Submitted:
02 February 2026
Posted:
05 February 2026
Read the latest preprint version here
Abstract
Central nervous system disorders drive disability, yet many neuroactive candidates fail because the brain is a hard compartment to dose. Plant derived molecules spanning polyphenols, alkaloids, terpenoids, and cannabinoids are attractive because their pleiotropic actions can engage oxidative stress, neuroinflammation, and circuit dysfunction. In practice, the blood-brain barrier (BBB) restricts most native phytochemicals through tight-junction selectivity, rapid metabolism, low solubility, and transporter-mediated efflux. Key gaps include poor standardization of exposure metrics, limited human relevant BBB models, and few head-to-head studies that compare delivery platforms on the same payload and outcome. This review tackles the mismatch between mechanistic promise and reliable brain exposure that stalls translation. The objectives are to link phytochemical liabilities to enabling strategies in nanomedicine, alternative routes, and transporter-targeted prodrugs, and to propose decision-grade endpoints for translation. We synthesize evidence on BBB transport logic, nanocarrier families, targeting ligands, intranasal delivery, focused ultrasound mediated opening, and prodrug approaches that hijack influx transporters, while foregrounding safety and chemistry, manufacturing, and controls (CMC) constraints. Here we highlight that effective neurotherapeutics emerge when chemistry, carrier, route, and measurement are co designed rather than optimized in isolation. This framework can guide platform selection, de-risk first in-human studies, and sharpen trial endpoints. More broadly, it offers a transferable playbook for barrier-limited drug development across neurology, psychiatry, and oncology.
Keywords:
central nervous system diseases (CNS)
; blood-brain barrier (BBB)
; drug delivery systems
; nanomedicine
; phytochemicals
; nanoparticles
; prodrugs
; drug administration
; intranasal
; ultrasonic therapy
; transferrin receptor
1. Introduction
1.1. Clinical Burden and Therapeutic Gap
Central nervous system (CNS) disorders such as depression, dementia, and chronic pain remain among the leading causes of global morbidity, disability, and economic burden [1,2]. Despite decades of intensive research, therapeutic outcomes remain unsatisfactory, with high relapse rates in major depressive disorder, limited disease-modifying options for dementia, and inadequate pain control across populations [2,3]. These shortcomings are amplified by the fact that most CNS-active drugs show poor penetration across the blood–brain barrier (BBB), resulting in suboptimal central exposure and attenuated efficacy [1,4]. Even when new compounds demonstrate preclinical promise, attrition rates during clinical translation remain staggering, with failure rates in neuropsychiatric drug development exceeding those in nearly all other therapeutic domains [5,6]. The net result is a widening therapeutic gap that leaves millions of patients reliant on outdated, partially effective, or poorly tolerated interventions [2,5,6,7].
This persistent impasse has renewed attention toward alternative sources of therapeutic innovation. Plant-derived molecules, particularly those rooted in neuroactive amino acid metabolism such as tryptophan, offer a compelling avenue [8,9,10,11]. These compounds are celebrated for their structural diversity, multitarget activity, and evolutionary compatibility with human physiology, making them attractive candidates for modulating complex CNS pathologies [8,10,12]. Yet, enthusiasm is tempered by major barriers. Many phytochemicals exhibit low bioavailability, poor stability, and unpredictable BBB permeability, which compromise their therapeutic impact [8,12,13,14]. Recent advances in nanotechnology, ranging from functionalized nanoparticles to receptor-assisted carriers, seek to overcome these pharmacokinetic and delivery hurdles, but their clinical translation is still in its infancy [8,12,13,14]. Against this backdrop, revisiting plant-derived tryptophan and its metabolic derivatives provides a unique opportunity to bridge neurobiology and psychiatry, while also testing the integration of phytochemistry with advanced delivery platforms to transform depression management [8,9,10,13,15].
1.2. Blood-Brain Barrier (BBB) as a Bottleneck for Phytochemicals
The BBB stands as the central checkpoint governing molecular access to the brain, designed to maintain homeostasis while excluding xenobiotics and potential toxins [12,16]. Its architecture is highly specialized: endothelial cells form continuous tight junctions that restrict paracellular flux, while efflux transporters such as P-glycoprotein (P-gp), breast cancer resistance protein, and multidrug resistance-associated proteins actively pump out diverse substrates [16]. This dual protection ensures neural integrity but also severely limits drug delivery [17]. For therapeutic compounds to penetrate effectively, they must navigate a gauntlet of physicochemical constraints. Molecules with poor aqueous solubility, inappropriate lipophilicity, or rapid metabolic breakdown are particularly disadvantaged, leading to negligible CNS exposure despite robust systemic availability [17,18].
Phytochemicals exemplify this paradox. Polyphenols such as resveratrol and curcumin exhibit potent antioxidant and neuroprotective activities in vitro, yet their hydrophilicity and metabolic instability limit their brain levels to trace amounts [12,14,19,20,21,22,23,24] Alkaloids, though often more lipophilic, encounter substantial efflux clearance, which nullifies their apparent permeability advantage [12]. Terpenoids and cannabinoids, despite their lipophilic structures, which favor passive diffusion, are hindered by rapid first-pass metabolism and limited bioavailability, yielding inconsistent central effects [14,19,25]. Studies consistently report that only a minority of native phytochemicals achieve detectable brain penetration, and even fewer reach concentrations required for therapeutic modulation of neurotransmission or neuroinflammation [12,26,27]. The recurring outcome is a stark disconnect between preclinical promise and clinical translation [19,28,29]. These limitations underscore the need for innovative delivery strategies that can re-engineer phytochemicals to evade efflux, improve stability, and optimize solubility [12,14,25,28]. Without such advances, the native molecular forms of these plant-derived agents remain ill-suited for reliable CNS targeting and fall short of their therapeutic potential in depression and related disorders [30,31].
1.3. Scope and Organizing Framework
This review is not a catalog of every BBB nanotechnology reported to date. Instead, we use an organizing framework that starts with payload liabilities, such as poor stability, rapid clearance, limited permeability, or off-target exposure, and then maps these constraints onto enabling strategies, from ligand-targeted carriers and intranasal systems to transporter-leveraging prodrugs and selected physical modulation [32,33,34]. We then judge platforms by pharmacological endpoints that matter for CNS translation: quantifiable brain exposure, target engagement, and a safety margin compatible with real world dosing [32,35,36,37].
Accordingly, we largely exclude systemic nanomedicine programs without explicit CNS intent, purely diagnostic nanomaterials, and highly speculative constructs lacking a plausible CMC and regulatory path. With that scope set, the next step is to ground these choices in the biological rules of the barrier itself, because delivery design only works when it respects architecture, transport routes, and disease-driven heterogeneity.
2. The Blood–Brain Barrier (BBB): Architecture, Transport, Heterogeneity
2.1. Neurovascular Unit Architecture
The neurovascular unit (NVU) forms the structural and functional foundation of the BBB, integrating multiple cellular and extracellular components into a finely tuned system that maintains CNS homeostasis [38,39,40]. Far from being a passive wall, the NVU is a dynamic interface whose architecture underpins both the protective selectivity of the barrier and its vulnerability in disease states [39,41,42].
At the core of this architecture are brain microvascular endothelial cells, which adopt a highly specialized phenotype distinct from systemic endothelia [39,43]. They exhibit extremely low rates of transcytosis and lack fenestrations, thereby minimizing nonspecific permeability [43]. Their intercellular contacts are enriched with tight junction proteins such as claudins, occludin, and ZO-1, creating an electrically resistant barrier that restricts paracellular diffusion while still permitting finely regulated transport of essential metabolites [44,45,46]. Adherens and gap junctions provide additional stability and communication, allowing endothelial cells to operate as a synchronized sheet rather than as isolated units [45,47].
Closely apposed to the endothelial layer, pericytes are embedded within the basement membrane and act as guardians of barrier integrity [42,48]. They regulate angiogenesis, modulate permeability, and secrete trophic factors such as angiopoietin I and vitronectin, which sustain endothelial survival and limit inflammatory activation [49]. Astrocytic endfeet ensheath nearly the entire capillary surface, releasing mediators, including vascular endothelial growth factor (VEGF) and glial-derived neurotrophic factor (GDNF), while their aquaporin-4 channels orchestrate water and ion balance, which are crucial for neuronal signaling [38,43,50,51].
The basement membrane itself, composed of extracellular matrix proteins secreted by both endothelial cells and astrocytes, provides not only structural stability but also biochemical cues that regulate cellular behavior and cross-talk within the NVU [39,52]. Central to barrier impermeability are tight and adherens junctions, which act as molecular rivets sealing adjacent endothelial cells [44,45]. These complexes are remarkably plastic, responding to oxidative stress, inflammation, and neurodegenerative insults by loosening or disassembling, thereby amplifying barrier leakiness [44,46,48]. Altogether, the NVU’s intricate cellular and extracellular architecture forms the scaffold upon which selective transport processes are built, ensuring both the protection and the metabolic supply of the CNS [38,39,40].
2.2. Transport Pathways and Efflux
The BBB operates under the constant tension of permitting the entry of essential nutrients while simultaneously excluding xenobiotics and potentially harmful agents. This balancing act defines its role as both protector and barrier, a duality that complicates CNS drug development [4,53,54]. Only a limited fraction of compounds traverse the BBB by passive diffusion, largely restricted to small, lipophilic molecules with low molecular weight [4,53,55]. Even for lipophilic drugs, passage is often curtailed by additional regulatory mechanisms that actively limit nonspecific entry [53,56].
To sustain brain metabolism, the BBB relies heavily on carrier-mediated transport [4,55]. Prominent examples include GLUT1 for glucose, LAT1 for large neutral amino acids, and monocarboxylate transporters (MCTs) for lactate and other energy substrates [56,57]. These carriers not only ensure nutrient delivery but also provide entry routes for select phytochemicals, though their activity is sensitive to pathological states and drug interactions [56,57]. For larger molecules, receptor-mediated transcytosis represents a critical pathway, with transferrin and insulin receptors serving as canonical examples [58,59]. Advances in nanomedicine are increasingly focused on exploiting these receptors to deliver therapeutic payloads across the barrier in a controlled manner [53,54,60]. Opposing these influx mechanisms are efflux pumps, the most formidable being P-gp, breast cancer resistance protein (BCRP), and multidrug resistance-associated proteins (MRPs) [53,56]. These ATP-binding cassette (ABC) transporters expel a vast array of xenobiotics and pharmacological compounds, shaping drug distribution within the brain [53,56]. Their expression is dynamic, influenced by neuronal activity, circadian rhythms, aging, and disease states, while interactions with polyphenols and phytochemicals may either inhibit or stimulate efflux, thereby altering drug bioavailability [4]. Ultimately, BBB transport functions as both a sentinel and a bottleneck, safeguarding the CNS but at the same time restricting the therapeutic reach of many promising neuroactive agents [53,54,61] (Table 1, Figure 1).
2.3. Disease- and Age-Driven Heterogeneity
The BBB is not a uniform structure but displays striking regional heterogeneity that shapes vulnerability and therapeutic access [73,74,75]. The hippocampus, for example, exhibits earlier and more pronounced permeability changes compared to the cortex, while the choroid plexus contains fenestrated vasculature that facilitates selective exchange with cerebrospinal fluid [73,75,76,77,78]. Such regional differences are further accentuated by pathological states: aging, neurodegeneration, and systemic inflammation remodel barrier integrity, producing spatially distinct patterns of leakage and dysfunction [75,79,80,81,82]. These dynamic alterations contribute to selective regional susceptibility in disorders such as Alzheimer’s and Parkinson’s disease [75,79,81,83]. Recognizing and integrating BBB heterogeneity is therefore critical for the rational design of nanomedicine and targeted neurotherapeutics [4,53,84,85,86].
Ultimately, BBB transport functions as both a sentinel and a bottleneck, safeguarding the CNS but at the same time restricting the therapeutic reach of many promising neuroactive agents [4,53,84,85,86]. Building on this transport landscape, the subsequent section turns to phytochemicals, profiling their distinct liabilities and highlighting how these molecular features dictate both formulation choices and prodrug design in the pursuit of effective neurotherapeutics [13,14,87,88].
3. Phytochemicals as Neurotherapeutics: Classes, Liabilities, MOAs
3.1. Polyphenols (Resveratrol, Quercetin, Curcumin)
Polyphenols, particularly resveratrol, quercetin, and curcumin, represent the most extensively studied class of neuroprotective phytochemicals, attracting attention due to their pleiotropic activities and broad preclinical support in models of neurodegenerative disease [89,90,91,92,93,94,95]. Their neurotherapeutic potential rests on a complex repertoire of mechanisms that extend beyond simple radical scavenging [92,93,95,96]. Resveratrol activates SIRT1 signaling, promoting mitochondrial biogenesis and synaptic resilience, while quercetin modulates AMPK and Nrf2 pathways to counter oxidative stress and restore redox balance [94,96,97,98]. Curcumin has been shown to suppress NF-κB and Toll-like receptor signaling, thereby dampening neuroinflammatory cascades and protecting neuronal networks [30,65,93,96]. Collectively, these pathways converge to enhance neuronal survival, preserve cognitive function, and mitigate disease-associated cellular stress [89,93,94,95].
Despite these appealing mechanisms, polyphenols suffer from profound pharmacokinetic limitations [99,100,101,102]. All three undergo extensive first-pass metabolism through uridine 5′-diphospho-glucuronosyltransferase (UGT) and sulfotransferase (SULT) pathways, producing conjugated metabolites with limited biological activity [92,101,102,103]. The predominance of glucuronides and sulfates in circulation sharply reduces the availability of free aglycones that are more pharmacologically active [92,99,101,102,103]. Moreover, oral bioavailability is poor, with plasma concentrations of parent compounds often remaining below therapeutic thresholds even at high dietary intake [99,100,101,104]. Such metabolic liabilities have fueled a parallel interest in prodrug approaches and nanoformulations aimed at preserving active moieties for CNS delivery [93,95,100,105].
Even when absorbed, polyphenol penetration into the brain is further constrained by efflux transporters at the BBB [94,95]. P-gp and breast cancer resistance protein (BCRP) actively restrict their accumulation in brain parenchyma, while a “permeability paradox” emerges from the discrepancy between promising in vitro BBB transport studies and the much lower exposures seen in vivo [32,94,95,106]. This discordance reflects not only transporter activity but also systemic metabolism and protein binding, which together limit CNS bioavailability [95,99,100,101,102]. Nevertheless, polyphenols continue to serve as reference scaffolds in neurotherapeutic research, inspiring innovative strategies to overcome BBB constraints while maintaining their broad pharmacodynamic advantages [93,94,95,100,105].
3.2. Alkaloids (Berberine and Galantamine)
Alkaloids represent a chemically diverse class of CNS–active molecules with deep roots in both ethnopharmacology and modern clinical medicine [107,108]. Among them, berberine has emerged as a compelling yet pharmacokinetically problematic candidate [109,110]. It interacts strongly with organic cation transporters and is a recognized substrate of P-gp, factors that severely limit its absorption and systemic distribution [109,110,111]. Berberine undergoes rapid first-pass metabolism, exhibits low oral bioavailability, and suffers from pronounced metabolic instability, resulting in extremely poor CNS exposure despite promising neuroprotective and anti-inflammatory effects demonstrated in cellular and animal models [109,110,112]. These challenges have spurred interest in nanoparticle formulations and prodrug strategies designed to bypass efflux transport and enhance brain uptake [112,113,114,115,116,117].
By contrast, galantamine provides an example of a plant-derived alkaloid that has successfully transitioned into clinical practice as an approved therapy for Alzheimer’s disease [107,108,118,119]. Acting as a selective acetylcholinesterase inhibitor, it improves cholinergic transmission and demonstrates measurable cognitive benefits [108,118,119]. Interestingly, its central activity is not strictly proportional to plasma exposure, as galantamine crosses the BBB primarily through passive diffusion with potential contributions from carrier-mediated processes [65,120,121,122]. This selective permeability enables therapeutic CNS engagement even at moderate systemic concentrations, underscoring the importance of pharmacodynamic targeting in addition to pharmacokinetics [65,120,121].
3.3. Terpenoids and Cannabinoids (CBD/THC, Ginkgolides)
Lipophilic terpenoids and cannabinoids such as cannabidiol (CBD), Δ9-tetrahydrocannabinol (THC), and ginkgolides display paradoxical behavior at the BBB, where their high hydrophobicity does not consistently translate into effective CNS delivery [123,124,125]. CBD and THC are both highly lipophilic molecules, yet their brain penetration is actively curtailed by efflux pumps including P-gp and BCRP, which lower their effective concentrations in neural tissue [125,126,127]. Despite these restrictions, clinical and preclinical evidence demonstrates robust antiepileptic, anxiolytic, and analgesic activity, leading to regulatory approval of CBD for severe childhood epilepsies and THC formulations for spasticity and pain management in multiple sclerosis [128,129,130,131]. Their mechanisms are diverse, encompassing CB1 and CB2 receptor modulation, serotonergic signaling through 5-HT1A receptors, and anti-inflammatory as well as antioxidant actions, though their oral bioavailability remains low and interindividual variability in CNS exposure is considerable [127,132,133,134].
Ginkgolides, diterpenoid lactones derived from Ginkgo biloba, present a different profile, achieving moderate penetration into the CNS. Their primary mechanism of action involves antagonism of platelet-activating factor, a pathway linked to neuroinflammation and ischemic injury [135,136]. Preclinical findings suggest neuroprotective and anti-inflammatory potential, yet clinical trials have yielded mixed results, with benefits often modest and outcomes limited by poor BBB permeability and variable bioavailability [135,136]. The discrepancy between mechanistic promise and inconsistent clinical performance reflects the difficulty of translating terpenoid pharmacology into effective CNS therapeutics.
Together, cannabinoids and ginkgolides illustrate the so-called lipophilicity trap, in which excessive hydrophobicity, combined with efflux and metabolic instability, can paradoxically hinder brain delivery rather than facilitate it [123,124,125,126]. This paradox underscores the need for nuanced drug design and advanced delivery systems when considering terpenoids as neurotherapeutic candidates [133,134,137].
3.4. Formulation-Relevant Liabilities and Structure–Activity Relationship (SAR) Flags
Beyond class-specific mechanisms, the physicochemical properties of phytochemicals largely dictate their ability to cross the BBB and achieve therapeutic relevance [65,138,139]. Optimal penetration is typically observed in compounds with a logD between 1 and 3, balanced pK_a values that minimize ionization at physiological pH, a hydrogen bond donor count of two or fewer, and a hydrogen bond acceptor count not exceeding five [138,140,141]. Molecular flexibility is equally important, with fewer than ten rotatable bonds generally favoring permeability and sustained CNS exposure [138,142]. These criteria extend Lipinski’s Rule of Five into the realm of CNS drug-likeness and provide practical benchmarks for evaluating natural products [138,140,141].
Conversely, several red flags frequently emerge among phytochemicals. Excessive polarity or a topological polar surface area greater than 90 Å2 strongly predicts poor CNS penetration [138,140,142]. Similarly, a high density of hydrogen bond donors, often in the form of phenolic hydroxyl groups, correlates with both poor permeability and metabolic vulnerability through glucuronidation or sulfation [65,138]. These metabolic soft spots, common in polyphenols and terpenoids, reduce bioavailability and amplify efflux transporter recognition [143,144,145].
Early recognition of these liabilities through structure–activity relationship analysis and computational screening is therefore essential [138,146,147]. Such insights can guide the rational design of prodrugs and nanoformulations, improving bioavailability and transforming suboptimal scaffolds into viable neurotherapeutic candidates [4,139,148] (Table 2).
4. Nanomedicine Platforms for Blood-Brain Barrier (BBB) Delivery
4.1. Polymeric Nanoparticles (PLGA, PEG-PLGA, and Chitosan)
Polymeric nanoparticles have emerged as highly versatile carriers for CNS delivery, with poly(lactic-co-glycolic acid) (PLGA) considered the gold standard due to its biocompatibility, biodegradability, and track record of clinical translation [156,157,158]. The incorporation of polyethylene glycol (PEG) into PLGA scaffolds provides “stealth” properties by shielding the carrier from opsonization and prolonging its circulation half-life, thereby substantially enhancing the probability of crossing the BBB [159,160,161]. Beyond their pharmacokinetic advantages, PLGA and PEG-PLGA matrices are particularly well suited for encapsulating hydrophobic phytocompounds, thereby improving aqueous solubility and enabling sustained release profiles that minimize burst effects while maintaining therapeutic concentrations within neural tissue [157,160,162]. Several studies have highlighted that surface modifications, ranging from peptide ligands such as Angiopep-2 to functional protein corona interactions, can further refine nanoparticle selectivity for BBB transport and neuronal uptake [33,163,164,165].
Chitosan-based systems represent a complementary and increasingly significant strategy, exploiting their intrinsic cationic nature and mucoadhesive capacity [159,166]. When used either as a surface coating or as a hybrid scaffold with PLGA, chitosan enables tight interaction with mucosal surfaces and facilitates paracellular transport [159,166,167]. This property is particularly advantageous for intranasal administration, as demonstrated by formulations where PLGA nanoparticles embedded within chitosan microparticles achieved enhanced uptake across the olfactory mucosa and direct delivery to the brain [159,167,168] Intranasal chitosan–PLGA carriers have been successfully applied to deliver repurposed chemotherapeutics such as gemcitabine for glioblastoma therapy, achieving tumor-selective release while bypassing systemic clearance [158,159,165].
Quantitative pharmacokinetic assessments underscore the translational potential of these approaches [156,161,167]. Enrichment analyses of brain-to-plasma distribution indicate that optimized PLGA- and chitosan-based delivery systems can increase cerebral accumulation by several fold relative to unformulated compounds. These findings support the use of polymeric nanomedicine not merely as a means to enhance phytocompound bioavailability, but as a deliberate strategy to harness transport mechanisms for targeted CNS therapy [157,160,161,167].
4.2. Lipid Carriers (Liposomes, Solid Lipid Nanoparticles (SLNs), Nanoemulsions)
Lipid-based nanocarriers have become central to brain-targeted delivery, with liposomes representing the archetypal bilayer system [169,170]. Their amphiphilic structure allows for simultaneous encapsulation of hydrophilic and hydrophobic compounds, while PEGylated liposomes confer stealth properties that prolong circulation time and enhance BBB penetration [169,171,172]. This versatility has been leveraged in multiple preclinical models, where PEGylated formulations not only improved stability but also demonstrated controlled biodistribution within brain parenchyma [169,171,173]. The bilayer’s modularity also facilitates functionalization with targeting ligands, thereby adding a level of precision that polymeric systems often struggle to replicate [169,173].
Solid lipid nanoparticles (SLNs) and nanostructured lipid carriers (NLCs) have advanced the field by addressing limitations of traditional liposomes, particularly with respect to stability and drug-loading capacity [155,169,174]. SLNs, composed of solid lipids at body temperature, offer biocompatibility and controlled release but are often restricted by lower payload efficiency [155,175]. In contrast, NLCs incorporate both solid and liquid lipids into their matrix, thereby providing greater drug accommodation and reducing the risk of expulsion during storage [151,155,176]. Comparative studies consistently highlight the superior stability and performance of NLCs over SLNs, particularly for long-term formulations aimed at chronic neurodegenerative diseases [151,176,177].
Nanoemulsions extend the potential of lipid systems by enabling rapid and direct intranasal delivery to the brain [176,178]. Their small droplet size promotes fast absorption through the olfactory epithelium, a pathway particularly attractive for bypassing systemic metabolism [176,178,179]. However, reproducibility in manufacturing, along with challenges in preventing aggregation and ensuring shelf-life stability, remains an unresolved hurdles [178,180,181]. As recent in vivo studies emphasize, the promise of nanoemulsions lies in their high uptake efficiency, yet their clinical translation will depend on improved standardization and stabilization strategies [176,178,181].
4.3. Dendrimers and Micelles
Dendrimers represent one of the most structurally sophisticated nanocarriers for CNS therapy [182,183]. Their branched, tree-like architecture provides internal cavities for drug encapsulation and a multivalent surface for ligand attachment, making them particularly effective in tuning brain-specific targeting [182,183,184] critical challenge, however, lies in balancing ligand density: while higher densities can enhance receptor-mediated transcytosis, excessive functionalization often increases steric hindrance or cytotoxicity [182,184,185]. Studies with polyamidoamine (PAMAM) and carbosilane dendrimers demonstrate that size and surface charge strongly dictate BBB penetration, with mixed-surface or PEGylated variants offering improved biocompatibility and reduced clearance [186,187,188,189]. Such design nuances highlight the delicate trade-off between maximizing efficacy and minimizing off-target toxicity in vivo [182,184,187].
Micelles, by contrast, exploit their amphiphilic organization to solubilize poorly water-soluble compounds, a feature particularly advantageous for phytochemicals and nucleic acids [190,191]. Their self-assembly into nanosized aggregates enables responsiveness to physiological triggers, such as acidic pH or enzyme activity, resulting in controlled drug release within the brain microenvironment [192,193,194]. Optimization of micelle size and zeta potential has been shown to extend circulation while facilitating BBB passage without rapid renal clearance [186,190,192]. Recent developments in cation-free micelles for siRNA delivery illustrate how fine-tuned charge control can reduce cytotoxicity while preserving efficient uptake in glioblastoma models [190,195]. Together, dendrimers and micelles provide complementary strategies: dendrimers excel at multivalent, targeted interactions, while micelles offer dynamic, responsive platforms for solubilization and triggered release [186,191].
4.4. Inorganic/Carbon Nanostructures
Carbon- and inorganic-based nanostructures have carved a distinct niche in BBB research due to their dual roles as carriers and imaging agents [196,197,198]. Carbon dots and quantum dots, for example, exhibit intrinsic fluorescence, enabling simultaneous drug delivery and real-time tracking [196,199,200,201]. This built-in diagnostic capacity underpins their promise for theranostic applications, particularly in neurodegenerative disease models where monitoring biodistribution is critical [196,202,203,204]. Magnetic nanoparticles extend this paradigm further by enabling magnetically guided delivery, offering external control over accumulation in targeted brain regions [198,203,205]. Such approaches not only enhance precision but also reduce systemic exposure, positioning these nanostructures as powerful candidates for next-generation CNS therapeutics [197,198,203].
Despite these advantages, their translation faces significant barriers [199,206]. The long-term biocompatibility of quantum dots and carbon nanodots remains uncertain, with concerns over oxidative stress, protein corona formation, and potential accumulation in neural tissues [196,199,206,207]. Magnetic nanoparticles, while effective in guidance and imaging, also raise questions about clearance and toxicity with repeated use [198,203,205,206]. Animal studies have demonstrated promising biodistribution patterns, yet discrepancies in chronic safety outcomes highlight the need for rigorous toxicological evaluation before clinical adoption [199,206,208,209]. In this context, the field is actively exploring polymer-coated and functionalized variants to mitigate oxidative damage while preserving the diagnostic and therapeutic potential [198,203,205,206]. Together, inorganic and carbon nanostructures embody both the allure and caution of theranostic nanomedicine: they provide unparalleled control and visibility, but demand equally careful assessment of their long-term biological footprint [197,199,206].
4.5. Hybrid/Biodegradable and Protein Corona Control
Hybrid nanomedicine platforms are increasingly recognized as promising strategies for crossing the BBB because they combine complementary features of polymeric, lipid, and inorganic scaffolds [67,210,211]. Polymeric–lipid hybrids, for example, integrate the structural stability of polymers with the biocompatibility and drug-loading flexibility of lipid layers, while inorganic–organic hybrids offer imaging capabilities alongside controlled drug release [67,212,213]. A recurring theme in these designs is the fine-tuning of surface charge: adjusting zeta potential can minimize opsonization and prolong circulation, yet excessive neutralization may compromise cellular uptake [67,149,214]. Dual-targeting systems, such as polyanionic polymalic acid nanodrugs conjugated with Angiopep-2, illustrate how surface chemistry can be leveraged to maintain stability while enabling efficient receptor-mediated transport into the brain [214,215,216].
A critical determinant of in vivo performance lies in protein corona formation, which reshapes nanoparticle identity immediately upon systemic entry [217]. Far from being an inert byproduct, the corona can hinder transcytosis, reduce tumor selectivity, or alternatively, be engineered to guide delivery [217]. Strategies to regulate this interface include pre-coating with tunable surfactants, exploiting biomimetic exosome-mimetic shells, or even deliberately co-opting serum proteins to enhance stealth [217]. Such approaches demonstrate that corona engineering is not merely defensive but can be actively harnessed to improve BBB passage and targeting precision.
At the same time, clinical translation of these platforms hinges on biodegradability and safety [67,149,210]. Polyanhydride-based carriers and bioinspired protein–polymer nanocapsules exemplify progress toward fully degradable designs, yet challenges remain regarding reproducibility, long-term safety, and scale-up [67,149]. As recent reviews emphasize, the success of hybrid systems will depend not only on their multifunctional design but also on overcoming regulatory hurdles by proving that corona control and biodegradability can coexist without compromising efficacy [67,210,211].
4.6. Literature Snapshot
Comparative evaluations of nanomedicine platforms highlight how material choice shapes brain delivery outcomes [61,218,219]. Head-to-head studies reveal that PLGA nanoparticles carrying flavonoids often outperform liposomes in terms of controlled release and systemic stability, while liposomes enable faster brain penetration but can be prone to leakage and reduced retention [220,221,222]. Similarly, dendrimers and micelles demonstrate complementary advantages when loaded with peptide cargos: dendrimers benefit from multivalency that enhances receptor-mediated uptake, whereas micelles provide stimulus-responsive release and better solubilization of hydrophobic payloads [223,224,225]. Endpoints such as BBB permeability ratios, neurobehavioral outcomes in stroke or neurodegeneration models, and toxicity remain the unifying benchmarks across these comparisons [61,218,219].
The collective evidence makes one principle clear: there is no universal best nanomedicine platform [218,219,222]. The optimal choice is highly context-driven, defined by the therapeutic payload, disease model, and the balance between efficacy and safety [61,218,219]. Surface chemistry and biological targeting often make or break brain delivery—hence a focused look at ligands, valency, and stimuli [221,223,224] (Table 3).
5. Targeting and Stimuli Strategies
5.1. Receptor-Mediated Transcytosis (RMT) Ligands
Receptor-mediated transcytosis (RMT) has emerged as the cornerstone of BBB targeting, providing a rational route for therapeutic entry into the CNS [72]. Among the most extensively characterized receptors, the transferrin receptor (TfR), low-density lipoprotein receptor-related protein 1 (LRP1), insulin receptor (IR), and LDL receptor (LDLR) have each been exploited to shuttle biologics, peptides, and nanocarriers across endothelial cells [58,236]. Ligands such as Angiopep-2, apolipoprotein E (ApoE)-mimetics, and engineered transferrin derivatives illustrate how endogenous trafficking machinery can be co-opted without major structural perturbations to the BBB [58,236]. This strategy has been validated in multiple preclinical models and increasingly in human-relevant systems, including iPSC-derived BBB platforms that closely recapitulate receptor dynamics [237,238].
The success of RMT hinges on a finely tuned balance between ligand affinity and avidity [72,239]. Excessively strong binding may lead to receptor saturation or sequestration in lysosomes, while weak interactions risk premature dissociation and suboptimal delivery [70,239]. Mathematical modeling and linker engineering have refined our understanding of these trade-offs, highlighting an “affinity window” that permits recycling and productive transcytosis [239,240]. Yet, competition with endogenous ligands imposes a physiological ceiling effect, particularly for receptors like TfR and IR that are critical for nutrient homeostasis [240,241]. This ceiling necessitates dosing strategies and ligand modifications that preserve BBB transport without displacing natural substrates.
Translational progress has been most visible in the diversification of ligand classes. Antibody fragments and bispecific constructs with optimized linkers now outperform earlier monoclonals in preclinical BBB assays [32,242,243]. In parallel, short peptides, nanobodies, and aptamers provide modular alternatives that reduce immunogenicity while retaining high receptor specificity [70,244,245]. Collectively, these innovations illustrate how the conceptual framework of RMT is being transformed into clinically viable neurotherapeutic strategies, setting the stage for next-generation delivery systems tailored to the diseased brain microenvironment.
5.2. Adsorptive and Cell-Penetrating Peptide (CPP) Strategies
Adsorptive-mediated transcytosis and cell-penetrating peptides (CPPs) represent alternative strategies to receptor-based delivery for crossing the BBB [71,246,247]. Their principle rests on cationic surfaces that engage in electrostatic interactions with the negatively charged glycocalyx of endothelial cells, thereby initiating uptake [246,248]. Classic CPPs such as Tat, penetratin, and rabies virus glycoprotein (RVG) have been widely studied and consistently demonstrate high internalization efficiency across a range of in vitro and in vivo models [247,249,250]. This strong uptake capacity has made them attractive tools for brain-directed delivery of proteins, nucleic acids, and nanocarriers [246,247,250]. However, the very same non-specificity that enables broad penetration also increases the risk of cytotoxicity and off-target accumulation in peripheral tissues, posing major challenges for clinical translation [251,252,253].
To overcome these limitations, newer designs exploit reversible or conditional activation of cationic charges [254,255]. Charge-switchable coatings that remain neutral in circulation but expose CPP activity in acidic or enzymatically active microenvironments offer a way to enhance specificity while reducing systemic toxicity [254,255]. Stimulus-responsive CPPs integrated into nanogels or liposomes can be selectively unveiled in glioma or inflamed brain regions, thereby combining the high uptake efficiency of adsorptive strategies with a more targeted therapeutic profile [254,256,257]. These innovations suggest that adsorptive and CPP approaches, once criticized for their lack of selectivity, may reemerge as valuable complements to receptor-mediated systems when combined with smart design principles [247,251,254].
5.3. Multivalent/Dual-Targeting Designs
Multivalent and dual-targeting strategies build on the idea that no single ligand can fully capture the complexity of BBB transport and disease-specific recognition [258,259,260]. By combining receptor-mediated transcytosis ligands with disease-associated epitopes, such as amyloid-binding motifs in Alzheimer’s disease or glioma-homing peptides, researchers aim to achieve both efficient barrier penetration and selective accumulation in pathological tissue [241,259,261]. This layered approach provides synergy, since one ligand optimizes endothelial uptake while the second guides delivery toward neuronal or tumor targets [261,262,263]. The result is not simply additive transport but enhanced fidelity of targeting, often translating into improved therapeutic outcomes in preclinical models [261,262,264].
Nonetheless, designing dual-decorated nanocarriers requires careful calibration [260,264,265]. Steric hindrance between ligands, suboptimal linker lengths, or excessive surface density can compromise binding efficiency and even trigger accelerated clearance [264,265]. Several studies illustrate both promise and pitfalls [259,261,262]. In Alzheimer’s disease models, nanoparticles bearing transferrin and neuron-targeting Tet1 peptides showed superior cognitive rescue compared with single-ligand systems [259,261]. In glioblastoma, lipid nanocarriers co-functionalized with Angiopep-2 and TAT achieved deeper tumor penetration and survival benefits [259,261]. These examples highlight how multivalent strategies, when optimized, can balance BBB entry with precision delivery, positioning them as one of the most forward-looking directions in neurotherapeutics [32] [32,70,258].
5.4. Stimuli-Responsive Systems
Stimuli-responsive systems harness both endogenous and exogenous cues to achieve precise control over drug delivery across the BBB [33,266,267]. Internal triggers such as acidic pH gradients, redox imbalances, and overexpressed enzymes in the tumor microenvironment have been successfully integrated into nanocarriers to enable controlled and site-specific release [266,268,269]. pH-sensitive polymers, disulfide-cleavable linkers, and enzyme-activated coatings exemplify this strategy, ensuring that therapeutic cargo remains stable in circulation yet becomes rapidly available once inside diseased brain regions [186,269,270]. These approaches not only enhance local efficacy but also reduce systemic exposure, thereby addressing one of the central challenges of neurotherapeutics [220,267].
External stimuli offer an additional dimension of spatiotemporal precision [220,267,268]. Magnetic fields, focused ultrasound, and light-based activation provide reversible and non-invasive triggers that can be synchronized with drug administration [267,271,272]. Such methods have been paired with polymeric and lipid nanocarriers to achieve on-demand release and deep penetration into glioblastoma tissue [4,267,272]. However, questions of safety, reproducibility, and clinical feasibility remain unresolved, particularly for modalities requiring specialized equipment or prolonged exposure [220,273,274]. Balancing innovation with practicality is crucial as these systems move [220] toward translation [220,266,275]. Beyond targeting, alternative routes and device-enabled openings can bypass or transiently relax the barrier [4,54]. (Table 4, Figure 2).
6. Alternative Routes and Device-Enabled Blood-Brain Barrier (BBB) Opening
6.1. Intranasal Nose-to-Brain
The intranasal route exploits the unique anatomical connectivity between the nasal cavity and the brain through the olfactory epithelium and the branches of the trigeminal nerve [176,287]. These pathways enable both intra- and extra-neuronal transport, providing rapid and direct access to the CNS while bypassing systemic circulation and hepatic first-pass metabolism [176,288]. Such direct trafficking has been demonstrated for a wide range of small molecules, peptides, and nanocarrier systems, reinforcing the potential of this route for delivering neuroprotective phytochemicals and engineered prodrugs [176,289,290].
Formulation science has been central to enhancing this delivery mode [287,289]. Mucoadhesive in situ gels, often thermo- or ion-responsive, prolong nasal residence time and counteract mucociliary clearance, while nanoemulsions improve solubility and stability of hydrophobic polyphenols such as curcumin or resveratrol [176,291,292]. Nanoparticulate systems—ranging from lipid-based carriers to chitosan-modified polymeric nanoparticles—further allow surface functionalization for improved permeability and targeted release [149,290,293]. Prodrug strategies that exploit enzymatic conversion within the nasal mucosa are being increasingly explored to improve bioavailability and sustain brain exposure [287].
Despite this promise, translational hurdles remain [289,294]. Anatomical variability, short retention time, and interindividual differences in nasal airflow complicate dosing precision and reproducibility in humans [176,294]. Ergonomic device design, accurate metered dosing, and integration with pharmacokinetic modeling will be essential for clinical translation [176,294]. Large-scale human trials, coupled with regulatory harmonization, are still required before intranasal nanoformulations can be considered reliable delivery systems for neurotherapeutics [287,289,295].
6.2. Focused Ultrasound + Microbubbles (FUS)
Focused ultrasound combined with circulating microbubbles has emerged as one of the most precise approaches to transiently opening the BBB [296,297]. The mechanism relies on acoustic cavitation, in which microbubbles oscillate in response to ultrasound exposure, producing localized shear stress on the vascular endothelium [296,298,299]. This process induces mechanoporation and loosening of tight junctions, thereby increasing paracellular permeability in a controlled and reversible manner [296,298,299]. Importantly, both stable and inertial cavitation contribute to permeability enhancement, yet parameters must be carefully tuned to avoid endothelial damage or hemorrhage [298,300,301]. The reversible nature of the opening distinguishes FUS from chemical osmotic methods, as the barrier typically restores within hours [296].
A major strength of this technology lies in its spatiotemporal precision [296,302,303]. MRI-guided or neuronavigation-based systems allow targeting of submillimeter brain regions, enabling localized drug accumulation with minimal off-target exposure [302,304,305]. Real-time cavitation monitoring, coupled with feedback-controlled ultrasound delivery, provides essential safety guardrails, reducing risks of edema, neuroinflammation, or microvascular injury [300,306,307]. Longitudinal studies in both primates and humans confirm the feasibility of repeated sessions without significant adverse cognitive effects, although vigilance for subtle inflammatory responses remains necessary [298,308,309].
Clinical translation is already well under way [302]. Phase I trials in gliomas, Alzheimer’s disease, and Parkinson’s dementia consistently report tolerability and transient BBB disruption, with imaging confirming enhanced delivery of chemotherapeutics, antibodies, and nanoparticles [298,302,310]. While no trials have yet evaluated phytochemicals or natural prodrugs directly, the compatibility of FUS with nanocarriers and controlled-release systems makes such applications plausible [297,299,310]. Integrating polyphenol-based therapeutics into FUS platforms could represent a novel frontier for noninvasive neuroprotection and disease modification [299,310] (Table 3).
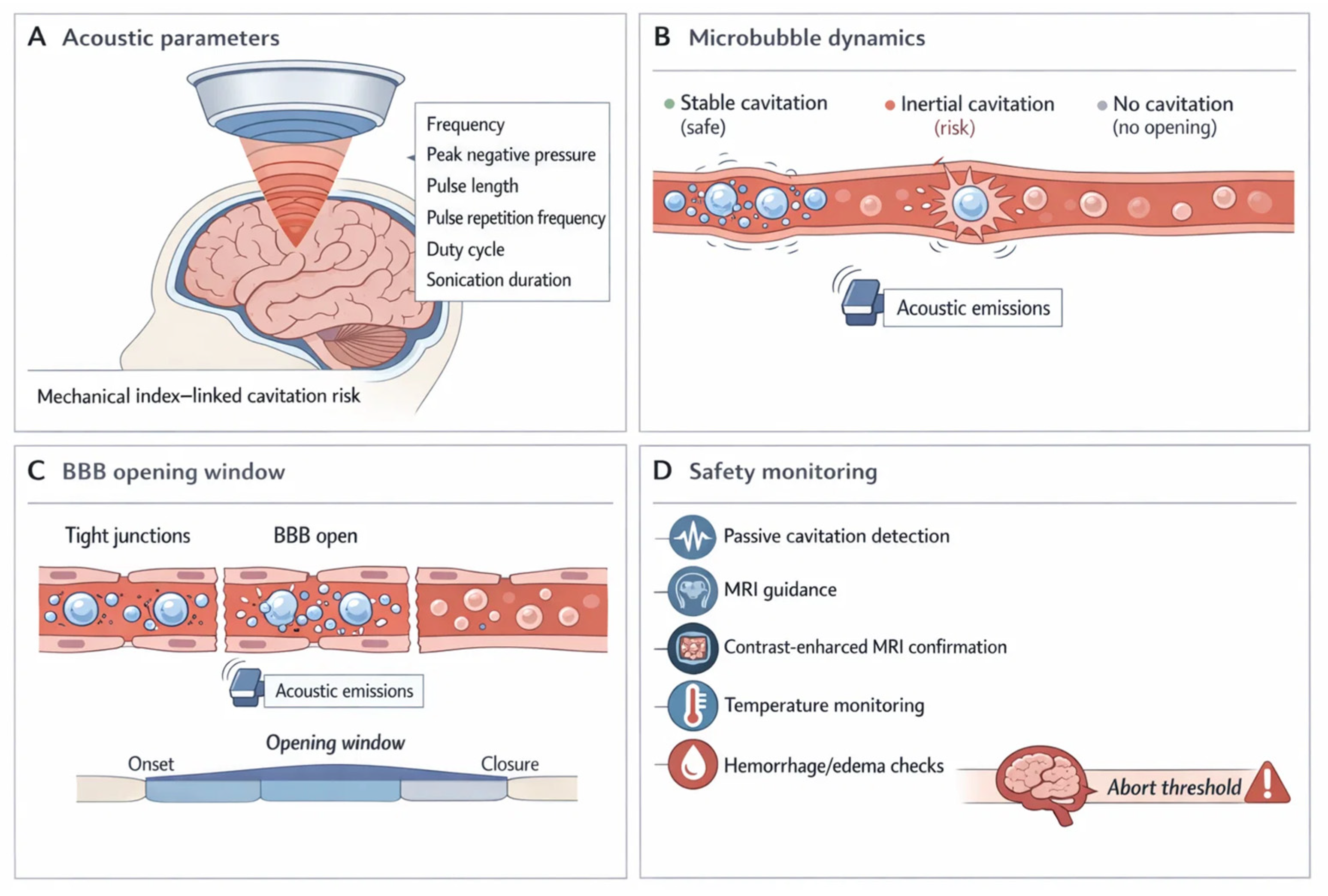
Figure 3.
Focused ultrasound (FUS) delivery paradigm: acoustic control, microbubble cavitation, transient blood–brain barrier (BBB) opening, and safety monitoring. The schematic summarizes the focused ultrasound (FUS)–microbubble paradigm for transient, spatially targeted BBB modulation to enable CNS delivery. (A) Acoustic parameters (frequency, peak negative pressure, pulse length, pulse repetition frequency, duty cycle, and sonication duration) set the cavitation regime and therefore the balance between efficacy and risk[311,312,313] 10.3171/2017.11.focus17627. 10.1016/j.ultsonch.2018.12.031. 10.1016/j.ultsonch.2016.09.016. (B) Microbubble dynamics span a spectrum from no response (insufficient opening) to stable cavitation (desired oscillation that increases permeability) and inertial cavitation (collapse associated with vascular damage risk). These regimes are inferred in real time using acoustic emissions and passive cavitation detection to support parameter adjustment and stop rules[307,314,315] 10.1038/s41598-020-66994-8. 10.1016/j.ultsonch.2023.106291. 10.1109/tbme.2018.2882337. (C) BBB opening window is depicted as a reversible permeability increase (minutes–hours) during which small molecules, prodrugs, and nanocarriers can cross the endothelium; closure restores barrier integrity[316,317,318] 10.1371/journal.pone.0125911. 10.1038/s41598-018-36340-0. 10.3171/2022.9.jns221565. (D) Safety monitoring includes image-guided targeting (MRI), confirmation of opening (contrast-enhanced MRI), surveillance for edema or microhemorrhage, and temperature/physiological checks where relevant. Together, the framework links controllable acoustic inputs to microbubble behavior, delivery timing, and verification steps required for reproducible, trial-grade BBB opening[307,316,319] 10.1073/pnas.2103280118. 10.1038/s41598-020-66994-8. 10.1371/journal.pone.0125911. BBB, blood–brain barrier; CNS, central nervous system; FUS, focused ultrasound; MRI, magnetic resonance imaging.
Figure 3.
Focused ultrasound (FUS) delivery paradigm: acoustic control, microbubble cavitation, transient blood–brain barrier (BBB) opening, and safety monitoring. The schematic summarizes the focused ultrasound (FUS)–microbubble paradigm for transient, spatially targeted BBB modulation to enable CNS delivery. (A) Acoustic parameters (frequency, peak negative pressure, pulse length, pulse repetition frequency, duty cycle, and sonication duration) set the cavitation regime and therefore the balance between efficacy and risk[311,312,313] 10.3171/2017.11.focus17627. 10.1016/j.ultsonch.2018.12.031. 10.1016/j.ultsonch.2016.09.016. (B) Microbubble dynamics span a spectrum from no response (insufficient opening) to stable cavitation (desired oscillation that increases permeability) and inertial cavitation (collapse associated with vascular damage risk). These regimes are inferred in real time using acoustic emissions and passive cavitation detection to support parameter adjustment and stop rules[307,314,315] 10.1038/s41598-020-66994-8. 10.1016/j.ultsonch.2023.106291. 10.1109/tbme.2018.2882337. (C) BBB opening window is depicted as a reversible permeability increase (minutes–hours) during which small molecules, prodrugs, and nanocarriers can cross the endothelium; closure restores barrier integrity[316,317,318] 10.1371/journal.pone.0125911. 10.1038/s41598-018-36340-0. 10.3171/2022.9.jns221565. (D) Safety monitoring includes image-guided targeting (MRI), confirmation of opening (contrast-enhanced MRI), surveillance for edema or microhemorrhage, and temperature/physiological checks where relevant. Together, the framework links controllable acoustic inputs to microbubble behavior, delivery timing, and verification steps required for reproducible, trial-grade BBB opening[307,316,319] 10.1073/pnas.2103280118. 10.1038/s41598-020-66994-8. 10.1371/journal.pone.0125911. BBB, blood–brain barrier; CNS, central nervous system; FUS, focused ultrasound; MRI, magnetic resonance imaging.
6.3. Chemical/Osmotic Opening and Convection-Enhanced Delivery (CED)
Classical BBB-disruption strategies such as intra-arterial mannitol, DMSO co-solvent effects, and bradykinin analogs can transiently loosen tight junctions, yet their clinical utility has waned [273,320]. The reasons are consistent across reviews: non-selective permeability increases, variable magnitude and duration of opening, and procedure-related risks, including seizures, edema, or ischemic events [321,322]. Lack of spatiotemporal control and systemic toxicities further erode risk–benefit in longitudinal care [320]. For phytochemicals and natural prodrugs, which often require sustained or repeated exposure, these invasive and poorly tunable methods are a poor fit; cumulative toxicity and patient burden compound the translational gap [320,323].
CED preserves a niche when focal, high-dose deposition is essential, as in gliomas or diffuse midline lesions, particularly with implantable ports and image-guided catheters, despite technical complexity and heterogeneity of distribution [324,325,326]. When chemistry can carry the payload, prodrugs simplify the problem, if transporter hijacking and cleavage are tuned correctly [327,328] (Table 5).
7. Prodrugs and Transporter Hijacking
7.1. LAT1/MCT1/GLUT1-Targeted Prodrugs
Among the influx transporters that shape small-molecule entry into the brain, the large neutral amino acid carrier LAT1 has emerged as the most exploited in prodrug design [336,337,338]. LAT1 is highly expressed on the luminal side of brain capillaries and recognizes aromatic and branched-chain amino acids as substrates [53,338,339]. By conjugating drugs with phenylalanine, tyrosine, or related promoieties, it is possible to achieve carrier-mediated uptake that circumvents passive BBB limitations [336,337,340]. LAT1-linked prodrugs of valproic acid, ferulic acid, and NSAIDs have shown superior brain penetration, and the Xiong 2021 dataset provides compelling evidence that conjugated neurotherapeutics not only cross the BBB but also accumulate within neurons, astrocytes, and microglia, confirming cellular specificity of uptake [341,342,343].
Beyond LAT1, other solute carriers are beginning to attract attention [56,338] Monocarboxylate transporter 1 (MCT1) recognizes lactate and pyruvate analogues, providing a scaffold for monocarboxylate-linked prodrugs, whereas GLUT1, the primary glucose transporter, can be hijacked via glucose conjugation [53,56,338]. Proof-of-principle studies demonstrate that indomethacin and ketoprofen conjugated to glucose traverse the BBB in rodents, though kinetic competition with endogenous glucose poses significant challenges [53,343]. These strategies illustrate the expanding toolkit for tailoring prodrug chemistry to align with the substrate repertoire of BBB carriers [53,338].
Transporter hijacking, however, is not without risk [336,344]. Kinetic constraints such as Km and Vmax dictate the efficiency of uptake, and saturation by high-affinity endogenous substrates can diminish drug delivery [53,337]. Moreover, transporter expression varies across species, complicating preclinical-to-clinical translation [56,338]. LAT1 prodrugs are generally selective, off-target interactions and potential saturation effects remain critical safety considerations [341,345,346]. The challenge now lies in fine-tuning conjugate chemistry to balance affinity, stability, and enzymatic cleavability in the brain while minimizing systemic exposure [339,340,342].
7.2. Lipidization, Soft Drugs, Self-Immolative Linkers
Lipidization remains one of the oldest yet most versatile strategies for enhancing drug penetration into the brain [347,348,349]. By appending lipophilic chains or glyceride motifs, polar APIs can acquire sufficient passive diffusion across endothelial membranes, provided the modifications are designed for efficient cleavage once in the CNS [347,348,349]. This balance between increased lipophilicity and metabolic lability is critical: too stable and the parent drug may not be released; too labile and systemic hydrolysis prevents brain delivery [347,349]. The approach has been applied successfully to small neuroactive agents, though reproducibility across species remains a central design challenge [348,350].
In parallel, soft drug concepts introduce the inverse logic: compounds are deliberately engineered for predictable inactivation outside the CNS, ensuring that only a fraction escapes rapid metabolism and reaches the brain [347,349]. Self-immolative linkers add yet another layer of sophistication, exploiting pH gradients, enzyme expression, or redox triggers to launch controlled cleavage cascades [351,352,353]. Modern designs favor traceless release, often with dual stimuli or cascade amplification to achieve brain-first activation while avoiding premature systemic leakage [351,352,354]. The guiding rule across these platforms is to harmonize stability, trigger sensitivity, and cleavage kinetics so that release occurs only under CNS-relevant conditions, minimizing off-target toxicity while maximizing therapeutic gain [351,352,354].
7.3. Solubility Boosters (Cyclodextrins, Co-Crystals, Ion Pairing)
Cyclodextrins have been widely used to improve aqueous solubility through inclusion complexes that sequester hydrophobic moieties within their cyclic cavities [142,355,356]. This strategy can significantly enhance systemic exposure and oral bioavailability, yet it offers little direct benefit for BBB permeation, as the bulky complexes rarely cross endothelial tight junctions intact [355,357,358]. Their role is therefore supportive: enabling consistent systemic levels that may feed into other brain-targeted strategies rather than acting as genuine CNS delivery enhancers [356,359,360].
Co-crystals and ion pairing occupy a more dynamic niche [361]. Co-crystals modify dissolution rates and solubility without altering the pharmacodynamic profile of the parent drug, creating opportunities for predictable exposure kinetics [361,362,363]. Ion pairing, in contrast, transiently adjusts lipophilicity by associating ionizable drugs with counterions, thereby improving membrane partitioning and yielding short-lived permeability gains [364,365,366]. The central distinction is crucial: while all three approaches may improve systemic bioavailability, only certain ion-pairing strategies directly modulate BBB permeability [364,365,366]. Recognizing this separation between systemic solubility enhancers and true BBB permeability modulators is essential when positioning such methods within prodrug pipelines [365,366,367].
7.4. Nano–Prodrug Conjugates
Nano–prodrug conjugates represent a convergence of nanomedicine and classical prodrug chemistry [368,369,370]. In these systems, nanocarriers such as polymers, liposomes, or albumin-binding constructs are covalently linked to prodrug moieties, creating assemblies that combine carrier stability with controlled release [368,370,371]. Examples include polymer–drug conjugates that self-assemble into micelles or nanoparticles, and liposome–prodrug hybrids that integrate covalently modified drugs into bilayer structures [227,368,372]. Activation is then triggered by tumor- or CNS-relevant stimuli such as redox gradients, pH shifts, or enzyme cleavage, ensuring spatially restricted release [269,372,373].
The rationale for this complexity is strongest when dealing with drugs that have narrow therapeutic windows or poor solubility, where conventional formulations risk systemic toxicity or inadequate exposure [370,374]. By embedding prodrug chemistry within nanocarriers, it becomes possible to synchronize delivery, minimize premature release, and improve therapeutic indices [368,374,375]. Yet translation remains challenging [375,376,377]. Manufacturing reproducibility, batch-to-batch stability, and regulatory pathways for hybrid entities blur the lines between drug and device, complicating approvals [376,377,378]. Scalability and quality control of multifunctional prodrug nanocarriers are further hurdles that limit current clinical penetration despite compelling preclinical evidence [376,377,379] Delivery vectors and prodrugs must be vetted in models that actually predict human exposure—next we align models with decision-grade endpoints [376,377] (Table 6).
8. Biogenic and Exosome-Mimetic Vesicles
8.1. Mammalian Exosomes
Mammalian exosomes have attracted intense interest as endogenous delivery vehicles, given their origin from neuronal, immune, and stem-cell lineages [392,393]. Neuron-derived vesicles display inherent neurotropism, while macrophage or dendritic cell exosomes often retain immunological signaling capabilities that can be leveraged for targeted delivery [393,394]. Stem cell-derived vesicles, particularly those from mesenchymal sources, exhibit regenerative properties and have been applied in models of neuroinflammation and tissue repair [395,396]. This natural diversity provides a menu of options for CNS-directed therapy, with the vesicle’s parent cell type influencing both tropism and therapeutic payload [395,397].
Several methods exist for incorporating cargo into exosomes [394,398]. Electroporation transiently disrupts vesicle membranes to load nucleic acids, while passive incubation exploits lipid bilayer partitioning [394,399]. Sonication and extrusion, though less subtle, can increase loading efficiency for small molecules and proteins [399,400]. More sophisticated approaches combine chemical conjugation or ligand decoration to engineer selective homing properties, extending beyond the vesicle’s innate targeting profile [398,401].
Despite these advantages, translational obstacles remain formidable [402,403]. Batch-to-batch variability complicates reproducibility, and large-scale production has yet to reach regulatory-grade consistency [392,402,403]. Issues of heterogeneity, yield, and purification standards pose barriers to clinical adoption, while classification of exosomes as biologics, devices, or drug–biologic hybrids remains unresolved [404,405]. Thus, mammalian exosomes stand at the intersection of promise and challenge, offering unmatched biocompatibility but demanding rigorous solutions in scalability and regulation before they can function as reliable neurotherapeutic vectors [392,393].
8.2. Plant-Derived Extracellular Vesicles
Plant-derived extracellular vesicles (PDEVs) are emerging as abundant, low-costnanocarriers harvested from edible sources such as ginger, grape, and citrus [406,407,408]. Their natural stability, low immunogenicity, and tolerance to gastrointestinal conditions make them particularly attractive for oral or intranasal administration, routes that remain challenging for mammalian exosomes [406,409,410]. PDEVs also carry intrinsic bioactive metabolites, adding antioxidant and anti-inflammatory potential to their delivery role [408,411,412].
Despite these advantages, several limitations temper enthusiasm [406,407]. Vesicle heterogeneity across plant species and even between batches complicates reproducibility, while the mechanisms by which PDEVs interact with or traverse the BBB remain poorly defined [406,412,413]. Preclinical studies demonstrate promising antioxidant and anti-inflammatory effects in models of neuroinflammation and oxidative stress, yet translation into predictable CNS uptake remains uncertain [414,415,416]. Thus, PDEVs occupy a unique space: safe, scalable, and bioactive, but requiring deeper mechanistic insight before they can be positioned as reliable neurotherapeutic vectors [407,413,417].
8.3. Synthetic Mimetics
Synthetic exosome-mimetic vesicles are designed to replicate the communication and delivery roles of natural exosomes while sidestepping their limitations of yield and heterogeneity [418,419]. Strategies include polymersomes with controllable membrane chemistry, membrane-coated nanoparticles that borrow cellular surface markers, and hybrid designs that combine synthetic scaffolds with natural membrane fragments [420,421,422]. Such constructs excel in tunability and scalability, making them better suited for standardized manufacturing compared to their mammalian counterparts [418,419,423].
Yet these advantages come with trade-offs [421,422]. Replacing native membranes often diminishes biocompatibility cues that exosomes naturally provide, raising concerns about immune activation and altered clearance [421,422,423]. Nonetheless, synthetic platforms allow for reproducible incorporation of targeting ligands or exosomal motifs, offering a level of precision that natural vesicles rarely achieve [418,419,420]. Early applications highlight their promise in oncology, regenerative medicine, and CNS targeting [421,424,425]. We now connect models to the endpoints that drive go/no-go decisions.
9. Translational Models and Decision-Enabling Endpoints
9.1. In Vitro Models
In vitro BBB models remain indispensable as early decision tools in neurotherapeutic development [426,427]. Classic Transwell systems with endothelial monolayers offer simplicity and throughput but often fail to reproduce the restrictive tight junctions of the human BBB [428,429]. Adding astrocytes or pericytes in co-culture improves fidelity, as astrocytic signals reinforce junctional protein expression and better align transendothelial electrical resistance (TEER) values with physiological ranges [427,430,431]. These refinements help distinguish passive permeability from transporter-mediated flux, although limitations in dynamic responses persist [428,432].
Human induced pluripotent stem cell (iPSC)-derived BBB organoids introduce greater biological relevance by capturing species-specific expression of transporters and efflux pumps [427,433]. However, the lack of standardized differentiation protocols results in variable permeability and metabolic profiles across laboratories [427,434]. Microfluidic BBB-on-chip systems address some of these issues by incorporating shear stress, nutrient gradients, and continuous flow, recapitulating the hemodynamic conditions that shape BBB integrity [428,432,435]. These dynamic constructs offer a closer physiological context but are more technically demanding and costly to implement [432,435].
Benchmarking against in vivo data remains essential [428,435]. Metrics such as apparent permeability (P_app) and TEER are routinely compared to animal and human datasets, yet over- or underestimation of drug transport is common [429,436]. The dataset reported by Aday and colleagues (2016) illustrates how carefully calibrated microfluidic models can achieve closer alignment with in vivo permeability coefficients [428,435]. Despite progress, no single in vitro system fully resolves the trade-off between scalability and predictive accuracy, underscoring the need for model selection tailored to the specific decision point in development [426,432].
9.2. In Vivo Models and Species Differences
Rodent models remain the workhorse of preclinical neurotherapeutics, offering high-throughput screening, ease of genetic manipulation, and well-characterized disease models [437] Yet their BBB exhibits greater paracellular leakiness than that of primates, which can overestimate drug penetrance [438,439]. This divergence partly explains why promising rodent data often fail to translate into clinical success [437,440]. Moreover, rodents display transporter expression patterns that differ in both abundance and substrate specificity compared to humans, adding further complexity to predictions of central exposure [440,441,442].
Non-human primates provide the closest approximation of human BBB integrity and regional perfusion characteristics [441,443]. Their barrier tightness, transporter repertoire, and cerebrovascular physiology more closely align with human data, making them critical for late-stage validation [439,444]. However, cost, ethical concerns, and limited availability restrict their widespread use [444]. Adding further complication, disease states reshape barrier permeability: ischemic stroke disrupts endothelial junctions, Alzheimer’s disease alters transporter activity, and glioblastoma induces localized leakiness that changes drug distribution [344,445,446].
These species and disease-dependent differences emphasize the translational gap between model systems and patients [437,438]. Cross-species network analyses and computational integration strategies are increasingly used to bridge this gap, but the fundamental challenge remains: no single in vivo model fully captures the nuances of human BBB physiology [429,447]. Strategic selection and careful benchmarking are therefore essential to guide go/no-go decisions in CNS drug development [429,439].
9.3. Quantitative PK Endpoints
Quantitative pharmacokinetic endpoints are central to linking drug exposure with CNS activity [448]. The unbound fraction in brain tissue (f_u,brain) defines the pharmacologically active pool, while the brain-to-plasma partition coefficient (K_p,brain) describes overall distribution across compartments [449]. A more precise index is K_p,uu,brain, the ratio of unbound brain to unbound plasma concentrations, which reflects true equilibrium between compartments and better predicts central efficacy [449,450]. These parameters guide whether a compound achieves sufficient free concentrations at its target site or is limited by efflux transporters and protein binding [451,452].
Measuring these endpoints remains technically demanding [448]. Microdialysis enables direct sampling of interstitial fluid, offering dynamic readouts of unbound concentrations, but it is invasive and limited to specialized settings [453] Homogenate binding assays, in contrast, are more accessible but prone to overestimation due to disrupted tissue architecture [450,454]. Cerebrospinal fluid is often used as a surrogate for interstitial concentrations, yet differences in turnover and compartmentalization mean CSF rarely mirrors brain extracellular fluid with high fidelity [455,456]. Regulators increasingly emphasize integration of such quantitative PK endpoints with pharmacodynamic measures, particularly through physiologically based pharmacokinetic and PK–PD models, to inform dose selection and reduce translational uncertainty [456,457,458]. By anchoring drug development in f_u,brain, K_p,brain, and K_p,uu,brain, researchers can more confidently bridge preclinical data with human predictions and make decision-grade assessments of CNS penetration [450,459].
9.4. Imaging and Biomarker Readouts
Imaging has become a cornerstone in evaluating how drugs and nanocarriers navigate the BBB [460,461]. Positron emission tomography (PET) tracers provide sensitive, quantitative assessments of permeability, while magnetic resonance imaging (MRI) with contrast agents captures dynamic leakage and regional perfusion in vivo [460,462]. Recent work extends these approaches to track nanocarrier fate over time, revealing how size, charge, and surface chemistry influence deposition within target regions [461,463]. Such dynamic readouts are invaluable not only for confirming delivery but also for ruling out vascular compromise or off-target accumulation that could cloud efficacy signals [464].
In parallel, biomarker development is beginning to complement and extend imaging [465]. Neuroinflammation markers such as GFAP or ICAM-1 flag astrocytic and endothelial responses, while circulating exosomal signatures hint at brain-specific injury or remodeling processes [466,467]. The true translational power lies in linking imaging and biomarkers simultaneously to efficacy—drug exposure within the intended region—and safety, including the detection of edema or inflammatory activation [468,469]. Emerging strategies combine multimodal imaging with panels of fluid biomarkers, offering a near real-time window into drug delivery, target engagement, and tissue response [465,470,471]. This convergence is setting the stage for decision frameworks that go beyond single endpoints and instead integrate orthogonal readouts to guide go/no-go calls with greater confidence [468,472]. What has actually reached patients? We summarize clinical traction and why certain bets are moving first [460] (Table 7).
10. Clinical Landscape and Case Snapshots
10.1. Neuro-Oncology
Focused ultrasound (FUS) with microbubbles is the most advanced clinical strategy for transiently opening the BBB in neuro-oncology [488,489,490] By generating localized acoustic cavitation, FUS temporarily loosens tight junctions, permitting chemotherapeutics to achieve higher intratumoral concentrations than with systemic dosing alone [488,489,490,491]. Early-phase trials in glioblastoma and brain metastases report encouraging safety signals, with most adverse events being transient edema or headaches rather than irreversible damage [489,491]. Imaging-confirmed increases in drug penetration, paired with pharmacokinetic analyses, have strengthened confidence that this approach is technically feasible and biologically impactful [489,491].
Parallel efforts explore ligand-targeted nanocarriers for glioblastoma, including transferrin- and integrin-directed liposomes, which are designed to selectively home to tumor vasculature or infiltrating glioma cells [492,493,494]. These systems aim not only to improve local accumulation but also to minimize systemic exposure [492,493]. Key endpoints now extend beyond radiographic progression-free survival to include intratumoral drug levels, pharmacodynamic signatures, and radiomic biomarkers that track response heterogeneity [274,489,490]. Yet regulatory progress remains uneven. Enrollment in neuro-oncology trials frequently lags behind projections, with disparities in infrastructure and patient access limiting study completion rates [317,495]. As adaptive designs and external control datasets gain traction, the field is moving toward more flexible, inclusive trial frameworks capable of sustaining momentum in a disease space with urgent unmet needs [317,490,495].
10.2. Neurodegeneration
Alzheimer’s disease trials have tested diverse delivery routes, with intranasal insulin standing out for its ability to bypass systemic metabolism and provide direct brain access [496,497]. Peptide- and polyphenol-loaded carriers are also under investigation to stabilize bioactive molecules while enhancing their penetration into hippocampal and cortical regions [498,499]. Although some studies report cognitive benefits and favorable biomarker shifts, variability in patient populations and endpoint sensitivity continues to limit clear conclusions [500,501,502]. Reliance on radiographic and cognitive scales alone often underestimates subtle, early effects, underscoring the need for multimodal biomarker panels [500,501,503].
In Parkinson’s disease, dopamine prodrugs and nanoparticle-based formulations represent strategies to extend half-life and reduce peripheral toxicity while restoring striatal dopamine tone [157,504,505]. Several trials show encouraging motor improvements, yet variability in absorption and BBB transport remains a barrier to consistency [504,505,506]. Lessons from both Alzheimer’s and Parkinson’s pipelines converge on the importance of robust biomarkers, sensitive endpoints, and trial designs that accommodate disease heterogeneity [500,501,503]. Without these refinements, even promising therapeutic concepts risk falling short in translation [500,503,507].
10.3. Psychiatric and Pain Indications
Early translational studies in psychiatry and pain have focused on phytochemicals such as curcumin and resveratrol, as well as terpenoids such as pinene and linalool, which show preclinical promise for mood regulation and analgesia [30,508,509]. Yet progress into robust clinical validation remains limited [30,508]. Subjective endpoints and the strong influence of placebo responses complicate signal detection, while modest funding and heterogeneous trial designs further slow momentum [510,511]. Current exploratory efforts in depression and chronic pain increasingly employ advanced delivery systems and biomarker-informed approaches, but most remain proof-of-concept [512,513,514,515]. The field illustrates both opportunity and fragility in translating natural compounds into psychiatric and pain therapeutics [30,508,516,517].
10.4. Snapshot
First-in-human studies with natural compounds illustrate both opportunity and limitation across therapeutic domains. Oncology has the deepest record, with plant-derived chemotherapeutics and semi-synthetic derivatives advancing into late-stage trials [518,519]. Yet many candidates stall due to safety uncertainties, inconsistent batch quality, or regulatory concerns related to chemistry and manufacturing controls [520]. In neurodegeneration, compounds such as curcumin, resveratrol, and quercetin have entered clinical testing, often showing bioactivity but hampered by poor bioavailability and heterogeneous outcomes [21,521,522]. Psychiatric indications remain the least mature, with strong preclinical rationale but scarce head-to-head trials against approved therapies [523,524].
This comparative landscape highlights clear gaps for phytochemicals: translation remains fragmented, efficacy signals are often modest, and reproducibility suffers without rigorous manufacturing standards [519,520]. The field now recognizes that scientific novelty alone is insufficient. Translation hinges on safety, manufacturability, and regulatory clarity—complex systems demand disciplined CMC [524] (Table 8).
11. Material Safety and Immunogenicity
11.1. Hemolysis, Complement Activation, Microglial Responses
Early material safety screening hinges on blood compatibility, because initial interactions with blood components often dictate downstream immune trajectories [536,537]. Hemolysis is not a benign artifact but an active trigger of innate immunity [538,539]. Cell-free heme and heme-bearing microvesicles directly activate the complement cascade, driving C3 cleavage, leukocyte activation, and cytokine release, thereby linking red blood cell damage to acute inflammatory toxicity and organ injury [538,539,540]. As a result, hemolysis assays are most informative when paired with measurements of complement split products and early cytokines in serum or whole-blood systems [536,540,541]. Across preclinical and clinical contexts, rising immune complexes, C3a generation, or depletion of C3 and C4 consistently correlate with infusion reactions and dose-limiting hypersensitivity, particularly during dose escalation [538,541,542].
For CNS-targeted materials, microglia and astrocytes represent a distinct and highly sensitive safety axis [111,543,544]. Complement opsonization can promote microglial uptake that is either neuroprotective or deleterious, depending on persistence and inflammatory tone [543,544]. Astrocyte-derived complement components, together with IL-1, TNF, and IL-6 signaling, shape microglial activation states and synaptic integrity [544,545,546]. Here, physicochemical parameters act as immune dials rather than binary switches [536]. Smaller size, higher dose, and increased positive surface charge enhance uptake and cytokine release, while excessive activation pushes glia toward chronic inflammatory phenotypes [546,547,548]. Early integration of these variables helps distinguish immunologically silent designs from those primed to provoke neuroimmune risk [547,548,549].
11.2. Hemocompatibility and Neuroinflammation Assays
Standardized hemocompatibility testing remains the first safety filter for blood-contacting and intravascular materials [550,551,552]. In vitro panels aligned with ISO 10993-4 typically assess platelet adhesion and aggregation, the intrinsic and extrinsic coagulation pathways, and complement activation, using through thrombin generation, aPTT, platelet surface markers, and C3a or C5b-9 formation [550,552,553]. Sequential whole-blood and platelet-rich plasma assays increasingly capture the cascade from protein adsorption to thrombogenicity and cytokine release, allowing mechanistic interpretation rather than binary pass-fail outcomes [550,554].
Neuroinflammation assays extend this logic into the CNS space. Human iPSC-derived microglia and astrocytes, cocultures, and emerging organoid systems enable multiplex cytokine profiling, complement C3 readouts, and neurotoxicity markers under controlled stimuli [555,556,557]. Critically, aligning these outputs with clinically validated biomarkers such as GFAP, IL-6, or TNF strengthens the bridge between in vitro signals and patient-level neuroinflammatory risk [558,559,560,561].
11.3. Biodistribution and Clearance
Biodistribution and clearance represent a central determinant of both efficacy and long-term safety for material-based therapeutics [562,563]. Following systemic administration, the majority of nanoscale materials are rapidly sequestered by the mononuclear phagocyte system, with liver and spleen often capturing most of the injected dose [564]. Uptake by Kupffer cells, splenic macrophages, and sinusoidal endothelium can markedly reduce target tissue exposure while establishing persistent intracellular reservoirs [564,565]. Such retention may remain clinically silent, yet it raises concerns under repeated dosing and complicates the interpretation of chronic toxicity risk [564,566].
Design choices strongly bias this balance between persistence and elimination [562,567]. Ultrasmall or biodegradable architectures favor renal or hepatobiliary clearance, shortening organ residence while preserving therapeutic exposure [563,568]. In contrast, larger or rigid constructs tend toward lysosomal trapping [562,569] De-risking strategies increasingly combine biodegradable scaffolds, surface chemistry optimization, and dose fractionation to limit cumulative burden without sacrificing pharmacological performance [570,571,572].
11.4. CMC/GMP and Critical Quality Attributes (CQAs)
Critical quality attributes anchor the translation of complex materials from bench to clinic [573,574]. Across quality by design frameworks, particle size, polydispersity, zeta potential, encapsulation efficiency, and release kinetics consistently emerge as core CQAs because they integrate manufacturability with exposure and immunogenicity risk [573,575]. Multivariate and machine learning driven designs show that modest shifts in process parameters can propagate into meaningful changes in these attributes, with downstream effects on stability and biological performance [576,577,578] 10.3390/ijms262010238. Release profiles, often first order or diffusion controlled, are increasingly treated as quantitative CQAs rather than descriptive outcomes [573,579].
For CNS administered products, sterility and endotoxin control are nonnegotiable [580,581] Endotoxin thresholds are substantially lower than for systemic routes, reflecting heightened neuroinflammatory sensitivity [582,583]. Routine lot release; therefore, couples validated BET or rFC assays with conservative specifications aligned with intrathecal exposure [582,584]. Stability programs add another layer of complexity [585,586]. Aggregation, content leakage, and loss of redispersibility during storage or lyophilization can silently erode CQAs unless cryoprotectants and freezing protocols are optimized [586,587].
11.5. Regulatory Expectations
Regulatory agencies approach nanomedicines through a risk-based, case-by-case lens that reflects their structural diversity and evolving biology [591,592]. Both the FDA and the EMA emphasize nanomedicine-specific risks that extend beyond those of conventional small molecules, including altered biodistribution, immune activation, and long-term tissue persistence [591,593]. Guidance increasingly calls for deeper physicochemical characterization, nano-relevant immunotoxicity testing, and justification when standard ICH assays lack sensitivity [594,595]. For complex biological nanoparticle hybrids such as lipid nanoparticles, polymer conjugates, or gene delivery systems, regulators treat products as non-biological complex drugs, limiting assumptions of equivalence and requiring product-specific clinical evidence [591]
Bridging preclinical data to first-in-human studies relies on standardized safety frameworks that integrate in vitro and ex vivo human blood assays with human blood, and targeted in vivo assays [594,596]. For CNS indications, expectations tighten further [597]. BBB interactions, neuroinflammation risk, and irreversible outcomes demand a transparent risk–benefit narrative grounded in mechanistic data rather than exposure alone [595,597]. Data science tightens design loops and right-sizes risk before first dose in humans [598,599].
12. Data Science, Modeling, Artificial Intelligence (AI)-Guided Design
12.1. BBB Permeability Prediction and Polypharmacology
Data science increasingly reframes CNS design from intuition to prediction [600,601] QSAR and machine learning models trained on large BBB datasets now capture both quantitative logBB and categorical permeability with accuracy that supports early triage [600,601]. Beyond simple lipophilicity, modern models incorporate nonlinear descriptors and explicitly account for transporter effects, with P-gp emerging as a dominant determinant of CNS variability [147,602]. This is especially relevant for polyphenols, where favorable passive diffusion can be offset by strong efflux liability [147]. Transporter-aware modeling, combined with PBPK frameworks, allows permeability to be interpreted as a balance of influx and clearance rather than a static property [602,603].
AI-guided polypharmacology further expands this view [604,605]. Network-level profiling distinguishes harmful off target promiscuity from coordinated multi target engagement, enabling rational exploitation of pleiotropic mechanisms that are often intrinsic to natural products and CNS therapeutics [606,607].
12.2. Multi-Objective Formulation Optimization
Multi objective optimization reframes formulation design as a data driven negotiation between competing constraints [608]. Machine learning models trained on design of experiment data now predict how size, zeta potential, and drug loading jointly shape potency, exposure, stability, and manufacturability [609,610]. Rather than chasing a single optimum, Bayesian and evolutionary algorithms explore Pareto fronts, revealing trade-offs that are invisible to one-factor-at-a-time approaches [608,611]. In practice, this enables probabilistic design spaces where acceptable formulations are defined by balanced desirability rather than maximal performance [609]. Such frameworks accelerate iteration, reduce experimental burden, and align early formulation choices with downstream safety and GMP feasibility [609,610].
12.3. PBPK/PKPD and Digital Twins
Physiologically based pharmacokinetic modeling has become a cornerstone for forecasting CNS exposure and BBB penetration in silico [603,612]. Modern CNS PBPK platforms resolve regional brain compartments, passive permeability, and active efflux, allowing human predictions to be extrapolated from limited preclinical or in vitro data [612,613]. Coupling these frameworks to PK PD models refines dose response by linking brain time courses to target engagement and effect kinetics, enabling virtual dose fractionation before first exposure [602]. Digital twin concepts extend this logic further [614]. By integrating PBPK, machine learning derived BBB parameters, and virtual populations, individualized predictions of permeability and response become feasible [612,614]. We synthesize the major gaps and convert them into concrete, testable strategies.
13. Research Gaps and Concrete Strategies
13.1. Standardized Human-Relevant PK Endpoints
A clear gap is the lack of standardized, human-relevant CNS PK endpoints that translate cleanly from animals to early clinical trials. Across conceptual surveys and candidate-selection frameworks, Kp,uu,brain repeatedly emerges as the most defensible common currency because it captures BBB transport and binding within an unbound metric. Yet human Kp,uu,brain data remain sparse, and many programs still rely on total brain concentrations or non-comparable surrogates. A concrete strategy is universal adoption of Kp,uu,brain, paired with PBPK-informed target-site exposure ratios that connect unbound concentrations to in vivo IC50-class benchmarks and pharmacodynamic effect.
Methodologically, the field needs harmonization of how Kp,uu,brain and related endpoints are measured in humans. Combined PET plus microdialysis can convert imaging signal into unbound interstitial exposure, while mechanistic PBPK platforms can reconcile compartmental and spatial heterogeneity when CSF is unreliable. Standardized PET endpoints for exposure and engagement, together with aligned CSF metrics such as AUC, Cmax, and Ctrough normalized to potency, would make datasets interoperable. Regulatory pressure could then drive CNS drug development toward quantifiable, auditable endpoints, replacing subjective “brain penetration” claims with decision-grade measures.
13.2. Humanized BBB Models with Disease Fidelity
Humanized BBB models still fall short when they trade biological realism for convenience. A priority gap is scaling iPSC derived BBB organoids, spheroids, and self assembled microvessels into flow conditioned systems that reproduce shear, polarization, and transport kinetics seen in vivo. Microfluidic BBB-on-a-chip platforms and perfusable 3D microvessels now achieve low paracellular permeability and strong junctional phenotypes, yet protocol and donor variability remain major sources of noise [615].
Concrete strategy: build disease fidelity into the neurovascular unit, not just the endothelium. Co culture designs that incorporate pericytes, astrocytes, neurons, and microglia, ideally from patient or isogenic iPSC backgrounds, capture inter-individual differences in maturation, transporter function, immune cell trafficking, and barrier breakdown [616]. The field then needs rigorous validation against human in vivo benchmarks, such as PET permeability proxies, CSF-to-plasma relationships, and clinical drug exposure patterns, so these models stop being “pretty biology” and start reducing translational attrition.
13.3. Prodrug Translation Playbook
A translational prodrug playbook has to start with transporter rigor, not transporter “positive” checkboxes. For carrier mediated designs, uptake should be quantified with Km and Vmax under physiologic substrate conditions, then stress tested for competition with endogenous ligands and likely co medications [337]. Pharmacoproteomic transporter expression can anchor these kinetics to realistic barrier capacity, while time course uptake modeling helps separate true transported substrates from high affinity binders that never meaningfully cross.
Next, de-risk activation and safety in parallel. Cleavage mapping should quantify where and how fast the promoiety is removed across plasma, liver, brain microvessels, parenchyma, and disease relevant compartments to enforce brain first activation and avoid premature systemic unmasking. Off target profiling can be expanded beyond cell lines using tissue thermal proteome profiling or ABPP style probes across organ panels [617]. Go/no go rules then become tangible: require a brain unbound exposure gain, a defined brain to plasma activation ratio, no dominant peripheral off target signals, and a pharmacodynamic effect that tracks brain exposure in 3D GBM models or organotypic brain slices.
13.4. Long-Term Safety and Immunogenicity Registries
Long-term safety remains a blind spot when CNS trials end at symptom curves rather than at biology and latency. A concrete strategy is post-trial registries that follow participants for years, capturing delayed toxicities, immune responses, and neuroinflammation through linked EHR and claims data, structured adverse event reports, and longitudinal fluid or imaging biomarkers such as GFAP, YKL-40, sTREM2, or neuroinflammatory PET [618]. These registries should interlock with pharmacovigilance databases via standardized, FAIR data models and privacy preserving linkage, so signals can be detected, replicated, and risk managed across systems. Biologics and cell or gene therapies offer the template: mandated long follow up, harmonized reporting, and global registries that turn rare late events into quantifiable risk [618].
13.5. Manufacturability and QC for Complex Carriers
A central translational gap for complex nanocarriers is that manufacturability and QC often lag behind formulation ingenuity. QbD should be treated as the organizing logic, starting with a clear QTPP and mapping CQAs to CMAs and CPPs so that a justified design space and control strategy survive scale-up [573]. Yet non-linear formulation process couplings and raw material drift still drive lot-to-lot variability, especially for surface functionalized systems where small chemistry changes reshape size, charge, corona, and bioactivity. Concrete fixes include PAT-enabled real-time monitoring (inline or online size sensing, turbidity, spectroscopy, multivariate analytics), semi-continuous or continuous lines, and tighter incoming material specifications plus stage-gated in-process controls to secure reproducible release quality [619].
13.6. Clinical Trial Design Upgrades
Clinical trials for BBB therapeutics need design upgrades that treat BBB heterogeneity as a core covariate rather than background noise. Adaptive platform, basket, and window-of-opportunity approaches can rapidly prune futile delivery strategies while learning which BBB modulation, timing, and dosing actually shift brain exposure [620]. Pair this with enrichment: stratify participants by BBB integrity or permeability status using DCE MRI, PET based uptake metrics, or fluid markers reflecting barrier leakage and clearance kinetics.
Endpoints should prove target engagement, not just clinical change [621]. Imaging derived cerebral PK, longitudinal PD imaging, and permeability limited PBPK models can define exposure response relationships and justify go or no-go decisions. To satisfy regulators for high-cost, high-complexity products, adaptations and estimands must be pre specified, bias controlled, and CMC and companion diagnostics aligned early. Finally, we outline what will likely materialize soon and what needs deeper tech maturation,
14. Roadmap: Near-Term vs Long-Term
14.1. Near-Term (2–4 Years)
In the near term (2 to 4 years), the most “deployable” polyphenol programs will likely be ligand targeted PLGA nanoparticles and liposomes carrying resveratrol and curcumin, chosen because their safety narratives are mature while formulation science can add real value. Practical targets include transferrin or RVG-style ligands for BBB facing delivery, plus dual loading to exploit complementary redox and anti-inflammatory pharmacology. The translation gate is not efficacy hype, it is reproducible particle size, drug loading, and stability under scalable unit operations [162].
Intranasal mucoadhesive nanoemulsions and nanoemulgels are even closer to early-phase readiness because dosing ergonomics can be engineered into a sprayable, residence-time-extending product. Chitosan-coated or thermotriggered in situ gel formats already map nicely onto trial-friendly endpoints: nasal tolerability, systemic exposure, and nose-to-brain PK surrogates such as regional brain concentrations in imaging-rich substudies or CSF exposure when justified [178]. Here, the CMC story must be crisp: droplet size distribution, rheology, spray plume metrics, and preservative compatibility.
LAT1 anchored prodrugs are the “biology first” option. The appeal is a validated transporter with design rules for aromatic promoieties and linker choices, enabling higher brain exposure with lower peripheral burden [336,381]. Near term success will come from leveraging known promoiety scaffolds, building a screening cascade that confirms LAT1 affinity, bioconversion kinetics, and intra brain distribution, then anchoring dose selection to target engagement readouts in neurons and glia.
Focused ultrasound-assisted regional delivery fits the same horizon when paired with drugs that already have a clinical path, such as chemotherapy for glioma margins or neuroprotectives with clean systemic safety profiles. It offers a controllable exposure window, but only if trials pre-specify imaging-based BBB opening, local PK confirmation, and safety monitoring that regulators recognize [622]. Across all these tracks, feasibility wins: scalable manufacturing, release tests that predict performance, and endpoints that prove delivery plus mechanism, not just symptomatic change [623] (Figure 4).
14.2. Longer-Term (5–10+ Years)
Looking 5 to 10 years out, exosome mimetics and hybrid vesicles could become the “biomimetic workhorses” of brain delivery, but only if GMP-scale-up stops being artisanal. The roadmap points to programmed assembly, extrusion, and liposome fusion approaches, followed by process intensification via microfluidics and bottom-up manufacturing to reduce heterogeneity while improving yield [624]. Cost control will hinge on standardized membrane sourcing, robust cargo-loading metrics, and shelf-stable storage protocols.
In parallel, AI-designed multi target nano prodrugs may unlock rational polypharmacology, not by adding more ligands, but by learning which combinations actually cooperate at the BBB. The key upgrade is transporter awareness in silico: prediction stacks that integrate passive permeability, efflux risk, and carrier or promoiety interactions with uptake transporters [625]. Regulatory credibility will depend on curated datasets, auditable models, and prospective validation rather than retrospective fits.
Remote triggered release platforms promise precision with fewer systemic side effects. Magnetic fields [626].
Finally, patient specific BBB digital twins could connect PBPK and PKPD to biomarker-informed adaptation, turning trial dosing into a learning loop [625]. That vision depends on humanized BBB models with functional readouts, such as real-time TEER, longitudinal safety registries for complex nanomedicines, and early alignment with regulators on what counts as validated exposure and engagement evidence [627] (Figure 4).
15. Clinical Applications and Translational Implications
The concepts synthesized in this study have direct clinical relevance for the development of next-generation neurotherapeutics targeting disorders with high unmet medical need, including neurodegenerative diseases, neuropsychiatric conditions, epilepsy, and brain tumors [628]. By systematically linking the physicochemical limitations of plant-derived compounds to rational delivery solutions—such as nanocarriers, transporter-targeted prodrugs, intranasal administration, and device-enabled BBB modulation—this framework provides actionable guidance for improving CNS drug exposure where conventional pharmacotherapy has failed [4]. In clinical contexts characterized by multifactorial pathophysiology, such as Alzheimer’s disease, Parkinson’s disease, depression, and chronic pain, phytochemicals with pleiotropic anti-inflammatory, antioxidant, and neuromodulatory actions may offer therapeutic advantages if reliable brain delivery can be achieved[629].
From a translational standpoint, the study supports a shift away from empiric compound selection toward delivery-first clinical development, in which candidate molecules are paired early with route, carrier, or prodrug strategies to achieve decision-grade CNS exposure [67]. Clinically, this approach may enable dose reduction, improved safety margins, and more predictable pharmacokinetics, particularly in vulnerable populations such as older adults or patients receiving polypharmacy [630]. Furthermore, the discussed platforms—especially intranasal delivery and focused ultrasound–mediated BBB opening—offer opportunities for region-specific or noninvasive treatment paradigms, which are increasingly relevant in precision neurology and psychiatry [631]. Collectively, these insights inform the design of early-phase clinical trials, guide biomarker and endpoint selection, and support regulatory-aligned translation of plant-derived neurotherapeutics from bench to bedside [632].
15. Conclusion and Translational Implications
RMT targeting via. Intranasal delivery enables potent, stable payloads to exploit the nose-to-brain pathways [155]. Focused ultrasound with microbubbles provides a reversible, local window of BBB opening [633]. Exosome mimetics and nano prodrugs stay promising but remain unproven. As authors, we argue that translation will speed up once Kp,uu,brain centric workflows become the common language across teams. The take home message is simple: treat delivery as a coupled system where chemistry, carrier, route, and measurement are co designed. Start with a liability map for each phytochemical, then choose the simplest strategy that can raise unbound brain exposure while meeting safety and chemistry, manufacturing, and controls constraints. Report fu,brain, unbound plasma fractions, and Kp,uu,brain, then connect them to target engagement, imaging, and functional outcomes. Future research should deliver head-to-head platform comparisons on identical payloads, validate human-relevant BBB models against in vivo benchmarks, and build PBPK guided dose projections that survive species shifts. Methodologically, the field also needs harmonized critical quality attributes and longitudinal safety panels that capture complement activation, microglial priming, and vascular repair. Done right, these principles can accelerate barrier-limited therapeutics far beyond phytochemicals.
Author Contributions
Conceptualization, M.T. and S.M.B.; methodology, A.C.A., E.L.G., V.E.V., R.S.A.H., R.A.G.; investigation; writing—original draft preparation, M.T., V.C.S.C., C.M.G., E.S.B.M.P., A.C.A.C.; writing—review and editing, M.T., V.E.V., S.M.B; visualization, M.T.; supervision, M.T. and S.M.B.; project administration, M.T. and S.M.B.; funding acquisition, M.T. and S.M.B. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ABC | ATP-binding cassette |
| ApoE | apolipoprotein E |
| AUC | area under the curve |
| BBB | blood-brain barrier |
| BCRP | breast cancer resistance protein |
| CBD | cannabidiol |
| CED | convection-enhanced delivery |
| CMC | chemistry, manufacturing, and controls |
| CMT | carrier-mediated transport |
| CNS | central nervous system |
| CPP | cell-penetrating peptide |
| CQA | critical quality attribute |
| CSF | cerebrospinal fluid |
| CYP | cytochrome P450 |
| DOI | digital object identifier |
| EVS | extracellular vesicles |
| FUS | focused ultrasound |
| GDNF | glial-derived neurotrophic factor |
| GLUT1 | glucose transporter 1 |
| GMP | good manufacturing practice |
| IPSC | induced pluripotent stem cell |
| IR | insulin receptor |
| ISF | interstitial fluid |
| KM | michaelis constant |
| KP | uu, unbound brain-to-plasma partition coefficient |
| KP,BRAIN | brain-to-plasma partition coefficient |
| KP,UU | unbound brain-to-plasma partition coefficient |
| K_D | dissociation constant |
| K_P,BRAIN | brain-to-plasma partition coefficient |
| K_P,UU | unbound brain-to-plasma partition coefficient |
| LAT1 | large neutral amino acid transporter 1 |
| LD | linear dichroism |
| LDL | low-density lipoprotein |
| LDLR | low-density lipoprotein receptor |
| LOGD | distribution coefficient |
| LRP1 | low-density lipoprotein receptor-related protein 1 |
| MCT1 | monocarboxylate transporter 1 |
| MCTS | monocarboxylate transporters |
| MRI | magnetic resonance imaging |
| MRPS | multidrug resistance-associated proteins |
| NHP | non-human primate |
| NLCS | nanostructured lipid carriers |
| NPS | nanoparticles |
| NR | not reported |
| NVU | neurovascular unit |
| P-gp | P-glycoprotein |
| PAMAM | poly(amidoamine) |
| PAT | process analytical technology |
| PBPK | physiologically based pharmacokinetic |
| PCD | passive cavitation detection |
| PD | pharmacodynamics |
| PDEVS | plant-derived extracellular vesicles |
| PDI | polydispersity index |
| PEG | polyethylene glycol |
| PET | positron emission tomography |
| PK | pharmacokinetics |
| PKA | acid dissociation constant |
| PKPD | pharmacokinetic-pharmacodynamic |
| PLGA | poly(lactic-co-glycolic acid) |
| QBD | quality by design |
| QTPP | quality target product profile |
| RESVERATROL/CURCUMIN | including dual loading) gated by reproducible size/PDI, loading, and stability under scalable unit operations |
| RMT | receptor-mediated transcytosis |
| RVG | rabies virus glycoprotein |
| SIRNA | small interfering RNA |
| SLCS | solute carrier transporters |
| SLNS | solid lipid nanoparticles |
| SULT | sulfotransferases |
| TAT | trans-activator of transcription |
| TEER | transendothelial electrical resistance |
| TF | transferrin |
| TFR | transferrin receptor |
| THC | Δ9-tetrahydrocannabinol |
| UGT | UDP-glucuronosyltransferases |
| VEGF | vascular endothelial growth factor |
| VMAX | maximum transport rate |
References
- Global, regional, and national burden of disorders affecting the nervous system, 1990-2021: a systematic analysis for the Global Burden of Disease Study 2021. Lancet Neurol 2024, 23, 344–381. [CrossRef] [PubMed]
- Arias, D.; Saxena, S.; Verguet, S. Quantifying the global burden of mental disorders and their economic value. EClinicalMedicine 2022, 54, 101675. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M. Parkinson's Disease: Bridging Gaps, Building Biomarkers, and Reimagining Clinical Translation. Cells 2025, 14. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Chen, Q.; Chen, X.; Han, F.; Chen, Z.; Wang, Y. The blood-brain barrier: structure, regulation, and drug delivery. Signal Transduct Target Ther 2023, 8, 217. [Google Scholar] [CrossRef]
- Gribkoff, V.K.; Kaczmarek, L.K. The need for new approaches in CNS drug discovery: Why drugs have failed, and what can be done to improve outcomes. Neuropharmacology 2017, 120, 11–19. [Google Scholar] [CrossRef]
- Howe, J.R.t.; Bear, M.F.; Golshani, P.; Klann, E.; Lipton, S.A.; Mucke, L.; Sahin, M.; Silva, A.J. The mouse as a model for neuropsychiatric drug development. Curr Biol 2018, 28, R909–r914. [Google Scholar] [CrossRef]
- Tanaka, M. From Monoamines to Systems Psychiatry: Rewiring Depression Science and Care (1960s–2025). Biomedicines 2025, 14, 35. [Google Scholar] [CrossRef]
- Naoi, M.; Wu, Y.; Maruyama, W.; Shamoto-Nagai, M. Phytochemicals Modulate Biosynthesis and Function of Serotonin, Dopamine, and Norepinephrine for Treatment of Monoamine Neurotransmission-Related Psychiatric Diseases. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Chen, X.; Xu, D.; Yu, J.; Song, X.J.; Li, X.; Cui, Y.L. Tryptophan Metabolism Disorder-Triggered Diseases, Mechanisms, and Therapeutic Strategies: A Scientometric Review. Nutrients 2024, 16. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, M.; Chen, X.; Zhang, R.; Le, A.; Hong, M.; Zhang, Y.; Jia, L.; Zang, W.; Jiang, C.; et al. Tryptophan Metabolism in Central Nervous System Diseases: Pathophysiology and Potential Therapeutic Strategies. Aging Dis 2023, 14, 858–878. [Google Scholar] [CrossRef]
- Szabó, Á.; Galla, Z.; Spekker, E.; Martos, D.; Szűcs, M.; Fejes-Szabó, A.; Fehér, Á.; Takeda, K.; Ozaki, K.; Inoue, H.; et al. Behavioral Balance in Tryptophan Turmoil: Regional Metabolic Rewiring in Kynurenine Aminotransferase II Knockout Mice. Cells 2025, 14. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Z.; Li, A.; Liu, R.; Yang, H.; Xia, X. The Phytochemical Potential for Brain Disease Therapy and the Possible Nanodelivery Solutions for Brain Access. Front Oncol 2022, 12, 936054. [Google Scholar] [CrossRef] [PubMed]
- Mishra, K.; Rana, R.; Tripathi, S.; Siddiqui, S.; Yadav, P.K.; Yadav, P.N.; Chourasia, M.K. Recent Advancements in Nanocarrier-assisted Brain Delivery of Phytochemicals Against Neurological Diseases. Neurochem Res 2023, 48, 2936–2968. [Google Scholar] [CrossRef] [PubMed]
- Rassu, G.; Sorrenti, M.; Catenacci, L.; Pavan, B.; Ferraro, L.; Gavini, E.; Bonferoni, M.C.; Giunchedi, P.; Dalpiaz, A. Conjugation, Prodrug, and Co-Administration Strategies in Support of Nanotechnologies to Improve the Therapeutic Efficacy of Phytochemicals in the Central Nervous System. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Chehadi, A.C.; Pereira de Lima, E.; Detregiachi, C.R.P.; Santos de Argollo Haber, R.; Catharin, V.; Fornari Laurindo, L.; Engracia Valenti, V.; Machado Galhardi, C.; Tanaka, M.; Maria Barbalho, S. Harnessing Dietary Tryptophan: Bridging the Gap Between Neurobiology and Psychiatry in Depression Management. Int J Mol Sci 2026, 27. [Google Scholar] [CrossRef]
- Endo, H.; Ogasawara, M.; Tega, Y.; Kubo, Y.; Hosoya, K.I.; Akanuma, S.I. Upregulation of P-Glycoprotein and Breast Cancer Resistance Protein Activity in Newly Developed in Vitro Rat Blood-Brain Barrier Spheroids Using Advanced Glycation End-Products. Biol Pharm Bull 2024, 47, 1893–1903. [Google Scholar] [CrossRef]
- Strazielle, N.; Ghersi-Egea, J.F. Factors affecting delivery of antiviral drugs to the brain. Rev Med Virol 2005, 15, 105–133. [Google Scholar] [CrossRef]
- Waterhouse, R.N. Determination of lipophilicity and its use as a predictor of blood-brain barrier penetration of molecular imaging agents. Mol Imaging Biol 2003, 5, 376–389. [Google Scholar] [CrossRef]
- Ayaz, M.; Mosa, O.F.; Nawaz, A.; Hamdoon, A.A.E.; Elkhalifa, M.E.M.; Sadiq, A.; Ullah, F.; Ahmed, A.; Kabra, A.; Khan, H.; et al. Neuroprotective potentials of Lead phytochemicals against Alzheimer's disease with focus on oxidative stress-mediated signaling pathways: Pharmacokinetic challenges, target specificity, clinical trials and future perspectives. Phytomedicine 2024, 124, 155272. [Google Scholar] [CrossRef]
- Lehoczki, A.; Fekete, M.; Jarecsny, T.; Zábó, V.; Szappanos, Á.; Csípő, T.; Lipécz, Á.; Major, D.; Fazekas-Pongor, V.; Varga, P.; et al. The Neuroprotective Role of Curcumin: From Molecular Pathways to Clinical Translation-A Narrative Review. Nutrients 2025, 17. [Google Scholar] [CrossRef]
- Nunes, Y.C.; Mendes, N.M.; Pereira de Lima, E.; Chehadi, A.C.; Lamas, C.B.; Haber, J.F.S.; Dos Santos Bueno, M.; Araújo, A.C.; Catharin, V.C.S.; Detregiachi, C.R.P.; et al. Curcumin: A Golden Approach to Healthy Aging: A Systematic Review of the Evidence. Nutrients 2024, 16. [Google Scholar] [CrossRef]
- Laurindo, L.F.; de Carvalho, G.M.; de Oliveira Zanuso, B.; Figueira, M.E.; Direito, R.; de Alvares Goulart, R.; Buglio, D.S.; Barbalho, S.M. Curcumin-Based Nanomedicines in the Treatment of Inflammatory and Immunomodulated Diseases: An Evidence-Based Comprehensive Review. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Marton, L.T.; Barbalho, S.M.; Sloan, K.P.; Sloan, L.A.; Goulart, R.A.; Araújo, A.C.; Bechara, M.D. Curcumin, autoimmune and inflammatory diseases: going beyond conventional therapy - a systematic review. Crit Rev Food Sci Nutr 2022, 62, 2140–2157. [Google Scholar] [CrossRef] [PubMed]
- Matias, J.N.; Achete, G.; Campanari, G.; Guiguer É, L.; Araújo, A.C.; Buglio, D.S.; Barbalho, S.M. A systematic review of the antidepressant effects of curcumin: Beyond monoamines theory. Aust N Z J Psychiatry 2021, 55, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Kaspute, G.; Ramanavicius, A.; Prentice, U. Natural drug delivery systems for the treatment of neurodegenerative diseases. Mol Biol Rep 2025, 52, 217. [Google Scholar] [CrossRef]
- Sánchez-Martínez, J.D.; Valdés, A.; Gallego, R.; Suárez-Montenegro, Z.J.; Alarcón, M.; Ibañez, E.; Alvarez-Rivera, G.; Cifuentes, A. Blood-Brain Barrier Permeability Study of Potential Neuroprotective Compounds Recovered From Plants and Agri-Food by-Products. Front Nutr 2022, 9, 924596. [Google Scholar] [CrossRef]
- Shimazu, R.; Anada, M.; Miyaguchi, A.; Nomi, Y.; Matsumoto, H. Evaluation of Blood-Brain Barrier Permeability of Polyphenols, Anthocyanins, and Their Metabolites. J Agric Food Chem 2021, 69, 11676–11686. [Google Scholar] [CrossRef]
- Musa, I.; Rotaru-Zavaleanu, A.D.; Sfredel, V.; Aldea, M.; Gresita, A.; Glavan, D.G. Post-Stroke Recovery: A Review of Hydrogel-Based Phytochemical Delivery Systems. Gels 2025, 11. [Google Scholar] [CrossRef]
- Tanaka, M.; Vécsei, L. From Microbial Switches to Metabolic Sensors: Rewiring the Gut-Brain Kynurenine Circuit. Biomedicines 2025, 13. [Google Scholar] [CrossRef]
- Figueiredo Godoy, A.C.; Frota, F.F.; Araújo, L.P.; Valenti, V.E.; Pereira, E.; Detregiachi, C.R.P.; Galhardi, C.M.; Caracio, F.C.; Haber, R.S.A.; Fornari Laurindo, L.; et al. Neuroinflammation and Natural Antidepressants: Balancing Fire with Flora. Biomedicines 2025, 13. [Google Scholar] [CrossRef]
- Karad, V.; Gupta, G.L. Phytochemicals encouraging neurotrophic pathways: brain-derived neurotrophic factors as molecular targets in depression. Naunyn Schmiedebergs Arch Pharmacol 2025, 398, 15075–15094. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M. Overcoming the Blood-Brain Barrier: Advanced Strategies in Targeted Drug Delivery for Neurodegenerative Diseases. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- Zha, S.; Liu, H.; Li, H.; Li, H.; Wong, K.L.; All, A.H. Functionalized Nanomaterials Capable of Crossing the Blood-Brain Barrier. ACS Nano 2024, 18, 1820–1845. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Ali, S.A.; Kumar, M.; Jahan, I.; Hak, J. Emerging Strategies for Targeted Drug Delivery across the Blood–Brain Barrier in Neurological Disorder. Current Pharmaceutical Research 2025, 1–14. [CrossRef]
- Zou, L.; Chien, H.C.; Pade, D.; Li, Y.; Nguyen, M.; Bhamidipati, R.K.; Wang, Z.; Enogieru, O.J.; Wahlstrom, J. Considerations in K(p,uu,brain)-based Strategy for Selecting CNS-targeted Drug Candidates with Sufficient Target Coverage and Substantial Pharmacodynamic Effect. Aaps j 2025, 27, 52. [Google Scholar] [CrossRef]
- Gonen, O.M.; Porter, T.; Wang, B.; Xue, F.; Ma, Y.; Song, L.; Sun, P.; Fan, W.; Shen, Y. Safety, Pharmacokinetics and Target Engagement of a Novel Brain Penetrant RIPK1 Inhibitor (SIR9900) in Healthy Adults and Elderly Participants. Clin Transl Sci 2025, 18, e70151. [Google Scholar] [CrossRef]
- Tanaka, M.; Szatmári, I.; Vécsei, L. Quinoline Quest: Kynurenic Acid Strategies for Next-Generation Therapeutics via Rational Drug Design. Pharmaceuticals (Basel) 2025, 18. [Google Scholar] [CrossRef]
- Gong, Y.; Wu, M.; Huang, Y.; He, X.; Yuan, J.; Dang, B. Research developments in the neurovascular unit and the blood-brain barrier (Review). Biomed Rep 2025, 22, 88. [Google Scholar] [CrossRef]
- McConnell, H.L.; Mishra, A. Cells of the Blood-Brain Barrier: An Overview of the Neurovascular Unit in Health and Disease. Methods Mol Biol 2022, 2492, 3–24. [Google Scholar] [CrossRef]
- Schaeffer, S.; Iadecola, C. Revisiting the neurovascular unit. Nat Neurosci 2021, 24, 1198–1209. [Google Scholar] [CrossRef]
- Gong, Y.; Wu, M.; Huang, Y.; He, X.; Yuan, J.; Dang, B. Research developments in the neurovascular unit and the blood-brain barrier. Biomedical Reports 2025, 22, 88. [Google Scholar] [CrossRef] [PubMed]
- Divecha, Y.A.; Rampes, S.; Tromp, S.; Boyanova, S.T.; Fleckney, A.; Fidanboylu, M.; Thomas, S.A. The microcirculation, the blood-brain barrier, and the neurovascular unit in health and Alzheimer disease: The aberrant pericyte is a central player. Pharmacol Rev 2025, 77, 100052. [Google Scholar] [CrossRef] [PubMed]
- Kugler, E.C.; Greenwood, J.; MacDonald, R.B. The "Neuro-Glial-Vascular" Unit: The Role of Glia in Neurovascular Unit Formation and Dysfunction. Front Cell Dev Biol 2021, 9, 732820. [Google Scholar] [CrossRef] [PubMed]
- Sasson, E.; Anzi, S.; Bell, B.; Yakovian, O.; Zorsky, M.; Deutsch, U.; Engelhardt, B.; Sherman, E.; Vatine, G.; Dzikowski, R. Nano-scale architecture of blood-brain barrier tight-junctions. Elife 2021, 10, e63253. [Google Scholar] [CrossRef]
- Alluri, H.; Peddaboina, C.S.; Tharakan, B. Evaluation of Tight Junction Integrity in Brain Endothelial Cells Using Confocal Microscopy. Methods Mol Biol 2024, 2711, 257–262. [Google Scholar] [CrossRef]
- Gowrikumar, S.; Tarudji, A.; McDonald, B.Z.; Balusa, S.S.; Kievit, F.M.; Dhawan, P. Claudin-1 impairs blood-brain barrier by downregulating endothelial junctional proteins in traumatic brain injury. Tissue Barriers 2025, 13, 2470482. [Google Scholar] [CrossRef]
- Maiuolo, J.; Gliozzi, M.; Musolino, V.; Scicchitano, M.; Carresi, C.; Scarano, F.; Bosco, F.; Nucera, S.; Ruga, S.; Zito, M.C.; et al. The "Frail" Brain Blood Barrier in Neurodegenerative Diseases: Role of Early Disruption of Endothelial Cell-to-Cell Connections. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Potjewyd, G.; Moxon, S.; Wang, T.; Domingos, M.; Hooper, N.M. Tissue Engineering 3D Neurovascular Units: A Biomaterials and Bioprinting Perspective. Trends Biotechnol 2018, 36, 457–472. [Google Scholar] [CrossRef]
- Mäe, M.A.; He, L.; Nordling, S.; Vazquez-Liebanas, E.; Nahar, K.; Jung, B.; Li, X.; Tan, B.C.; Chin Foo, J.; Cazenave-Gassiot, A.; et al. Single-Cell Analysis of Blood-Brain Barrier Response to Pericyte Loss. Circ Res 2021, 128, e46–e62. [Google Scholar] [CrossRef]
- Guérit, S.; Fidan, E.; Macas, J.; Czupalla, C.J.; Figueiredo, R.; Vijikumar, A.; Yalcin, B.H.; Thom, S.; Winter, P.; Gerhardt, H.; et al. Astrocyte-derived Wnt growth factors are required for endothelial blood-brain barrier maintenance. Prog Neurobiol 2021, 199, 101937. [Google Scholar] [CrossRef]
- Tanaka, M. Neurogenesis and Neuroinflammation in Dialogue: Mapping Gaps, Modulating Microglia, Rewiring Aging. Cells 2026, 15. [Google Scholar] [CrossRef]
- Reed, M.J.; Damodarasamy, M.; Banks, W.A. The extracellular matrix of the blood-brain barrier: structural and functional roles in health, aging, and Alzheimer's disease. Tissue Barriers 2019, 7, 1651157. [Google Scholar] [CrossRef]
- Ding, L.; Kshirsagar, P.; Agrawal, P.; Murry, D.J. Crossing the Blood-Brain Barrier: Innovations in Receptor- and Transporter-Mediated Transcytosis Strategies. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- Terstappen, G.C.; Meyer, A.H.; Bell, R.D.; Zhang, W. Strategies for delivering therapeutics across the blood-brain barrier. Nat Rev Drug Discov 2021, 20, 362–383. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Transport of small molecules through the blood-brain barrier: biology and methodology. Advanced drug delivery reviews 1995, 15, 5–36. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Wu, J.; Xie, Y.; Kim, S.; Michelhaugh, S.; Jiang, J.; Mittal, S.; Sanai, N.; Li, J. Protein Expression and Functional Relevance of Efflux and Uptake Drug Transporters at the Blood-Brain Barrier of Human Brain and Glioblastoma. Clin Pharmacol Ther 2020, 107, 1116–1127. [Google Scholar] [CrossRef]
- Latif, S.; Kang, Y.S. Blood-Brain Barrier Solute Carrier Transporters and Motor Neuron Disease. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Tashima, T. Smart Strategies for Therapeutic Agent Delivery into Brain across the Blood-Brain Barrier Using Receptor-Mediated Transcytosis. Chem Pharm Bull (Tokyo) 2020, 68, 316–325. [Google Scholar] [CrossRef]
- Stanimirovic, D.B.; Sandhu, J.K.; Costain, W.J. Emerging Technologies for Delivery of Biotherapeutics and Gene Therapy Across the Blood-Brain Barrier. BioDrugs 2018, 32, 547–559. [Google Scholar] [CrossRef]
- Kucharz, K.; Kristensen, K.; Johnsen, K.B.; Lund, M.A.; Lønstrup, M.; Moos, T.; Andresen, T.L.; Lauritzen, M.J. Post-capillary venules are the key locus for transcytosis-mediated brain delivery of therapeutic nanoparticles. Nat Commun 2021, 12, 4121. [Google Scholar] [CrossRef]
- Hersh, A.M.; Alomari, S.; Tyler, B.M. Crossing the Blood-Brain Barrier: Advances in Nanoparticle Technology for Drug Delivery in Neuro-Oncology. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Lochhead, J.J.; Yang, J.; Ronaldson, P.T.; Davis, T.P. Structure, Function, and Regulation of the Blood-Brain Barrier Tight Junction in Central Nervous System Disorders. Front Physiol 2020, 11, 914. [Google Scholar] [CrossRef]
- Keaney, J.; Walsh, D.M.; O'Malley, T.; Hudson, N.; Crosbie, D.E.; Loftus, T.; Sheehan, F.; McDaid, J.; Humphries, M.M.; Callanan, J.J.; et al. Autoregulated paracellular clearance of amyloid-β across the blood-brain barrier. Sci Adv 2015, 1, e1500472. [Google Scholar] [CrossRef] [PubMed]
- Nozohouri, E.; Noorani, B.; Patel, D.; Ahn, Y.; Zoubi, S.; Bickel, U. Assessing blood-brain barrier (BBB) integrity in an Alzheimer's disease mouse model: is the BBB globally or locally disrupted? Fluids Barriers CNS 2025, 22, 79. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, F.M.G.; Markert, G.; Deutsch, G.; Antonara, M.; Faaij, N.; Bartelink, I.; Noske, D.; Vandertop, W.P.; Bender, A.; Westerman, B.A. Explaining Blood-Brain Barrier Permeability of Small Molecules by Integrated Analysis of Different Transport Mechanisms. J Med Chem 2023, 66, 7253–7267. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.; Barras, A.; Boukherroub, R.; Tseng, C.L.; Devos, D.; Burnouf, T.; Neuhaus, W.; Szunerits, S. Enhancing paracellular and transcellular permeability using nanotechnological approaches for the treatment of brain and retinal diseases. Nanoscale Horiz 2023, 9, 14–43. [Google Scholar] [CrossRef]
- Nájera-Maldonado, L.; Parra-González, M.; Peralta-Cuevas, E.; Gutierrez-Onofre, A.J.; Garcia-Atutxa, I.; Villanueva-Flores, F. Cracking the Blood-Brain Barrier Code: Rational Nanomaterial Design for Next-Generation Neurological Therapies. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Smith, Q.R. Carrier-mediated transport to enhance drug delivery to brain. In Proceedings of the International Congress Series, 2005; pp. 63–74. [Google Scholar]
- Ozgür, B.; Puris, E.; Brachner, A.; Appelt-Menzel, A.; Oerter, S.; Balzer, V.; Holst, M.R.; Christiansen, R.F.; Hyldig, K.; Buckley, S.T.; et al. Characterization of an iPSC-based barrier model for blood-brain barrier investigations using the SBAD0201 stem cell line. Fluids Barriers CNS 2023, 20, 96. [Google Scholar] [CrossRef]
- Li, Y.; Liu, R.; Zhao, Z. Targeting Brain Drug Delivery with Macromolecules Through Receptor-Mediated Transcytosis. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Hervé, F.; Ghinea, N.; Scherrmann, J.M. CNS delivery via adsorptive transcytosis. Aaps j 2008, 10, 455–472. [Google Scholar] [CrossRef]
- Baghirov, H. Receptor-mediated transcytosis of macromolecules across the blood-brain barrier. Expert Opin Drug Deliv 2023, 20, 1699–1711. [Google Scholar] [CrossRef] [PubMed]
- Pfau, S.J.; Langen, U.H.; Fisher, T.M.; Prakash, I.; Nagpurwala, F.; Lozoya, R.A.; Lee, W.A.; Wu, Z.; Gu, C. Characteristics of blood-brain barrier heterogeneity between brain regions revealed by profiling vascular and perivascular cells. Nat Neurosci 2024, 27, 1892–1903. [Google Scholar] [CrossRef] [PubMed]
- Noumbissi, M.E.; Galasso, B.; Stins, M.F. Brain vascular heterogeneity: implications for disease pathogenesis and design of in vitro blood-brain barrier models. Fluids Barriers CNS 2018, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Beltran-Velasco, A.I.; Clemente-Suárez, V.J. Impact of Peripheral Inflammation on Blood-Brain Barrier Dysfunction and Its Role in Neurodegenerative Diseases. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Wilhelm, I.; Nyúl-Tóth, Á.; Suciu, M.; Hermenean, A.; Krizbai, I.A. Heterogeneity of the blood-brain barrier. Tissue Barriers 2016, 4, e1143544. [Google Scholar] [CrossRef]
- Ha, I.H.; Lim, C.; Kim, Y.; Moon, Y.; Han, S.H.; Moon, W.J. Regional Differences in Blood-Brain Barrier Permeability in Cognitively Normal Elderly Subjects: A Dynamic Contrast-Enhanced MRI-Based Study. Korean J Radiol 2021, 22, 1152–1162. [Google Scholar] [CrossRef]
- Schaffenrath, J.; Huang, S.F.; Wyss, T.; Delorenzi, M.; Keller, A. Characterization of the blood-brain barrier in genetically diverse laboratory mouse strains. Fluids Barriers CNS 2021, 18, 34. [Google Scholar] [CrossRef]
- Kurz, C.; Walker, L.; Rauchmann, B.S.; Perneczky, R. Dysfunction of the blood-brain barrier in Alzheimer's disease: Evidence from human studies. Neuropathol Appl Neurobiol 2022, 48, e12782. [Google Scholar] [CrossRef]
- Bravo-Ferrer, I.; Gaasdal-Bech, K.; Colvin, C.; Vaughan, H.J.; Moss, J.; Williams, A.; Díaz Castro, B. Multiregional blood-brain barrier phenotyping identifies the prefrontal cortex as the most vulnerable region to ageing in mice. Brain Commun 2025, 7, fcaf332. [Google Scholar] [CrossRef]
- Chen, T.; Dai, Y.; Hu, C.; Lin, Z.; Wang, S.; Yang, J.; Zeng, L.; Li, S.; Li, W. Cellular and molecular mechanisms of the blood-brain barrier dysfunction in neurodegenerative diseases. Fluids Barriers CNS 2024, 21, 60. [Google Scholar] [CrossRef]
- de Lima, E.P.; Tanaka, M.; Lamas, C.B.; Quesada, K.; Detregiachi, C.R.P.; Araújo, A.C.; Guiguer, E.L.; Catharin, V.; de Castro, M.V.M.; Junior, E.B.; et al. Vascular Impairment, Muscle Atrophy, and Cognitive Decline: Critical Age-Related Conditions. Biomedicines 2024, 12. [Google Scholar] [CrossRef]
- Bhattacharya, T.; Soares, G.; Chopra, H.; Rahman, M.M.; Hasan, Z.; Swain, S.S.; Cavalu, S. Applications of Phyto-Nanotechnology for the Treatment of Neurodegenerative Disorders. Materials (Basel) 2022, 15. [Google Scholar] [CrossRef] [PubMed]
- Kirit, E.; Gokce, C.; Altun, B.; Yilmazer, A. Nanotherapeutic Strategies for Overcoming the Blood-Brain Barrier: Applications in Disease Modeling and Drug Delivery. ACS Omega 2025, 10, 32606–32625. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Hu, D.; Gao, D.; Butch, C.J.; Wang, Y.; Zheng, H.; Sheng, Z. Recent advances of engineering cell membranes for nanomedicine delivery across the blood-brain barrier. J Nanobiotechnology 2025, 23, 493. [Google Scholar] [CrossRef] [PubMed]
- Pedder, J.H.; Sonabend, A.M.; Cearns, M.D.; Michael, B.D.; Zakaria, R.; Heimberger, A.B.; Jenkinson, M.D.; Dickens, D. Crossing the blood-brain barrier: emerging therapeutic strategies for neurological disease. Lancet Neurol 2025, 24, 246–260. [Google Scholar] [CrossRef]
- Dirir, A.M.; Ali, A.; Hachem, M. Recent Advancements in Lipid Nanoparticles-Based Phytoactives Delivery Systems for Neurodegenerative Diseases. Int J Nanomedicine 2025, 20, 10279–10300. [Google Scholar] [CrossRef]
- Kamath, A.P.; Nayak, P.G.; John, J.; Mutalik, S.; Balaraman, A.K.; Krishnadas, N. Revolutionizing neurotherapeutics: Nanocarriers unveiling the potential of phytochemicals in Alzheimer's disease. Neuropharmacology 2024, 259, 110096. [Google Scholar] [CrossRef]
- de Lima, E.P.; Laurindo, L.F.; Catharin, V.C.S.; Direito, R.; Tanaka, M.; Jasmin Santos German, I.; Lamas, C.B.; Guiguer, E.L.; Araújo, A.C.; Fiorini, A.M.R.; et al. Polyphenols, Alkaloids, and Terpenoids Against Neurodegeneration: Evaluating the Neuroprotective Effects of Phytocompounds Through a Comprehensive Review of the Current Evidence. Metabolites 2025, 15. [Google Scholar] [CrossRef]
- Laurindo, L.F.; Santos, A.; Carvalho, A.C.A.; Bechara, M.D.; Guiguer, E.L.; Goulart, R.A.; Vargas Sinatora, R.; Araújo, A.C.; Barbalho, S.M. Phytochemicals and Regulation of NF-kB in Inflammatory Bowel Diseases: An Overview of In Vitro and In Vivo Effects. Metabolites 2023, 13. [Google Scholar] [CrossRef]
- Buglio, D.S.; Marton, L.T.; Laurindo, L.F.; Guiguer, E.L.; Araújo, A.C.; Buchaim, R.L.; Goulart, R.A.; Rubira, C.J.; Barbalho, S.M. The Role of Resveratrol in Mild Cognitive Impairment and Alzheimer's Disease: A Systematic Review. J Med Food 2022, 25, 797–806. [Google Scholar] [CrossRef]
- Ciupei, D.; Colişar, A.; Leopold, L.; Stănilă, A.; Diaconeasa, Z.M. Polyphenols: From Classification to Therapeutic Potential and Bioavailability. Foods 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Dong, Y.; Wang, F.; Zhang, Y. Nanoformulations to Enhance the Bioavailability and Physiological Functions of Polyphenols. Molecules 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Grabska-Kobyłecka, I.; Szpakowski, P.; Król, A.; Książek-Winiarek, D.; Kobyłecki, A.; Głąbiński, A.; Nowak, D. Polyphenols and Their Impact on the Prevention of Neurodegenerative Diseases and Development. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Arias-Sánchez, R.A.; Torner, L.; Fenton Navarro, B. Polyphenols and Neurodegenerative Diseases: Potential Effects and Mechanisms of Neuroprotection. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Mamun, A.A.; Shao, C.; Geng, P.; Wang, S.; Xiao, J. Polyphenols Targeting NF-κB Pathway in Neurological Disorders: What We Know So Far? Int J Biol Sci 2024, 20, 1332–1355. [Google Scholar] [CrossRef]
- Kaluza, M.; Ksiazek-Winiarek, D.; Szpakowski, P.; Czpakowska, J.; Fijalkowska, J.; Glabinski, A. Polyphenols in the Central Nervous System: Cellular Effects and Liposomal Delivery Approaches. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Biswas, P.; Dey, D.; Biswas, P.K.; Rahaman, T.I.; Saha, S.; Parvez, A.; Khan, D.A.; Lily, N.J.; Saha, K.; Sohel, M.; et al. A Comprehensive Analysis and Anti-Cancer Activities of Quercetin in ROS-Mediated Cancer and Cancer Stem Cells. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Teng, H.; Chen, L. Polyphenols and bioavailability: an update. Crit Rev Food Sci Nutr 2019, 59, 2040–2051. [Google Scholar] [CrossRef]
- Williamson, G. Bioavailability of Food Polyphenols: Current State of Knowledge. Annu Rev Food Sci Technol 2025, 16, 315–332. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am J Clin Nutr 2005, 81, 230s–242s. [Google Scholar] [CrossRef]
- Hu, M.; Wu, B.; Liu, Z. Bioavailability of Polyphenols and Flavonoids in the Era of Precision Medicine. Mol Pharm 2017, 14, 2861–2863. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Hu, M. Natural polyphenol disposition via coupled metabolic pathways. Expert Opin Drug Metab Toxicol 2007, 3, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Kapetanovic, I.M.; Muzzio, M.; Huang, Z.; Thompson, T.N.; McCormick, D.L. Pharmacokinetics, oral bioavailability, and metabolic profile of resveratrol and its dimethylether analog, pterostilbene, in rats. Cancer Chemother Pharmacol 2011, 68, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, J.W.; Cong, R.; Park, J.S.; Nguyen, C.H.B.; Park, K.; Kang, K.; Shim, S.M. Exploring absorption indices for a variety of polyphenols through Caco-2 cell model: insights from permeability studies and principal component analysis. J Sci Food Agric 2025, 105, 6243–6253. [Google Scholar] [CrossRef]
- Martos, D.; Lőrinczi, B.; Szatmári, I.; Vécsei, L.; Tanaka, M. Decoupling Behavioral Domains via Kynurenic Acid Analog Optimization: Implications for Schizophrenia and Parkinson's Disease Therapeutics. Cells 2025, 14. [Google Scholar] [CrossRef]
- Dos Santos, T.C.; Gomes, T.M.; Pinto, B.A.S.; Camara, A.L.; Paes, A.M.A. Naturally Occurring Acetylcholinesterase Inhibitors and Their Potential Use for Alzheimer's Disease Therapy. Front Pharmacol 2018, 9, 1192. [Google Scholar] [CrossRef]
- Kabir, M.T.; Uddin, M.S.; Begum, M.M.; Thangapandiyan, S.; Rahman, M.S.; Aleya, L.; Mathew, B.; Ahmed, M.; Barreto, G.E.; Ashraf, G.M. Cholinesterase Inhibitors for Alzheimer's Disease: Multitargeting Strategy Based on Anti-Alzheimer's Drugs Repositioning. Curr Pharm Des 2019, 25, 3519–3535. [Google Scholar] [CrossRef]
- Murakami, T.; Bodor, E.; Bodor, N. Approaching strategy to increase the oral bioavailability of berberine, a quaternary ammonium isoquinoline alkaloid: Part 1. Physicochemical and pharmacokinetic properties. Expert Opin Drug Metab Toxicol 2023, 19, 129–137. [Google Scholar] [CrossRef]
- Abo El-Enin, H.A.; Elkomy, M.H.; Naguib, I.A.; Ahmed, M.F.; Alsaidan, O.A.; Alsalahat, I.; Ghoneim, M.M.; Eid, H.M. Lipid Nanocarriers Overlaid with Chitosan for Brain Delivery of Berberine via the Nasal Route. Pharmaceuticals (Basel) 2022, 15. [Google Scholar] [CrossRef]
- Kwon, M.; Lim, D.Y.; Lee, C.H.; Jeon, J.H.; Choi, M.K.; Song, I.S. Enhanced Intestinal Absorption and Pharmacokinetic Modulation of Berberine and Its Metabolites through the Inhibition of P-Glycoprotein and Intestinal Metabolism in Rats Using a Berberine Mixed Micelle Formulation. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- El-Nahas, A.E.; Elbedaiwy, H.M.; Masoud, I.M.; Aly, R.G.; Helmy, M.W.; El-Kamel, A.H. Berberine-loaded zein/hyaluronic acid composite nanoparticles for efficient brain uptake to alleviate neuro-degeneration in the pilocarpine model of epilepsy. Eur J Pharm Biopharm 2023, 188, 182–200. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Sang, W.; Linghu, K.G.; Zhong, Z.F.; Cheang, W.S.; Li, J.; Hu, Y.J.; Yu, H.; Wang, Y.T. Dual-functional Brij-S20-modified nanocrystal formulation enhances the intestinal transport and oral bioavailability of berberine. Int J Nanomedicine 2018, 13, 3781–3793. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.C.; Yang, M.Y.; Hsieh, W.Y.; Tsay, G.J.; Yang, Y.C.; Huang, Y.F.; Liu, S.Y.; Lai, C.M.; Lee, C.H.; Tang, C.M.; et al. Berberine's Impact on Apoptosis, Proliferation, Uptake Efficiency, and Nanoparticle-Based Therapy in DBTRG Cells. ACS Nanosci Au 2025, 5, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Elsheikh, M.A.; Elnaggar, Y.S.R.; Hamdy, D.A.; Abdallah, O.Y. Novel cremochylomicrons for improved oral bioavailability of the antineoplastic phytomedicine berberine chloride: Optimization and pharmacokinetics. Int J Pharm 2018, 535, 316–324. [Google Scholar] [CrossRef]
- Bian, X.; Guo, Q.; Yau, L.F.; Yang, L.; Wang, X.; Zhao, S.; Wu, S.; Qin, X.; Jiang, Z.H.; Li, C. Berberine-inspired ionizable lipid for self-structure stabilization and brain targeting delivery of nucleic acid therapeutics. Nat Commun 2025, 16, 2368. [Google Scholar] [CrossRef]
- Saleh, S.R.; Abd-Elmegied, A.; Aly Madhy, S.; Khattab, S.N.; Sheta, E.; Elnozahy, F.Y.; Mehanna, R.A.; Ghareeb, D.A.; Abd-Elmonem, N.M. Brain-targeted Tet-1 peptide-PLGA nanoparticles for berberine delivery against STZ-induced Alzheimer's disease in a rat model: Alleviation of hippocampal synaptic dysfunction, Tau pathology, and amyloidogenesis. Int J Pharm 2024, 658, 124218. [Google Scholar] [CrossRef]
- Marucci, G.; Buccioni, M.; Ben, D.D.; Lambertucci, C.; Volpini, R.; Amenta, F. Efficacy of acetylcholinesterase inhibitors in Alzheimer's disease. Neuropharmacology 2021, 190, 108352. [Google Scholar] [CrossRef]
- Moss, D.E. Improving Anti-Neurodegenerative Benefits of Acetylcholinesterase Inhibitors in Alzheimer's Disease: Are Irreversible Inhibitors the Future? Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Ebert, A.; Goss, K.U. Blood-brain barrier permeability revisited: Predicting intrinsic passive BBB permeability using the Solubility-diffusion model. Eur J Pharm Sci 2025, 215, 107354. [Google Scholar] [CrossRef]
- Fong, C.W. Permeability of the Blood-Brain Barrier: Molecular Mechanism of Transport of Drugs and Physiologically Important Compounds. J Membr Biol 2015, 248, 651–669. [Google Scholar] [CrossRef]
- Vilar, S.; Sobarzo-Sanchez, E.; Santana, L.; Uriarte, E. Ligand and Structure-based Modeling of Passive Diffusion through the Blood-Brain Barrier. Curr Med Chem 2018, 25, 1073–1089. [Google Scholar] [CrossRef] [PubMed]
- Calapai, F.; Cardia, L.; Sorbara, E.E.; Navarra, M.; Gangemi, S.; Calapai, G.; Mannucci, C. Cannabinoids, Blood-Brain Barrier, and Brain Disposition. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Basavarajappa, B.S.; Subbanna, S. Unveiling the Potential of Phytocannabinoids: Exploring Marijuana's Lesser-Known Constituents for Neurological Disorders. Biomolecules 2024, 14. [Google Scholar] [CrossRef]
- Chatterjee, S.; Deshpande, A.A.; Shen, H. Recent advances in the in vitro and in vivo methods to assess impact of P-glycoprotein and breast cancer resistance protein transporters in central nervous system drug disposition. Biopharm Drug Dispos 2023, 44, 7–25. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.C.; Feng, B.; Hou, X.; West, M.A.; Trapa, P.E.; Sciabola, S.; Verhoest, P.; Liras, J.L.; Maurer, T.S.; Wager, T.T. Harnessing Preclinical Data as a Predictive Tool for Human Brain Tissue Targeting. ACS Chem Neurosci 2021, 12, 1007–1017. [Google Scholar] [CrossRef]
- Śmiarowska, M.; Białecka, M.; Machoy-Mokrzyńska, A. Cannabis and cannabinoids: pharmacology and therapeutic potential. Neurol Neurochir Pol 2022, 56, 4–13. [Google Scholar] [CrossRef]
- Friedman, D.; French, J.A.; Maccarrone, M. Safety, efficacy, and mechanisms of action of cannabinoids in neurological disorders. Lancet Neurol 2019, 18, 504–512. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat Rev Neurol 2020, 16, 9–29. [Google Scholar] [CrossRef]
- Blebea, N.M.; Pricopie, A.I.; Vlad, R.A.; Hancu, G. Phytocannabinoids: Exploring Pharmacological Profiles and Their Impact on Therapeutical Use. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Pagano, C.; Navarra, G.; Coppola, L.; Avilia, G.; Bifulco, M.; Laezza, C. Cannabinoids: Therapeutic Use in Clinical Practice. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Masteikova, R.; Lazauskas, R.; Bernatoniene, J. Cannabis sativa L. Bioactive Compounds and Their Protective Role in Oxidative Stress and Inflammation. Antioxidants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, L.; Landucci, E.; Pieraccini, G.; Mazzantini, C.; Bergonzi, M.C.; Pellegrini-Giampietro, D.E.; Bilia, A.R. Development and Blood-Brain Barrier Penetration of Nanovesicles Loaded with Cannabidiol. Pharmaceuticals (Basel) 2025, 18. [Google Scholar] [CrossRef] [PubMed]
- Al-Khazaleh, A.K.; Zhou, X.; Bhuyan, D.J.; Münch, G.W.; Al-Dalabeeh, E.A.; Jaye, K.; Chang, D. The Neurotherapeutic Arsenal in Cannabis sativa: Insights into Anti-Neuroinflammatory and Neuroprotective Activity and Potential Entourage Effects. Molecules 2024, 29. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, L.; Liu, G.; Fan, W.; Li, B.; Liu, R.; Wang, Z.; Fan, Q.; Xiao, W.; Li, Y.; et al. Ginkgo diterpene lactones inhibit cerebral ischemia/reperfusion induced inflammatory response in astrocytes via TLR4/NF-κB pathway in rats. J Ethnopharmacol 2020, 249, 112365. [Google Scholar] [CrossRef]
- Yu, T.; Wei, Z.; Wang, J.; Song, C.; Huang, W.; Zhang, P.; Shi, J.; Zhang, R.; Jiang, M.; Wang, D.; et al. Ginkgo biloba Extract GBE50 ameliorates cerebrovascular dysfunction and cognitive impairment in a mouse model of Alzheimer's disease. Phytomedicine 2025, 141, 156646. [Google Scholar] [CrossRef]
- Upadhyay, G.; Fihurka, O.; Patel, P.; Sanchez-Ramos, J. Quantitation of Cannabidiol (CBD) in brain regions and plasma following intranasal administration of a CBD nanoformulation. J Cannabis Res 2025, 7, 63. [Google Scholar] [CrossRef]
- Dehnbostel, F.O.; Dixit, V.A.; Preissner, R.; Banerjee, P. Non-animal models for blood-brain barrier permeability evaluation of drug-like compounds. Sci Rep 2024, 14, 8908. [Google Scholar] [CrossRef]
- Mohan Kumar, D.; Talwar, P. Neurotherapeutics across blood-brain barrier: screening of BBB-permeable and CNS-active molecules for neurodegenerative disease. Front Pharmacol 2025, 16, 1616144. [Google Scholar] [CrossRef]
- Etukudo, E.M.; Usman, I.M.; Oviosun, A.; Ojiakor, V.O.; Makena, W.; Owembabazi, E.; Aja, P.M.; Mutume Nzanzu Vivalya, B.; Archibong, V.B.; Anyanwu, E. Exploring the Neuroprotective Potentials of Flavonoid Metabolites in Syzygium aromaticum: A Review with in-silico Insight to Therapeutic Potential. J Exp Pharmacol 2025, 17, 587–611. [Google Scholar] [CrossRef]
- Meng, F.; Xi, Y.; Huang, J.; Ayers, P.W. A curated diverse molecular database of blood-brain barrier permeability with chemical descriptors. Sci Data 2021, 8, 289. [Google Scholar] [CrossRef]
- Janicka, M.; Sztanke, M.; Sztanke, K. Modeling the Blood-Brain Barrier Permeability of Potential Heterocyclic Drugs via Biomimetic IAM Chromatography Technique Combined with QSAR Methodology. Molecules 2024, 29. [Google Scholar] [CrossRef]
- Kocsis, A.E.; Kucsápszky, N.; Santa-Maria, A.R.; Hunyadi, A.; Deli, M.A.; Walter, F.R. Much More than Nutrients: The Protective Effects of Nutraceuticals on the Blood-Brain Barrier in Diseases. Nutrients 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- Isabel, U.-V.; Belén, A.d.l.R.M.; Elena, G.-B. A new frontier in neuropharmacology: Recent progress in natural products research for blood–brain barrier crossing. Current Research in Biotechnology 2024, 8, 100235. [Google Scholar] [CrossRef]
- Kato, R.; Zhang, L.; Kinatukara, N.; Huang, R.; Asthana, A.; Weber, C.; Xia, M.; Xu, X.; Shah, P. Investigating blood-brain barrier penetration and neurotoxicity of natural products for central nervous system drug development. Sci Rep 2025, 15, 7431. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Banerjee, A.; Roy, K. Breaking the Barriers: Machine-Learning-Based c-RASAR Approach for Accurate Blood-Brain Barrier Permeability Prediction. J Chem Inf Model 2024, 64, 4298–4309. [Google Scholar] [CrossRef]
- Spielvogel, C.P.; Schindler, N.; Schröder, C.; Stellnberger, S.L.; Wadsak, W.; Mitterhauser, M.; Papp, L.; Hacker, M.; Pichler, V.; Vraka, C. Enhancing Blood-Brain Barrier Penetration Prediction by Machine Learning-Based Integration of Novel and Existing, In Silico and Experimental Molecular Parameters from a Standardized Database. J Chem Inf Model 2025, 65, 2773–2784. [Google Scholar] [CrossRef]
- Liu, S.; Jin, X.; Ge, Y.; Dong, J.; Liu, X.; Pei, X.; Wang, P.; Wang, B.; Chang, Y.; Yu, X.A. Advances in brain-targeted delivery strategies and natural product-mediated enhancement of blood-brain barrier permeability. J Nanobiotechnology 2025, 23, 382. [Google Scholar] [CrossRef]
- Maher, R.; Moreno-Borrallo, A.; Jindal, D.; Mai, B.T.; Ruiz-Hernandez, E.; Harkin, A. Intranasal Polymeric and Lipid-Based Nanocarriers for CNS Drug Delivery. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Ilić, T.; Đoković, J.B.; Nikolić, I.; Mitrović, J.R.; Pantelić, I.; Savić, S.D.; Savić, M.M. Parenteral Lipid-Based Nanoparticles for CNS Disorders: Integrating Various Facets of Preclinical Evaluation towards More Effective Clinical Translation. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Maeng, H.J. Pharmacokinetics and Pharmacodynamics of Intranasal Solid Lipid Nanoparticles and Nanostructured Lipid Carriers for Nose-to-Brain Delivery. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, Z.; Zhang, H.; Wei, H.; Wang, T.; Du, S.; Li, P. Transnasal PLGA Nanoparticles with Terpene Permeation Enhancers: Membrane Remodeling and Tight Junction Modulation for Enhanced Brain Drug Delivery. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef] [PubMed]
- Reddy, T.S.; Zomer, R.; Mantri, N. Nanoformulations as a strategy to overcome the delivery limitations of cannabinoids. Phytother Res 2023, 37, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Pires, P.C.; Rodrigues, M.; Alves, G.; Santos, A.O. Strategies to Improve Drug Strength in Nasal Preparations for Brain Delivery of Low Aqueous Solubility Drugs. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Koo, J.; Lim, C.; Oh, K.T. Recent Advances in Intranasal Administration for Brain-Targeting Delivery: A Comprehensive Review of Lipid-Based Nanoparticles and Stimuli-Responsive Gel Formulations. Int J Nanomedicine 2024, 19, 1767–1807. [Google Scholar] [CrossRef] [PubMed]
- Akpinar Adscheid, S.; Rojas-Rodríguez, M.; Abdel-Hafez, S.M.; Pavone, F.S.; Schneider, M.; Türeli, A.E.; Calamai, M.; Günday-Türeli, N. Scalable Manufacturing Method for Model Protein-Loaded PLGA Nanoparticles: Biocompatibility, Trafficking and Release Properties. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Lababidi, J.M.; Azzazy, H.M.E. Revamping Parkinson's disease therapy using PLGA-based drug delivery systems. NPJ Parkinsons Dis 2025, 11, 248. [Google Scholar] [CrossRef]
- Yang, J.; Zeng, H.; Luo, Y.; Chen, Y.; Wang, M.; Wu, C.; Hu, P. Recent Applications of PLGA in Drug Delivery Systems. Polymers (Basel) 2024, 16. [Google Scholar] [CrossRef]
- Sheffey, V.V.; Siew, E.B.; Tanner, E.E.L.; Eniola-Adefeso, O. PLGA's Plight and the Role of Stealth Surface Modification Strategies in Its Use for Intravenous Particulate Drug Delivery. Adv Healthc Mater 2022, 11, e2101536. [Google Scholar] [CrossRef]
- Kesharwani, P.; Kumar, V.; Goh, K.W.; Gupta, G.; Alsayari, A.; Wahab, S.; Sahebkar, A. PEGylated PLGA nanoparticles: unlocking advanced strategies for cancer therapy. Mol Cancer 2025, 24, 205. [Google Scholar] [CrossRef]
- Sánchez-López, E.; Ettcheto, M.; Egea, M.A.; Espina, M.; Cano, A.; Calpena, A.C.; Camins, A.; Carmona, N.; Silva, A.M.; Souto, E.B.; et al. Memantine loaded PLGA PEGylated nanoparticles for Alzheimer's disease: in vitro and in vivo characterization. J Nanobiotechnology 2018, 16, 32. [Google Scholar] [CrossRef]
- De Soricellis, C.; Amante, C.; Russo, P.; Aquino, R.P.; Del Gaudio, P. Prilling as an Effective Tool for Manufacturing Submicrometric and Nanometric PLGA Particles for Controlled Drug Delivery to Wounds: Stability and Curcumin Release. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- Hoyos-Ceballos, G.P.; Ruozi, B.; Ottonelli, I.; Da Ros, F.; Vandelli, M.A.; Forni, F.; Daini, E.; Vilella, A.; Zoli, M.; Tosi, G.; et al. PLGA-PEG-ANG-2 Nanoparticles for Blood-Brain Barrier Crossing: Proof-of-Concept Study. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- Zhang, W.; Refaat, A.; Li, H.; Zhu, D.; Tong, Z.; Nicolazzo, J.A.; Peng, B.; Bai, H.; Esser, L.; Voelcker, N.H. Optimizing Angiopep-2 Density on Polymeric Nanoparticles for Enhanced Blood–Brain Barrier Penetration and Glioblastoma Targeting: Insights From In Vitro and In Vivo Experiments. Advanced Functional Materials 2025, 2425165. [CrossRef]
- Chen, Z.A.; Wu, C.H.; Wu, S.H.; Huang, C.Y.; Mou, C.Y.; Wei, K.C.; Yen, Y.; Chien, I.T.; Runa, S.; Chen, Y.P.; et al. Receptor Ligand-Free Mesoporous Silica Nanoparticles: A Streamlined Strategy for Targeted Drug Delivery across the Blood-Brain Barrier. ACS Nano 2024, 18, 12716–12736. [Google Scholar] [CrossRef]
- Ojeda-Hernández, D.D.; Canales-Aguirre, A.A.; Matias-Guiu, J.; Gomez-Pinedo, U.; Mateos-Díaz, J.C. Potential of Chitosan and Its Derivatives for Biomedical Applications in the Central Nervous System. Front Bioeng Biotechnol 2020, 8, 389. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Wang, A.; Hua, H.; Jiang, Y.; Wang, Y.; Mu, H.; Wu, Z.; Sun, K. Intranasal delivery of Huperzine A to the brain using lactoferrin-conjugated N-trimethylated chitosan surface-modified PLGA nanoparticles for treatment of Alzheimer's disease. Int J Nanomedicine 2018, 13, 705–718. [Google Scholar] [CrossRef]
- O'Donnell, A.; Moollan, A.; Baneham, S.; Ozgul, M.; Pabari, R.M.; Cox, D.; Kirby, B.P.; Ramtoola, Z. Intranasal and intravenous administration of octa-arginine modified poly(lactic-co-glycolic acid) nanoparticles facilitates central nervous system delivery of loperamide. J Pharm Pharmacol 2015, 67, 525–536. [Google Scholar] [CrossRef]
- Jang, Y.J.; Kang, S.J.; Park, H.S.; Lee, D.H.; Kim, J.H.; Kim, J.E.; Kim, D.I.; Chung, C.H.; Yoon, J.K.; Bhang, S.H. Drug delivery strategies with lipid-based nanoparticles for Alzheimer's disease treatment. J Nanobiotechnology 2025, 23, 99. [Google Scholar] [CrossRef]
- Aslam, M.; Javed, M.N.; Deeb, H.H.; Nicola, M.K.; Mirza, M.A.; Alam, M.S.; Akhtar, M.H.; Waziri, A. Lipid Nanocarriers for Neurotherapeutics: Introduction, Challenges, Blood-brain Barrier, and Promises of Delivery Approaches. CNS Neurol Disord Drug Targets 2022, 21, 952–965. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Z.; R.S., M; Owens, T.; M.H.K., R; Wu, C. PEGylated liposomes via ATRP for brain drug delivery. J Liposome Res 2025, 35, 283–289. [Google Scholar] [CrossRef]
- Du, Q.; Liu, Y.; Fan, M.; Wei, S.; Ismail, M.; Zheng, M. PEG length effect of peptide-functional liposome for blood brain barrier (BBB) penetration and brain targeting. J Control Release 2024, 372, 85–94. [Google Scholar] [CrossRef]
- Zuberi, A.; Rehman, U.; Gupta, G.; Ghazwani, M.; Hani, U.; Kesharwani, P. Smart liposomal systems for brain cancer: Technological innovations in drug delivery. Colloids Surf B Biointerfaces 2025, 255, 114904. [Google Scholar] [CrossRef]
- Tapeinos, C.; Battaglini, M.; Ciofani, G. Advances in the design of solid lipid nanoparticles and nanostructured lipid carriers for targeting brain diseases. J Control Release 2017, 264, 306–332. [Google Scholar] [CrossRef] [PubMed]
- Mehrdadi, S. Drug Delivery of Solid Lipid Nanoparticles (SLNs) and Nanostructured Lipid Carriers (NLCs) to Target Brain Tumors. Adv Pharm Bull 2023, 13, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.P.; Moreira, J.N.; Sousa Lobo, J.M.; Silva, A.C. Intranasal delivery of nanostructured lipid carriers, solid lipid nanoparticles and nanoemulsions: A current overview of in vivo studies. Acta Pharm Sin B 2021, 11, 925–940. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Cui, L.; Lu, H.; Liu, Z.; Zhai, Z.; Wang, H.; Shao, L.; Lu, Z.; Song, X.; Zhang, Y. Nose to Brain: Exploring the Progress of Intranasal Delivery of Solid Lipid Nanoparticles and Nanostructured Lipid Carriers. Int J Nanomedicine 2024, 19, 12343–12368. [Google Scholar] [CrossRef]
- Bonferoni, M.C.; Rossi, S.; Sandri, G.; Ferrari, F.; Gavini, E.; Rassu, G.; Giunchedi, P. Nanoemulsions for "Nose-to-Brain" Drug Delivery. Pharmaceutics 2019, 11. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Duong, V.A. Advancements in Nanocarrier Systems for Nose-to-Brain Drug Delivery. Pharmaceuticals (Basel) 2025, 18. [Google Scholar] [CrossRef]
- Bahadur, S.; Pardhi, D.M.; Rautio, J.; Rosenholm, J.M.; Pathak, K. Intranasal Nanoemulsions for Direct Nose-to-Brain Delivery of Actives for CNS Disorders. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- Chatterjee, B.; Gorain, B.; Mohananaidu, K.; Sengupta, P.; Mandal, U.K.; Choudhury, H. Targeted drug delivery to the brain via intranasal nanoemulsion: Available proof of concept and existing challenges. Int J Pharm 2019, 565, 258–268. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, C.; Pang, Z. Dendrimer-Based Drug Delivery Systems for Brain Targeting. Biomolecules 2019, 9. [Google Scholar] [CrossRef]
- Li, H.; Zha, S.; Li, H.; Liu, H.; Wong, K.L.; All, A.H. Polymeric Dendrimers as Nanocarrier Vectors for Neurotheranostics. Small 2022, 18, e2203629. [Google Scholar] [CrossRef]
- Janaszewska, A.; Lazniewska, J.; Trzepiński, P.; Marcinkowska, M.; Klajnert-Maculewicz, B. Cytotoxicity of Dendrimers. Biomolecules 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, P.; Roullin, V.G.; Chain, J.L. Crossing the blood-brain barrier: advances in dendrimer-based nanocarriers for central nervous system delivery. Nanoscale 2025, 17, 23202–23227. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, D.; Yapa, P.; Dabare, S.; Munaweera, I. Precision targeting of the CNS: recent progress in brain-directed nanodrug delivery. RSC Adv 2025, 15, 25910–25928. [Google Scholar] [CrossRef] [PubMed]
- Zawadzki, S.; Martín-Serrano, Á.; Okła, E.; Kędzierska, M.; Garcia-Gallego, S.; López, P.O.; de la Mata, F.J.; Michlewska, S.; Makowski, T.; Ionov, M.; et al. Synthesis and biophysical evaluation of carbosilane dendrimers as therapeutic siRNA carriers. Sci Rep 2024, 14, 1615. [Google Scholar] [CrossRef]
- Santos, S.D.; Xavier, M.; Leite, D.M.; Moreira, D.A.; Custódio, B.; Torrado, M.; Castro, R.; Leiro, V.; Rodrigues, J.; Tomás, H.; et al. PAMAM dendrimers: blood-brain barrier transport and neuronal uptake after focal brain ischemia. J Control Release 2018, 291, 65–79. [Google Scholar] [CrossRef]
- Yan, X.; Chen, Q. Polyamidoamine Dendrimers: Brain-Targeted Drug Delivery Systems in Glioma Therapy. Polymers (Basel) 2024, 16. [Google Scholar] [CrossRef]
- Jiang, T.; Qiao, Y.; Ruan, W.; Zhang, D.; Yang, Q.; Wang, G.; Chen, Q.; Zhu, F.; Yin, J.; Zou, Y.; et al. Cation-Free siRNA Micelles as Effective Drug Delivery Platform and Potent RNAi Nanomedicines for Glioblastoma Therapy. Adv Mater 2021, 33, e2104779. [Google Scholar] [CrossRef]
- Bhagat, N.; Nalawala, Z.; Patel, J.; Das, D.; Baldha, R.; Sarolia, J.; Rathod, S. Self-Assembled systems for Nose-to-Brain delivery of Temozolamide (TMZ) in brain tumor therapy. Int J Pharm 2025, 675, 125540. [Google Scholar] [CrossRef]
- Kadekar, S.; Nawale, G.N.; Rangasami, V.K.; Le Joncour, V.; Laakkonen, P.; Hilborn, J.; Varghese, O.P.; Oommen, O.P. Redox responsive Pluronic micelle mediated delivery of functional siRNA: a modular nano-assembly for targeted delivery. Biomater Sci 2021, 9, 3939–3944. [Google Scholar] [CrossRef]
- Wei, H.X.; Liu, M.H.; Wang, T.Y.; Shih, M.H.; Yu, J.; Yeh, Y.C. Fabrication of pH- and Ultrasound-Responsive Polymeric Micelles: The Effect of Amphiphilic Block Copolymers with Different Hydrophilic/Hydrophobic Block Ratios for Self-Assembly and Controlled Drug Release. Biomacromolecules 2025, 26, 2116–2130. [Google Scholar] [CrossRef] [PubMed]
- Convertine, A.J.; Diab, C.; Prieve, M.; Paschal, A.; Hoffman, A.S.; Johnson, P.H.; Stayton, P.S. pH-responsive polymeric micelle carriers for siRNA drugs. Biomacromolecules 2010, 11, 2904–2911. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, J.; Li, G.; Ni, B.; Liang, Z.; Chen, H.; Xu, C.; Zhou, J.; Huang, J.; Deng, S. Cation-free siRNA-cored nanocapsules for tumor-targeted RNAi therapy. Acta Biomater 2023, 161, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Pechnikova, N.A.; Domvri, K.; Porpodis, K.; Istomina, M.S.; Iaremenko, A.V.; Yaremenko, A.V. Carbon quantum dots in biomedical applications: advances, challenges, and future prospects. Aggregate 2025, 6, e707. [Google Scholar] [CrossRef]
- Perini, G.; Palmieri, V.; Ciasca, G.; De Spirito, M.; Papi, M. Unravelling the Potential of Graphene Quantum Dots in Biomedicine and Neuroscience. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Qiao, R.; Fu, C.; Forgham, H.; Javed, I.; Huang, X.; Zhu, J.; Whittaker, A.K.; Davis, T.P. Magnetic iron oxide nanoparticles for brain imaging and drug delivery. Adv Drug Deliv Rev 2023, 197, 114822. [Google Scholar] [CrossRef]
- Truskewycz, A.; Yin, H.; Halberg, N.; Lai, D.T.H.; Ball, A.S.; Truong, V.K.; Rybicka, A.M.; Cole, I. Carbon Dot Therapeutic Platforms: Administration, Distribution, Metabolism, Excretion, Toxicity, and Therapeutic Potential. Small 2022, 18, e2106342. [Google Scholar] [CrossRef]
- Molaei, M.J. Carbon quantum dots and their biomedical and therapeutic applications: a review. RSC Adv 2019, 9, 6460–6481. [Google Scholar] [CrossRef]
- Das, S.; Mondal, S.; Ghosh, D. Carbon quantum dots in bioimaging and biomedicines. Front Bioeng Biotechnol 2023, 11, 1333752. [Google Scholar] [CrossRef]
- Araújo, C.; Rodrigues, R.O.; Bañobre-López, M.; Silva, A.M.T.; Ribeiro, R.S. Carbon Dots as a Fluorescent Nanosystem for Crossing the Blood-Brain Barrier with Plausible Application in Neurological Diseases. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Chaparro, C.I.P.; Simões, B.T.; Borges, J.P.; Castanho, M.; Soares, P.I.P.; Neves, V. A Promising Approach: Magnetic Nanosystems for Alzheimer's Disease Theranostics. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Mohebichamkhorami, F.; Faizi, M.; Mahmoudifard, M.; Hajikarim-Hamedani, A.; Mohseni, S.S.; Heidari, A.; Ghane, Y.; Khoramjouy, M.; Khayati, M.; Ghasemi, R.; et al. Microfluidic Synthesis of Ultrasmall Chitosan/Graphene Quantum Dots Particles for Intranasal Delivery in Alzheimer's Disease Treatment. Small 2023, 19, e2207626. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, M.; Basu, S.M.; Qasim, M.; Giri, J. Polypropylene sulphide coating on magnetic nanoparticles as a novel platform for excellent biocompatible, stimuli-responsive smart magnetic nanocarriers for cancer therapeutics. Nanoscale 2023, 15, 7384–7402. [Google Scholar] [CrossRef] [PubMed]
- Raja, I.S.; Song, S.J.; Kang, M.S.; Lee, Y.B.; Kim, B.; Hong, S.W.; Jeong, S.J.; Lee, J.C.; Han, D.W. Toxicity of Zero- and One-Dimensional Carbon Nanomaterials. Nanomaterials (Basel) 2019, 9. [Google Scholar] [CrossRef]
- Sharma, M.; Kumar, C.; Arya, S.K.; Puri, S.; Khatri, M. Neurological effects of carbon quantum dots on zebrafish: A review. Neuroscience 2024, 560, 334–346. [Google Scholar] [CrossRef]
- Jacobsen, N.R.; Møller, P.; Clausen, P.A.; Saber, A.T.; Micheletti, C.; Jensen, K.A.; Wallin, H.; Vogel, U. Biodistribution of Carbon Nanotubes in Animal Models. Basic Clin Pharmacol Toxicol 2017, 121 Suppl 3, 30–43. [Google Scholar] [CrossRef]
- Galassi, T.V.; Antman-Passig, M.; Yaari, Z.; Jessurun, J.; Schwartz, R.E.; Heller, D.A. Long-term in vivo biocompatibility of single-walled carbon nanotubes. PLoS One 2020, 15, e0226791. [Google Scholar] [CrossRef]
- Clementino, A.R.; Pellegrini, G.; Banella, S.; Colombo, G.; Cantù, L.; Sonvico, F.; Del Favero, E. Structure and Fate of Nanoparticles Designed for the Nasal Delivery of Poorly Soluble Drugs. Mol Pharm 2021, 18, 3132–3146. [Google Scholar] [CrossRef]
- Haro-Martínez, E.; Muscolino, E.; Moral, N.; Duran, J.; Fornaguera, C. Crossing the blood-brain barrier: nanoparticle-based strategies for neurodegenerative disease therapy. In Drug Deliv Transl Res; 2025. [Google Scholar] [CrossRef]
- Omar, S.H.; Osman, R.; Mamdouh, W.; Abdel-Bar, H.M.; Awad, G.A.S. Bioinspired lipid-polysaccharide modified hybrid nanoparticles as a brain-targeted highly loaded carrier for a hydrophilic drug. Int J Biol Macromol 2020, 165, 483–494. [Google Scholar] [CrossRef]
- Ishak, R.A.H.; Mostafa, N.M.; Kamel, A.O. Stealth lipid polymer hybrid nanoparticles loaded with rutin for effective brain delivery - comparative study with the gold standard (Tween 80): optimization, characterization and biodistribution. Drug Deliv 2017, 24, 1874–1890. [Google Scholar] [CrossRef]
- Yuan, T.; Gao, L.; Zhan, W.; Dini, D. Effect of Particle Size and Surface Charge on Nanoparticles Diffusion in the Brain White Matter. Pharm Res 2022, 39, 767–781. [Google Scholar] [CrossRef]
- Israel, L.L.; Galstyan, A.; Cox, A.; Shatalova, E.S.; Sun, T.; Rashid, M.H.; Grodzinski, Z.; Chiechi, A.; Fuchs, D.T.; Patil, R.; et al. Signature Effects of Vector-Guided Systemic Nano Bioconjugate Delivery Across Blood-Brain Barrier of Normal, Alzheimer's, and Tumor Mouse Models. ACS Nano 2022, 16, 11815–11832. [Google Scholar] [CrossRef]
- Sharma, S.; Lee, D.; Maity, S.; Singh, P.; Chadokiya, J.; Mohaghegh, N.; Hassani, A.; Kim, H.; Gangarade, A.; Ljubimova, J.Y.; et al. Antibody-Free Immunopeptide Nanoconjugates for Brain-Targeted Drug Delivery in Glioblastoma Multiforme. Bioconjug Chem 2025, 36, 2132–2144. [Google Scholar] [CrossRef]
- Kim, W.; Ly, N.K.; He, Y.; Li, Y.; Yuan, Z.; Yeo, Y. Protein corona: Friend or foe? Co-opting serum proteins for nanoparticle delivery. Adv Drug Deliv Rev 2023, 192, 114635. [Google Scholar] [CrossRef] [PubMed]
- Kashapov, R.; Ibragimova, A.; Pavlov, R.; Gabdrakhmanov, D.; Kashapova, N.; Burilova, E.; Zakharova, L.; Sinyashin, O. Nanocarriers for Biomedicine: From Lipid Formulations to Inorganic and Hybrid Nanoparticles. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Alshawwa, S.Z.; Kassem, A.A.; Farid, R.M.; Mostafa, S.K.; Labib, G.S. Nanocarrier Drug Delivery Systems: Characterization, Limitations, Future Perspectives and Implementation of Artificial Intelligence. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Beach, M.A.; Nayanathara, U.; Gao, Y.; Zhang, C.; Xiong, Y.; Wang, Y.; Such, G.K. Polymeric Nanoparticles for Drug Delivery. Chem Rev 2024, 124, 5505–5616. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, W.; Wang, Z.; Lu, J. Lipid-Based Nanotechnology: Liposome. Pharmaceutics 2023, 16. [Google Scholar] [CrossRef]
- Roszkowski, S.; Durczynska, Z. Advantages and limitations of nanostructures for biomedical applications. Adv Clin Exp Med 2025, 34, 447–456. [Google Scholar] [CrossRef]
- Kesharwani, P.; Gothwal, A.; Iyer, A.K.; Jain, K.; Chourasia, M.K.; Gupta, U. Dendrimer nanohybrid carrier systems: an expanding horizon for targeted drug and gene delivery. Drug Discov Today 2018, 23, 300–314. [Google Scholar] [CrossRef] [PubMed]
- Das, S.S.; Bharadwaj, P.; Bilal, M.; Barani, M.; Rahdar, A.; Taboada, P.; Bungau, S.; Kyzas, G.Z. Stimuli-Responsive Polymeric Nanocarriers for Drug Delivery, Imaging, and Theragnosis. Polymers (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Wakaskar, R.R. General overview of lipid-polymer hybrid nanoparticles, dendrimers, micelles, liposomes, spongosomes and cubosomes. J Drug Target 2018, 26, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Yanar, F.; Carugo, D.; Zhang, X. Hybrid Nanoplatforms Comprising Organic Nanocompartments Encapsulating Inorganic Nanoparticles for Enhanced Drug Delivery and Bioimaging Applications. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Mehta, M.; Bui, T.A.; Yang, X.; Aksoy, Y.; Goldys, E.M.; Deng, W. Lipid-Based Nanoparticles for Drug/Gene Delivery: An Overview of the Production Techniques and Difficulties Encountered in Their Industrial Development. ACS Mater Au 2023, 3, 600–619. [Google Scholar] [CrossRef]
- Viegas, C.; Patrício, A.B.; Prata, J.M.; Nadhman, A.; Chintamaneni, P.K.; Fonte, P. Solid Lipid Nanoparticles vs. Nanostructured Lipid Carriers: A Comparative Review. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Scioli Montoto, S.; Muraca, G.; Ruiz, M.E. Solid Lipid Nanoparticles for Drug Delivery: Pharmacological and Biopharmaceutical Aspects. Front Mol Biosci 2020, 7, 587997. [Google Scholar] [CrossRef]
- Duan, Y.; Dhar, A.; Patel, C.; Khimani, M.; Neogi, S.; Sharma, P.; Siva Kumar, N.; Vekariya, R.L. A brief review on solid lipid nanoparticles: part and parcel of contemporary drug delivery systems. RSC Adv 2020, 10, 26777–26791. [Google Scholar] [CrossRef]
- Ranjbar, S.; Emamjomeh, A.; Sharifi, F.; Zarepour, A.; Aghaabbasi, K.; Dehshahri, A.; Sepahvand, A.M.; Zarrabi, A.; Beyzaei, H.; Zahedi, M.M.; et al. Lipid-Based Delivery Systems for Flavonoids and Flavonolignans: Liposomes, Nanoemulsions, and Solid Lipid Nanoparticles. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Dhiman, N.; Awasthi, R.; Sharma, B.; Kharkwal, H.; Kulkarni, G.T. Lipid Nanoparticles as Carriers for Bioactive Delivery. Front Chem 2021, 9, 580118. [Google Scholar] [CrossRef]
- Hassan, A.A.A.; Ramadan, E.; Kristó, K.; Regdon, G., Jr.; Sovány, T. Lipid-Polymer Hybrid Nanoparticles as a Smart Drug Delivery System for Peptide/Protein Delivery. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Sivadasan, D.; Sultan, M.H.; Madkhali, O.; Almoshari, Y.; Thangavel, N. Polymeric Lipid Hybrid Nanoparticles (PLNs) as Emerging Drug Delivery Platform-A Comprehensive Review of Their Properties, Preparation Methods, and Therapeutic Applications. Pharmaceutics 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Seyyedi-Mansour, S.; Carpena, M.; Barciela, P.; Perez-Vazquez, A.; Assadpour, E.; Prieto, M.A.; Jafari, S.M. Lipid-based nanocarriers loaded with bioactive compounds in active food packaging: Fabrication, characterization, and applications. Adv Colloid Interface Sci 2025, 340, 103457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, Q.Y.; Haqqani, A.S.; Leclerc, S.; Liu, Z.; Fauteux, F.; Baumann, E.; Delaney, C.E.; Ly, D.; Star, A.T.; et al. Differential expression of receptors mediating receptor-mediated transcytosis (RMT) in brain microvessels, brain parenchyma and peripheral tissues of the mouse and the human. Fluids Barriers CNS 2020, 17, 47. [Google Scholar] [CrossRef]
- Piantino, M.; Louis, F.; Shigemoto-Mogami, Y.; Kitamura, K.; Sato, K.; Yamaguchi, T.; Kawabata, K.; Yamamoto, S.; Iwasaki, S.; Hirabayashi, H.; et al. Brain microvascular endothelial cells derived from human induced pluripotent stem cells as in vitro model for assessing blood-brain barrier transferrin receptor-mediated transcytosis. Mater Today Bio 2022, 14, 100232. [Google Scholar] [CrossRef] [PubMed]
- Katt, M.E.; Waters, E.A.; Gastfriend, B.D.; Herrin, B.R.; Cooper, M.D.; Shusta, E.V. Identification of Variable Lymphocyte Receptors That Target the Human Blood-Brain Barrier. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Kuhn, P.; Petralla, S.; Dabbagh, F.; Pegoretti, V.; Muranyi, W.; Ishikawa, H.; Schroten, H.; Fischer, R.; Frenzel, A.; Schirrmann, T.; et al. A pH-sensitive binding modality allows successful transferrin receptor-mediated transcytosis of a bivalent antibody across brain barriers. MAbs 2025, 17, 2563758. [Google Scholar] [CrossRef]
- Pardridge, W.M.; Chou, T. Mathematical Models of Blood-Brain Barrier Transport of Monoclonal Antibodies Targeting the Transferrin Receptor and the Insulin Receptor. Pharmaceuticals (Basel) 2021, 14. [Google Scholar] [CrossRef]
- Shen, X.; Li, H.; Zhang, B.; Li, Y.; Zhu, Z. Targeting Transferrin Receptor 1 for Enhancing Drug Delivery Through the Blood-Brain Barrier for Alzheimer's Disease. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Morrison, J.I.; Petrovic, A.; Metzendorf, N.G.; Rofo, F.; Yilmaz, C.U.; Stenler, S.; Laudon, H.; Hultqvist, G. Standardized Preclinical In Vitro Blood-Brain Barrier Mouse Assay Validates Endocytosis-Dependent Antibody Transcytosis Using Transferrin-Receptor-Mediated Pathways. Mol Pharm 2023, 20, 1564–1576. [Google Scholar] [CrossRef]
- Khoury, N.; Pizzo, M.E.; Discenza, C.B.; Joy, D.; Tatarakis, D.; Todorov, M.I.; Negwer, M.; Ha, C.; De Melo, G.L.; Sarrafha, L.; et al. Fc-engineered large molecules targeting blood-brain barrier transferrin receptor and CD98hc have distinct central nervous system and peripheral biodistribution. Nat Commun 2025, 16, 1822. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-López, E.; Schuhmacher, A.J. Transportation of Single-Domain Antibodies through the Blood-Brain Barrier. Biomolecules 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Pompe, S.; Xenaki, K.T.; Di Maggio, A.; Moreno, C.B.; van Bergen En Henegouwen, P.M.P.; Mastrobattista, E.; Oliveira, S.; Caiazzo, M. Receptor-mediated transcytosis of nanobodies targeting the heparin-binding EGF-like growth factor in human blood-brain barrier models. J Control Release 2025, 383, 113852. [Google Scholar] [CrossRef] [PubMed]
- Szecskó, A.; Mészáros, M.; Simões, B.; Cavaco, M.; Chaparro, C.; Porkoláb, G.; Castanho, M.; Deli, M.A.; Neves, V.; Veszelka, S. PepH3-modified nanocarriers for delivery of therapeutics across the blood-brain barrier. Fluids Barriers CNS 2025, 22, 31. [Google Scholar] [CrossRef]
- Ghorai, S.M.; Deep, A.; Magoo, D.; Gupta, C.; Gupta, N. Cell-Penetrating and Targeted Peptides Delivery Systems as Potential Pharmaceutical Carriers for Enhanced Delivery across the Blood-Brain Barrier (BBB). Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Fu, L.; Bridges, C.A.; Kim, H.N.; Ding, C.; Bao Hou, N.C.; Yeow, J.; Fok, S.; Macmillan, A.; Sterling, J.D.; Baker, S.M.; et al. Cationic Polysaccharides Bind to the Endothelial Cell Surface Extracellular Matrix Involving Heparan Sulfate. Biomacromolecules 2024, 25, 3850–3862. [Google Scholar] [CrossRef]
- Reveret, L.; Leclerc, M.; Morin, F.; Émond, V.; Calon, F. Pharmacokinetics, biodistribution and toxicology of novel cell-penetrating peptides. Sci Rep 2023, 13, 11081. [Google Scholar] [CrossRef]
- Habault, J.; Poyet, J.L. Recent Advances in Cell Penetrating Peptide-Based Anticancer Therapies. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Nam, S.H.; Park, J.; Koo, H. Recent advances in selective and targeted drug/gene delivery systems using cell-penetrating peptides. Arch Pharm Res 2023, 46, 18–34. [Google Scholar] [CrossRef]
- Polderdijk, S.G.I.; Limzerwala, J.F.; Spiess, C. Plasma membrane damage limits cytoplasmic delivery by conventional cell penetrating peptides. PLoS One 2024, 19, e0305848. [Google Scholar] [CrossRef]
- Ghaemi, B.; Tanwar, S.; Singh, A.; Arifin, D.R.; McMahon, M.T.; Barman, I.; Bulte, J.W.M. Cell-Penetrating and Enzyme-Responsive Peptides for Targeted Cancer Therapy: Role of Arginine Residue Length on Cell Penetration and In Vivo Systemic Toxicity. ACS Appl Mater Interfaces 2024, 16, 11159–11171. [Google Scholar] [CrossRef]
- Mendes, M.; Nunes, S.; Cova, T.; Branco, F.; Dyrks, M.; Koksch, B.; Vale, N.; Sousa, J.; Pais, A.; Vitorino, C. Charge-switchable cell-penetrating peptides for rerouting nanoparticles to glioblastoma treatment. Colloids Surf B Biointerfaces 2024, 241, 113983. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Zaro, J.L.; Shen, Y.; Chen, Q.; Yu, Y.; Sun, P.; Wang, Y.; Shen, W.C.; Tu, J.; Sun, C. Acid-sensitive hybrid polymeric micelles containing a reversibly activatable cell-penetrating peptide for tumor-specific cytoplasm targeting. J Control Release 2018, 279, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Du, J.J.; Zhang, R.Y.; Jiang, S.; Xiao, S.; Liu, Y.; Niu, Y.; Zhao, W.X.; Wang, D.; Ma, X. Applications of cell penetrating peptide-based drug delivery system in immunotherapy. Front Immunol 2025, 16, 1540192. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Mi, G.; Shen, Y.; Webster, T.J. Glioma-targeted dual functionalized thermosensitive Ferri-liposomes for drug delivery through an in vitro blood-brain barrier. Nanoscale 2019, 11, 15057–15071. [Google Scholar] [CrossRef]
- Porro, G.; Basile, M.; Xie, Z.; Tuveri, G.M.; Battaglia, G.; Lopes, C.D.F. A new era in brain drug delivery: Integrating multivalency and computational optimisation for blood-brain barrier permeation. Adv Drug Deliv Rev 2025, 224, 115637. [Google Scholar] [CrossRef]
- Moreira, R.; Nóbrega, C.; de Almeida, L.P.; Mendonça, L. Brain-targeted drug delivery - nanovesicles directed to specific brain cells by brain-targeting ligands. J Nanobiotechnology 2024, 22, 260. [Google Scholar] [CrossRef]
- Luo, F.; Zhong, T.; Chen, Y.; Guo, Q.; Tao, L.; Shen, X.; Fan, Y.; Wu, X. Dual-Ligand Synergistic Targeting Anti-Tumor Nanoplatforms with Cascade-Responsive Drug Release. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Guo, Q.; Xu, S.; Yang, P.; Wang, P.; Lu, S.; Sheng, D.; Qian, K.; Cao, J.; Lu, W.; Zhang, Q. A dual-ligand fusion peptide improves the brain-neuron targeting of nanocarriers in Alzheimer's disease mice. J Control Release 2020, 320, 347–362. [Google Scholar] [CrossRef]
- Chen, C.; Duan, Z.; Yuan, Y.; Li, R.; Pang, L.; Liang, J.; Xu, X.; Wang, J. Peptide-22 and Cyclic RGD Functionalized Liposomes for Glioma Targeting Drug Delivery Overcoming BBB and BBTB. ACS Appl Mater Interfaces 2017, 9, 5864–5873. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhai, M.; Chen, Z.; Han, X.; Yu, F.; Li, Z.; Xie, X.; Han, C.; Yu, L.; Yang, Y.; et al. Dual-modified liposome codelivery of doxorubicin and vincristine improve targeting and therapeutic efficacy of glioma. Drug Deliv 2017, 24, 1045–1055. [Google Scholar] [CrossRef]
- Ling, C.; Chen, L.; Liang, J.; Li, X.; Wei, H.; Yu, C.; Wang, J. Neutrophil/monocyte-targeted dual-ligands modified liposomes delivering puerarin for ischemia stroke treatment. Mater Today Bio 2025, 33, 102077. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, Q.; Chen, Z.; Liu, J.; Zhang, Z.; Tian, J.; Wang, X.; Yang, X.; Chen, Q.; Huang, X.; et al. Rational Design of Dual-Targeted Nanomedicines for Enhanced Vascular Permeability in Low-Permeability Tumors. ACS Nano 2025, 19, 3424–3438. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Yang, L.; Wang, R.; Song, G.; Fu, J.; Wang, J.; Gao, N.; Wang, H. Drug Delivery Across the Blood-Brain Barrier: A New Strategy for the Treatment of Neurological Diseases. Pharmaceutics 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Toader, C.; Dumitru, A.V.; Eva, L.; Serban, M.; Covache-Busuioc, R.A.; Ciurea, A.V. Nanoparticle Strategies for Treating CNS Disorders: A Comprehensive Review of Drug Delivery and Theranostic Applications. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Lv, Z.; Yang, Q.; Chang, R.; Wu, J. Research Progress on Stimulus-Responsive Polymer Nanocarriers for Cancer Treatment. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Deirram, N.; Zhang, C.; Kermaniyan, S.S.; Johnston, A.P.R.; Such, G.K. pH-Responsive Polymer Nanoparticles for Drug Delivery. Macromol Rapid Commun 2019, 40, e1800917. [Google Scholar] [CrossRef]
- Meng, X.; Shen, Y.; Zhao, H.; Lu, X.; Wang, Z.; Zhao, Y. Redox-manipulating nanocarriers for anticancer drug delivery: a systematic review. J Nanobiotechnology 2024, 22, 587. [Google Scholar] [CrossRef]
- Hersh, A.M.; Bhimreddy, M.; Weber-Levine, C.; Jiang, K.; Alomari, S.; Theodore, N.; Manbachi, A.; Tyler, B.M. Applications of Focused Ultrasound for the Treatment of Glioblastoma: A New Frontier. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- He, C.; Wu, Z.; Zhuang, M.; Li, X.; Xue, S.; Xu, S.; Xu, J.; Wu, Z.; Lu, M. Focused ultrasound-mediated blood-brain barrier opening combined with magnetic targeting cytomembrane based biomimetic microbubbles for glioblastoma therapy. J Nanobiotechnology 2023, 21, 297. [Google Scholar] [CrossRef]
- Teleanu, R.I.; Preda, M.D.; Niculescu, A.G.; Vladâcenco, O.; Radu, C.I.; Grumezescu, A.M.; Teleanu, D.M. Current Strategies to Enhance Delivery of Drugs across the Blood-Brain Barrier. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Sonabend, A.M.; Gould, A.; Amidei, C.; Ward, R.; Schmidt, K.A.; Zhang, D.Y.; Gomez, C.; Bebawy, J.F.; Liu, B.P.; Bouchoux, G.; et al. Repeated blood-brain barrier opening with an implantable ultrasound device for delivery of albumin-bound paclitaxel in patients with recurrent glioblastoma: a phase 1 trial. Lancet Oncol 2023, 24, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Manzari, M.T.; Shamay, Y.; Kiguchi, H.; Rosen, N.; Scaltriti, M.; Heller, D.A. Targeted drug delivery strategies for precision medicines. Nat Rev Mater 2021, 6, 351–370. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, K.B.; Burkhart, A.; Thomsen, L.B.; Andresen, T.L.; Moos, T. Targeting the transferrin receptor for brain drug delivery. Prog Neurobiol 2019, 181, 101665. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.S.; Johnsen, K.B.; Kucharz, K.; Lauritzen, M.; Moos, T. Blood-Brain Barrier Transport of Transferrin Receptor-Targeted Nanoparticles. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Bien-Ly, N.; Yu, Y.J.; Bumbaca, D.; Elstrott, J.; Boswell, C.A.; Zhang, Y.; Luk, W.; Lu, Y.; Dennis, M.S.; Weimer, R.M.; et al. Transferrin receptor (TfR) trafficking determines brain uptake of TfR antibody affinity variants. J Exp Med 2014, 211, 233–244. [Google Scholar] [CrossRef]
- Arguello, A.; Mahon, C.S.; Calvert, M.E.K.; Chan, D.; Dugas, J.C.; Pizzo, M.E.; Thomsen, E.R.; Chau, R.; Damo, L.A.; Duque, J.; et al. Molecular architecture determines brain delivery of a transferrin receptor-targeted lysosomal enzyme. J Exp Med 2022, 219. [Google Scholar] [CrossRef]
- Bonvicini, G.; Singh, S.; Sandersjöö, L.; Sehlin, D.; Syvänen, S.; Andersson, K.G. The effects of dose, valency, and affinity on TfR-mediated brain delivery in vivo. Fluids Barriers CNS 2025, 22, 36. [Google Scholar] [CrossRef]
- di Polidoro, A.C.; Cafarchio, A.; Vecchione, D.; Donato, P.; De Nola, F.; Torino, E. Revealing Angiopep-2/LRP1 Molecular Interaction for Optimal Delivery to Glioblastoma (GBM). Molecules 2022, 27. [Google Scholar] [CrossRef]
- Khan, N.U.; Ni, J.; Ju, X.; Miao, T.; Chen, H.; Han, L. Escape from abluminal LRP1-mediated clearance for boosted nanoparticle brain delivery and brain metastasis treatment. Acta Pharm Sin B 2021, 11, 1341–1354. [Google Scholar] [CrossRef]
- Kafa, H.; Wang, J.T.; Rubio, N.; Klippstein, R.; Costa, P.M.; Hassan, H.A.; Sosabowski, J.K.; Bansal, S.S.; Preston, J.E.; Abbott, N.J.; et al. Translocation of LRP1 targeted carbon nanotubes of different diameters across the blood-brain barrier in vitro and in vivo. J Control Release 2016, 225, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Yamada, S.; Suzuki, R.; Iida, Y.; Matsumoto, M.; Fumoto, S.; Arima, H.; Mukai, H.; Kawakami, S. Development of an apolipoprotein E mimetic peptide-lipid conjugate for efficient brain delivery of liposomes. Drug Deliv 2023, 30, 2173333. [Google Scholar] [CrossRef] [PubMed]
- Benitez Amaro, A.; Solanelles Curco, A.; Garcia, E.; Julve, J.; Rives, J.; Benitez, S.; Llorente Cortes, V. Apolipoprotein and LRP1-Based Peptides as New Therapeutic Tools in Atherosclerosis. J Clin Med 2021, 10. [Google Scholar] [CrossRef]
- Hjelm, L.C.; Lindberg, H.; Ståhl, S.; Löfblom, J. Affibody Molecules Intended for Receptor-Mediated Transcytosis via the Transferrin Receptor. Pharmaceuticals (Basel) 2023, 16. [Google Scholar] [CrossRef] [PubMed]
- Crowe, T.P.; Hsu, W.H. Evaluation of Recent Intranasal Drug Delivery Systems to the Central Nervous System. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Crowe, T.P.; Greenlee, M.H.W.; Kanthasamy, A.G.; Hsu, W.H. Mechanism of intranasal drug delivery directly to the brain. Life Sci 2018, 195, 44–52. [Google Scholar] [CrossRef]
- Islam, S.U.; Shehzad, A.; Ahmed, M.B.; Lee, Y.S. Intranasal Delivery of Nanoformulations: A Potential Way of Treatment for Neurological Disorders. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, C.; Huang, Y.; Ma, Y.; Song, Q.; Chen, H.; Jiang, G.; Gao, X. Intranasal drug delivery: The interaction between nanoparticles and the nose-to-brain pathway. Adv Drug Deliv Rev 2024, 207, 115196. [Google Scholar] [CrossRef]
- Agrawal, M.; Saraf, S.; Saraf, S.; Dubey, S.K.; Puri, A.; Gupta, U.; Kesharwani, P.; Ravichandiran, V.; Kumar, P.; Naidu, V.G.M.; et al. Stimuli-responsive In situ gelling system for nose-to-brain drug delivery. J Control Release 2020, 327, 235–265. [Google Scholar] [CrossRef]
- Vigani, B.; Rossi, S.; Sandri, G.; Bonferoni, M.C.; Caramella, C.M.; Ferrari, F. Recent Advances in the Development of In Situ Gelling Drug Delivery Systems for Non-Parenteral Administration Routes. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- Xu, K.; Duan, S.; Wang, W.; Ouyang, Q.; Qin, F.; Guo, P.; Hou, J.; He, Z.; Wei, W.; Qin, M. Nose-to-brain delivery of nanotherapeutics: Transport mechanisms and applications. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2024, 16, e1956. [Google Scholar] [CrossRef] [PubMed]
- Lofts, A.; Abu-Hijleh, F.; Rigg, N.; Mishra, R.K.; Hoare, T. Using the Intranasal Route to Administer Drugs to Treat Neurological and Psychiatric Illnesses: Rationale, Successes, and Future Needs. CNS Drugs 2022, 36, 739–770. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Chen, Y.; Zhang, W.; Xia, X.; Li, H.; Qin, M.; Gao, H. Nanotechnology for enhanced nose-to-brain drug delivery in treating neurological diseases. J Control Release 2024, 366, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Gorick, C.M.; Breza, V.R.; Nowak, K.M.; Cheng, V.W.T.; Fisher, D.G.; Debski, A.C.; Hoch, M.R.; Demir, Z.E.F.; Tran, N.M.; Schwartz, M.R.; et al. Applications of focused ultrasound-mediated blood-brain barrier opening. Adv Drug Deliv Rev 2022, 191, 114583. [Google Scholar] [CrossRef]
- Chen, K.T.; Wei, K.C.; Liu, H.L. Theranostic Strategy of Focused Ultrasound Induced Blood-Brain Barrier Opening for CNS Disease Treatment. Front Pharmacol 2019, 10, 86. [Google Scholar] [CrossRef]
- Zhu, H.; Allwin, C.; Bassous, M.G.; Pouliopoulos, A.N. Focused ultrasound-mediated enhancement of blood-brain barrier permeability for brain tumor treatment: a systematic review of clinical trials. J Neurooncol 2024, 170, 235–252. [Google Scholar] [CrossRef]
- Gosselet, F.; Loiola, R.A.; Roig, A.; Rosell, A.; Culot, M. Central nervous system delivery of molecules across the blood-brain barrier. Neurochem Int 2021, 144, 104952. [Google Scholar] [CrossRef]
- Sun, T.; Samiotaki, G.; Wang, S.; Acosta, C.; Chen, C.C.; Konofagou, E.E. Acoustic cavitation-based monitoring of the reversibility and permeability of ultrasound-induced blood-brain barrier opening. Phys Med Biol 2015, 60, 9079–9094. [Google Scholar] [CrossRef]
- Xie, Y.; Hu, J.; Lei, W.; Qian, S. Prediction of vascular injury by cavitation microbubbles in a focused ultrasound field. Ultrason Sonochem 2022, 88, 106103. [Google Scholar] [CrossRef]
- Meng, Y.; Kalia, L.V.; Kalia, S.K.; Hamani, C.; Huang, Y.; Hynynen, K.; Lipsman, N.; Davidson, B. Current Progress in Magnetic Resonance-Guided Focused Ultrasound to Facilitate Drug Delivery across the Blood-Brain Barrier. Pharmaceutics 2024, 16. [Google Scholar] [CrossRef]
- Timbie, K.F.; Mead, B.P.; Price, R.J. Drug and gene delivery across the blood-brain barrier with focused ultrasound. J Control Release 2015, 219, 61–75. [Google Scholar] [CrossRef]
- Burgess, A.; Shah, K.; Hough, O.; Hynynen, K. Focused ultrasound-mediated drug delivery through the blood-brain barrier. Expert Rev Neurother 2015, 15, 477–491. [Google Scholar] [CrossRef]
- Wu, S.Y.; Aurup, C.; Sanchez, C.S.; Grondin, J.; Zheng, W.; Kamimura, H.; Ferrera, V.P.; Konofagou, E.E. Efficient Blood-Brain Barrier Opening in Primates with Neuronavigation-Guided Ultrasound and Real-Time Acoustic Mapping. Sci Rep 2018, 8, 7978. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Zhang, Y.; Power, C.; Alexander, P.M.; Sutton, J.T.; Aryal, M.; Vykhodtseva, N.; Miller, E.L.; McDannold, N.J. Closed-loop control of targeted ultrasound drug delivery across the blood-brain/tumor barriers in a rat glioma model. Proc Natl Acad Sci U S A 2017, 114, E10281–e10290. [Google Scholar] [CrossRef] [PubMed]
- Novell, A.; Kamimura, H.A.S.; Cafarelli, A.; Gerstenmayer, M.; Flament, J.; Valette, J.; Agou, P.; Conti, A.; Selingue, E.; Aron Badin, R.; et al. A new safety index based on intrapulse monitoring of ultra-harmonic cavitation during ultrasound-induced blood-brain barrier opening procedures. Sci Rep 2020, 10, 10088. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Liu, K.; Pouliopoulos, A.N.; Ji, R.; Jiménez-Gambín, S.; Yousefian, O.; Kline-Schoder, A.R.; Batts, A.J.; Tsitsos, F.N.; Kokossis, D.; et al. Transcranial blood-brain barrier opening in Alzheimer's disease patients using a portable focused ultrasound system with real-time 2-D cavitation mapping. Theranostics 2024, 14, 4519–4535. [Google Scholar] [CrossRef]
- Todd, N.; Angolano, C.; Ferran, C.; Devor, A.; Borsook, D.; McDannold, N. Secondary effects on brain physiology caused by focused ultrasound-mediated disruption of the blood-brain barrier. J Control Release 2020, 324, 450–459. [Google Scholar] [CrossRef]
- Wang, J.; Li, Z.; Pan, M.; Fiaz, M.; Hao, Y.; Yan, Y.; Sun, L.; Yan, F. Ultrasound-mediated blood-brain barrier opening: An effective drug delivery system for theranostics of brain diseases. Adv Drug Deliv Rev 2022, 190, 114539. [Google Scholar] [CrossRef]
- Shin, J.; Kong, C.; Cho, J.S.; Lee, J.; Koh, C.S.; Yoon, M.S.; Na, Y.C.; Chang, W.S.; Chang, J.W. Focused ultrasound-mediated noninvasive blood-brain barrier modulation: preclinical examination of efficacy and safety in various sonication parameters. Neurosurg Focus 2018, 44, E15. [Google Scholar] [CrossRef]
- Cheng, M.; Li, F.; Han, T.; Yu, A.C.H.; Qin, P. Effects of ultrasound pulse parameters on cavitation properties of flowing microbubbles under physiologically relevant conditions. Ultrason Sonochem 2019, 52, 512–521. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, L.; Cheng, M.; Jin, L.; Du, L.; Han, T.; Xu, L.; Yu, A.C.H.; Qin, P. Effect of acoustic parameters on the cavitation behavior of SonoVue microbubbles induced by pulsed ultrasound. Ultrason Sonochem 2017, 35, 176–184. [Google Scholar] [CrossRef]
- Zhao, X.; Wright, A.; Goertz, D.E. An optical and acoustic investigation of microbubble cavitation in small channels under therapeutic ultrasound conditions. Ultrason Sonochem 2023, 93, 106291. [Google Scholar] [CrossRef]
- Patel, A.; Schoen, S.J., Jr.; Arvanitis, C.D. Closed Loop Spatial and Temporal Control of Cavitation Activity with Passive Acoustic Mapping. IEEE Trans Biomed Eng 2018. [Google Scholar] [CrossRef] [PubMed]
- Downs, M.E.; Buch, A.; Sierra, C.; Karakatsani, M.E.; Teichert, T.; Chen, S.; Konofagou, E.E.; Ferrera, V.P. Long-Term Safety of Repeated Blood-Brain Barrier Opening via Focused Ultrasound with Microbubbles in Non-Human Primates Performing a Cognitive Task. PLoS One 2015, 10, e0125911. [Google Scholar] [CrossRef]
- Mainprize, T.; Lipsman, N.; Huang, Y.; Meng, Y.; Bethune, A.; Ironside, S.; Heyn, C.; Alkins, R.; Trudeau, M.; Sahgal, A.; et al. Blood-Brain Barrier Opening in Primary Brain Tumors with Non-invasive MR-Guided Focused Ultrasound: A Clinical Safety and Feasibility Study. Sci Rep 2019, 9, 321. [Google Scholar] [CrossRef]
- Rezai, A.R.; Ranjan, M.; Haut, M.W.; Carpenter, J.; D'Haese, P.F.; Mehta, R.I.; Najib, U.; Wang, P.; Claassen, D.O.; Chazen, J.L.; et al. Focused ultrasound-mediated blood-brain barrier opening in Alzheimer's disease: long-term safety, imaging, and cognitive outcomes. J Neurosurg 2023, 139, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Anastasiadis, P.; Gandhi, D.; Guo, Y.; Ahmed, A.K.; Bentzen, S.M.; Arvanitis, C.; Woodworth, G.F. Localized blood-brain barrier opening in infiltrating gliomas with MRI-guided acoustic emissions-controlled focused ultrasound. Proc Natl Acad Sci U S A 2021, 118. [Google Scholar] [CrossRef]
- Pinkiewicz, M.; Zaczyński, A.; Walecki, J.; Zawadzki, M. Beyond the Walls of Troy: A Scoping Review on Pharmacological Strategies to Enhance Drug Delivery Across the Blood-Brain Barrier and Blood-Tumor Barrier. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Priest, R.; Ambady, P.; Neweult, E. ACTR-23. safety of intra-arterial chemotherapy with osmotic opening of the blood-brain barrier. Neuro-Oncology 2018, 20, vi16. [Google Scholar] [CrossRef]
- Ferreira, C.; Ferreira, M.Y.; Cardoso, L.J.C.; Scarramal, J.P.L.; Nogueira, A.; Wong, T.; Bokil, S.; Singh, F.; Massimo, S.; Albers, O.; et al. Safety and efficacy of selective and superselective intra-arterial cerebral infusion with blood-brain barrier disruption for glioma: a systematic review and meta-analysis. J Neurointerv Surg 2025. [Google Scholar] [CrossRef]
- Lei, K.; Zhou, L.; Dan, M.; Yang, F.; Jian, T.; Xin, J.; Yu, Z.; Wang, Y. Trojan Horse Delivery Strategies of Natural Medicine Monomers: Challenges and Limitations in Improving Brain Targeting. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- D'Amico, R.S.; Aghi, M.K.; Vogelbaum, M.A.; Bruce, J.N. Convection-enhanced drug delivery for glioblastoma: a review. J Neurooncol 2021, 151, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Jahangiri, A.; Chin, A.T.; Flanigan, P.M.; Chen, R.; Bankiewicz, K.; Aghi, M.K. Convection-enhanced delivery in glioblastoma: a review of preclinical and clinical studies. J Neurosurg 2017, 126, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.M.; Sonabend, A.M.; Bruce, J.N. Convection-Enhanced Delivery. Neurotherapeutics 2017, 14, 358–371. [Google Scholar] [CrossRef]
- Huttunen, K.M. Improving drug delivery to the brain: the prodrug approach. Expert Opin Drug Deliv 2024, 21, 683–693. [Google Scholar] [CrossRef]
- Puris, E.; Fricker, G.; Gynther, M. Targeting Transporters for Drug Delivery to the Brain: Can We Do Better? Pharm Res 2022, 39, 1415–1455. [Google Scholar] [CrossRef]
- Papakyriakopoulou, P.; Valsami, G. The nasal route for nose-to-brain drug delivery: advanced nasal formulations for CNS disorders. Expert Opin Drug Deliv 2025, 22, 823–839. [Google Scholar] [CrossRef]
- Chu, C.; Jablonska, A.; Gao, Y.; Lan, X.; Lesniak, W.G.; Liang, Y.; Liu, G.; Li, S.; Magnus, T.; Pearl, M.; et al. Hyperosmolar blood-brain barrier opening using intra-arterial injection of hyperosmotic mannitol in mice under real-time MRI guidance. Nat Protoc 2022, 17, 76–94. [Google Scholar] [CrossRef]
- Virtanen, P.S.; Ortiz, K.J.; Patel, A.; Blocher, W.A., 3rd; Richardson, A.M. Blood-Brain Barrier Disruption for the Treatment of Primary Brain Tumors: Advances in the Past Half-Decade. Curr Oncol Rep 2024, 26, 236–249. [Google Scholar] [CrossRef]
- Chen, T.; Wang, W.; Ramos, N.M.; Schonthal, A. EXTH-66. NEO100 TRANSIENTLY OPENS UP THE BLOOD BRAIN BARRIER VIA TIGHT JUNCTION INHIBITION. Neuro-Oncology 2020, 22, ii101. [Google Scholar] [CrossRef]
- Trevisani, M.; Berselli, A.; Alberini, G.; Centonze, E.; Vercellino, S.; Cartocci, V.; Millo, E.; Ciobanu, D.Z.; Braccia, C.; Armirotti, A.; et al. A claudin5-binding peptide enhances the permeability of the blood-brain barrier in vitro. Sci Adv 2025, 11, eadq2616. [Google Scholar] [CrossRef]
- Parvar, S.J.; Wong, C.I.; Lewis, A.; Szychot, E.; Morris, C.J.; Shorthouse, D.; Dziemidowicz, K. Convection-enhanced delivery for brain malignancies: Technical parameters, formulation strategies and clinical perspectives. Adv Drug Deliv Rev 2025, 224, 115657. [Google Scholar] [CrossRef]
- Lonser, R.R.; Sarntinoranont, M.; Morrison, P.F.; Oldfield, E.H. Convection-enhanced delivery to the central nervous system. J Neurosurg 2015, 122, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Puris, E.; Gynther, M.; Auriola, S.; Huttunen, K.M. L-Type amino acid transporter 1 as a target for drug delivery. Pharm Res 2020, 37, 88. [Google Scholar] [CrossRef] [PubMed]
- Chien, H.C.; Colas, C.; Finke, K.; Springer, S.; Stoner, L.; Zur, A.A.; Venteicher, B.; Campbell, J.; Hall, C.; Flint, A.; et al. Reevaluating the Substrate Specificity of the L-Type Amino Acid Transporter (LAT1). J Med Chem 2018, 61, 7358–7373. [Google Scholar] [CrossRef] [PubMed]
- Parvez, M.M.; Sadighi, A.; Ahn, Y.; Keller, S.F.; Enoru, J.O. Uptake Transporters at the Blood-Brain Barrier and Their Role in Brain Drug Disposition. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Bahrami, K.; Järvinen, J.; Laitinen, T.; Reinisalo, M.; Honkakoski, P.; Poso, A.; Huttunen, K.M.; Rautio, J. Structural Features Affecting the Interactions and Transportability of LAT1-Targeted Phenylalanine Drug Conjugates. Mol Pharm 2023, 20, 206–218. [Google Scholar] [CrossRef]
- Hugele, A.; Löffler, S.; Molina, B.H.; Guillon, M.; Montaser, A.B.; Auriola, S.; Huttunen, K.M. Aminopeptidase B can bioconvert L-type amino acid transporter 1 (LAT1)-utilizing amide prodrugs in the brain. Front Pharmacol 2022, 13, 1034964. [Google Scholar] [CrossRef]
- Huttunen, J.; Peltokangas, S.; Gynther, M.; Natunen, T.; Hiltunen, M.; Auriola, S.; Ruponen, M.; Vellonen, K.S.; Huttunen, K.M. L-Type Amino Acid Transporter 1 (LAT1/Lat1)-Utilizing Prodrugs Can Improve the Delivery of Drugs into Neurons, Astrocytes and Microglia. Sci Rep 2019, 9, 12860. [Google Scholar] [CrossRef]
- Montaser, A.B.; Järvinen, J.; Löffler, S.; Huttunen, J.; Auriola, S.; Lehtonen, M.; Jalkanen, A.; Huttunen, K.M. L-Type Amino Acid Transporter 1 Enables the Efficient Brain Delivery of Small-Sized Prodrug across the Blood-Brain Barrier and into Human and Mouse Brain Parenchymal Cells. ACS Chem Neurosci 2020, 11, 4301–4315. [Google Scholar] [CrossRef]
- Tampio, J.; Huttunen, J.; Montaser, A.; Huttunen, K.M. Targeting of Perforin Inhibitor into the Brain Parenchyma Via a Prodrug Approach Can Decrease Oxidative Stress and Neuroinflammation and Improve Cell Survival. Mol Neurobiol 2020, 57, 4563–4577. [Google Scholar] [CrossRef] [PubMed]
- Al Rihani, S.B.; Darakjian, L.I.; Deodhar, M.; Dow, P.; Turgeon, J.; Michaud, V. Disease-Induced Modulation of Drug Transporters at the Blood-Brain Barrier Level. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Huttunen, J.; Gynther, M.; Vellonen, K.S.; Huttunen, K.M. L-Type amino acid transporter 1 (LAT1)-utilizing prodrugs are carrier-selective despite having low affinity for organic anion transporting polypeptides (OATPs). Int J Pharm 2019, 571, 118714. [Google Scholar] [CrossRef] [PubMed]
- Huttunen, J.; Gynther, M.; Vellonen, K.-S.; Huttunen, K.M. L-Type amino acid transporter 1 (LAT1)-utilizing prodrugs are carrier-selective despite having low affinity for organic anion transporting polypeptides (OATPs). International Journal of Pharmaceutics 2019, 571, 118714. [Google Scholar] [CrossRef]
- van de Waterbeemd, H.; Smith, D.A.; Jones, B.C. Lipophilicity in PK design: methyl, ethyl, futile. J Comput Aided Mol Des 2001, 15, 273–286. [Google Scholar] [CrossRef]
- Patel, M.M.; Patel, B.M. Crossing the Blood-Brain Barrier: Recent Advances in Drug Delivery to the Brain. CNS Drugs 2017, 31, 109–133. [Google Scholar] [CrossRef]
- Sethi, B.; Kumar, V.; Mahato, K.; Coulter, D.W.; Mahato, R.I. Recent advances in drug delivery and targeting to the brain. J Control Release 2022, 350, 668–687. [Google Scholar] [CrossRef]
- Juhairiyah, F.; de Lange, E.C.M. Understanding Drug Delivery to the Brain Using Liposome-Based Strategies: Studies that Provide Mechanistic Insights Are Essential. Aaps j 2021, 23, 114. [Google Scholar] [CrossRef]
- Gonzaga, R.V.; do Nascimento, L.A.; Santos, S.S.; Machado Sanches, B.A.; Giarolla, J.; Ferreira, E.I. Perspectives About Self-Immolative Drug Delivery Systems. J Pharm Sci 2020, 109, 3262–3281. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, S.; Cheng, G.; Yu, P.; Chang, J.; Chen, X. Cascade Drug-Release Strategy for Enhanced Anticancer Therapy. Matter 2021, 4, 26–53. [Google Scholar] [CrossRef]
- Muñoz-Sánchez, S.; Gong, J.; de la Mata, F.J.; Gillies, E.R.; García-Gallego, S. Functional Self-Immolative Hydrogels with Dendritic Cross-Linkers for Controlled Drug Delivery. Chem Mater 2025, 37, 5814–5824. [Google Scholar] [CrossRef] [PubMed]
- van der Westhuyzen, A.E.; Hodson, L.E.; Pashikanti, G.; Fritzemeier, R.; Yeung, S.B.; Mancia, A.; Lian, D.R.; Tholath, P.J.; Paez, A.C.; Chavan, L.N.; et al. Rotamer-Controlled Self-Immolative Linkers Enable Tunable Release of Neurosteroid Oxime Prodrugs. ACS Med Chem Lett 2025, 16, 2022–2031. [Google Scholar] [CrossRef]
- Wang, H.; Zheng, C.; Tian, F.; Xiao, Z.; Sun, Z.; Lu, L.; Dai, W.; Zhang, Q.; Mei, X. Improving the Dissolution Rate and Bioavailability of Curcumin via Co-Crystallization. Pharmaceuticals (Basel) 2024, 17. [Google Scholar] [CrossRef] [PubMed]
- Nicolaescu, O.E.; Belu, I.; Mocanu, A.G.; Manda, V.C.; Rău, G.; Pîrvu, A.S.; Ionescu, C.; Ciulu-Costinescu, F.; Popescu, M.; Ciocîlteu, M.V. Cyclodextrins: Enhancing Drug Delivery, Solubility and Bioavailability for Modern Therapeutics. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- Lanevskij, K.; Japertas, P.; Didziapetris, R.; Petrauskas, A. Ionization-specific prediction of blood-brain permeability. J Pharm Sci 2009, 98, 122–134. [Google Scholar] [CrossRef]
- Xing, Y.; Meng, B.; Chen, Q. Cyclodextrin-Containing Drug Delivery Systems and Their Applications in Neurodegenerative Disorders. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Văruț, R.M.; Popescu, A.I.S.; Gaman, S.; Niculescu, C.E.; Niculescu, A.; Dop, D.; Stepan, M.D.; Ionovici, N.; Singer, C.E.; Popescu, C. Cyclodextrin-Based Drug Delivery Systems for Depression: Improving Antidepressant Bioavailability and Targeted Central Nervous System Delivery. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Thiberville, L.; Faivre, V.; Sizun, C.; Dehouck, M.P.; Landry, C.; Baati, R.; Tsapis, N. Cyclodextrin-based formulations for delivering broad-spectrum nerve agent antidote to the central nervous system: stability, physicochemical characterization and application in a human blood-brain barrier model. Int J Pharm 2025, 674, 125505. [Google Scholar] [CrossRef]
- Guo, M.; Sun, X.; Chen, J.; Cai, T. Pharmaceutical cocrystals: A review of preparations, physicochemical properties and applications. Acta Pharm Sin B 2021, 11, 2537–2564. [Google Scholar] [CrossRef]
- Spiridon, I.; Anghel, N. Cyclodextrins as Multifunctional Platforms in Drug Delivery and Beyond: Structural Features, Functional Applications, and Future Trends. Molecules 2025, 30. [Google Scholar] [CrossRef]
- Duan, C.; Liu, W.; Tao, Y.; Liang, F.; Chen, Y.; Xiao, X.; Zhang, G.; Chen, Y.; Hao, C. Two Novel Palbociclib-Resorcinol and Palbociclib-Orcinol Cocrystals with Enhanced Solubility and Dissolution Rate. Pharmaceutics 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.; Weyers, M.; Steenekamp, J.H.; Steyn, J.D.; Gouws, C.; Hamman, J.H. Drug Bioavailability Enhancing Agents of Natural Origin (Bioenhancers) that Modulate Drug Membrane Permeation and Pre-Systemic Metabolism. Pharmaceutics 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Fu, B.M.; Zhang, Z.J. Borneol, a novel agent that improves central nervous system drug delivery by enhancing blood-brain barrier permeability. Drug Deliv 2017, 24, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Neaz, S.; Alam, M.M.; Imran, A.B. Advancements in cyclodextrin-based controlled drug delivery: Insights into pharmacokinetic and pharmacodynamic profiles. Heliyon 2024, 10, e39917. [Google Scholar] [CrossRef]
- de Souza, M.M.; Gini, A.L.R.; Moura, J.A.; Scarim, C.B.; Chin, C.M.; Dos Santos, J.L. Prodrug Approach as a Strategy to Enhance Drug Permeability. Pharmaceuticals (Basel) 2025, 18. [Google Scholar] [CrossRef]
- Dong, X.; Brahma, R.K.; Fang, C.; Yao, S.Q. Stimulus-responsive self-assembled prodrugs in cancer therapy. Chem Sci 2022, 13, 4239–4269. [Google Scholar] [CrossRef]
- Kopeček, J.; Yang, J. Polymer nanomedicines. Adv Drug Deliv Rev 2020, 156, 40–64. [Google Scholar] [CrossRef]
- Guo, H.; Mi, P. Polymer-drug and polymer-protein conjugated nanocarriers: Design, drug delivery, imaging, therapy, and clinical applications. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2024, 16, e1988. [Google Scholar] [CrossRef]
- Jiang, Y.; Stenzel, M. Drug Delivery Vehicles Based on Albumin-Polymer Conjugates. Macromol Biosci 2016, 16, 791–802. [Google Scholar] [CrossRef]
- Mi, P. Stimuli-responsive nanocarriers for drug delivery, tumor imaging, therapy and theranostics. Theranostics 2020, 10, 4557–4588. [Google Scholar] [CrossRef]
- Lee, Y.; Thompson, D.H. Stimuli-responsive liposomes for drug delivery. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2017, 9. [Google Scholar] [CrossRef]
- Wu, X.; Shu, Y.; Zheng, Y.; Zhang, P.; Cong, H.; Zou, Y.; Cai, H.; Zha, Z. Recent Advances in Nanomedicine: Cutting-Edge Research on Nano-PROTAC Delivery Systems for Cancer Therapy. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Li, X.; Peng, X.; Zoulikha, M.; Boafo, G.F.; Magar, K.T.; Ju, Y.; He, W. Multifunctional nanoparticle-mediated combining therapy for human diseases. Signal Transduct Target Ther 2024, 9, 1. [Google Scholar] [CrossRef]
- Đorđević, S.; Gonzalez, M.M.; Conejos-Sánchez, I.; Carreira, B.; Pozzi, S.; Acúrcio, R.C.; Satchi-Fainaro, R.; Florindo, H.F.; Vicent, M.J. Current hurdles to the translation of nanomedicines from bench to the clinic. Drug Deliv Transl Res 2022, 12, 500–525. [Google Scholar] [CrossRef]
- Ahmad, A.; Imran, M.; Sharma, N. Precision Nanotoxicology in Drug Development: Current Trends and Challenges in Safety and Toxicity Implications of Customized Multifunctional Nanocarriers for Drug-Delivery Applications. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Buya, A.B.; Mahlangu, P.; Witika, B.A. From lab to industrial development of lipid nanocarriers using quality by design approach. Int J Pharm X 2024, 8, 100266. [Google Scholar] [CrossRef]
- Sammasagi, S.S.; Sutar, K.P.; Hooli, S. Scale-up and quality control challenges in the industrial manufacturing of nanoformulations: current trends and future perspectives. IJSAT-International Journal on Science and Technology 2025, 16. [Google Scholar]
- Gyimesi, G.; Hediger, M.A. Transporter-Mediated Drug Delivery. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Puris, E.; Gynther, M.; Huttunen, J.; Petsalo, A.; Huttunen, K.M. L-type amino acid transporter 1 utilizing prodrugs: How to achieve effective brain delivery and low systemic exposure of drugs. J Control Release 2017, 261, 93–104. [Google Scholar] [CrossRef]
- Montaser, A.; Lehtonen, M.; Gynther, M.; Huttunen, K.M. L-Type Amino Acid Transporter 1-Utilizing Prodrugs of Ketoprofen Can Efficiently Reduce Brain Prostaglandin Levels. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- Patching, S.G. Glucose Transporters at the Blood-Brain Barrier: Function, Regulation and Gateways for Drug Delivery. Mol Neurobiol 2017, 54, 1046–1077. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Miura, Y.; Mochida, Y.; Miyazaki, T.; Toh, K.; Anraku, Y.; Melo, V.; Liu, X.; Ishii, T.; Nagano, O.; et al. Glucose transporter 1-mediated vascular translocation of nanomedicines enhances accumulation and efficacy in solid tumors. J Control Release 2019, 301, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Vijay, N.; Morris, M.E. Role of monocarboxylate transporters in drug delivery to the brain. Curr Pharm Des 2014, 20, 1487–1498. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, L.; Jiang, Q.; Sun, Y.; Zhao, D.; Sun, M.; He, Z.; Sun, J.; Wang, Y. Intestinal OCTN2- and MCT1-targeted drug delivery to improve oral bioavailability. Asian J Pharm Sci 2020, 15, 158–173. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, D.; Wang, G.; Jiang, Q.; Guo, M.; Kan, Q.; He, Z.; Sun, J. A novel oral prodrug-targeting transporter MCT 1: 5-fluorouracil-dicarboxylate monoester conjugates. Asian J Pharm Sci 2019, 14, 631–639. [Google Scholar] [CrossRef]
- Hu, C.; Tao, L.; Cao, X.; Chen, L. The solute carrier transporters and the brain: Physiological and pharmacological implications. Asian J Pharm Sci 2020, 15, 131–144. [Google Scholar] [CrossRef]
- Huttunen, J.; Adla, S.K.; Markowicz-Piasecka, M.; Huttunen, K.M. Increased/Targeted Brain (Pro)Drug Delivery via Utilization of Solute Carriers (SLCs). Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Begley, D.J. ABC transporters and the blood-brain barrier. Curr Pharm Des 2004, 10, 1295–1312. [Google Scholar] [CrossRef]
- Colclough, N.; Alluri, R.V.; Tucker, J.W.; Gozalpour, E.; Li, D.; Du, H.; Li, W.; Harlfinger, S.; O'Neill, D.J.; Sproat, G.G.; et al. Utilizing a Dual Human Transporter MDCKII-MDR1-BCRP Cell Line to Assess Efflux at the Blood Brain Barrier. Drug Metab Dispos 2024, 52, 95–105. [Google Scholar] [CrossRef]
- Tian, J.; Han, Z.; Song, D.; Peng, Y.; Xiong, M.; Chen, Z.; Duan, S.; Zhang, L. Engineered Exosome for Drug Delivery: Recent Development and Clinical Applications. Int J Nanomedicine 2023, 18, 7923–7940. [Google Scholar] [CrossRef]
- Koh, H.B.; Kim, H.J.; Kang, S.W.; Yoo, T.H. Exosome-Based Drug Delivery: Translation from Bench to Clinic. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Zeng, H.; Guo, S.; Ren, X.; Wu, Z.; Liu, S.; Yao, X. Current Strategies for Exosome Cargo Loading and Targeting Delivery. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Hade, M.D.; Suire, C.N.; Suo, Z. Mesenchymal Stem Cell-Derived Exosomes: Applications in Regenerative Medicine. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Wu, J.; Zhang, L.; Huang, N.; Luo, Y. A New Strategy for the Regulation of Neuroinflammation: Exosomes Derived from Mesenchymal Stem Cells. Cell Mol Neurobiol 2024, 44, 24. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Abouelnazar, F.A.; Iqbal, M.A.; Pan, J.; Zheng, X.; Chen, T.; Shen, W.; Yin, J.; Yan, Y.; Liu, P.; et al. Mesenchymal stem cell-derived exosomes as a plausible immunomodulatory therapeutic tool for inflammatory diseases. Front Cell Dev Biol 2025, 13, 1563427. [Google Scholar] [CrossRef]
- Fu, S.; Wang, Y.; Xia, X.; Zheng, J.C. Exosome engineering: Current progress in cargo loading and targeted delivery. NanoImpact 2020, 20, 100261. [Google Scholar] [CrossRef]
- Xi, X.M.; Xia, S.J.; Lu, R. Drug loading techniques for exosome-based drug delivery systems. Pharmazie 2021, 76, 61–67. [Google Scholar] [CrossRef]
- Chen, C.; Sun, M.; Wang, J.; Su, L.; Lin, J.; Yan, X. Active cargo loading into extracellular vesicles: Highlights the heterogeneous encapsulation behaviour. J Extracell Vesicles 2021, 10, e12163. [Google Scholar] [CrossRef]
- Ahmed, W.; Mushtaq, A.; Ali, S.; Khan, N.; Liang, Y.; Duan, L. Engineering Approaches for Exosome Cargo Loading and Targeted Delivery: Biological versus Chemical Perspectives. ACS Biomater Sci Eng 2024, 10, 5960–5976. [Google Scholar] [CrossRef]
- Kimiz-Gebologlu, I.; Oncel, S.S. Exosomes: Large-scale production, isolation, drug loading efficiency, and biodistribution and uptake. J Control Release 2022, 347, 533–543. [Google Scholar] [CrossRef]
- Ahn, S.H.; Ryu, S.W.; Choi, H.; You, S.; Park, J.; Choi, C. Manufacturing Therapeutic Exosomes: from Bench to Industry. Mol Cells 2022, 45, 284–290. [Google Scholar] [CrossRef]
- Wang, C.K.; Tsai, T.H.; Lee, C.H. Regulation of exosomes as biologic medicines: Regulatory challenges faced in exosome development and manufacturing processes. Clin Transl Sci 2024, 17, e13904. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Arora, S. Navigating the Global Regulatory Landscape for Exosome-Based Therapeutics: Challenges, Strategies, and Future Directions. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- Lian, M.Q.; Chng, W.H.; Liang, J.; Yeo, H.Q.; Lee, C.K.; Belaid, M.; Tollemeto, M.; Wacker, M.G.; Czarny, B.; Pastorin, G. Plant-derived extracellular vesicles: Recent advancements and current challenges on their use for biomedical applications. J Extracell Vesicles 2022, 11, e12283. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.J.; Wang, M.H.; Ho, C.T.; Pan, M.H. Plant-Derived Extracellular Vesicles: A New Revolutionization of Modern Healthy Diets and Biomedical Applications. J Agric Food Chem 2024, 72, 2853–2878. [Google Scholar] [CrossRef]
- Kim, M.; Jang, H.; Kim, W.; Kim, D.; Park, J.H. Therapeutic Applications of Plant-Derived Extracellular Vesicles as Antioxidants for Oxidative Stress-Related Diseases. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Karamanidou, T.; Tsouknidas, A. Plant-Derived Extracellular Vesicles as Therapeutic Nanocarriers. Int J Mol Sci 2021, 23. [Google Scholar] [CrossRef]
- Alzahrani, F.A.; Khan, M.I.; Kameli, N.; Alsahafi, E.; Riza, Y.M. Plant-Derived Extracellular Vesicles and Their Exciting Potential as the Future of Next-Generation Drug Delivery. Biomolecules 2023, 13. [Google Scholar] [CrossRef]
- Shao, M.; Jin, X.; Chen, S.; Yang, N.; Feng, G. Plant-derived extracellular vesicles -a novel clinical anti-inflammatory drug carrier worthy of investigation. Biomed Pharmacother 2023, 169, 115904. [Google Scholar] [CrossRef]
- Calzoni, E.; Bertoldi, A.; Cusumano, G.; Buratta, S.; Urbanelli, L.; Emiliani, C. Plant-derived extracellular vesicles: natural nanocarriers for biotechnological drugs. Processes 2024, 12, 2938. [Google Scholar] [CrossRef]
- Zuo, Y.; Zhang, J.; Sun, B.; Wang, X.; Wang, R.; Tian, S.; Miao, M. A New Perspective on Regenerative Medicine: Plant-Derived Extracellular Vesicles. Biomolecules 2025, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhao, J.; Ding, H.; Qiu, M.; Xue, L.; Ge, D.; Wen, G.; Ren, H.; Li, P.; Wang, J. Applications of plant-derived extracellular vesicles in medicine. MedComm (2020) 2024, 5, e741. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Chen, L.; Li, W.; Chang, C.J. Anti-inflammatory and antioxidative effects of Perilla frutescens-derived extracellular vesicles: Insights from Zebrafish models. Mol Immunol 2025, 182, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.Y.; Chuang, C.W.; Huang, Y.C.; Huang, C.J. Therapeutic potential of plant-derived small extracellular vesicles in sepsis: A network meta-analysis. Pharmacol Res 2025, 217, 107795. [Google Scholar] [CrossRef]
- Cui, L.; Perini, G.; Minopoli, A.; Palmieri, V.; De Spirito, M.; Papi, M. Plant-derived extracellular vesicles as a natural drug delivery platform for glioblastoma therapy: A dual role in preserving endothelial integrity while modulating the tumor microenvironment. Int J Pharm X 2025, 10, 100349. [Google Scholar] [CrossRef]
- Xu, X.; Xu, L.; Wen, C.; Xia, J.; Zhang, Y.; Liang, Y. Programming assembly of biomimetic exosomes: An emerging theranostic nanomedicine platform. Mater Today Bio 2023, 22, 100760. [Google Scholar] [CrossRef]
- Lu, M.; Huang, Y. Bioinspired exosome-like therapeutics and delivery nanoplatforms. Biomaterials 2020, 242, 119925. [Google Scholar] [CrossRef]
- Chen, Y.; Douanne, N.; Wu, T.; Kaur, I.; Tsering, T.; Erzingatzian, A.; Nadeau, A.; Juncker, D.; Nerguizian, V.; Burnier, J.V. Leveraging nature's nanocarriers: Translating insights from extracellular vesicles to biomimetic synthetic vesicles for biomedical applications. Sci Adv 2025, 11, eads5249. [Google Scholar] [CrossRef]
- Poinsot, V.; Pizzinat, N.; Ong-Meang, V. Engineered and Mimicked Extracellular Nanovesicles for Therapeutic Delivery. Nanomaterials (Basel) 2024, 14. [Google Scholar] [CrossRef]
- Antimisiaris, S.G.; Mourtas, S.; Marazioti, A. Exosomes and Exosome-Inspired Vesicles for Targeted Drug Delivery. Pharmaceutics 2018, 10. [Google Scholar] [CrossRef]
- Kooijmans, S.A.; Vader, P.; van Dommelen, S.M.; van Solinge, W.W.; Schiffelers, R.M. Exosome mimetics: a novel class of drug delivery systems. Int J Nanomedicine 2012, 7, 1525–1541. [Google Scholar] [CrossRef]
- Rayamajhi, S.; Nguyen, T.D.T.; Marasini, R.; Aryal, S. Macrophage-derived exosome-mimetic hybrid vesicles for tumor targeted drug delivery. Acta Biomater 2019, 94, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Ailuno, G.; Baldassari, S.; Lai, F.; Florio, T.; Caviglioli, G. Exosomes and Extracellular Vesicles as Emerging Theranostic Platforms in Cancer Research. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Jagtiani, E.; Yeolekar, M.; Naik, S.; Patravale, V. In vitro blood brain barrier models: An overview. J Control Release 2022, 343, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Sivandzade, F.; Cucullo, L. In-vitro blood-brain barrier modeling: A review of modern and fast-advancing technologies. J Cereb Blood Flow Metab 2018, 38, 1667–1681. [Google Scholar] [CrossRef]
- Wang, Y.I.; Abaci, H.E.; Shuler, M.L. Microfluidic blood-brain barrier model provides in vivo-like barrier properties for drug permeability screening. Biotechnol Bioeng 2017, 114, 184–194. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Humle, N.; Hede, E.; Moos, T.; Burkhart, A.; Thomsen, L.B. The blood-brain barrier studied in vitro across species. PLoS One 2021, 16, e0236770. [Google Scholar] [CrossRef]
- Bernard-Patrzynski, F.; Lécuyer, M.A.; Puscas, I.; Boukhatem, I.; Charabati, M.; Bourbonnière, L.; Ramassamy, C.; Leclair, G.; Prat, A.; Roullin, V.G. Isolation of endothelial cells, pericytes and astrocytes from mouse brain. PLoS One 2019, 14, e0226302. [Google Scholar] [CrossRef]
- Park, J.S.; Choe, K.; Khan, A.; Jo, M.H.; Park, H.Y.; Kang, M.H.; Park, T.J.; Kim, M.O. Establishing Co-Culture Blood-Brain Barrier Models for Different Neurodegeneration Conditions to Understand Its Effect on BBB Integrity. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Jiang, L.; Li, S.; Zheng, J.; Li, Y.; Huang, H. Recent Progress in Microfluidic Models of the Blood-Brain Barrier. Micromachines (Basel) 2019, 10. [Google Scholar] [CrossRef]
- Goldeman, C.; Andersen, M.; Al-Robai, A.; Buchholtz, T.; Svane, N.; Ozgür, B.; Holst, B.; Shusta, E.; Hall, V.J.; Saaby, L.; et al. Human induced pluripotent stem cells (BIONi010-C) generate tight cell monolayers with blood-brain barrier traits and functional expression of large neutral amino acid transporter 1 (SLC7A5). Eur J Pharm Sci 2021, 156, 105577. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.R.; Gromnicova, R.; Brachner, A.; Kraev, I.; Romero, I.A.; Neuhaus, W.; Male, D. A hydrogel model of the human blood-brain barrier using differentiated stem cells. PLoS One 2023, 18, e0283954. [Google Scholar] [CrossRef] [PubMed]
- Campisi, M.; Shin, Y.; Osaki, T.; Hajal, C.; Chiono, V.; Kamm, R.D. 3D self-organized microvascular model of the human blood-brain barrier with endothelial cells, pericytes and astrocytes. Biomaterials 2018, 180, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER measurement techniques for in vitro barrier model systems. J Lab Autom 2015, 20, 107–126. [Google Scholar] [CrossRef]
- Andjelkovic, A.V.; Stamatovic, S.M.; Phillips, C.M.; Martinez-Revollar, G.; Keep, R.F. Modeling blood-brain barrier pathology in cerebrovascular disease in vitro: current and future paradigms. Fluids Barriers CNS 2020, 17, 44. [Google Scholar] [CrossRef]
- O'Brown, N.M.; Pfau, S.J.; Gu, C. Bridging barriers: a comparative look at the blood-brain barrier across organisms. Genes Dev 2018, 32, 466–478. [Google Scholar] [CrossRef]
- Watanabe, D.; Nakagawa, S.; Morofuji, Y.; Tóth, A.E.; Vastag, M.; Aruga, J.; Niwa, M.; Deli, M.A. Characterization of a Primate Blood-Brain Barrier Co-Culture Model Prepared from Primary Brain Endothelial Cells, Pericytes and Astrocytes. Pharmaceutics 2021, 13. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Q.Y.; Haqqani, A.S.; Liu, Z.; Sodja, C.; Leclerc, S.; Baumann, E.; Delaney, C.E.; Brunette, E.; Stanimirovic, D.B. Differential Expression of ABC Transporter Genes in Brain Vessels vs. Peripheral Tissues and Vessels from Human, Mouse and Rat. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Hoshi, Y.; Uchida, Y.; Tachikawa, M.; Inoue, T.; Ohtsuki, S.; Terasaki, T. Quantitative atlas of blood-brain barrier transporters, receptors, and tight junction proteins in rats and common marmoset. J Pharm Sci 2013, 102, 3343–3355. [Google Scholar] [CrossRef]
- Hussner, J.; Foletti, A.; Seibert, I.; Fuchs, A.; Schuler, E.; Malagnino, V.; Grube, M.; Meyer Zu Schwabedissen, H.E. Differences in transport function of the human and rat orthologue of the Organic Anion Transporting Polypeptide 2B1 (OATP2B1). Drug Metab Pharmacokinet 2021, 41, 100418. [Google Scholar] [CrossRef]
- Strazielle, N.; Blondel, S.; Confais, J.; El Khoury, R.; Contamin, H.; Ghersi-Egea, J.F. Molecular determinants of neuroprotection in blood-brain interfaces of the cynomolgus monkey. Front Pharmacol 2025, 16, 1523819. [Google Scholar] [CrossRef] [PubMed]
- Chaves, C.; Do, T.M.; Cegarra, C.; Roudières, V.; Tolou, S.; Thill, G.; Rocher, C.; Didier, M.; Lesuisse, D. Non-Human Primate Blood-Brain Barrier and In Vitro Brain Endothelium: From Transcriptome to the Establishment of a New Model. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, M.B.; Anishchenko, A.M.; Khlebnikov, A.I.; Schepetkin, I.A. Regulation of Blood-Brain Barrier Permeability via JNK Signaling Pathway: Mechanisms and Potential Therapeutic Strategies for Ischemic Stroke, Alzheimer's Disease and Brain Tumors. Molecules 2025, 30. [Google Scholar] [CrossRef] [PubMed]
- Vazana, U.; Veksler, R.; Pell, G.S.; Prager, O.; Fassler, M.; Chassidim, Y.; Roth, Y.; Shahar, H.; Zangen, A.; Raccah, R.; et al. Glutamate-Mediated Blood-Brain Barrier Opening: Implications for Neuroprotection and Drug Delivery. J Neurosci 2016, 36, 7727–7739. [Google Scholar] [CrossRef]
- Brynildsen, J.K.; Rajan, K.; Henderson, M.X.; Bassett, D.S. Network models to enhance the translational impact of cross-species studies. Nat Rev Neurosci 2023, 24, 575–588. [Google Scholar] [CrossRef]
- de Lange, E.C.M.; van den Brink, W.; Yamamoto, Y.; de Witte, W.E.A.; Wong, Y.C. Novel CNS drug discovery and development approach: model-based integration to predict neuro-pharmacokinetics and pharmacodynamics. Expert Opin Drug Discov 2017, 12, 1207–1218. [Google Scholar] [CrossRef]
- Vallianatou, T.; Tsopelas, F.; Tsantili-Kakoulidou, A. Prediction Models for Brain Distribution of Drugs Based on Biomimetic Chromatographic Data. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Loryan, I.; Reichel, A.; Feng, B.; Bundgaard, C.; Shaffer, C.; Kalvass, C.; Bednarczyk, D.; Morrison, D.; Lesuisse, D.; Hoppe, E.; et al. Unbound Brain-to-Plasma Partition Coefficient, K(p,uu,brain)-a Game Changing Parameter for CNS Drug Discovery and Development. Pharm Res 2022, 39, 1321–1341. [Google Scholar] [CrossRef]
- Kalvass, J.C.; Maurer, T.S.; Pollack, G.M. Use of plasma and brain unbound fractions to assess the extent of brain distribution of 34 drugs: comparison of unbound concentration ratios to in vivo p-glycoprotein efflux ratios. Drug Metab Dispos 2007, 35, 660–666. [Google Scholar] [CrossRef]
- Li, J.; Jiang, J.; Wu, J.; Bao, X.; Sanai, N. Physiologically Based Pharmacokinetic Modeling of Central Nervous System Pharmacokinetics of CDK4/6 Inhibitors to Guide Selection of Drug and Dosing Regimen for Brain Cancer Treatment. Clin Pharmacol Ther 2021, 109, 494–506. [Google Scholar] [CrossRef]
- Sundheimer, J.K.; Benzel, J.; Longuespée, R.; Burhenne, J.; Pfister, S.M.; Maaß, K.K.; Sauter, M.; Pajtler, K.W. Experimental Insights and Recommendations for Successfully Performing Cerebral Microdialysis With Hydrophobic Drug Candidates. Clinical and translational science 2025, 18, e70226. [Google Scholar] [CrossRef] [PubMed]
- Fridén, M.; Gupta, A.; Antonsson, M.; Bredberg, U.; Hammarlund-Udenaes, M. In vitro methods for estimating unbound drug concentrations in the brain interstitial and intracellular fluids. Drug Metab Dispos 2007, 35, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Van Natta, K.; Yeo, H.; Vilenski, O.; Weller, P.E.; Worboys, P.D.; Monshouwer, M. Unbound drug concentration in brain homogenate and cerebral spinal fluid at steady state as a surrogate for unbound concentration in brain interstitial fluid. Drug Metab Dispos 2009, 37, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Välitalo, P.A.; Wong, Y.C.; Huntjens, D.R.; Proost, J.H.; Vermeulen, A.; Krauwinkel, W.; Beukers, M.W.; Kokki, H.; Kokki, M.; et al. Prediction of human CNS pharmacokinetics using a physiologically-based pharmacokinetic modeling approach. Eur J Pharm Sci 2018, 112, 168–179. [Google Scholar] [CrossRef]
- Mu, R.J.; Liu, T.L.; Liu, X.D.; Liu, L. PBPK-PD model for predicting morphine pharmacokinetics, CNS effects and naloxone antagonism in humans. Acta Pharmacol Sin 2024, 45, 1752–1764. [Google Scholar] [CrossRef]
- Wu, K.; Li, X.; Zhou, Z.; Zhao, Y.; Su, M.; Cheng, Z.; Wu, X.; Huang, Z.; Jin, X.; Li, J.; et al. Predicting pharmacodynamic effects through early drug discovery with artificial intelligence-physiologically based pharmacokinetic (AI-PBPK) modelling. Front Pharmacol 2024, 15, 1330855. [Google Scholar] [CrossRef]
- Zou, Y.; Yuan, H.; Guo, Z.; Guo, T.; Fu, Z.; Wang, R.; Xu, D.; Wang, Q.; Wang, T.; Chen, L. Predicting the Brain-To-Plasma Unbound Partition Coefficient of Compounds via Formula-Guided Network. J Chem Inf Model 2025, 65, 5099–5112. [Google Scholar] [CrossRef]
- Chung, K.J.; Abdelhafez, Y.G.; Spencer, B.A.; Jones, T.; Tran, Q.; Nardo, L.; Chen, M.S., Jr.; Sarkar, S.; Medici, V.; Lyo, V.; et al. Quantitative PET imaging and modeling of molecular blood-brain barrier permeability. Nat Commun 2025, 16, 3076. [Google Scholar] [CrossRef]
- Mishra, V.; Kumari, N.; Vyas, M.; Aljabali, A.A.A.; Chattaraj, A.; Mishra, Y. Advances in multimodal imaging techniques in nanomedicine: enhancing drug delivery precision. RSC Adv 2025, 15, 27187–27209. [Google Scholar] [CrossRef]
- Varatharaj, A.; Liljeroth, M.; Darekar, A.; Larsson, H.B.W.; Galea, I.; Cramer, S.P. Blood-brain barrier permeability measured using dynamic contrast-enhanced magnetic resonance imaging: a validation study. J Physiol 2019, 597, 699–709. [Google Scholar] [CrossRef]
- Mishra, A.; Payne, C.; Carrascal-Miniño, A.; Sunassee, K.; Halbherr, S.; Pouliopoulos, A.N.; de Rosales, R.T. PET imaging for non-invasive monitoring of 89Zr-Talidox delivery to the brain following focused ultrasound-mediated blood-brain barrier opening. Journal of Controlled Release 2025, 387, 114183. [Google Scholar] [CrossRef]
- Hugon, G.; Goutal, S.; Dauba, A.; Breuil, L.; Larrat, B.; Winkeler, A.; Novell, A.; Tournier, N. [(18)F]2-Fluoro-2-deoxy-sorbitol PET Imaging for Quantitative Monitoring of Enhanced Blood-Brain Barrier Permeability Induced by Focused Ultrasound. Pharmaceutics 2021, 13. [Google Scholar] [CrossRef]
- Pa, C.; Shen, S.; Dai, Y.; Wu, M. Bibliometric analysis of neural injury biomarkers in neurodegenerative diseases: research trends and future perspectives. Front Hum Neurosci 2025, 19, 1614132. [Google Scholar] [CrossRef] [PubMed]
- Gerber, K.S.; Alvarez, G.; Alamian, A.; Behar-Zusman, V.; Downs, C.A. Biomarkers of Neuroinflammation in Traumatic Brain Injury. Clin Nurs Res 2022, 31, 1203–1218. [Google Scholar] [CrossRef] [PubMed]
- Mondello, S.; Guedes, V.A.; Lai, C.; Czeiter, E.; Amrein, K.; Kobeissy, F.; Mechref, Y.; Jeromin, A.; Mithani, S.; Martin, C.; et al. Circulating Brain Injury Exosomal Proteins following Moderate-To-Severe Traumatic Brain Injury: Temporal Profile, Outcome Prediction and Therapy Implications. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Lopez, B.G.C.; Kohale, I.N.; Du, Z.; Korsunsky, I.; Abdelmoula, W.M.; Dai, Y.; Stopka, S.A.; Gaglia, G.; Randall, E.C.; Regan, M.S.; et al. Multimodal platform for assessing drug distribution and response in clinical trials. Neuro Oncol 2022, 24, 64–77. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Leme Boaro, B.; da Silva Camarinha Oliveira, J.; Patočka, J.; Barbalho Lamas, C.; Tanaka, M.; Laurindo, L.F. Molecular Mechanisms Underlying Neuroinflammation Intervention with Medicinal Plants: A Critical and Narrative Review of the Current Literature. Pharmaceuticals (Basel) 2025, 18. [Google Scholar] [CrossRef]
- Tanaka, M.; Battaglia, S.; Liloia, D. Navigating Neurodegeneration: Integrating Biomarkers, Neuroinflammation, and Imaging in Parkinson's, Alzheimer's, and Motor Neuron Disorders. Biomedicines 2025, 13. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Laurindo, L.F.; de Oliveira Zanuso, B.; da Silva, R.M.S.; Gallerani Caglioni, L.; Nunes Junqueira de Moraes, V.B.F.; Fornari Laurindo, L.; Dogani Rodrigues, V.; da Silva Camarinha Oliveira, J.; Beluce, M.E.; et al. AdipoRon's Impact on Alzheimer's Disease-A Systematic Review and Meta-Analysis. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Tanaka, M. From Serendipity to Precision: Integrating AI, Multi-Omics, and Human-Specific Models for Personalized Neuropsychiatric Care. Biomedicines 2025, 13. [Google Scholar] [CrossRef]
- Shamul, J.G.; Wang, Z.; Gong, H.; Ou, W.; White, A.M.; Moniz-Garcia, D.P.; Gu, S.; Clyne, A.M.; Quiñones-Hinojosa, A.; He, X. Meta-analysis of the make-up and properties of in vitro models of the healthy and diseased blood-brain barrier. Nat Biomed Eng 2025, 9, 566–598. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.L.; England, T.J.; O'Sullivan, S.E. A Novel Transwell Blood Brain Barrier Model Using Primary Human Cells. Front Cell Neurosci 2019, 13, 230. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, Z.; Chen, F.; Su, J.; Han, J.; Liu, S. Using Regular Porous Membrane-Based Blood-Brain Barrier Model to Screen Brain-Targeted Drugs with Nanochannel Electrochemistry. Anal Chem 2025, 97, 7968–7977. [Google Scholar] [CrossRef] [PubMed]
- Burgio, F.; Gaiser, C.; Brady, K.; Gatta, V.; Class, R.; Schrage, R.; Suter-Dick, L. A Perfused In Vitro Human iPSC-Derived Blood-Brain Barrier Faithfully Mimics Transferrin Receptor-Mediated Transcytosis of Therapeutic Antibodies. Cell Mol Neurobiol 2023, 43, 4173–4187. [Google Scholar] [CrossRef]
- Ohshima, M.; Kamei, S.; Fushimi, H.; Mima, S.; Yamada, T.; Yamamoto, T. Prediction of Drug Permeability Using In Vitro Blood-Brain Barrier Models with Human Induced Pluripotent Stem Cell-Derived Brain Microvascular Endothelial Cells. Biores Open Access 2019, 8, 200–209. [Google Scholar] [CrossRef]
- Workman, M.J.; Svendsen, C.N. Recent advances in human iPSC-derived models of the blood-brain barrier. Fluids Barriers CNS 2020, 17, 30. [Google Scholar] [CrossRef]
- Mathew-Schmitt, S.; Oerter, S.; Reitenbach, E.; Gätzner, S.; Höchner, A.; Jahnke, H.G.; Piontek, J.; Neuhaus, W.; Brachner, A.; Metzger, M.; et al. Generation of Advanced Blood-Brain Barrier Spheroids Using Human-Induced Pluripotent Stem Cell-Derived Brain Capillary Endothelial-Like Cells. Adv Biol (Weinh) 2025, 9, e2400442. [Google Scholar] [CrossRef]
- Cho, C.F.; Wolfe, J.M.; Fadzen, C.M.; Calligaris, D.; Hornburg, K.; Chiocca, E.A.; Agar, N.Y.R.; Pentelute, B.L.; Lawler, S.E. Blood-brain-barrier spheroids as an in vitro screening platform for brain-penetrating agents. Nat Commun 2017, 8, 15623. [Google Scholar] [CrossRef]
- Logan, S.; Arzua, T.; Canfield, S.G.; Seminary, E.R.; Sison, S.L.; Ebert, A.D.; Bai, X. Studying Human Neurological Disorders Using Induced Pluripotent Stem Cells: From 2D Monolayer to 3D Organoid and Blood Brain Barrier Models. Compr Physiol 2019, 9, 565–611. [Google Scholar] [CrossRef]
- Hajal, C.; Offeddu, G.S.; Shin, Y.; Zhang, S.; Morozova, O.; Hickman, D.; Knutson, C.G.; Kamm, R.D. Engineered human blood-brain barrier microfluidic model for vascular permeability analyses. Nat Protoc 2022, 17, 95–128. [Google Scholar] [CrossRef]
- Oddo, A.; Peng, B.; Tong, Z.; Wei, Y.; Tong, W.Y.; Thissen, H.; Voelcker, N.H. Advances in Microfluidic Blood-Brain Barrier (BBB) Models. Trends Biotechnol 2019, 37, 1295–1314. [Google Scholar] [CrossRef]
- Vetter, J.; Palagi, I.; Waisman, A.; Blaeser, A. Recent advances in blood-brain barrier-on-a-chip models. Acta Biomater 2025, 197, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Chaulagain, B.; Gothwal, A.; Lamptey, R.N.L.; Trivedi, R.; Mahanta, A.K.; Layek, B.; Singh, J. Experimental Models of In Vitro Blood-Brain Barrier for CNS Drug Delivery: An Evolutionary Perspective. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Park, T.E.; Mustafaoglu, N.; Herland, A.; Hasselkus, R.; Mannix, R.; FitzGerald, E.A.; Prantil-Baun, R.; Watters, A.; Henry, O.; Benz, M.; et al. Hypoxia-enhanced Blood-Brain Barrier Chip recapitulates human barrier function and shuttling of drugs and antibodies. Nat Commun 2019, 10, 2621. [Google Scholar] [CrossRef] [PubMed]
- Katt, M.E.; Shusta, E.V. In vitro Models of the Blood-Brain Barrier: Building in physiological complexity. Curr Opin Chem Eng 2020, 30, 42–52. [Google Scholar] [CrossRef]
- Filieri, S.; Miciaccia, M.; Armenise, D.; Baldelli, O.M.; Liturri, A.; Ferorelli, S.; Sardanelli, A.M.; Perrone, M.G.; Scilimati, A. Can Focused Ultrasound Overcome the Failure of Chemotherapy in Treating Pediatric Diffuse Intrinsic Pontine Glioma Due to a Blood-Brain Barrier Obstacle? Pharmaceuticals (Basel) 2025, 18. [Google Scholar] [CrossRef]
- Tian, H.; Zhang, T.; Qin, S.; Huang, Z.; Zhou, L.; Shi, J.; Nice, E.C.; Xie, N.; Huang, C.; Shen, Z. Enhancing the therapeutic efficacy of nanoparticles for cancer treatment using versatile targeted strategies. J Hematol Oncol 2022, 15, 132. [Google Scholar] [CrossRef]
- Rahman, R.; Ventz, S.; McDunn, J.; Louv, B.; Reyes-Rivera, I.; Polley, M.C.; Merchant, F.; Abrey, L.E.; Allen, J.E.; Aguilar, L.K.; et al. Leveraging external data in the design and analysis of clinical trials in neuro-oncology. Lancet Oncol 2021, 22, e456–e465. [Google Scholar] [CrossRef]
- Chen, K.T.; Chai, W.Y.; Lin, Y.J.; Lin, C.J.; Chen, P.Y.; Tsai, H.C.; Huang, C.Y.; Kuo, J.S.; Liu, H.L.; Wei, K.C. Neuronavigation-guided focused ultrasound for transcranial blood-brain barrier opening and immunostimulation in brain tumors. Sci Adv 2021, 7. [Google Scholar] [CrossRef]
- Liu, Y.; Castro Bravo, K.M.; Liu, J. Targeted liposomal drug delivery: a nanoscience and biophysical perspective. Nanoscale Horiz 2021, 6, 78–94. [Google Scholar] [CrossRef]
- McDannold, N.; Wen, P.Y.; Reardon, D.A.; Fletcher, S.M.; Golby, A.J. Cavitation monitoring, treatment strategy, and acoustic simulations of focused ultrasound blood-brain barrier disruption in patients with glioblastoma. J Control Release 2024, 372, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Armstrong, T.S.; Gilbert, M.R.; Celiku, O. A critical analysis of neuro-oncology clinical trials. Neuro Oncol 2023, 25, 1658–1671. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Q.; Feng, Z.; Qin, M.; Zhang, Z.; Jiang, J.; Ren, T.; Liu, X.; Jeffrey Brinker, C.; Zhao, Y.; et al. Unlocking tumor barrier: annexin A2-mediated transcytosis boosts drug delivery in pancreatic and breast tumors. Nat Commun 2025, 16, 6531. [Google Scholar] [CrossRef] [PubMed]
- Hallschmid, M. Intranasal Insulin for Alzheimer's Disease. CNS Drugs 2021, 35, 21–37. [Google Scholar] [CrossRef]
- Wong, C.Y.J.; Baldelli, A.; Hoyos, C.M.; Tietz, O.; Ong, H.X.; Traini, D. Insulin Delivery to the Brain via the Nasal Route: Unraveling the Potential for Alzheimer's Disease Therapy. Drug Deliv Transl Res 2024, 14, 1776–1793. [Google Scholar] [CrossRef]
- Chen, L.; Guan, Y.; Wang, S.; Han, X.; Guo, F.; Wang, Y. Engineered nanoplatforms for brain-targeted co-delivery of phytochemicals in Alzheimer's disease: Rational design, blood-brain barrier penetration, and multi-target therapeutic synergy. Neurotherapeutics 2025, 22, e00722. [Google Scholar] [CrossRef]
- Wei, Y.; Xia, X.; Wang, X.; Yang, W.; He, S.; Wang, L.; Chen, Y.; Zhou, Y.; Chen, F.; Li, H.; et al. Enhanced BBB penetration and microglia-targeting nanomodulator for the two-pronged modulation of chronically activated microglia-mediated neuroinflammation in Alzheimer's disease. Acta Pharm Sin B 2025, 15, 1098–1111. [Google Scholar] [CrossRef]
- Oosthoek, M.; Vermunt, L.; de Wilde, A.; Bongers, B.; Antwi-Berko, D.; Scheltens, P.; van Bokhoven, P.; Vijverberg, E.G.B.; Teunissen, C.E. Utilization of fluid-based biomarkers as endpoints in disease-modifying clinical trials for Alzheimer's disease: a systematic review. Alzheimers Res Ther 2024, 16, 93. [Google Scholar] [CrossRef]
- Elghanam, Y.; Purja, S.; Kim, E.Y. Biomarkers as Endpoints in Clinical Trials for Alzheimer's Disease. J Alzheimers Dis 2024, 99, 693–703. [Google Scholar] [CrossRef]
- Pascoal, T.A.; Aguzzoli, C.S.; Lussier, F.Z.; Crivelli, L.; Suemoto, C.K.; Fortea, J.; Rosa-Neto, P.; Zimmer, E.R.; Ferreira, P.C.L.; Bellaver, B. Insights into the use of biomarkers in clinical trials in Alzheimer's disease. EBioMedicine 2024, 108, 105322. [Google Scholar] [CrossRef]
- Petersen, R.C.; Graf, A.; Brady, C.; De Santi, S.; Florian, H.; Landen, J.; Pontecorvo, M.; Randolph, C.; Sink, K.; Carrillo, M.; et al. Operationalizing selection criteria for clinical trials in Alzheimer's disease: Biomarker and clinical considerations: Proceedings from the Alzheimer's Association Research Roundtable (AARR) Fall 2021 meeting. Alzheimers Dement (N Y) 2025, 11, e70038. [Google Scholar] [CrossRef] [PubMed]
- Monge-Fuentes, V.; Biolchi Mayer, A.; Lima, M.R.; Geraldes, L.R.; Zanotto, L.N.; Moreira, K.G.; Martins, O.P.; Piva, H.L.; Felipe, M.S.S.; Amaral, A.C.; et al. Dopamine-loaded nanoparticle systems circumvent the blood-brain barrier restoring motor function in mouse model for Parkinson's Disease. Sci Rep 2021, 11, 15185. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.; Yang, L.; Kang, J.; Liu, B.; Zhang, D.; Wang, L.; Wang, W.; Wang, Q. Improving treatment for Parkinson's disease: Harnessing photothermal and phagocytosis-driven delivery of levodopa nanocarriers across the blood-brain barrier. Asian J Pharm Sci 2024, 19, 100963. [Google Scholar] [CrossRef] [PubMed]
- Leta, V.; Klingelhoefer, L.; Longardner, K.; Campagnolo, M.; Levent, H.; Aureli, F.; Metta, V.; Bhidayasiri, R.; Chung-Faye, G.; Falup-Pecurariu, C.; et al. Gastrointestinal barriers to levodopa transport and absorption in Parkinson's disease. Eur J Neurol 2023, 30, 1465–1480. [Google Scholar] [CrossRef]
- Qiu, Y.; Jacobs, D.M.; Messer, K.; Salmon, D.P.; Wellington, C.L.; Stukas, S.; Revta, C.; Brewer, J.B.; Léger, G.C.; Askew, B.; et al. Prognostic value of plasma biomarkers for informing clinical trial design in mild-to-moderate Alzheimer's disease. Alzheimers Res Ther 2025, 17, 97. [Google Scholar] [CrossRef]
- Asgharian, P.; Quispe, C.; Herrera-Bravo, J.; Sabernavaei, M.; Hosseini, K.; Forouhandeh, H.; Ebrahimi, T.; Sharafi-Badr, P.; Tarhriz, V.; Soofiyani, S.R.; et al. Pharmacological effects and therapeutic potential of natural compounds in neuropsychiatric disorders: An update. Front Pharmacol 2022, 13, 926607. [Google Scholar] [CrossRef]
- Küpeli Akkol, E.; Tatlı Çankaya, I.; Şeker Karatoprak, G.; Carpar, E.; Sobarzo-Sánchez, E.; Capasso, R. Natural Compounds as Medical Strategies in the Prevention and Treatment of Psychiatric Disorders Seen in Neurological Diseases. Front Pharmacol 2021, 12, 669638. [Google Scholar] [CrossRef]
- McIntyre, R.S.; Kwan, A.T.H.; Mansur, R.B.; Oliveira-Maia, A.J.; Teopiz, K.M.; Maletic, V.; Suppes, T.; Stahl, S.M.; Rosenblat, J.D. Psychedelics for the Treatment of Psychiatric Disorders: Interpreting and Translating Available Evidence and Guidance for Future Research. Am J Psychiatry 2025, 182, 21–32. [Google Scholar] [CrossRef]
- Barakji, J.; Korang, S.K.; Feinberg, J.; Maagaard, M.; Mathiesen, O.; Gluud, C.; Jakobsen, J.C. Cannabinoids versus placebo for pain: A systematic review with meta-analysis and Trial Sequential Analysis. PLoS One 2023, 18, e0267420. [Google Scholar] [CrossRef]
- Jiang, P.; Li, J. Recent advances in biomimetic nanodelivery systems for the treatment of depression. Mater Today Bio 2025, 32, 101781. [Google Scholar] [CrossRef]
- Davis, K.D.; Aghaeepour, N.; Ahn, A.H.; Angst, M.S.; Borsook, D.; Brenton, A.; Burczynski, M.E.; Crean, C.; Edwards, R.; Gaudilliere, B.; et al. Discovery and validation of biomarkers to aid the development of safe and effective pain therapeutics: challenges and opportunities. Nat Rev Neurol 2020, 16, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M. Beyond the boundaries: Transitioning from categorical to dimensional paradigms in mental health diagnostics. Adv Clin Exp Med 2024, 33, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Battaglia, S. Dualistic Dynamics in Neuropsychiatry: From Monoaminergic Modulators to Multiscale Biomarker Maps. Biomedicines 2025, 13. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Szabó, Á.; Vécsei, L. Redefining Roles: A Paradigm Shift in Tryptophan-Kynurenine Metabolism for Innovative Clinical Applications. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Szabó, Á.; Galla, Z.; Spekker, E.; Szűcs, M.; Martos, D.; Takeda, K.; Ozaki, K.; Inoue, H.; Yamamoto, S.; Toldi, J.; et al. Oxidative and Excitatory Neurotoxic Stresses in CRISPR/Cas9-Induced Kynurenine Aminotransferase Knockout Mice: A Novel Model for Despair-Based Depression and Post-Traumatic Stress Disorder. Front Biosci (Landmark Ed) 2025, 30, 25706. [Google Scholar] [CrossRef]
- Naeem, A.; Hu, P.; Yang, M.; Zhang, J.; Liu, Y.; Zhu, W.; Zheng, Q. Natural Products as Anticancer Agents: Current Status and Future Perspectives. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Chunarkar-Patil, P.; Kaleem, M.; Mishra, R.; Ray, S.; Ahmad, A.; Verma, D.; Bhayye, S.; Dubey, R.; Singh, H.N.; Kumar, S. Anticancer Drug Discovery Based on Natural Products: From Computational Approaches to Clinical Studies. Biomedicines 2024, 12. [Google Scholar] [CrossRef]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in Cancer Treatment: From Preclinical Studies to Clinical Practice. Front Pharmacol 2019, 10, 1614. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, X.; Pang, X.; Ma, K.; Li, L.; Song, Y.; Hou, D.; Wang, X. Pharmacological effects, molecular mechanisms and strategies to improve bioavailability of curcumin in the treatment of neurodegenerative diseases. Front Pharmacol 2025, 16, 1625821. [Google Scholar] [CrossRef]
- Grabarczyk, M.; Justyńska, W.; Czpakowska, J.; Smolińska, E.; Bielenin, A.; Glabinski, A.; Szpakowski, P. Role of Plant Phytochemicals: Resveratrol, Curcumin, Luteolin and Quercetin in Demyelination, Neurodegeneration, and Epilepsy. Antioxidants (Basel) 2024, 13. [Google Scholar] [CrossRef]
- Ahn-Horst, R.Y.; Turner, E.H.; Kesselheim, A.S. Characteristics of Trials Preceding FDA Approval of Novel Psychiatric Drugs. JAMA Netw Open 2025, 8, e2456588. [Google Scholar] [CrossRef]
- Markowitz, J.C.; Milrod, B.L. Lost in Translation: The Value of Psychiatric Clinical Trials. J Clin Psychiatry 2022, 83. [Google Scholar] [CrossRef]
- Lipsman, N.; Meng, Y.; Bethune, A.J.; Huang, Y.; Lam, B.; Masellis, M.; Herrmann, N.; Heyn, C.; Aubert, I.; Boutet, A.; et al. Blood-brain barrier opening in Alzheimer's disease using MR-guided focused ultrasound. Nat Commun 2018, 9, 2336. [Google Scholar] [CrossRef]
- Gasca-Salas, C.; Fernández-Rodríguez, B.; Pineda-Pardo, J.A.; Rodríguez-Rojas, R.; Obeso, I.; Hernández-Fernández, F.; Del Álamo, M.; Mata, D.; Guida, P.; Ordás-Bandera, C.; et al. Blood-brain barrier opening with focused ultrasound in Parkinson's disease dementia. Nat Commun 2021, 12, 779. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, H.; Pandey, M.; Chin, P.X.; Phang, Y.L.; Cheah, J.Y.; Ooi, S.C.; Mak, K.K.; Pichika, M.R.; Kesharwani, P.; Hussain, Z.; et al. Transferrin receptors-targeting nanocarriers for efficient targeted delivery and transcytosis of drugs into the brain tumors: a review of recent advancements and emerging trends. Drug Deliv Transl Res 2018, 8, 1545–1563. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, M.J.; Sevin, E.; Gosselet, F.; Lima, J.; Coelho, M.A.N.; Loureiro, J.A.; Pereira, M.C. Receptor-mediated PLGA nanoparticles for glioblastoma multiforme treatment. Int J Pharm 2018, 545, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Kreatsoulas, D.; Damante, M.; Cua, S.; Lonser, R.R. Adjuvant convection-enhanced delivery for the treatment of brain tumors. J Neurooncol 2024, 166, 243–255. [Google Scholar] [CrossRef]
- Upadhyay, R.; Ghosh, P.; Desavathu, M. Advancement in the Nose-to-Brain Drug delivery of FDA-approved drugs for the better management of Depression and Psychiatric disorders. Int J Pharm 2024, 667, 124866. [Google Scholar] [CrossRef]
- Yang, H.; Liu, R.; Huang, J.; Zhao, Y.; Zou, C.; Wu, D.; Zhao, H. Nose-to-brain delivery of biomineralized silk fibroin nanoparticles synergistically extinguish neuroinflammation for efficient depressive-like symptoms alleviation. Int J Biol Macromol 2025, 322, 146761. [Google Scholar] [CrossRef]
- Zünkeler, B.; Carson, R.E.; Olson, J.; Blasberg, R.G.; DeVroom, H.; Lutz, R.J.; Saris, S.C.; Wright, D.C.; Kammerer, W.; Patronas, N.J.; et al. Quantification and pharmacokinetics of blood-brain barrier disruption in humans. J Neurosurg 1996, 85, 1056–1065. [Google Scholar] [CrossRef]
- Burks, S.R.; Kersch, C.N.; Witko, J.A.; Pagel, M.A.; Sundby, M.; Muldoon, L.L.; Neuwelt, E.A.; Frank, J.A. Blood-brain barrier opening by intracarotid artery hyperosmolar mannitol induces sterile inflammatory and innate immune responses. Proc Natl Acad Sci U S A 2021, 118. [Google Scholar] [CrossRef]
- Wang, W.; Marín-Ramos, N.I.; He, H.; Zeng, S.; Cho, H.Y.; Swenson, S.D.; Zheng, L.; Epstein, A.L.; Schönthal, A.H.; Hofman, F.M.; et al. NEO100 enables brain delivery of blood‒brain barrier impermeable therapeutics. Neuro Oncol 2021, 23, 63–75. [Google Scholar] [CrossRef]
- Pinkiewicz, M.; Pinkiewicz, M.; Walecki, J.; Zaczyński, A.; Zawadzki, M. Breaking Barriers in Neuro-Oncology: A Scoping Literature Review on Invasive and Non-Invasive Techniques for Blood-Brain Barrier Disruption. Cancers (Basel) 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Mollnes, T.E.; Storm, B.S.; Brekke, O.L.; Nilsson, P.H.; Lambris, J.D. Application of the C3 inhibitor compstatin in a human whole blood model designed for complement research - 20 years of experience and future perspectives. Semin Immunol 2022, 59, 101604. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Hu, D.; Wang, M.; Zipfel, P.F.; Hu, Y. Complement in Hemolysis- and Thrombosis- Related Diseases. Front Immunol 2020, 11, 1212. [Google Scholar] [CrossRef] [PubMed]
- Roumenina, L.T.; Bartolucci, P.; Pirenne, F. The role of Complement in Post-Transfusion Hemolysis and Hyperhemolysis Reaction. Transfus Med Rev 2019, 33, 225–230. [Google Scholar] [CrossRef]
- Poillerat, V.; Gentinetta, T.; Leon, J.; Wassmer, A.; Edler, M.; Torset, C.; Luo, D.; Tuffin, G.; Roumenina, L.T. Hemopexin as an Inhibitor of Hemolysis-Induced Complement Activation. Front Immunol 2020, 11, 1684. [Google Scholar] [CrossRef]
- Nguyen-Peyre, K.-A.; Meuleman, M.-S.; Bodivit, G.; Kassasseya, C.; Decrouy, X.; Vingert, B.; Bencheikh, L.; Lambris, J.; Pirenne, F.; Roumenina, L.T. Red Blood Cell-Derived Particles Induce Endothelial Damage Via the Alternative Complement Pathway. 2024.
- Kang, H.J.; Roh, J.; Lee, H.; Park, E.M.; Lee, H.W.; Lee, J.Y.; Hwang, J.H.; Shim, J.; Choi, K. Complement Activation and Hemolysis in Non-human Primates Following Transfusion of Genetically Modified Pig Red Blood Cells. Ann Lab Med 2025, 45, 509–519. [Google Scholar] [CrossRef]
- Alharbi, Y.M. Phenotype- and age-associated variations in non-specific agglutinins and complement components (C3 and C5a) in camels: Implications for transfusion compatibility and immune function. Vet World 2025, 18, 2811–2822. [Google Scholar] [CrossRef]
- Singh, D. Astrocytic and microglial cells as the modulators of neuroinflammation in Alzheimer's disease. J Neuroinflammation 2022, 19, 206. [Google Scholar] [CrossRef]
- Müller, L.; Di Benedetto, S. Neuroimmune crosstalk in chronic neuroinflammation: microglial interactions and immune modulation. Front Cell Neurosci 2025, 19, 1575022. [Google Scholar] [CrossRef]
- West, P.K.; McCorkindale, A.N.; Guennewig, B.; Ashhurst, T.M.; Viengkhou, B.; Hayashida, E.; Jung, S.R.; Butovsky, O.; Campbell, I.L.; Hofer, M.J. The cytokines interleukin-6 and interferon-α induce distinct microglia phenotypes. J Neuroinflammation 2022, 19, 96. [Google Scholar] [CrossRef]
- Fornari Laurindo, L.; Aparecido Dias, J.; Cressoni Araújo, A.; Torres Pomini, K.; Machado Galhardi, C.; Rucco Penteado Detregiachi, C.; Santos de Argollo Haber, L.; Donizeti Roque, D.; Dib Bechara, M.; Vialogo Marques de Castro, M.; et al. Immunological dimensions of neuroinflammation and microglial activation: exploring innovative immunomodulatory approaches to mitigate neuroinflammatory progression. Front Immunol 2023, 14, 1305933. [Google Scholar] [CrossRef] [PubMed]
- Alaei, M.; Koushki, K.; Taebi, K.; Yousefi Taba, M.; Keshavarz Hedayati, S.; Keshavarz Shahbaz, S. Metal nanoparticles in neuroinflammation: impact on microglial dynamics and CNS function. RSC Adv 2025, 15, 5426–5451. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Francis, N.L.; Calvelli, H.R.; Moghe, P.V. Microglia-targeting nanotherapeutics for neurodegenerative diseases. APL Bioeng 2020, 4, 030902. [Google Scholar] [CrossRef] [PubMed]
- Benita, B.A.; Koss, K.M. Peptide discovery across the spectrum of neuroinflammation; microglia and astrocyte phenotypical targeting, mediation, and mechanistic understanding. Front Mol Neurosci 2024, 17, 1443985. [Google Scholar] [CrossRef]
- Weber, M.; Steinle, H.; Golombek, S.; Hann, L.; Schlensak, C.; Wendel, H.P.; Avci-Adali, M. Blood-Contacting Biomaterials: In Vitro Evaluation of the Hemocompatibility. Front Bioeng Biotechnol 2018, 6, 99. [Google Scholar] [CrossRef]
- Nalezinková, M. In vitro hemocompatibility testing of medical devices. Thromb Res 2020, 195, 146–150. [Google Scholar] [CrossRef]
- van Oeveren, W. Obstacles in haemocompatibility testing. Scientifica (Cairo) 2013, 2013, 392584. [Google Scholar] [CrossRef]
- Seyfert, U.T.; Biehl, V.; Schenk, J. In vitro hemocompatibility testing of biomaterials according to the ISO 10993-4. Biomol Eng 2002, 19, 91–96. [Google Scholar] [CrossRef]
- Tienda-Vazquez, M.A.; Arredondo, P.; Mejía-Delgadillo, X.; Rodríguez-González, J.A.; Soto-Cajiga, J.A.; Sabath, E.; Lozano, O.; Almanza-Arjona, Y.C. Biological testing unification for hemodialysis membranes evaluation: A step towards standardization. Biomater Adv 2025, 169, 214165. [Google Scholar] [CrossRef]
- Tujula, I.; Hyvärinen, T.; Lotila, J.; Rogal, J.; Voulgaris, D.; Sukki, L.; Tornberg, K.; Korpela, K.; Jäntti, H.; Malm, T.; et al. Modeling neuroinflammatory interactions between microglia and astrocytes in a human iPSC-based coculture platform. Cell Commun Signal 2025, 23, 298. [Google Scholar] [CrossRef] [PubMed]
- Guttikonda, S.R.; Sikkema, L.; Tchieu, J.; Saurat, N.; Walsh, R.M.; Harschnitz, O.; Ciceri, G.; Sneeboer, M.; Mazutis, L.; Setty, M.; et al. Fully defined human pluripotent stem cell-derived microglia and tri-culture system model C3 production in Alzheimer's disease. Nat Neurosci 2021, 24, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Yuan, N.Y.; Richards, W.D.; Parham, K.T.; Clark, S.G.; Greuel, K.; Polzin, B.; Smith, S.W.; Lebakken, C.S. Neural organoids incorporating microglia to assess neuroinflammation and toxicities induced by known developmental neurotoxins. Curr Res Toxicol 2025, 9, 100252. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Gauthier, S.; Chandekar, S.A.; Hviid Hahn-Pedersen, J.; Bentsen, M.A.; Zetterberg, H. Neuroinflammatory fluid biomarkers in patients with Alzheimer's disease: a systematic literature review. Mol Psychiatry 2025, 30, 2783–2798. [Google Scholar] [CrossRef]
- Roveta, F.; Bonino, L.; Piella, E.M.; Rainero, I.; Rubino, E. Neuroinflammatory Biomarkers in Alzheimer's Disease: From Pathophysiology to Clinical Implications. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Battaglia, S.; Tanaka, M. Screen, Sample, Stratify: Biomarkers and Machine Learning Compress Dementia Pathways. 2026, 14, 159. [Google Scholar] [CrossRef]
- Tanaka, M.; Battaglia, S. From Biomarkers to Behavior: Mapping the Neuroimmune Web of Pain, Mood, and Memory. Biomedicines 2025, 13. [Google Scholar] [CrossRef]
- Wei, Y.; Quan, L.; Zhou, C.; Zhan, Q. Factors relating to the biodistribution & clearance of nanoparticles & their effects on in vivo application. Nanomedicine (Lond) 2018, 13, 1495–1512. [Google Scholar] [CrossRef]
- Dogra, P.; Adolphi, N.L.; Wang, Z.; Lin, Y.S.; Butler, K.S.; Durfee, P.N.; Croissant, J.G.; Noureddine, A.; Coker, E.N.; Bearer, E.L.; et al. Establishing the effects of mesoporous silica nanoparticle properties on in vivo disposition using imaging-based pharmacokinetics. Nat Commun 2018, 9, 4551. [Google Scholar] [CrossRef]
- Gustafson, H.H.; Holt-Casper, D.; Grainger, D.W.; Ghandehari, H. Nanoparticle Uptake: The Phagocyte Problem. Nano Today 2015, 10, 487–510. [Google Scholar] [CrossRef] [PubMed]
- Ngo, W.; Ahmed, S.; Blackadar, C.; Bussin, B.; Ji, Q.; Mladjenovic, S.M.; Sepahi, Z.; Chan, W.C.W. Why nanoparticles prefer liver macrophage cell uptake in vivo. Adv Drug Deliv Rev 2022, 185, 114238. [Google Scholar] [CrossRef] [PubMed]
- Lérida-Viso, A.; Estepa-Fernández, A.; García-Fernández, A.; Martí-Centelles, V.; Martínez-Máñez, R. Biosafety of mesoporous silica nanoparticles; towards clinical translation. Adv Drug Deliv Rev 2023, 201, 115049. [Google Scholar] [CrossRef] [PubMed]
- Baek, M.J.; Hur, W.; Kashiwagi, S.; Choi, H.S. Design Considerations for Organ-Selective Nanoparticles. ACS Nano 2025, 19, 14605–14626. [Google Scholar] [CrossRef]
- Zhu, G.H.; Gray, A.B.C.; Patra, H.K. Nanomedicine: controlling nanoparticle clearance for translational success. Trends Pharmacol Sci 2022, 43, 709–711. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, X.; Chen, X.; Fang, W.; Yu, K.; Gu, W.; Wei, Y.; Zheng, H.; Piao, J.; Li, F. Strategies to Regulate the Degradation and Clearance of Mesoporous Silica Nanoparticles: A Review. Int J Nanomedicine 2024, 19, 5859–5878. [Google Scholar] [CrossRef]
- Wang, L.; Quine, S.; Frickenstein, A.N.; Lee, M.; Yang, W.; Sheth, V.M.; Bourlon, M.D.; He, Y.; Lyu, S.; Garcia-Contreras, L.; et al. Exploring and Analyzing the Systemic Delivery Barriers for Nanoparticles. Adv Funct Mater 2024, 34. [Google Scholar] [CrossRef]
- Zelepukin, I.V.; Shevchenko, K.G.; Deyev, S.M. Rediscovery of mononuclear phagocyte system blockade for nanoparticle drug delivery. Nat Commun 2024, 15, 4366. [Google Scholar] [CrossRef]
- Bresinsky, M.; Goepferich, A. Control of biomedical nanoparticle distribution and drug release in vivo by complex particle design strategies. Eur J Pharm Biopharm 2025, 208, 114634. [Google Scholar] [CrossRef]
- Hidayat, A.F.; Wardhana, Y.W.; Suwendar, S.; Mohammed, A.F.A.; Mahmoud, S.A.; Elamin, K.M.; Wathoni, N. A Review on QbD-Driven Optimization of Lipid Nanoparticles for Oral Drug Delivery: From Framework to Formulation. Int J Nanomedicine 2025, 20, 8611–8651. [Google Scholar] [CrossRef]
- Yang, S.; Hu, X.; Zhu, J.; Zheng, B.; Bi, W.; Wang, X.; Wu, J.; Mi, Z.; Wu, Y. Aspects and Implementation of Pharmaceutical Quality by Design from Conceptual Frameworks to Industrial Applications. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Pielenhofer, J.; Meiser, S.L.; Gogoll, K.; Ciciliani, A.M.; Denny, M.; Klak, M.; Lang, B.M.; Staubach, P.; Grabbe, S.; Schild, H.; et al. Quality by Design (QbD) Approach for a Nanoparticulate Imiquimod Formulation as an Investigational Medicinal Product. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Walsh, I.; Myint, M.; Nguyen-Khuong, T.; Ho, Y.S.; Ng, S.K.; Lakshmanan, M. Harnessing the potential of machine learning for advancing "Quality by Design" in biomanufacturing. MAbs 2022, 14, 2013593. [Google Scholar] [CrossRef] [PubMed]
- Suriyaamporn, P.; Kansom, T.; Pamornpathomkul, B.; Ngawhirunpat, T.; Opanasopit, P.; Ramjan, S. Predictive modeling approach using machine learning-integrated design of experiments in quality by design for optimizing resveratrol-loaded polymeric nanoparticle formulation. Int J Pharm 2025, 683, 126080. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M. Special Issue "Translating Molecular Psychiatry: From Biomarkers to Personalized Therapies. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef] [PubMed]
- Palakurthi, S.S.; Charbe, N.B.; Kapre, S.; Zheng, W.; Thalla, M.; Palaniappan, D.; Lu, D.; Palakurthi, S. Evaluating critical quality attributes and novel drug release testing of difluprednate nanoemulsions. Int J Pharm 2025, 674, 125431. [Google Scholar] [CrossRef]
- Soncin, S.; Lo Cicero, V.; Astori, G.; Soldati, G.; Gola, M.; Sürder, D.; Moccetti, T. A practical approach for the validation of sterility, endotoxin and potency testing of bone marrow mononucleated cells used in cardiac regeneration in compliance with good manufacturing practice. J Transl Med 2009, 7, 78. [Google Scholar] [CrossRef]
- Vuillemenot, B.R.; Korte, S.; Wright, T.L.; Adams, E.L.; Boyd, R.B.; Butt, M.T. Safety Evaluation of CNS Administered Biologics-Study Design, Data Interpretation, and Translation to the Clinic. Toxicol Sci 2016, 152, 3–9. [Google Scholar] [CrossRef]
- Cooper, J.F.; Latta, K.S.; Smith, D. Automated endotoxin testing program for high-risk-level compounded sterile preparations at an institutional compounding pharmacy. Am J Health Syst Pharm 2010, 67, 280–286. [Google Scholar] [CrossRef]
- Cooper, J.F.; Thoma, L.A. Screening extemporaneously compounded intraspinal injections with the bacterial endotoxins test. Am J Health Syst Pharm 2002, 59, 2426–2433. [Google Scholar] [CrossRef]
- Dubczak, J.; Reid, N.; Tsuchiya, M. Evaluation of limulus amebocyte lysate and recombinant endotoxin alternative assays for an assessment of endotoxin detection specificity. Eur J Pharm Sci 2021, 159, 105716. [Google Scholar] [CrossRef]
- Secretan, P.H.; Annereau, M.; Do, B. A Risk-Based Framework for Hospital Compounding: Integrating Degradation Mechanisms and Predictive Toxicology. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Pallerla, S.; Pires, I.S.; Melo, M.B.; Yun, D.; Wagner, A.; Budai, M.; Kumar, D.; Katinger, D.; Sayeed, E.; Lombardo, A.; et al. Scale-up and cGMP manufacturing of next-generation vaccine adjuvant saponin/MPLA nanoParticles (SMNP). J Pharm Sci 2025, 114, 103913. [Google Scholar] [CrossRef] [PubMed]
- Cunha, S.; Costa, C.P.; Loureiro, J.A.; Alves, J.; Peixoto, A.F.; Forbes, B.; Sousa Lobo, J.M.; Silva, A.C. Double Optimization of Rivastigmine-Loaded Nanostructured Lipid Carriers (NLC) for Nose-to-Brain Delivery Using the Quality by Design (QbD) Approach: Formulation Variables and Instrumental Parameters. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Laggner, M.; Gugerell, A.; Bachmann, C.; Hofbauer, H.; Vorstandlechner, V.; Seibold, M.; Gouya Lechner, G.; Peterbauer, A.; Madlener, S.; Demyanets, S. Reproducibility of GMP-compliant production of therapeutic stressed peripheral blood mononuclear cell-derived secretomes, a novel class of biological medicinal products. Stem cell research & therapy 2020, 11, 9. [Google Scholar]
- Sanyal, G.; Särnefält, A.; Kumar, A. Considerations for bioanalytical characterization and batch release of COVID-19 vaccines. NPJ Vaccines 2021, 6, 53. [Google Scholar] [CrossRef]
- Laggner, M.; Gugerell, A.; Bachmann, C.; Hofbauer, H.; Vorstandlechner, V.; Seibold, M.; Gouya Lechner, G.; Peterbauer, A.; Madlener, S.; Demyanets, S.; et al. Reproducibility of GMP-compliant production of therapeutic stressed peripheral blood mononuclear cell-derived secretomes, a novel class of biological medicinal products. Stem Cell Res Ther 2020, 11, 9. [Google Scholar] [CrossRef]
- Csóka, I.; Ismail, R.; Jójárt-Laczkovich, O.; Pallagi, E. Regulatory Considerations, Challenges and Risk-based Approach in Nanomedicine Development. Curr Med Chem 2021, 28, 7461–7476. [Google Scholar] [CrossRef]
- Ali, F.; Neha, K.; Parveen, S. Current regulatory landscape of nanomaterials and nanomedicines: A global perspective. Journal of Drug Delivery Science and Technology 2023, 80, 104118. [Google Scholar] [CrossRef]
- Dobrovolskaia, M.A. Lessons learned from immunological characterization of nanomaterials at the Nanotechnology Characterization Laboratory. Front Immunol 2022, 13, 984252. [Google Scholar] [CrossRef]
- Giannakou, C.; Park, M.; Bosselaers, I.E.M.; de Jong, W.H.; van der Laan, J.W.; van Loveren, H.; Vandebriel, R.J.; Geertsma, R.E. Nonclinical regulatory immunotoxicity testing of nanomedicinal products: Proposed strategy and possible pitfalls. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2020, 12, e1633. [Google Scholar] [CrossRef] [PubMed]
- Hofer, S.; Hofstätter, N.; Punz, B.; Hasenkopf, I.; Johnson, L.; Himly, M. Immunotoxicity of nanomaterials in health and disease: Current challenges and emerging approaches for identifying immune modifiers in susceptible populations. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2022, 14, e1804. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaia, M.A.; McNeil, S.E. Understanding the correlation between in vitro and in vivo immunotoxicity tests for nanomedicines. J Control Release 2013, 172, 456–466. [Google Scholar] [CrossRef] [PubMed]
- van Gerven, J.; Bonelli, M. Commentary on the EMA Guideline on strategies to identify and mitigate risks for first-in-human and early clinical trials with investigational medicinal products. Br J Clin Pharmacol 2018, 84, 1401–1409. [Google Scholar] [CrossRef]
- Musazzi, U.M.; Franzè, S.; Condorelli, F.; Minghetti, P.; Caliceti, P. Feeding Next-Generation Nanomedicines to Europe: Regulatory and Quality Challenges. Adv Healthc Mater 2023, 12, e2301956. [Google Scholar] [CrossRef]
- Zhuang, Y.; Mendes, B.B.; Menon, D.; Oliveira, J.; Chen, X.; Duman, F.D.; Conniot, J.; Mercado, S.; Liu, X.; Zhang, S.Y.; et al. Multiscale Profiling of Nanoscale Metal-Organic Framework Biocompatibility and Immune Interactions. Adv Healthc Mater 2025, 14, e01809. [Google Scholar] [CrossRef]
- Nabi, A.E.; Pouladvand, P.; Liu, L.; Hua, N.; Ayubcha, C. Machine Learning in Drug Development for Neurological Diseases: A Review of Blood Brain Barrier Permeability Prediction Models. Mol Inform 2025, 44, e202400325. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, L.; Feng, H.; Li, S.; Liu, M.; Zhao, J.; Liu, H. Prediction of the Blood-Brain Barrier (BBB) Permeability of Chemicals Based on Machine-Learning and Ensemble Methods. Chem Res Toxicol 2021, 34, 1456–1467. [Google Scholar] [CrossRef]
- Gülave, B.; van den Maagdenberg, H.W.; van Boven, L.; van Westen, G.J.P.; de Lange, E.C.M.; van Hasselt, J.G.C. Prediction of the Extent of Blood-Brain Barrier Transport Using Machine Learning and Integration into the LeiCNS-PK3.0 Model. Pharm Res 2025, 42, 281–289. [Google Scholar] [CrossRef]
- Mehta, P.; Soliman, A.; Rodriguez-Vera, L.; Schmidt, S.; Muniz, P.; Rodriguez, M.; Forcadell, M.; Gonzalez-Perez, E.; Vozmediano, V. Interspecies Brain PBPK Modeling Platform to Predict Passive Transport through the Blood-Brain Barrier and Assess Target Site Disposition. Pharmaceutics 2024, 16. [Google Scholar] [CrossRef]
- Cichońska, A.; Ravikumar, B.; Rahman, R. AI for targeted polypharmacology: The next frontier in drug discovery. Curr Opin Struct Biol 2024, 84, 102771. [Google Scholar] [CrossRef] [PubMed]
- Abdelsayed, M. AI-Driven Polypharmacology in Small-Molecule Drug Discovery. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef] [PubMed]
- Noor, F.; Asif, M.; Ashfaq, U.A.; Qasim, M.; Tahir Ul Qamar, M. Machine learning for synergistic network pharmacology: a comprehensive overview. Brief Bioinform 2023, 24. [Google Scholar] [CrossRef]
- Chaudhari, R.; Fong, L.W.; Tan, Z.; Huang, B.; Zhang, S. An up-to-date overview of computational polypharmacology in modern drug discovery. Expert Opin Drug Discov 2020, 15, 1025–1044. [Google Scholar] [CrossRef] [PubMed]
- Lambrinidis, G.; Tsantili-Kakoulidou, A. Multi-objective optimization methods in novel drug design. Expert Opin Drug Discov 2021, 16, 647–658. [Google Scholar] [CrossRef]
- Waibel, I.; Schneider, T.N.; Fischer, F.J.; Dumnoenchanvanit, P.; Kulakova, A.; Nguyen, T.D.; Egebjerg, T.; Bertelsen, S.; Lorenzen, N.; Arosio, P. Bayesian Optimization for Efficient Multiobjective Formulation Development of Biologics. Mol Pharm 2025, 22, 6636–6645. [Google Scholar] [CrossRef]
- Li, L.; Back, S.I.; Ma, J.; Guo, Y.; Galeandro-Diamant, T.; Clénet, D. Bayesian optimization and machine learning for vaccine formulation development. PLoS One 2025, 20, e0324205. [Google Scholar] [CrossRef]
- Luukkonen, S.; van den Maagdenberg, H.W.; Emmerich, M.T.M.; van Westen, G.J.P. Artificial intelligence in multi-objective drug design. Curr Opin Struct Biol 2023, 79, 102537. [Google Scholar] [CrossRef]
- Wickramasinghe, C.D.; Kim, S.; Li, J. SpatialCNS-PBPK: An R/Shiny Web-Based Application for Physiologically Based Pharmacokinetic Modeling of Spatial Pharmacokinetics in the Human Central Nervous System and Brain Tumors. CPT Pharmacometrics Syst Pharmacol 2025, 14, 864–880. [Google Scholar] [CrossRef]
- Bowman, C.; Ma, F.; Mao, J.; Plise, E.; Chen, E.; Liu, L.; Zhang, S.; Chen, Y. Evaluation of bottom-up modeling of the blood-brain barrier to improve brain penetration prediction via physiologically based pharmacokinetic modeling. Biopharm Drug Dispos 2023, 44, 60–70. [Google Scholar] [CrossRef]
- Tregub, P.P.; Bystrov, D.A.; Kushnir, I.A.; Korsakova, S.A.; Yurchenko, S.O.; Salmina, A.B. Blood-brain barriers and drug pharmacokinetics: mechanisms and models. Eur J Pharmacol 2025, 1003, 177872. [Google Scholar] [CrossRef]
- Linville, R.M.; DeStefano, J.G.; Sklar, M.B.; Xu, Z.; Farrell, A.M.; Bogorad, M.I.; Chu, C.; Walczak, P.; Cheng, L.; Mahairaki, V.; et al. Human iPSC-derived blood-brain barrier microvessels: validation of barrier function and endothelial cell behavior. Biomaterials 2019, 190-191, 24–37. [Google Scholar] [CrossRef]
- Canfield, S.G.; Stebbins, M.J.; Faubion, M.G.; Gastfriend, B.D.; Palecek, S.P.; Shusta, E.V. An isogenic neurovascular unit model comprised of human induced pluripotent stem cell-derived brain microvascular endothelial cells, pericytes, astrocytes, and neurons. Fluids Barriers CNS 2019, 16, 25. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, L.; Vermond, S.; Hill, S.; de Munnik, S.; van Puijvelde, G.; Quist, B.; Pooley, R.; Carr, K.; Brown, N.; Aspinall-O'Dea, M. Abstract LB009: Safety profile assessment for IND-enabling studies: Bispecific antibodies and antibody-drug conjugates (ADCs). Cancer Research 2025, 85, LB009–LB009. [Google Scholar] [CrossRef]
- Youssef, E.; Weddle, K.; Zimmerman, L.; Palmer, D. Pharmacovigilance in Cell and Gene Therapy: Evolving Challenges in Risk Management and Long-Term Follow-Up. Drug Saf 2026, 49, 27–53. [Google Scholar] [CrossRef] [PubMed]
- Glader, C.; Jeitler, R.; Wang, Y.; van Tuijn, R.; Grau-Carbonell, A.; Tetyczka, C.; Mesite, S.; Caisse, P.; Khinast, J.; Roblegg, E. Process Analytical Strategies for Size Monitoring: Offline, At-Line, Online, and Inline Methods in a Top-Down Nano-Manufacturing Line. Pharmaceutics 2025, 17. [Google Scholar] [CrossRef]
- Adaptive platform trials: definition, design, conduct and reporting considerations. Nat Rev Drug Discov 2019, 18, 797–807. [CrossRef]
- Narsinh, K.H.; Perez, E.; Haddad, A.F.; Young, J.S.; Savastano, L.; Villanueva-Meyer, J.E.; Winkler, E.; de Groot, J. Strategies to Improve Drug Delivery Across the Blood-Brain Barrier for Glioblastoma. Curr Neurol Neurosci Rep 2024, 24, 123–139. [Google Scholar] [CrossRef]
- Zhu, M.; Zhuang, J.; Li, Z.; Liu, Q.; Zhao, R.; Gao, Z.; Midgley, A.C.; Qi, T.; Tian, J.; Zhang, Z.; et al. Machine-learning-assisted single-vessel analysis of nanoparticle permeability in tumour vasculatures. Nat Nanotechnol 2023, 18, 657–666. [Google Scholar] [CrossRef]
- Pan, F.; Xia, Y.; Zhang, B.; Mohammed, A.; Zhao, X. Microfluidic Fabrication of Peptide-Functionalized Poly(lactic-co-glycolic acid) Nanoparticles for Targeted Curcumin Delivery in Breast Cancer. Langmuir 2025, 41, 19514–19525. [Google Scholar] [CrossRef]
- Kim, J.; Lee, D.H.; Seo, H.; Lee, H.J.; Kim, G.; Nie, C.; Hong, Y.; Yoon, Y.J.; Lim, J.Y.; Park, S.J.; et al. Microfluidic Generation of Exosome-Mimetic Nanoparticles for Scalable Production and Enhanced Therapeutic Efficacy. Small 2025, 21, e06162. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Wang, W.; Che, W.; Xu, Y.; Jin, C.; Dong, L.; Xia, Q. Nanomedicines Targeting Metabolic Pathways in the Tumor Microenvironment: Future Perspectives and the Role of AI. Metabolites 2025, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kohane, D.S. External triggering and triggered targeting strategies for drug delivery. Nature Reviews Materials 2017, 2, 1–14. [Google Scholar] [CrossRef]
- Sanadgol, N.; Abedi, M.; Hashemzaei, M.; Kamran, Z.; Khalseh, R.; Beyer, C.; Voelz, C. Exosomes as nanocarriers for brain-targeted delivery of therapeutic nucleic acids: advances and challenges. J Nanobiotechnology 2025, 23, 453. [Google Scholar] [CrossRef]
- Schreiner, T.G.; Menéndez-González, M.; Schreiner, O.D.; Ciobanu, R.C. Intrathecal Therapies for Neurodegenerative Diseases: A Review of Current Approaches and the Urgent Need for Advanced Delivery Systems. Biomedicines 2025, 13. [Google Scholar] [CrossRef]
- Sebghatollahi, Z.; Yogesh, R.; Mahato, N.; Kumar, V.; Mohanta, Y.K.; Baek, K.H.; Mishra, A.K. Signaling Pathways in Oxidative Stress-Induced Neurodegenerative Diseases: A Review of Phytochemical Therapeutic Interventions. Antioxidants (Basel) 2025, 14. [Google Scholar] [CrossRef]
- Mehta, R.S.; Kochar, B.D.; Kennelty, K.; Ernst, M.E.; Chan, A.T. Emerging approaches to polypharmacy among older adults. Nat Aging 2021, 1, 347–356. [Google Scholar] [CrossRef]
- de Koning, L.A.; Vazquez-Matias, D.A.; Beaino, W.; Vugts, D.J.; van Dongen, G.; van der Flier, W.M.; Ries, M.; van Vuurden, D.G.; Vijverberg, E.G.B.; van de Giessen, E. Drug delivery strategies to cross the blood-brain barrier in Alzheimer's disease: a comprehensive review on three promising strategies. J Prev Alzheimers Dis 2025, 12, 100204. [Google Scholar] [CrossRef]
- Vargas, R.; Martinez-Martinez, N.; Lizano-Barrantes, C.; Pacheco-Molina, J.A.; García-Montoya, E.; Pérez-Lozano, P.; Suñé-Negre, J.M.; Suñé, C.; Suñé-Pou, M. Advancing through the blood-brain barrier: mechanisms, challenges and drug delivery strategies. Admet dmpk 2025, 13, 2988. [Google Scholar] [CrossRef]
- Durham, P.G.; Butnariu, A.; Alghorazi, R.; Pinton, G.; Krishna, V.; Dayton, P.A. Current clinical investigations of focused ultrasound blood-brain barrier disruption: A review. Neurotherapeutics 2024, 21, e00352. [Google Scholar] [CrossRef]
Figure 1.
Blood-brain barrier (BBB) transport routes and where delivery platforms intervene: passive limits, engineered influx, and efflux ceilings. The schematic summarizes dominant BBB transport routes relevant to phytochemicals and highlights where delivery platforms intervene. At an intact BBB, paracellular diffusion across tight junctions is negligible and disease-dependent; controlled opening is best treated as a localized modulation strategy (e.g., focused ultrasound (FUS) with microbubbles) rather than a baseline assumption. Transcellular passive diffusion depends on size, polarity, hydrogen bonding, and lipophilicity, but apparent permeability is often capped by metabolism and active efflux. ABC transporters—P-gp, BCRP, MRPs—frequently set the ceiling for unbound brain exposure, motivating efflux-evading prodrugs, stealth/corona control for nanocarriers, and early efflux-liability screening. For polar phytochemicals, carrier-mediated transport (CMT) via GLUT1, LAT1, and MCT1 enables engineered influx using nutrient-mimetic promoieties, requiring Km/Vmax-aware design and brain-selective cleavage. For larger cargos, receptor-mediated transcytosis (RMT) via TfR and LRP1 supports ligand-decorated nanocarriers, but demands an affinity “sweet spot” to avoid endothelial sequestration and lysosomal routing. Adsorptive/CPP uptake can boost entry yet trades specificity for off-target risk; charge-switchable or stimulus-unmasked coatings can mitigate this. Intranasal delivery provides a complementary bypass for suitable payloads. ABC, ATP-binding cassette; BBB, blood–brain barrier; BCRP, breast cancer resistance protein; CMT, carrier-mediated transport; CPP, cell-penetrating peptide; FUS, focused ultrasound; GLUT1, glucose transporter 1; Km, Michaelis constant; LAT1, L-type amino acid transporter 1; LRP1, low-density lipoprotein receptor–related protein 1; MCT1, monocarboxylate transporter 1; MRPs, multidrug resistance–associated proteins; P-gp, P-glycoprotein; RMT, receptor-mediated transcytosis; TfR, transferrin receptor; Vmax, maximum transport rate.
Figure 1.
Blood-brain barrier (BBB) transport routes and where delivery platforms intervene: passive limits, engineered influx, and efflux ceilings. The schematic summarizes dominant BBB transport routes relevant to phytochemicals and highlights where delivery platforms intervene. At an intact BBB, paracellular diffusion across tight junctions is negligible and disease-dependent; controlled opening is best treated as a localized modulation strategy (e.g., focused ultrasound (FUS) with microbubbles) rather than a baseline assumption. Transcellular passive diffusion depends on size, polarity, hydrogen bonding, and lipophilicity, but apparent permeability is often capped by metabolism and active efflux. ABC transporters—P-gp, BCRP, MRPs—frequently set the ceiling for unbound brain exposure, motivating efflux-evading prodrugs, stealth/corona control for nanocarriers, and early efflux-liability screening. For polar phytochemicals, carrier-mediated transport (CMT) via GLUT1, LAT1, and MCT1 enables engineered influx using nutrient-mimetic promoieties, requiring Km/Vmax-aware design and brain-selective cleavage. For larger cargos, receptor-mediated transcytosis (RMT) via TfR and LRP1 supports ligand-decorated nanocarriers, but demands an affinity “sweet spot” to avoid endothelial sequestration and lysosomal routing. Adsorptive/CPP uptake can boost entry yet trades specificity for off-target risk; charge-switchable or stimulus-unmasked coatings can mitigate this. Intranasal delivery provides a complementary bypass for suitable payloads. ABC, ATP-binding cassette; BBB, blood–brain barrier; BCRP, breast cancer resistance protein; CMT, carrier-mediated transport; CPP, cell-penetrating peptide; FUS, focused ultrasound; GLUT1, glucose transporter 1; Km, Michaelis constant; LAT1, L-type amino acid transporter 1; LRP1, low-density lipoprotein receptor–related protein 1; MCT1, monocarboxylate transporter 1; MRPs, multidrug resistance–associated proteins; P-gp, P-glycoprotein; RMT, receptor-mediated transcytosis; TfR, transferrin receptor; Vmax, maximum transport rate.
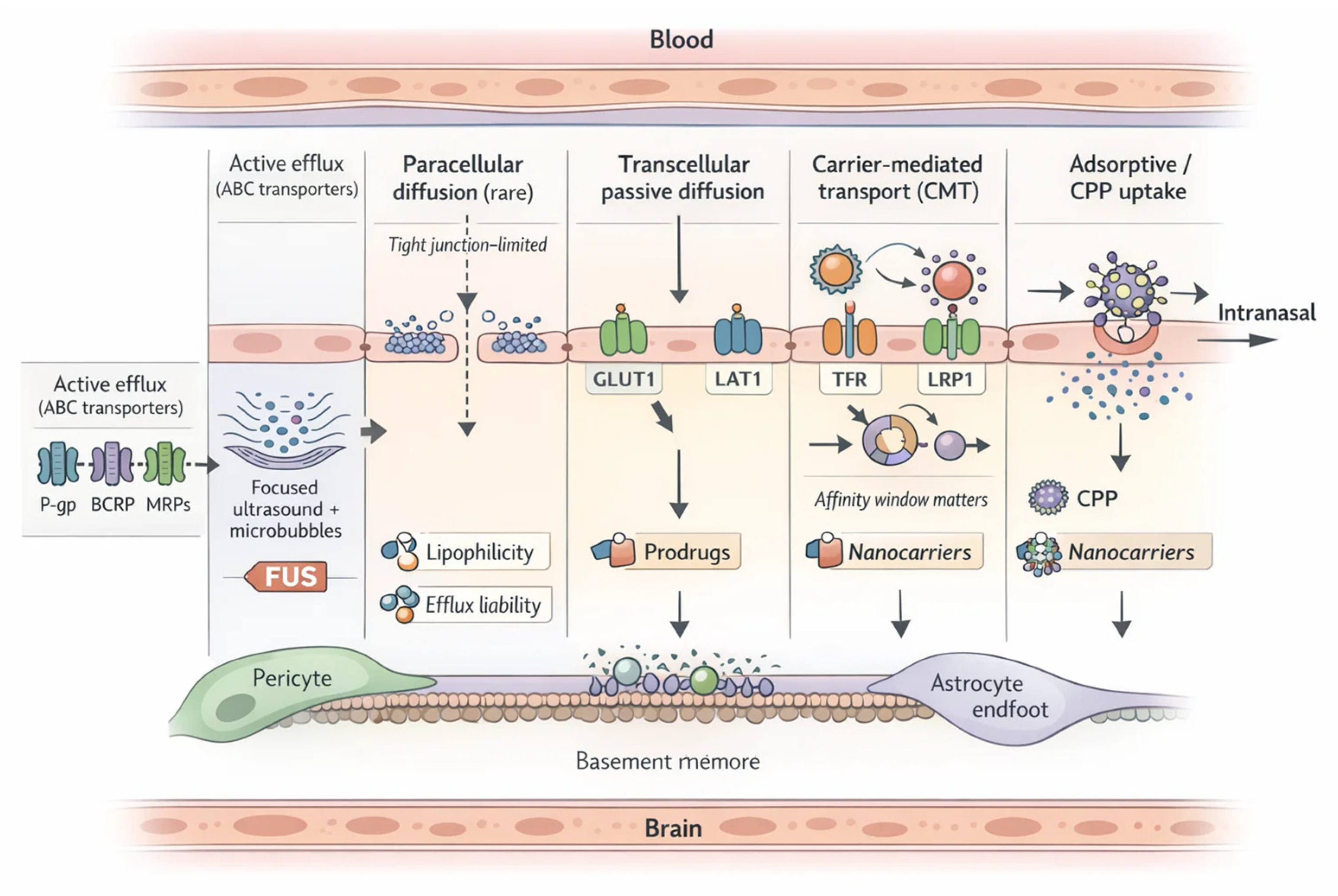
Figure 2.
Decision tree for selecting CNS delivery strategies by payload liabilities and clinical context. The schematic provides a pragmatic decision tree to match a CNS payload to the most efficient delivery strategy. The workflow begins with dominant liabilities: (i) solubility/dissolution limitation, (ii) efflux liability (e.g., P-gp/BCRP/MRPs), and (iii) chemical or metabolic instability. Solubility-driven failure routes first to formulation solutions (solubilizing vehicles, lipid systems, polymeric nanoparticles, mucoadhesive/intranasal formats when appropriate). Predominant efflux liability routes to prodrug design (efflux-evading promoieties or transporter-hijacking approaches such as GLUT1/LAT1/MCT1) and/or carrier shielding/targeting (stealth/corona control; RMT ligands such as TfR/LRP1). Instability-driven failure routes to protective encapsulation or stability-optimized prodrugs with controlled release. Clinical context modifiers (need for rapid onset, diffuse vs focal pathology, and tolerability constraints) direct selection of route (e.g., intranasal bypass) or focused ultrasound (FUS) + microbubbles for localized BBB modulation. Each terminal node specifies decision-grade endpoints: brain exposure (AUC, Kp,brain, Kp,uu), target engagement, safety margin, and CMC scalability. AUC, area under the curve; BBB, blood–brain barrier; BCRP, breast cancer resistance protein; CMC, chemistry, manufacturing, and controls; CNS, central nervous system; FUS, focused ultrasound; GLUT1, glucose transporter 1; Kp,brain, brain-to-plasma partition coefficient; Kp,uu, unbound brain-to-plasma partition coefficient; LAT1, L-type amino acid transporter 1; MCT1, monocarboxylate transporter 1; MRPs, multidrug resistance–associated proteins; P-gp, P-glycoprotein; RMT, receptor-mediated transcytosis; TfR, transferrin receptor; LRP1, low-density lipoprotein receptor–related protein 1.
Figure 2.
Decision tree for selecting CNS delivery strategies by payload liabilities and clinical context. The schematic provides a pragmatic decision tree to match a CNS payload to the most efficient delivery strategy. The workflow begins with dominant liabilities: (i) solubility/dissolution limitation, (ii) efflux liability (e.g., P-gp/BCRP/MRPs), and (iii) chemical or metabolic instability. Solubility-driven failure routes first to formulation solutions (solubilizing vehicles, lipid systems, polymeric nanoparticles, mucoadhesive/intranasal formats when appropriate). Predominant efflux liability routes to prodrug design (efflux-evading promoieties or transporter-hijacking approaches such as GLUT1/LAT1/MCT1) and/or carrier shielding/targeting (stealth/corona control; RMT ligands such as TfR/LRP1). Instability-driven failure routes to protective encapsulation or stability-optimized prodrugs with controlled release. Clinical context modifiers (need for rapid onset, diffuse vs focal pathology, and tolerability constraints) direct selection of route (e.g., intranasal bypass) or focused ultrasound (FUS) + microbubbles for localized BBB modulation. Each terminal node specifies decision-grade endpoints: brain exposure (AUC, Kp,brain, Kp,uu), target engagement, safety margin, and CMC scalability. AUC, area under the curve; BBB, blood–brain barrier; BCRP, breast cancer resistance protein; CMC, chemistry, manufacturing, and controls; CNS, central nervous system; FUS, focused ultrasound; GLUT1, glucose transporter 1; Kp,brain, brain-to-plasma partition coefficient; Kp,uu, unbound brain-to-plasma partition coefficient; LAT1, L-type amino acid transporter 1; MCT1, monocarboxylate transporter 1; MRPs, multidrug resistance–associated proteins; P-gp, P-glycoprotein; RMT, receptor-mediated transcytosis; TfR, transferrin receptor; LRP1, low-density lipoprotein receptor–related protein 1.
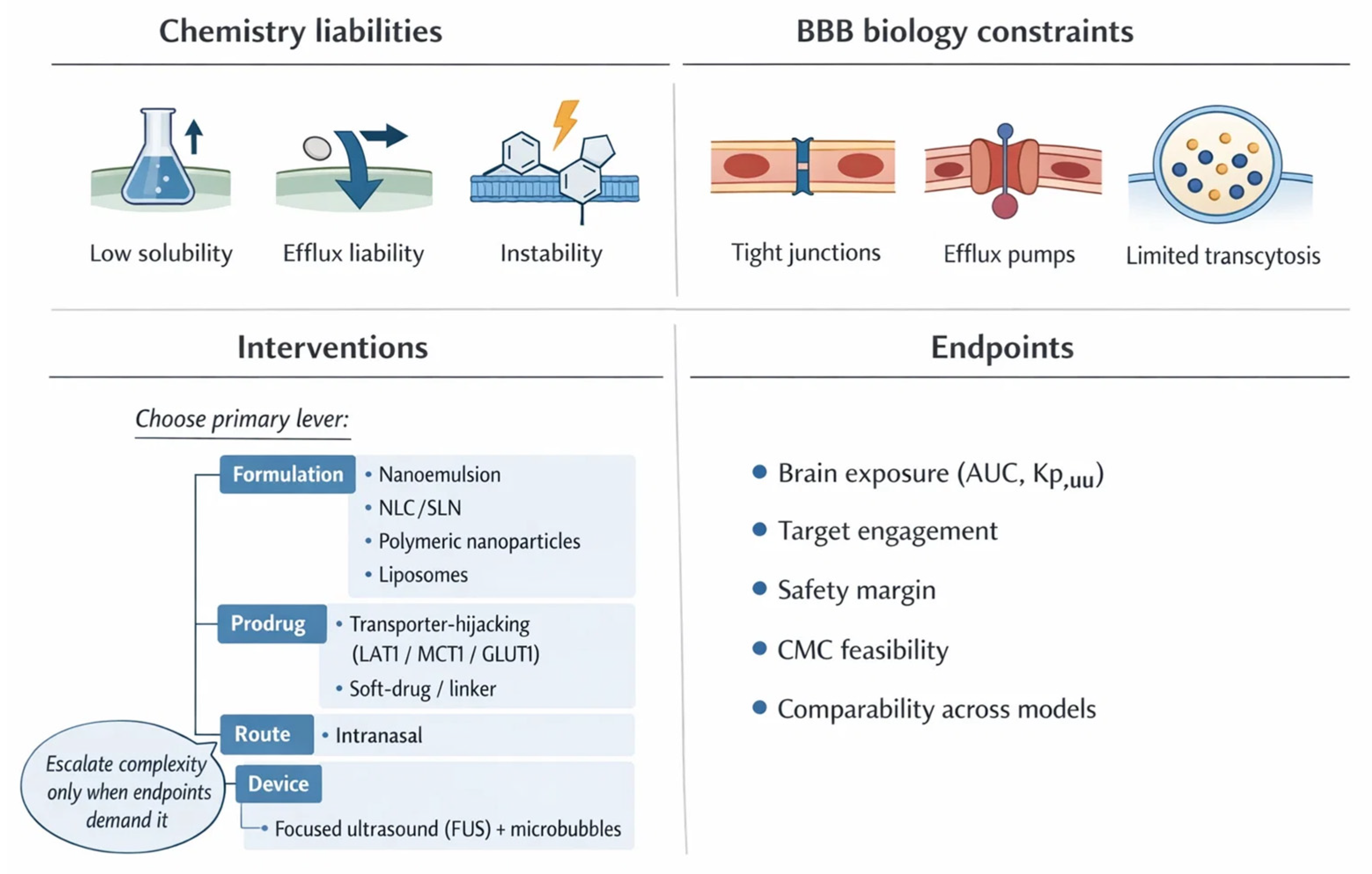
Figure 4.
Roadmap timeline for brain delivery platforms: near-term deployables versus long-term maturation, with validation gates and dependencies. Gantt-style bands summarize expected maturation windows for CNS delivery strategies and the dependencies that determine whether programs advance from feasibility to translation. Near-term (2–4 years) tracks emphasize “deployable” options with the clearest CMC and endpoint pathways: ligand-targeted PLGA nanoparticles/liposomes (e.g., transferrin or RVG; resveratrol/curcumin, including dual loading) gated by reproducible size/PDI, loading, and stability under scalable unit operations; intranasal mucoadhesive nanoemulsions/nanoemulgels (chitosan or thermo/in situ gels) gated by droplet-size distribution, rheology, spray plume metrics, preservative compatibility, and tolerability plus PK surrogates (CSF or imaging-rich substudies where justified); LAT1-anchored prodrugs gated by LAT1 affinity, bioconversion kinetics, intrabrain distribution, and target engagement in neurons/glia; and focused ultrasound–assisted regional delivery gated by imaging-confirmed BBB opening, local PK confirmation, and regulator-recognized safety monitoring. Longer-term (5–10+ years) bands capture exosome mimetics/hybrid vesicles (GMP scale-up and heterogeneity control), AI-designed multi-target nano–prodrugs (auditable datasets and prospective validation), remote-triggered release (dosimetry and repeat-use safety standards), and BBB “digital twins” integrating PBPK/PKPD with real-time TEER and registries. AI, artificial intelligence; BBB, blood–brain barrier; CMC, chemistry, manufacturing, and controls; CNS, central nervous system; CSF, cerebrospinal fluid; GMP, good manufacturing practice; LAT1, large neutral amino acid transporter 1; PBPK, physiologically based pharmacokinetic; PDI, polydispersity index; PK, pharmacokinetics; PKPD, pharmacokinetic–pharmacodynamic; PLGA, poly(lactic-co-glycolic acid); RVG, rabies virus glycoprotein; TEER, transendothelial electrical resistance.
Figure 4.
Roadmap timeline for brain delivery platforms: near-term deployables versus long-term maturation, with validation gates and dependencies. Gantt-style bands summarize expected maturation windows for CNS delivery strategies and the dependencies that determine whether programs advance from feasibility to translation. Near-term (2–4 years) tracks emphasize “deployable” options with the clearest CMC and endpoint pathways: ligand-targeted PLGA nanoparticles/liposomes (e.g., transferrin or RVG; resveratrol/curcumin, including dual loading) gated by reproducible size/PDI, loading, and stability under scalable unit operations; intranasal mucoadhesive nanoemulsions/nanoemulgels (chitosan or thermo/in situ gels) gated by droplet-size distribution, rheology, spray plume metrics, preservative compatibility, and tolerability plus PK surrogates (CSF or imaging-rich substudies where justified); LAT1-anchored prodrugs gated by LAT1 affinity, bioconversion kinetics, intrabrain distribution, and target engagement in neurons/glia; and focused ultrasound–assisted regional delivery gated by imaging-confirmed BBB opening, local PK confirmation, and regulator-recognized safety monitoring. Longer-term (5–10+ years) bands capture exosome mimetics/hybrid vesicles (GMP scale-up and heterogeneity control), AI-designed multi-target nano–prodrugs (auditable datasets and prospective validation), remote-triggered release (dosimetry and repeat-use safety standards), and BBB “digital twins” integrating PBPK/PKPD with real-time TEER and registries. AI, artificial intelligence; BBB, blood–brain barrier; CMC, chemistry, manufacturing, and controls; CNS, central nervous system; CSF, cerebrospinal fluid; GMP, good manufacturing practice; LAT1, large neutral amino acid transporter 1; PBPK, physiologically based pharmacokinetic; PDI, polydispersity index; PK, pharmacokinetics; PKPD, pharmacokinetic–pharmacodynamic; PLGA, poly(lactic-co-glycolic acid); RVG, rabies virus glycoprotein; TEER, transendothelial electrical resistance.
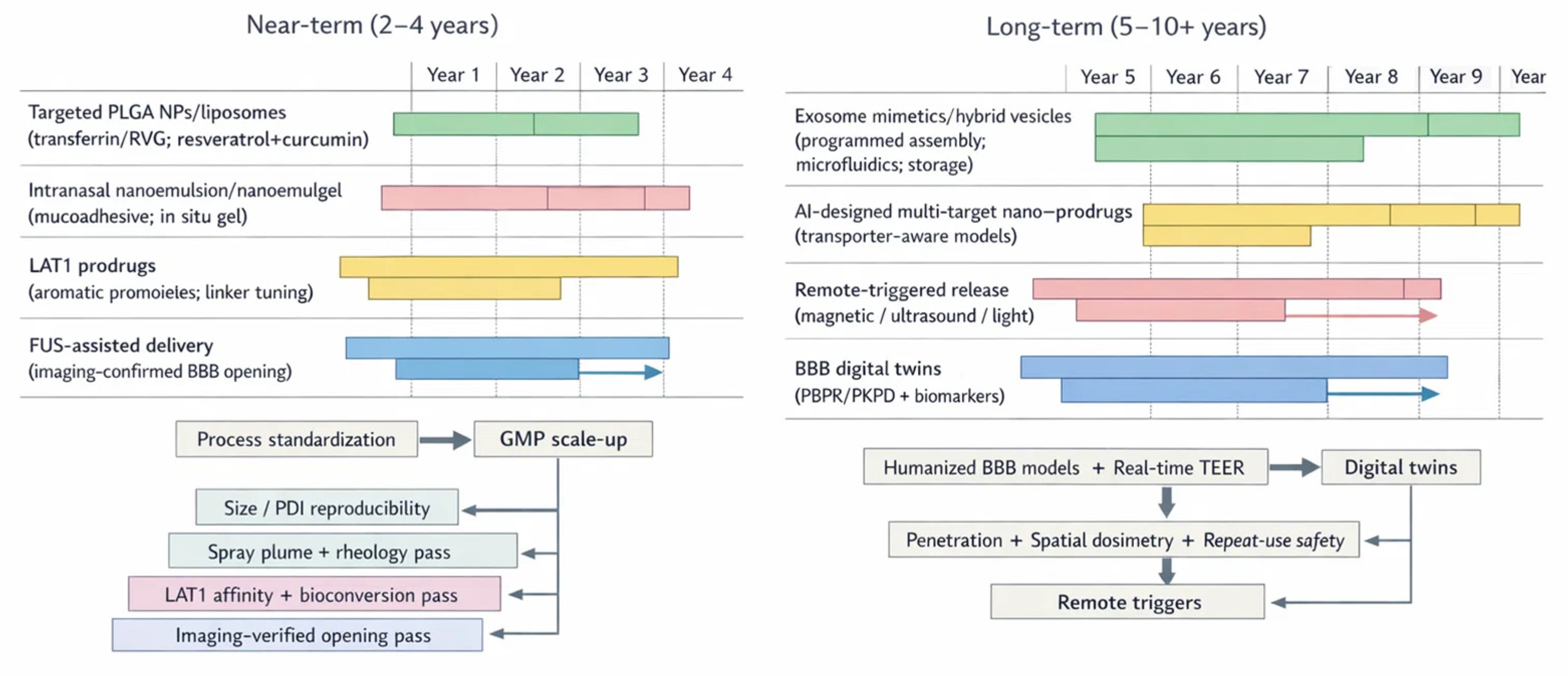

Table 1.
Blood-brain barrier (BBB) transport machineries and implications for phytochemicals. This table summarizes the dominant BBB transport routes relevant to plant-derived neurotherapeutics and links each route to pragmatic design levers. Pathways are framed as decision levers: passive diffusion is constrained by physicochemical liabilities common in polyphenols; carrier-mediated transport and receptor-mediated transcytosis provide engineered influx opportunities; adsorptive and cell-penetrating peptide strategies can amplify uptake but trade specificity for risk; and efflux pumps (P-gp, BCRP, MRPs) often set the ceiling for unbound brain exposure even when in vitro permeability appears promising.
Table 1.
Blood-brain barrier (BBB) transport machineries and implications for phytochemicals. This table summarizes the dominant BBB transport routes relevant to plant-derived neurotherapeutics and links each route to pragmatic design levers. Pathways are framed as decision levers: passive diffusion is constrained by physicochemical liabilities common in polyphenols; carrier-mediated transport and receptor-mediated transcytosis provide engineered influx opportunities; adsorptive and cell-penetrating peptide strategies can amplify uptake but trade specificity for risk; and efflux pumps (P-gp, BCRP, MRPs) often set the ceiling for unbound brain exposure even when in vitro permeability appears promising.
| Pathway | Molecular prerequisites | Exemplars | Impact on phytochemicals | Engineering lever(s) | References |
|---|---|---|---|---|---|
| Paracellular diffusion (tight junction limited) | Effectively negligible at an intact BBB; requires transient junction loosening or pathological leak | Small hydrophiles in disease-associated leak states | Native polyphenols remain largely excluded; leak is disease- and region-dependent and poorly controllable | Localized opening approaches (e.g., focused ultrasound with microbubbles); avoid programs that depend on nonspecific leak | [62,63,64] |
| Transcellular passive diffusion | Small size, low polarity, limited H-bonding; favorable lipophilicity; minimal efflux liability | CNS-permeable small molecules; selected alkaloids | Many phytochemicals exceed polarity and H-bonding windows; metabolism and efflux can negate apparent permeability | Prodrug or soft-drug design; tune logD and polar surface area; stabilize against first-pass metabolism; solubility enabling formulations | [65,66,67] |
| Carrier-mediated transport (CMT) | Structural mimicry of endogenous nutrients; transporter affinity plus adequate chemical stability | GLUT1 (glucose), LAT1 (large neutral amino acids), MCTs (monocarboxylates) | Provides an influx handle for polar phytochemicals, but competition with endogenous substrates and species differences can limit delivery | Transporter-hijacking prodrugs (amino acid, glucose, monocarboxylate promoieties); Km/Vmax-aware design; brain-selective cleavage | [68,69,70] |
| Receptor-mediated transcytosis (RMT) | Ligand engagement within a productive affinity window; excessive avidity increases sequestration and lysosomal routing | Transferrin receptor, insulin receptor, LRP1 (targeting designs) | Enables macromolecular and nanoparticle shuttling, but ligand density and valency control release into brain parenchyma | Ligand-decorated nanocarriers; optimize affinity and ligand density; cleavable linkers; designs that favor recycling over degradation | [53,71,72] |
| Adsorptive-mediated transcytosis and CPP uptake | Net positive charge and/or CPP motifs; electrostatic interactions with endothelial glycocalyx | Tat, penetratin, RVG-derived peptides (as CPP/targeting motifs) | High uptake can trade specificity for off-target accumulation and cytotoxicity; "more cationic" is not always better | Charge-switchable coatings; stimulus-unmasking CPPs; cap surface charge; combine with targeting ligands to improve selectivity | [53,67,71] |
| Active efflux (ABC transporters) | Substrate recognition by ATP-driven pumps; efflux can dominate even when passive permeability is favorable | P-gp, BCRP, MRPs | A key barrier for many polyphenols; inhibition or induction can shift CNS exposure unpredictably across age, disease, and comedication | Efflux-evading prodrugs; corona control and stealth coatings; carrier strategies that reduce free substrate at the luminal membrane; early efflux liability screening | [65,66,67] |
ABC ATP-binding cassette; BBB, blood–brain barrier; BCRP, breast cancer resistance protein; CMT, carrier-mediated transport; CNS, central nervous system; CPP, cell-penetrating peptide; GLUT1, glucose transporter 1; Km, Michaelis constant; LAT1, L-type amino acid transporter 1; LRP1, low-density lipoprotein receptor–related protein 1; MCTs, monocarboxylate transporters; MRPs, multidrug resistance–associated proteins; P-gp, P-glycoprotein; RVG, rabies virus glycoprotein; RMT, receptor-mediated transcytosis; Vmax, maximum transport rate.
Table 2.
Translational map from phytochemical class to key delivery liabilities and practical enabling strategies for central nervous system (CNS) development across blood-brain barrier (BBB) constrained programs. Phytochemical scaffolds share recurring developability bottlenecks at the BBB, yet the dominant liability differs by class. The table links representative compounds discussed in the manuscript to the most common physicochemical and biopharmaceutical constraints, then pairs each class with a preferred enabling strategy that is compatible with scale-up and safety screening.
Table 2.
Translational map from phytochemical class to key delivery liabilities and practical enabling strategies for central nervous system (CNS) development across blood-brain barrier (BBB) constrained programs. Phytochemical scaffolds share recurring developability bottlenecks at the BBB, yet the dominant liability differs by class. The table links representative compounds discussed in the manuscript to the most common physicochemical and biopharmaceutical constraints, then pairs each class with a preferred enabling strategy that is compatible with scale-up and safety screening.
| Class | Representative compounds | Primary liabilities | Preferred enabling strategy | Notes (e.g., stability, taste, ionization) | References |
|---|---|---|---|---|---|
| Polyphenols (flavonoids, stilbenes, curcuminoids) | Resveratrol; quercetin; curcumin | Low brain exposure despite in vitro potency; extensive phase II metabolism (UGT/SULT); efflux liability (P-gp/BCRP); often high polarity or poor solubility; chemical instability (oxidation or hydrolysis). | Prodrug or transporter-hijacking promoieties; nanoencapsulation (polymeric NPs, liposomes, SLNs) to protect the scaffold and modulate release; intranasal or BBB opening adjuncts when justified. | Phenolic acids are weak acids with context-dependent ionization; many are light and pH sensitive; bitter or astringent taste can limit oral dosing and adherence. | [149,150,151] |
| Alkaloids | Berberine; galantamine | Ionization and strong transporter interactions can cap CNS entry; variable oral bioavailability; efflux driven exposure variability; class dependent safety margins and CYP interactions. | Salt selection plus lipid-based carriers or polymeric micelles; efflux bypass via prodrug or nanocarrier shielding; leverage high target potency with lower systemic exposure through controlled release or alternative routes. | Often strongly bitter; typical basic pKa yields cationic fraction at physiological pH; galantamine illustrates that pharmacodynamic targeting can partly offset limited brain partitioning. | [110,116,149] |
| Terpenoids (mono-, sesqui-, diterpenes) | Ginkgolides; pinene; linalool | High lipophilicity with low aqueous solubility; volatility for some monoterpenes; oxidative degradation; high protein binding and rapid metabolism leading to variable CNS exposure. | Self-emulsifying systems, nanoemulsions, cyclodextrin inclusion, or lipid nanoparticles to raise apparent solubility and stabilize the payload; intranasal formulations for rapid onset when appropriate. | Usually neutral with limited ionization; aroma and taste are prominent; irritation potential should be checked for concentrated essential-oil-like components. | [149,151,152] |
| Cannabinoids | Cannabidiol (CBD); THC | Formulation-limited absorption and marked individual variability; extensive hepatic metabolism; drug-drug interactions; psychoactivity and regulatory constraints for THC; long tissue residence due to lipophilicity. | Lipid vehicles, nanoemulsions, polymeric carriers, or controlled release depots; route optimization (oromucosal, intranasal) and dose fractionation to reduce peak related adverse effects while maintaining exposure. | Light and oxygen sensitive; taste can be limiting; largely neutral but highly lipophilic; legal status and labeling requirements can shape trial design. | [153,154,155] |
BBB, blood–brain barrier; BCRP, breast cancer resistance protein; CBD, cannabidiol; CNS, central nervous system; CYP, cytochrome P450; NPs, nanoparticles; pKa, acid dissociation constant; P-gp, P-glycoprotein; SLNs, solid lipid nanoparticles; SULT, sulfotransferases; THC, Δ9-tetrahydrocannabinol; UGT, UDP-glucuronosyltransferases.
Table 3.
Nanocarrier platforms for blood-brain barrier (BBB)-constrained phytochemicals: design levers, translational performance, and CMC/GMP-critical considerations. This table offers a decision-oriented snapshot of major nanocarrier platforms for brain delivery, linking each to core/shell materials, surface strategies (PEG, receptor ligands, corona control), loading modes, and release logic. Release is categorized as constitutive (diffusion/erosion) or stimulus-enabled (pH/redox/enzymes; external triggers such as magnetic fields or focused ultrasound). Key trade-offs include leakage versus retention (liposomes), payload expulsion (SLNs), reproducibility/stability limits (nanoemulsions), multivalency versus toxicity (dendrimers/CPP-like surfaces), long-term safety uncertainty (inorganic systems), and batch heterogeneity/regulatory issues (exosomes). CMC/GMP notes emphasize QbD priorities: size/PDI, zeta potential, encapsulation efficiency, release kinetics, sterility/endotoxin, and scalable manufacture.
Table 3.
Nanocarrier platforms for blood-brain barrier (BBB)-constrained phytochemicals: design levers, translational performance, and CMC/GMP-critical considerations. This table offers a decision-oriented snapshot of major nanocarrier platforms for brain delivery, linking each to core/shell materials, surface strategies (PEG, receptor ligands, corona control), loading modes, and release logic. Release is categorized as constitutive (diffusion/erosion) or stimulus-enabled (pH/redox/enzymes; external triggers such as magnetic fields or focused ultrasound). Key trade-offs include leakage versus retention (liposomes), payload expulsion (SLNs), reproducibility/stability limits (nanoemulsions), multivalency versus toxicity (dendrimers/CPP-like surfaces), long-term safety uncertainty (inorganic systems), and batch heterogeneity/regulatory issues (exosomes). CMC/GMP notes emphasize QbD priorities: size/PDI, zeta potential, encapsulation efficiency, release kinetics, sterility/endotoxin, and scalable manufacture.
| Platform | Core/shell materials | Size/PDI | Surface (PEG/ligand) | Loading | Release trigger | Pros/Cons | Exemplar payloads | Notes (CMC/GMP) | References |
|---|---|---|---|---|---|---|---|---|---|
| Polymeric nanoparticles (PLGA, PEG-PLGA; chitosan-coated/hybrids) | PLGA or PEG-PLGA matrix; optional chitosan coating or chitosan microparticle embedding | Tunable; low PDI targeted as a core CQA | PEG “stealth”; Angiopep-2 and other RMT ligands; corona-engineering approaches | Encapsulation of hydrophobic phytocompounds; co-loading feasible; drug protected from metabolism | Diffusion + polymer erosion; can add pH/redox/enzyme-responsive elements | Pros: biodegradable, sustained release, strong stability. Cons: MPS uptake; burst-release risk if not tuned; process sensitivity | Resveratrol; curcumin; flavonoids; intranasal chitosan–PLGA example: gemcitabine | QbD/QTPP with CQAs (size, PDI, zeta potential, encapsulation efficiency, release kinetics); sterility/endotoxin control; scale-up reproducibility | [61,220,226] |
| Liposomes (including PEGylated and ligand-decorated) | Phospholipid bilayer ± cholesterol; aqueous core + hydrophobic bilayer domain | Tunable; control needed to limit leakage and maintain uniformity | PEGylation for circulation; modular ligand functionalization for BBB targeting | Hydrophilic cargo in core; hydrophobic in bilayer; co-encapsulation possible | Constitutive leakage/partitioning; can be engineered for thermosensitive or pH-triggered release | Pros: versatile loading; rapid brain access in some models. Cons: leakage and reduced retention; stability/shelf-life constraints | Resveratrol; curcumin; peptide cargos (as discussed in platform comparisons) | Composition and lipid raw-material controls; filtration-compatible sterility where feasible; stability/lyophilization programs to preserve CQAs | [221,222,227] |
| Solid lipid nanoparticles (SLNs) and nanostructured lipid carriers (NLCs) | SLN: solid lipid matrix. NLC: mixed solid + liquid lipids to increase drug accommodation | Typically robust colloids when optimized; low PDI targeted as a translation enabler | Surfactant-stabilized; PEG and BBB ligands can be incorporated as needed | Best for lipophilic phytochemicals; NLCs improve loading vs SLNs | Diffusion from lipid matrix; matrix reorganization influences kinetics | Pros: biocompatible, controlled release, improved stability. Cons: SLNs limited loading; storage-driven drug expulsion risk reduced in NLCs | Polyphenols; terpenoids; chronic neurodegeneration-oriented formulations | Control lipid polymorphism, surfactant system, and storage conditions; monitor size/PDI, encapsulation efficiency, leakage/expulsion during stability testing | [228,229,230] |
| Nanoemulsions / nanoemulgels (often intranasal) | Oil-in-water droplets with surfactants/co-surfactants; optional mucoadhesive gel phase | Small droplet size emphasized for uptake; distribution width is a key CQA | Mucoadhesive polymers (e.g., chitosan coatings) possible; ligand targeting less common than for NPs | Solubilization of hydrophobic polyphenols and terpenoids; formulation-driven payload stabilization | Rapid absorption via olfactory epithelium; residence-time extension via in situ gels (thermo/ion-responsive) | Pros: fast nose-to-brain access; bypass first-pass metabolism. Cons: manufacturing reproducibility and aggregation/shelf-life challenges | Curcumin; resveratrol; terpenoids; cannabinoid-class examples referenced under lipid vehicles | CMC focuses on droplet-size distribution, rheology (for gels), spray plume/metered dosing, preservative compatibility, and long-term stability | [227,231,232] |
| Dendrimers (e.g., PAMAM, carbosilane) | Monodisperse branched polymers; generation-defined architecture; optional PEGylation/mixed surfaces | Defined by generation; surface charge is a dominant performance/safety dial | Multivalent ligand attachment (RMT or disease-homing); PEG/mixed-surface designs to reduce toxicity | Internal cavity encapsulation and/or covalent conjugation; suitable for peptides and nucleic acids | Cleavable linkers (pH/redox/enzyme) for conjugates; diffusion for encapsulated small molecules | Pros: programmable multivalency and targeting density. Cons: cationic toxicity risk and clearance trade-offs with high functionalization | Peptide cargos; nucleic acids (siRNA-class examples) | Tight control of generation, residual monomers/solvents, surface substitution ratio; sterility/endotoxin; charge-related immunotoxicity screening | [223,224,225] |
| Polymeric micelles | Amphiphilic block copolymers that self-assemble (hydrophobic core + hydrophilic corona) | Nanosized aggregates; stability to dilution is a critical attribute | PEG corona common; charge tuning (including cation-free designs) to reduce cytotoxicity | Solubilizes poorly water-soluble phytochemicals; can carry nucleic acids with tailored chemistry | Physiologic triggers (acidic pH, enzymes) that destabilize micelles and release cargo | Pros: strong solubilization; triggerable release. Cons: disassembly risk in vivo; formulation sensitivity to concentration and serum interactions | Hydrophobic phytochemicals; siRNA delivery examples discussed in glioblastoma context | Define CMC/CQA set including size/PDI, critical micelle concentration, loading, and release; assess serum stability and storage (lyophilization) effects | [220,224,225] |
| Inorganic/carbon nanostructures (carbon dots/quantum dots; magnetic cores) | Carbon dots or quantum-dot-like cores; magnetic nanoparticles; polymer/PEG coatings for biocompatibility | Engineered nanoscale; surface chemistry dominates biodistribution and clearance | Functional coatings (polymer/PEG) and ligands for targeting; magnetic guidance as an external control layer | Conjugation/adsorption of small molecules; intrinsic imaging signal enables theranostics | External triggers (magnetic fields/light) and surface chemistry-dependent release | Pros: tracking + delivery; controllable guidance. Cons: long-term safety/retention uncertainty; oxidation and accumulation concerns | Theranostic small molecules; imaging-enabled CNS delivery constructs | Extended toxicology and clearance characterization; strict control of surface chemistry and impurities; justification of long-term tissue persistence risk | [218,219,226] |
| Hybrid/biomimetic systems and corona-controlled designs | Polymer–lipid hybrids; inorganic–organic hybrids; membrane-coated nanoparticles; polymalic acid nanodrugs | Tunable; multi-component systems amplify sensitivity to process drift | Dual-targeting (e.g., Angiopep-2 + CPP/TAT); corona engineering via pre-coating or biomimetic shells | Multi-cargo capability; can integrate prodrug chemistry with carrier features | Layered control: diffusion/erosion plus internal (pH/redox/enzyme) or external (ultrasound/light) triggers | Pros: integrates stability, targeting, and triggerable release. Cons: CMC complexity; regulatory ambiguity for multifunctional hybrids | Angiopep-2-conjugated nanodrug examples; dual-ligand lipid nanocarriers in glioblastoma models | PAT-enabled monitoring; tight incoming material specs; control strategy for surface functionalization and corona; batch-to-batch fidelity prioritized | [218,233,234] |
| Biogenic vesicles (mammalian exosomes; plant-derived EVs; synthetic mimetics) | Natural EV lipid bilayers (mammalian or plant-derived) or synthetic mimetics (polymersomes/membrane-coated NPs) | Often heterogeneous; purity and population definition are key | Innate tropism plus optional ligand decoration; mimetics allow controlled ligand presentation | Electroporation/incubation/sonication/extrusion for small molecules, proteins, and nucleic acids; PDEVs may carry intrinsic bioactives | Cellular uptake/endosomal trafficking; release depends on uptake and intracellular routing | Pros: high biocompatibility; potential tropism; PDEVs are abundant and low immunogenic. Cons: batch variability, yield/purification and regulatory classification; BBB mechanisms for PDEVs less defined | Small molecules, proteins, nucleic acids; antioxidant/anti-inflammatory cargos in neuroinflammation models | Standardize source, isolation, and potency assays; define acceptable heterogeneity; scale-up and regulatory pathway planning are decisive | [219,225,235] |
BBB, blood–brain barrier; CMC, chemistry, manufacturing, and controls; CNS, central nervous system; CPP, cell-penetrating peptide; CQA, critical quality attribute; EVs, extracellular vesicles; NLCs, nanostructured lipid carriers; NPs, nanoparticles; PAMAM, poly(amidoamine); PAT, process analytical technology; PDEVs, plant-derived extracellular vesicles; PEG, polyethylene glycol; PLGA, poly(lactic-co-glycolic acid); QbD, quality by design; QTPP, quality target product profile; RMT, receptor-mediated transcytosis; siRNA, small interfering RNA; SLNs, solid lipid nanoparticles; TAT, trans-activator of transcription.
Table 4.
Receptor-mediated transcytosis (RMT) ligands for BBB delivery: binding/valency design levers, species caveats, and reported brain-exposure deltas. This table summarizes RMT ligand classes by target receptor, binding strength (K_d), and valency/ligand density—key determinants of productive transcytosis versus endothelial sequestration and lysosomal routing. Species caveats flag translation pitfalls, including endogenous ligand competition (e.g., transferrin/insulin), regional and disease-dependent receptor expression, and limited rodent–human cross-reactivity. Δ brain exposure records fold-changes (brain area under the curve (AUC), K_p,brain, or preferably K_p,uu) versus matched non-targeted controls under comparable dosing and sampling windows. “NR” indicates values not specified in the current draft and to be completed during final reference curation.
Table 4.
Receptor-mediated transcytosis (RMT) ligands for BBB delivery: binding/valency design levers, species caveats, and reported brain-exposure deltas. This table summarizes RMT ligand classes by target receptor, binding strength (K_d), and valency/ligand density—key determinants of productive transcytosis versus endothelial sequestration and lysosomal routing. Species caveats flag translation pitfalls, including endogenous ligand competition (e.g., transferrin/insulin), regional and disease-dependent receptor expression, and limited rodent–human cross-reactivity. Δ brain exposure records fold-changes (brain area under the curve (AUC), K_p,brain, or preferably K_p,uu) versus matched non-targeted controls under comparable dosing and sampling windows. “NR” indicates values not specified in the current draft and to be completed during final reference curation.
| Ligand/target | K_d range | Valency | Species caveats | Δ brain exposure (fold) | Notes | References |
|---|---|---|---|---|---|---|
| Transferrin (Tf) → TfR | NR in manuscript; productive “sweet spot” emphasized (avoid very high avidity) | Mono- to multivalent (ligand density-dependent) | Endogenous Tf competition; receptor expression varies by region/disease; rodent–human differences | NR; enter as fold vs non-targeted control (specify metric: AUC, K_p,brain, K_p,uu) | Canonical BBB shuttle; format and density govern recycling vs lysosomal routing; can be paired with parenchymal motifs | [276,277] |
| Anti-TfR antibodies / fragments / bispecific shuttles → TfR | NR in manuscript; affinity and epitope selection critical | Often monovalent/low-avidity formats preferred; bispecific designs common | Epitope-specific species cross-reactivity; high affinity can increase trapping; saturation effects | NR; report with dosing window and comparator | Design goal is efficient transcytosis with minimal TfR downregulation and reduced endothelial retention | [278,279,280] |
| Angiopep-2 → LRP1 | NR in manuscript; ligand density and avidity tuning highlighted | Typically multivalent on nanocarriers; density optimized to avoid sequestration | LRP1 expression/context dependence; human relevance must be confirmed; tumor vs healthy BBB differences | NR; report relative to non-targeted carrier | Widely used peptide shuttle for nanoparticles and conjugates; can support glioblastoma-directed constructs | [281,282,283] |
| ApoE-mimetic peptides → LDLR (±LRP1) | NR in manuscript; affinity window and release kinetics emphasized | Mono- or multivalent; avidity increases uptake but can increase trapping | Strong endogenous ApoE/LDL competition; lipid-state effects; species differences in lipoprotein biology | NR; specify endpoint (brain/plasma ratio, AUC) | Leverages lipoprotein trafficking; cleavable linkers and controlled valency can aid parenchymal release | [284,285] |
| Insulin / engineered IR ligands / anti-IR formats → IR | NR in manuscript; avoid receptor saturation | Low-avidity designs generally favored | Physiological ceiling and safety constraints (glucose homeostasis); high endogenous competition; species differences | NR; report alongside safety/tolerability | Attractive but constrained by homeostatic receptor function; format and dosing are decisive | [240] |
| Aptamers / alternative binders (e.g., TfR- or LRP1-binding) → RMT receptors | NR in manuscript; receptor-specific values to be inserted | Usually monovalent; multimerization possible | Cross-reactivity and epitope mapping required; stability in plasma and nuclease resistance differ by species | NR; populate with harmonized assay definitions | Modular alternatives to peptides/antibodies; can reduce immunogenicity but require robust CMC characterization | [276,286] |
ApoE, apolipoprotein E; AUC, area under the curve; BBB, blood–brain barrier; CMC, chemistry, manufacturing, and controls; IR, insulin receptor; K_d, dissociation constant; K_p,brain, brain-to-plasma partition coefficient; K_p,uu, unbound brain-to-plasma partition coefficient; LDL, low-density lipoprotein; LDLR, low-density lipoprotein receptor; LRP1, low-density lipoprotein receptor–related protein 1; NR, not reported; RMT, receptor-mediated transcytosis; Tf, transferrin; TfR, transferrin receptor.
Table 5.
Alternative routes and device-enabled blood-brain barrier (BBB) modulation: evidence and risk–mitigation matrix for improving central nervous system (CNS) exposure. This table summarizes alternative routes and device-enabled strategies to bypass or locally modulate the BBB when systemic nanocarriers or prodrugs are insufficient. Evidence level (preclinical to clinical use) is paired with key translational risks. Intranasal delivery enables olfactory/trigeminal access but is limited by variability and dose volume. Focused ultrasound (FUS) with microbubbles provides precise, reversible BBB opening requiring cavitation-aware control and imaging verification. Osmotic/chemical disruption increases permeability at the cost of safety and control. Convection-enhanced delivery (CED) enables focal infusion but is invasive. External-field approaches remain exploratory.
Table 5.
Alternative routes and device-enabled blood-brain barrier (BBB) modulation: evidence and risk–mitigation matrix for improving central nervous system (CNS) exposure. This table summarizes alternative routes and device-enabled strategies to bypass or locally modulate the BBB when systemic nanocarriers or prodrugs are insufficient. Evidence level (preclinical to clinical use) is paired with key translational risks. Intranasal delivery enables olfactory/trigeminal access but is limited by variability and dose volume. Focused ultrasound (FUS) with microbubbles provides precise, reversible BBB opening requiring cavitation-aware control and imaging verification. Osmotic/chemical disruption increases permeability at the cost of safety and control. Convection-enhanced delivery (CED) enables focal infusion but is invasive. External-field approaches remain exploratory.
| Modality | Mechanism | Evidence level | Clinical status | Advantages | Risks/mitigations | References |
|---|---|---|---|---|---|---|
| Intranasal nose-to-brain (sprays, gels, nanoemulsions) | Direct transport along olfactory and trigeminal pathways; reduced first-pass metabolism; mucoadhesion prolongs residence | Preclinical strong; early clinical emerging (context-dependent) | Used clinically for some CNS-active small molecules; delivery platforms under evaluation for neurodegeneration/oncology | Noninvasive; rapid onset potential; bypasses systemic dilution for suitable payloads; compatible with solubility-enabling formulations | High inter-individual variability (anatomy, mucociliary clearance); limited dose volume; nasal irritation—mitigate with device optimization, deposition mapping, mucoadhesive/in situ gels, and PK endpoints (AUC, Kp,uu when feasible) | [4,155,329] |
| Focused ultrasound (FUS) + microbubbles | Pulsed acoustic exposure drives stable cavitation–mediated mechanoporation and transient tight-junction/transport changes enabling local BBB opening | Robust preclinical; multiple early clinical studies | Clinical translation under way (MRI-guided protocols in neuro-oncology and neurodegeneration) | Spatially targeted, reversible opening; compatible with diverse payloads (small molecules, prodrugs, nanocarriers, biologics); enables region-specific dosing | Hemorrhage/edema risk with inertial cavitation; off-target opening—mitigate via cavitation monitoring (acoustic emissions/PCD), conservative parameter sets, MRI guidance, contrast-enhanced confirmation, and predefined abort thresholds | [148,330,331] |
| Osmotic BBB disruption (intra-arterial mannitol) | Hyperosmolar shrinkage of endothelial cells transiently widens tight junctions and increases permeability | Established concept; variable evidence by indication and protocol | Applied in select centers/indications; invasive and less commonly used than device-guided opening | Can increase delivery of otherwise excluded agents; compatible with intra-arterial co-administration | Poor spatial control; seizure/edema/hemorrhage risk; procedure-related risks—mitigate with stringent patient selection, hemodynamic monitoring, imaging surveillance, and avoidance of programs relying on nonspecific leak | [148,330,332] |
| Chemical permeability modulation (selected permeabilizers/co-solvents) | Transiently alters membrane integrity, tight-junction signaling, or transporter function to raise permeability | Limited to mixed; often preclinical or adjunctive | Not routine for broad CNS delivery; used cautiously as adjuncts in narrow settings | Potentially simple to implement; can be paired with systemic dosing when local devices are unavailable | Nonspecific barrier disruption and systemic toxicity; unpredictable PK and inflammation—mitigate with minimal-effective exposure, local delivery where possible, tight safety biomarkers, and preference for controllable modalities | [148,332,333] |
| Convection-enhanced delivery (CED) | Pressure-driven interstitial infusion via intracranial catheter achieves high local concentrations independent of BBB transport | Strong preclinical/clinical experience in focal indications | Clinical use and trials in neuro-oncology and focal CNS targets; procedure-dependent | High local dose; bypasses efflux and systemic barriers; controllable infusion profiles; suitable for macromolecules and particles | Invasive; catheter placement errors, reflux/backflow, heterogeneous distribution, infection—mitigate with image-guided planning, real-time distribution tracking, optimized cannula design, and sterility controls | [326,334,335] |
| External-field targeting/triggering (magnetic guidance; remote release) | Magnetic gradients concentrate magnetically responsive carriers; external fields can trigger release from stimuli-responsive constructs | Primarily preclinical; exploratory translation | Investigational; requires specialized hardware and long-term safety data | Adds spatiotemporal control without barrier-wide opening; can pair with imaging-enabled carriers for tracking | Uncertain long-term retention/clearance; heating and off-target accumulation; device standardization gaps—mitigate with biocompatible coatings, rigorous dosimetry, biodistribution/clearance studies, and conservative escalation | [4,148] |
auc, area under the curve; BBB, blood–brain barrier; ced, convection-enhanced delivery; cns, central nervous system; fus, focused ultrasound; mri, magnetic resonance imaging; pcd, passive cavitation detection; pk, pharmacokinetics; kp,uu, unbound brain-to-plasma partition coefficient.
Table 6.
Prodrug design playbook for BBB delivery: transporter-hijacking promoieties, brain-selective cleavage logic, and translation risks. This table condenses transporter-hijacking prodrug strategies into a practical design checklist for BBB delivery. It centers on key influx carriers (LAT1, GLUT1, MCT1), outlining common promoieties, linker chemistries, and brain-selective cleavage triggers that mitigate polarity, efflux, and instability while limiting premature systemic activation. Exposure gain is reported as fold-change versus parent or non-targeted controls using harmonized endpoints (brain AUC, Kp,brain, preferably Kp,uu), with “NR” for values not specified. Off-target risks include substrate competition, peripheral uptake, species differences, and unintended metabolite activity; notes emphasize Km/Vmax-aware design, linker stability, and assay/QC needs.
Table 6.
Prodrug design playbook for BBB delivery: transporter-hijacking promoieties, brain-selective cleavage logic, and translation risks. This table condenses transporter-hijacking prodrug strategies into a practical design checklist for BBB delivery. It centers on key influx carriers (LAT1, GLUT1, MCT1), outlining common promoieties, linker chemistries, and brain-selective cleavage triggers that mitigate polarity, efflux, and instability while limiting premature systemic activation. Exposure gain is reported as fold-change versus parent or non-targeted controls using harmonized endpoints (brain AUC, Kp,brain, preferably Kp,uu), with “NR” for values not specified. Off-target risks include substrate competition, peripheral uptake, species differences, and unintended metabolite activity; notes emphasize Km/Vmax-aware design, linker stability, and assay/QC needs.
| Transporter | Promoiety/linker | Cleavage trigger | Exposure gain | Off-target risks | Notes | References |
|---|---|---|---|---|---|---|
| LAT1 (large neutral amino acid transporter) | L-amino acid promoieties (e.g., phenylalanine/leucine/tyrosine analogs); ester, amide, or carbamate linkers; optional self-immolative spacers | Brain-enriched esterases/peptidases; linker-enabled self-immolation after enzymatic trigger | NR (populate with fold-change in brain AUC, Kp,brain or Kp,uu) | Competition with endogenous amino acids; saturation at high dose; peripheral uptake (gut, kidney); rodent–human affinity/epitope differences | Prefer moderate affinity to favor flux over trapping; design should be Km/Vmax-aware; verify brain-selective cleavage and low systemic conversion; include efflux liability screening for released parent | [380,381,382] |
| GLUT1 (glucose transporter) | Glucose or glucosyl-like promoieties; O- or C-linked glycosides; carbonate/carbamate/ester linkers for release | Glycosidase-assisted unmasking (where applicable) and/or esterase-triggered cleavage of linkers; self-immolative release modules | NR (report with matched control and dosing window) | High peripheral distribution (erythrocytes/endothelium); competition with glucose; risk of rapid systemic cleavage; potential metabolic liabilities | Aim for productive transport without excessive binding; validate stability in plasma and nasal/intestinal matrices; monitor impact on glucose handling only where pharmacologically plausible | [380,383,384] |
| MCT1 (monocarboxylate transporter) | Monocarboxylate promoieties (e.g., lactate/pyruvate/acetate-like); ester linkers; soft-drug variants to tune logD | Carboxylesterase-mediated cleavage; pH/enzyme-sensitive linkers can bias release toward brain compartments | NR (capture as fold-change in brain exposure and unbound fraction when available) | Peripheral uptake (muscle, liver); competition with endogenous monocarboxylates; acidosis-related confounding in sensitive settings; species differences in transporter expression | Useful for polar acids/phenolics; quantify competition effects under physiological substrate levels; include brain-selective cleavage validation and metabolite profiling | [385,386,387] |
| Multiple SLCs (exploratory / case-by-case) | Nutrient-mimetic fragments matched to a selected transporter’s substrate space; modular linkers (esters/amides/carbamates) | Enzyme-labile trigger + self-immolative release (design-dependent) | NR (insert when transporter, affinity, and PK endpoints are specified) | Uncertain selectivity; off-target tissue uptake; unpredictable metabolism; model-to-human translation risk | Use only with strong transporter evidence (expression at BBB + uptake assays); pair with orthogonal confirmation (inhibitors/knockdown, saturability, competitive substrates) | [380,388,389] |
| Efflux-evading (non-transporter-hijacking) prodrugs | Mask H-bond donors/acceptors; increase logD modestly; promoieties that reduce recognition by P-gp/BCRP; soft-drug linkers | Systemic or brain esterases (must be tuned to avoid premature conversion) | NR (report brain exposure and safety vs parent) | Premature systemic activation; altered distribution and toxicity; active metabolite formation; drug–drug interactions | Useful when influx carriers are not practical; requires early efflux screening (P-gp/BCRP) and rigorous metabolite ID; prioritize Kp,uu as decision endpoint | [389,390,391] |
AUC, area under the curve; BBB, blood–brain barrier; BCRP, breast cancer resistance protein; GLUT1, glucose transporter 1; km, Michaelis constant; kp,brain, brain-to-plasma partition coefficient; kp,uu, unbound brain-to-plasma partition coefficient; LAT1, large neutral amino acid transporter 1; logD, distribution coefficient; MCT1, monocarboxylate transporter 1; nr, not reported; P-gp, P-glycoprotein; pk, pharmacokinetics; SLCs, solute carrier transporters; vmax, maximum transport rate.
Table 7.
Models-to-endpoints crosswalk for blood-brain barrier (BBB) delivery: throughput, predictive scope, artifacts, and best-fit development decisions. This crosswalk links commonly used BBB/central nervous system (CNS) delivery models discussed in the manuscript to the endpoints they most reliably support. For each model, throughput is contrasted with predictive scope (barrier integrity, transporter effects, regional delivery, or human translation), and key artifacts that bias interpretation are flagged. The Best-fit decision column indicates where each approach is most informative in a development workflow—from early screening to go/no-go based on decision-grade brain exposure (e.g., Kp,uu) and target engagement.
Table 7.
Models-to-endpoints crosswalk for blood-brain barrier (BBB) delivery: throughput, predictive scope, artifacts, and best-fit development decisions. This crosswalk links commonly used BBB/central nervous system (CNS) delivery models discussed in the manuscript to the endpoints they most reliably support. For each model, throughput is contrasted with predictive scope (barrier integrity, transporter effects, regional delivery, or human translation), and key artifacts that bias interpretation are flagged. The Best-fit decision column indicates where each approach is most informative in a development workflow—from early screening to go/no-go based on decision-grade brain exposure (e.g., Kp,uu) and target engagement.
| Model | Throughput | What it predicts | Key artifacts | Best-fit decision | References |
|---|---|---|---|---|---|
| Transwell BBB (endothelial mono-/co-culture; static TEER/permeability) | High | Relative permeability and gross barrier integrity; early ranking of formulations/prodrugs; qualitative efflux effects (context-dependent) | Static conditions; nonphysiologic shear; variable tight junction maturation; transporter expression drift; adsorption to plastics | Early screen and rank-order; eliminate non-starters before costly models | [473,474,475] |
| iPSC-derived BBB endothelium (Transwell) | Medium | More human-relevant tight junctions/transporters; better prediction of human-like permeability windows | Differentiation variability; batch effects; incomplete neurovascular unit (NVU) signaling unless co-cultured | Mid-stage confirmation of BBB-relevant transport and efflux liability | [476,477,478] |
| iPSC BBB organoids / spheroids (NVU-like) | Medium–low | 3D cell–cell interactions, uptake and penetration trends; neuroinflammation-compatible testing | Size heterogeneity; diffusion limits; limited perfusion; measurement standardization gaps | Mechanism prioritization and safety/uptake profiling; compare targeting vs non-targeting designs | [479,480,481] |
| Microfluidic BBB-on-chip (flow/shear; NVU co-culture) | Low–medium | Dynamic barrier responses under flow; transporter-mediated flux; inflammation-dependent permeability shifts | Device-to-device variability; bubble/absorption effects; complex operation; limited throughput | Late preclinical de-risking for mechanism and context dependence (inflammation, disease cues) | [482,483,484] |
| Rodent in vivo PK (brain + plasma; brain/plasma ratios) | Medium | System-level exposure, metabolism, distribution; initial signal of CNS delivery improvement | Species differences in BBB properties and transporters; confounding by vascular space and binding; anesthesia effects | Go/no-go based on integrated exposure; prioritize candidates for quantitative endpoints (Kp,uu) | [473,477,485] |
| Rodent microdialysis (ISF sampling) | Low | Unbound interstitial exposure and time-course; closest preclinical readout to target-site pharmacology | Invasive; recovery calibration; regional restriction; limited to specialized setups | Decision-grade confirmation of CNS penetration (Kp,uu-like inference) and PK/PD linkage | [473,477,485] |
| CSF sampling (preclinical/clinical) | Medium | Surrogate exposure trends when ISF is unavailable; supports translational sampling designs | CSF ≠ ISF; compartmental delays; protein binding differences; disease-state confounding | Clinical feasibility planning; supportive evidence alongside imaging or modeling | [473,477,485] |
| PET imaging (labeled payload or marker) | Low | Whole-brain/spatial distribution; target engagement surrogates; longitudinal kinetics in vivo | Radiolabel alters properties; metabolite signal; resolution limits; tracer-specific assumptions | Translation-facing biodistribution and engagement readouts; de-risk regional delivery claims | [478,486,487] |
| Non-human primate (NHP) studies | Very low | Closest approximation to human BBB transport and PK; de-risks scale and delivery paradigm | Cost/ethics; small n; limited disease modeling; procedural constraints | Preclinical-to-clinical bridge for top candidates and delivery devices/targeting ligands | [473,477,485] |
| Mechanistic PBPK / BBB models (incl. efflux and binding) | High (in silico) | Scenario testing; dose-to-exposure translation; integrates binding, efflux, and tissue partitioning | Parameter uncertainty; requires high-quality input data; model misspecification risk | Study design, endpoint selection, and translation planning; interpret CSF/ISF and imaging outputs | [473,477,485] |
BBB, blood–brain barrier; CNS, central nervous system; CSF, cerebrospinal fluid; ISF, interstitial fluid; iPSC, induced pluripotent stem cell; Kp,uu, unbound brain-to-plasma partition coefficient; NHP, non-human primate; NVU, neurovascular unit; PBPK, physiologically based pharmacokinetic; PD, pharmacodynamics; PET, positron emission tomography; PK, pharmacokinetics; TEER, transendothelial electrical resistance.
Table 8.
Clinical snapshot of central nervous system (CNS) delivery strategies in the manuscript: indication-by-modality overview, endpoints, and exposure evidence. This table provides a high-level clinical snapshot of the delivery modalities discussed in the manuscript, organized by representative CNS indications. For each entry, the development phase and primary clinical endpoint are summarized alongside the type of exposure evidence available (e.g., imaging-verified BBB opening, CSF/PK surrogates, or decision-grade brain exposure metrics when reported). “NR” denotes details not specified in the current draft and can be completed during final reference curation.
Table 8.
Clinical snapshot of central nervous system (CNS) delivery strategies in the manuscript: indication-by-modality overview, endpoints, and exposure evidence. This table provides a high-level clinical snapshot of the delivery modalities discussed in the manuscript, organized by representative CNS indications. For each entry, the development phase and primary clinical endpoint are summarized alongside the type of exposure evidence available (e.g., imaging-verified BBB opening, CSF/PK surrogates, or decision-grade brain exposure metrics when reported). “NR” denotes details not specified in the current draft and can be completed during final reference curation.
| Indication | Modality | Phase | Primary endpoint | Exposure evidence | Status | References |
|---|---|---|---|---|---|---|
| Neurodegeneration (Alzheimer’s / Parkinson’s) | Focused ultrasound (FUS) + microbubbles | Phase I (as noted in manuscript) | Safety/tolerability; imaging-confirmed BBB opening | MRI guidance + contrast-enhanced confirmation; BBB resealing within hours (NR details) | Early clinical translation under way | [296,525,526] |
| Glioblastoma / focal CNS tumors | RMT-targeted nanocarriers (e.g., TfR/LRP1 ligands; Angiopep-2-type designs) | Preclinical → early clinical (NR) | Tumor response / progression metrics (NR) | Biodistribution/brain uptake signals; comparator vs non-targeted carrier (NR) | Investigational; target/format-dependent | [215,527,528] |
| Glioblastoma / focal CNS tumors | Convection-enhanced delivery (CED) | Clinical use/trials (NR) | Local control and safety (procedure-specific) | High local concentration by direct interstitial infusion; distribution tracking (NR) | Procedure-dependent; used in specialized settings | [324,325,529] |
| Depression / neuropsychiatric disorders | Intranasal nose-to-brain formulations (sprays, gels, nanoemulsions) | Preclinical → early clinical signals (NR) | Symptom scales and tolerability (NR) | PK/PD signals; CSF or surrogate exposure where available (NR) | Emerging; high variability and formulation-sensitive | [329,530,531] |
| Broad CNS indications (adjunct permeability strategies) | Osmotic BBB disruption (intra-arterial mannitol) | Selective clinical application (NR) | Feasibility/safety; delivery enhancement (NR) | Increased permeability by protocol; exposure quantification variable (NR) | Invasive; limited use due to safety/control trade-offs | [321,532,533] |
| Exploratory / device-enabled targeting | External-field targeting/triggering (magnetic guidance; remote release) | Preclinical | Proof-of-concept delivery and safety | Tracking-enabled carriers; biodistribution and clearance studies (NR) | Exploratory; hardware and long-term safety gaps | [296,534,535] |
BBB, blood–brain barrier; CED, convection-enhanced delivery; CNS, central nervous system; CSF, cerebrospinal fluid; FUS, focused ultrasound; LRP1, low-density lipoprotein receptor–related protein 1; MRI, magnetic resonance imaging; NR, not reported; PD, pharmacodynamics; PK, pharmacokinetics; RMT, receptor-mediated transcytosis; TfR, transferrin receptor.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



