Liang, Y; Li, Y; Lee, C; Yu, Z; Chen, C; Liang, C. Ulcerative colitis: molecular insights and intervention therapy. Mol Biomed 2024, 5, 42. [Google Scholar] [CrossRef]
- Xu, Q; Huang, K; Shi, Y; Song, Y; Ying, T; Yang, Z. Therapeutic antibodies targeting G protein-coupled receptors. Trends Pharmacol Sci 2026. [Google Scholar] [CrossRef]
- Essa, K; Ortiz, ZN; Roth, S; Xin, BT; de Winter, S; van der Horst, C; et al. Leveraging Targeted Protein Degradation for G Protein-Coupled Receptors: The Development of CCR2 Molecular Degraders. J Med Chem 2025, 68, 26525–46. [Google Scholar] [CrossRef] [PubMed]
- Otkur, W; Wang, J; Hou, T; Liu, F; Yang, R; Li, Y; et al. Aminosalicylates target GPR35, partly contributing to the prevention of DSS-induced colitis. Eur J Pharmacol 2023, 949, 175719. [Google Scholar] [CrossRef] [PubMed]
- Elias, JE; Debela, M; Sewell, GW; Stopforth, RJ; Partl, H; Heissbauer, S; et al. GPR35 prevents osmotic stress induced cell damage. Commun Biol 2025, 8, 478. [Google Scholar] [CrossRef]
- Wu, Y; Zhang, P; Fan, H; Zhang, C; Yu, P; Liang, X; et al. GPR35 acts a dual role and therapeutic target in inflammation. Front Immunol 2023, 14, 1254446. [Google Scholar] [CrossRef]
- Bi, Z; Li, H; Liang, Y; Sun, D; Liu, S; Chen, W; et al. Emerging paradigms for target discovery of traditional medicines: A genome-wide pan-GPCR perspective. Innovation (Camb) 2025, 6, 100774. [Google Scholar] [CrossRef] [PubMed]
- Guo, S; Zhao, T; Yun, Y; Xie, X. Recent progress in assays for GPCR drug discovery. Am J Physiol Cell Physiol 2022, 323, C583–94. [Google Scholar] [CrossRef] [PubMed]
- Duan, J; Liu, Q; Yuan, Q; Ji, Y; Zhu, S; Tan, Y; et al. Insights into divalent cation regulation and G(13)-coupling of orphan receptor GPR35. Cell Discov 2022, 8, 135. [Google Scholar] [CrossRef]
- de Angelo, RM; Nascimento, LA; Encide, J; Barbosa, H; Ghilardi, LJ; Da, SEF; et al. Advances and Challenges in Molecular Docking Applied to Neglected Tropical Diseases. Curr Med Chem 2025, 32, 5939–59. [Google Scholar] [CrossRef]
- Dewaker, V; Prabhakar, YS. Molecular Dynamics Simulations of HDAC-ligand Complexes Towards the Design of New Anticancer Compounds. Curr Top Med Chem 2023, 23, 2743–64. [Google Scholar] [CrossRef]
- Aguiar, C; Camps, I. Molecular Docking in Drug Discovery: Techniques, Applications, and Advancements. Curr Med Chem 2025, 32, 5924–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y; Ma, Z; Li, G; Meng, X; Duan, S; Liu, Z; et al. The Structural Deciphering of the α3 Helix Within ZmHsfA2'S DNA-Binding Domain for the Recognition of Heat Shock Elements in Maize. Plants (Basel) 2025, 14. [Google Scholar] [CrossRef]
- Ning, Q; Yang, P; Guan, Y; Cai, Y; Chen, D; Gao, Y. Deoxycholic Acid Promotes Persistent Low-Level Viremia in Chronic Hepatitis B via Enhancing HBV Particle Formation. J Gastroenterol Hepatol 2025, 40, 2997–3008. [Google Scholar] [CrossRef]
- Wu, X; Xu, LY; Li, EM; Dong, G. Application of molecular dynamics simulation in biomedicine. Chem Biol Drug Des 2022, 99, 789–800. [Google Scholar] [CrossRef]
- Wang, Y; Li, Y; Chen, J; Lai, L. Modeling protein-ligand interactions for drug discovery in the era of deep learning. Chem Soc Rev 2025, 54, 11141–83. [Google Scholar] [CrossRef]
- Kong, C; Yan, X; Liu, Y; Huang, L; Zhu, Y; He, J; et al. Ketogenic diet alleviates colitis by reduction of colonic group 3 innate lymphoid cells through altering gut microbiome. Signal Transduct Target Ther 2021, 6, 154. [Google Scholar] [CrossRef]
- Yi, L; Han, Y; Shen, P; Du, H; Guo, X; Zhou, Z; et al. Dietary Porphyra tenera ameliorated dextran sodium sulfate-induced colitis in mice via modulating gut microbiota dysbiosis. Food Chem 2024, 461, 140832. [Google Scholar] [CrossRef]
- Miyake, Y; Tanaka, K; Nagata, C; Furukawa, S; Andoh, A; Yokoyama, T; et al. Dietary intake of vegetables, fruit, and antioxidants and risk of ulcerative colitis: A case-control study in Japan. Nutrition 2021, 91-92, 111378. [Google Scholar] [CrossRef]
- Zhong, X; Dai, Q; Li, J; Yang, J; Zheng, X. Research progress on the use of polyphenols in the treatment of Crohn's disease. Front Pharmacol 2025, 16, 1704901. [Google Scholar] [CrossRef]
- Wen, X; Tang, S; Wan, F; Zhong, R; Chen, L; Zhang, H. The PI3K/Akt-Nrf2 Signaling Pathway and Mitophagy Synergistically Mediate Hydroxytyrosol to Alleviate Intestinal Oxidative Damage. Int J Biol Sci 2024, 20, 4258–76. [Google Scholar] [CrossRef]
- Lv, W; Jin, W; Lin, J; Wang, Z; Ma, Y; Zhang, W; et al. Forsythia suspensa polyphenols regulate macrophage M1 polarization to alleviate intestinal inflammation in mice. Phytomedicine 2024, 125, 155336. [Google Scholar] [CrossRef] [PubMed]
- Zhong, K; Yao, Y; Shi, B; Li, S; Zhao, Y; Wang, X; et al. EGCG alleviates colitis in inflammatory bowel disease via gut microbiota-derived LCA and inhibition of NLRP3 inflammasome. Int Immunopharmacol 2026, 168, 115962. [Google Scholar] [CrossRef] [PubMed]
- Otkur, W; Liu, X; Chen, H; Li, S; Ling, T; Lin, H; et al. GPR35 antagonist CID-2745687 attenuates anchorage-independent cell growth by inhibiting YAP/TAZ activity in colorectal cancer cells. Front Pharmacol 2023, 14, 1126119. [Google Scholar] [CrossRef]
- Yang, C; Merlin, D. Unveiling Colitis: A Journey through the Dextran Sodium Sulfate-induced Model. Inflamm Bowel Dis 2024, 30, 844–53. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y; Zhu, X; Shen, J; Chen, H; Zhou, G. Mechanism of Nicotiflorin in San-Ye-Qing rhizome for anti-inflammatory effect in ulcerative colitis. Phytomedicine 2024, 129, 155564. [Google Scholar] [CrossRef]
- Li, Z; Wen, Q; Pi, J; Zhang, D; Nie, J; Wei, W; et al. An inulin-type fructan isolated from Serratula chinensis alleviated the dextran sulfate sodium-induced colitis in mice through regulation of intestinal barrier and gut microbiota. Carbohydr Polym 2023, 320, 121206. [Google Scholar] [CrossRef]
- Zhang, M; Yang, D; Yu, H; Li, Q. MicroRNA-497 inhibits inflammation in DSS-induced IBD model mice and lipopolysaccharide-induced RAW264.7 cells via Wnt/β-catenin pathway. Int Immunopharmacol 2021, 101, 108318. [Google Scholar] [CrossRef]
- Yu, Z; Ruan, G; Bai, X; Sun, Y; Yang, H; Qian, J. Growing burden of inflammatory bowel disease in China: Findings from the Global Burden of Disease Study 2021 and predictions to 2035. Chin Med J (Engl) 2024, 137, 2851–59. [Google Scholar] [CrossRef]
- Yang, X; Guo, H; Zou, M. Inflammatory bowel diseases: pathological mechanisms and therapeutic perspectives. Mol Biomed 2026, 7, 2. [Google Scholar] [CrossRef]
- Mansouri, P; Mansouri, P; Behmard, E; Najafipour, S; Kouhpayeh, A; Farjadfar, A. Novel targets for mucosal healing in inflammatory bowel disease therapy. Int Immunopharmacol 2025, 144, 113544. [Google Scholar] [CrossRef]
- Christensen, C; Knudsen, A; Arnesen, EK; Hatlebakk, JG; Sletten, IS; Fadnes, LT. Diet, Food, and Nutritional Exposures and Inflammatory Bowel Disease or Progression of Disease: an Umbrella Review. Adv Nutr 2024, 15, 100219. [Google Scholar] [CrossRef] [PubMed]
- Fraga, CG; Croft, KD; Kennedy, DO; Tomás-Barberán, FA. The effects of polyphenols and other bioactives on human health. Food Funct 2019, 10, 514–28. [Google Scholar] [CrossRef]
- Foata, F; Sprenger, N; Rochat, F; Damak, S. Activation of the G-protein coupled receptor GPR35 by human milk oligosaccharides through different pathways. Sci Rep 2020, 10, 16117. [Google Scholar] [CrossRef] [PubMed]
- Xie, B; Wang, M; Xiao, Y; Zhang, X; Liu, M; Miao, J; et al. Tryptophan metabolic gatekeeping in epithelial repair: GPR35-KLF5 circuitry decodes mucosal damage signals for repair programming. Cell Death Dis 2026, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- Kozielewicz, P; Schulte, G. NanoBRET and NanoBiT/BRET-Based Ligand Binding Assays Permit Quantitative Assessment of Small Molecule Ligand Binding to Smoothened. Methods Mol Biol 2022, 2374, 195–204. [Google Scholar] [CrossRef]
- Tekin, GG; Deveci, B; Deveci, E. Ellagic acid protected the gingival tissue via fibroblast and epidermal growth factors in rats. Acta Cir Bras 2024, 39, e391224. [Google Scholar] [CrossRef] [PubMed]
- Tian, L; Wang, Z; Chen, S; Guo, K; Hao, Y; Ma, L; et al. Ellagic Acid-Loaded sEVs Encapsulated in GelMA Hydrogel Accelerate Diabetic Wound Healing by Activating EGFR on Skin Repair Cells. Cell Prolif 2025, 58, e70064. [Google Scholar] [CrossRef]
- Talevi, A. Computer-Aided Drug Discovery and Design: Recent Advances and Future Prospects. Methods Mol Biol 2024, 2714, 1–20. [Google Scholar] [CrossRef]
- Wu, Q; Li, Y; Zhao, Y; Zhang, R; Tong, J; Ji, C; et al. Astragaloside IV Alleviates Fructose-Induced Intestinal Metabolic Senescence by Targeting Ketohexokinase Asn261/Ala226 to Preserve Intestinal Stem Cell Homeostasis. ACS Cent Sci 2025, 11, 1682–99. [Google Scholar] [CrossRef]
- Zhang, M; Cui, S; Mao, B; Zhang, Q; Zhao, J; Zhang, H; et al. Ellagic acid and intestinal microflora metabolite urolithin A: A review on its sources, metabolic distribution, health benefits, and biotransformation. Crit Rev Food Sci Nutr 2023, 63, 6900–22. [Google Scholar] [CrossRef]
- Li, X; Xu, L; Peng, X; Zhang, H; Kang, M; Jiang, Y; et al. The alleviating effect of ellagic acid on DSS-induced colitis via regulating gut microbiomes and gene expression of colonic epithelial cells. Food Funct 2023, 14, 7550–61. [Google Scholar] [CrossRef]
- Xiong, Y; Cheng, Z; Zhang, Y; Liu, T; Wan, Z; Xia, C; et al. Ellagic acid alleviates DSS-induced ulcerative colitis by inhibiting ROS/NLRP3 pathway activation and modulating gut microbiota in mice. Eur J Nutr 2025, 64, 64. [Google Scholar] [CrossRef]
- Ye, X; Chen, T; Chen, L; Wu, D; Du, Y; Hu, J. Hydrogel-Shielded Ellagic Acid Nanoparticles Prolong Colonic Retention and Mitigate DSS-Induced Colitis via Reactive Oxygen Species Scavenging. Foods 2025, 14. [Google Scholar] [CrossRef]
- Jin, H; Che, S; Wu, K; Wu, M. Ellagic acid prevents gut damage via ameliorating microbe-associated intestinal lymphocyte imbalance. Food Funct 2022, 13, 9822–31. [Google Scholar] [CrossRef]
- Kim, DH; Kim, JS; Kwon, JH; Kwun, IS; Baek, MC; Kwon, GS; et al. Ellagic Acid Prevented Dextran-Sodium-Sulfate-Induced Colitis, Liver, and Brain Injury through Gut Microbiome Changes. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Tsukahara, T; Hamouda, N; Utsumi, D; Matsumoto, K; Amagase, K; Kato, S. G protein-coupled receptor 35 contributes to mucosal repair in mice via migration of colonic epithelial cells. Pharmacol Res 2017, 123, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Lan, A; Blachier, F; Benamouzig, R; Beaumont, M; Barrat, C; Coelho, D; et al. Mucosal healing in inflammatory bowel diseases: is there a place for nutritional supplementation? Inflamm Bowel Dis 2015, 21, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Kong, C; Yang, M; Yue, N; Zhang, Y; Tian, C; Wei, D; et al. Restore Intestinal Barrier Integrity: An Approach for Inflammatory Bowel Disease Therapy. J Inflamm Res 2024, 17, 5389–413. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



