Submitted:
29 January 2026
Posted:
30 January 2026
You are already at the latest version
Abstract
Responses to cutaneous injury differ fundamentally across developmental stages. During early human gestation, when the fetus is less than 24 weeks old, wounds are capable of restoring normal tissue architecture without forming fibrotic scars. In con-trast, postnatal and adult injuries typically resolve through the process of fibrosis. This divergence reflects coordinated differences in epidermal and dermal compartments, inflammatory signaling, extracellular matrix (ECM) composition, mechanical cues, and gene regulation. Recent studies have demonstrated that dermal fibroblasts are no longer considered a uniform population but instead arise from distinct developmental lineages with stable functional identities. Engrailed-1–negative fibroblasts (ENFs) predominate in early fetal skin and support regenerative repair, while Engrailed-1–positive fibroblasts (EPFs) emerge later in development and are the principal contrib-utors to fibrotic matrix deposition following injury. The developmental shift between these fibroblast populations coincides with the loss of scar-free healing capacity. This review examines current understanding of fibroblast lineage specification, with par-ticular emphasis on the roles of mechanotransduction, ECM cues, and epigenetic reg-ulation. Elucidating how these lineage-encoded programs are established and main-tained may enable strategies to reprogram adult fibroblasts toward a fetal-like regen-erative state and thereby promote scar-free tissue repair.
Keywords:
skin
; wound healing
; scar
; regeneration
; fibroblast
1. Introduction
The skin functions as a multifunctional organ that provides mechanical protection, regulates fluid balance, and mediates immune surveillance. Structurally, it consists of a stratified epidermis anchored to an underlying dermis rich in ECM and mesenchymal cells. Following injury, the skin initiates a coordinated repair response aimed at restoring barrier integrity and preventing infection or fluid loss [1].
Cutaneous wound healing is traditionally described as a sequence of partially overlapping phases, including inflammation, proliferation characterized by tissue formation, and remodeling [2]. Injuries sustained during early fetal development (<24 weeks) typically heal through the complete restoration of native tissue architecture, including regeneration of hair follicles and glands, without visible scar formation [3,4,5,6]. After mid-gestation, however, this regenerative capacity is progressively lost, and healing increasingly resembles the fibrotic response observed in adults.
Skin repair is the result of dynamic and interactive processes that involve soluble factors, ECM components, blood elements, and cells. Re-epithelialization is a critical step in the wound healing process since defects in the formation of new epidermis lead to chronic non-healing wounds [7]. In adults, there is wound contraction due to fibroblasts pulling on the ECM [8]. As the inflammatory phase concludes, the fibroblasts enter the wound to initiate granulation tissue formation, which in adults is the precursor to the mature scar. Due to exposure to transforming growth factor-β (TGF-β) and the ECM of the adult wounds, fibroblasts take on a contractile phenotype named myofibroblasts. This is a critical step in the adult wound healing process, which contributes to wound closure via contraction [9]. Despite initial thoughts that embryonic fibroblasts were incapable of turning into myofibroblasts, experimental data concluded that, although ephemeral, a transition of embryonic fibroblasts to myofibroblasts was possible after exposure to large amounts of TGF-β1, in vitro [10,11]. Nevertheless, induced embryonic myofibroblasts produced less collagen, whereas the transplantation of embryonic fibroblasts to adult wounds resulted in healing with a reticular collagen pattern, indistinguishable from that of healthy skin [12]. In fact, mammalian embryos use rapid re-epithelialization in the absence of extensive inflammation and granulation tissue formation. This is a scarless process driven by the rapid formation of a contractile actin-myosin “purse-string” cable at the wound margins that pulls cells together to seal the gap [13,14].
Hence, a central challenge in regenerative medicine is to elucidate the mechanisms underlying fetal skin regeneration following injury, as opposed to adult wound scarring, and to reactivate these regenerative mechanisms for therapeutic purposes. Historically, research has focused on studying and analyzing the differences in inflammatory responses, ECM composition, growth factor profiles, and levels of mechanical tension between fetal and adult skin [5,15,16,17]. Although each of the mentioned factors contributes to the overall outcome, they do not completely explain why the fetal environment allows regeneration, nor how these permissive conditions are lost during development.
Recent studies have excluded environmental factors as the cause of scarless repair and suggest that scarless healing is most likely an intrinsic property of the early embryonic skin [4]. This suggestion is in line with the relatively recent evidence that distinct fibroblasts lineages exist in all mammals’ skin that represent unique cell types, and their switch from one type to the other during development imposes the transition from regeneration to scarring [18,19,20]. Since the early 1990s, the influence of the environment has been assessed as the principal factor that may influence fibroblast behavior and the initiation of scarring in adult healing. Recent lineage-tracing studies have introduced a different concept. Fibroblast identity —and more specifically, the developmental switch from ENFs to EPFs— is most likely a primary determinant of whether a wound heals through regeneration or fibrosis. ENFs, which dominate early fetal skin, are intrinsically pro-regenerative. EPFs, which emerge later in development and dominate postnatally, are intrinsically pro-fibrotic and responsible for the scar ECM deposition. This lineage switch appears to coincide with the loss of scarless healing property, suggesting that fibroblast ontogeny may be a compelling mechanistic driver of the regenerative-to-fibrotic transition [21,20,19,22].
Understanding the signals behind fibroblast lineage specification may therefore unlock the ability to reprogram adult fibroblasts into a fetal-like state. Such reprogramming would challenge the presumed irreversibility of adult fibrosis and enable therapeutic strategies that promote true tissue regeneration.
This review summarizes current knowledge on the developmental origin of fibroblast heterogeneity, the molecular and mechanical factors driving lineage switching, and emerging strategies for recapitulating fetal-like healing in adults.
2. Overview of Adult Wound Healing vs. Fetal Regenerative Healing
2.1. Fetal vs. Adult Wound Healing: A Comparative Overview
Scarless healing in humans is observed before approximately 24 weeks of gestation, after which injuries begin to heal with progressively more adult-like fibrosis [23]. Healing of the fetal wounds withheld during this early window demonstrate the following typical characteristics: 1. rapid coverage with new epithelium without formation of granulation tissue which postnatally is correlated with scar tissue formation [24], 2. restoration of skin appendages, including hair follicles [25] and 3. complete collagen architecture regeneration rather than deposition of aligned bundles [26,27]. Consequently, adult cutaneous wound healing results in incomplete regeneration of the original tissue, excessive production of an unorganized collagen meshwork that forms the scar, loss of appendages, and a flattened epidermis. In fact, the newly formed tissue has a lower tensile strength of less than 70-80% [25,28].
The advancement of biological events during the wound healing process, i.e., hemostasis, inflammation, proliferation, and remodeling, does not depend on age. However, the quality, magnitude, and timing of these processes are profoundly different [16,29].
Hemostasis, by definition, means to stop the hemorrhage. When a trauma occurs, this is achieved via the formation of the fibrin clot, which is created by the platelets leaking from the disrupted vasculature. At the same time, the platelets degranulate and release cytokines and growth factors that attract other types of inflammatory cells to the wound site. The latter is what induces the next phase, namely the inflammatory phase. The first inflammatory cells to be recruited are neutrophils and monocytes, followed by the macrophages [4]. These inflammatory cells clear the wound bed from microorganisms and tissue debris but also release chemoattractant signals that act on adaptive immune cells, such as mast cells, dendritic cells, and T-lymphocytes [30], fibroblasts, and endothelial cells to form the granulation tissue [4,9]. As the inflammatory phase progresses, fibroblasts from the reticular dermis [31] adhere to fibronectin (FN)deposited within the fibrin clot and reach the wound. There, they start the production and deposition of the granulation tissue [32] rich in collagens and other ECM components, such as hyaluronic acid (HA), FN, and tenascin C [33]. Subsequently, in response to TGF-β1 signaling originating from the immune cells and likely Wnt/β-catenin signaling [31,34], and the ECM of the wound bed, fibroblasts from the reticular dermis are activated and turn into contractile myofibroblasts [10,31,35]. In postnatal wound healing, the transition of fibroblasts to myofibroblasts is critical for wound closure via contraction of the wound edges achieved through interaction with the ECM [34]. The formation of granulation tissue is a hallmark of the initiation of the proliferative phase that aims to replace the destroyed tissue. One essential event of this phase is the activation of keratinocytes, which simultaneously migrate over the granulation tissue and proliferate to compensate for the loss in cell number, leading to the formation of new epithelium, the so-called re-epithelialization phase [4,36]. The wound healing process concludes with the remodeling phase, which occurs 2 to 3 weeks post-injury and results in the formation of scar tissue. This phase is characterized by a reduction in the cell component of the wound milieu via apoptosis, rearrangement of collagen fibres, regression of neovascularization and the deposition of components of the ECM [23,30]. The scar tissue presents a different orientation of collagen, a disorganized elastic fiber network, more dermatan sulfate, and less HA as compared to the normal skin, making it less extensible [23].
As for the early-gestation fetal wounds, a markedly different trajectory is followed. Scar formation is considered as a solution to the age-related, delayed re-epithelialization process. However, the fetal wound healing is characterized by the remarkably rapid regeneration of the injured epithelium [29]. Overall, inflammation is limited in both scale and persistence, granulation tissue formation is minimal or absent, and tissue replacement occurs without the prolonged activation of fibrogenic pathways. Evidence suggests that fetal keratinocytes present different properties and use a different mechanism to migrate and resurface the disrupted area. Not only do they proliferate faster, but they also form a cable that runs from cell to cell and contracts the keratinocytes together to close the wound within hours after wounding [29]. As a result, fetal wounds resolve with restoration of normal dermal and epidermal organization, an outcome that has been observed across multiple mammalian species [21]. Early assumptions attributed this regenerative outcome to environmental factors such as reduced immune activation or the sterile intrauterine milieu. However, experimental evidence demonstrating scarless healing in fetal skin transplanted into adult environments has challenged this view, suggesting that regenerative capacity is governed primarily by intrinsic properties of the tissue itself [37]. Interestingly, strong evidence suggests that scarless wound healing in the oral mucosa is also derived from intrinsic differences of the tissue rather than from the environment [25].
As described in detail in the following sections, fetal wound healing reflects a coordinated biological state in which inflammation is muted, the ECM is soft and hydrated, cytokine signals favor regeneration, and fibroblasts exist in a plastic, pro-regenerative state. In adult skin, these parameters shift toward inflammation, mechanical stiffness, and pro-fibrotic signaling, creating an environment that supports scarring. Therefore, the capacity of early-gestation human embryos’ skin to heal without scarring reflects a fundamentally different biological program as compared to the fibrotic wound healing observed in adults [5,6,29]. The contrasting outcomes of fetal and adult repair therefore, reflect not merely quantitative differences in wound healing responses, but the engagement of fundamentally different tissue programs.
2.2. Inflammatory Response
One of the most prominent biological distinctions between regenerative fetal and fibrotic postnatal wound healing is the nature of the inflammatory response [23,24,25].
The inflammatory response is a prerequisite for successful wound healing in adults [2] that initiates within a few minutes to hours upon injury. The postnatal healing initiates with a robust inflammatory response with high neutrophil and macrophage infiltration and the release of pro-inflammatory cytokines and growth factors with pleiotropic roles in activating and/or regulating cellular responses. Initially, disruption of the vasculature drives platelet aggregation from the circulation into the wound site and formation of the clot [23]. The latter is accompanied by degranulation of platelets, the release of platelet-derived growth factor (PDGF), TGF-β1, tumor necrosis factor α (TNFα), and interleukin-1 (IL-1) that activate resident cells and recruit macrophages to the wound site [24,25]. The pro-inflammatory cytokines IL-6 and -8 are also produced during the inflammatory phase. These signals further induce activation of the migratory phenotype in keratinocytes at the wound edges and fibroblasts’ recruitment and transformation to myofibroblasts, which in turn, collectively release more cytokines and growth factors acting in an autocrine or paracrine manner [2,16,23]. Vascular endothelial growth factor (VEGF) is also upregulated in adult wounds, mediating wound angiogenesis [25]. Therefore, inflammatory cells may have the central role during the inflammatory stage; nevertheless, non-inflammatory cells and the ECM they secrete contribute as well.
The fetal wound healing process is characterized by a minimal inflammatory response. The main differences are recapitulated as follows. To begin with, platelets exhibit a variation depending on the stage of gestation. There is evidence suggesting that platelet aggregation and degranulation are decreased in early gestation embryos. As a result, lower levels of TGF-β and PDGF are released at the wound site. Some in vitro studies have shown that platelet-rich plasma (PRP) produced by platelets possibly induces dermal fibroblast differentiation to myofibroblasts [38]. However, contradictory results from different studies do not allow us to conclude. In turn, less TGF-β1 has been correlated with reduced neutrophil recruitment [39]. Moreover, levels of the pro-inflammatory IL-6 and IL-8 are characteristically lower in scarless healing, contributing to the lower acute inflammatory infiltrate at the wound bed. Overall, fewer macrophages, neutrophils, dendritic cells, mast cells (degranulating less effectively), T-cells, and Langerhans cells are detected within the fetal tissue, while macrophages are mostly represented by the M2 (anti-inflammatory) phenotype [4,24,38,39]. In early gestation fetuses, macrophages are not recruited to fetal wound sites due to lower expression levels of TGF-β1 to which contributes to the conversion of circulating monocytes to macrophages [25]. Instead, in scarless wounds, there is another version of TGF-β, i.e., TGF-β3, elevated, and its presence has been linked to hypoxia [40] and as an inhibitory signal for terminal differentiation [41]. It is likely that the presence of TGF-β3 keeps the tissue in an immature state that impacts the release of proinflammatory cytokines and eventually favors healing with minimal scar formation [42]. Another cytokine detected in scar-forming mice that induced scar formation into scarless embryo wounds in an experimental setting is IL-33 [39]. On the contrary, the anti-inflammatory cytokine IL-10 is highly expressed in fetal wounds. Finally, fibroblasts that actively participate in the immune regulation present age-related differences in the expression of major histocompatibility complex (MHC) class I and II in humans [23].
The relative absence of inflammation in fetal skin is thought to protect fibroblasts from adopting a pro-fibrotic identity and to maintain a pro-regenerative milieu.
2.3. Extracellular Matrix Composition
Marked differences in the composition and structural organization of the ECM characterize fetal versus adult wound healing. These variations are critical because they fundamentally alter the biochemical milieu and biomechanical properties of the matrix. In doing so, they shape the quality and identity of the newly formed tissue, influence the behavior and phenotypic state of the cells participating in repair, and modulate the complex network of mechanical and molecular cues that orchestrate the wound-healing response [17].
Collagen is a key structural component of the skin, both in the fetal and later stages, with type one collagen predominating in all phases. Its mechanical robustness is derived from a three-stranded helical arrangement of polypeptide chains, whose crosslinking and stabilization are mediated by the activity of lysyl oxidase [32]. However, in fetal skin, there is a predominance of collagen III και V over collagen I, which decreases over time. Correspondingly, fetal fibroblasts show an increased ratio of collagen III και V to collagen I, compared to neonatal and adult fibroblasts [43]. Additionally, increased expression was observed in collagens IV and XIV in fetal versus neonatal and adult fibroblasts [43]. The specific collagen profile within the ECM plays a central role in directing fibrillogenesis, thereby determining both the diameter of individual fibrils and the organization of collagen bundles. Collagen type I promotes the formation of relatively thick fibrils and is associated with a slower turnover compared with type III. Type V is required for the proper assembly of collagen I and III fibrils, while collagens V and XIV function as regulatory elements that constrain the ultimate diameter of the collagen fibers [44,45]. This explains the fact that in fetuses, collagen deposition occurs in thin, reticular and more organized bundles, while in adults it occurs in denser and parallel bundles [46]. By extension, it is understood that while cross-linking of type I collagen is critical for conferring tensile strength during adult wound repair, the resulting matrix stiffness can restrict the mobility of key cellular mediators. In fetal tissues, this increased rigidity may hinder the swift cellular dynamics required for accelerated regenerative responses [47]. Moreover, data support that the more rapid wound healing in fetuses as compared to adults is achieved through the earlier secretion of higher amounts of total collagen (collagen types I, III, IV, V and VI) [17].
Another major component of the ECM are glycosaminoglycans (GAGs), particularly HA and chondroitin sulfate (CS) [5,48]. HA is a negatively charged, unsulfated GAG found in soluble form or complexed with proteoglycans, which increases during rapid cellular migration and angiogenesis. The overall negative charge of HA enables it to attract and transiently restrain water molecules, a property that contributes to resistance against mechanical deformation while simultaneously supporting efficient cell migration through the matrix and stimulating collagen synthesis by fibroblasts [32]. The HA content of scarred fetal wounds increases more rapidly compared to adults, while fetal fibroblasts also present higher amounts of HA receptors, which allows fibroblasts to maintain the amount of HA produced longer and promotes their migration [49]. Furthermore, increased expression of HA in embryos reduces the uptake of inflammatory cytokines such as IL-1 and TNF-a [50].
Proteoglycan-associated ECM regulators such as decorin and fibromodulin, along with enzymes including lysyl oxidase and the matrix metalloproteinases (MMPs), collectively participate in orchestrating the processes of collagen production, post-translational maturation, and controlled degradation [51]. Decreased expression of decorin and increased expression of fibromodulin have been observed after injury in early gestation compared to later developmental stages [52]. Reduced decorin has been shown to cause reduced tensile strength [53] and regulate collagen fiber formation, while fibromodulin has been associated with anti-inflammatory function and increased cellular migration [54]. In general, embryonic fibroblasts exhibit elevated levels of enzymes involved in collagen cross-linking—including lysyl hydroxylase (LH), prolyl hydroxylase (PH), low lysyl oxidase (LOX) [55], and transglutaminase 2 (TGM2) [56]—a pattern of expression that is accompanied by enhanced collagen production [17]. LOX is expressed at higher levels during adult tissue repair, and its upregulation has been associated with the development of fibrotic pathologies [57]. Nevertheless, findings from another investigation reported that fetal wounds exhibit higher LOX expression relative to neonatal wounds, indicating that additional studies are needed to clarify this discrepancy [58]. Furthermore, the expressions of MMPs, which are necessary for the release and migration of cells from the ECM, have been observed to be higher and faster in early embryonic wounds, while at the same time, the higher expression of MMPs/TIMPs (tissue inhibitors of metalloproteinases) ratio may facilitate the recycling of the ECM towards scar-free healing [17].
Fetal wounds that heal without scarring exhibit a faster induction of ECM adhesion molecules and a distinct pattern of integrin expression on the cell surface. During early human embryogenesis, elastin is present at minimal levels because its synthesis peaks in the late embryonic and neonatal periods and subsequently declines, resulting in limited regenerative capacity in adult tissues [5,48]. Conversely, FN is more abundant in fetal skin, preceding and facilitating the deposition of tenascin, a matrix component that suppresses cell adhesion and emerges earlier in fetal repair [5,59]. Laminin expression remains stable across developmental stages in human fibroblasts [60]. Moreover, fetal fibroblasts display elevated levels of integrin α2 and reduced levels of α1 and α3 compared with adult cells, a profile associated with their diminished ability to contract collagen matrices [61].
It is important to recognize that scarless fetal healing is not uniform across all tissues. During early gestation—when the skin is capable of regenerating without fibrosis—organs such as the fetal stomach, intestine, and diaphragm nonetheless undergo scar formation following injury. This disparity implies that specific skin-resident cell populations play a crucial role in directing the localized wound-healing response [62,63]. Conversely, evidence indicates that the uterine (endometrial) environment alone does not determine, nor can it independently induce, a scarless pattern of repair [64,65].
The fetal ECM is compositionally distinct from the adult ECM and plays a major role in influencing fibroblast behavior. The ECM most likely does not merely reflect fibroblast activity but actively drives it: fetal ECM provides cues that support regenerative fibroblast phenotypes, while adult ECM reinforces pro-fibrotic behavior. The question that arises here, though, is who and through which mechanism produces this ECM after all that favors one population of fibroblasts over the other.
2.4. Biochemical Signaling Through the ECM
Beyond its structural role, the ECM functions as a dynamic biochemical signaling platform that instructs fibroblast fate and behavior. Fibroblasts actively sense ECM composition, organization, and stiffness through integrins, proteoglycan receptors (such as syndecans), and mechanosensitive ion channels, translating extracellular cues into intracellular signaling cascades that regulate proliferation, migration, differentiation, and fibrotic versus regenerative outcomes.
Members of the transforming growth factor family play a central role in the regulation of wound repair. TGF-β functions as a potent chemoattractant for fibroblasts, keratinocytes, and immune cells, while also promoting collagen type I synthesis by fibroblasts [66]. In fetal wounds, TGF-β3 is expressed at markedly elevated levels; this factor, produced mainly by keratinocytes and fibroblasts, is closely associated with skin morphogenesis. By contrast, the expression of TGF-β1 and TGF-β2 remains minimal in fetal repair. Adult wounds display an opposing profile, being dominated by TGF-β1 and TGF-β2, which initially originates from platelet degranulation and is later supplied by infiltrating inflammatory cells, including monocytes and macrophages [67].
In this context, TGF-β acts as a regulator of MMP expression. TGF-β1 inhibits MMPs, mainly MMP1 [68]. MMPs mediate the proteolytic breakdown of ECM, and their activity within tissues is tightly controlled by TIMPs. The dynamic equilibrium between these enzymes and their inhibitors is a critical determinant of tissue remodeling, as it governs the turnover and persistence of the ECM. Embryonic wounds are characterized by an elevated MMP-to-TIMP ratio, resulting in enhanced degradation of ECM components. This enzymatic environment promotes dynamic matrix remodeling rather than excessive ECM deposition at the site of injury [69]. Similarly, adult wound environments are enriched in PDGF, a factor that is largely absent from embryonic wounds due to minimal platelet degranulation. In contrast, embryonic wounds exhibit elevated levels of endogenous fibroblast growth factors (FGFs) that are associated with skin morphogenetic processes [70]. VEGF, a potent mitogen for endothelial cells, is upregulated approximately two-fold in wounds that heal without scarring, whereas its expression remains unchanged in fetal wounds that undergo fibrotic repair. This enhanced angiogenic and permeability-promoting signal may contribute to the accelerated healing observed in scarless fetal wounds [71].
ILs constitute a class of cytokines that play key roles in directing inflammatory cell recruitment and activation, thereby contributing to the regulation of wound repair. Notably, early embryonic fibroblasts exhibit markedly reduced expression of IL-6 and IL-8, both under basal conditions and following PDGF stimulation, when compared with adult fibroblasts [72]. In addition, fetal skin demonstrates higher levels of IL-10 expression relative to adult tissue [73].
The integrin–focal adhesion kinase (FAK), a key cytoplasmic tyrosine kinase, regulates cellular processes such as proliferation, survival, and signaling and so represents a key regulator of mechanotransduction in the skin [74]. During wound repair, FAK becomes activated in response to mechanical stimuli and subsequently modulates intracellular signaling through multiple downstream effectors, including Phosphatidylinositol 3-Kinase (PI3K) and Mitogen-Activated Protein Kinase (MAPK) pathways, which are closely linked to fibrotic outcomes [75]. Inhibition of the FAK signaling pathway suppresses the activity of the MAPK pathway, reduces glycolytic metabolism in trophoblast cells, and negatively affects the healing process [74]. FAK upregulation is mainly observed in mature tissues and is associated with increased scar formation [76] The transcriptional co-activators Yes-associated protein (YAP) and Transcriptional coactivator with PDZ-binding motif (TAZ), targets of the Hippo pathway, are activated by mechanical stimulation transmitted through FAK and RhoA/ROCK, leading to their translocation to the nucleus and activation of genes associated with fibroblast activation, such as connective tissue growth factor (CTGF) [77]. In skin fibroblasts, YAP is nuclear in proliferating cells and is mainly cytoplasmic in quiescent cells after birth, while in the early fetal it is mainly localized in the nucleus [78]. Accordingly, YAP/TAZ signaling is preferentially activated in adult skin as a consequence of elevated mechanical tension and increased ECM stiffness—conditions that are largely absent in fetal and neonatal tissues [79]. During early developmental stages, YAP/TAZ activity is stringently controlled and remains attenuated within mechanically compliant, low-stiffness microenvironments [80]. Moreover, Rho/ROCK signaling converges with the Hippo pathway through its role in mechanically regulating YAP and TAZ activity. ROCK-driven actomyosin contractility increases cytoskeletal tension, facilitating the nuclear translocation of YAP/TAZ and the subsequent activation of transcriptional programs linked to cell proliferation and lineage commitment. Compliant extracellular matrices are associated with reduced ROCK signaling and lower intracellular tension, conditions that favor maintenance of stem cell characteristics. In stiffer adult matrices, enhanced cellular contractility promotes YAP/TAZ activation, biasing stem cell fate toward epithelial or mesenchymal differentiation [81].
Wnt signaling plays a critical role in embryonic development and organogenesis. Wnt-responsive cells in dermal tissue include hair follicle bulge cells, basal interfollicular epidermal cells, and dermal fibroblasts [15]. In adult skin tissue, several Wnt proteins (Wnt 1, 3, 4, 5, and 10) are activated during normal wound healing, similar to cell proliferation signals for dermal fibroblasts and keratinocytes. In contrast, in fetal skin, Wnt4, Wnt5a, and Wnt11 are expressed in the dermis and play an important role in hair follicle morphogenesis [82].
2.5. Gene Expression and Transcriptional Programs
The transition from regenerative to fibrotic wound healing has been proposed to reflect, at least in part, a developmental compromise that compensates for the comparatively delayed re-epithelialization observed in adult skin relative to fetal skin. However, unveiling cause–and–effect relationships in this process remains challenging. Here, we provide an overview of differences in gene expression, transcriptional programs, and signaling cues that influence keratinocyte and fibroblast behavior in fetal versus adult skin. Keratinocytes are discussed because of their capacity to modulate fibrotic responses, whereas fibroblasts represent the principal ECM–producing cells and play a central role during the proliferative and remodeling phases of wound healing.
During fetal skin development, the epidermis transitions from a single-layered structure at early gestation into a stratified and keratinized epithelium by gestational weeks 22-24 [51]. At approximately 14 weeks of gestation, the fetal epidermis consists of a basal layer, one or two intermediate layers, and the periderm [5]. At this early stage, highly proliferative keratinocytes progressively commit to stratification, a process reflected by dynamic changes in keratin expression. Keratins K5 and K14 are selectively expressed in proliferating basal keratinocytes, K1 and K10 are induced at weeks 9-10 and mark keratinocytes committed to terminal differentiation in the suprabasal layers [5]. Additional keratins, such as K8, K17, and K19, are preferentially expressed in fetal keratinocytes and used as markers to distinguish them from adult keratinocytes in vitro [5,83].
Functionally, fetal keratinocytes display enhanced proliferative capacity in vivo [5] and increased clonogenicity in vitro, characterized by longer telomeres and decreased expression of MHC proteins [83]. Notably, fetal keratinocytes do not induce proliferation of naïve T cells, suggesting a mechanism of immune tolerance at the maternal-fetal interface during pregnancy. Consistent with this observation, fetal keratinocytes produce higher levels antimicrobial peptides compared to postnatal keratinocytes, including β-defensins, S100 protein family members, and cathelicidin, indicating that innate antimicrobial defense represents a critical protective strategy in the embryo [84].
At the transcriptional and epigenetic level, fetal keratinocytes exhibit distinct chromatin landscapes. Reduced levels of trimethylated histone H3k27 and increased expression of histone demethylase JMJD3 have been reported in fetal compared with postnatal and adult keratinocytes [84,85,86]. In contrast, postnatal keratinocytes display increased activation of Wnt signaling pathways [87]. Nuclear accumulation of β-catenin following Wnt-3α signaling promotes transcriptional activation via TCF/LEF enhancers and has been linked to profibrotic gene expression [88,89]. Additionally, members of the SRY-related high-mobility-group (HMG) box (SOX) family of transcription factors, particularly SOX-4 and SOX-11, regulate epidermal differentiation during embryogenesis through mechanisms that involve AP-1 transcription activity [90]. During adult wound healing, reactivation of embryonic gene programs, regulating keratinocyte migration has been shown to involve these transcription factors [91].
Accumulating evidence suggests that many of the fundamental differences between fetal and adult skin reside within the dermal compartment, implicating fibroblasts as key effectors of scarless repair. Supporting this concept, Besides, keratinocytes cultured on fetal human dermal fibroblasts present increased expression of cell-cycle-associated genes, while adult fibroblasts promote keratinocyte differentiation, reinforcing the idea that age-dependent changes in the ECM critically influence wound healing outcomes [92]. Fetal fibroblasts proliferate and migrate more rapidly than adult fibroblasts, while simultaneously synthesizing collagen [17,48]. These functional differences are partially attributable to distinct ECM composition and deposition kinetics. Fetal fibroblasts secrete higher amounts of total collagen, including collagen types I, III, V, IV, and XIV, with collagen I deposited more rapidly and at a lower relative percentage, resulting in thinner collagen fibres that favor scarless healing [17].
Proteomic analyses further revealed differences in cytoskeletal regulation and protein turnover between fetal and adult fibroblasts. Fetal fibroblasts express significantly higher levels of tubulin alpha 1 and relatively higher levels of actin, whereas adult fibroblasts exhibit increased expression of actin-regulating proteins such as fructose-biphosphate aldolase A, cofilin-1, profilin-1, which may impair ubiquitin-mediated protein degradation and delay protein turnover. Moreover, fetal fibroblasts demonstrate enhanced superoxide radical degradation and early activation of apoptosis-related signaling pathways following injury, suggesting that efficient detoxification of reactive oxygen species and timely removal of damaged cells contribute to scarless repair [88,93]. In contrast, adult fibroblasts overexpress proteins associated with pro-inflammatory signaling, granulation tissue formation, and cell adhesion, such as lactotransferrin, galectin-1, and calreticulin-1 [94]. More recently, fibrotic repair has been linked to fibroblast expression of the transcription factor EN-1, increased YAP activity, and suppression of Trichorhinophalangeal Syndrome Type I (Trps1), highlighting the contribution of mechanosensitive and lineage-associated transcriptional programs to scarring outcomes [89,95].
Together, these findings indicate that developmental stage–dependent differences in gene expression, mechanotransduction, and epigenetic regulation progressively restrict transcriptional plasticity in skin cells. Rather than acting as transient modulators of wound repair, these processes establish stable transcriptional states that shape how dermal cells respond to injury. Such transcriptional and epigenetic priming provides the foundation for the emergence of fibroblast populations with distinct functional identities, which are discussed in the following section.
The dermal layer consists of three compartments with different architecture and collagen deposition patterns, namely the papillary, the reticular, and the hypodermis, each containing different fibroblast populations that arise from varying lineages. Increasing evidence suggests that the dermis comprises at least two functionally distinct lineages of fibroblasts with various morphological and functional properties, as well as distinct roles in the wound healing process [reviewed in 94]. Evidence suggests that papillary fibroblasts are likely important contributors to the scarless healing process [97]. Neonatal papillary fibroblasts lose their ability to form hair follicle mesenchyme upon injury, and in turn, they are adapted to acquire their rapid wound healing properties and become specialized in the fibrous deposition matrix once major tissue developmental processes are completed [98]. This phenomenon can only be converted at the center of large wounds. It appears that genetic upregulation of developmental pathways, such as Wnt and Shh, in fibroblasts enhances their ability to form hair follicles in adult skin. Evidence suggests that histone modifications are responsible for the regulation of the developmental maturation of fibroblasts. For example, Kim et al showed that the transcription factor Twist 2 drives the decrease in acetylation of histone H3K27, which subsequently inactivates Wnt signaling, ultimately leading to postnatal fibroblast differentiation [98]. Other histone modifications, such as methylation of H3K27, seem to participate in the regulation of fibroblast maturation. Methylation seems to control the expression of αSMA and the differentiation to myofibroblasts [97]. Another possibility is that fibroblasts, similarly to keratinocytes, may carry epigenetic “memory” modifications that dictate their behavior upon injury. Interestingly, adult fibroblasts exhibit at the same time, remarkable plasticity and can convert to other cell types [97], although they demonstrate strong positional and behavioral stability when they are not perturbed [98]. Elucidating the origins of the distinct fibroblast subpopulations and the mechanisms that are responsible for their great plasticity is therefore of major importance.
3. Fibroblast Subpopulations and Lineage-Dependent Regulation of Wound Healing
The dermal layer of the skin is structurally and functionally heterogeneous, comprising distinct compartments—including the papillary dermis, reticular dermis, and hypodermis—that differ in ECM organization, mechanical properties, and cellular composition [99,100,101]. Fibroblasts residing within these compartments exhibit distinct transcriptional, epigenetic, and functional identities that reflect their developmental origins [96,101,102,103,104,105]. Rather than constituting a homogeneous population, dermal fibroblasts comprise multiple developmentally encoded lineages whose relative abundance and activity change across ontogeny and in response to injury [94]. Understanding these lineage relationships is therefore central to explaining the transition from regenerative to fibrotic wound healing during development.
3.1. Embryonic Origin and Identification Markers of Dermal Fibroblasts Sub-Populations
Fibroblast heterogeneity arises from both embryonic origin and anatomical localization [97,99]. In most body regions, fibroblasts derive from mesodermal progenitors—originating from the lateral plate mesoderm or dermomyotome—whereas craniofacial fibroblasts arise from neural crest–derived ectoderm [100]. During skin development, multipotent mesenchymal progenitors populate the dermis and progressively differentiate into discrete fibroblast subsets under the influence of positional cues and morphogenetic signaling pathways, including Wnt, BMP, FGF, and Shh [101,102]. This maturation process coincides with compartmentalization of the dermis into papillary and reticular layers and with changes in ECM composition, mechanical properties, and appendage-forming capacity [101,103,104].
Many previous studies revealed the presence of multiple functionally distinct subtypes of fibroblasts in developing mouse skin. A combination of markers for the identification of multipotent mesenchymal cells with the ability to differentiate into all types of dermal fibroblasts includes PDGF receptor A (PDGFRA), delta-like homology -1 (DLK-1), EN-1, and leucine-rich repeat protein (LRIG1) [19,105,106]. After establishing a nascent connective tissue, skin progenitors undergo progressive specification into three main distinct groups, while these three groups are further separated into ten subgroups that display different combinations of gene expression patterns [97,100]. Currently, papillary fibroblasts (CD26+/SCA1+) and reticular fibroblasts (DLK1+/SCA1-) are established populations with distinct expression patterns and properties [97].
Although papillary and reticular fibroblasts can now be clearly discriminated from one another, studies reveal that there are transcriptionally overlapped fibroblasts across skin layers [100]. Papillary fibroblasts seem to share a common progenitor with the erector pili muscle of the hair follicle and the dermal papilla fibroblasts that participate in the formation of hair follicle [107]. Moreover, papillary fibroblasts are most likely related to scarless healing. Papillary fibroblasts express fap, CD26, Lrig1, integrin Itga8, and Blimp1. Adipocytes, adipocyte progenitor cells and reticular fibroblasts are derived from common fibroblast progenitors expressing PDGFRA, twist-related protein-2 (Twist-2/Dermo-1), and EN-1 [97]. To sum up, while papillary fibroblasts are associated with fine fibrillar matrix deposition and support of hair follicle morphogenesis, reticular fibroblasts are more strongly linked to dense ECM deposition and fibrotic repair. Importantly, transcriptional overlap between fibroblasts across dermal layers suggests that lineage identity is not strictly defined by location alone but reflects shared developmental trajectories [100].
Recent single-cell analyses have confirmed fibroblast heterogeneity in human fetal and adult skin [99,104,108]. In fetal dermis, multiple fibroblast subtypes with distinct metabolic and biosynthetic profiles coexist, including progenitor-like populations that dominate early gestation and more differentiated fibroblasts resembling adult papillary and reticular populations [104]. These findings indicate that lineage diversification precedes birth and that fibroblast identities associated with fibrotic repair are established during development rather than arising solely in response to injury.
Fibroblast heterogeneity has been recently confirmed in adult human skin as well [109,108,99]. In a study by Morioka et al, it has been shown that at least eight subpopulations of fibroblasts reside within the fetal dermis with specific locations that are correlated to their functions in developing skin. Two subpopulations designated as precursor (HOX5+) and early (PLAT+) were found as the predominant cell types at 7-8 weeks of gestation and exhibited highly active protein biosynthesis. Another subtype with high metabolic activity was related to the growth and development of HFs during embryogenesis. In contrast, the rest of the identified subtypes included vasculature–related fibroblasts, papillary and reticular fibroblasts with localization and gene expression similar to those of adult skin, and a distinct cluster of proliferative fibroblasts only identified in the fetal dermis [104].
3.2. Fibroblast Subpopulations Contribute Differently to Wound Healing
Lineage-tracing and functional studies have demonstrated that fibroblast subpopulations contribute differentially to wound healing outcomes [19,21]. Fibroblasts located in the papillary dermis have been associated with regenerative functions, including fine fibrillar matrix deposition and support of hair follicle neogenesis. In contrast, fibroblasts derived from deeper dermal compartments are more strongly linked to fibrotic matrix production and scar formation reviewed in [96]. These lineage-dependent behaviors are progressively established during development and reinforced after birth by epigenetic mechanisms that limit transcriptional plasticity. Recently, two pairs of embryonic fibroblast lineages have been identified with a significant role in scar formation in dorsal and ventral wounds, respectively. The first pair includes EPFs and ENFs. EPFs have been associated with scar formation in the dorsal wound. The second pair includes Pair-related homeobox 1 (Prrx1) positive (PPFs) and negative fibroblasts (PNFs), with PPFs being responsible for scar formation on the ventral dermis of the mouse [19].
Functional lineage-tracing studies have provided direct evidence that fibroblast subpopulations contribute differentially to wound healing outcomes (Figure 1). In mouse skin, ENFs predominate in early embryonic dermis and exhibit transcriptional programs associated with tissue plasticity, ECM remodeling, and regenerative competence. ENFs contribute to restoration of normal dermal architecture and appendage formation following injury. In contrast, EPFs, which arise later in development and become dominant postnatally, display strong pro-fibrotic behavior, robust ECM deposition, and a high propensity for myofibroblast differentiation, making them principal contributors to scar formation [19,20,95].
Lineage tracing reveals that ENFs and EPFs differ not only in developmental timing but also in spatial distribution, proliferative capacity, and sensitivity to mechanotransduction. In Table 1, the most well-established differences between ENFs and EPFs properties are presented. Notably, postnatal mechanical cues can drive ENF-to-EPF conversion through YAP/TAZ-dependent signaling pathways, indicating that fibroblast lineage identity is stabilized—but not irreversibly fixed—by mechanical and epigenetic inputs [110,111]. Complementary studies have identified analogous lineage-dependent fibroblast populations in ventral skin, including Prrx1-positive fibroblasts that similarly contribute to fibrotic repair [19].
4. Conclusions
Accumulating evidence suggests that the divergence between regenerative and fibrotic skin repair reflects developmentally encoded differences in fibroblast lineage identity that are likely enhanced by ECM composition and mechanical stimuli. Fetal wound healing is characterized by a compliant, collagen type III–rich, growth factor–retentive matrix that supports fibroblast plasticity and regenerative capacity, whereas adult repair occurs within a stiffer, collagen type I–dominant environment that promotes pro-fibrotic fibroblast programs in response to mechanotransduction signaling [113,114,115]. Emerging biomaterial strategies, including heparin-functionalized collagen I/III scaffolds and engineered matrices designed to recapitulate embryonic ECM properties, demonstrate that it is possible to partially reconstitute key biochemical and mechanical cues of the regenerative niche [116,117]. By stabilizing and spatially presenting growth factors, modulating matrix composition, and tuning mechanical stiffness, such platforms can bias fibroblast behavior toward fetal-like, pro-regenerative states. These approaches underscore the concept that scarring is not an inevitable outcome of adult wound healing, but rather the result of lineage-restricted and mechanically reinforced cellular programs. Future regenerative therapies will likely require integrated strategies that combine biomaterial design with targeted modulation of mechanotransduction and epigenetic regulation to reprogram adult fibroblasts and restore regenerative healing capacity. Cell line reprogramming procedures may enable the production of pluripotent human cell lines without the use of an embryo. Specifically, the plasticity of fibroblasts can be exploited to restore ENF-like regenerative behavior [118].
Author Contributions
Conceptualization, A.M.; writing—original draft preparation and editing A.M. and A.N.; writing—review and editing, K.K.; writing, All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
This study did not involve the collection or processing of primary data from humans or animals and was therefore exempt from ethical review by the Committee for Research Ethics (Social Sciences).
Informed Consent Statement
Not applicable.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments). Where GenAI has been used for purposes such as generating text, data, or graphics, or for study design, data collection, analysis, or interpretation of data, please add “During the preparation of this manuscript/study, the author(s) used [tool name, version information] for the purposes of [description of use]. The authors have reviewed and edited the output and take full responsibility for the content of this publication.”
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ECM | extracellular matrix |
| EPFs | Engrailed-1–positive fibroblasts |
| TGF-β | transforming growth factor-β |
| HA | hyaluronic acid |
| FN | fibronectin |
| PDGF | platelet-derived growth factor |
| TNFα | tumor necrosis factor α |
| IL | interleukin |
| VEGF | vascular endothelial growth factor |
| PRP | platelet-rich plasma |
| MHC | major histocompatibility complex |
| GAGs | glycosaminoglycans |
| CS | chondroitin sulfate |
| MMPs | matrix metalloproteinases |
| LH | lysyl hydroxylase |
| PH | prolyl hydroxylase |
| LOX | low lysyl oxidase |
| TGM2 | transglutaminase 2 |
| FGFs | fibroblast growth factors |
| FAK | focal adhesion kinase |
| PI3K | Phosphatidylinositol 3-Kinase |
| MAPK | Mitogen-Activated Protein Kinase |
| YAP | Yes-associated protein |
| TAZ | Transcriptional coactivator with PDZ-binding motif |
| CTGF | connective tissue growth factor |
| SOX | SRY-related high-mobility-group (HMG) box |
| Trps1 | Trichorhinophalangeal Syndrome Type I |
| PDGFRA | PDGF receptor A |
| DLK-1 | delta-like homology -1 |
| LRIG1 | leucine-rich repeat protein |
| Twist-2/Dermo-1 | twist-related protein-2 |
| Prrx1 | Pair-related homeobox 1 |
| PPFs | Pair-related homeobox 1 (Prrx1) positive |
| PNFs | Pair-related homeobox 1 (Prrx1) negative fibroblasts |
References
- Lim, K.-M. Skin Epidermis and Barrier Function. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Michopoulou, A.; Montmasson, M.; Garnier, C.; Lambert, E.; Dayan, G.; Rousselle, P. How Do Epidermal Matrix Metalloproteinases Support Re-Epithelialization during Skin Healing? Matrix Biol. 2020, 94, 1–17. [Google Scholar] [CrossRef]
- Hu, M.S.; Borrelli, M.R.; Hong, W.X.; Malhotra, S.; Cheung, A.T.M.; Ransom, R.C.; Rennert, R.C.; Morrison, S.D.; Lorenz, H.P.; Longaker, M.T. Embryonic Skin Development and Repair. Organogenesis 2018, 14, 46–63. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, M.M.W. Fetal Wound Healing BT - Textbook on Scar Management: State of the Art Management and Emerging Technologies; Téot, L., Mustoe, T.A., Middelkoop, E., Gauglitz, G.G., Eds.; Springer International Publishing: Cham, 2020; pp. 3–9. ISBN 978-3-030-44766-3. [Google Scholar]
- Coolen, N.A.; Schouten, K.C.W.M.; Middelkoop, E.; Ulrich, M.M.W. Comparison between Human Fetal and Adult Skin. Arch. Dermatol. Res. 2010, 302, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Park, S. Building vs. Rebuilding Epidermis: Comparison Embryonic Development and Adult Wound Repair. Front. Cell Dev. Biol. 2022, 9. [Google Scholar] [CrossRef]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in Wound Healing: A Comprehensive Review. Adv. Wound Care 2014. [Google Scholar] [CrossRef]
- Rousselle, P.; Montmasson, M.; Garnier, C. Extracellular Matrix Contribution to Skin Wound Re-Epithelialization. Matrix Biol. 2019, 75–76, 12–26. [Google Scholar] [CrossRef]
- Degen, K.E.; Gourdie, R.G. Embryonic Wound Healing: A Primer for Engineering Novel Therapies for Tissue Repair. Birth Defects Res. C. Embryo Today 2012, 96, 258–270. [Google Scholar] [CrossRef]
- Desmoulière, A.; Geinoz, A.; Gabbiani, F.; Gabbiani, G. Transforming Growth Factor-Beta 1 Induces Alpha-Smooth Muscle Actin Expression in Granulation Tissue Myofibroblasts and in Quiescent and Growing Cultured Fibroblasts. J. Cell Biol. 1993, 122, 103–111. [Google Scholar] [CrossRef]
- Rolfe, K.J.; Richardson, J.; Vigor, C.; Irvine, L.M.; Grobbelaar, A.O.; Linge, C. A Role for TGF-Beta1-Induced Cellular Responses during Wound Healing of the Non-Scarring Early Human Fetus? J. Invest. Dermatol 2007, 127, 2656–2667. [Google Scholar] [CrossRef]
- Lorenz, H.P.; Lin, R.Y.; Longaker, M.T.; Whitby, D.J.; Adzick, N.S. The Fetal Fibroblast: The Effector Cell of Scarless Fetal Skin Repair. Plast. Reconstr. Surg. 1995, 96, 1251. [Google Scholar] [CrossRef]
- Martin, P.; Lewis, J. Actin Cables and Epidermal Movement in Embryonic Wound Healing. Nature 1992, 360, 179–183. [Google Scholar] [CrossRef]
- McCluskey, J.; Martin, P. Analysis of the Tissue Movements of Embryonic Wound Healing—DiI Studies in the Limb Bud Stage Mouse Embryo. Dev. Biol. 1995, 170, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.L.; Marshall, C.D.; Barnes, L.A.; Murphy, M.P.; Ransom, R.C.; Longaker, M.T. Scarless Wound Healing: Transitioning from Fetal Research to Regenerative Healing. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Larson, B.J.; Longaker, M.T.; Lorenz, H.P. Scarless Fetal Wound Healing: A Basic Science Review. Plast. Reconstr. Surg. 2010, 126, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.L.N.P.; Krijnen, P.A.J.; Middelkoop, E.; Niessen, H.W.M.; Boekema, B.K.H.L. Fetal Skin Wound Healing: Key Extracellular Matrix Components and Regulators in Scarless Healing. J. Invest. Dermatol. 2025, 145, 280–302. [Google Scholar] [CrossRef]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct Fibroblast Lineages Determine Dermal Architecture in Skin Development and Repair. Nature 2013. [Google Scholar] [CrossRef]
- Rinkevich, Y.; Walmsley, G.G.; Hu, M.S.; Maan, Z.N.; Newman, A.M.; Drukker, M.; Januszyk, M.; Krampitz, G.W.; Gurtner, G.C.; Lorenz, H.P.; et al. Identification and Isolation of a Dermal Lineage with Intrinsic Fibrogenic Potential. Science (80-. ) 2015. [Google Scholar] [CrossRef]
- Jiang, D.; Correa-Gallegos, D.; Christ, S.; Stefanska, A.; Liu, J.; Ramesh, P.; Rajendran, V.; De Santis, M.M.; Wagner, D.E.; Rinkevich, Y. Two Succeeding Fibroblastic Lineages Drive Dermal Development and the Transition from Regeneration to Scarring. Nat. Cell Biol. 2018. [Google Scholar] [CrossRef]
- Jiang, D.; Rinkevich, Y. Distinct Fibroblasts in Scars and Regeneration. Curr. Opin. Genet. Dev. 2021, 70, 7–14. [Google Scholar] [CrossRef]
- Ye, H.; Yu, Q.; Lee, A.J.K.S.; Dong, W.; Chang, Z.; Correa-Gallegos, D.; Jiang, D.; Dai, R.; Schorpp, K.; Hadian, K.; et al. Distinct Fibroblast Assemblies Establish Scarless Regeneration. Cell Rep. 2025, 45, 116767. [Google Scholar] [CrossRef] [PubMed]
- Lorant, J.; Poinas, A.; Nerriere, O.; Vrignaud, F.; Frenard, C.; Winer, N.; Khammari, A.; Dréno, B. Foetal Skin Cells in Wound Healing: A Promising Tool for Clinical Application. Eur. J. Dermatology 2019, Vol. 29, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Redd, M.J.; Cooper, L.; Wood, W.; Stramer, B.; Martin, P. Wound Healing and Inflammation: Embryos Reveal the Way to Perfect Repair. Philos. Trans. R. Soc. London. Ser. B, Biol. Sci. 2004, 359, 777–784. [Google Scholar] [CrossRef]
- Lo, D.D.; Hu, M.S.; Zimmermann, A.S.; Longaker, M.T.; Peter Lorenz, H. Chapter 51 - Differences in Foetal, Adult Skin and Mucosal Repair; Vishwakarma, A., Sharpe, P., Shi, S., Ramalingam, M.B.T.-S.C.B., Eds.; Academic Press: Boston, 2015; pp. 691–702. ISBN 978-0-12-397157-9. [Google Scholar]
- Beanes, S.R.; Hu, F.-Y.; Soo, C.; Dang, C.M.H.; Urata, M.; Ting, K.; Atkinson, J.B.; Benhaim, P.; Hedrick, M.H.; Lorenz, H.P. Confocal Microscopic Analysis of Scarless Repair in the Fetal Rat: Defining the Transition. Plast. Reconstr. Surg. 2002, 109, 160–170. [Google Scholar] [CrossRef]
- Whitby, D.J.; Ferguson, M.W. The Extracellular Matrix of Lip Wounds in Fetal, Neonatal and Adult Mice. Development 1991, 112, 651–668. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound Repair and Regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Ulrich MMW Fetal Wound Healing; Téot, L.; Mustoe, T.A.; Middelkoop, E.; et al. (Eds.) Textb. Scar Manag. State Art Manag. Emerg. Technol. [Internet]; Springer: Cham, 2020; Chapter 1. [Google Scholar]
- Wang, Z.; Qi, F.; Luo, H.; Xu, G.; Wang, D. Inflammatory Microenvironment of Skin Wounds. In Front. Immunol.; 2022; Volume 13. [Google Scholar]
- Frech, S.; Lichtenberger, B.M. Modulating Embryonic Signaling Pathways Paves the Way for Regeneration in Wound Healing. In Front. Physiol.; 2024; Volume 15. [Google Scholar]
- Clark, R.A.F. Wound Repair Overview and General Considerations. In The Molecular, Cellular Biology of Wound Repair; Clark, R.A.F., Ed.; Plenum Press: New York; - References - Scientific Research Publishing, 1996; pp. 3–35. [Google Scholar]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin Wound Healing: An Update on the Current Knowledge and Concepts. Eur. Surg. Res. Eur. Chir. Forschung. Rech. Chir. Eur. 2017, 58, 81–94. [Google Scholar] [CrossRef]
- Mastrogiannaki, M.; Lichtenberger, B.M.; Reimer, A.; Collins, C.A.; Driskell, R.R.; Watt, F.M. β-Catenin Stabilization in Skin Fibroblasts Causes Fibrotic Lesions by Preventing Adipocyte Differentiation of the Reticular Dermis. J. Invest. Dermatol. 2016. [Google Scholar] [CrossRef]
- Bochaton-Piallat, M.-L.; Gabbiani, G.; Hinz, B. The Myofibroblast in Wound Healing and Fibrosis: Answered and Unanswered Questions. F1000Research 2016, 5. [Google Scholar] [CrossRef]
- Michopoulou, A.; Montmasson, M.; Garnier, C.; Lambert, E.; Dayan, G.; Rousselle, P. A Novel Mechanism in Wound Healing: Laminin 332 Drives MMP9/14 Activity by Recruiting Syndecan-1 and CD44. Matrix Biol. 2020, 94, 1–17. [Google Scholar] [CrossRef]
- Longaker, M.T.; Whitby, D.J.; Ferguson, M.W.; Lorenz, H.P.; Harrison, M.R.; Adzick, N.S. Adult Skin Wounds in the Fetal Environment Heal with Scar Formation. Ann. Surg. 1994, 219, 65–72. [Google Scholar] [CrossRef]
- Monavarian, M.; Kader, S.; Moeinzadeh, S.; Jabbari, E. Regenerative Scar-Free Skin Wound Healing. Tissue Eng. Part B Rev. 2019, 25, 294–311. [Google Scholar] [CrossRef]
- Yates, C.C.; Hebda, P.; Wells, A. Skin Wound Healing and Scarring: Fetal Wounds and Regenerative Restitution. Birth Defects Res. C. Embryo Today 2012, 96, 325–333. [Google Scholar] [CrossRef]
- Scheid, A.; Wenger, R.H.; Schäffer, L.; Camenisch, I.; Distler, O.; Ferenc, A.; Cristina, H.; Ryan, H.E.; Johnson, R.S.; Wagner, K.F.; et al. Physiologically Low Oxygen Concentrations in Fetal Skin Regulate Hypoxia-Inducible Factor 1 and Transforming Growth Factor-Beta3. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2002, 16, 411–413. [Google Scholar] [CrossRef]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth Factors and Cytokines in Wound Healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef]
- Lo, D.D.; Zimmermann, A.S.; Nauta, A.; Longaker, M.T.; Lorenz, H.P. Scarless Fetal Skin Wound Healing Update. Birth Defects Res. Part C Embryo Today Rev. 2012, 96, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.Y.; Chi, J.T.; Dudoit, S.; Bondre, C.; Van De Rijn, M.; Botstein, D.; Brown, P.O. Diversity, Topographic Differentiation, and Positional Memory in Human Fibroblasts. In Proc. Natl. Acad. Sci. U. S. A.; 2002; Volume 99, pp. 12877–12882. [Google Scholar] [CrossRef]
- Singh, D.; Rai, V.; Agrawal, D.K. Regulation of Collagen I and Collagen III in Tissue Injury and Regeneration. Cardiol. Cardiovasc. Med. 2023, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Leeming, D.J.; Karsdal, M.A. Type V Collagen. In Biochem. Collagens, Laminins Elastin Struct. Funct. Biomarkers, Third Ed. ed; 2024; pp. 55–60. [Google Scholar] [CrossRef]
- Smith, L.T.; Holbrook, K.A.; Madri, J.A. Collagen Types I, III, and V in Human Embryonic and Fetal Skin. Am. J. Anat. 1986, 175, 507–521. [Google Scholar] [CrossRef]
- Buchanan, E.P.; Longaker, M.T.; Lorenz, H.P. Chapter 6 Fetal Skin Wound Healing. Adv. Clin. Chem. 2009, 48, 137–161. [Google Scholar] [CrossRef]
- Coolen, N.A.; Schouten, K.C.W.M.; Boekema, B.K.H.L.; Middelkoop, E.; Ulrich, M.M.W. Wound Healing in a Fetal, Adult, and Scar Tissue Model: A Comparative Study. In Wound Repair Regen.; 2010; Volume 18, pp. 291–301. [Google Scholar] [CrossRef]
- Mast, B.A.; Flood, L.C.; Haynes, J.H.; Depalma, R.L.; Cohen, I.K.; Diegelmann, R.F.; Krummel, T.M. Hyaluronic Acid Is a Major Component of the Matrix of Fetal Rabbit Skin and Wounds: Implications for Healing by Regeneration. Matrix 1991, 11, 63–68. [Google Scholar] [CrossRef]
- Kennedy, C.I.; Diegelmann, R.F.; Haynes, J.H.; Yager, D.R. Proinflammatory Cytokines Differentially Regulate Hyaluronan Synthase Isoforms in Fetal and Adult Fibroblasts. J. Pediatr. Surg. 2000, 35, 874–879. [Google Scholar] [CrossRef]
- Colwell, A.S.; Longaker, M.T.; Lorenz, H.P. Fetal Wound Healing. Front. Biosci. 2003, 8. [Google Scholar] [CrossRef]
- Soo, C.; Hu, F.Y.; Zhang, X.; Wang, Y.; Beanes, S.R.; Lorenz, H.P.; Hedrick, M.H.; Mackool, R.J.; Plaas, A.; Kim, S.J.; et al. Differential Expression of Fibromodulin, a Transforming Growth Factor-β Modulator, in Fetal Skin Development and Scarless Repair. Am. J. Pathol. 2000, 157, 423–433. [Google Scholar] [CrossRef]
- Danielson, K.G.; Baribault, H.; Holmes, D.F.; Graham, H.; Kadler, K.E.; Iozzo, R. V. Targeted Disruption of Decorin Leads to Abnormal Collagen Fibril Morphology and Skin Fragility. J. Cell Biol. 1997, 136, 729–743. [Google Scholar] [CrossRef]
- Pang, X.; Dong, N.; Zheng, Z. Small Leucine-Rich Proteoglycans in Skin Wound Healing. Front. Pharmacol. 2020, 10, 501915. [Google Scholar] [CrossRef]
- Bullard, K.M.; Longaker, M.T.; Lorenz, H.P. Fetal Wound Healing: Current Biology. World J. Surg. 2003, 27, 54–61. [Google Scholar] [CrossRef]
- Ramelet, A.A.; Hirt-Burri, N.; Raffoul, W.; Scaletta, C.; Pioletti, D.P.; Offord, E.; Mansourian, R.; Applegate, L.A. Chronic Wound Healing by Fetal Cell Therapy May Be Explained by Differential Gene Profiling Observed in Fetal versus Old Skin Cells. Exp. Gerontol. 2009, 44, 208–218. [Google Scholar] [CrossRef]
- Soo, C.; Beanes, S.R.; Hu, F.Y.; Zhang, X.; Dang, C.; Chang, G.; Wang, Y.; Nishimura, I.; Freymiller, E.; Longaker, M.T.; et al. Ontogenetic Transition in Fetal Wound Transforming Growth Factor-β Regulation Correlates with Collagen Organization. Am. J. Pathol. 2003, 163, 2459–2476. [Google Scholar] [CrossRef] [PubMed]
- Colwell, A.S.; Longaker, M.T.; Lorenz, H.P. Identification of Differentially Regulated Genes in Fetal Wounds during Regenerative Repair. Wound Repair Regen. 2008, 16, 450–459. [Google Scholar] [CrossRef]
- Whitby, D.J.; Ferguson, M.W.J. The Extracellular Matrix of Lip Wounds in Fetal, Neonatal and Adult Mice. Development 1991, 112, 651–668. [Google Scholar] [CrossRef] [PubMed]
- Hirt-Burri, N.; Scaletta, C.; Gerber, S.; Pioletti, D.P.; Applegate, L.A. Wound-Healing Gene Family Expression Differences between Fetal and Foreskin Cells Used for Bioengineered Skin Substitutes. Artif. Organs 2008, 32, 509–518. [Google Scholar] [CrossRef]
- Moulin, V.; Plamondon, M. Differential Expression of Collagen Integrin Receptor on Fetal vs. Adult Skin Fibroblasts: Implication in Wound Contraction during Healing. Br. J. Dermatol. 2002, 147, 886–892. [Google Scholar] [CrossRef] [PubMed]
- Meuli, M.; Lorenz, H.P.; Hedrick, M.H.; Sullivan, K.M.; Harrison, M.R.; Adzick, N.S. Scar Formation in the Fetal Alimentary Tract. J. Pediatr. Surg. 1995, 30, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Longaker, M.T.; Whitby, D.J.; Jennings, R.W.; Duncan, B.W.; Ferguson, M.W.J.; Harrison, M.R.; Adzick, N.S. Fetal Diaphragmatic Wounds Heal with Scar Formation. J. Surg. Res. 1991, 50, 375–385. [Google Scholar] [CrossRef]
- Longaker, M.T.; Whitby, D.J.; Ferguson, M.W.J.; Lorenz, H.P.; Harrison, M.R.; Adzick, N.S. Adult Skin Wounds in the Fetal Environment Heal with Scar Formation. Ann. Surg. 1994, 219, 65–72. [Google Scholar] [CrossRef]
- Armstrong, J.R.; Ferguson, M.W.J. Ontogeny of the Skin and the Transition from Scar-Free to Scarring Phenotype during Wound Healing in the Pouch Young of a Marsupial, Monodelphis Domestica. Dev. Biol. 1995, 169, 242–260. [Google Scholar] [CrossRef]
- Yang, L.; Qui, C.X.; Ludlow, A.; Ferguson, M.W.J.; Brunner, G. Active Transforming Growth Factor-β in Wound Repair: Determination Using a New Assay. Am. J. Pathol. 1999, 154, 105. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M.W.J.; O’Kane, S. Scar-Free Healing: From Embryonic Mechanisms to Adult Therapeutic Intervention. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2004, 359, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Bullard, K.M.; Cass, D.L.; Banda, M.J.; Adzick, N.S. Transforming Growth Factor Beta-1 Decreases Interstitial Collagenase in Healing Human Fetal Skin. J. Pediatr. Surg. 1997, 32, 1023–1027. [Google Scholar] [CrossRef]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef]
- Whitby, D.J.; Ferguson, M.W.J. Immunohistochemical Localization of Growth Factors in Fetal Wound Healing. Dev. Biol. 1991, 147, 207–215. [Google Scholar] [CrossRef]
- Marini, M.; Bertolai, R.; Ambrosini, S.; Sarchielli, E.; Vannelli, G.B.; Sgambati, E. Differential Expression of Vascular Endothelial Growth Factor in Human Fetal Skeletal Site-Specific Tissues: Mandible versus Femur. Acta Histochem. 2015, 117, 228–234. [Google Scholar] [CrossRef]
- Ono, I.; Gunji, H.; Zhang, J.Z.; Maruyama, K.; Kaneko, F. A Study of Cytokines in Burn Blister Fluid Related to Wound Healing. Burns 1995, 21, 352–355. [Google Scholar] [CrossRef]
- Gordon, A.; Kozin, E.D.; Keswani, S.G.; Vaikunth, S.S.; Katz, A.B.; Zoltick, P.W.; Favata, M.; Radu, A.P.; Soslowsky, L.J.; Herlyn, M.; et al. Permissive Environment in Postnatal Wounds Induced by Adenoviral-Mediated Overexpression of the Anti-Inflammatory Cytokine Interleukin-10 Prevents Scar Formation. Wound Repair Regen. 2008, 16, 70–79. [Google Scholar] [CrossRef]
- Liang, H.X.; Yi, C.; He, S.H.; Zhou, L.Y.; Li, F.F.; Tang, J.; Chen, E.X.; Fu, L.J.; Wang, Y.X.; Xie, Y.L.; et al. The Role of FAK Signaling in Early Placental Development and Trophoblast Lineage Specification in Human Pregnancy. Cell. Signal. 2025, 134, 111946. [Google Scholar] [CrossRef] [PubMed]
- Barnes, L.A.; Marshall, C.D.; Leavitt, T.; Hu, M.S.; Moore, A.L.; Gonzalez, J.G.; Longaker, M.T.; Gurtner, G.C. Mechanical Forces in Cutaneous Wound Healing: Emerging Therapies to Minimize Scar Formation. Adv. wound care 2018, 7, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Panciera, T.; Azzolin, L.; Cordenonsi, M.; Piccolo, S. Mechanobiology of YAP and TAZ in Physiology and Disease. Nat. Rev. Mol. Cell Biol. 2017 1812 2017, 18, 758–770. [Google Scholar] [CrossRef]
- Liu, Z.; Li, S.; Qian, X.; Li, L.; Zhang, H.; Liu, Z. RhoA/ROCK-YAP/TAZ Axis Regulates the Fibrotic Activity in Dexamethasone-Treated Human Trabecular Meshwork Cells. Front. Mol. Biosci. 2021, 8, 728932. [Google Scholar] [CrossRef]
- Walko, G.; Woodhouse, S.; Pisco, A.O.; Rognoni, E.; Liakath-Ali, K.; Lichtenberger, B.M.; Mishra, A.; Telerman, S.B.; Viswanathan, P.; Logtenberg, M.; et al. A Genome-Wide Screen Identifies YAP/WBP2 Interplay Conferring Growth Advantage on Human Epidermal Stem Cells. Nat. Commun. 2017 81 2017, 8, 14744. [Google Scholar] [CrossRef] [PubMed]
- Rognoni, E.; Walko, G. The Roles of YAP/TAZ and the Hippo Pathway in Healthy and Diseased Skin. Cells 2019, 8, 411. [Google Scholar] [CrossRef]
- Heng, B.C.; Zhang, X.; Aubel, D.; Bai, Y.; Li, X.; Wei, Y.; Fussenegger, M.; Deng, X. Role of YAP/TAZ in Cell Lineage Fate Determination and Related Signaling Pathways. Front. Cell Dev. Biol. 2020, 8, 559813. [Google Scholar] [CrossRef]
- Lim, J.H.; Kang, H.M.; Jung, C.-R. The Role of RhoA/ROCK Singaling Pathway in Organoid Research. Organoid 2025, 5, e4. [Google Scholar] [CrossRef]
- Whyte, J.L.; Smith, A.A.; Liu, B.; Manzano, W.R.; Evans, N.D.; Dhamdhere, G.R.; Fang, M.Y.; Chang, H.Y.; Oro, A.E.; Helms, J.A. Augmenting Endogenous Wnt Signaling Improves Skin Wound Healing. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Tan, K.K.B.; Salgado, G.; Connolly, J.E.; Chan, J.K.Y.; Lane, E.B. Characterization of Fetal Keratinocytes, Showing Enhanced Stem Cell-like Properties: A Potential Source of Cells for Skin Reconstruction. Stem cell reports 2014, 3, 324–338. [Google Scholar] [CrossRef]
- Gschwandtner, M.; Zhong, S.; Tschachler, A.; Mlitz, V.; Karner, S.; Elbe-Bürger, A.; Mildner, M. Fetal Human Keratinocytes Produce Large Amounts of Antimicrobial Peptides: Involvement of Histone-Methylation Processes. J. Invest. Dermatol. 2014, 134, 2192–2201. [Google Scholar] [CrossRef] [PubMed]
- Shaw, T.; Martin, P. Epigenetic Reprogramming during Wound Healing: Loss of Polycomb-Mediated Silencing May Enable Upregulation of Repair Genes. EMBO Rep. 2009, 10, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Yuen, R.K.; Jiang, R.; Peñaherrera, M.S.; McFadden, D.E.; Robinson, W.P. Genome-Wide Mapping of Imprinted Differentially Methylated Regions by DNA Methylation Profiling of Human Placentas from Triploidies. Epigenetics Chromatin 2011, 4, 10. [Google Scholar] [CrossRef]
- Carre, A.L.; James, A.W.; MacLeod, L.; Kong, W.; Kawai, K.; Longaker, M.T.; Lorenz, H.P. Interaction of Wingless Protein (Wnt), Transforming Growth Factor-Beta1, and Hyaluronan Production in Fetal and Postnatal Fibroblasts. Plast. Reconstr. Surg. 2010, 125, 74–88. [Google Scholar] [CrossRef]
- Hu, M.S.; Januszyk, M.; Hong, W.X.; Walmsley, G.G.; Zielins, E.R.; Atashroo, D.A.; Maan, Z.N.; McArdle, A.; Takanishi, D.M.; Gurtner, G.C.; et al. Gene Expression in Fetal Murine Keratinocytes and Fibroblasts. J. Surg. Res. 2014, 190, 344–357. [Google Scholar] [CrossRef]
- Yang, W.; Liu, L.; Cheng, X. Research Progress on Scar-Free Healing. Ann. Med. 2025, 57, 2575109. [Google Scholar] [CrossRef]
- Wissmüller, S.; Kosian, T.; Wolf, M.; Finzsch, M.; Wegner, M. The High-Mobility-Group Domain of Sox Proteins Interacts with DNA-Binding Domains of Many Transcription Factors. Nucleic Acids Res. 2006, 34, 1735–1744. [Google Scholar] [CrossRef]
- Miao, Q.; Hill, M.C.; Chen, F.; Mo, Q.; Ku, A.T.; Ramos, C.; Sock, E.; Lefebvre, V.; Nguyen, H. SOX11 and SOX4 Drive the Reactivation of an Embryonic Gene Program during Murine Wound Repair. Nat. Commun. 2019, 10, 4042. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.W.; LeGrand, C.F.; Kinnear, B.F.; Denil, S.L.I.J.; Dye, D.E.; Benny, P.; Chan, J.K.Y.; Lane, E.B.; Coombe, D.R. Keratinocyte Self-Renewal and Differentiation Is Dictated by Extrinsic Signals from Dermal Extracellular Matrices. Open Biol. 2025, 15, 240417. [Google Scholar] [CrossRef]
- Hu, M.S.; Hong, W.X.; Januszyk, M.; Walmsley, G.G.; Luan, A.; Maan, Z.N.; Moshrefi, S.; Tevlin, R.; Wan, D.C.; Gurtner, G.C.; et al. Pathway Analysis of Gene Expression in Murine Fetal and Adult Wounds. Adv. wound care 2018, 7, 262–275. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.; Marçal, H.; Foster, L.J.R. Towards Scarless Wound Healing: A Comparison of Protein Expression between Human, Adult and Foetal Fibroblasts. Biomed Res. Int. 2014, 2014, 676493. [Google Scholar] [CrossRef]
- Hao, A.; Dong, X.; Gou, Y.; Li, A.; Li, J.; Xiang, H.; Rahaman, S.; Zhu, Y.; Zhang, H.; You, W.; et al. Engrailed-1 Inactivation Leads to Scarless Skin Wound Healing through Extracellular Matrix Remodeling. Genes Dis. 2025, 12, 101484. [Google Scholar] [CrossRef]
- Korosec, A.; Frech, S.; Gesslbauer, B.; Vierhapper, M.; Radtke, C.; Petzelbauer, P.; Lichtenberger, B.M. Lineage Identity and Location within the Dermis Determine the Function of Papillary and Reticular Fibroblasts in Human Skin. J. Invest. Dermatol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.B.; Valencia, C.; Akras, D.; DiIorio, S.E.; Griffin, M.F.; Longaker, M.T.; Wan, D.C. Understanding Fibroblast Heterogeneity in Form and Function. Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, M.; Ohn, J.; Seong, R.H.; Chung, J.H.; Kim, K.H.; Jo, S.J.; Kwon, O. Twist2-Driven Chromatin Remodeling Governs the Postnatal Maturation of Dermal Fibroblasts. Cell Rep. 2022, 39, 110821. [Google Scholar] [CrossRef]
- Gary, R.; Peter, V.; James, F.; P.E.F., M.; Emily, S.; Issac, G.; B.R., A.; Ni, H.; Bayanne, O.; Anna, D.; et al. Developmental Cell Programs Are Co-Opted in Inflammatory Skin Disease. Science (80-. ). 2021, 371, eaba6500. [Google Scholar] [CrossRef]
- Plikus, M. V; Wang, X.; Sinha, S.; Forte, E.; Thompson, S.M.; Herzog, E.L.; Driskell, R.R.; Rosenthal, N.; Biernaskie, J.; Horsley, V. Fibroblasts: Origins, Definitions, and Functions in Health and Disease. Cell 2021, 184, 3852–3872. [Google Scholar] [CrossRef]
- Thulabandu, V.; Chen, D.; Atit, R.P. Dermal Fibroblast in Cutaneous Development and Healing. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7. [Google Scholar] [CrossRef]
- Myung, P.; Andl, T.; Atit, R. The Origins of Skin Diversity: Lessons from Dermal Fibroblasts. Development 2022, 149, dev200298. [Google Scholar] [CrossRef]
- Rognoni, E.; Pisco, A.O.; Hiratsuka, T.; Sipilä, K.H.; Belmonte, J.M.; Mobasseri, S.A.; Philippeos, C.; Dilão, R.; Watt, F.M. Fibroblast State Switching Orchestrates Dermal Maturation and Wound Healing. Mol. Syst. Biol. 2018, 14, e8174. [Google Scholar] [CrossRef]
- Morioka, N.; Ganier, C.; Watt, F.M. Fetal Fibroblast Heterogeneity Defines Dermal Architecture during Human Embryonic Skin Development. J. Invest. Dermatol. 2025, 145, 1081–1091.e7. [Google Scholar] [CrossRef]
- Driskell, R.R.; Watt, F.M. Understanding Fibroblast Heterogeneity in the Skin. Trends Cell Biol. 2015, 25, 92–99. [Google Scholar] [CrossRef]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct Fibroblast Lineages Determine Dermal Architecture in Skin Development and Repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct Fibroblast Lineages Determine Dermal Architecture in Skin Development and Repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Philippeos, C.; Telerman, S.B.; Oulès, B.; Pisco, A.O.; Shaw, T.J.; Elgueta, R.; Lombardi, G.; Driskell, R.R.; Soldin, M.; Lynch, M.D.; et al. Spatial and Single-Cell Transcriptional Profiling Identifies Functionally Distinct Human Dermal Fibroblast Subpopulations. J. Invest. Dermatol. 2018, 138, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Ganier, C.; Rognoni, E.; Goss, G.; Lynch, M.; Watt, F.M. Fibroblast Heterogeneity in Healthy and Wounded Skin. Cold Spring Harb. Perspect. Biol. 2022, 14, a041238. [Google Scholar] [CrossRef]
- Mascharak, S.; Griffin, M.; Talbott, H.E.; Guo, J.L.; Parker, J.; Morgan, A.G.; Valencia, C.; Kuhnert, M.M.; Li, D.J.; Liang, N.E.; et al. Inhibiting Mechanotransduction Prevents Scarring and Yields Regeneration in a Large Animal Model. Sci. Transl. Med. 2026, 17, eadt6387. [Google Scholar] [CrossRef]
- Györfi, A.-H.; Matei, A.-E.; Fuchs, M.; Liang, C.; Rigau, A.R.; Hong, X.; Zhu, H.; Luber, M.; Bergmann, C.; Dees, C.; et al. Engrailed 1 Coordinates Cytoskeletal Reorganization to Induce Myofibroblast Differentiation. J. Exp. Med. 2021, 218, e20201916. [Google Scholar] [CrossRef]
- Jiang, D.; Rinkevich, Y. Defining Skin Fibroblastic Cell Types beyond CD90. Front. Cell Dev. Biol. 2018, 6, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Des Parkin, J.; San Antonio, J.D.; Persikov, A. V.; Dagher, H.; Dalgleish, R.; Jensen, S.T.; Jeunemaitre, X.; Savige, J. The Collαgen III Fibril Has a “Flexi-Rod” Structure of Flexible Sequences Interspersed with Rigid Bioactive Domains Including Two with Hemostatic Roles. PLoS One 2017, 12. [Google Scholar] [CrossRef]
- Cuttle, L.; Nataatmadja, M.; Fraser, J.F.; Kempf, M.; Kimble, R.M.; Hayes, M.T. Collagen in the Scarless Fetal Skin Wound: Detection with Picrosirius-Polarization. Wound Repair Regen. 2005, 13, 198–204. [Google Scholar] [CrossRef]
- Sriram, G.; Bigliardi, P.L.; Bigliardi-Qi, M. Fibroblast Heterogeneity and Its Implications for Engineering Organotypic Skin Models in Vitro. Eur. J. Cell Biol. 2015. [Google Scholar] [CrossRef]
- Wang, F.; Wang, M.; She, Z.; Fan, K.; Xu, C.; Chu, B.; Chen, C.; Shi, S.; Tan, R. Collagen/Chitosan Based Two-Compartment and Bi-Functional Dermal Scaffolds for Skin Regeneration. Mater. Sci. Eng. C 2015, 52, 155–162. [Google Scholar] [CrossRef]
- Michopoulou, A.; Koliakou, E.; Terzopoulou, Z.; Rousselle, P.; Palamidi, A.; Doxakis, A.; Konstantinidou, P.; Roig-Rosello, E.; Demiri, E.; Bikiaris, D. Benefit of Coupling Heparin to Crosslinked Collagen I/III Scaffolds for Human Dermal Fibroblast Subpopulations’ Tissue Growth No Title. J. Biomed. Mater. Res. Part A 2022, 110. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.-M. Skin Epidermis and Barrier Function. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
Figure 1.
Activation of Engrailed-1 in response to mechanical tension in adult wounds. Mechanical tension applied to wounds leads to suppression of Trps 1 and induces transcriptional programs related to scarring outcomes. Wnt activation in the dermis is related to regeneration and hair follicle morphogenesis. Green fibroblasts: regenerative / ENF-like / papillary-associated, purple fibroblasts: fibrotic/EPF-like / reticular-associated.
Figure 1.
Activation of Engrailed-1 in response to mechanical tension in adult wounds. Mechanical tension applied to wounds leads to suppression of Trps 1 and induces transcriptional programs related to scarring outcomes. Wnt activation in the dermis is related to regeneration and hair follicle morphogenesis. Green fibroblasts: regenerative / ENF-like / papillary-associated, purple fibroblasts: fibrotic/EPF-like / reticular-associated.
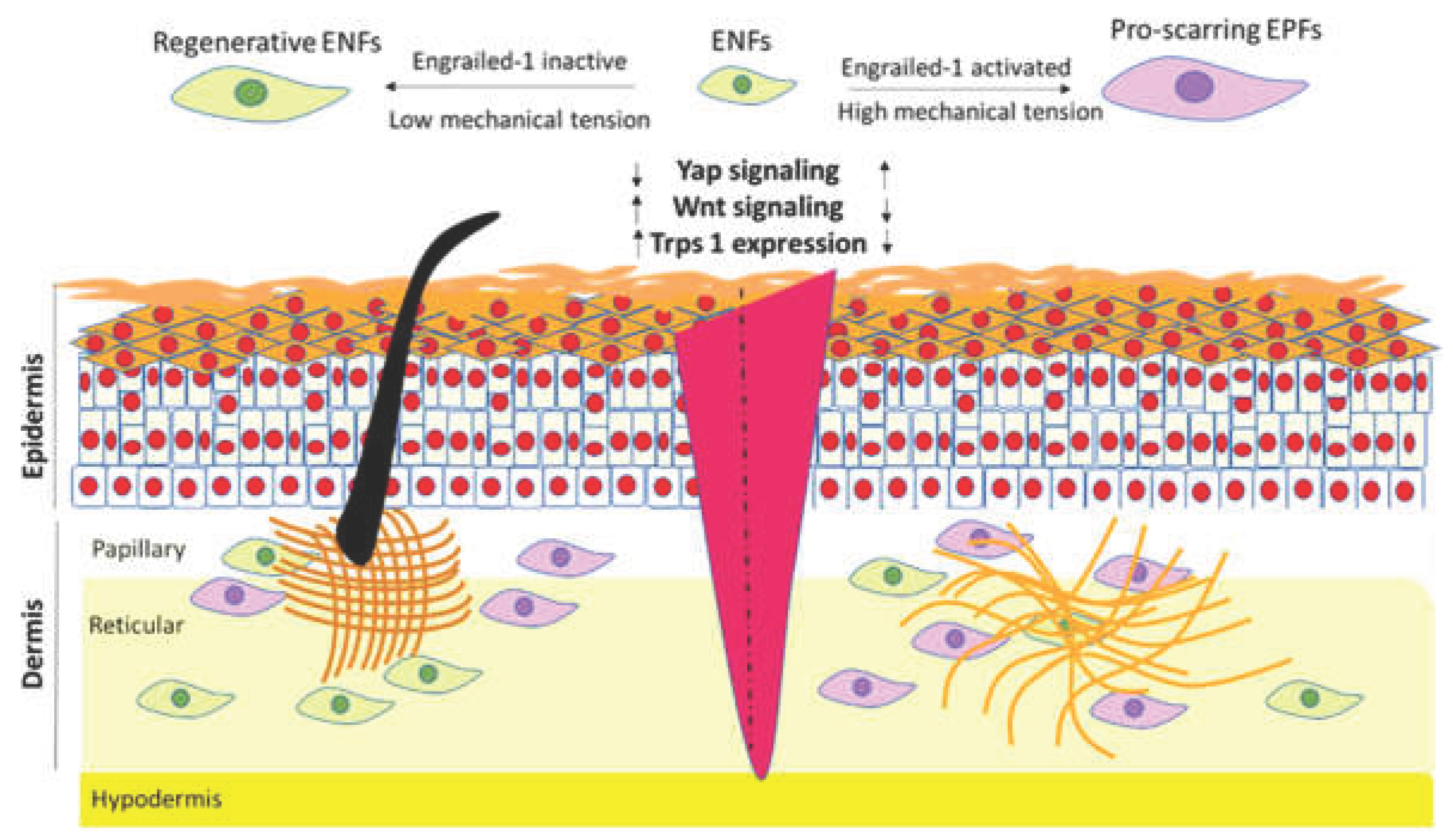

Table 1.
The most well-established differences between ENFs and EPFs properties.
| Properties | ENFs (En1– Lineage-Negative Fibroblasts) | EPFs (En1+ Lineage-Positive Fibroblasts) |
|---|---|---|
| Developmental Timing | Early embryonic dermis; predominant before mid-gestation [19,20] | Emerge later in development; dominant postnatally [19,20] |
| Functional Role in Wounds | Regenerative; support restoration of dermal architecture and appendages [19,21,110] | Pro-fibrotic; major contributors to scar ECM and myofibroblast formation [19,110,111,112] |
| Transcriptional Signatures | Developmental genes, ECM remodeling, and reduced contractile gene expression [19] | High collagen I expression, cross-linking enzymes, contractile machinery, myofibroblast markers [19] |
| Mechanotransduction Sensitivity | Lower YAP/TAZ activation; tolerant of soft/low-tension environments [110] | High YAP/TAZ activation; responsive to stiffness and tension; mechanosensitive profibrotic signaling [110] |
| Typical ECM deposition | Produce loose, fetal-like matrix rich in HA and collagen III [17] | Produce dense, aligned, collagen-I–rich ECM with greater cross-linking [19] |
| Spatial Localization (Adult) | Reduced pool; more papillary-like transcriptional identity [31] | Expanded pool; more reticular-like / deep dermal identity [31] |
| Plasticity / Reprogrammability | High plasticity, fetal-like | Relatively fixed, stabilized by epigenetic and mechanical cues [110] |
| Response to Injury | Promote remodeling and reconstitution [19,20,95] | Drive fibrosis, contraction, and scar deposition [19,20,95] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



