Submitted:
29 January 2026
Posted:
30 January 2026
You are already at the latest version
Abstract
Heart failure with preserved ejection fraction (HFpEF) represents the predominant form of heart failure, affecting over 50% of all heart failure patients with increasing prevalence in aging populations. Despite significant advances in cardiovascular medicine, HFpEF remains a complex clinical syndrome with poorly understood pathophysiology and limited treatment options. While most studies traditionally focused on the renin-angiotensin-aldosterone system (RAAS) and other related mechanisms, emerging evidence has unveiled a critical bidirectional relationship between dysregulation of gut microbiota and HFpEF development. This phenomenon, mediated through microbiome-driven inflammation and endothelial dysfunction, introduces a novel concept and potential paradigm shift in understanding HFpEF. This comprehensive review explores this novel gut-heart axis by synthesizing the latest evidence from original studies and clinical trials. We discuss novel mechanisms involving bacterial metabolites, including short-chain fatty acids (SCFAs), trimethylamine N-oxide (TMAO), bile acids, and amino acid derivatives. We also examine how gut dysbiosis contributes to systemic inflammation through lipopolysaccharide translocation, NLRP3 inflammasome activation, and endothelial dysfunction. Furthermore, clinical trials investigating microbiome-targeted interventions, including probiotics, fecal microbiota transplantation, metabolite supplementation, and precision medicine approaches, are critically evaluated for their therapeutic potential. This review provides a roadmap for future research and therapeutic strategies targeting the gut-heart axis in HFpEF management.
Keywords:
heart failure with preserved ejection fraction
; gut microbiota
; microbiome-driven inflammation
; endothelial dysfunction
; TMAO
; short-chain fatty acids
; precision medicine
1. Introduction
Heart failure with preserved ejection fraction has emerged as the dominant form of heart failure in the 21st century, accounting for over 50% of all heart failure cases with a rising prevalence that parallels global demographic shifts toward aging populations and increasing prevalence of metabolic disorders [1,2]. This complex syndrome is characterized by preserved systolic function (ejection fraction >50%) but impaired diastolic performance, often accompanied by multiple comorbidities, including obesity, diabetes mellitus, hypertension, and metabolic syndrome. Unlike heart failure with reduced ejection fraction, which has established therapeutic frameworks, HFpEF presents unique pathophysiological challenges that have confounded traditional treatment approaches [3,4]. The syndrome’s heterogeneous nature, coupled with complex interactions between cardiac dysfunction and systemic comorbidities, has led to disappointing results in clinical trials using conventional heart failure therapeutics [5]. This therapeutic gap has intensified the search for novel mechanistic pathways and treatment targets.
This inflammatory burden, often exacerbated by prevalent comorbidities such as obesity, diabetes, hypertension, and chronic kidney disease, initiates a cascade of events leading to endothelial dysfunction, myocardial stiffening, and impaired diastolic function [6]. Given the multisystemic nature of HFpEF, where conditions like diabetes and metabolic syndrome are increasingly recognized as primary drivers, understanding the underlying inflammatory-metabolic phenotype is crucial for developing effective therapeutic strategies [7,8]. Indeed, despite its rising prevalence, HFpEF lacks evidence-based therapies, highlighting a significant unmet need for novel approaches to improve patient outcomes [5,9].
The concept of the gut-heart axis has emerged as a paradigm-shifting framework for understanding HFpEF pathogenesis. Recent investigations have revealed significant alterations in gut microbiome composition in HFpEF patients, characterized by reduced microbial diversity, depletion of beneficial bacteria, and enrichment of pro-inflammatory species [10,11]. These changes are associated with altered production of bacterial metabolites, creating a systemic inflammatory milieu that promotes endothelial dysfunction, cardiac remodeling, and diastolic dysfunction [12,13]. The gut microbiota-inflammation-HFpEF axis represents a bidirectional relationship where gut dysbiosis contributes to cardiovascular dysfunction while cardiac dysfunction further exacerbates intestinal barrier impairment and microbial translocation [14]. This vicious cycle offers novel therapeutic opportunities through microbiome modulation, metabolite replacement therapy, and precision medicine approaches based on individual microbial signatures.
This comprehensive review synthesizes the most recent evidence on the gut-heart axis in HFpEF, with particular emphasis on microbiome-driven inflammation and endothelial dysfunction. We critically evaluate emerging therapeutic strategies and provide a framework for future research directions that could transform HFpEF management through precision microbiome medicine. By bridging experimental and clinical evidence, this review proposes gut–heart axis modulation as a promising frontier for innovative HFpEF therapies.
1.1. Gut Microbiome Alterations in HFpEF:
Recent investigations into the gut microbiome of patients with heart failure with preserved ejection fraction (HFpEF) have unveiled significant alterations that differentiate these individuals from healthy controls. Notably, a consistent pattern of microbial dysbiosis has been observed, characterized by a decline in beneficial, anti-inflammatory bacterial populations and an increase in potentially pathogenic species. Studies indicate that HFpEF patients display a marked decrease in α-diversity, particularly in species richness. Metrics such as the Chao index show significant reductions in HFpEF patients compared to those without the condition [11,15]. This reduction in microbial diversity has been shown to correlate with disease severity and functional capacity among HFpEF patients [15,16].
One of the most critical findings regarding HFpEF-related dysbiosis is the depletion of specific beneficial genera that play vital roles in gut health and inflammatory modulation. Genera such as Butyricicoccus, Lachnospira, and Ruminiclostridium, recognized for their production of short-chain fatty acids (SCFAs) and anti-inflammatory properties, are often reduced in HFpEF patients [11,15]. This loss is thought to contribute to systemic inflammation and cardiac dysfunction, highlighting the pathogenic potential of altered gut microbiota. Concurrently, there is an observed increase in pathogenic microorganisms such as Enterococcus, which can be linked to inflammatory responses [10,15].
Importantly, emerging evidence suggests that these microbial shifts in HFpEF patients occur independently of traditional cardiovascular risk factors, such as age, body mass index (BMI), and hypertension, implying that HFpEF itself may uniquely drive specific patterns of gut dysbiosis [10,16]. The work by Zhou et al. indicates that these gut microbiome alterations may warrant consideration in the management and treatment of HFpEF, potentially opening pathways for therapeutic interventions aimed at restoring gut health to improve heart function [10].
1.2. Metabolomic Consequences:
The observed compositional changes within the gut microbiome profoundly alter the gut metabolome, with significant implications for systemic inflammation and cardiovascular function.
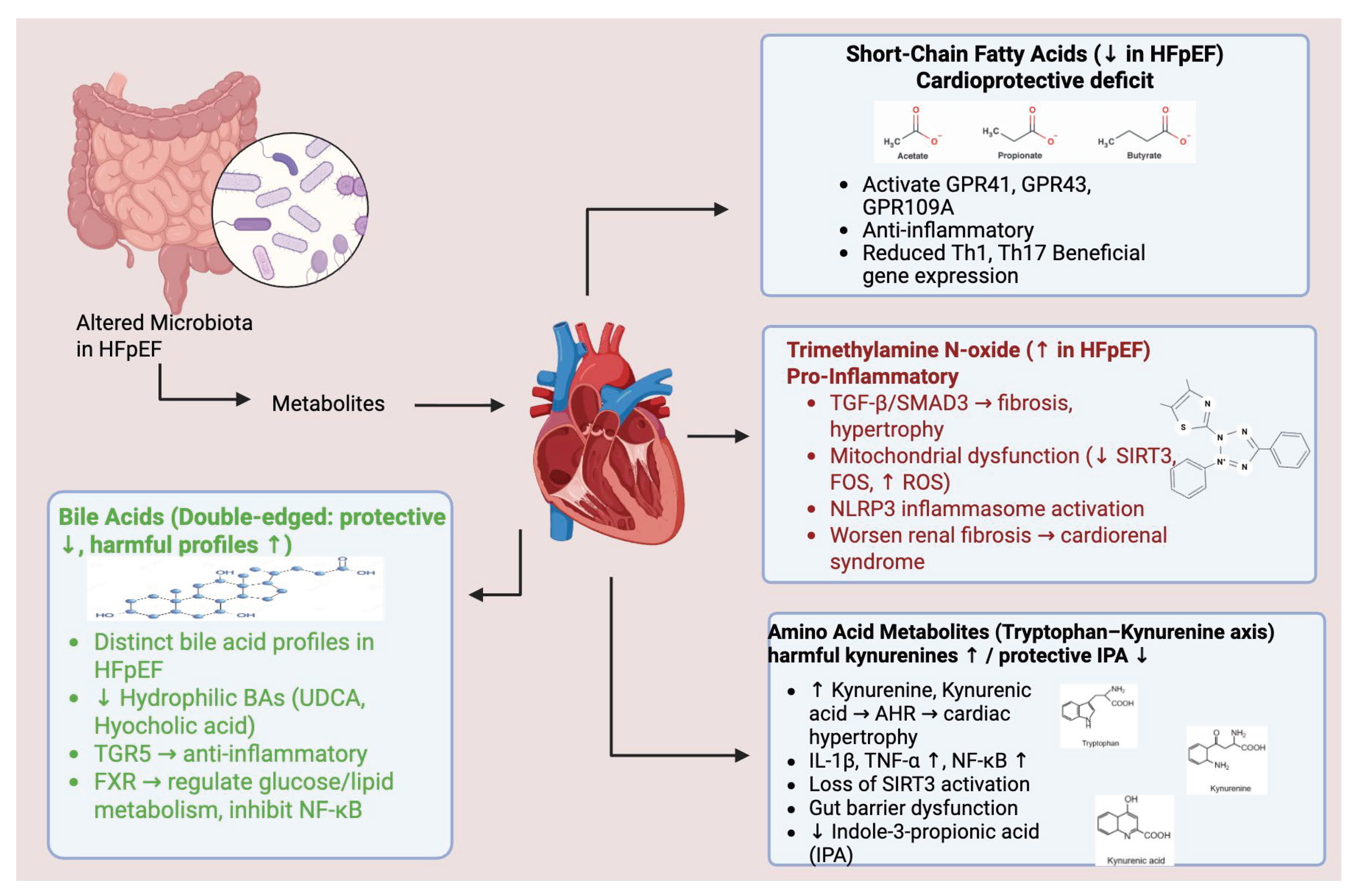
SCFA Depletion: A direct consequence of the reduction in short-chain fatty acid-producing bacteria is the diminished circulating levels of key SCFAs such as acetate, propionate, and butyrate. This depletion is particularly significant given the established cardioprotective properties of SCFAs and their crucial role in maintaining intestinal barrier integrity. [17]
Harmful Metabolite Accumulation: Patients with HFpEF often exhibit elevated levels of potentially toxic bacterial metabolites. This includes an increase in trimethylamine N-oxide production and altered amino acid metabolism products. These metabolic shifts collectively contribute to a systemic environment characterized by heightened inflammation and pro-fibrotic processes. [18]
Bile Acid Dysregulation: Alterations in gut microbiome composition also lead to a disruption in bile acid metabolism. Such dysregulation has broad implications for both cardiac function and overall metabolic homeostasis. Specifically, certain bile acid profiles, including reduced levels of ursodeoxycholic acid and hyocholic acid, have been associated with an increased risk of HFpEF. [19].
2. Microbiome-Derived Metabolites: Key Players in HFpEF Pathogenesis
The observed alterations in gut microbiota composition lead to profound changes in the gut metabolome, generating various metabolites that significantly impact systemic inflammation and cardiovascular function, thereby playing a key role in HFpEF pathogenesis. [20]
2.1. Short-Chain Fatty Acids: The Cardioprotective Deficit
Short-chain fatty acids, particularly acetate, propionate, and butyrate, are crucial beneficial metabolites produced by gut microbiota, serving as vital mediators of gut-heart communication. A profound depletion of SCFA-producing bacteria in HFpEF patients directly contributes to disease pathogenesis. SCFAs exert multiple cardioprotective effects through diverse molecular pathways [20]. They activate G-protein coupled receptors (GPR41, GPR43, GPR109A), triggering anti-inflammatory signaling cascades and metabolic regulation [17]. Furthermore, SCFAs act as endogenous histone deacetylase inhibitors, promoting beneficial gene expression patterns and reducing inflammatory transcription. Specifically, propionate diminishes NF-κB, IL-6, STAT1, and STAT3 activity, thereby reducing Th1 and Th17 immune responses, while butyrate significantly inhibits NF-κB activity via PPAR-γ stimulation [17,20]. Beyond their anti-inflammatory roles, SCFAs serve as direct cardiac energy substrates and promote beneficial metabolic adaptations, including enhanced branched-chain amino acid catabolism. Experimental studies underscore their importance, demonstrating that SCFA supplementation, especially butyrate, can reverse cardiac dysfunction in HFpEF models. For instance, tributyrin treatment in obese pre-HFpEF mice improved early cardiac dysfunction by upregulating protein phosphatase 2cm and enhancing branched chain amino acid metabolism [21]. This evidence highlights the SCFA deficit as a clear therapeutic target, with interventions encompassing direct supplementation, prebiotic therapy to promote endogenous production, and dietary modifications to increase fiber intake [15].
2.2. Trimethylamine N-Oxide: The Inflammatory Culprit
Trimethylamine N-oxide (TMAO) has emerged as a significant pathogenic metabolite in the context of the gut-heart axis, particularly relating to heart failure with preserved ejection fraction (HFpEF). Elevated levels of TMAO have been consistently linked to adverse clinical outcomes in HFpEF patients [22,23,24]. This metabolite is produced by the metabolic activity of gut bacteria, which convert dietary choline and carnitine into trimethylamine (TMA), subsequently oxidized to TMAO in the liver. This illustrates a direct mechanistic relationship between dietary intake, gut microbiome dynamics, and cardiovascular dysfunction [25,26].
TMAO contributes to the pathophysiology of HFpEF through various interconnected pathways. One critical mechanism involves the activation of the TGF-β/SMAD3 signaling pathway, which is implicated in promoting cardiac fibrosis and hypertrophy [27]. Furthermore, TMAO has been shown to induce mitochondrial dysfunction by inhibiting the expression of sirtuin 3 and suppressing the activities of superoxide dismutase 2 and mitochondrial aldehyde dehydrogenase 2, resulting in the accumulation of reactive oxygen species and consequent cellular damage [28]. Additionally, TMAO triggers NLRP3 inflammasome activation, promoting cardiovascular inflammation and endothelial dysfunction, which are pivotal processes in the progression of heart failure [29].
The implications of elevated TMAO extend beyond direct cardiac effects; it has also been observed to exacerbate renal interstitial fibrosis, thereby accelerating the progression of HFpEF through mechanisms associated with cardiorenal syndrome [23]. Clinically, cross-sectional studies have identified significantly elevated TMAO levels in HFpEF patients compared to healthy controls, underscoring its role as an independent risk factor for negative cardiovascular outcomes. Elevated TMAO correlates with increased mortality and major adverse cardiovascular events and provides valuable stratification in risk assessments, particularly in scenarios where natriuretic peptides do not reflect disease severity accurately [30,31].
Ongoing research is focused on exploring multiple strategies to reduce TMAO levels, including dietary modifications, microbiome interventions, and direct inhibition of TMA-producing microbial pathways or the flavin monooxygenase 3 enzyme involved in TMAO synthesis [26]. This multifaceted approach holds promise for mitigating the pro-inflammatory and harm-promoting effects of TMAO, potentially improving the clinical management of HFpEF patients.
2.3. Amino Acid Metabolites: The Tryptophan-Kynurenine Axis
Dysregulated amino acid metabolism, particularly the tryptophan-kynurenine pathway, is a significant aspect of the gut-heart axis in heart failure with preserved ejection fraction (HFpEF), representing a potential therapeutic target. Patients with HFpEF have been reported to exhibit increased levels of tryptophan catabolism products such as kynurenine and kynurenic acid, which could be associated with mechanisms like left ventricular remodeling and increased pro-inflammatory cytokines [32]. These metabolites may contribute to HFpEF pathogenesis through several mechanisms, including the activation of the aryl hydrocarbon receptor (AHR), which has been implicated in cardiac hypertrophy [33]. Specifically, kynurenic acid is understood to induce the expression of pro-inflammatory cytokines like IL-1β and TNF-α via AHR signaling [34].
Conversely, indole-3-propionic acid (IPA), a beneficial metabolite derived from tryptophan, is observed to be significantly reduced in HFpEF patients [33]. IPA has illustrated therapeutic potential by activating SIRT3, thus enhancing the NAD+ pathway and improving gut barrier integrity [35]. Moreover, kynurenine pathway metabolites are of growing interest as potential biomarkers for the diagnosis and prognosis of HFpEF. Therapeutic approaches aimed at modulating tryptophan metabolism, which include IPA supplementation and inhibition of the kynurenine pathway, are currently under investigation to explore their potential utility in HFpEF management [36].
2.4. Bile Acids: The Double-Edged Metabolites
Bile acids are complex signaling molecules that exert both beneficial and detrimental effects on cardiovascular health. Recent research has identified specific bile acid profiles associated with HFpEF risk and therapeutic potential. HFpEF patients, particularly those with metabolic dysfunction-associated fatty liver disease, exhibit distinct bile acid signatures, including significantly lower levels of ursodeoxycholic acid and hyocholic acid species [37,38].
Hydrophilic bile acids, including ursodeoxycholic acid, may confer protective effects on cardiac tissues through mechanisms such as TGR5 receptor activation. Activation of TGR5 has been linked to anti-inflammatory signaling pathways that support cardiac function and mitigate inflammation [39]. Additionally, bile acids also engage the FXR pathway, playing a significant role in regulating glucose and lipid metabolism while inhibiting NF-κB signaling, which reduces inflammatory responses [40]. The therapeutic potential of bile acids is under investigation, with strategies focusing on bile acid receptor agonists and supplementation with cardioprotective bile acids, such as taurine-conjugated bile acids, showing promise in experimental models of HFpEF [41]. Emerging evidence also indicates that bile acid signaling may be leveraged to improve metabolic outcomes in HFpEF patients, suggesting that interventions designed to modulate bile acid metabolism could represent a novel therapeutic approach in the management of HFpEF [42].
3. Endothelial Dysfunction in the Gut-Heart Axis
Endothelial dysfunction represents a critical pathophysiological component of HFpEF, with gut microbiome alterations serving as a major driver of this impairment. The gut-heart axis contributes to endothelial dysfunction through multiple interconnected pathways that ultimately compromise coronary microvascular function and promote diastolic dysfunction.
Mechanisms of Microbiome-Mediated Endothelial Impairment.
Microbiome-derived factors contribute significantly to endothelial damage:
- TMAO-Induced Endothelial Damage: Trimethylamine N-oxide directly impairs endothelial function through several mechanisms. It enhances endothelial cell apoptosis and inflammatory activation, promotes oxidative stress, and reduces nitric oxide bioavailability. Furthermore, TMAO facilitates foam cell formation, accelerates atherosclerosis, and impairs endothelium-dependent vasodilation, all contributing to endothelial dysfunction. [43,44].
- LPS-Mediated Endothelial Activation: Bacterial lipopolysaccharides, crossing the compromised gut barrier, activate endothelial cells via Toll-like receptor 4 signaling. This activation leads to the upregulation of adhesion molecules (ICAM-1, VCAM-1) and chemokines, increasing vascular permeability and inflammatory cell recruitment. This process ultimately contributes to a pro-thrombotic endothelial phenotype and activates complement and coagulation cascades. [45,46,47].
- SCFA Deficiency Effects: A reduction in beneficial short-chain fatty acid levels also contributes to endothelial dysfunction. This deficiency results in decreased G-protein coupled receptor-mediated protective signaling in endothelial cells, impairs endothelial barrier function leading to increased permeability, and reduces the production of anti-inflammatory mediators. Consequently, it compromises endothelial repair and regeneration mechanisms, exacerbating dysfunction. [48,49].
4. Clinical Implications of Endothelial Dysfunction and Therapeutic Targets
The clinical implications of endothelial dysfunction, particularly in the context of heart failure with preserved ejection fraction (HFpEF), are significant, impacting patient management and therapeutic strategies. Endothelial dysfunction has been associated with coronary microvascular dysfunction, exercise intolerance, and the overall progression of heart failure pathology. Understanding these implications reinforces the need for targeted therapeutic interventions aimed at these mechanisms.
Endothelial dysfunction severely compromises coronary microvascular function, leading to impaired coronary flow reserve and increased microvascular resistance, which restricts oxygen delivery to cardiac tissues [50]. These factors are central to the pathophysiology of HFpEF and contribute to exercise intolerance in affected patients. The inadequate ability of both peripheral and coronary vasculature to dilate during physical activity exacerbates hemodynamic instability and diminishes exercise capacity, ultimately impairing quality of life [50]. Furthermore, chronic endothelial dysfunction can accelerate the progression of HFpEF, resulting in increased arterial stiffness, cardiac remodeling, and potential onset of cardiorenal syndrome, thereby worsening patient outcomes [48,50].
Given these challenges, several therapeutic strategies targeting endothelial dysfunction present opportunities for improving patient outcomes. For instance, restoring gut microbiome balance through approaches such as fecal microbiota transplantation (FMT) or the administration of probiotics can enhance endothelial function by decreasing harmful metabolite production and promoting beneficial microbial profiles [51]. Additionally, direct interventions focusing on short-chain fatty acids (SCFAs) supplementation and strategies aimed at reducing trimethylamine N-oxide (TMAO) levels may positively influence endothelial integrity and cardiovascular health [50]. Anti-inflammatory strategies, including modulation of the NLRP3 inflammasome and NF-κB pathways, may offer additional avenues to counteract the pro-inflammatory state associated with gut-derived endotoxemia and endothelial dysfunction [50,52].
The complex nature of the gut-heart axis emphasizes the urgent need for comprehensive strategies to address endothelial dysfunction in HFpEF patients. By implementing microbiome-focused interventions, healthcare professionals may improve patient outcomes and better manage this intricate condition.
5. Therapeutic Interventions Targeting the Gut-Heart Axis in HFpEF
Fecal Microbiota Transplantation: Comprehensive Microbiome Restoration:
FMT represents a comprehensive approach to gut microbiome restoration, offering significant potential for fundamentally correcting the dysbiosis underlying HFpEF. Recent experimental evidence strongly supports its therapeutic promise. Preclinical studies, such as those conducted in obesity-associated pre-HFpEF mouse models, have demonstrated remarkable therapeutic effects of FMT from lean donors. These effects include improvement in systolic and diastolic function, along with reductions in cardiac hypertrophy and pathological remodeling [53]. These outcomes are thought to be linked to an increase in butyrate-producing bacteria, highlighting the microbiome’s role in cardiovascular health [54]. Moreover, investigations into diet-induced dysbiosis reveal that FMT can improve metabolic health, suggesting that gastrointestinal health restoration may yield significant cardiovascular benefits [55].
The transition from preclinical success to clinical application for FMT in HFpEF faces several challenges. These include the need for standardization of donor selection and microbiome characterization, optimization of delivery methods and dosing protocols, and rigorous safety evaluation in cardiovascular disease populations. Furthermore, the development of synthetic microbial consortiums is being explored as potential alternatives to whole microbiome transfer [56,57,58,59].
The future of FMT in treating cardiovascular conditions appears promising, particularly as personalized medicine approaches develop. By tailoring donor selection to match recipient microbiome profiles and creating oral formulations for easier patient acceptance, FMT could potentially transform treatment strategies for HFpEF and related cardiometabolic disorders. As research advances, rigorous clinical trials will be essential to assess the long-term safety and efficacy of FMT interventions in diverse cardiovascular patient populations [60,61].
5.1. Probiotic Interventions: Targeted Microbial Supplementation
Probiotic interventions present a targeted strategy for microbiome modulation, particularly with promising implications for heart failure with preserved ejection fraction (HFpEF) populations. Ongoing clinical studies are exploring the efficacy of specific probiotic strains on metabolic improvements and heart failure outcomes. Notably, the trial titled "Influence of Probiotics on Metabolome and Heart Failure" is investigating the effects of Lactobacillus rhamnosus supplementation, which has been associated with changes in metabolic profiles and beneficial cardiac function biomarkers [62]. This emphasis on specific strains highlights the need for tailored probiotic applications to enhance therapeutic outcomes in HFpEF.
Advanced formulations combining multiple probiotic strains are also being researched for their potential synergistic effects on inflammation and cardiovascular function [63,64]. Evidence suggests that probiotics may address HFpEF outcomes through a multitude of mechanisms, including restoring microbial diversity, enhancing the production of beneficial metabolites, and competitively excluding pathogenic bacteria—all of which can positively influence gut health [65,66]. Probiotics are especially promising in reducing trimethylamine N-oxide (TMAO) synthesis, which is linked to cardiovascular risk, as well as enhancing gut barrier function to mitigate bacterial translocation into circulation [66,67].
The direct anti-inflammatory effects of probiotics, facilitated by immune modulation, further underline their therapeutic relevance in managing HFpEF-related pathophysiology [68,69]. As ongoing research continues to unravel the complexities of probiotics in cardiovascular health, their application in clinical settings appears increasingly viable. The findings from these investigations may pave the way for refined probiotic formulations that can be strategically integrated into treatment regimens for patients managing HFpEF.
5.2. Metabolite-Based Therapies and Precision Medicine Approach
Direct supplementation of beneficial metabolites or inhibition of harmful metabolite production represents a precision medicine approach to targeting the gut-heart axis, particularly in heart failure with preserved ejection fraction (HFpEF). Emerging studies have highlighted the potential of short-chain fatty acid (SCFA) supplementation in HFpEF models. Specifically, tributyrin, a butyrate prodrug, has been found to improve cardiac function and reduce fibrosis [70]. Additionally, sodium butyrate supplementation has enhanced cardiac energy metabolism, and propionate supplementation has been shown to reduce systemic inflammation and myocardial fibrosis [71].
Indole-3-propionic acid (IPA) is gaining attention as a therapeutic target with potential benefits for HFpEF. Its mechanisms include activation of SIRT3, enhancement of NAD+ pathways, reduction of oxidative stress and inflammation, and improvement of gut barrier function and microbiome composition [72]. These processes collectively support its cardioprotective effects through metabolic modulation, positioning IPA as a beneficial agent in managing HFpEF.
In parallel, strategies aimed at reducing harmful trimethylamine N-oxide (TMAO) levels are being investigated. Approaches include dietary modifications that lower choline and carnitine intake, microbiome modulation to decrease TMA-producing bacteria, and inhibition of the flavin monooxygenase 3 (FMO3) enzyme, crucial in TMAO synthesis [73,74]. The development of TMAO-binding compounds for enhanced elimination is also in progress, indicating a multifaceted strategy to mitigate cardiovascular risks associated with TMAO [75].
Looking ahead, deploying precision medicine approaches tailored to individual microbiome responses and metabolic profiles is crucial. Ongoing efforts to create biomarker panels incorporating gut microbiome-derived metabolites aim to facilitate early detection of HFpEF, risk stratification, and personalized intervention strategies [76]. The integration of multi-omics data—comprising genomics, metabolomics, and microbiomics promises to identify novel therapeutic targets and optimize combination therapies for individual patient needs [77]. Furthermore, advancements in digital health technologies, such as wearable devices integrated with microbiome analysis, will enable real-time monitoring of therapeutic responses, enhancing personalized dietary recommendations and optimizing intervention timing [78]. Therefore, precision medicine approaches focusing on metabolic and microbiome modulation concerning the gut-heart axis hold promise for improving outcomes in patients with HFpEF.
Figure 1.
Gut–heart axis mechanisms in HFpEF.
6. Future Directions and Research Opportunities:
The gut-heart axis in HFpEF presents numerous unexplored avenues for research, offering transformative opportunities that could revolutionize treatment approaches and patient outcomes.
6.1. Novel Therapeutic Targets
Beyond conventional approaches, several novel therapeutic targets are emerging from research into the gut-heart axis. Recent evidence suggests that Paneth cell dysfunction contributes significantly to gut-heart axis disruption. Paneth cells, specialized intestinal epithelial cells that produce antimicrobial peptides, may represent a novel therapeutic target for maintaining gut barrier function and preventing bacterial translocation [79,80,81]. Recent studies have elucidated the impact of Paneth cell dysfunction in conditions like obesity and inflammatory bowel disease (IBD), highlighting their role in regulating gut microbiota and preventing dysbiosis [82,83,84]. For instance, a report noted that bile acid toxicity within Paneth cells can exacerbate dysbiosis, suggesting a potential target for therapeutic intervention [85]. Additionally, research indicates that supplementation of products derived from Paneth cells, such as defensins, can restore gut microbiota balance after dysregulation [80].
Understanding microbiome-drug interactions represents another crucial aspect of optimizing therapies for heart failure with preserved ejection fraction (HFpEF). Current medications, particularly SGLT2 inhibitors, show promise not only for their cardioprotective effects but also for their ability to modulate gut microbiome composition, which may contribute to their therapeutic efficacy [86,87]. The complex relationship between medications and the gut microbiome necessitates further exploration to optimize treatment strategies effectively. Additionally, integrating circadian rhythm research offers a unique perspective on gut-heart signaling, given that the gut microbiome exhibits circadian rhythms that can profoundly influence cardiovascular function. Chronotherapy approaches that consider the timing of interventions may enhance therapeutic outcomes for HFpEF patients [88].
Moreover, phage therapy is emerging as a precision medicine strategy to modulate the gut microbiome selectively. Phage therapy’s ability to target specific pathogenic bacteria while preserving beneficial microbial populations could lead to personalized treatments that mitigate dysbiosis-related issues in HFpEF patients [81,89].
6.2. Advanced Diagnostic Approaches:
The development of advanced diagnostic tools is essential for harnessing the full potential of gut-heart axis research in heart failure with preserved ejection fraction (HFpEF). Metabolomic profiling, combined with comprehensive clinical assessments, can facilitate the early identification of HFpEF risk in asymptomatic individuals and monitor treatment efficacy through metabolite tracking [90,91]. This advancement may foster the creation of point-of-care diagnostic tools that enhance patient management in clinical settings.
Artificial intelligence applications to multi-omics datasets integrating genomics, metabolomics, and microbiomics hold promise for identifying novel biomarker combinations that could improve diagnostic accuracy. These approaches may aid in predicting optimal therapeutic interventions for individual patients, discovering previously unrecognized pathogenic pathways, and enabling real-time treatment optimization based on continuous monitoring [92,93,94]. For instance, systems utilizing AI to analyze extensive omics data can uncover complex interactions that might otherwise remain hidden, thus refining treatment strategies and improving outcomes for HFpEF patients [95].
Moreover, the integration of advanced cardiac imaging techniques with microbiome analysis could yield valuable insights into correlations between microbiome signatures and specific cardiac phenotypes. This dual approach may facilitate the identification of subclinical disease progression markers and guide imaging-informed microbiome interventions [91].
6.3. Therapeutic Innovation:
Innovative therapeutic strategies are being explored to directly target the gut-heart axis in HFpEF. Advances in gene therapy, such as trials involving SERCA2a gene therapy for HFpEF, could be further enhanced through combinations with microbiome interventions focused on inflammation and metabolism [96,97]. Regenerative medicine approaches, including stem cell therapies and tissue engineering, could significantly benefit from concomitant microbiome optimization, creating favorable inflammatory environments conducive to cardiac repair and regeneration.
Rational design of combination therapies is also crucial, which includes integrating traditional HFpEF medications with microbiome modulators, combining multiple microbiome-targeting approaches for synergistic effects, and incorporating dietary interventions alongside pharmacological microbiome modulation [98,99]. Furthermore, the exploration of the integration of device-based therapies with microbiome optimization represents an exciting frontier in therapeutic innovation [100]. Such multi-faceted strategies are likely to enhance the efficacy of existing treatments while paving the way for novel interventions tailored to the unique needs of HFpEF patients.

Table 1.
Hypothetical Research Concepts for Future Investigation into the Gut-Heart Axis in HFpEF. The following research concepts are proposed to address major gaps identified in this review. These ideas are hypothetical and intended to stimulate future investigation into the gut–heart axis in HFpEF, rather than describe ongoing or completed clinical trials.
Table 1.
Hypothetical Research Concepts for Future Investigation into the Gut-Heart Axis in HFpEF. The following research concepts are proposed to address major gaps identified in this review. These ideas are hypothetical and intended to stimulate future investigation into the gut–heart axis in HFpEF, rather than describe ongoing or completed clinical trials.
| Proposed Study (Acronym) | Objective | Proposed Design | Primary Endpoints | Secondary Endpoints | Key Innovation |
|---|---|---|---|---|---|
| Precision Microbiome Medicine for HFpEF | Develop and validate a precision medicine platform for personalized microbiome interventions in HFpEF patients | Multi-center RCT; personalized interventions vs. standard care; n=1,000 | Composite CV outcomes, QoL, exercise capacity | Microbiome restoration, metabolite profiles, inflammatory biomarkers | ML-guided real-time microbiome monitoring for treatment optimization |
| Gut–Heart Metabolite Intervention Trial | Assess efficacy/safety of combined SCFA + IPA replacement vs. placebo in metabolite-deficient HFpEF | Phase II/III double-blind RCT; n=500 | Peak VO2, NT-proBNP, diastolic function | Endothelial function, inflammation, gut barrier integrity | First precision metabolite replacement therapy trial in CVD |
| Early HFpEF Detection Through Microbiome Analysis | Develop microbiome biomarkers for early HFpEF detection in at-risk individuals | Prospective cohort; 5-year follow-up; n=2,000 | Time to HFpEF diagnosis, predictive accuracy | Intermediate phenotypes, cost-effectiveness | First large-scale microbiome-based CV prediction study |
| Microbiome-Targeted Combination Therapy | Evaluate synergy of FMT, probiotics, and metabolite supplementation in refractory HFpEF | Adaptive randomized trial; n=300 | Clinical improvement, HF hospitalization rates, QoL | Microbiome restoration, safety, biomarkers | First systematic combination microbiome therapy trial in HFpEF |
| Pediatric-to-Adult Microbiome Trajectory Study | Determine early-life microbiome influence on adult HFpEF risk | 20-year longitudinal cohort; n=1,000 | Adult CV risk profile, HFpEF incidence | Metabolic health, inflammatory profiles, intervention windows | First lifespan microbiome–cardiovascular link study |
| Environmental Modulation of Gut–Heart Axis | Investigate environmental factors affecting gut-heart axis and HFpEF risk | Multi-cohort; varied exposures; n=5,000 | Environmental associations with microbiome disruption and HFpEF | Mechanistic pathways, reversibility | First comprehensive study of environmental determinants of CV-relevant microbiome changes |
| Artificial Intelligence–Guided Microbiome Therapy | Develop AI algorithms for real-time microbiome therapy optimization | Single-arm pilot; wearable and microbiome monitoring; n=100 | Feasibility, clinical improvement | Algorithm performance, patient acceptability, cost-effectiveness | First AI-driven real-time microbiome therapy approach in CVD |
| Microbiome–Heart Device Integration Study | Test whether microbiome optimization enhances device-based HFpEF therapy efficacy | RCT: device therapy vs. device + microbiome optimization; n=200 | Device efficacy enhancement, clinical outcomes | Inflammatory profiles, device complications, mechanisms | First integration of microbiome therapy with device-based HFpEF treatment |
6.4. Population Health Implications:
Understanding the gut-heart axis carries significant implications for population health, enabling novel prevention strategies through early interventions tailored to high-risk individuals based on their microbiome signatures. This knowledge fosters public health initiatives promoting gut microbiome health and the development of dietary guidelines optimized for cardiovascular benefits. Research indicates that diets high in fiber, which are essential for gut health, can lower the risk of cardiovascular diseases. For example, studies have shown that dietary patterns such as the Dietary Approaches to Stop Hypertension (DASH) and Mediterranean diets significantly improve gut health and subsequently cardiovascular outcomes [75].
Furthermore, public health efforts must consider environmental interventions aimed at reducing exposures that disrupt the microbiome. For instance, the Dutch Microbiome Project has highlighted various factors that shape a healthy gut microbiome, emphasizing the importance of maintaining these conditions to support public health [101]. Additionally, understanding how social determinants of health influence the gut-heart axis is essential for addressing health disparities. Variations in microbiome patterns have been observed across different populations, influenced by access to microbiome-supporting foods and interventions [102]. Cultural factors and economic barriers also play significant roles in dietary patterns and gut health, underscoring the need to consider these factors in public health strategies [103].
As evidenced by recent research, there are intrinsic relationships between gut microbiota composition and chronic health conditions such as hypertension, which shows variations based on dietary and lifestyle changes across populations [104]. This reinforces the potential of leveraging microbiome insights into effective public health strategies tailored to specific demographic groups to optimize cardiovascular health on a larger scale [105].
Moreover, ongoing studies investigate the impact of maternal diet and the consequent gut microbiota composition in offspring, which may predispose them to cardiovascular diseases later in life [106]. This longitudinal perspective highlights the importance of dietary interventions that foster healthy gut microbiomes from early stages to mitigate future health risks.
7. Conclusion
The emerging understanding of the gut-heart axis marks a significant paradigm shift in HFpEF research and treatment, presenting unprecedented avenues for therapeutic intervention in a condition that has historically been challenging to manage with traditional methods. Mounting evidence increasingly points to gut microbiota dysregulation not merely as an accompanying feature of HFpEF, but as a foundational driver of its pathogenesis, primarily through microbiome-induced inflammation and endothelial dysfunction.
Recent research highlights consistent patterns of gut dysbiosis in HFpEF patients, characterized by diminished microbial diversity, a reduction in beneficial short-chain fatty acid-producing bacteria, and an increase in pro-inflammatory species. This microbial imbalance profoundly alters the gut metabolome, leading to lower levels of cardioprotective metabolites like SCFAs and indole-3-propionic acid, alongside elevated production of detrimental compounds such as trimethylamine N-oxide. Such metabolic disturbances initiate systemic inflammation, notably via NLRP3 inflammasome activation, contribute to endothelial dysfunction, and ultimately result in cardiac remodeling and impaired diastolic function.
The reciprocal relationship within the gut-heart axis introduces both complexities and opportunities. While HFpEF can worsen gut function due to reduced cardiac output and intestinal hypoperfusion, this very bidirectionality offers numerous points for therapeutic intervention. Initial results from ongoing clinical trials exploring fecal microbiota transplantation, probiotic supplementation, and metabolite replacement therapies are encouraging, though large-scale validation studies remain essential.
The future landscape of HFpEF treatment will increasingly integrate precision medicine approaches, combining gut microbiome analysis with conventional cardiovascular assessments. This will facilitate the development of personalized therapeutic strategies tailored to individual microbiome signatures, metabolic profiles, and inflammatory patterns. Advancements such as AI-guided therapy optimization, real-time monitoring technologies, and innovative biomarker panels are poised to further refine our capacity to deliver precise and effective interventions.
This review identifies several research priorities poised to reshape HFpEF care:
- (a)
- establishing precision microbiome medicine platforms;
- (b)
- validating combination microbiome therapies;
- (c)
- developing microbiome-based systems for early disease detection;
- (d)
- integrating environmental determinants into treatment planning; and
- (e)
- leveraging artificial intelligence to optimize outcomes.
The gut-heart axis paradigm also underscores the importance of adopting a holistic approach to HFpEF management, extending beyond traditional cardiovascular interventions. Future treatment strategies may encompass dietary counseling, prebiotic supplementation, targeted antibiotic therapies, and lifestyle modifications aimed at optimizing gut microbiome health, alongside standard heart failure medications.
As we move into an era of personalized cardiovascular medicine, the gut-heart axis provides a crucial framework for addressing the intricate, multi-system nature of HFpEF. By targeting the fundamental inflammatory and metabolic dysfunctions underlying this syndrome, microbiome-based interventions offer significant promise for improving outcomes for millions of patients globally afflicted by this challenging condition.
The convergence of gut microbiome science with cardiovascular medicine represents more than just a therapeutic advancement; it signifies a profound paradigm shift towards understanding human health as a deeply interconnected ecosystem. Within this system, the gut and heart communicate through complex molecular networks, opening new frontiers for research, therapeutic development, and, critically, enhanced patient outcomes in HFpEF and other cardiovascular diseases.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Acknowledgments
The authors thank Hafiz Asad Mukhtar, CA, for his assistance with proofreading the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Clark, K; Velazquez, EJ. Heart failure with preserved ejection fraction. JAMA 2020, 324(15), 1506. [Google Scholar] [CrossRef] [PubMed]
- Guo, X; Huang, C; Zhang, L; Hu, G; Du, Y; Chen, X; et al. Lymphatic endothelial branched-chain amino acid catabolic defects undermine cardiac lymphatic integrity and drive HFpEF. Circulation 2025, 151(23), 1651–1666. [Google Scholar] [CrossRef] [PubMed]
- Tah, S; Valderrama, M; Afzal, M; Iqbal, J; Farooq, A; Lak, MA; et al. Heart failure with preserved ejection fraction: an evolving understanding. Cureus 2023, 15(7), e46152. [Google Scholar] [CrossRef] [PubMed]
- Lucà, F; Oliva, F; Abrignani, MG; Fusco, SAD; Gori, M; Giubilato, S; et al. Heart failure with preserved ejection fraction: how to deal with this chameleon. J Clin Med. 2024, 13(5), 1375. [Google Scholar] [CrossRef]
- Vaz-Salvador, P; Adão, R; Vasconcelos, I; Leite-Moreira, A; Brás-Silva, C. Heart failure with preserved ejection fraction: a pharmacotherapeutic update. Cardiovasc Drugs Ther. 2022, 37(4), 815–832. [Google Scholar] [CrossRef]
- Boulet, J; Sridhar, VS; Bouabdallaoui, N; Tardif, JC; White, M. Inflammation in heart failure: pathophysiology and therapeutic strategies. Inflamm Res. 2024, 73(5), 709–723. [Google Scholar] [CrossRef]
- Clemenza, F; Citarrella, R; Patti, AM; Rizzo, M. Obesity and HFpEF. J Clin Med. 2022, 11(13), 3858. [Google Scholar] [CrossRef]
- Lin, Y; Fu, S; Yao, Y; Li, Y; Zhao, Y; Luo, L. Heart failure with preserved ejection fraction based on aging and comorbidities. J Transl Med. 2021, 19(1), 291. [Google Scholar] [CrossRef]
- Inácio, JM; Cristo, F; Pinheiro, M; Vasques-Nóvoa, F; Saraiva, F; Nunes, MM; et al. Myocardial RNA sequencing reveals new potential therapeutic targets in heart failure with preserved ejection fraction. Biomedicines 2023, 11(8), 2131. [Google Scholar] [CrossRef]
- Zhou, S; Zhou, X; Zhang, P; et al. The gut microbiota-inflammation-HFpEF axis: deciphering the role of gut microbiota dysregulation in the pathogenesis and management of HFpEF. Front Cell Infect Microbiol. 2025, 15, 1537576. [Google Scholar] [CrossRef]
- Huang, Z; Mei, X; Jiang, Y; Chen, T; Zhou, Y. Gut microbiota in heart failure patients with preserved ejection fraction (GUMPTION Study). Front Cardiovasc Med. 2022, 8, 803744. [Google Scholar] [CrossRef] [PubMed]
- Trøseid, M; Andersen, GØ; Broch, K; Hov, JR. The gut microbiome in coronary artery disease and heart failure: current knowledge and future directions. EBioMedicine 2020, 52, 102649. [Google Scholar] [CrossRef] [PubMed]
- Andreadou, I; Ghigo, A; Nikolaou, PE; et al. Immunometabolism in heart failure. Nat Rev Cardiol. Published online. 22 June 2025. [CrossRef]
- Shen, S; Tian, B; Zhang, H; Wang, YC; Li, T; Cao, Y. Heart failure and gut microbiota: what is cause and effect? Research (Wash D C) 2025, 8, 0610. [Google Scholar] [CrossRef] [PubMed]
- Beale, A; O’Donnell, J; Nakai, M; Nanayakkara, S; Vizi, D; Carter, K; et al. The gut microbiome of heart failure with preserved ejection fraction. J Am Heart Assoc. 2021, 10(13), e020654. [Google Scholar] [CrossRef]
- Perticone, M; Gigliotti, S; Shehaj, E; Maio, R; Suraci, E; Miceli, S; et al. Gut permeability and immune-mediated inflammation in heart failure. Biomedicines 2024, 12(6), 1217. [Google Scholar] [CrossRef]
- Yukino-Iwashita, M; Nagatomo, Y; Kawai, A; Taruoka, A; Yumita, Y; Kagami, K; et al. Short-chain fatty acids in gut-heart axis: their role in the pathology of heart failure. J Pers Med. 2022, 12(11), 1805. [Google Scholar] [CrossRef]
- Liu, J; Ge, P; Luo, Y; Sun, Z; Luo, X; Li, H; et al. Decoding TMAO in the gut-organ axis: from biomarkers and cell death mechanisms to therapeutic horizons. Drug Des Devel Ther. 2025, 19, 3363–3393. [Google Scholar] [CrossRef]
- Zhou, XD; Xu, CF; Chen, QF; Shapiro, MD; Lip, GYH; Chen, LL; et al. Serum bile acid profiles are associated with heart failure with preserved ejection fraction in patients with metabolic dysfunction-associated fatty liver disease: an exploratory study. Diabetes Obes Metab. 2024, 26(9), 3684–3695. [Google Scholar] [CrossRef]
- Palm, CL; Nijholt, KT; Bakker, BM; Westenbrink, BD. Short-chain fatty acids in the metabolism of heart failure: rethinking the fat stigma. Front Cardiovasc Med. 2022, 9, 915102. [Google Scholar] [CrossRef]
- Hatahet, J; Cook, TM; Bonomo, RR; Elshareif, N; Gavini, CK; White, CR; et al. Fecal microbiome transplantation and tributyrin improves early cardiac dysfunction and modifies the BCAA metabolic pathway in a diet-induced pre-HFpEF mouse model. Front Cardiovasc Med. 2023, 10, 1105581. [Google Scholar] [CrossRef]
- Dong, Z; Zheng, S; Shen, Z; Luo, Y; Hai, X. Trimethylamine N-oxide is associated with heart failure risk in patients with preserved ejection fraction. Lab Med. 2021, 52(4), 346–351. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Giménez, R; Ahmed-Khodja, W; Molina-Castro, Y; Peiró, ÓM; Bonet, G; Carrasquer, A; et al. Gut microbiota-derived metabolites and cardiovascular disease risk: a systematic review of prospective cohort studies. Nutrients 2022, 14(13), 2654. [Google Scholar] [CrossRef] [PubMed]
- Roncal, C; Martínez-Aguilar, E; Orbe, J; Ravassa, S; Fernández-Montero, A; Saenz-Pipaon, G; et al. Trimethylamine N-oxide predicts cardiovascular mortality in peripheral artery disease. Sci Rep. 2019, 9(1), 15580. [Google Scholar] [CrossRef] [PubMed]
- Almer, G; Semeraro, MD; Meinitzer, A; Enko, D; Blanco, GR; Gallè, B; et al. Impact of long-term high dietary fat intake and regular exercise on serum TMAO and microbiome composition in female rats. Nutr Healthy Aging 2023, 8(1), 157–170. [Google Scholar] [CrossRef]
- Hsu, C; Lu, P; Lo, M; Lin, I; Chang-Chien, G; Lin, S; et al. Gut microbiota-dependent trimethylamine N-oxide pathway associated with cardiovascular risk in children with early-stage chronic kidney disease. Int J Mol Sci. 2018, 19(12), 3699. [Google Scholar] [CrossRef]
- Liu, L; Lin, Y; Zhuang, J; Ren, J; Jiang, X; Chen, M; et al. Analysis of serum metabolite profiles in syphilis patients by untargeted metabolomics. J Eur Acad Dermatol Venereol. 2019, 33(7), 1378–1385. [Google Scholar] [CrossRef]
- James, K; Gertz, E; Kirschke, CP; Allayee, H; Huang, L; Kable, ME; et al. Trimethylamine N-oxide response to a mixed macronutrient tolerance test in a cohort of healthy United States adults. Int J Mol Sci. 2023, 24(3), 2074. [Google Scholar] [CrossRef]
- Wang, Q; Sun, Y; Zhou, T; Jiang, C; A, L; Xu, W. Gut microbiota-dependent trimethylamine N-oxide pathway contributes to the bidirectional relationship between intestinal inflammation and periodontitis. Front Cell Infect Microbiol. 2022, 12, 1125463. [Google Scholar] [CrossRef]
- Dong, Z; Liang, Z; Guo, M; Hu, S; Shen, Z; Hai, X. The association between plasma levels of trimethylamine N-oxide and the risk of coronary heart disease in Chinese patients with or without type 2 diabetes mellitus. Dis Markers 2018, 2018, 1578320. [Google Scholar] [CrossRef]
- Andraos, S; Jones, B; Lange, K; Clifford, S; Thorstensen, EB; Kerr, JA; et al. Trimethylamine N-oxide (TMAO) is not associated with cardiometabolic phenotypes and inflammatory markers in children and adults. Curr Dev Nutr. 2021, 5(1), nzaa179. [Google Scholar] [CrossRef]
- Hahn, VS; Petucci, C; Kim, M; Bedi, K; Wang, H; Mishra, S; et al. Myocardial metabolomics of human heart failure with preserved ejection fraction. Circulation 2023, 147(15), 1147–1161. [Google Scholar] [CrossRef]
- Benítez, TM; Vanderwoude, E; Han, Y; Byun, J; Konje, VC; Gillespie, BW; et al. Kynurenine pathway metabolites predict subclinical atherosclerotic disease and new cardiovascular events in chronic kidney disease. Clin Kidney J. 2022, 15(10), 1952–1965. [Google Scholar] [CrossRef] [PubMed]
- Hakan, H; Kurhan, F; Akbay, Hİ. Predictive value of kynurenine pathway metabolites in the severity of patients with obsessive-compulsive disorder. Psychiatry Clin Neurosci. 2025, 79(2), 13819. [Google Scholar] [CrossRef]
- Lewkowicz, J; Tankiewicz-Kwedlo, A; Pawlak, D; Kiluk, M; Lagoda, K; Kowalska, I. Kynurenines in heart failure with preserved ejection fraction: an influence of type 2 diabetes. Res Square [Preprint] 2024. [Google Scholar] [CrossRef] [PubMed]
- Regan, JA; Truby, LK; Tahir, UA; Katz, DH; Nguyen, M; Kwee, LC; et al. Protein biomarkers of cardiac remodeling and inflammation associated with HFpEF and incident events. Sci Rep. 2022, 12(1), 20488. [Google Scholar] [CrossRef]
- Tian, Y; Cai, J; Gui, W; Nichols, RG; Koo, I; Zhang, J; et al. Berberine directly affects the gut microbiota to promote intestinal farnesoid X receptor activation. Drug Metab Dispos. 2019, 47(2), 86–93. [Google Scholar] [CrossRef]
- Pathak, P; Xie, C; Nichols, RG; Ferrell, JM; Boehme, S; Krausz, KW; et al. Intestine farnesoid X receptor agonist and the gut microbiota activate G-protein bile acid receptor-1 signaling to improve metabolism. Hepatology 2018, 68(4), 1574–1588. [Google Scholar] [CrossRef]
- Kim, H; Fang, S. Crosstalk between FXR and TGR5 controls glucagon-like peptide 1 secretion to maintain glycemic homeostasis. Lab Anim Res. 2018, 34(4), 140–147. [Google Scholar] [CrossRef]
- Li, Y; Hou, H; Wang, X; Dai, X; Zhang, W; Tang, Q; et al. Diammonium glycyrrhizinate ameliorates obesity through modulation of gut microbiota-conjugated BAS-FXR signaling. Front Pharmacol. 2021, 12, 796590. [Google Scholar] [CrossRef]
- Ding, L; Zhang, E; Yang, Q; Jin, L; Sousa, KM; Dong, B; et al. Vertical sleeve gastrectomy confers metabolic improvements by reducing intestinal bile acids and lipid absorption in mice. Proc Natl Acad Sci U S A 2021, 118(6), e2019388118. [Google Scholar] [CrossRef]
- Zhang, C; Fang, R; Lu, X; Zhang, Y; Yang, M; Su, Y; et al. Lactobacillus reuteri J1 prevents obesity by altering the gut microbiota and regulating bile acid metabolism in obese mice. Food Funct. 2022, 13(12), 6688–6701. [Google Scholar] [CrossRef]
- Lee, S; Han, A; Kim, B; Sung, M; Hong, S. Lactococcus lactis-fermented spinach juice suppresses LPS-induced expression of adhesion molecules and inflammatory cytokines through the NF-κB pathway in HUVECs. Exp Ther Med. 2022, 23(6), 393. [Google Scholar] [CrossRef]
- Kim, D; Unenkhuu, B; Kim, G; Kim, S; Kim, H. Cynarin attenuates LPS-induced endothelial inflammation via upregulation of the negative regulator MKP-3. Anim Cells Syst (Seoul) 2022, 26(3), 119–128. [Google Scholar] [CrossRef] [PubMed]
- Gram, A; Kowalewski, M. Molecular mechanisms of lipopolysaccharide (LPS)-induced inflammation in an immortalized ovine luteal endothelial cell line (OLENDO). Vet Sci. 2022, 9(3), 99. [Google Scholar] [CrossRef] [PubMed]
- Tsioti, I; Steiner, B; Escher, P; Zinkernagel, M; Benz, P; Kokona, D. Endothelial toll-like receptor 4 is required for microglia activation in the murine retina after systemic lipopolysaccharide exposure. J Neuroinflammation 2023, 20(1), 162. [Google Scholar] [CrossRef] [PubMed]
- Cai, G; Yang, Z; Guo, D; Hu, C; Yan, M; Yan, J. Macrophages enhance lipopolysaccharide-induced apoptosis via ANG1 and NF-κB pathways in human umbilical vein endothelial cells. Sci Rep. 2021, 11(1), 3024. [Google Scholar] [CrossRef]
- Kutikhin, A; Shishkova, D; Velikanova, E; Sinitsky, M; Sinitskaya, A; Markova, V. Endothelial dysfunction in the context of blood-brain barrier modeling. J Evol Biochem Physiol. 2022, 58(3), 781–806. [Google Scholar] [CrossRef]
- Wong, E; Xu, F; Joffre, J; Nguyen, N; Wilhelmsen, K; Hellman, J. ERK1/2 has divergent roles in LPS-induced microvascular endothelial cell cytokine production and permeability. Shock. 2020, 55(3), 349–356. [Google Scholar] [CrossRef]
- Robles-Vera, I; Toral, M; Visitación, N; Aguilera-Sánchez, N; Redondo, J; Duarte, J. Protective effects of short-chain fatty acids on endothelial dysfunction induced by angiotensin II. Front Physiol. 2020, 11, 277. [Google Scholar] [CrossRef]
- Oh, J; Vasquez, R; Kim, S; Hwang, I; Song, J; Park, J; et al. Multispecies probiotics alter fecal short-chain fatty acids and lactate levels in weaned pigs by modulating gut microbiota. J Anim Sci Technol. 2021, 63(5), 1142–1158. [Google Scholar] [CrossRef]
- Colombo, A; Sadler, R; Llovera, G; Singh, V; Roth, S; Heindl, S; et al. Microbiota-derived short chain fatty acids modulate microglia and promote Aβ plaque deposition. Elife 2021, 10, e59826. [Google Scholar] [CrossRef]
- Xu, X; Jin, H; Li, X; Yan, C; Zhang, Q; Yu, X; et al. Fecal microbiota transplantation regulates blood pressure by altering gut microbiota composition and intestinal mucosal barrier function in spontaneously hypertensive rats. Res Square [Preprint] 2024. [Google Scholar] [CrossRef]
- Paramsothy, S; Nielsen, S; Kamm, MA; Deshpande, N; Faith, JJ; Clemente, JC; et al. Specific bacteria and metabolites associated with response to fecal microbiota transplantation in patients with ulcerative colitis. Gastroenterology 2019, 156(5), 1440–1454.e2. [Google Scholar] [CrossRef] [PubMed]
- Guirro, M; Costa, A; Gual-Grau, A; Herrero, P; Torrell-Galceran, H; Canela-Canela, N; et al. Effects from diet-induced gut microbiota dysbiosis and obesity can be ameliorated by fecal microbiota transplantation: a multiomics approach. PLoS One 2019, 14(9), e0218143. [Google Scholar] [CrossRef] [PubMed]
- Huang, X; Zhang, Y; Huang, J; Gao, W; Xie, Y; Zeng, C; et al. The effect of FMT and vitamin C on immunity-related genes in antibiotic-induced dysbiosis in mice. PeerJ. 2023, 11, e15356. [Google Scholar] [CrossRef] [PubMed]
- Cammarota, G; Ianiro, G; Kelly, C; Mullish, BH; Allegretti, JR; Kassam, Z; et al. International consensus conference on stool banking for faecal microbiota transplantation in clinical practice. Gut 2019, 68(12), 2111–2121. [Google Scholar] [CrossRef]
- Sun, L; Shang, B; Lv, S; Liu, G; Qiu, W; Geng, Y. Effects of semaglutide on metabolism and gut microbiota in high-fat diet-induced obese mice. Front Pharmacol. 2025, 16, 1562896. [Google Scholar] [CrossRef]
- Dupas, T; Pelé, T; Dhot, J; Burban, M; Persello, A; Aillerie, V; et al. The endothelial dysfunction could be a cause of heart failure with preserved ejection fraction development in a rat model. Oxid Med Cell Longev. 2022, 2022, 7377877. [Google Scholar] [CrossRef]
- Drekonja, D; Shaukat, A; Zhang, JH; Reinink, AR; Nugent, S; Dominitz, JA; et al. Microbiota or placebo after antimicrobial therapy for recurrent Clostridioides difficile at home: a clinical trial with novel home-based enrollment. Clin Trials 2021, 18(5), 622–629. [Google Scholar] [CrossRef]
- Ooijevaar, RE; Terveer, EM; Verspaget, HW; Kuijper, EJ; Keller, JJ. Clinical application and potential of fecal microbiota transplantation. Annu Rev Med. 2019, 70, 335–351. [Google Scholar] [CrossRef]
- Szulińska, M; Łoniewski, I; van Hemert, S; Sobieska, M; Bogdański, P. Dose-dependent effects of multispecies probiotic supplementation on the lipopolysaccharide (LPS) level and cardiometabolic profile in obese postmenopausal women: a 12-week randomized clinical trial. Nutrients 2018, 10(6), 773. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z; Chen, X; Sheng, H; Shen, X; Sun, X; Yan, Y; et al. Engineering probiotics as living diagnostics and therapeutics for improving human health. Microb Cell Fact. 2020, 19(1), 56. [Google Scholar] [CrossRef] [PubMed]
- Grumet, L; Tromp, Y; Stiegelbauer, V. The development of high-quality multispecies probiotic formulations: from bench to market. Nutrients 2020, 12(8), 2453. [Google Scholar] [CrossRef] [PubMed]
- Przerwa, F; Kukowka, A; Kotrych, K; Uzar, I. Probiotics in prevention and treatment of cardiovascular diseases. Herba Pol. 2021, 67(4), 77–85. [Google Scholar] [CrossRef]
- Khan, S; Ahmad, F; Khalid, N. Applications of strain-specific probiotics in the management of cardiovascular diseases: a systematic review. Mol Nutr Food Res. 2024, 68(8), e2300675. [Google Scholar] [CrossRef]
- Dixon, A; Robertson, K; Yung, A; Que, MS; Randall, H; Wellalagodage, T; et al. Efficacy of probiotics in patients at cardiovascular disease risk: a systematic review and meta-analysis. Curr Hypertens Rep. 2020, 22(9), 66. [Google Scholar] [CrossRef]
- Singh, RP; Shadan, A; Ma, Y. Biotechnological applications of probiotics: a multifarious weapon to disease and metabolic abnormality. Probiotics Antimicrob Proteins 2022, 14(6), 1184–1210. [Google Scholar] [CrossRef]
- Suez, J; Zmora, N; Elinav, E. Probiotics in the next-generation sequencing era. Gut Microbes 2020, 11(1), 77–93. [Google Scholar] [CrossRef]
- Videja, M; Sevostjanovs, E; Upmale-Engela, S; Liepinsh, E; Konrāde, I; Dambrova, M. Fasting-mimicking diet reduces trimethylamine N-oxide levels and improves serum biochemical parameters in healthy volunteers. Nutrients 2022, 14(5), 1093. [Google Scholar] [CrossRef]
- Goldiș, A; Dragomir, R; Mercioni, MA; Goldiș, C; Sirca, D; Enătescu, I; et al. Personalized microbiome modulation to improve clinical outcomes in pediatric inflammatory bowel disease: a multi-omics and interventional approach. Microorganisms 2025, 13(5), 1047. [Google Scholar] [CrossRef]
- Zamparette, CP; Teixeira, BL; Cruz, GNF; Filho, VB; Oliveira, LFV. Individual effect of diet on postprandial glycemic response and its relationship with gut microbiome profile in healthy subjects: protocol for a series of randomized N-of-1 trials. medRxiv [Preprint] 2023. [Google Scholar] [CrossRef]
- Shoer, S; Shilo, S; Godneva, A; Ben-Yacov, O; Rein, M; Wolf, BC; et al. Impact of dietary interventions on pre-diabetic oral and gut microbiome, metabolites and cytokines. Nat Commun. 2023, 14(1), 5716. [Google Scholar] [CrossRef] [PubMed]
- Iorra, FQ; Rodrigues, PG; Bock, PM; Guahnon, MP; Eller, S; Oliveira, TF; et al. Gut microbiota metabolite TMAO and adolescent cardiometabolic health: a cross-sectional analysis. J Endocr Soc. 2025, 9(5), bvaf055. [Google Scholar] [CrossRef] [PubMed]
- Hill, EB; Chen, L; Bailey, MT; Khalsa, AS; Maltz, R; Kelleher, KJ; et al. Facilitating a high-quality dietary pattern induces shared microbial responses linking diet quality, blood pressure, and microbial sterol metabolism in caregiver-child dyads. Gut Microbes 2022, 14(1), 2150502. [Google Scholar] [CrossRef]
- Saxena, R; Sharma, V; Saxena, AR; Patel, A. Harnessing AI and gut microbiome research for precision health. J Artif Intell Gen Sci. 2024, 3(1), 74–88. [Google Scholar] [CrossRef]
- Karwowska, Z; Szczerbiak, P; Kościółek, T. Microbiome time series data reveal predictable patterns of change. Microbiol Spectr. 2024, 12(10), e04109-23. [Google Scholar] [CrossRef]
- Mendes-Soares, H; Raveh-Sadka, T; Azulay, S; Edens, K; Ben-Shlomo, Y; Cohen, Y; et al. Assessment of a personalized approach to predicting postprandial glycemic responses to food among individuals without diabetes. JAMA Netw Open. 2019, 2(2), e188102. [Google Scholar] [CrossRef]
- Ye, Y; More, SK; Hamade, H; Alexeev, EE; Shimodaira, Y; Blackwood, AY; et al. TL1A overexpression in Crohn’s disease and mice alters Paneth cells and microbiota promoting ileal inflammation. bioRxiv [Preprint] 2025. [Google Scholar] [CrossRef]
- Fu, Y; Mei, Q; Yin, N; Huang, Z; Li, B; Luo, S; et al. Paneth cells protect against acute pancreatitis via modulating gut microbiota dysbiosis. mSystems 2022, 7(3), e01507-21. [Google Scholar] [CrossRef]
- Lee, VH; Gulati, A. Implications of Paneth cell dysfunction on gastrointestinal health and disease. Curr Opin Gastroenterol. 2022, 38(6), 535–540. [Google Scholar] [CrossRef]
- Du, W; Wang, J; Kuo, T; Wang, L; McKimpson, WM; Son, J; et al. Pharmacological conversion of gut epithelial cells into insulin-producing cells lowers glycemia in diabetic animals. J Clin Invest. 2022, 132(24), e162720. [Google Scholar] [CrossRef] [PubMed]
- Wang, J; Tian, F; Wang, P; Zheng, H; Zhang, Y; Tian, H; et al. Gut microbiota as a modulator of Paneth cells during parenteral nutrition in mice. JPEN J Parenter Enteral Nutr. 2018, 42(8), 1280–1287. [Google Scholar] [CrossRef] [PubMed]
- Sun, S; Hodel, M; Wang, X; Vicente, J; Haritunians, T; Debebe, A; et al. Macrophage LRRK2 hyperactivity impairs autophagy and induces Paneth cell dysfunction. Sci Immunol. 2024, 9(101), eadi7907. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H; Zhou, S; Gillilland, M; Li, J; Lee, A; Gao, J; et al. Bile acid toxicity in Paneth cells contributes to gut dysbiosis induced by high-fat feeding. JCI Insight 2020, 5(20), e138881. [Google Scholar] [CrossRef]
- Ahlawat, S; Asha, A; Sharma, KK. Gut–organ axis: A microbial outreach and networking. Lett Appl Microbiol. 2021, 72(6), 636–668. [Google Scholar] [CrossRef]
- Srugo, SA; Bloise, E; Nguyen, T; Connor, KL. Impact of maternal malnutrition on gut barrier defense: implications for pregnancy health and fetal development. Nutrients 2019, 11(6), 1375. [Google Scholar] [CrossRef]
- Rivera, K; González, L; Bravo, L; Manjarres, L; Andía, ME. The gut–heart axis: molecular perspectives and implications for myocardial infarction. Int J Mol Sci. 2024, 25(22), 12465. [Google Scholar] [CrossRef]
- Jaimez-Alvarado, S; López-Tenorio, II; Santos, J; Bello-Vega, DC; Gómez, FJR; Amedei, A; et al. Gut-heart axis: microbiome involvement in restrictive cardiomyopathies. Biomedicines 2025, 13(1), 144. [Google Scholar] [CrossRef]
- Kessler, EL; Oerlemans, MI; van den Hoogen, P; Yap, C; Sluijter, JPG; de Jager, SC. Immunomodulation in heart failure with preserved ejection fraction: current state and future perspectives. J Cardiovasc Transl Res. 2021, 14(1), 63–74. [Google Scholar] [CrossRef]
- Shi, Y; Perez-Bonilla, P; Chen, X; Tam, K; Marshall, M; Morin, J; et al. Metabolic syndrome nonalcoholic steatohepatitis male mouse with adeno-associated viral renin as a novel model for heart failure with preserved ejection fraction. J Am Heart Assoc. Published online. 2024. [CrossRef]
- Doiron, JE; Xia, H; Yu, X; Nevins, A; LaPenna, KB; Sharp, TE; et al. Adjunctive therapy with an oral H2S donor provides additional therapeutic benefit beyond SGLT2 inhibition in cardiometabolic heart failure with preserved ejection fraction. Br J Pharmacol. 2024, 181(21), 4294–4310. [Google Scholar] [CrossRef]
- Tong, D; Schiattarella, GG; Jiang, N; Altamirano, F; Szweda, PA; Elnwasany, A; et al. NAD+ repletion reverses heart failure with preserved ejection fraction. Circ Res. 2021, 128(11), 1629–1641. [Google Scholar] [CrossRef] [PubMed]
- Parajuli, P; Lara-Garcia, O; Regmi, MR; Skoza, W; Bhattarai, M; Kulkarni, A; et al. Heart failure drug class effects on 30-day readmission rates in patients with heart failure with preserved ejection fraction: a retrospective single-center study. Medicines (Basel) 2020, 7(5), 30. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G; Chen, J; Wu, C; Jiang, P; Wang, Y; Zhang, Y; et al. Deciphering the protein, modular connections and precision medicine for heart failure with preserved ejection fraction and hypertension based on TMT quantitative proteomics and molecular docking. Front Physiol. 2021, 12, 607089. [Google Scholar] [CrossRef] [PubMed]
- Aimo, A; Senni, M; Barison, A; Panichella, G; Passino, C; Bayés-Genís, A; et al. Management of heart failure with preserved ejection fraction: from neurohormonal antagonists to empagliflozin. Heart Fail Rev. Published online. 2022. [CrossRef]
- Herisson, FM; Cluzel, G; Llopis-Grimalt, MA; O’Donovan, AN; Koç, F; Karnik, K; et al. Targeting the gut-heart axis improves cardiac remodeling in a clinical scale model of cardiometabolic syndrome. JACC Basic Transl Sci. 2025, 10(1), 1–15. [Google Scholar] [CrossRef]
- Withaar, C; Meems, LMG; Markousis-Mavrogenis, G; Boogerd, CJ; Silljé, HH; Schouten, EM; et al. The effects of liraglutide and dapagliflozin on cardiac function and structure in a multi-hit mouse model of heart failure with preserved ejection fraction. Cardiovasc Res. 2021, 117(9), 2108–2124. [Google Scholar] [CrossRef]
- Momot, K; Krauz, K; Czarzasta, K; Zarębiński, M; Puchalska, L; Wojciechowska, M. Evaluation of nitrosative/oxidative stress and inflammation in heart failure with preserved and reduced ejection fraction. Int J Mol Sci. 2023, 24(21), 15944. [Google Scholar] [CrossRef]
- Guivala, SJ; Bode, KA; Okun, JG; Kartal, E; Pohl, LV; Werner, S; et al. Interactions between the gut microbiome, associated metabolites and the manifestation and progression of heart failure with preserved ejection fraction in ZSF1 rats. Res Square [Preprint] 2024. [Google Scholar] [CrossRef]
- Gaćeša, R; Kurilshikov, A; Vila, AV; Sinha, T; Klaassen, MAY; Bolte, LA; et al. The Dutch Microbiome Project defines factors that shape the healthy gut microbiome. bioRxiv [Preprint] 2020. [Google Scholar] [CrossRef]
- Wagenaar, CA; van de Put, M; Bisschops, M; Walrabenstein, W; de Jonge, CS; Herrema, H; et al. The effect of dietary interventions on chronic inflammatory diseases in relation to the microbiome: a systematic review. Nutrients 2021, 13(9), 3208. [Google Scholar] [CrossRef]
- Krüger-Genge, A; Jung, F; Hufert, F; Küpper, J; Storsberg, J. Effects of gut microbial metabolite trimethylamine N-oxide (TMAO) on platelets and endothelial cells. Clin Hemorheol Microcirc. 2020, 76(2), 309–316. [Google Scholar] [CrossRef]
- Joishy, TK; Jha, AR; Oudah, M; Das, S; Adak, A; Deb, D; et al. Human gut microbes associated with systolic blood pressure. Int J Hypertens. 2022, 2022, 2923941. [Google Scholar] [CrossRef]
- Valencia, S; Zuluaga, M; Pérez, M; Montoya-Quintero, KF; Candamil-Cortés, MS; Robledo, S. Human gut microbiome: a connecting organ between nutrition, metabolism, and health. Int J Mol Sci. 2025, 26(9), 4112. [Google Scholar] [CrossRef]
- Jama, H; Dona, MS; Dinakis, E; Nakai, M; Paterson, M; Shihata, W; et al. Maternal diet and gut microbiota influence predisposition to cardiovascular disease in the offspring. bioRxiv [Preprint] 2022. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



