
Submitted:
28 January 2026
Posted:
29 January 2026
You are already at the latest version
Abstract
3D cell systems for in vitro experimental studies are able to mimic the in vivo efficacy of drugs, before testing on animals, however many studies are still needed for mimicking the physiological environment with 3D cell growth systems. The mechano-physical properties of the microenvironment are relevant for the invasiveness of cancer cells and also for their drug resistance. 3D tumoroids of human oral squamous cell carcinoma (OSCC) CAL27 cells of different stiffness are here produced using a tunable PEG-silk fibroin hydrogel (PSF) and the anti-tumor activity of the CIGB300 peptide, an anticancer therapeutic peptide, on these 3D tumoroids models is assessed. Moreover, spectroscopical studies on CIGB300 peptide are here also reported for analyzing its structure, stability, aggregation and diffusion properties. For the first time, the CIGB300 peptide diffusion in tunable silk fibroin hydrogels of different stiffness is investigated over-time by fluorescence spectroscopy as potential tool in drug-screening using hydrogel-based 3D tumoroids.
Keywords:
anticancer peptides
; spheroids
; hydrogels
; stiffness
; CAL27
; 3D cell culture systems
1. Introduction
The 3D cell growth systems give the possibility to greatly expand knowledge on the effects of the microenvironment on the invasiveness of cancer cells and their response to mechano-physical stimuli and drugs [1,2,3,4,5]. Recently, a valid alternative to simple spheroids obtainable through commercial kits, in which the cell density and rheology of the spheroid are not tunable, is represented by the use of cellular microspheres systems produced by photopolymerizable semi-synthetic hydrogels [6,7,8,9]. This strategy allows to varying the chemical and mechano-physical properties of the system, thus to assess different conditions and effects on proliferation, cell invasiveness, as well as on the expression of genes of metabolic pathways relevant to tumorigenicity and drug resistance [10,11,12,13]. The effects of the stiffness on the drug response of cancer cells were here investigated on 3D PEG-silk-fibroin (PSF) hydrogel-based tumoroids and using an antitumor peptide, named CIGB300. This is a cyclic peptide (GRKKRRQRRRPPQ-β-alaCWMSPRHLGTC, see Figure 1A) characterized by a disulphide bond between the two cysteine residues, and by the presence of a cell-penetrating peptide sequence derived from the HIV-Tat protein at the N-terminus. This compound was obtained by the Center of Genetic Engineering and Biotechnology of Havana (Cuba) screening a random cyclic peptide phage library against the HPV-16 E7 oncoprotein site targeted by CK2 for phosphorylation [14,15,16]. The anticancer activity of CIGB300 is linked to the ability of this peptide to inhibit the casein kinase 2 (CK2) and it has been investigated since the peptide was first developed [15,16]. Several studies have demonstrated its ability to modulate critical hallmarks of tumor biology [17,18,19,20,21]. In lung cancer line NCI-H82, CIGB300 was shown to interact with B23/nucleophosmin 1 (B23/NPM1) in the nucleolus, thereby disrupting its CK2-mediated phosphorylation and triggering rapid apoptotic cell death, a process apparently preceded by nucleolar disassembly [22,23]. Consistently, CIGB300 impaired CK2-dependent phosphorylation of NPM1, leading to cell cycle arrest and apoptosis. Moreover, CIGB300 induced a rapid, dose-dependent apoptosis in cancer cells and displayed a broad antiproliferative activity within the micromolar range (20–300 μM) across lung, cervical, prostate, and colon cancer-derived cell lines [15,16,24,25]. Moreover, intravenous and intraperitoneal administration of CIGB300 significantly reduced primary tumor growth in heterotopic models of cervical and lung cancer [24]. Consistent with these findings, CIGB300 exhibited pronounced anti-angiogenic and pro-apoptotic effects both in vitro and in vivo [26]. Importantly, these studies have provided the basis for the initiation of clinical trials in patients with cervical cancer [15,16,24,27]. Following the completion of multiple phase I clinical trials, which established the safety and tolerability of intra-tumoral administration, a phase II clinical trial is currently underway to evaluate the efficacy of CIGB300 in patients with stage IB2/II cervical cancer [28] CIGB300 markedly reduced the adhesion, migration, and invasion capacities of human H125 and murine 3LL lung cancer cells. In vivo, studies employing the syngeneic 3LL lung cancer model in C57BL/6 mice demonstrated robust anticancer activity, including inhibition of lung colonization, metastasis, and tumor-induced neovascularization [24]. Further analyses revealed that CIGB300 impaired migration, invasion, and tumor cell–secreted protease activity. Mechanistically, CK2 inhibition by CIGB300 was associated with reduced motility and invasiveness of lung cancer cells, concomitant with suppression of the proteolytic activity of tumor cell-secreted urokinase plasminogen activator (uPA) and matrix metalloproteinase2 (MMP2) [24,29]. In this study the anti-cancer effects of CIGB300 on the proliferation and less invasive oral squamous cell carcinoma (OSCC) cell line CAL27 were here investigated in 2D and 3D (CALPSF) cell culture systems. The effects of the different stiffness of the 3D-CALPSF system on the B23/NPM1 expression and drug resistance were assessed, as well as the effects of CIGB300 treatment on the cancer cell viability. Moreover, the aggregation and diffusion properties of CIGB300 in the silk fibroin (SF) hydrogels were investigated by molecular spectroscopy.
2. Results and Discussion
2.1. Molecular Characterization of CIGB300 Peptide Aggregation by UV-Vis Absorption, and Fluorescence Spectroscopy and Circular Dichroism
The UV-vis spectra of CIGB300 recorded at different concentrations, ranging from 48 µM to 326 µM, are shown in Figure 1A. The absorbance at 280 nm increased proportionally to the peptide concentration, as shown in Figure 1Ab by linear regression, an ε280 molar extinction coefficient of 4468 (S.D.± 284) M-1 cm-1 was obtained. In water solution the peptide showed a circular dichroic spectrum characteristic of cyclic peptides [30,31,32], with a minimum of molar ellipticity at 200 nm, characteristic by deconvolution of a beta strand with turn structure (Figure 1B). Deconvolution of the CD curve by the BestSel code [33] provided a 30% of antiparallel β-sheet, 17% β-turn and 53% of random conformations.
The fluorescence emission spectrum of CIGB300, obtained by excitation at λex=280 nm, showed a maximum intensity at λem=350 nm, characteristic of the tryptophan residue in the sequence. Also in this case the fluorescence emission intensity increased proportionally to the peptide concentration, suggesting that peptide aggregation was negligible in the range of used concentrations.
2.2. CIGB300 Affects the Cell Viability and Migration of CAL 27 Cancer Cell Line in 2D Cultures
Firstly, the effects of CIGB300 peptide on the cell viability of human head and neck squamous carcinoma adherent cells of the CAL 27 cancer cell line were assessed using 2D cell cultures. In the Figure 2A is shown the cell viability of CAL 27 cells after 24h of treatment with 0, 80, 160, 240 and 320 µM of CIGB300. A decrease in cell viability of CAL 27 cancer cells in a concentration dependent manner was observed, which was significant at CIGB300 concentrations higher than 80 µM in the cell culture medium. The optical micrographs of the cells after 24h of cell growth in the absence and in the presence of 160 µM of CIGB300 are shown in the Figure 2B. Moreover, the peptide was also able to reduce the cell migration/invasiveness in concentration dependent manner, as shown in the Figure 2C. A significant reduction of the migration was observable also after treatment with 80 µM of CIGB300 by scratch assay, that was not due to an inhibition of the cell viability.
2.3. CAL 27 Tumoroids Production
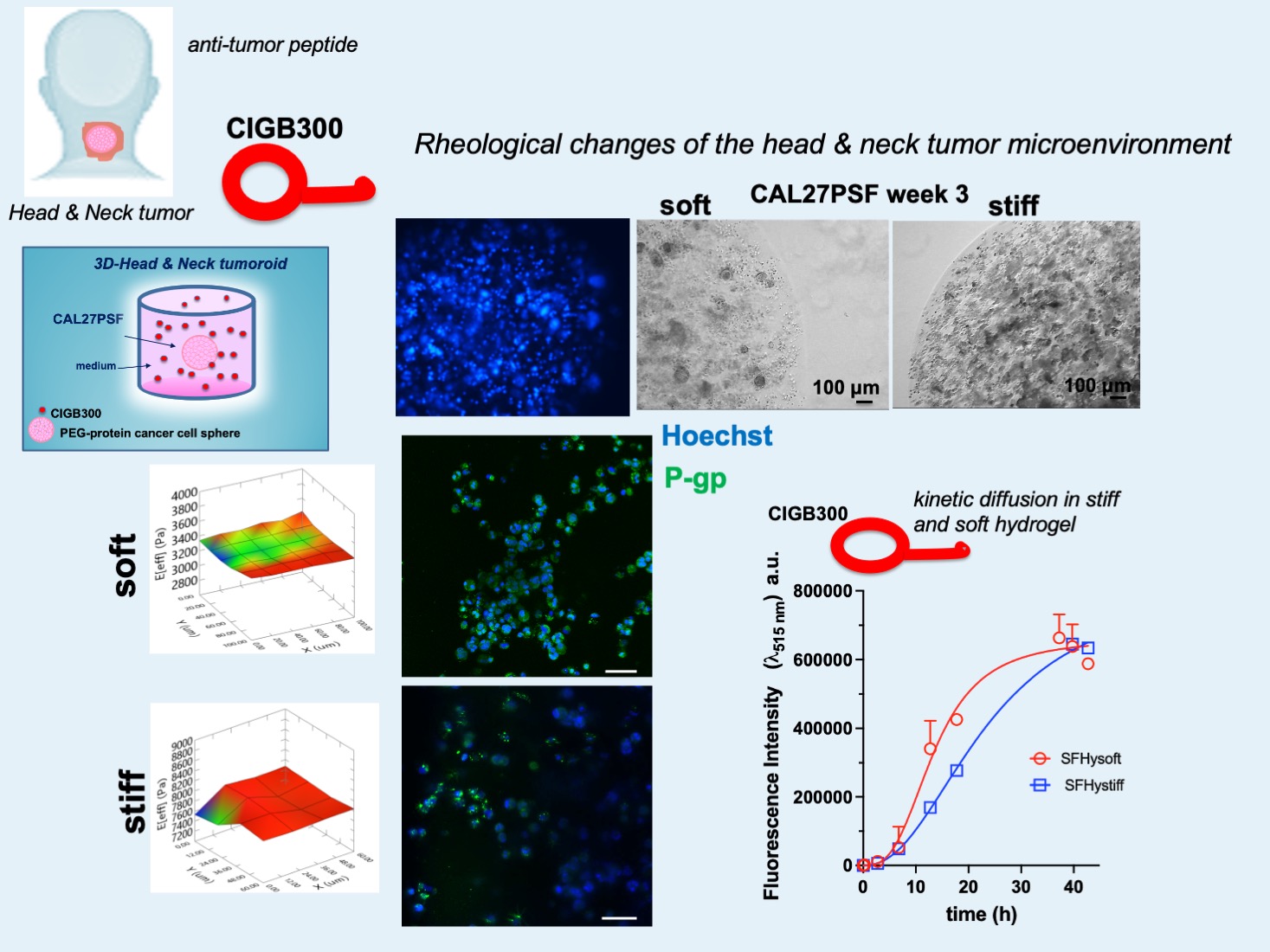
PEG-silk fibroin hydrogel (PSF) was obtained according to previous studies [4,34]. The viscoelastic properties of the PSFs were previously assessed using oscillatory shear rheometric analysis [4]. PSFs with 4.5% (PSFHysoft) and 9% (PSFHystiff) (w/v) of PEGDa (10kDa), without embedded cells were characterized by rheological nanoindentation methodology showing shear storage moduli (Eeff, Effective Young moduli) of 3.37 (± 0.03) kPa and 7.81 (± 0.01) kPa, respectively (Figure 3B). The higher stiffness was comparable to that one of in vivo tumor tissue of invasive carcinoma that was observed to be 6-10 kPa [35]. Spheres of CAL 27 cancers cells in PSF (CAL27PSF) were produced using photopolymerization with UVA of drops of 3 or 5 µl of PSF precursor solution on the superhydrophobic nano-structured polydimethylsiloxane (PDMS) surface (Figure 3A).
The micrographs of the CAL 27 spheroids (CAL27PSF) are showed in Figure 3C. After 1 week of cell growth CAL27PSFs were stained with Hoechst 33342 in living conditions and the fluorescence of the nuclei was observed after fixing by confocal fluorescence microscopy (see Figure 3D). A good viability was also observed using WST-1 metabolic assay after 24h of cell culture of CAL27PSFstiff (Figure 3E) and after three days of cell culture of CAL27PSFsoft and CAL27PSFstiff by LIVE/DEAD fluorescence assay (Figure 4A). The fluorescence micrographs of the CAL27PSF in soft and stiff systems obtained by Hoechst staining under living conditions after 4 days of culture are shown in the Figure 4B. No significant differences between the two systems were observed.
However, after 3 weeks of cell culture in the CAL27PSFsoft was visible the presence of cell clusters that were not present in the CAL27PSFstiff (Figure 4C). This finding indicates that the higher stiffness of the microenvironment affects the ability of the cells to form carcinogenic cell clusters. The cyclin D1 expression was evaluated in CAL27PSFstiff produced using concentrations of 9% w/v PEGDa (Figure 4D). No significant changes in cyclin D1 expression were observed over time, indicating a good invasion and proliferation of the cancer cells in this system. Moreover, the acetylation of the histone H3 in 3D at high stiffness was also similar to the cell grown on TCP and over time. This result is in agreement with the existing link between cyclin D1 and the histone acetylation [36].
2.4. Effects of CIGB300 Peptide on the Cell Viability of CAL27PSF Tumoroids in Dependence on the Stiffness
The cell viability of the CAL27PSFsoft and CAL27PSFstiff treated at different concentrations of CIGB300 was assessed by LIVE/DEAD (L/D) assay (see Figure 5B).
After 3 days of treatment the cell viability L/D assay showed an increase of the cell death depending on the CIGB300 concentration in the cell medium either in the CAL27PSFsoft or in the CAL27PSFstiff. However, a major effect was observed in the CAL27PSFstiff tumoroids. The cell viability of the tumoroids in the presence of CIGB300 in the cell culture medium was also evaluated by metabolic assay (Figure 5C) observing a decrease of the 24.2% (S.D. ± 6.59) of the cell viability of CAL27PSFstiff tumoroid after 72h of treatment with 80 µM of CIGB300.
2.5. Diffusion Studies of the CIGB300 in SF Hydrogels by Fluorescence Spectroscopy
The tunable rheological cellular models allow to mimic the complexity of natural architecture and the physiological drug administration. Diffusion of molecules in the hydrogel-based tumoroid has a crucial role in drug administration. In general, the majority of drug permeation mechanism is mediated by diffusion kinetics and is mainly considered to be Fickian [37]. The CIGB300 peptide diffusion in the hydrogels of different stiffness was here evaluated by fluorescence spectroscopy. Soft and stiff silk fibroin (SF) hydrogels were produced, using 4.5 and 9% of PEGDA respectively, and their Young’s modulus was evaluated by nanoindentation, as shown in the Figure 6A, resulting in 2.29 (± 0.05) kPa and 7.57 (± 0.09) kPa, respectively.
The peptide was labelled with FITC and 100 µL of 3 mM of FITC-CIGB300 was added on the top of the SF hydrogel (900 µl) previously polymerized directly in the cuvette. The variation of the fluorescence emission intensity at λem= 515 nm was monitored over time. Fluorescence emission of the hydrogels at λem=515 nm was performed before the addition of the peptide solution on the top and subtracted as background signal. The diffusion of the FITC molecule in the stiffer SF hydrogel over time is shown in Figure 6 B. The diffusion kinetics of FITC was very fast, showing a hyperbolic trend. In 1 hour, the fluorescence emission intensity of FITC was at its maximum and remained almost constant over the time demonstrating that small molecules like FITC are able to rapidly diffuse in the SF hydrogels and interact with the cells of PSF spheroids. By contrast the diffusion rate of FITC-CIGB300 was slow and dependent on the SF hydrogel stiffness (Figure 6C and S1). The profiles of the diffusion kinetics of FITC-CIGB300 were sigmoidal for both the hydrogels, indicating the presence of different steps in the diffusion process, and the Hill coefficients of FITC-CIGB300 were 2.94 and 2.24 in SFHy soft and stiff, respectively. The calculated diffusion rates were approximately 11.5 and 6.35 μM/hour in SFHy soft and stiff, respectively. This behavior is most likely related to the polymeric phase of the hydrogel and to its swelling [38,39], and to the interaction of the peptide with the protein component of the hydrogel [40,41,42].
2.6. The Tumoroid Stiffness Affects the Drug Resistance of the CAL 27 Cancer Cells
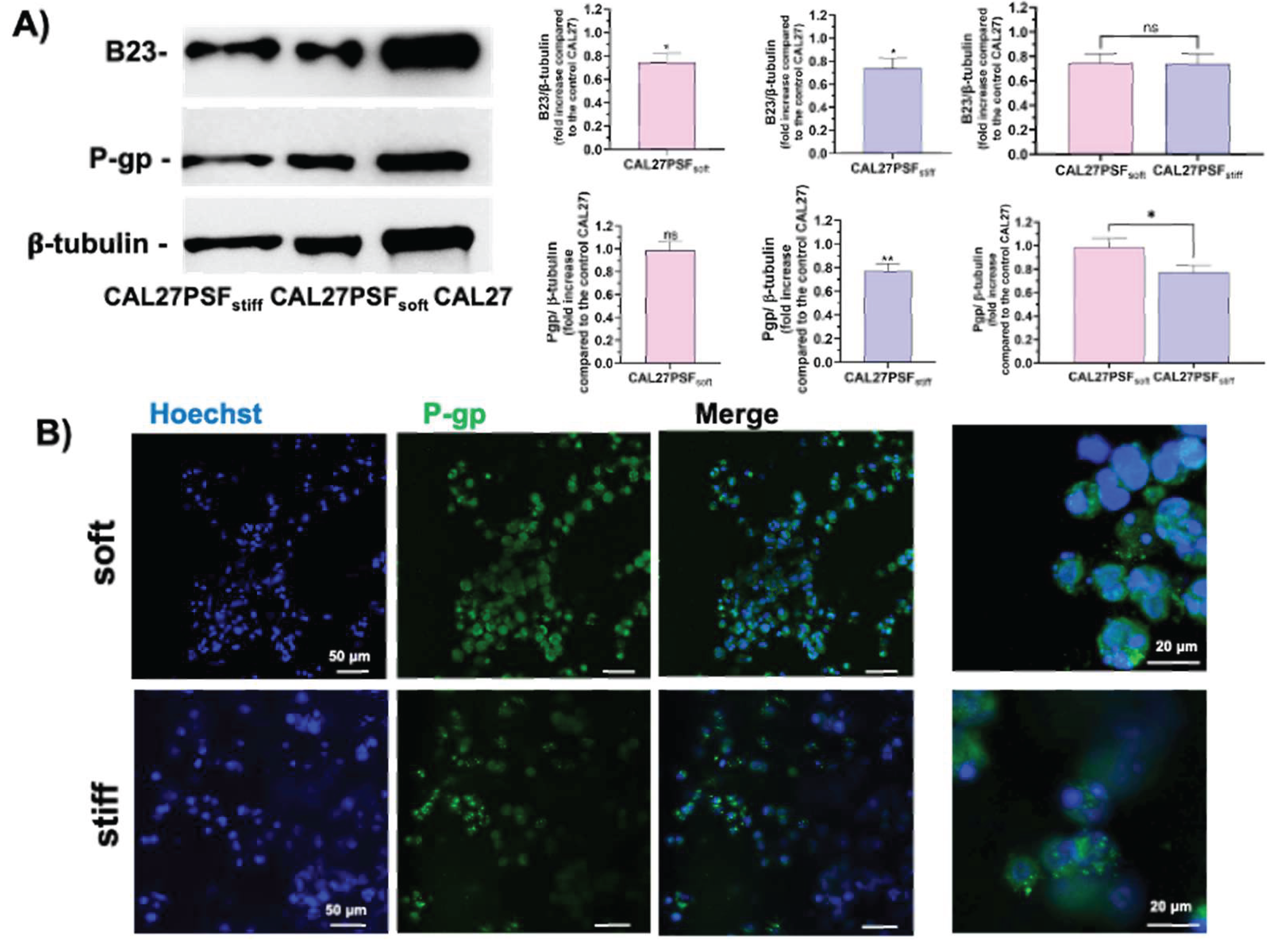
Considering the faster diffusion kinetic of CIGB300 into the soft SF hydrogel, the effects of the major cell death observed in the tumoroids at higher stiffness could be due to an increased sensitivity to the antitumor peptide of the cells grown in the stiffer conditions. Therefore, the antitumor property of the CIGB300 peptide can be increased by the physical rheological factor in 3D head and neck tumoroids models in a synergistic way. As described above the antitumor effects of CIGB300 are related to the down-regulation of the B23/NPM1, which is an important tumor marker [15,22,43], by physical interaction and inhibition of the activation by phosphorylation at Ser 125 from the casein-kinase 2 (CK2) in the tumour cells [44,45,46,47]. Several studies have shown that B23/NPM1 is over-expressed in solid tumours with different histological origin, including: pancreas [48], prostate [49], liver [50], glioma and glioblastoma [51,52], astrocytoma [53], thyroid [54], gastric [55], colon [56] and others. Further, the NPM1 has been proposed as an adverse prognostic marker in a number of such malignancies [57,58] due to its overexpression often correlated with high mitotic and metastatization index. On these bases, the B23/NPM1 protein expression in different rheological conditions was here assessed. In the Figure 7A are shown the western blot analyses for evaluating the expression of B23/NPM1 in CAL27 cultured in both 2D and 3D conditions and at different stiffness (CAL27PSFsoft and CAL27PSFstiff). A significant decrease of the B23/NPM1 expression was observed in 3D conditions compared to the 2D culture, indicating that the B23/NPM1 expression change in response to the microenvironment of the cell culture system. However, no significant differences in the B23/NPM1 expression were observed for the two stiffness (CAL27PSFsoft and CAL27PSFstiff). Therefore, the B23/NPM1 expression in the 3D- CAL27 cell culture system does not change in the 3.4- 7.8 kPa stiffness range. Moreover, also the P-gp expression in both soft and stiff tumoroids was here evaluated. P-glycoprotein (P-gp) is an ABC transporter associated with resistance to cancer chemotherapy [59]. In the Figure 6 are shown the representative confocal fluorescence micrographs of the CALPSFsoft and CAL27PSFstiff, after 3 days of cell culture. A major expression of the P-gp expression was observable in CAL27PSFsoft with respect to CAL27PSFstiff in the confocal fluorescence analyses and it was also confirmed by western blotting analysis (Figure 7B), where a statistically significant difference of the P-gp expression at the two different stiffness was observed.
Although it is not known if the P-gp protein is implied in the transport of the CIGB300 out the cell, some studies indicate that the cancer resistance to some antitumor cyclic peptides, such as FK228 and apicidin, are related to the P-gp expression [60]. These preliminary data could open the way to the study of the correlation of the activity of this cyclic antitumor-peptide and the drug resistance. Moreover, the P-gp function is relevant in the endogenous detoxification system from metabolites therefore its down-regulation could be extremely important for sensibilizing the cell to the drugs. The P-gp expression has been also associated to the B23/NPM1 expression [61], the knockdown of B23/NPM1 reduces the drug-resistance by down-regulation of P-gp. In agreement, we also observed a decrease in the P-gp expression associated with a B23/NPM1 down-regulation in stiff conditions in respect to cells grown on TCP.
These results indicate relevant effects of the stiffness on the drug resistance in head and neck tumoroids, therefore, the increase in the sensitivity of the tumor cells to the CIGB300 peptide treatment may be due to a synergistic effect with the physical rheological factor that induces a change of the cancer cells.
3. Materials and Methods
3.1. Molecular Studies
3.1.1. Fluorescence Spectroscopy
Steady-state fluorescence spectra were measured through a spectrofluorimeter “Fluoromax-4” (Horiba Jobon Yvon), in a thermostated (25 °C) quartz cell (1 cm). The fluorescence emission spectra of W residues in 163 μM CIGB300 aqueous solution (λex= 280 nm, λem= 290-450 nm ) have been measured with both excitation and emission slits set at 2 nm, integrating the signal for 0.6 s at each nanometer. FITC and FITC-CIGB300 fluorescence intensities were measured using λex= 495 nm, and λem= 515 nm , with excitation and emission slits set at 3 and 5 nm, respectively, integrating the signal for 0.4 s at each nanometer. The data were fitted using the following sigmoidal curve equation:
ΔF =ΔFmax*tS/(tΔFmax/2+t)S (1)
where ΔF was (F1-F0), ΔFmax was (Fmax-F0), t was the time, tΔFmax/2 was time where the ΔF was half of the ΔFmax and S was the sigmoid Hill coefficient.
3.1.2. Circular Dichroism of CIGB300 Peptide
Far-UV CD measurements were performed using a Jasco 1500 spectropolarimeter (Jasco, Tokyo, Japan) calibrated with camphorsulfonic acid. CD spectra were recorded between 190 and 260 nm using 0.1 cm path length, 1.0 s time constant, 2 nm band width and 0.2 nm/ min of scan rate and 20 mdeg of sensitivity. The CD measurements were performed on 67 μM CIGB300 in aqueous solutions using a quartz cell 0.1 cm optical length thermostated at 25 °C.
3.2. Rheological Analyses of the Hydrogels
The microrheological analyses of SF and PSF hydrogels at high and low stiffness (without cells) were performed by nanoindentation using PIUMA nano-indenter equipment (Optics11 Life, Amsterdam, The Netherlands). SFHy and PSFHy samples were produced as follows. SF was purified from Bombix mori cocoon and lyophilized through a published protocol and PSF was obtained by PEGylation of SF as previously described [62]. The hydrogels were prepared by photopolymerization (5 min, 365 nm, 5 mW/cm2) of 100 μl of the gel precursor solutions (3 mg/ml of SF or 70 µl of 6 mg/ml PSF in PBS buffer, 30 µl (stiff) or 15 µl (soft) of 30% w/v PEGDa 10kDa, 1% w/v of Irgacure 2959 for a total volume of 100 µl in PBS) using o-rings as molds. The samples were then analyzed in liquid environment using PBS buffer. A probe having a 0.45 N/m cantilever stiffness and a 49.5 µm radius spherical tip was selected for the measurements. The effective Young’s Modulus (Eeff, Pa) was measured in peak load poking mode performing a matrix scan of 6x6 and 4x4 with a displacement on X and Y axes of 20 μm (max load 0.6-1.40 µN and piezo speed 30.0 µm/s). Data analysis was performed setting an Hertzian contact model and obtaining a 3D plot of Eeff values across the matrix. One of the registered Load-Indentation curves and the matrix scan are reported for each sample analyzed.
3.3. Cellular Studies
3.3.1. Cell Culture Systems Production and Cell Viability Assay
Cell studies were performed using the human head and neck squamous cell carcinoma (HNSCC), adherent cells of the CAL27 cell line (Istituto Zooprofilattico, Brescia, Italy). Cell cultures were grown in Dulbecco’s Modified Eagle Medium (DMEM) (Gibco, Italy), containing 10% v/v Fetal Bovine Serum (FBS) (Gibco, Italy), 1% w/v penicillin-streptomycin (Sigma-Aldrich, Italy), 1% w/v of a 200 mM L-Glutamine solution (Gibco, Italy) and 1% v/v non-essential amino acids solution (Sigma-Aldrich, Italy). CAL27PSF hydrogel spheres were obtained using the PSF precursor solution (70 µl of 6 mg/ml PSF in PBS buffer, 30 µl (stiff) or 15 µl (soft) of 30% w/v PEGDa 10kDa, 1% w/v of Irgacure 2959 for a total volume of 100 µl in PBS). A nanostructured superhydrophobic surface of polydimethylsiloxane (PDMS) was produced using the protocol described by Patent LDO0252 [4,5,63]. CAL27 cells at a density of 104 cells/µl were resuspended in the gel precursor solution, dropped (drops of 3-5 µl) on the superhydrophobic surface and photopolymerized by exposure under UV light (365 nm, 5 mW/cm2) for 2 minutes. 3D cell systems were cultured and monitored over time by brightfield microscopy using a Zeiss microscope (Primovert, Zeiss, Italy). CIGB300 peptide was diluted in Dulbecco’s Modified Eagle Medium (DMEM) without FBS at 100 mM, aliquoted and stored at -20 °C, and used for the treatments at the different concentrations.
Cell viability and proliferation after each cell treatment with CIGB300 were tested by WST-1(4-[3-(4-lodophenyl)-2-(4-nitrophenyl)-2H-5-tetrazolium]-1,3-benzene disulfonate (Cell Proliferation Reagent WST-1, Roche, Mannheim, Germany) assay [64]. CAL27 cells were seeded at 1.6x104 cells/cm2 in a 96w. multiwell and the day after treated for 24 hours with CIGB300 (80, 160, 240 and 320 μM). 3D CAL27PSF spheroids were produced as described above and, the day after their production, were treated with CIGB300 (80, 160, 240 and 320 μM) for 72 hours. After each treatment, the medium was replaced with fresh DMEM high glucose without phenol-red (GIBCO, Italy) containing tetrazolium salt WST-1 (5% v/v). 2D-seeded cells and 3D cell samples were then incubated for 2h and 4 h respectively at 37 °C, 5% CO2. Absorbance of the medium was recorded at 450 nm using a microplate reader (iMark™ Microplate Absorbance Reader, Bio-Rad). The cell viability of the CAL27PSF systems was also monitored by fluorescence microscopy using a LIVE/DEAD® Cell Imaging Kit (L/D assay) (488/570) (Molecular Probes, Life Technologies, Thermo Fisher Scientific, Milan, Italy) and using a Zeiss fluorescence microscope (Zeiss, Axio Observer 7).
3.3.2. Immunofluorescence Microscopy Analyses
Fluorescence microscopy of CAL27PSFs was performed by nuclei staining of live cells with Hoechst 33342 (Sigma-Aldrich, Italy) for 4 hours; then, the spheres were washed with PBS and fixed with 4% paraformaldehyde (PFA) for 30 min at room temperature and the fluorescence was observed using a Zeiss fluorescence microscope (Zeiss, Axio Observer 7). Immunofluorescence analysis of CAL27PSF spheres was performed as follows. After staining the nuclei of live cells with Hoechst 33342 (Sigma-Aldrich, Italy), the spheres were then fixed with 4% PFA in PBS for 30 min at room temperature, permeabilized with 0.3% Triton X-100 for 15 min and maintained in a blocking buffer (10% of BSA (w/v), 0.1% of Triton X- 100 (v/v) and 1% of glycine (w/v) in PBS) overnight at 4 °C. CAL27PSF were then incubated overnight with 1% BSA and 20 mM Gly solution in PBS containing the primary antibody for P-gp (rabbit) (Abcam, Cambridge, UK) followed by the appropriate Alexa fluorochrome-conjugated secondary antibody (488 nm, green) (Thermo Fisher Scientific, Invitrogen, USA). Confocal microscopy was performed using a Stellaris Leica microscope platform.
3.3.3. Scratch Wound Healing Assay
CAL27 cells were seeded into a 24-well plate (6.5 x 104 cells/cm2) and cultured at 37 °C 5% CO2 so that the cells would reach confluency the next day. A scratch-wound was created and the area of the scratch-wound at time 0 and after 24h was measured. Percentage of wound closure was measured as follows:
Wound closure(%)= (Wound surface area after 24h/Wound surface area at time 0)x 100 (2)
3.3.4. Protein Expression by Western Blot Analysis
Proteins were extracted by lysis of CAL27 cells and CALPSF using 100 μl of sample buffer for SDS-PAGE. Samples were vortexed and boiled for 5 min, centrifuged for 5 minutes at 10000 rpm and stored at -20 °C. The cell extracts were electrophoresed on 12 or 15% polyacrylamide gel and electro-blotted on PVDF membrane (Sigma-Aldrich, Italy). The membranes were then blocked and probed with primary monoclonal antibodies: Ab-Acetyl-Histone H3 [Ac-Lys9] (rabbit) (Sigma-Aldrich, Italy), Ab-CyclinD1 (rabbit) (Cell Signaling Technology, USA), Ab-B23/NPM1 (rabbit) (Merck, Sigma Aldrich, Italy) or Ab-P-gp (rabbit) (Abcam, Cambridge, UK) overnight at 4 °C. Immunoblots were next incubated with the secondary antibodies (dilution 1:3000) (Cell Signaling Technology, USA) for 4 hours at room temperature. Immunoblot with Ab-β-tubulin mouse (Sigma-Aldrich, Milan Italy) were also probed for controlling the protein loading. Immunoblots were probed with a Super Signal West Pico kit (Thermo Scientific, USA) to visualize signal, followed by exposure to a Fluorchem Imaging system (Alpha Innotech Corporation-Analitica De Mori, Milan, Italy). The quantitative evaluation of protein expression of CAL27 tumoroids was obtained by 3-5 microspheres for each sample and is therefore also inherently mediated.
3.3.5. Statistical Analysis
The statistical analysis was performed using GraphPad Prism version 8.0 for Windows (GraphPad Software, San Diego, CA, USA). Data from three to six independent experiments were quantified and analyzed for each variable using a one-tailed Student’s t-test or One-way ANOVA test. A p value of < 0.05 was considered to be statistically significant. Standard deviations or standard error means were calculated and presented for each experiment.
4. Conclusions
The head-neck tumoroids were here produced and characterized for in vitro studies of the antitumor activity of the peptide CIGB300, taking into account the mechanical properties of the cellular microenvironment. A spectroscopical characterization of CIGB300 was performed observing absence of peptide aggregation in the concentration range used for the cell treatment. An inhibition of the cell viability and migration of oral squamous cell carcinoma cell line CAL27 were observed on 2D cell culture system in agreement with previous studies on different cancer cell lines. The antitumor effects were here also validated using 3D CAL27 culture systems. Tunable CAL27PSF tumoroids were here produced and characterized and the anti-tumor activity of the CIGB300 was also assessed in relation to the mechanical properties changing of the cellular microenvironment. The major anti-tumor effect was observed using a stiffer microenvironment. The diffusion over time of CIGB300 peptide was investigated on silk fibroin hydrogels at the same Young’s moduli by fluorescence spectroscopy. A cooperative diffusion behavior of the peptide for both the hydrogels at different stiffness was observed, in agreement to previous studies on the diffusion of molecules in the hydrogels [38,39]. Significant differences in the diffusion rate of the peptide in the two hydrogels were here observed in agreement with their Young’s moduli. Our results demonstrate an effect of the stiffness on the increase of the sensitivity of the tumor cells to the CIGB300 peptide treatment, which was independent by the diffusion rate in the system of the peptide in the microenvironment. The B23/NPM1 and P-gp expressions either 2D versus 3D and soft versus stiff were here assessed, observing only significant decrease of B23/NPM1 expression by 2D versus 3D at the rheological conditions used, but a significant decrease of the P-gp expression by soft versus stiff suggesting a correlation between cyclic antitumor-peptides activity and the drug resistance. In conclusion, these results suggest that the increase of the sensitivity of the CAL27 cells to the CIGB300 peptide treatment may be due to a synergistic effect with the physical rheological factor that induces a change of the drug resistance of the cancer cells, targeting the microenvironment rheology for optimized anticancer therapies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Diffusion kinetics of CIGB300 in the SF hydrogels.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, S.M. and M.V.; methodology, S.M., S.B., M.V., B.M.; validation, S.B., S.M.; formal analysis, S.M, S.B., V.S., G.P.; investigation, S.M., S.B., V.S., G.P.; resources, S.M., M.V., B.M.; data curation, S.M, S.B., M.V.; writing—original draft preparation, S..M., S.B.; writing—review and editing, S.M., M.V., B.M., S.B., S.P., H.G., V.S.; visualization, S.M.; supervision, S.M., M.V., B.M.; project administration, S.M.; funding acquisition, S.M., M.V., B.M . All authors have read and agreed to the published version of the manuscript.:.
Funding
The research leading to these results has received funding from University of Rome “Tor Vergata” for the project: Tunable 3D-Cancer cell systems for potential application in drug discovery. The research was also supported by European Union – Next Generation EU through the Italian Ministry of University and Research under PNRR - M4C2-I1.3 Project PE_00000019 “HEAL ITALIA”. The views and opinions expressed are those of the authors only and do not necessarily reflect those of the European Union or the European Commission. Neither the European Union nor the European Commission can be held responsible for them.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors declare that all data generated in this study are available within the article or the Supplementary Information. Other data related to this work are available from the corresponding authors upon request.
Conflicts of Interest
The authors declare no conflicts of financial interest or personal relationships that could have influenced the work presented in this paper.
References
- Vaupel, P. Tumor Microenvironmental Physiology and Its Implications for Radiation Oncology. Seminars in Radiation Oncology 2004, 14, 198–206. [Google Scholar] [CrossRef]
- Law, A.M.K.; Rodriguez de la Fuente, L.; Grundy, T.J.; Fang, G.; Valdes-Mora, F.; Gallego-Ortega, D. Advancements in 3D Cell Culture Systems for Personalizing Anti-Cancer Therapies. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, J.; Srivastava, T.P.; Sahoo, O.S.; Karmakar, A.; Rai, A.K.; Sarma, A.; Gogoi, G.; Alqahtani, M.S.; Abbas, M.; Dhar, R.; et al. Cancer Stem Cells: Signaling Pathways and Therapeutic Targeting. MedComm – Oncology 2023, 2, e62. [Google Scholar] [CrossRef]
- Buonvino, S.; Arciero, I.; Martinelli, E.; Seliktar, D.; Melino, S. Modelling the Disease: H2S-Sensitivity and Drug-Resistance of Triple Negative Breast Cancer Cells Can Be Modulated by Embedding in Isotropic Micro-Environment. Materials Today Bio 2023, 23, 100862. [Google Scholar] [CrossRef]
- Buonvino, S.; Di Giuseppe, D.; Filippi, J.; Martinelli, E.; Seliktar, D.; Melino, S. 3D Cell Migration Chip (3DCM-Chip): A New Tool toward the Modeling of 3D Cellular Complex Systems. Advanced Healthcare Materials n/a 2024, 2400040. [Google Scholar] [CrossRef] [PubMed]
- Ivascu, A.; Kubbies, M. Rapid Generation of Single-Tumor Spheroids for High-Throughput Cell Function and Toxicity Analysis. SLAS Discovery 2006, 11, 922–932. [Google Scholar] [CrossRef]
- Stein, A.M.; Nowicki, M.O.; Demuth, T.; Berens, M.E.; Lawler, S.E.; Chiocca, E.A.; Sander, L.M. Estimating the Cell Density and Invasive Radius of Three-Dimensional Glioblastoma Tumor Spheroids Grown in Vitro. Applied Optics 2007, 46, 5110–5118. [Google Scholar] [CrossRef] [PubMed]
- Chatzinikolaidou, M. Cell Spheroids: The New Frontiers in in Vitro Models for Cancer Drug Validation. Drug Discovery Today 2016, 21, 1553–1560. [Google Scholar] [CrossRef]
- Buonvino, S.; Ciocci, M.; Seliktar, D.; Melino, S. Photo-Polymerization Damage Protection by Hydrogen Sulfide Donors for 3d-Cell Culture Systems Optimization. International journal of molecular sciences 2021, 22, 6095. [Google Scholar] [CrossRef]
- Olive, P.L.; Durand, R.E. Drug and Radiation Resistance in Spheroids: Cell Contact and Kinetics. Cancer and Metastasis Reviews 1994, 13, 121–138. [Google Scholar] [CrossRef]
- Trédan, O.; Galmarini, C.M.; Patel, K.; Tannock, I.F. Drug Resistance and the Solid Tumor Microenvironment. JNCI: Journal of the National Cancer Institute 2007, 99, 1441–1454. [Google Scholar] [CrossRef]
- Wenzel, C.; Riefke, B.; Gründemann, S.; Krebs, A.; Christian, S.; Prinz, F.; Osterland, M.; Golfier, S.; Räse, S.; Ansari, N.; et al. 3D High-Content Screening for the Identification of Compounds That Target Cells in Dormant Tumor Spheroid Regions. Experimental Cell Research 2014, 323, 131–143. [Google Scholar] [CrossRef]
- Grimes, D.R.; Kelly, C.; Bloch, K.; Partridge, M. A Method for Estimating the Oxygen Consumption Rate in Multicellular Tumour Spheroids. Journal of The Royal Society Interface 2014, 11, 20131124. [Google Scholar] [CrossRef] [PubMed]
- CIGB Centro de Ingeniería Genética y Biotecnología. Available online: https://www.cigb.edu.cu/en/project/cigb-300-lung/ (accessed on 10th December 2025).
- Perea, S.E.; Reyes, O.; Baladron, I.; Perera, Y.; Farina, H.; Gil, J.; Rodriguez, A.; Bacardi, D.; Marcelo, J.L.; Cosme, K.; et al. CIGB-300, a Novel Proapoptotic Peptide That Impairs the CK2 Phosphorylation and Exhibits Anticancer Properties Both in Vitro and in Vivo. Molecular and Cellular Biochemistry 2008, 316, 163–167. [Google Scholar] [CrossRef]
- Perea, S.E.; Baladrón, I.; Valenzuela, C.; Perera, Y. CIGB-300: A Peptide-Based Drug That Impairs the Protein Kinase CK2-Mediated Phosphorylation. Seminars in Oncology 2018, 45, 58–67. [Google Scholar] [CrossRef]
- Borgo, C.; Ruzzene, M. Role of Protein Kinase CK2 in Antitumor Drug Resistance. Journal of Experimental & Clinical Cancer Research 2019, 38, 287. [Google Scholar] [CrossRef] [PubMed]
- Firnau, M.-B.; Brieger, A. CK2 and the Hallmarks of Cancer. Biomedicines 2022, 10, 1987. [Google Scholar] [CrossRef]
- Bruserud, Ø.; Reikvam, H. Casein Kinase 2 (CK2): A Possible Therapeutic Target in Acute Myeloid Leukemia. Cancers 2023, 15, 3711. [Google Scholar] [CrossRef] [PubMed]
- Trembley, J.H.; Kren, B.T.; Afzal, M.; Scaria, G.A.; Klein, M.A.; Ahmed, K. Protein Kinase CK2–Diverse Roles in Cancer Cell Biology and Therapeutic Promise. Molecular and cellular biochemistry 2023, 478, 899–926. [Google Scholar] [CrossRef]
- Ghani, M.U.; Shi, J.; Du, Y.; Zhong, L.; Cui, H. A Comprehensive Review on the Dynamics of Protein Kinase CK2 in Cancer Development and Optimizing Therapeutic Strategies. International Journal of Biological Macromolecules 2024, 280, 135814. [Google Scholar] [CrossRef]
- Perera, Y.; Farina, H.G.; Gil, J.; Rodriguez, A.; Benavent, F.; Castellanos, L.; Gómez, R.E.; Acevedo, B.E.; Alonso, D.F.; Perea, S.E. Anticancer Peptide CIGB-300 Binds to Nucleophosmin/B23, Impairs Its CK2-Mediated Phosphorylation, and Leads to Apoptosis through Its Nucleolar Disassembly Activity. Molecular Cancer Therapeutics 2009, 8, 1189–1196. [Google Scholar] [CrossRef]
- Perera, Y.; Costales, H.C.; Diaz, Y.; Reyes, O.; Farina, H.G.; Mendez, L.; Gómez, R.E.; Acevedo, B.E.; Gomez, D.E.; Alonso, D.F.; et al. Sensitivity of Tumor Cells towards CIGB-300 Anticancer Peptide Relies on Its Nucleolar Localization. Journal of Peptide Science 2012, 18, 215–223. [Google Scholar] [CrossRef]
- Benavent Acero, F.; Capobianco, C.S.; Garona, J.; Cirigliano, S.M.; Perera, Y.; Urtreger, A.J.; Perea, S.E.; Alonso, D.F.; Farina, H.G. CIGB-300, an Anti-CK2 Peptide, Inhibits Angiogenesis, Tumor Cell Invasion and Metastasis in Lung Cancer Models. Lung Cancer 2017, 107, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Dai, L.; Lan, Y.; Tan, C.; Vázquez-Blomquist, D.M.; Zeng, G.; Jiang, D.; Yang, K.; Perea, S.E.; Perera, Y. CIGB-300 Internalizes and Impairs Viability of NSCLC Cells Lacking Actionable Targets by Inhibiting Casein Kinase-2 Signaling. Scientific Reports 2024, 14, 26038. [Google Scholar] [CrossRef] [PubMed]
- Farina, H.G.; Benavent Acero, F.; Perera, Y.; Rodríguez, A.; Perea, S.E.; Castro, B.A.; Gomez, R.; Alonso, D.F.; Gomez, D.E. CIGB-300, a Proapoptotic Peptide, Inhibits Angiogenesis in Vitro and in Vivo. Experimental Cell Research 2011, 317, 1677–1688. [Google Scholar] [CrossRef] [PubMed]
- Perea, S.E.; Baladron, I.; Garcia, Y.; Perera, Y.; Lopez, A.; Soriano, J.L.; Batista, N.; Palau, A.; Hernández, I.; Farina, H.; et al. CIGB-300, a Synthetic Peptide-Based Drug That Targets the CK2 Phosphoaceptor Domain. Translational and Clinical Research. Molecular and Cellular Biochemistry 2011, 356, 45–50. [Google Scholar] [CrossRef]
- Open Clinical Trial, Uncontrolled in Patients Bearing Squamous Cell Carcinoma or Adenocarcinoma of the Cervix Stage IIA and IIB FIGO Classification Treated With Radiotherapy External Endocavitary Brachytherapy + Concurrent Hemotherapy Weekly Systemic and Local Application of CIGB-300 Dose Escalation 2012. Also . Available online: https://go.drugbank.com/drugs/DB16464/clinical_trials?conditions=DBCOND0029359%2CDBCOND0029361&phase=2&status=completed (accessed on 5th December 2025).
- Gottardo, M.F.; Capobianco, C.S.; Sidabra, J.E.; Garona, J.; Perera, Y.; Perea, S.E.; Alonso, D.F.; Farina, H.G. Preclinical Efficacy of CIGB-300, an Anti-CK2 Peptide, on Breast Cancer Metastasic Colonization. Scientific Reports 2020, 10, 14689. [Google Scholar] [CrossRef]
- Bellomaria, A.; Barbato, G.; Melino, G.; Paci, M.; Melino, S. Recognition of P63 by the E3 Ligase ITCH: Effect of an Ectodermal Dysplasia Mutant. Cell cycle 2010, 9, 3754–3763. [Google Scholar] [CrossRef]
- Bellomaria, A.; Barbato, G.; Melino, G.; Paci, M.; Melino, S. Recognition Mechanism of P63 by the E3 Ligase Itch: Novel Strategy in the Study and Inhibition of This Interaction. Cell cycle 2012, 11, 3638–3648. [Google Scholar] [CrossRef]
- Savioli, M.; Antonelli, L.; Bocchinfuso, G.; Cavalieri, F.; Cimino, R.; Gatto, E.; Placidi, E.; Fernandez Masso, J.R.; Garay Perez, H.; Santana, H.; et al. Formulation Matters! A Spectroscopic and Molecular Dynamics Investigation on the Peptide CIGB552 as Itself and in Its Therapeutical Formulation. Journal of Peptide Science 2022, 28, e3356. [Google Scholar] [CrossRef]
- Micsonai, A.; Wien, F.; Bulyáki, É.; Kun, J.; Moussong, É.; Lee, Y.-H.; Goto, Y.; Réfrégiers, M.; Kardos, J. BeStSel: A Web Server for Accurate Protein Secondary Structure Prediction and Fold Recognition from the Circular Dichroism Spectra. Nucleic Acids Research 2018, 46, W315–W322. [Google Scholar] [CrossRef]
- Arciero, I.; Buonvino, S.; Palumbo, V.; Scimeca, M.; Melino, S. A 3D-Printable Cell Array for In Vitro Breast Cancer Modeling. International Journal of Molecular Sciences 2024, 25. [Google Scholar] [CrossRef]
- Pradhan, S.; Hassani, I.; Seeto, W.J.; Lipke, E.A. PEG-Fibrinogen Hydrogels for Three-Dimensional Breast Cancer Cell Culture. Journal of Biomedical Materials Research Part A 2017, 105, 236–252. [Google Scholar] [CrossRef]
- You, H.; Li, Q.; Kong, D.; Liu, X.; Kong, F.; Zheng, K.; Tang, R. The Interaction of Canonical Wnt/β-Catenin Signaling with Protein Lysine Acetylation. Cellular & Molecular Biology Letters 2022, 27, 7. [Google Scholar] [CrossRef]
- Lavrentev, F.V.; Shilovskikh, V.V.; Alabusheva, V.S.; Yurova, V.Yu.; Nikitina, A.A.; Ulasevich, S.A.; Skorb, E.V. Diffusion-Limited Processes in Hydrogels with Chosen Applications from Drug Delivery to Electronic Components. Molecules 2023, 28, 5931. [Google Scholar] [CrossRef] [PubMed]
- Sheth, S.; Barnard, E.; Hyatt, B.; Rathinam, M.; Zustiak, S.P. Predicting Drug Release From Degradable Hydrogels Using Fluorescence Correlation Spectroscopy and Mathematical Modeling. Frontiers in Bioengineering and Biotechnology 2019, 7–2019. [Google Scholar] [CrossRef] [PubMed]
- Strachota, B.; Strachota, A.; Šlouf, M.; Brus, J.; Cimrová, V. Monolithic Intercalated PNIPAm/Starch Hydrogels with Very Fast and Extensive One-Way Volume and Swelling Responses to Temperature and pH: Prospective Actuators and Drug Release Systems. Soft Matter 2019, 15, 752–769. [Google Scholar] [CrossRef]
- Koetting, M.C.; Peters, J.T.; Steichen, S.D.; Peppas, N.A. Stimulus-Responsive Hydrogels: Theory, Modern Advances, and Applications. Materials Science and Engineering: R: Reports 2015, 93, 1–49. [Google Scholar] [CrossRef]
- Caccavo, D.; Cascone, S.; Lamberti, G.; Barba, A.A. Hydrogels: Experimental Characterization and Mathematical Modelling of Their Mechanical and Diffusive Behaviour. Chem. Soc. Rev. 2018, 47, 2357–2373. [Google Scholar] [CrossRef]
- Omar, J.; Ponsford, D.; Dreiss, C.A.; Lee, T.-C.; Loh, X.J. Supramolecular Hydrogels: Design Strategies and Contemporary Biomedical Applications. Chemistry – An Asian Journal 2022, 17, e202200081. [Google Scholar] [CrossRef]
- Ye, K. Nucleophosmin/B23, a Multifunctional Protein That Can Regulate Apoptosis. Cancer Biology & Therapy 2005, 4, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Tawfic, S.; Olson, M.O.; Ahmed, K. Role of Protein Phosphorylation in Post-Translational Regulation of Protein B23 during Programmed Cell Death in the Prostate Gland (∗). Journal of Biological Chemistry 1995, 270, 21009–21015. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ulloa, A.; Ramos, Y.; Gil, J.; Perera, Y.; Castellanos-Serra, L.; García, Y.; Betancourt, L.; Besada, V.; González, L.J.; Fernández-de-Cossio, J. Proteomic Profile Regulated by the Anticancer Peptide CIGB-300 in Non-Small Cell Lung Cancer (NSCLC) Cells. Journal of proteome research 2010, 9, 5473–5483. [Google Scholar] [CrossRef]
- Di Matteo, A.; Franceschini, M.; Chiarella, S.; Rocchio, S.; Travaglini-Allocatelli, C.; Federici, L. Molecules That Target Nucleophosmin for Cancer Treatment: An Update. Oncotarget 2016, 7, 44821. [Google Scholar] [CrossRef]
- Cela, I.; Di Matteo, A.; Federici, L. Nucleophosmin in Its Interaction with Ligands. International Journal of Molecular Sciences 2020, 21, 4885. [Google Scholar] [CrossRef]
- Zhu, Y.; Shi, M.; Chen, H.; Gu, J.; Zhang, J.; Shen, B.; Deng, X.; Xie, J.; Zhan, X.; Peng, C. NPM1 Activates Metabolic Changes by Inhibiting FBP1 While Promoting the Tumorigenicity of Pancreatic Cancer Cells. Oncotarget 2015, 6, 21443. [Google Scholar] [CrossRef]
- Léotoing, L.; Meunier, L.; Manin, M.; Mauduit, C.; Decaussin, M.; Verrijdt, G.; Claessens, F.; Benahmed, M.; Veyssière, G.; Morel, L.; et al. Influence of Nucleophosmin/B23 on DNA Binding and Transcriptional Activity of the Androgen Receptor in Prostate Cancer Cell. Oncogene 2008, 27, 2858–2867. [Google Scholar] [CrossRef]
- Yun, J.; Miao, J.; Chen, G.; Tian, Q.; Zhang, C.; Xiang, J.; Fu, J.; Lai, P. Increased Expression of Nucleophosmin/B23 in Hepatocellular Carcinoma and Correlation with Clinicopathological Parameters. British journal of cancer 2007, 96, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Sun, J.; Yang, L.; Yan, Y.; Shi, W.; Shi, J.; Huang, Q.; Chen, J.; Lan, Q. Upregulation of B23 Promotes Tumor Cell Proliferation and Predicts Poor Prognosis in Glioma. Biochemical and Biophysical Research Communications 2015, 466, 124–130. [Google Scholar] [CrossRef]
- Holmberg Olausson, K.; Elsir, T.; Moazemi Goudarzi, K.; Nistér, M.; Lindström, M.S. NPM1 Histone Chaperone Is Upregulated in Glioblastoma to Promote Cell Survival and Maintain Nucleolar Shape. Scientific reports 2015, 5, 16495. [Google Scholar] [CrossRef]
- Kuo, Y.; Chen, Y.; Tsai, H.; Chai, C.; Kwan, A. Nucleophosmin Overexpression Is Associated with Poor Survival in Astrocytoma. APMIS 2015, 123, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Pianta, A.; Puppin, C.; Franzoni, A.; Fabbro, D.; Di Loreto, C.; Bulotta, S.; Deganuto, M.; Paron, I.; Tell, G.; Puxeddu, E. Nucleophosmin Is Overexpressed in Thyroid Tumors. Biochemical and biophysical research communications 2010, 397, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Sasaki, H.; Kino, I.; Sugimura, T.; Terada, M. Genes Preferentially Expressed in Embryo Stomach Are Predominantly Expressed in Gastric Cancer. Cancer research 1992, 52, 3372–3377. [Google Scholar] [PubMed]
- Nozawa, Y.; Van Belzen, N.; Van Der Made, A.C.; Dinjens, W.N.; Bosman, F.T. Expression of Nucleophosmin/B23 in Normal and Neoplastic Colorectal Mucosa. The Journal of pathology 1996, 178, 48–52. [Google Scholar] [CrossRef]
- Yung, B.Y.M. Oncogenic Role of Nucleophosmin/B23. Chang Gung Medical Journal 2007, 30, 285–293. [Google Scholar] [PubMed]
- Colombo, E.; Alcalay, M.; Pelicci, P. Nucleophosmin and Its Complex Network: A Possible Therapeutic Target in Hematological Diseases. Oncogene 2011, 30, 2595–2609. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.-H. ABC Transporters as Multidrug Resistance Mechanisms and the Development of Chemosensitizers for Their Reversal. Cancer Cell International 2005, 5, 30. [Google Scholar] [CrossRef]
- Okada, T.; Tanaka, K.; Nakatani, F.; Sakimura, R.; Matsunobu, T.; Li, X.; Hanada, M.; Nakamura, T.; Oda, Y.; Tsuneyoshi, M. Involvement of P-glycoprotein and MRP1 in Resistance to Cyclic Tetrapeptide Subfamily of Histone Deacetylase Inhibitors in the Drug-resistant Osteosarcoma and Ewing’s Sarcoma Cells. International journal of cancer 2006, 118, 90–97. [Google Scholar] [CrossRef]
- Wang, L.; Chen, B.; Lin, M.; Cao, Y.; Chen, Y.; Chen, X.; Liu, T.; Hu, J. Decreased Expression of Nucleophosmin/B23 Increases Drug Sensitivity of Adriamycin-Resistant Molt-4 Leukemia Cells through Mdr-1 Regulation and Akt/mTOR Signaling. Immunobiology 2015, 220, 331–340. [Google Scholar] [CrossRef]
- Ciocci, M.; Cacciotti, I.; Seliktar, D.; Melino, S. Injectable Silk Fibroin Hydrogels Functionalized with Microspheres as Adult Stem Cells-Carrier Systems. International Journal of Biological Macromolecules 2018, 108, 960–971. [Google Scholar] [CrossRef]
- Garibbo, A.; Boragno, C.; Gagliardi, F. Patent LDO0252 Film super idrofobico–Mesap IPC–B08B17/06; B29C39/14|B29C39/148 US20120181717A1 2019. Available online: https://patents.google.com/patent/US20120181717A1/en?oq=US20120181717A1 (accessed on 20th December 2025).
- Koyanagi, M.; Kawakabe, S.; Arimura, Y. A Comparative Study of Colorimetric Cell Proliferation Assays in Immune Cells. Cytotechnology 2016, 68, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Spectroscopical characterization of the CIGB300 peptide. A) UV-vis absorption spectra of CIGB300 recorded at different concentrations, ranging from 48 µM to 326 µM. B) circular dichroic spectrum obtained at 65.5 µM of CIGB-300 C) Fluorescence spectra a) obtained at different concentrations of CIGB300 (48, 54, 65, 81, 108, 136, 145 and 163 µM) with excitation at λex=280 nm and emission at λem=300- 440 nm and b) linear regression of the fluorescence intensity at 350 nm at different concentrations of CIGB300.
Figure 1.
Spectroscopical characterization of the CIGB300 peptide. A) UV-vis absorption spectra of CIGB300 recorded at different concentrations, ranging from 48 µM to 326 µM. B) circular dichroic spectrum obtained at 65.5 µM of CIGB-300 C) Fluorescence spectra a) obtained at different concentrations of CIGB300 (48, 54, 65, 81, 108, 136, 145 and 163 µM) with excitation at λex=280 nm and emission at λem=300- 440 nm and b) linear regression of the fluorescence intensity at 350 nm at different concentrations of CIGB300.
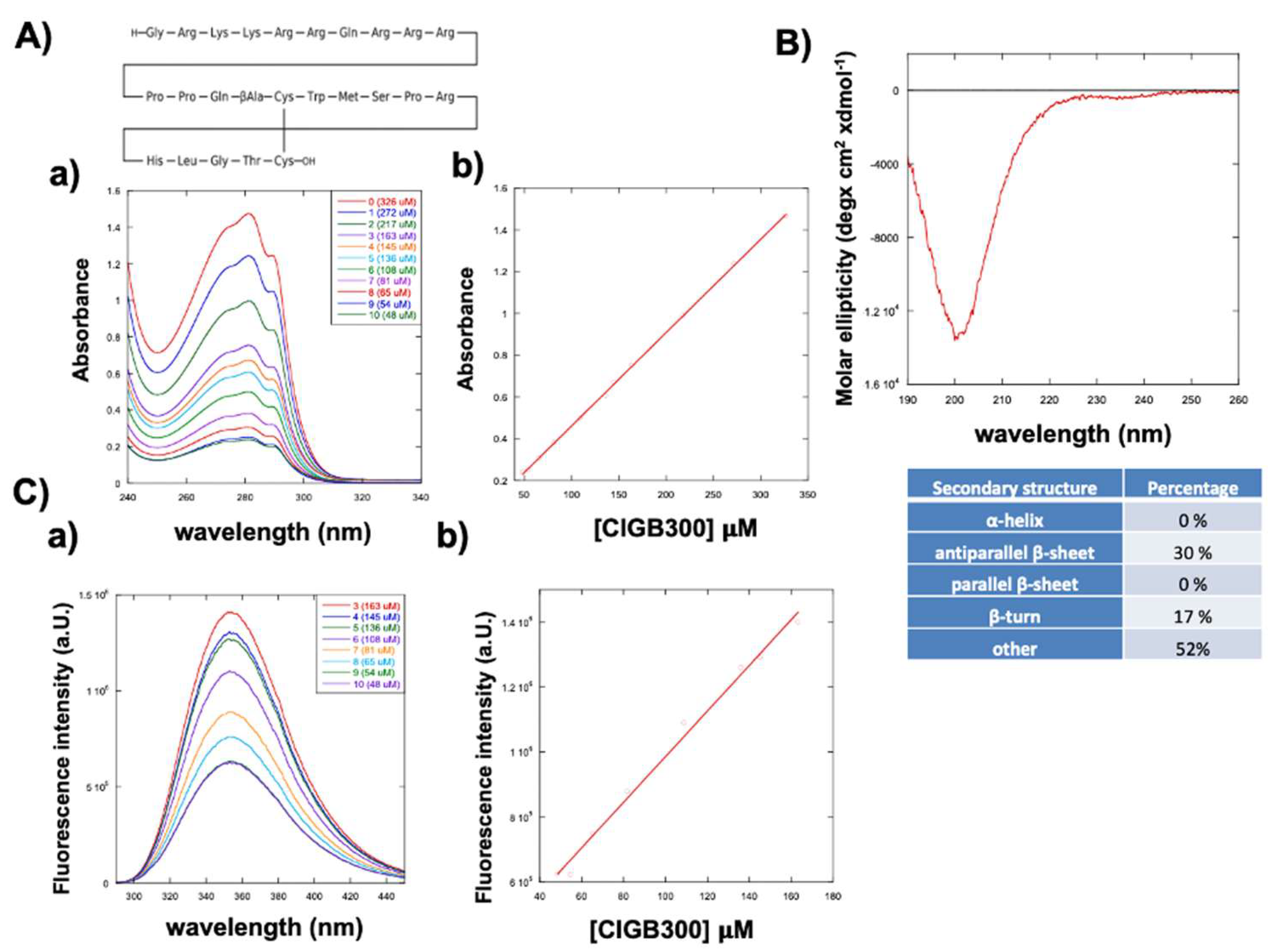
Figure 2.
Effects of CIGB300 on cell viability and migration of CAL27 cell line in 2D cell culture system. A) Inhibition of metabolic activity assessed by WST-1 assay in CAL27 after 24h of treatment with 0, 80, 160, 240 and 320 µM of CIGB300. B) Brightfield micrographs of CAL27 treated with 160 µM CIGB300 for 24 h; C) Scratch wound healing assay performed on CAL27 after 24h of treatment with 0, 80, 160 and 320 µM of CIGB300. The results are the mean of three or six biological replicates. Error bars indicate S.D. ns: non-significant, * p ≤ 0.05, *** p value ≤ 0.005, **** p < 0.001. Scale bars are of 100 µm.
Figure 2.
Effects of CIGB300 on cell viability and migration of CAL27 cell line in 2D cell culture system. A) Inhibition of metabolic activity assessed by WST-1 assay in CAL27 after 24h of treatment with 0, 80, 160, 240 and 320 µM of CIGB300. B) Brightfield micrographs of CAL27 treated with 160 µM CIGB300 for 24 h; C) Scratch wound healing assay performed on CAL27 after 24h of treatment with 0, 80, 160 and 320 µM of CIGB300. The results are the mean of three or six biological replicates. Error bars indicate S.D. ns: non-significant, * p ≤ 0.05, *** p value ≤ 0.005, **** p < 0.001. Scale bars are of 100 µm.
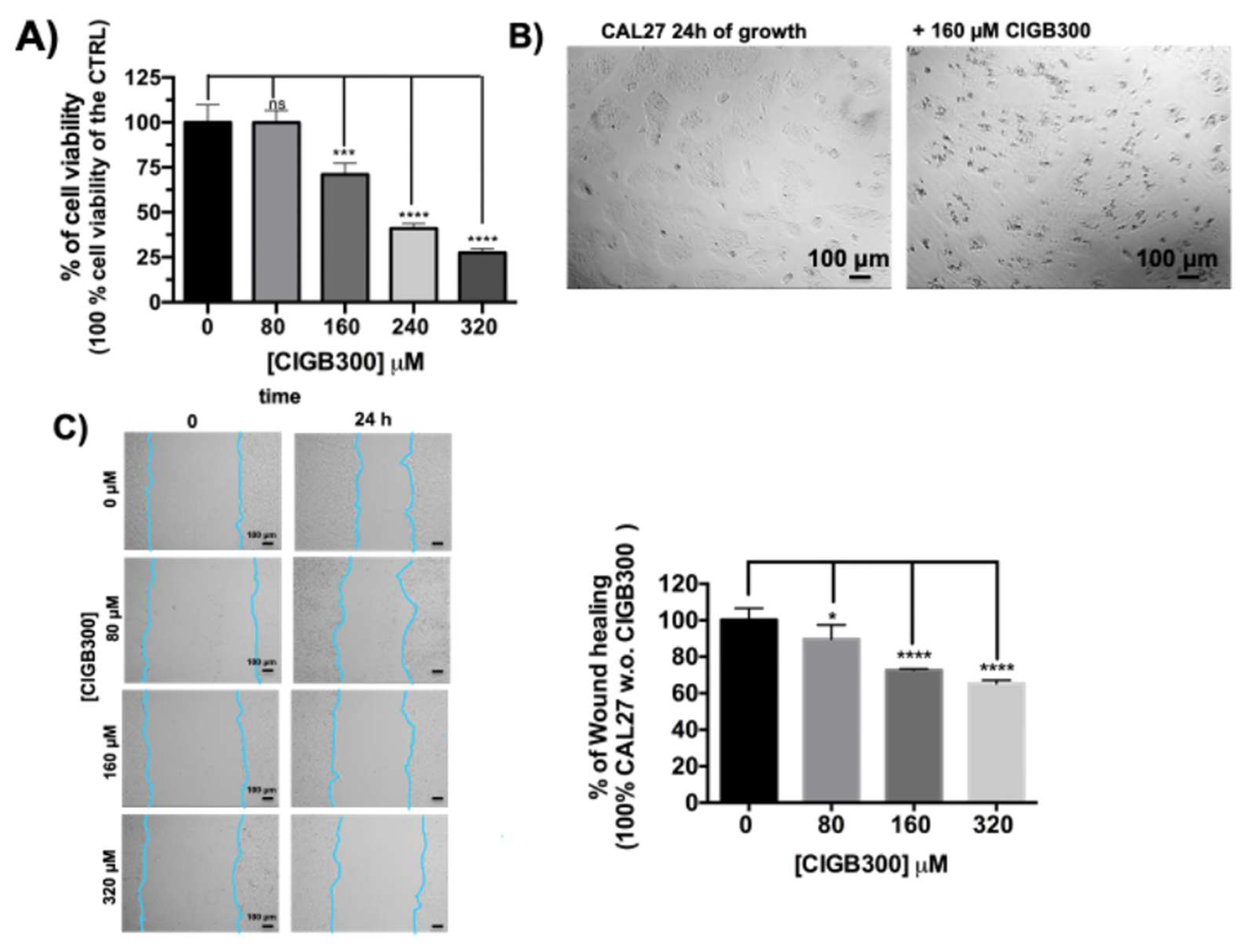
Figure 3.
CAL27 tumoroids production using PSF hydrogels. A) Schematic production of CAL27PSF tumoroids by photopolymerization of cell-spheres on the super-hydrophobic surface; B) rheological analyses of the PSFHys by nanoindentation, the Effective Young moduli (Eeff) of PSFHysoft and PSFHystiff were 3.37 (±0.03) kPa and 7.81 (±0.01) kPa respectively; C) Brightfield micrographs of CAL27PSF spheroids obtained at cell density of 1x104 cells/ µl; D) Confocal fluorescence micrographs of CAL27PSF spheroids stained with Hoechst 33342 in living conditions; E) WST-1 cell viability assay performed on CAL27PSFstiff (3µl) after 24h of cell culture. The results were obtained by three biological replicates. Error bars indicate S.D. * p ≤ 0.05. Scale bars are of 100 µm.
Figure 3.
CAL27 tumoroids production using PSF hydrogels. A) Schematic production of CAL27PSF tumoroids by photopolymerization of cell-spheres on the super-hydrophobic surface; B) rheological analyses of the PSFHys by nanoindentation, the Effective Young moduli (Eeff) of PSFHysoft and PSFHystiff were 3.37 (±0.03) kPa and 7.81 (±0.01) kPa respectively; C) Brightfield micrographs of CAL27PSF spheroids obtained at cell density of 1x104 cells/ µl; D) Confocal fluorescence micrographs of CAL27PSF spheroids stained with Hoechst 33342 in living conditions; E) WST-1 cell viability assay performed on CAL27PSFstiff (3µl) after 24h of cell culture. The results were obtained by three biological replicates. Error bars indicate S.D. * p ≤ 0.05. Scale bars are of 100 µm.
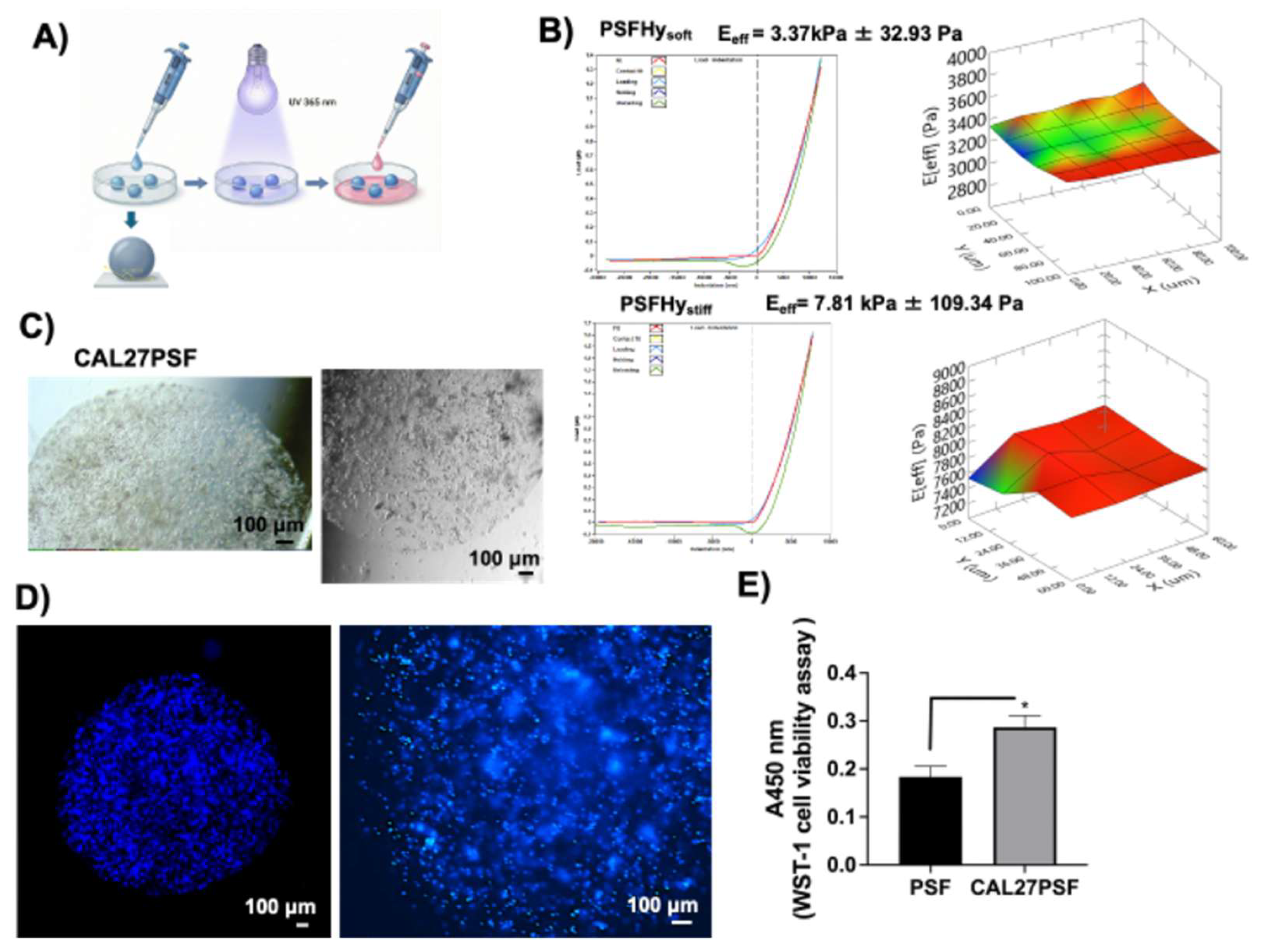
Figure 4.
CAL27PSF tumoroids cultures at different stiffness. A) Fluorescence micrographs of the LIVE/DEAD assay performed on CAL27PSFsoft and CAL27PSFstiff spheres after 3 days of cell culture; B) Fluorescence micrographs of the CAL27PSF spheres after four days of cell culture and Hoechst 33342 staining in live condition. C) Brightfield micrographs of CAL27PSFsoft and CAL27PSFstiff after 3 weeks of cell growth. Western blot analysis of CAL27PSFstiff for the evaluation of D) Cyclin D1 and E) AcHisH3 expression after 24 and 72h of cell growth. Error bars indicate S.D. ns: non-significant. Scale bars are of 50 and 100 µm.
Figure 4.
CAL27PSF tumoroids cultures at different stiffness. A) Fluorescence micrographs of the LIVE/DEAD assay performed on CAL27PSFsoft and CAL27PSFstiff spheres after 3 days of cell culture; B) Fluorescence micrographs of the CAL27PSF spheres after four days of cell culture and Hoechst 33342 staining in live condition. C) Brightfield micrographs of CAL27PSFsoft and CAL27PSFstiff after 3 weeks of cell growth. Western blot analysis of CAL27PSFstiff for the evaluation of D) Cyclin D1 and E) AcHisH3 expression after 24 and 72h of cell growth. Error bars indicate S.D. ns: non-significant. Scale bars are of 50 and 100 µm.
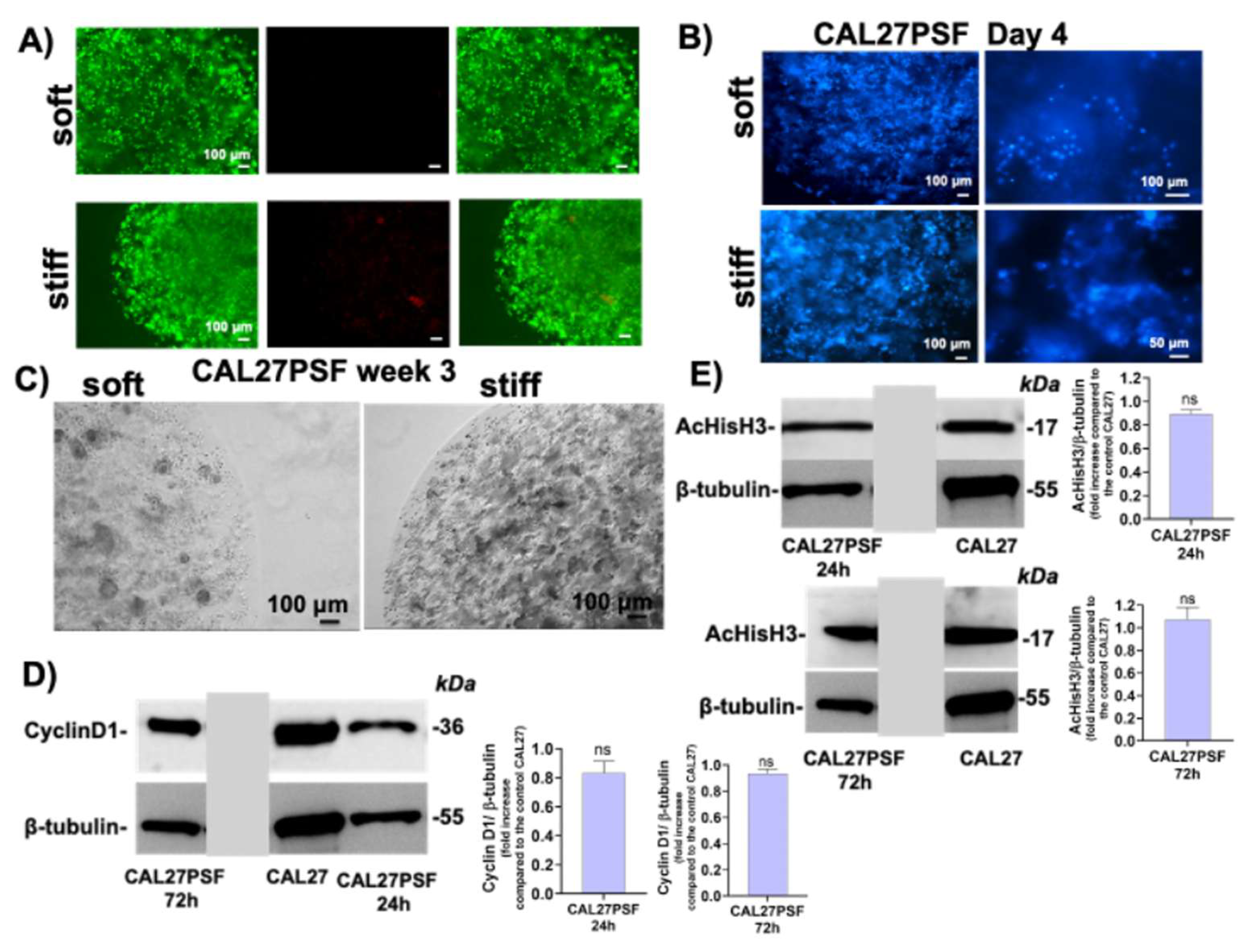
Figure 5.
Effects of the CIGB300 on the cell viability of CAL27PSF tumoroids. A) Schematic representation of the tumoroids treated with CIGB300 peptide; B) representative micrographs of the LIVE/DEAD assay of the CAL27PSFsoft and CAL27PSFstiff grown in the presence of different concentrations of CIGB300 (0, 80, 160, 320 µM) after three days of cell culture; the experiments were performed using three CAL27PSF spheres (3µl) in each well and using three biological replicates; C) WST-1 cell viability assay of the CAL27PSFstiff after three days of culture in the presence or in the absence of CIGB300 (80, 160 and 320 µM). The results were obtained by three biological replicates. Error bars indicate S.D. ** p value ≤ 0.01 *** p value ≤ 0.005, **** p < 0.001. Scale bars are of 100 µm.
Figure 5.
Effects of the CIGB300 on the cell viability of CAL27PSF tumoroids. A) Schematic representation of the tumoroids treated with CIGB300 peptide; B) representative micrographs of the LIVE/DEAD assay of the CAL27PSFsoft and CAL27PSFstiff grown in the presence of different concentrations of CIGB300 (0, 80, 160, 320 µM) after three days of cell culture; the experiments were performed using three CAL27PSF spheres (3µl) in each well and using three biological replicates; C) WST-1 cell viability assay of the CAL27PSFstiff after three days of culture in the presence or in the absence of CIGB300 (80, 160 and 320 µM). The results were obtained by three biological replicates. Error bars indicate S.D. ** p value ≤ 0.01 *** p value ≤ 0.005, **** p < 0.001. Scale bars are of 100 µm.
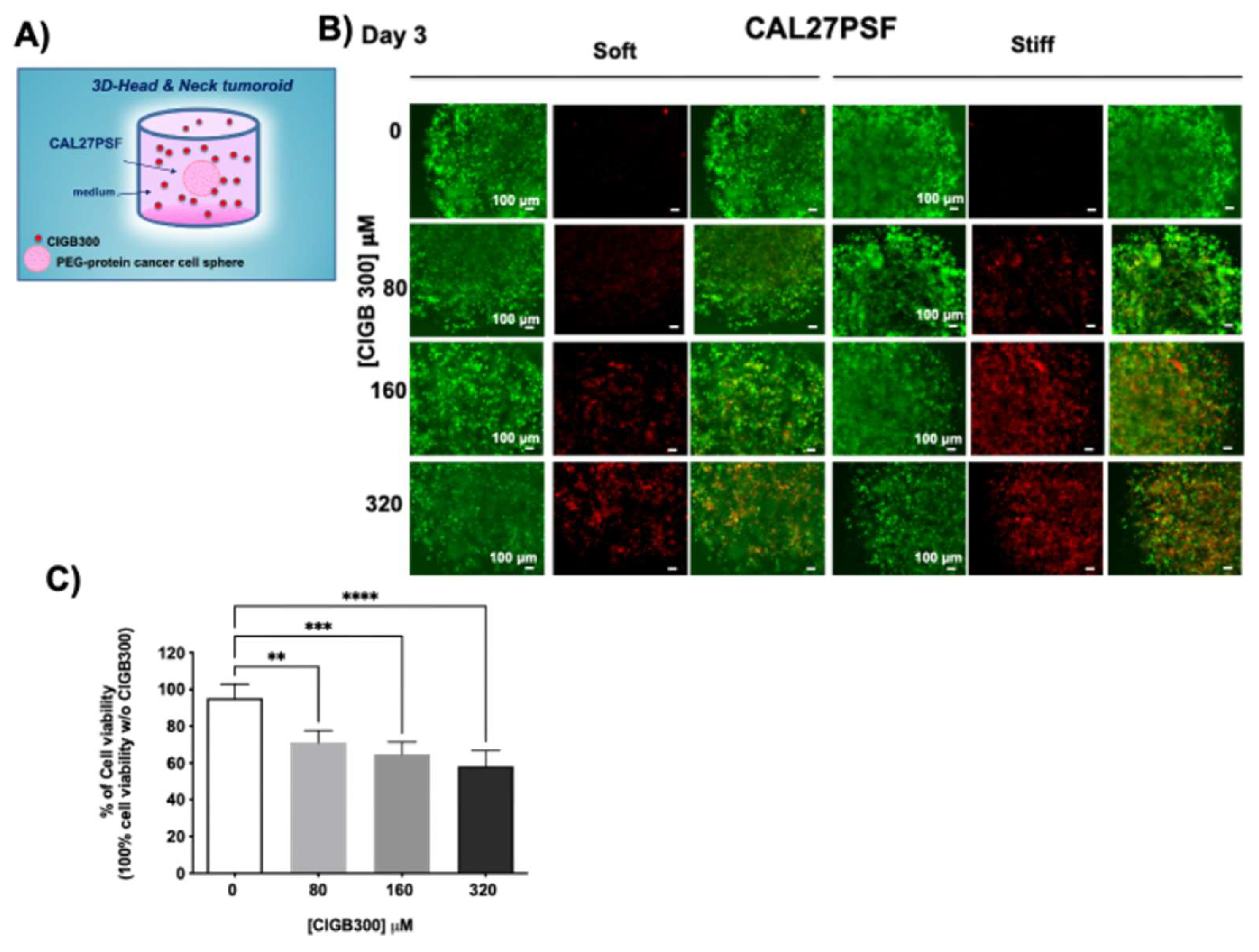
Figure 6.
Diffusion kinetics of CIGB300 in the SF hydrogels. A) Nanoindentation analysis of the soft and stiff SF hydrogels; B) Diffusion profile of FITC (11.8 nM) in the SFHystiff obtained by fluorescence spectroscopy using a wavelength excitation at 495 nm and monitoring the emission at 515 nm; C) Diffusion profiles of FITC-CIGB300 (300 µM) in the SFHysoft and SFHystiff by fluorescence spectroscopy using a wavelength excitation at λex= 495 nm and monitoring the wavelength emission at λem= 515 nm. The results were obtained by two or three experimental replicates. Error bars indicate S.D.
Figure 6.
Diffusion kinetics of CIGB300 in the SF hydrogels. A) Nanoindentation analysis of the soft and stiff SF hydrogels; B) Diffusion profile of FITC (11.8 nM) in the SFHystiff obtained by fluorescence spectroscopy using a wavelength excitation at 495 nm and monitoring the emission at 515 nm; C) Diffusion profiles of FITC-CIGB300 (300 µM) in the SFHysoft and SFHystiff by fluorescence spectroscopy using a wavelength excitation at λex= 495 nm and monitoring the wavelength emission at λem= 515 nm. The results were obtained by two or three experimental replicates. Error bars indicate S.D.
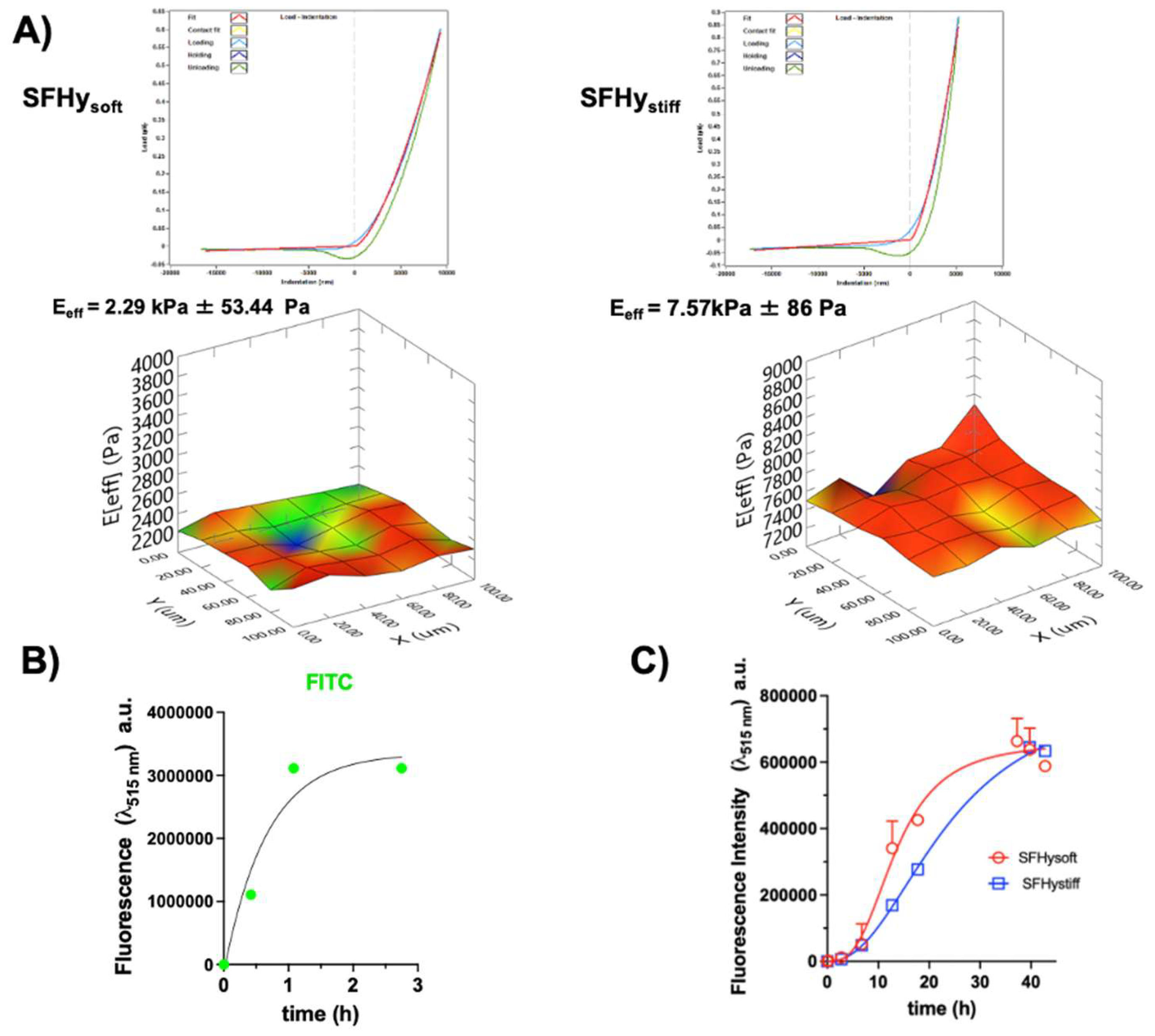
Figure 7.
Effects of the tumoroid stiffness on the P-gp and B23 expression. A) Western blotting analysis of the expression of both B23/nucleophosmin and P-gp; B) Confocal fluorescence micrographs of CAL27PSFsoft and CAL27PSFstiff after 3 days of cell culture. The P-gp expression was detected by Alexa-fluor-488-Ab-P-gp (in green) and the nuclei were stained with Hoechst 33342 in live conditions (in blue). Error bars indicate S.D. ns: non-significant; * p value ≤ 0.05; ** p value ≤ 0.01. Scale bars are of 20 and 50 µm.
Figure 7.
Effects of the tumoroid stiffness on the P-gp and B23 expression. A) Western blotting analysis of the expression of both B23/nucleophosmin and P-gp; B) Confocal fluorescence micrographs of CAL27PSFsoft and CAL27PSFstiff after 3 days of cell culture. The P-gp expression was detected by Alexa-fluor-488-Ab-P-gp (in green) and the nuclei were stained with Hoechst 33342 in live conditions (in blue). Error bars indicate S.D. ns: non-significant; * p value ≤ 0.05; ** p value ≤ 0.01. Scale bars are of 20 and 50 µm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



